Embed Size (px)
Citation preview

PAK1 tyrosine phosphorylation is required to induce epithelial-mesenchymal transition
and radioresistance in lung cancer cells
EunGi Kim1, HyeSook Youn2,3, TaeWoo Kwon1, Beomseok Son1, JiHoon Kang1, Hee
Jung Yang1, Ki Moon Seong4, Wanyeon Kim2,3 and BuHyun Youn1,2,3,*
1Department of Integrated Biological Science, Pusan National University, Busan, 609-735,
South Korea, 2Department of Biological Sciences, Pusan National University, Busan, 609-
735, South Korea, 3Nuclear Physics and Radiation Technology Institute, Pusan National
University, Busan, 609-735, South Korea, 4Department of Planning & Research for
Radiological Emergency, National Radiation Emergency Medical Center, Korea Institute of
Radiological & Medical Sciences, Seoul, 139-706, South Korea
Running title: Radioresistance mediated by Tyr phosphorylation of PAK1
Key words: PAK1, Tyrosine phosphorylation, Protein stability, Epithelial-mesenchymal
transition, Radioresistance
Grant Support: This work was supported by the Radiation Technology R&D Program
through the National Research Foundation of Korea funded by the Ministry of Science, ITC
& Future Planning (2013M2A2A7042502), a grant from the National R&D Program for
Cancer Control, Ministry for Health and Welfare, Republic of Korea (1320100), and the
Basic Science Research Program through the National Research Foundation of Korea funded
by the Ministry of Education, Science and Technology (2013R1A1A2059832 to W Kim).
*Correspondence should be addressed to
BuHyun Youn, Ph.D.
Department of Biological Sciences, Pusan National University
Busandaehak-ro 63beon-gil, Geumjeong-gu, Busan 609-735, South Korea
Tel: 82-51-510-2264; Fax: 82-51-581-2962; E-mail: [email protected]
Word count: 4996
Total numbers of figures and tables: 7
Conflict of interest: The authors declare that they have no conflict of interest.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Abstract
PAK1 kinase has an essential role in tumorigenesis and cell survival in many cancers
but its regulation is not fully understood. In this study, we showed that in response to
irradiation of lung cancer cells, PAK1 was upregulated, tyrosine phosphorylated and
translocated to the nucleus. Tyrosine phosphorylation relied upon JAK2 kinase activity and
was essential for PAK1 protein stability and binding to Snail. This radiation-induced
JAK2/PAK1/Snail signaling pathway increased epithelial-mesenchymal transition (EMT) by
regulating epithelial and mesenchymal cell markers. Notably, JAK2 inhibitors mediated
radiosensitization and EMT blockade in a mouse xenograft model of lung cancer. Taken
together, our findings offered evidence that JAK2 phosphorylates and stabilizes functions of
PAK1 that promote EMT and radioresistance in lung cancer cells, with additional
implications for the use of JAK2 inhibitors as radiosensitizers in lung cancer treatment.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Introduction
Radiotherapy can be a preferential management strategy for patients with inoperable
cancer types, including advanced stage of non-small cell lung cancer (NSCLC), in the
absence of more effective targeted therapies (1). Nevertheless, therapeutic outcomes are not
fully satisfactory due to the emergence of radioresistance which is considered to be a critical
obstacle that leads to the failure of radiation therapy and consequently increases mortality in
NSCLC patients (2). So far, a large number of studies have been conducted to find way to
control radioresistance and develop potent adjuvants for radiotherapy that enhance treatment
efficacy (3, 4). A profound understanding of the molecular events associated with therapeutic
resistance would greatly advance the discovery of drugs enabling to modulate radiation-
induced signaling responses to improve the prognosis of NSCLC patient (5-7).
We previously found that the p21-activated Ser/Thr kinase 1 (PAK1) is
overexpressed in radioresistant NSCLC cells exposed to ionizing radiation (IR) (8). PAK1 is
known to be broadly expressed in various normal tissues, but PAK1 is also significantly
overexpressed in some types of cancer, such as ovarian, breast, and bladder cancers (9).
Several functional studies have reported that PAK1 is highly associated with cell
transformation and tumorigenesis, supported by the development of premalignant lesions and
tumor formation in mammary glands induced by PAK1 overexpression and hyperactivation
(10, 11). This evidence implies that PAK1 may contribute to tumor initiation and acquisition
of tumor aggressiveness under certain cellular conditions. In addition, other reports have
shown that activated PAK1 can protect against chemotherapy- and anoikis-induced cell death
(12, 13). Genomic analysis has shown that gene amplification and overexpression of PAK1
can be detected in the cytoplasm and nucleus of cells from NSCLC patients, and selective
inhibition targeting PAK1 can negatively regulate cell proliferation both in vitro and in vivo
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

(13). Many undefined mechanisms may be involved, and the effects of PAK1 on the
apoptosis of human NSCLC cells have to be thoroughly investigated.
It is known that PAK1 activity is dependent on phosphorylation of Ser/Thr residues
(9). The role of phosphorylation at Tyr residues in the regulation of PAK1 function has not
been fully elucidated compared to phosphorylation at Ser/Thr sites. Prolactin-activated JAK2
phosphorylates the three Tyr residues in PAK1 and subsequently increases PAK1 kinase
activity in Nb2 cells (14). In addition to JAK2, the non-receptor tyrosine kinase ETK/Bmx, a
member of the Tec family, can interact with PAK1, leading to the phosphorylation and
activation of PAK1 (15). Furthermore, PAK1 can form multifactor complexes with several
proteins, including Shc, Grb2, and Nck, and mediates specific responses governed by
signaling induced by Epithelial growth factor receptor (EGFR), a well-known receptor Tyr
kinase (16). Altogether, these data serves as the critical evidence demonstrating that the
modulation of PAK1 activity by Tyr phosphorylation may play a significant role in the
control of cellular responses to various stimuli.
JAK2 is a Tyr kinase, a Janus kinase family member. JAKs are well-known to
activate STAT signaling, leading to regulation of the gene expression that influences
processes associated with tumor malignancy including cell cycle progression, angiogenesis,
metastasis, and immune evasion (17, 18). Many investigations have demonstrated that JAKs
and STATs are highly activated in cases of lung cancer and implied that JAKs as upstream
regulators can be promising targets for lung cancer therapy (19, 20). For this, small-molecule
compounds targeting JAKs and their signaling activity have been currently developed as
chemotherapeutic reagents. However, only a few studies have reported a relationship JAKs
and radiation-associated signaling. It has been demonstrated that the induction of c-jun
expression in irradiated cells requires the activation of JAK3, but is impervious to the
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

activation status of JAK1 and JAK2 (21). A recent study using TG101209, a small-molecule
inhibitor of JAK2, found that JAK2 inhibition could be a promising strategy for lung cancer
treatment (22). Therefore, it is important to define the role of the JAK2 pathway in irradiated
solid tumors, since better therapies are desperately needed to treat these malignancies.
The purpose of the present study was to elucidate the mechanisms and key molecules
that confer radioresistance in NSCLC cells. We demonstrated that IR-dependent
phosphorylation of PAK1 on Tyr residues by JAK2 and its subsequently increased stability
play an important role in survival signal transduction in NSCLC cells. Our findings provide a
possible explanation of how NSCLC cells can acquire and regulate resistance to radiation.
Furthermore, we suggest that JAK2 and PAK1 could be attractive pharmaceutical targets for
overcoming radioresistance of NSCLC and ultimately contribute to the effective radiation
treatment of lung cancer.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Materials and Methods
Chemicals, antibodies and reagents
Chemicals, antibodies, and reagents used are described in Supplementary Materials
and Methods.
Cell lines, cell culture, irradiation and drug treatment
A549 and NCI-H460 cells were acquired from the American Type Culture Collection
(ATCC, Manassas, VA), authenticated, and maintained in early passages, no more than 6
months after receipt from ATCC. Cells were grown in RPMI-1640 medium supplemented
with 10% FBS, 100 U/mL penicillin and 100 �g/mL streptomycin at 37°C in 95% air/5%
CO2. The cells were exposed to a single dose of γ-rays using a Gamma Cell-40 Exactor
(Nordion International, Inc., Kanata, Ontario, Canada) at a dose rate of 0.81 Gy/min. Flasks
containing the control cells were placed in the irradiation chamber but not exposed to
radiation. The cells were treated with the indicated drug dissolved in dimethyl sulfoxide
(DMSO) for 4 h.
Northern blot analysis
Northern blot analysis was performed as described previously (23). The detailed
conditions and primers used for mRNA expression are listed in Supplementary Materials and
Methods.
Western blot analysis, immunoprecipitation (IP), and in vivo kinase assay
Following the experimental treatment, Western blot analysis, IP, and in vivo kinase
assay studies were performed as previously described (4, 6, 23). Detailed procedures are
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

