Embed Size (px)
Citation preview

complex disrupts dynein-dependent maintenanceof membrane organelle distribution. J. Cell Biol.139, 469–484
23 Skop, A.R. and White, J.G. (1998) The dynactincomplex is required for cleavage plane specificationin early Caenorhabditis elegans embryos. Curr. Biol.8, 1110–1116
24 Adames, N.R. and Cooper, J.A. (2000) Microtubuleinteractions with the cell cortex causing nuclearmovements in Saccharomyces cerevisiae. J. CellBiol. 149, 863–874
25 Carminati, J.L. and Stearns, T. (1997) Microtubulesorient the mitotic spindle in yeast through dynein-dependent interactions with the cell cortex. J. CellBiol. 138, 629–641
26 Beckwith, S.M. et al. (1998) The ‘8-kD’ cytoplasmicdynein light chain is required for nuclear migration
and for dynein heavy chain localization inAspergillus nidulans. J. Cell Biol. 143, 1239–1247
27 Morris, N.R. (2000) Nuclear migration. From fungi tothe mammalian brain. J. Cell Biol.148, 1097–1101
28 Vaughan, K.T. etal. (1999) Co-localization ofdynactin and cytoplasmic dynein with CLIP-170 atmicrotubule distal ends. J. Cell Sci. 112, 1437–1447
29 Sawin, K.E. (2000) Microtubule dynamics: theview from the tip. Curr. Biol. 10, R860–R862
30 Rieder, C.L. and Alexander, S.P. (1990) Kinetochoresare transported poleward along a single astralmicrotubule during chromosome attachment to thespindle in newt lung cells. J.Cell Biol.110, 81–95
31 Morris, S.M. etal. (1998) The lissencephaly geneproduct Lis1, a protein involved in neuronalmigration, interacts with a nuclear movementprotein, NudC. Curr. Biol.8,603–606
32 Efimov, V.P. and Morris, N.R. (2000) The LIS1-related NUDF protein of Aspergillus nidulansinteracts with the coiled-coil domain of theNUDE/RO11 protein. J. Cell Biol. 150, 681–688
33 Kitagawa, M. etal. (2000) Direct association of LIS1,the lissencephaly gene product, with a mammalianhomologue of a fungal nuclear distribution protein,rNUDE. FEBS Lett.479,57–62
34 Sasaki, S. et al. (2000) A LIS1/NUDEL/cytoplasmicdynein heavy chain complex in the developing andadult nervous system. Neuron 28, 681–696
35 Niethammer, M. et al. (2000) NUDEL is a novelCdk5 substrate that associates with LIS1 andcytoplasmic dynein. Neuron 28, 697–711
36 Feng, Y. et al. (2000) LIS1 regulates CNSlamination by interacting with mNudE, a centralcomponent of the centrosome. Neuron 28, 665–679
TRENDS in Cell Biology Vol.11 No.4 April 2001
http://tcb.trends.com 0962-8924/01/$ – see front matter © 2001 Elsevier Science Ltd. All rights reserved. PII: S0962-8924(01)01954-7
160 Opinion
The orientation of the mitotic spindle has a profoundimpact on correct chromosomal segregation andspecification of the division plane in metazoan cells.Additionally, regulated orientation of the spindle candictate asymmetric cell divisions, a strategy exploitedthroughout development to generate cell-type diversity1.The budding yeast Saccharomyces cerevisiaedividesasymmetrically along a polarity axis defined by the site
of bud emergence, generating a larger mother and asmaller daughter cell. Orientation of the intranuclearspindle along this polarity axis prior to anaphase iscrucial for accurate chromosomal segregation betweenmother and daughter cells. Thus, S. cerevisiaeserves asa genetically tractable model to understand how spindleorientation is attained.