described in Supplementary Materials and Methods.
Transient transfection
For transient transfection, cells were plated at a density of 5 × 105 cells in 6-well
dishes and incubated for 4 h. The cells were transiently transfected with the indicated plasmid
using Lipofectin (Invitrogen, Carlsbad, CA) or the siRNA oligonucleotides (10 nM) targeting
EGFR, c-Abl, JAK1, JAK2, and JAK3 using DharmaFECT 1 (Dharmacon, Chicago, IL)
according to the manufacturers’ instructions.
Interactome analysis
For interactome analysis, cells were transfected with Myc-PAK1 and harvested after
being irradiated (4 Gy, 2 h). Nuclear extract was then used for double immunoaffinity
purification of PAK1-containing complexes as previously described (24). Detailed
procedures are summarized in Supplementary Materials and Methods.
Luciferase reporter gene assay
Following co-transfection with E-cadherin (E-cad-Luc) or occludin Luciferase
reporter gene (Occlu-Luc) and specific combination of plasmids as indicated, the medium
was changed and the cells were treated with siRNA, drug and/or irradiation. After 1 h, the
cells were washed twice with cold PBS and lysed in reporter lysis buffer (Promega, Madison,
WI). After vortexing the lysates and centrifugation at 12,000 g for 1 min at 4°C, the cell
extract and luciferase assay reagent (Promega) were mixed at room temperature, and placed
in a luminometer (AutoLumat LB 953, EG & G Berthold, Bad Widbad, Germany) to measure
luciferase activity.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Cell assay (3D culture) and immunofluorescence (IF) staining
The 3D acini culture was conducted to detect morphologic changes of the epithelial
acini as previously described (25). Detailed conditions are described in Supplementary
Materials and Methods.
Transwell cell migration assay
A cell migration assay was performed using a 24-well Transwell chamber (Corning,
Corning, NY). Cells (1 × 104 in serum-free RPMI-1640 medium) cultured with or without
specific treatments (transfection, IR and/or drugs) for 60 h (A549) or 72 h (NCI-H460) were
seeded in the upper chamber of a 5 �m pore-size insert. The lower chamber was filled with
RPMI-1640 medium containing 2% FBS. After 6 h, the upper membrane surface was wiped
with a cotton swab to remove cells that had not migrated into the lower chamber. Migrated
cells attached to the lower membrane surface were fixed with 4% paraformaldehyde, stained
with hematoxylin, and counted. The migration index was calculated and normalized to the
number of untreated cells that had migrated and the results were based on the relative number
of cells in a randomly selected field from three representative experiments.
Wound healing assay
Cell monolayers were scratched using a 200 �L pipette tip after the cells had reached
70% confluency in RPMI-1640 medium supplemented with 1% FBS. The cells were further
incubated with fresh medium with or without treatment for 18 h and 30 h (A549) or 24 h and
48 h (NCI-H460). Photomicrographs were then taken at 100× magnification with an Olympus
IX71 fluorescence microscope (Olympus Optical Co. Ltd., Tokyo, Japan).
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Real-time quantitative RT-PCR (qRT-PCR)
The level of EMT-related gene expression was measured using qRT-PCR as
previously described (23). The detailed conditions and primers used for mRNA expression
are listed in Supplementary Materials and Methods including Table S1.
Tumor xenografts in nude mice
Six-week-old male BALB/c athymic nude mice (Central Lab Animals Inc., Seoul,
South Korea) were injected with 2 × 106 A549 or NCI-H460 cells into the flank and tumors
were allowed to develop. Upon identification of a palpable tumor (minimal volume of 200
mm3), DMSO or drug (200 �g/kg body weight) was administered intraperitoneally every day
for 25 d. The animals were also irradiated with 10 Gy once a week for 3 wk. Tumor length (L)
and width (W) were measured with a caliper and tumor volumes were calculated with the
formula (L × W2)/2. At the end of the treatment period, the animals were euthanized and the
tumors were used for biochemical studies. Animal care protocol is detailed in Supplementary
Materials and Methods.
Statistical analysis
All numeric data are presented as the mean ± SD or SEM from at least three
independent experiments. The results were analyzed using a one-way ANOVA for ranked
data followed by Tukey's honestly significant difference test, and the two-way ANOVA for
ranked data followed by a Bonferroni post test. Prism 4 software (GraphPad Software,
SanDiego, CA) was used to conduct all statistical analyses. A p-value < 0.05 was considered
to be statistically significant.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Results
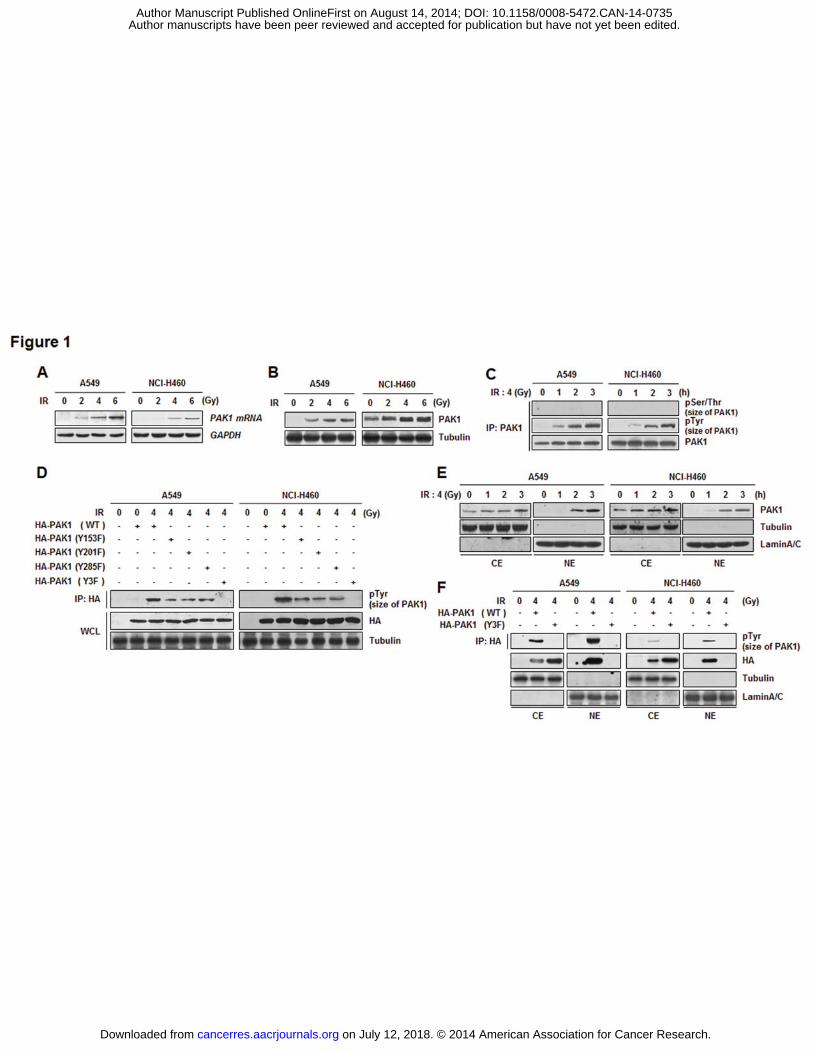
IR induces overexpression, Tyr phosphorylation, and nuclear localization of PAK1 in
NSCLC cells
Nuclear and cytoplasmic PAK1 is strongly expressed in squamous NSCLC cells (13).
Based on this information and our previous transcriptome analysis (8), we measured the
expression of PAK1 in irradiated A549 and NCI-H460 cells in the present study. As shown in
Fig. 1A and B, IR-exposure of A549 and NCI-H460 cells dramatically induced both mRNA
and protein expression of PAK1 in a dose-dependent manner. Next, IR-dependent molecular
modification of PAK1 was investigated since PAK1 was known to be phosphorylated for its
activity. In both NSCLC cells, IR-induced PAK1 phosphorylation on Tyr sites was observed,
whereas phosphorylation at Ser/Thr sites was not detected (Fig. 1C and Supplementary Fig.
S1A). Based on the literature and 3-dimensional structure of PAK1 (PDB ID: 1F3M), to
further determine which residues are the targets of Tyr kinase in PAK1, we used each mutant
(Y153F, Y201F, Y295F, or Y3F) as a substrate for an in vivo kinase assay (14, 26). Tyr
phosphorylation was reduced in Y153F, Y201F, or Y285F mutants of PAK1, and completely
eliminated from the PAK1 Y3F (Y153F/Y201F/Y285F) mutant (Fig. 1D and Supplementary
Fig. S1B). These findings indicated that Y153, Y201, and Y285 of PAK1 are required for Tyr
phosphorylation in irradiated A549 and NCI-H460 cells. It has been reported that PAK1
contains nuclear localizing signal sequences and signal-dependent nuclear localization is
required for transcriptional modulation of PAK1 (27, 28). To determine whether IR can
change the subcellular localization of PAK1, the protein levels of PAK1 in the cytoplasmic or
nuclear region were measured in irradiated A549 and NCI-H460 cells. As shown in Fig. 1E,
IR increased the amount of nuclear PAK1 in a time-dependent manner. To further examine
whether IR-induced Tyr phosphorylation affects the nuclear localization of PAK1, we
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

conducted an IP assay of the cytoplasmic or nuclear fraction from irradiated A549 and NCI-
H460 cells after transfection with HA-PAK1 (WT or Y3F). We found that IR-induced nuclear
localization of the PAK1 Y3F mutant was significantly inhibited compared to that of PAK1
WT (Fig. 1F). Collectively, these data suggest that IR induces overexpression, Tyr
phosphorylation, and, subsequently, nuclear localization of PAK1.
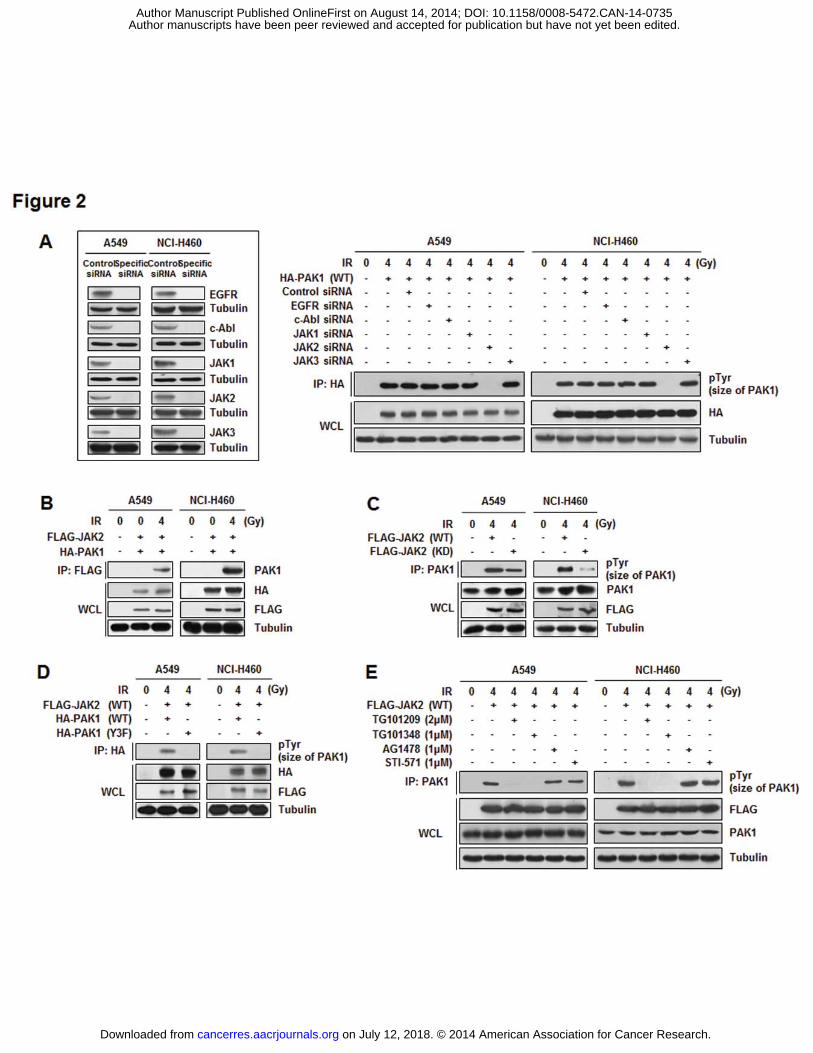
JAK2 phosphorylates PAK1 in irradiated NSCLC cells
Since we found that IR induced the phosphorylation of PAK1 on Tyr residues in
NSCLC cells, we further examined upstream kinases of PAK1. SiRNA specific for each Tyr
kinase (EGFR, c-Abl, JAK1, JAK2, and JAK3) was prepared and an in vivo kinase assay was
performed on the cells expressing these siRNA molecules (Fig. 2A, inset). Inhibition of JAK2
expression led to complete abolishment of PAK1 phosphorylation in irradiated A549 and
NCI-H460 cells; JAK2 was therefore selected as a candidate for upstream kinase of PAK1
(Fig. 2A). We next assessed the interaction between JAK2 and PAK1 in both irradiated
NSCLC cells. It was observed that JAK2 was activated by irradiation and interacted with
PAK1 in cytoplasm (Fig. 2B and Supplementary Fig. S2). Moreover, PAK1 was
phosphorylated at Tyr sites by JAK2 WT but not by JAK2 kinase-dead (KD) after IR
exposure (Fig. 2C). To further investigate whether the Y153, Y201, and Y285
phosphorylation sites of PAK1 are targets of JAK2, an in vivo kinase assay was performed
with the PAK1 Y3F mutant. As shown in Fig. 2D, JAK2-induced PAK1 phosphorylation was
eliminated by the Y3F mutation in PAK1, indicating that the suggested three residues of
PAK1 are required for JAK2-mediated phosphorylation in irradiated A549 and NCI-H460
cells. Direct phosphorylation of PAK1 by JAK2 was confirmed by treating A549 and NCI-
H460 cells with two JAK2-specific inhibitors, TG101209 and TG101348. IR-induced PAK1
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

phosphorylation by JAK2 was significantly diminished following treatment with TG101209
and TG101348, but not AG1478 (an EGFR inhibitor) or STI-571 (a c-Abl inhibitor) (Fig. 2E).
Therefore, we propose that IR-activated PAK1 phosphorylation at Tyr residues is directly
mediated by JAK2 in NSCLC cells.
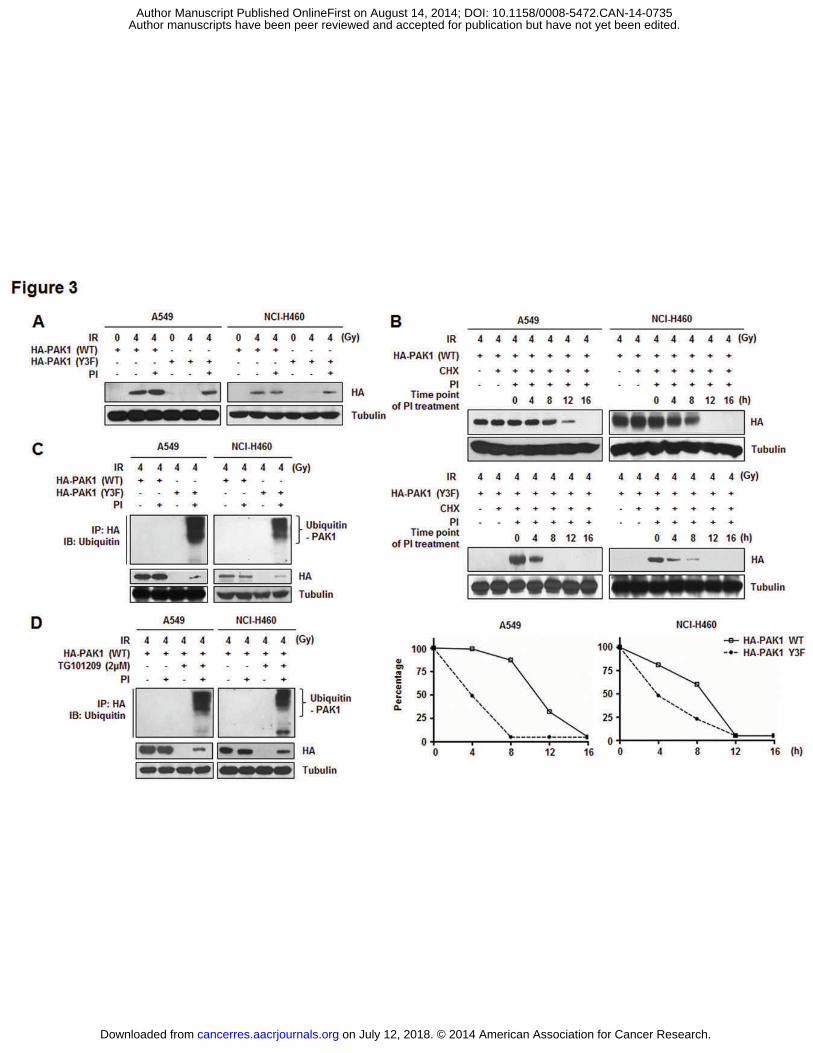
Tyr phosphorylation is required for maintaining PAK1 protein stability in irradiated
NSCLC cells
To explore the biological effects of PAK1 Tyr phosphorylation, both HA-PAK1 WT
and HA-PAK1 Y3F were expressed in NSCLC cells by transient transfection. As shown in
Fig. 3A, HA-PAK1 Y3F was expressed at a lower level under IR compared to the WT
construct. Treatment with proteasome inhibitors (PI) restored the mutant protein level,
indicating that Tyr phosphorylation of PAK1 modulates protein stability. Indeed, the
estimated half-life of PAK1 WT (12 h) was significantly longer than that of the Y3F mutant
(4 h; Fig. 3B). These results indicate that cellular PAK1 is subject to proteasome-dependent
degradation and that phosphorylation on Tyr residues prevents PAK1 degradation. In line
with these findings, proteasome inhibitors caused a more pronounced accumulation of poly-
ubiquitinated PAK1 Y3F than PAK1 WT (Fig. 3C). Previously, we observed that PAK1 is
phosphorylated by JAK2 after irradiation (Fig. 2). To test whether the same mechanism also
mediates the poly-ubiquitination that is associated with PAK1 Y3F, JAK2 activation was
blocked by a JAK2-specific inhibitor. The results showed that JAK2 inhibition did increase
the level of poly-ubiquitinated PAK1 WT, indicating that the protein stability of PAK1 is
controlled by a signaling pathway that is dependent on Tyr phosphorylation of PAK1 induced
by JAK2 after irradiation in NSCLC cells (Fig. 3D). According to these data, we propose that
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