Establishment of spindle polarity stems from events that enforce daughter-bound fate to one spindlepole and mother-bound fate to the other pole duringspindle morphogenesis. This entails integrating cell-cycle-regulated processes with cytoskeletal structures(actin and microtubules) that impart correct identity to each spindle pole and accordingly dictate spindlealignment along the cell polarity axis.
Control of spindle orientation is therefore complex and operates at several different levels: microtubule-organizing center (MTOC) function, spatial cues from the cell cortex, motors and additional factors regulatingmicrotubule dynamic instability. Correct orchestration of these various levels of control is absolutely crucial notonly for imparting correct polarity and orientation to themitotic spindle but also to ensure proper spindle dynamicsand, in turn, timely progression of the cell cycle.
Here, we review recent evidence and propose amodel for coordinated spindle assembly, polarity andorientation in budding yeast. For the purpose of thisarticle, spindle polarity denotes the commitment toan asymmetric fate for each spindle pole (mother-versus daughter-bound), whereas spindle orientationrepresents the ensuing positioning and alignment ofthe spindle at the bud neck prior to anaphase onset.
Spindle assembly and orientation in S. cerevisiae
Spindle morphogenesis in S. cerevisiae is initiated by the execution of START at the G1–S transition of the cell cycle2,3. Progression through START triggers budemergence, DNA replication and the duplication of theyeast MTOC – the spindle pole body (SPB; Fig.1).Because budding yeast cells undergo a closed mitosis(i.e. without nuclear envelope breakdown), the SPBsremain embedded in the nuclear envelope throughoutthe cell cycle and organize cytoplasmic or astral
Control of spindle
polarity and orientation
in SaccharomycescerevisiaeMarisa Segal and Kerry Bloom
Control of mitotic spindle orientation represents a major strategy for the
generation of cell diversity during development of metazoans.Studies in the
budding yeast Saccharomyces cerevisiae have contributed towards our present
understanding of the general principles underlying the regulation of spindle
positioning in an asymmetrically dividing cell. In S.cerevisiae, the mitotic spindle
must orient along the cell polarity axis,defined by the site of bud emergence, to
ensure correct nuclear division between the mother and daughter cells.
Establishment of spindle polarity dictates this process and relies on the concerted
control of spindle pole function and a precise program of cues originating from the
cell cortex that directs cytoplasmic microtubule attachments during spindle
morphogenesis.These cues cross talk with the machinery responsible for bud-site
selection, indicating that orientation of the spindle in yeast cells is mechanistically
coupled to the definition of a polarity axis and the division plane.Here,we propose
a model integrating the inherently asymmetric properties of the spindle pathway
with the program of positional information contributing towards orienting the
spindle in budding yeast.Because the basic machinery orienting the spindle in
higher-eukaryotic cells appears to be conserved, it might be expected that similar
principles govern centrosome asymmetry in the course of metazoan development.

microtubules from the cytosolic face of the SPB, theouter plaque, as well as intranuclear microtubules fromthe nuclear face, the inner plaque. Towards completionof DNA replication and SPB separation, spindleassembly and nuclear migration to the bud neck ensue.Prior to chromosome segregation and the onset ofanaphase, orientation of the spindle along themother–bud axis is a direct consequence of astralmicrotubule-mediated interactions established betweenone pole (SPBdaughter) with the bud cell cortex and thesecond pole (SPBmother) with the mother cell cortex4–6.
Spindle morphogenesis is subjected to cell-cyclecontrol. In S. cerevisiae, a single cyclin-dependent kinase– Cdc28 (also known as Cdk1) – regulates cell-cycleprogression by associating with different stage-specificcyclins7. Passage through START requires Cdc28activation by the G1 cyclins Cln1, Cln2 and Cln3.Accordingly, SPB duplication depends on Cln-dependentkinase activity. Progression through S, G2 and M phasesis driven by Cdc28 activation by the B-type cyclinsClb1–Clb6. Strains containing multiple CLBdeletionsfail to form a bipolar spindle8,9, implicating Clb functionin spindle assembly. The contribution of individual Clbsto various aspects of spindle morphogenesis is beginningto emerge and will be discussed below.