IR-activated PAK1 Tyr phosphorylation by JAK2 is directly related to PAK1 protein
stabilization in NSCLC cells.
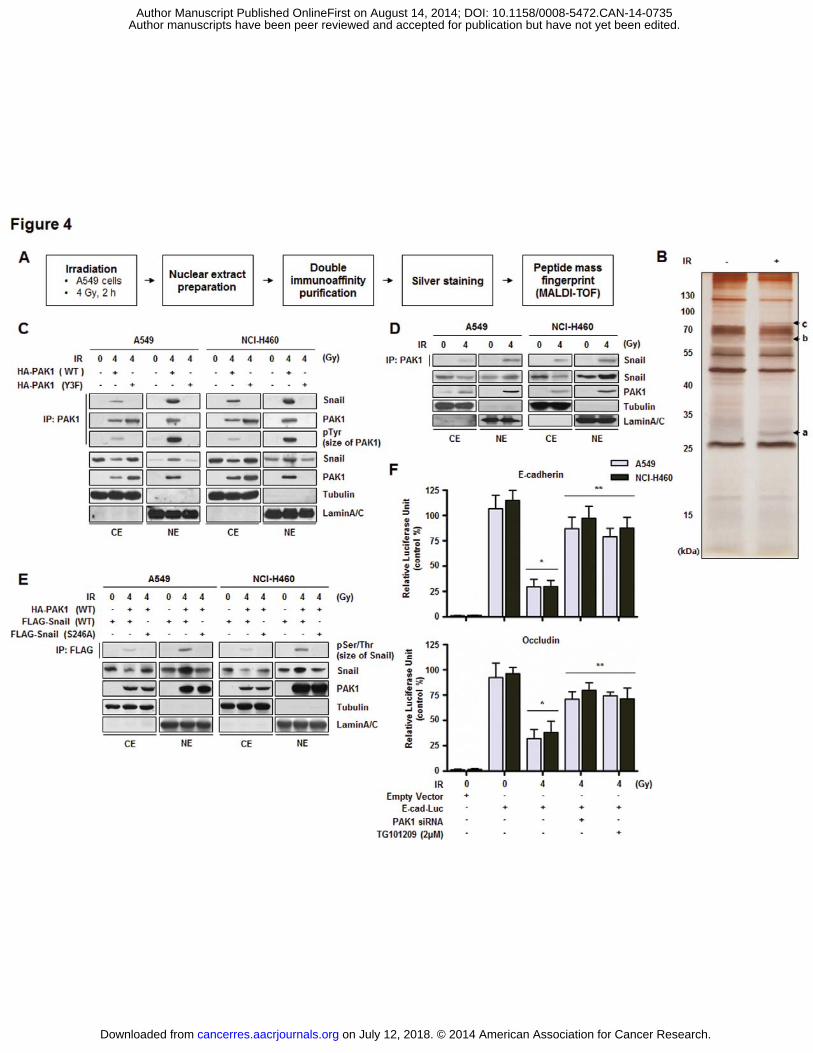
PAK1 interacts with Snail in irradiated NSCLC cells
Since we found that PAK1 was phosphorylated by IR-activated JAK2 and localized
to the nucleus, we further set out to identify a PAK1-interacting proteome in irradiated
NSCLC cells to elucidate the molecular functions of PAK1 in the nucleus. The strategy used
to identify nuclear proteins associated with nuclear PAK1 is presented in Fig. 4A. Nuclear
extracts were prepared from A549 cells cultured with IR and soluble nuclear protein
complexes were separated to reduce sample complexity. Complexes containing PAK1 were
purified using double immunoaffinity purification, and the identity of PAK1 interactors was
determined by repeated rounds of Peptide mass fingerprint (PMF) (29). As shown in Fig. 4B,
several IR-dependent PAK1 interacting proteins were identified with high confidence. Based
on information from published databases of physical and functional interactions, we focused
on Snail (indicated band (‘a’) in Fig. 4B), a key regulator of the epithelial-mesenchymal
transition (EMT) (30). We subsequently confirmed the interaction between PAK1 and Snail
(Fig. 4C and Fig. 4D). Unlike PAK1 Y3F, PAK1 WT interacted with Snail, suggesting that
the interaction of PAK1 with Snail in irradiated NSCLC cells is phosphorylation-dependent.
A previous report has shown that the phosphorylation of Snail on S246 by PAK1 is essential
for Snail function by promoting transcriptional repression of several genes, including
epithelial marker proteins (E-cadherin), in breast cancer cell lines (30). As shown in Fig. 4E,
PAK1-mediated Snail phosphorylation was detected and eliminated by an S246A mutation in
Snail, indicating that S246 of Snail is required for PAK1-mediated phosphorylation in
irradiated NSCLC cells. To explore a potential effect of PAK1 signaling on the functions of
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Snail, we next examined the influence of PAK1 on the ability of Snail to repress E-cadherin
promoter activity. Our results demonstrated that IR-activated PAK1 had an inhibitory effect
on E-cadherin promoter activity (Fig. 4F). We next showed that inhibition of the PAK1
pathway by PAK1-specific siRNA resulted in increased transcriptional activity of the E-
cadherin promoter in irradiated NSCLC cells. The involvement of phosphorylated PAK1 in
the ability of Snail to repress E-cadherin promoter activity was confirmed by treatment with a
JAK2-specific inhibitor, TG101209 (Fig. 4F). To support the generality of our findings, we
assessed whether PAK1 activity affects another Snail-regulated gene, Occludin (31). It was
observed that IR-activated PAK1 also promoted the Snail-induced transcriptional repression
of Occludin-promoter reporter constructs (Fig. 4F). According to these results, we propose
that IR-activated PAK1 directly interacts with Snail and regulates transcriptional repression
activity of Snail by Ser phosphorylation in NSCLC cells.
Inhibition of Tyr phosphorylation of PAK1 reduces IR-dependent EMT in NSCLC cells
Several lines of evidence have demonstrated the critical role of PAK1 and Snail in
the EMT (30, 32). To explore a potential role for PAK1 phosphorylation in Snail-mediated
EMT and the effect of a JAK2 inhibitor as an EMT suppressor, we monitored morphologic
changes of NSCLC cells. A549 and NCI-H460 cells were grown on a thick layer of Matrigel
to form epithelial acini (3D culture model). This system has been used extensively to identify
morphological changes that can perturb the normal cancer growth architecture (33, 34). In
this 3D culture model, irradiated NSCLC cells were distinguishable from the control cells and
formed an increased number of acini that invaded the Matrigel-containing 3D matrix.
Interestingly, the observed morphological changes of the epithelial acini were abrogated by
the addition of TG101209, indicating that the inhibition of PAK1 Tyr phosphorylation
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

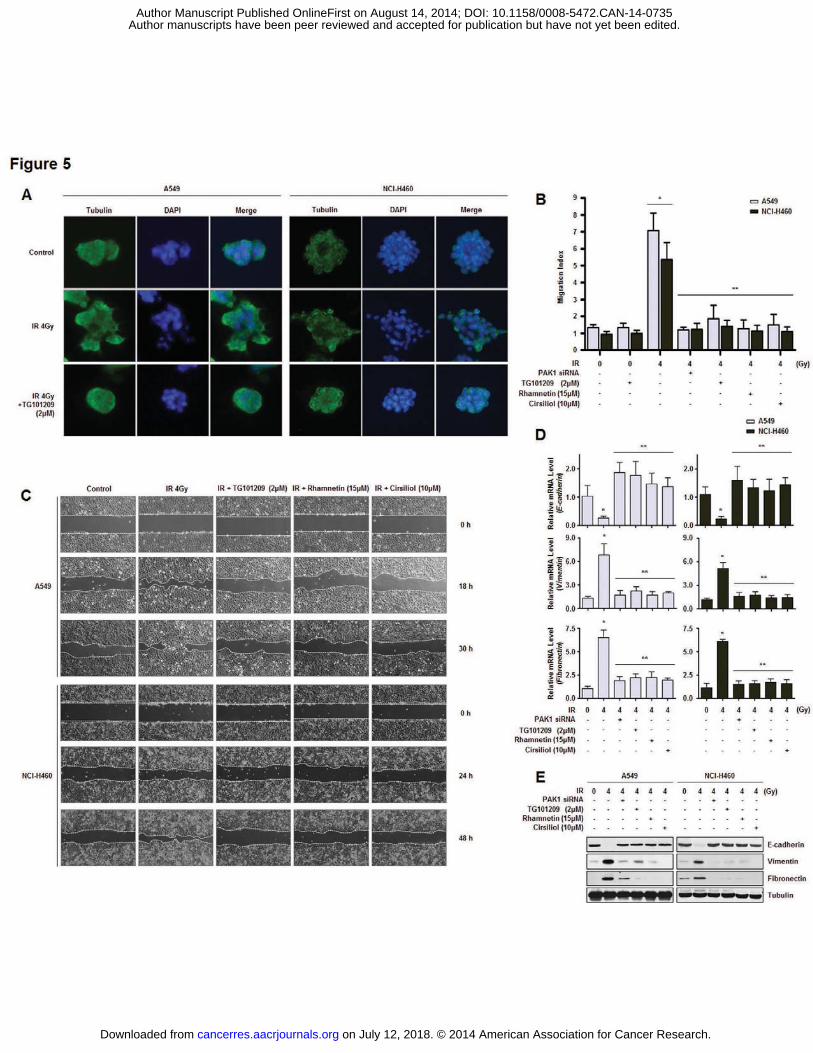
reduced IR-dependent EMT in the NSCLC cells (Fig. 5A). Next, we measured the migration
capacity and EMT marker expression of NSCLC cells treated with TG101209. NSCLC cells
treated with the JAK2 inhibitor exhibited reduction of motility as determined by both
Transwell cell migration and wound healing assays (Fig. 5B and C). Rhamnetin and cirsiliol
were administered as positive controls of EMT inhibitors (35). TG101209 treatment
alleviated IR-induced EMT by increasing the expression of E-cadherin (an epithelial marker)
while decreasing the expression of Vimentin and Fibronectin (two mesenchymal markers) at
both mRNA and protein levels (Fig. 5D and E). Reduced EMT was also observed in the
NSCLC cells in which PAK1 expression was knocked down. Taken together, these results
suggest that the inhibition of PAK1 activation in NSCLC cells suppresses the EMT.
JAK2-inhibitor increases in vivo radiosensitization and decreases in vivo EMT in a
xenograft mouse model
In our wound healing assay, we observed that TG101209 treatment prevented not
only the EMT but also the proliferation of IR-activated NSCLC cells (Fig. 5C). To evaluate
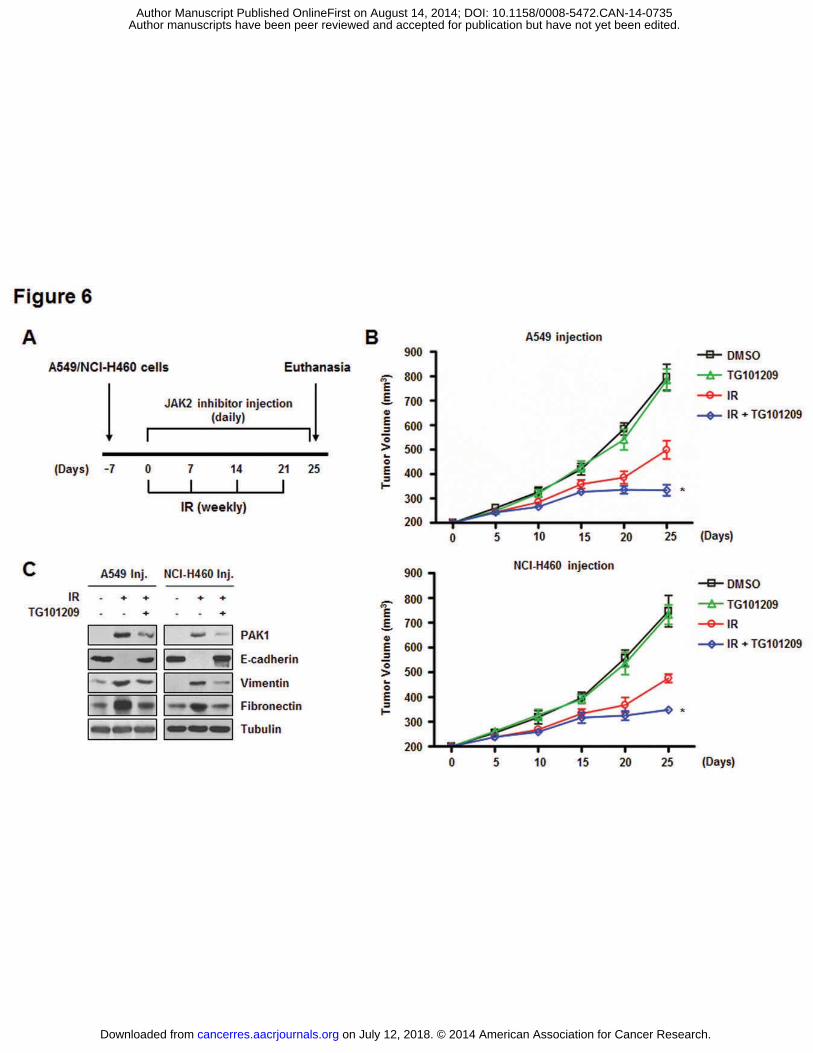
the combined effects of JAK2 inhibition with IR on tumor growth in vivo, a xenograft mouse
model was established (Fig. 6A). In vivo data from nude mice bearing tumors formed by
A549 and NCI-H460 cells indicated that TG101209 had an in vivo radiosensitization effect
(Fig. 6B). Tumor volumes of the mice treated with IR and TG101209 were significantly
reduced by approximately 57% (the group with tumors formed by A549 cells) and 50% (the
group injected with NCI-H460 cells) on day 25 compared to mice receiving radiation alone.
Moreover, IR-induced expression of PAK1 and EMT-related proteins was significantly
reduced in the extracted tumor tissue lysates when TG101209 was administered in the mice
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