Cytoplasmic microtubule behavior during
spindle assembly
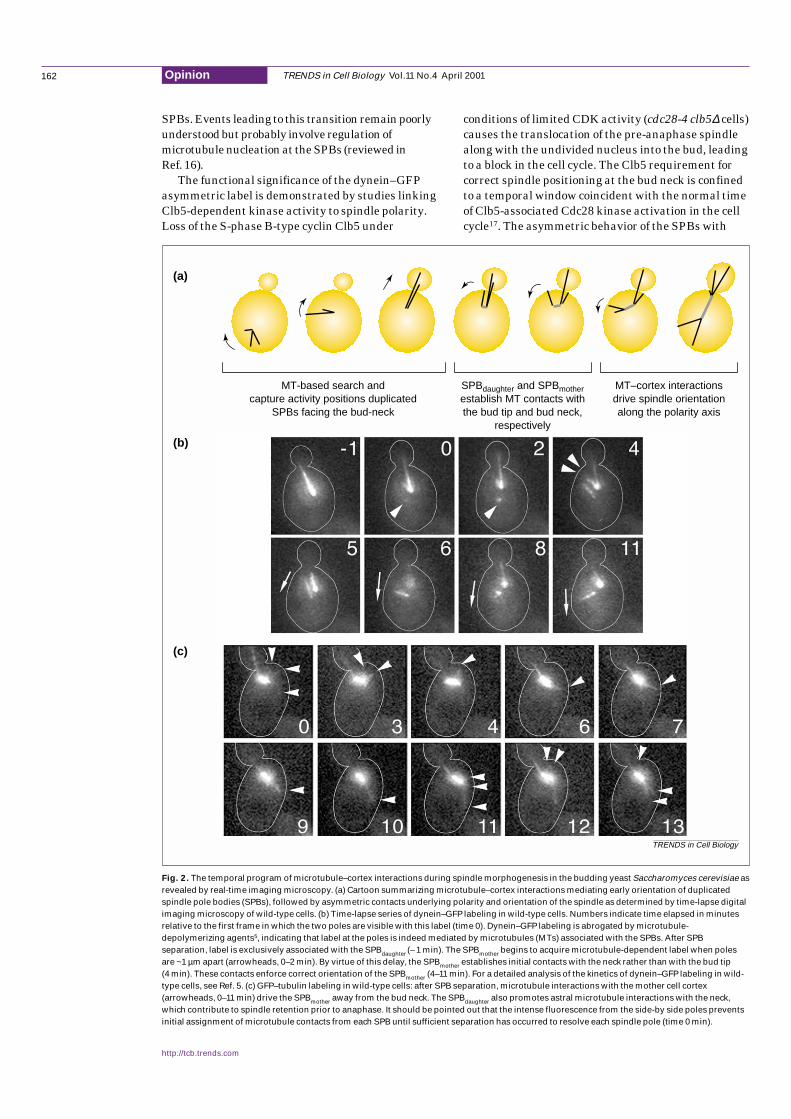
Pre-anaphase spindle positioning is specified by aprecise temporal program of microtubule–cortexinteractions in concert with spindle morphogenesis as seen by real-time imaging microscopy4–6,10 (Fig.2).Cytoplasmic microtubules emerging from the side-by-side SPBs exhibit multiple cycles of growth andshortening. Microtubules grow towards the cell
periphery, where they persist, on average, for less than 3 min. Microtubule penetration into the bud most likelyreflects the stochastic properties of growth andshortening, as well as interactions with the actincytoskeleton (through Kar9, see below)5,11,12. Theseevents orient the duplicated SPBs facing the bud neck inpreparation for spindle assembly (Fig. 2a). During SPBseparation, astral microtubules emanating from theSPBdaughter dynamically interact preferentially with thebud cortex. As separation proceeds, new microtubulecontacts occur with the bud neck region (Fig.2a,b).These contacts direct the remaining, mother-bound,SPB (SPBmother) to interact first with the mother cortexin the vicinity of the bud neck. At this point, spindleassembly (~1 µm-long spindle) and specification ofdaughter- and mother-bound fate, respectively, havebeen completed5,6,10. As the spindle establishes position,the astral microtubules emanating from the SPBmotherinteract dynamically with the mother cell cortex atpoints progressively further away, while the SPBdaughterremains at the bud neck (Fig.2a–c). These interactions,in concert with SPB separation and spindle assembly,drive the SPBmother away from the bud neck, ultimatelyorienting the spindle along the mother–bud axis. Onceoriented, the mobility of the SPBdaughter is restricted tothe neck, whereas the SPBmother is free to rotate10. Thissequence of events points to the importance of initialastral microtubule contacts with the vicinity of the neckin imparting correct fate of the SPBmother (Ref. 6).