(Fig. 6C). Thus, we suggest that the JAK2 inhibitor significantly increased in vivo
radiosensitization while inhibiting the EMT.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Discussion
Many researchers have focused on developing potent radiosensitizers to overcome
radioresistance and to increase the therapeutic efficacy in NSCLC patients. Given the
inherent insensitivity to cytotoxic agents, identifying molecules that drive the growth,
survival and metastasis of lung cancer cells is critical for the development of novel
therapeutic reagents. In the same context with identification of the biomarkers, we found that
JAK2 and PAK1 are promising pharmacological targets for the radiosensitization of NSCLC
cells because Tyr phosphorylation of PAK1 by JAK2 was shown to prevent cell death and to
promote cell invasion. In the present study, PAK1 mRNA and protein expression was up-
regulated, and the protein localized to the nucleus in irradiated NSCLC cells. In addition to
nuclear localization, the stability of PAK1 protein increased through IR-dependent Tyr
phosphorylation of PAK1 by JAK2. We also found that activation of the JAK2/PAK1/Snail
pathway by IR was important for radioresistance primarily via EMT induction (Fig. 7). Based
on these findings, we suggest that JAK2 can be a potent target as an upstream factor for Tyr
phosphorylation of PAK1 associated with radioresistant signaling in NSCLC cells.
PAK1 has an autoregulatory region (including a p21-binding domain, dimerization
segment, and inhibitory switch domain) and several motifs for interaction with adaptor
proteins (Nck, Grb2, and PIX) at the N-terminus, and a catalytic domain at the C-terminus
(26). PAK1 activity is known to be inhibited by the formation of a resting homodimer
through interaction of the inhibitory switch domain (26). The inhibitory PAK1 homodimer
can be disrupted and rearranged by interaction with activated Cdc42/Rac1 (26, 36).
Monomeric PAK1 can then undergo autophosphorylation at several residues including S57,
S144, S149, S199/S203, and T423, leading to PAK1 activation (26, 36-38). In the present
investigation, we could only observe PAK1 phosphorylation on Tyr residues and not Ser/Thr
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

residues in response to irradiation (Fig. 1). According to our data and those from several
studies, we assumed that Tyr phosphorylation of PAK1 could provide additional properties
compared to Ser/Thr phosphorylation. First of all, PAK1 phosphorylation on three Tyr
residues, especially Y201, might sufficiently disrupt the inhibitory conformation similar to
phosphorylation at S199/S203. Additionally, maintenance of the PAK1 active conformation
could be achieved, at least partially, by phosphorylation at Y153 since this residue also
resides nearby the autophosphorylation sites, S144 and S149 (especially S144 is responsible
for maximal kinase activity of PAK1) (39). Phosphorylation at Y285 could also affect kinase
activity since this residue is present in the N-lobe of the C-terminal catalytic domain. A
previous structural study revealed that PAK1 could form an active conformation through an
interaction between K299 at the N-lobe, E315 at the �C-helix, and ATP, leading to exposure
to a substrate binding site and phosphorylation of Ser/Thr residues of substrates (40). Based
on the location of Y285 at the ATP binding site, we expect that phosphorylation of this
residue might help stabilize the interaction between K299 and an ATP molecule. Although it
should be confirmed by further structural studies, phosphorylation of PAK1 at three Tyr
residues might directly or indirectly help PAK1 to form and maintain its catalytically active
conformation, resulting in the phosphorylation of downstream substrates in response to
irradiation.
We observed that Tyr phosphorylation of PAK1 was mediated by JAK2, a Tyr kinase
(Fig. 2). Most substrates bind to Tyr kinases through their SH2 domains. According to
previous structural studies, PAK1 does not contain an SH2 domain but has a proline-rich
motif for binding to SH3 domains (26, 36, 37). This structural information implies that PAK1
could be phosphorylated on Tyr residues by JAK2 through interaction with SH2/SH3-
containing adaptor proteins that provide a bridge for facilitating spatial proximity between
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

JAK2 primarily in near membrane and PAK1 in the cytoplasm. PAK1 has several known
adaptor proteins including Nck, Grb2, and PIX (41, 42). These proteins enable membrane
localization of PAK1 to be activated by other kinases including JAK2, Cdc42, and Rac1 (41,
43). Based on previous studies, we believed that Nck might be involved in JAK2-mediated
PAK1 Tyr phosphorylation in response to irradiation (28, 41). Nck shuttles PAK1 from the
cytoplasm to the plasma membrane. After membrane localization, PAK1 would be
susceptible to binding to JAK2 and, subsequently activated PAK1 would dissociate from Nck.
PAK1 activated by JAK2 could be therefore released from the plasma membrane into the
cytoplasm and even translocate into the nuclear. In this study, we observed that activated
PAK1 affected Snail-dependent cell migration in the nucleus (Fig. 4 and 5).
In addition to activating PAK1, we showed that Tyr phosphorylation increased PAK1
stability as well (Fig. 3). To understand how PAK1 stabilization is mediated by Tyr
phosphorylation, it was important to determine which E3 ligase is responsible for PAK1
degradation. Although a PAK1-specific E3 ligase has not been identified, Cdc42 homologous
protein (Chp) is a Rho-family GTPase that mediates ubiquitination-dependent degradation of
PAK1 (44). A previous report indicated that kinase activity of PAK1 is critical for PAK1
degradation (44). According to this study, Chp could mediate PAK1 degradation only when
PAK1 was activated by autophosphorylation on several Ser/Thr residues including S57, S144,
S149, S199/S203, and T423. Alanine substitutions at these sites led to increased PAK1
stability. Thus, we believe that Y201 phosphorylation of PAK1 might have structurally
similar property of phosphorylated moiety as phosphorylation of S199/S203, expecting
PAK1 destabilization. However, PAK1 was stabilized after being Tyr-phosphorylated as
shown in Fig. 3. One possible explanation is that Chp might not recognize Tyr-
phosphorylated PAK1 due to different autophosphorylation patterns. Thus, JAK2-induced
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Tyr-phosphorylation, especially at Y201, increased PAK1 activity and stability instead of
promoting PAK1 degradation. Our data are supported by findings from other investigations
demonstrating that protein degradation could be inhibited by phosphorylation at Tyr residues
(45, 46). For example, ubiquitination and degradation were more rapid for a mutant form of
c-Kit (Y823F) than in c-Kit WT (45). In this case, mutant c-Kit-mediated downstream
pathways involving Akt and Erk were negatively affected, leading to the suppression of cell
survival and proliferation. However, the Y823F mutation has no effect on kinase activity of
c-Kit. Another study showed that proliferating cell nuclear antigen (PCNA) was stabilized by
phosphorylation on Y211, leading to increased functional activity on DNA replication (46).
We showed that PAK1 without Tyr phosphorylation is susceptible to proteasome-dependent
degradation in Fig. 3. Similar to c-Kit and PCNA, phosphorylation on Tyr residues is
responsible for maintaining PAK1 stability and activity, leading to cell proliferation and
invasion induced by Snail phosphorylation.
Several investigations conducted by Diakonova et al. have suggested that Tyr-
phosphorylated PAK1 could be crucially involved in the regulation of cancer cell migration
(14, 28, 47). Based on the mechanisms underlying EMT regulation, we focused on a nuclear
factor as a downstream target of PAK1 that could confer radioresistant properties on NSCLC
cells. We found that Snail interacted with PAK1 in the nucleus according to our interactome
analysis in Fig. 4. Although this interaction was observed in the cytoplasm as well, it is
unclear whether PAK1 and Snail translocated into the nucleus together or individually.
Nevertheless, the interaction between PAK1 and Snail could be responsible for EMT
induction through the suppression of E-cadherin expression in irradiated NSCLC cells. This
finding could be supported by a previous study demonstrating that Snail phosphorylation at
S246 by PAK1 led to nuclear accumulation of Snail and promotes cell invasiveness (30).
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Additionally, constitutively active PAK1 (T423E mutant) enhanced Snail activation
compared to wild-type PAK1. T423 of PAK1 is located in the catalytic domain necessary for
kinase activity. As we mentioned above, phosphorylation of PAK1 on three Tyr residues
might have similar effects on kinase activity due to exposure of the catalytic domain through
phosphorylation of residues in the inhibitory switch domain. In this study, even when
phosphorylation at Ser/Thr residues (such as T423) of PAK1 was not detected, PAK1 could
interact with Snail in response to irradiation, consequently leading to increasing cell motility
through reduced E-cadherin expression.
In addition to regulating cell motility, Snail also promotes cell survival and
proliferation. Snail regulates the expression of several genes mainly involved in cell invasion
and migration including matrix metalloproteinase (MMP) family members (MMP1, MMP2,
and MMP7) and cell-cell or cell-matrix interacting proteins (E-cadherin, integrin �6, and
tissue inhibitor of metalloproteinase 3) (48). Furthermore, this factor is responsible for
suppressing the expression of other genes including caspase-8 and tumor suppressor CYLD
(49). It has been reported that CYLD could be repressed in a Snail-dependent manner
resulting in increased expression of cyclin D1 and N-cadherin, thus enhancing the
proliferation and migration of melanoma cells (49). These findings could explain, at least in
part, the hyper-growth of cells (especially NCI-H460 cells at 48 h) treated with irradiation
alone compared to that of cells treated with irradiation and each drug observed in the present
study (Fig. 5C). Taken together, our findings and data from the literature indicate that PAK1-
activated Snail might directly or indirectly confer radioresistance on NSCLC cells by
inducing EMT and cell proliferation.
The exact molecular mechanism underlying radioresistance in NSCLC has been still
largely elusive. In the present study, we demonstrated that Tyr phosphorylation of PAK1
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

could be a critical factor for conferring radioresistance. We have provided the first evidence
of a novel regulatory mechanism of radioresistance including the functional involvement of
JAK2, PAK1, and Snail in irradiated NSCLC cells (Fig. 7). We also propose that JAK2
inhibitors might be potential radiosensitizers that suppress Tyr phosphorylation of PAK1. In
this study, our results demonstrated that regulation of Tyr phosphorylation of PAK1 with
pharmacological agents in combination with radiotherapy could overcome radioresistance
and eventually enhance the efficiency of radiotherapy for treating NSCLC.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Authors’ Contributions
Conception and design: E. Kim, B. Youn
Development of methodology: E. Kim, H. Youn, W. Kim, B. Youn
Acquisition of data (provided animals, acquired and managed patients, provided facilities,
etc.): E. Kim, H. Youn, T. Kwon, B. Son, J. Kang, H.J. Yang, K.M. Seong
Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computational
analysis): E. Kim, H. Youn, H.J. Yang, K.M. Seong, W. Kim, B. Youn
Writing, review, and/or revision of the manuscript: E. Kim, W. Kim, B. Youn
Administrative, technical, or material support (i.e., reporting or organizing data,
constructing databases): W. Kim, B. Youn
Study supervision: B. Youn
Acknowledgements
The authors thank Dr. Maria Diakonova (University of Toledo, Toledo, OH) for
providing the mutant PAK1 constructs.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

References
1. Koh PK, Faivre-Finn C, Blackhall FH, De Ruysscher D. Targeted agents in non-small cell lung cancer (NSCLC): clinical developments and rationale for the combination with thoracic radiotherapy. Cancer Treat Rev 2012;38:626-40.
2. Bussink J, van der Kogel AJ, Kaanders JH. Activation of the PI3-K/AKT pathway and implications for radioresistance mechanisms in head and neck cancer. Lancet Oncol 2008;9:288-96.
3. Nishimura Y, Nakagawa K, Takeda K, Tanaka M, Segawa Y, Tsujino K, et al. Phase I/II trial of sequential chemoradiotherapy using a novel hypoxic cell radiosensitizer, doranidazole (PR-350), in patients with locally advanced non-small-cell lung Cancer (WJTOG-0002). Int J Radiat Oncol Biol Phys 2007;69:786-92.
4. Yang HJ, Youn H, Seong KM, Jin YW, Kim J, Youn B. Phosphorylation of ribosomal protein S3 and antiapoptotic TRAF2 protein mediates radioresistance in non-small cell lung cancer cells. J Biol Chem 2013;288:2965-75.
5. Hillman GG, Singh-Gupta V, Runyan L, Yunker CK, Rakowski JT, Sarkar FH, et al. Soy isoflavones radiosensitize lung cancer while mitigating normal tissue injury. Radiother Oncol 2011;101:329-36.
6. Kim W, Youn H, Seong KM, Yang HJ, Yun YJ, Kwon T, et al. PIM1-activated PRAS40 regulates radioresistance in non-small cell lung cancer cells through interplay with FOXO3a, 14-3-3 and protein phosphatases. Radiat Res 2011;176:539-52.
7. Kim W, Youn H, Kwon T, Kang J, Kim E, Son B, et al. PIM1 kinase inhibitors induce radiosensitization in non-small cell lung cancer cells. Pharmacol Res 2013;70:90-101.
8. Yang HJ, Kim N, Seong KM, Youn H, Youn B. Investigation of radiation-induced transcriptome profile of radioresistant non-small cell lung cancer A549 cells using RNA-seq. PloS one 2013;8:e59319.
9. Molli PR, Li DQ, Murray BW, Rayala SK, Kumar R. PAK signaling in oncogenesis. Oncogene 2009;28:2545-55.
10. Vadlamudi RK, Adam L, Wang RA, Mandal M, Nguyen D, Sahin A, et al. Regulatable expression of p21-activated kinase-1 promotes anchorage-independent growth and abnormal organization of mitotic spindles in human epithelial breast cancer cells. J Biol Chem 2000;275:36238-44.
11. Wang RA, Zhang H, Balasenthil S, Medina D, Kumar R. PAK1 hyperactivation is sufficient for mammary gland tumor formation. Oncogene 2006;25:2931-6.
12. Menard RE, Jovanovski AP, Mattingly RR. Active p21-activated kinase 1 rescues MCF10A breast epithelial cells from undergoing anoikis. Neoplasia (New York, NY) 2005;7:638-45.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

13. Ong CC, Jubb AM, Haverty PM, Zhou W, Tran V, Truong T, et al. Targeting p21-activated kinase 1 (PAK1) to induce apoptosis of tumor cells. Proceedings of the National Academy of Sciences of the United States of America 2011;108:7177-82.
14. Rider L, Shatrova A, Feener EP, Webb L, Diakonova M. JAK2 tyrosine kinase phosphorylates PAK1 and regulates PAK1 activity and functions. J Biol Chem 2007;282:30985-96.
15. Bagheri-Yarmand R, Mandal M, Taludker AH, Wang RA, Vadlamudi RK, Kung HJ, et al. Etk/Bmx tyrosine kinase activates Pak1 and regulates tumorigenicity of breast cancer cells. J Biol Chem 2001;276:29403-9.
16. McManus MJ, Boerner JL, Danielsen AJ, Wang Z, Matsumura F, Maihle NJ. An oncogenic epidermal growth factor receptor signals via a p21-activated kinase-caldesmon-myosin phosphotyrosine complex. J Biol Chem 2000;275:35328-34.
17. Doucette TA, Kong LY, Yang Y, Ferguson SD, Yang J, Wei J, et al. Signal transducer and activator of transcription 3 promotes angiogenesis and drives malignant progression in glioma. Neuro-oncology 2012;14:1136-45.
18. Sansone P, Bromberg J. Targeting the interleukin-6/Jak/stat pathway in human malignancies. Journal of clinical oncology : official journal of the American Society of Clinical Oncology 2012;30:1005-14.
19. Govindan R, Ding L, Griffith M, Subramanian J, Dees ND, Kanchi KL, et al. Genomic landscape of non-small cell lung cancer in smokers and never-smokers. Cell 2012;150:1121-34.
20. Yin ZJ, Jin FG, Liu TG, Fu EQ, Xie YH, Sun RL. Overexpression of STAT3 potentiates growth, survival, and radioresistance of non-small-cell lung cancer (NSCLC) cells. The Journal of surgical research 2011;171:675-83.
21. Goodman PA, Niehoff LB, Uckun FM. Role of tyrosine kinases in induction of the c-jun proto-oncogene in irradiated B-lineage lymphoid cells. J Biol Chem 1998;273:17742-8.
22. Sun Y, Moretti L, Giacalone NJ, Schleicher S, Speirs CK, Carbone DP, et al. Inhibition of JAK2 signaling by TG101209 enhances radiotherapy in lung cancer models. Journal of thoracic oncology : official publication of the International Association for the Study of Lung Cancer 2011;6:699-706.
23. Yang HJ, Youn H, Seong KM, Yun YJ, Kim W, Kim YH, et al. Psoralidin, a dual inhibitor of COX-2 and 5-LOX, regulates ionizing radiation (IR)-induced pulmonary inflammation. Biochem Pharmacol 2011;82:524-34.
24. Jirawatnotai S, Hu Y, Michowski W, Elias JE, Becks L, Bienvenu F, et al. A function for cyclin D1 in DNA repair uncovered by protein interactome analyses in human cancers. Nature 2011;474:230-4.
25. Muthuswamy SK, Li D, Lelievre S, Bissell MJ, Brugge JS. ErbB2, but not ErbB1,
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