Temporal, asymmetric association of dynein–GFP
hallmarks correct fate of the SPBs
The asymmetric nature of cytoplasmicmicrotubule–cortex interactions with the bud and themother cell cortex throughout SPB separation indicatesthat polarity is in fact established early in spindlemorphogenesis. Immunofluorescence experiments havesuggested that it is the newly synthesized SPB that isdestined for the daughter cell13. More recently, studiesbased on in vivo labeling by a dynein heavy chain fusionto green-fluorescent protein (dynein–GFP) haveprovided valuable information on astral microtubulebehavior and SPB polarity during spindlemorphogenesis and karyogamy4–6,14,15. Dynein–GFPdecorates astral microtubules mediating the earlyorientation of duplicated SPBs facing the bud neck inbudded cells (Fig.2b). As the spindle assembles, the bulkof dynein–GFP label remains associated with thedaughter-bound SPB (SPBdaughter). Once the spindlepoles are approximately 1 µm apart, label begins toassociate with the mother-bound pole (SPBmother). This ‘lag’ in dynein–GFP accumulation at the SPBmotherpresumably reflects a delay in astral microtubuleorganization by the SPB outer plaque relative to SPBseparation5,6 and is independent of the orientation ofmicrotubules into the bud or mother cell15. Indeed,following SPB duplication, astral microtubules emergefrom the bridge region between the side-by-side poles2.Throughout the rest of the cell cycle, however, astralmicrotubules are organized by the outer plaques of the
TRENDS in Cell Biology Vol.11 No.4 April 2001
http://tcb.trends.com
161Opinion
Marisa Segal
Dept of MolecularBiology, MB7, The ScrippsResearch Institute, 10550North Torrey Pines Road,La Jolla, CA 92037, USA.
Kerry Bloom
Dept of Biology, 623Fordham Hall CB#3280,University of NorthCarolina at Chapel Hill,Chapel Hill, NC27599-3280, USA.e-mail:[email protected]
TRENDS in Cell Biology
Astralmicrotubule
(a)
(b)
SPB
Spindle
G1 G2START S M
Fig. 1. The spindle pathway in the budding yeast Saccharomyces cerevisiae. (a) Landmark events ofthe spindle pathway. (b) Live images of cells expressing a GFP–α-tubulin fusion (GFP–Tub1),corresponding to the stages depicted in (a). The label is incorporated into cytoplasmic as well asintranuclear microtubules. Overlays of fluorescence and differential interference contrast microscopyare shown. Unbudded G1-phase cells contain a single spindle pole body (SPB). Duplication is primedby formation of a satellite in late G1 (not shown). SPB duplication is completed in early S phase, andSPBs remain side-by-side during DNA replication. Following SPB separation and spindle assembly,the nucleus positions at the bud neck, with the spindle oriented along the mother–daughter axis. Ascells proceed into anaphase of mitosis, the spindle elongates. Once chromosome segregation iscompleted, the spindle disassembles and the cells complete cytokinesis.