reinitiates proliferation and induces luminal repopulation in epithelial acini. Nat Cell Biol 2001;3:785-92.
26. Lei M, Lu W, Meng W, Parrini MC, Eck MJ, Mayer BJ, et al. Structure of PAK1 in an autoinhibited conformation reveals a multistage activation switch. Cell 2000;102:387-97.
27. Singh RR, Song C, Yang Z, Kumar R. Nuclear localization and chromatin targets of p21-activated kinase 1. J Biol Chem 2005;280:18130-7.
28. Tao J, Oladimeji P, Rider L, Diakonova M. PAK1-Nck regulates cyclin D1 promoter activity in response to prolactin. Molecular endocrinology (Baltimore, Md) 2011;25:1565-78.
29. Nakatani Y, Ogryzko V. Immunoaffinity purification of mammalian protein complexes. Methods in enzymology 2003;370:430-44.
30. Yang Z, Rayala S, Nguyen D, Vadlamudi RK, Chen S, Kumar R. Pak1 phosphorylation of snail, a master regulator of epithelial-to-mesenchyme transition, modulates snail's subcellular localization and functions. Cancer research 2005;65:3179-84.
31. Ikenouchi J, Matsuda M, Furuse M, Tsukita S. Regulation of tight junctions during the epithelium-mesenchyme transition: direct repression of the gene expression of claudins/occludin by Snail. Journal of cell science 2003;116:1959-67.
32. Kudo-Saito C, Shirako H, Takeuchi T, Kawakami Y. Cancer metastasis is accelerated through immunosuppression during Snail-induced EMT of cancer cells. Cancer cell 2009;15:195-206.
33. Debnath J, Brugge JS. Modelling glandular epithelial cancers in three-dimensional cultures. Nature reviews Cancer 2005;5:675-88.
34. Henry LA, Johnson DA, Sarrio D, Lee S, Quinlan PR, Crook T, et al. Endoglin expression in breast tumor cells suppresses invasion and metastasis and correlates with improved clinical outcome. Oncogene 2011;30:1046-58.
35. Kang J, Kim E, Kim W, Seong KM, Youn H, Kim JW, et al. Rhamnetin and Cirsiliol Induce Radiosensitization and Inhibition of Epithelial-Mesenchymal Transition (EMT) by miR-34a-mediated Suppression of Notch-1 Expression in Non-small Cell Lung Cancer Cell Lines. J Biol Chem 2013;288:27343-57.
36. Lei M, Robinson MA, Harrison SC. The active conformation of the PAK1 kinase domain. Structure 2005;13:769-78.
37. Wang J, Wu JW, Wang ZX. Structural insights into the autoactivation mechanism of p21-activated protein kinase. Structure 2011;19:1752-61.
38. Kichina JV, Goc A, Al-Husein B, Somanath PR, Kandel ES. PAK1 as a therapeutic target. Expert Opin Ther Targets 2010;14:703-25.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

39. Chong C, Tan L, Lim L, Manser E. The mechanism of PAK activation. Autophosphorylation events in both regulatory and kinase domains control activity. J Biol Chem 2001;276:17347-53.
40. Ng YW, Raghunathan D, Chan PM, Baskaran Y, Smith DJ, Lee CH, et al. Why an A-loop phospho-mimetic fails to activate PAK1: understanding an inaccessible kinase state by molecular dynamics simulations. Structure 2010;18:879-90.
41. Zhao ZS, Manser E, Lim L. Interaction between PAK and nck: a template for Nck targets and role of PAK autophosphorylation. Mol Cell Biol 2000;20:3906-17.
42. Puto LA, Pestonjamasp K, King CC, Bokoch GM. p21-activated kinase 1 (PAK1) interacts with the Grb2 adapter protein to couple to growth factor signaling. J Biol Chem 2003;278:9388-93.
43. Lu W, Mayer BJ. Mechanism of activation of Pak1 kinase by membrane localization. Oncogene 1999;18:797-806.
44. Weisz Hubsman M, Volinsky N, Manser E, Yablonski D, Aronheim A. Autophosphorylation-dependent degradation of Pak1, triggered by the Rho-family GTPase, Chp. Biochem J 2007;404:487-97.
45. Agarwal S, Kazi JU, Ronnstrand L. Phosphorylation of the activation loop tyrosine 823 in c-Kit is crucial for cell survival and proliferation. J Biol Chem 2013;288:22460-8.
46. Wang SC, Nakajima Y, Yu YL, Xia W, Chen CT, Yang CC, et al. Tyrosine phosphorylation controls PCNA function through protein stability. Nat Cell Biol 2006;8:1359-68.
47. Hammer A, Rider L, Oladimeji P, Cook L, Li Q, Mattingly RR, et al. Tyrosyl phosphorylated PAK1 regulates breast cancer cell motility in response to prolactin through filamin A. Molecular endocrinology (Baltimore, Md) 2013;27:455-65.
48. Jin H, Yu Y, Zhang T, Zhou X, Zhou J, Jia L, et al. Snail is critical for tumor growth and metastasis of ovarian carcinoma. Int J Cancer 2010;126:2102-11.
49. Massoumi R, Kuphal S, Hellerbrand C, Haas B, Wild P, Spruss T, et al. Down-regulation of CYLD expression by Snail promotes tumor progression in malignant melanoma. J Exp Med 2009;206:221-32.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Figure legends
Figure 1. IR induces overexpression, Tyr phosphorylation, and nuclear localization of
PAK1 in NSCLC cells. A, IR-induced PAK1 mRNA expression was detected by Northern
blot analysis. After irradiation (0, 2, 4, or 6 Gy), the cells were harvested and total RNA was
extracted. Total RNA was analyzed using an [�-32P]-labeled PAK1 cDNA probe. GAPDH was
used for normalization. B, IR-induced PAK1 protein expression was detected by Western blot
analysis. C, IR-induced PAK1 phosphorylation was verified by an in vivo kinase assay. After
irradiation (4 Gy) for 0, 1, 2, or 3 h, the cells were harvested and cell lysates were subjected
to an IP assay with an anti-PAK1 antibody followed by Western blot analysis for pSer/Thr
and pTyr. D, IR-induced PAK1 phosphorylation at three Tyr residues (Y153, Y201, and Y285)
was confirmed by an in vivo kinase assay using PAK1 Y153F, Y201F, Y285F, and Y3F
mutants (WCL, whole cell lysates). Cells were harvested after irradiation (4 Gy) for 2 h and
cell lysates were immunoprecipitated with an anti-HA antibody and analyzed by Western blot
analysis for pTyr. E, IR-induced translocation of PAK1 from the cytoplasm into the nucleus
was assayed by Western blot analysis after cytoplasmic or nuclear fractionation (CE,
cytoplasmic extract; NE, nuclear extract). Tubulin and Lamin A/C were used as markers for
the CE and NE, respectively. F, Involvement of IR-activated nuclear localization and
phosphorylation of PAK1 was determined by an IP assay and Western blot analysis. After
irradiation, the cells transiently transfected with PAK1 (WT or Y3F mutant) were harvested
to conduct cytoplasmic or nuclear fractionation. Each extract was immunoprecipitated with
an anti-HA antibody and detected by Western blot analysis using an anti-pTyr antibody.
Figure 2. JAK2 phosphorylates PAK1 in irradiated NSCLC cells. A, Tyr phosphorylation
of PAK1 by JAK2 was measured by an in vivo kinase assay using each Tyr kinase-specific
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

siRNA molecule (EGFR, c-Abl, JAK1, JAK2, and JAK3). The inset shows that siRNA
oligonucleotides specific for Tyr kinase significantly reduced Tyr kinase expression. B, In
vivo binding of IR-activated PAK1 with JAK2 was measured with an IP assay. After
irradiation for 2 h, cell lysates were immunoprecipitated with an anti-FLAG antibody and
subjected to Western blot analysis using an anti-PAK1 antibody. C, Phosphorylation of PAK1
by JAK2 was measured with an in vivo kinase assay with a JAK2 KD mutant. After
irradiation for 2 h, cells transfected with JAK2 WT or KD were harvested and
immunoprecipitated with an anti-PAK1 antibody followed by Western blot analysis using an
anti-pTyr antibody. D, JAK2-mediated PAK1 phosphorylation at Y153, Y201, and Y285 was
confirmed by an in vivo kinase assay with a PAK1 Y3F mutant. Cells were harvested after
irradiation (4 Gy) for 2 h. Immunoprecipitant of HA (PAK1) was subjected to Western blot
analysis using an anti-pTyr antibody. E, JAK2-mediated PAK1 phosphorylation was
reconfirmed using JAK2-specific inhibitors (TG101209 or TG101348), an EGFR inhibitor
(AG1478), and a c-Abl inhibitor (STI-571). After treatment of each inhibitor and irradiation
for 2 hr, cells were harvested and were immunoprecipitated with an anti-PAK1 antibody
followed by Western blot analysis using an anti-pTyr antibody.
Figure 3. Tyr phosphorylation is required for maintaining PAK1 protein stability in
irradiated NSCLC cells. A, Effects of Tyr phosphorylation on the protein stability of PAK1
were measured by Western blot analysis. Ectopic expression of PAK1 WT or the Y3F mutant
in NSCLC cells with or without IR (4 Gy; 6 h incubation) and PI (a mix of 5 μM MG115 and
5 μM MG262, 24 h incubation). B, The half-life of IR-activated PAK1 WT or Y3F mutant
with cycloheximide (CHX; 50 μM) in the presence of PI was estimated by Western blot
analysis. Cells were treated with PI at different time points after irradiation (0, 4, 8, 12, or 16
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

h). Cells were harvested and cell lysates were detected by Western blot analysis using an anti-
HA antibody. The level of HA-PAK1 WT or Y3F mutant protein at each time point measured
in Fig. 3B was plotted by densitometric analysis. The level of HA-PAK1 WT or Y3F mutant
expression at time point 0 (lane 3) was set at 100%. C, The accumulation of poly-
ubiquitinated IR-activated PAK1 Y3F mutant was measured. NSCLC cells expressing PAK1
WT or Y3F mutant were treated with or without PI as described in Fig. 3A. HA-PAK1 WT or
Y3F mutant protein was then immunoprecipitated with an anti-HA antibody and Western blot
analysis was performed with an anti-ubiquitin antibody. D, The direct relationship between
JAK2-dependent phosphorylation of PAK1 and poly-ubiquitination with irradiation in
NSCLC was confirmed by the same experiment described in Fig. 3C with a JAK2-specific
inhibitor, TG101209.
Figure 4. PAK1 interacts with Snail in the nucleus in irradiated NSCLC cells. A,
Schematic presentation of the strategy used to identify the PAK1 nuclear interactome in A549
cells. B, Purified PAK1 complexes in non-irradiated or irradiated A549 cells were separated
by PAGE and visualized by silver staining. The silver stained gel was analyzed by mass
spectrometry (peptide mass fingerprint). It was shown that three bands especially increased
by irradiation. The bands indicated by an arrowhead (a, b, and c) correspond to Snail, ER�,
and FKHR, respectively. C, In vivo binding of IR-activated PAK1 with Snail in the nucleus of
NSCLC cells was confirmed by an IP assay. After irradiation, the cells transiently transfected
with PAK1 WT or Y3F mutant were harvested to conduct cytoplasmic or nuclear
fractionation. Each extract was subjected to an IP assay with an anti-PAK1 antibody followed
by Western blot analysis for Snail. D, Interaction of endogenous PAK1 with endogenous
Snail was measured by an IP assay. After irradiation, the cells were harvested to conduct
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

cytoplasmic or nuclear fractionation. Each extract was subject to an IP assay with an anti-
PAK1 antibody followed by Western blot analysis using an anti-Snail antibody. E, PAK1-
mediated Snail phosphorylation at S246 was confirmed by an in vivo kinase assay with a
Snail S246A mutant. The cells transiently transfected with Snail (WT or S246A mutant) were
harvested to conduct cytoplasmic or nuclear fractionation. Each extract was subjected to an IP
assay with an anti-FLAG antibody (Snail) followed by Western blot analysis using an anti-
pSer/Thr antibody. F, Effect of PAK1 on the ability of Snail to repress E-cadherin (or
Occludin) promoter activity was assessed with a luciferase assay. Error bars represent ± SD
(n = 3); ∗p < 0.05 compared with E-cad (or Occlu)-Luc plasmid-transfected non-irradiated
cells, ∗∗p < 0.05 compared with E-cad (or Occlu)-Luc plasmid-transfected irradiated cells
(two-way ANOVA, Bonferroni post test).
Figure 5. Inhibition of PAK1 Tyr phosphorylation reduces IR-dependent EMT in
NSCLC cells. A, Morphological changes of NSCLC cells treated with a JAK2 inhibitor were
monitored with a 3D culture model. NSCLC cells treated with or without IR and TG101209
were grown in 3D cultures and fixed after 20 h (A549) or 28 h (NCI-H460). The cells were
permeabilized and stained for tubulin (green) and with DAPI (blue). B, The inhibitory effects
of a JAK2 inhibitor on IR-induced migration of NSCLC cells were measured with a
Transwell migration assay. The results are expressed as fold-increase of migration compared
to the control group and are based on the relative number of cells in a randomly selected field
from three representative experiments. Rhamnetin and cirsiliol were used as positive EMT
inhibitor controls. C, The inhibitory effects of a JAK2 inhibitor on IR-induced migration of
NSCLC cells were measured with a wound healing assay. After the treatment with each
inhibitor for 4 h and irradiation, cells were wounded and incubated for 18 h and 30 h (A549)
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

or 24 h and 48 h (NCI-H460). D, Effects of a JAK2 inhibitor on mRNA expression of E-
cadherin, Vimentin, and Fibronectin in irradiated NSCLC cells were analyzed by qRT-PCR.
E, Effects of a JAK2 inhibitor on the protein expression of E-cadherin, Vimentin, and
Fibronectin in irradiated NSCLC cells were analyzed by Western blot analysis. Error bars
represent ± SD (n = 3); *p < 0.05 compared with non-irradiated cells, **p < 0.05 compared
with irradiated cells (one-way ANOVA, Tukey's honestly significant difference test, or two-
way ANOVA, Bonferron post test).
Figure 6. JAK2-inhibitor acts as an in vivo radiosensitizer and decreases in vivo EMT in
a xenograft mouse model. A, An experimental protocol to determine whether JAK2-
inhibitor acts as an in vivo radiosensitizer and decreases in vivo EMT in a xenograft mouse
model. B, The effect of a JAK2 inhibitor on in vivo radiosensitization was measured in a
xenograft mouse model. C, The in vivo effects of a JAK2 inhibitor on the expression of PAK1
and EMT-related proteins were evaluated by Western blot analysis. Error bars represent ±
SEM (n = 3 with four animals per group); *p < 0.05 compared with tumor volume on day 25
in mice treated with radiation alone (one-way ANOVA, Tukey's honestly significant
difference test).
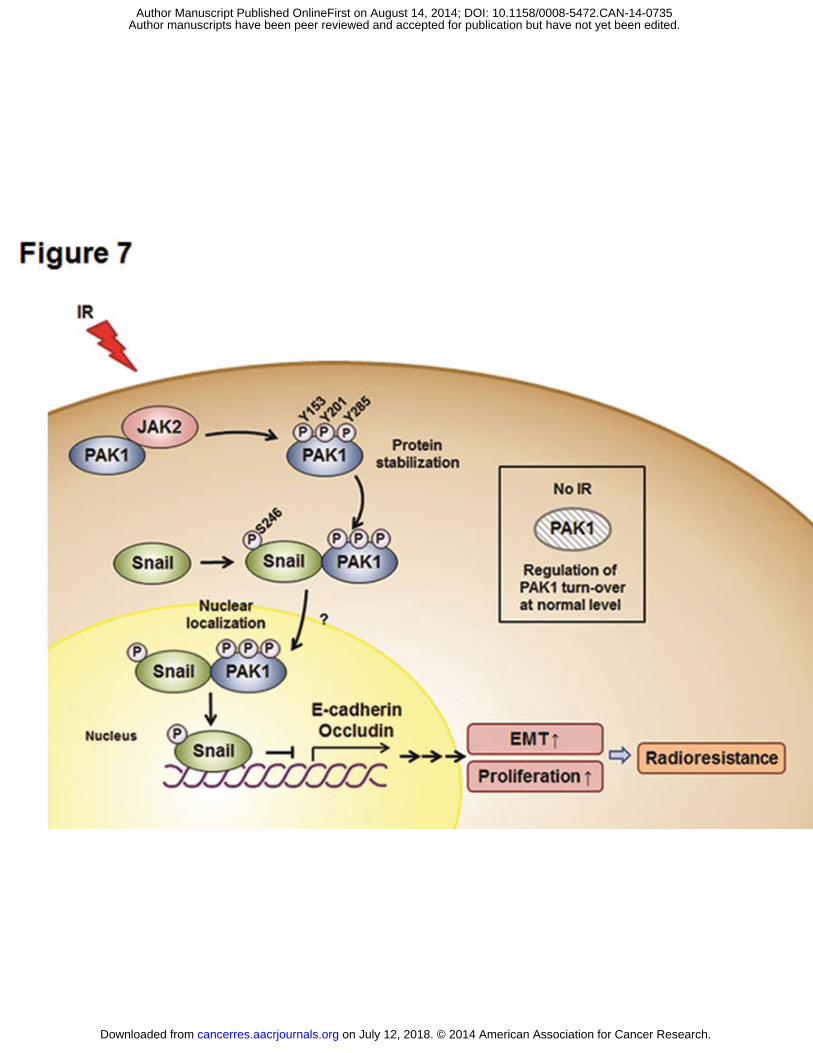
Figure 7. A schematic diagram illustrates JAK2-pPAK1-pSnail signaling in response to
irradiation and the radiosensitizing effects of a JAK2 inhibitor in NSCLC cells. PAK1 is
up-regulated and phosphorylated on Tyr residues by JAK2, consequently leading to
translocation into the nucleus after radiation exposure. IR-induced Tyr phosphorylation of
PAK1 is required to maintain PAK1 protein stability in NSCLC cells. After phosphorylation
on Tyr residues, PAK1 binds with Snail, thus promoting the transcriptional repression
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

activity of Snail by Ser phosphorylation. Additionally, activation of the JAK2-pPAK1-pSnail
signaling axis by irradiation promotes the EMT and cell proliferation. Treatment with a JAK2
inhibitor enhances EMT inhibition and radiosensitization in vitro and in vivo.
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735

Published OnlineFirst August 14, 2014.Cancer Res EunGi Kim, HyeSook Youn, TaeWoo Kwon, et al. cancer cellsepithelial-mesenchymal transition and radioresistance in lung PAK1 tyrosine phosphorylation is required to induce
Updated version
10.1158/0008-5472.CAN-14-0735doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2014/08/16/0008-5472.CAN-14-0735.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/early/2014/08/14/0008-5472.CAN-14-0735To request permission to re-use all or part of this article, use this link
on July 12, 2018. © 2014 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 14, 2014; DOI: 10.1158/0008-5472.CAN-14-0735























![codin9cafe[2015.05.13]안드로이드 개발하기 3rd week - 김태우(Kim Taewoo)](https://img.pdfslide.net/doc/110x75/58ed739c1a28ab29078b4675/codin9cafe20150513-3rd-week-kim.jpg)