SPBs. Events leading to this transition remain poorlyunderstood but probably involve regulation ofmicrotubule nucleation at the SPBs (reviewed inRef. 16).
The functional significance of the dynein–GFPasymmetric label is demonstrated by studies linkingClb5-dependent kinase activity to spindle polarity.Loss of the S-phase B-type cyclin Clb5 under
conditions of limited CDK activity (cdc28-4 clb5∆ cells)causes the translocation of the pre-anaphase spindlealong with the undivided nucleus into the bud, leadingto a block in the cell cycle. The Clb5 requirement forcorrect spindle positioning at the bud neck is confinedto a temporal window coincident with the normal timeof Clb5-associated Cdc28 kinase activation in the cellcycle17. The asymmetric behavior of the SPBs with
TRENDS in Cell Biology Vol.11 No.4 April 2001
http://tcb.trends.com
162 Opinion
TRENDS in Cell Biology
MT-based search andcapture activity positions duplicated
SPBs facing the bud-neck
SPBdaughter and SPBmotherestablish MT contacts withthe bud tip and bud neck,
respectively
MT–cortex interactionsdrive spindle orientationalong the polarity axis
(a)
(b)
(c)
Fig. 2. The temporal program of microtubule–cortex interactions during spindle morphogenesis in the budding yeast Saccharomyces cerevisiae asrevealed by real-time imaging microscopy. (a) Cartoon summarizing microtubule–cortex interactions mediating early orientation of duplicatedspindle pole bodies (SPBs), followed by asymmetric contacts underlying polarity and orientation of the spindle as determined by time-lapse digitalimaging microscopy of wild-type cells. (b) Time-lapse series of dynein–GFP labeling in wild-type cells. Numbers indicate time elapsed in minutesrelative to the first frame in which the two poles are visible with this label (time 0). Dynein–GFP labeling is abrogated by microtubule-depolymerizing agents5, indicating that label at the poles is indeed mediated by microtubules (MTs) associated with the SPBs. After SPBseparation, label is exclusively associated with the SPBdaughter (– 1 min). The SPBmother begins to acquire microtubule-dependent label when polesare ~1 µm apart (arrowheads, 0–2 min). By virtue of this delay, the SPBmother establishes initial contacts with the neck rather than with the bud tip(4 min). These contacts enforce correct orientation of the SPBmother (4–11 min). For a detailed analysis of the kinetics of dynein–GFP labeling in wild-type cells, see Ref. 5. (c) GFP–tubulin labeling in wild-type cells: after SPB separation, microtubule interactions with the mother cell cortex(arrowheads, 0–11 min) drive the SPBmother away from the bud neck. The SPBdaughter also promotes astral microtubule interactions with the neck,which contribute to spindle retention prior to anaphase. It should be pointed out that the intense fluorescence from the side-by side poles preventsinitial assignment of microtubule contacts from each SPB until sufficient separation has occurred to resolve each spindle pole (time 0 min).

regard to dynein–GFP labeling is lost in cdc28-4 clb5∆cells. Both poles display microtubule-mediated labelduring separation and therefore behave as daughter-bound. Thus, the lack of built-in delay of microtubuleorganization resulting in simultaneous SPB labelingtranslates into failure to assign mother-bound fate toone SPB. By contrast, the program is not perturbed byloss of either Clb3 or Clb4 B-type cyclins. Instead,these cyclins preferentially promote SPB separationand spindle assembly6. Genetic analysis has alsoassigned distinct roles for B-type cyclins in restrictingSPB duplication to once per cell cycle9. Thus, variousaspects of spindle morphogenesis are differentiallysubjected to cell-cycle control.
The cortical program underlying correct establishment
of spindle polarity
Functional cytoplasmic microtubule attachments aredrawn to specialized cortical sites of the mother andbud cells to achieve pre-anaphase spindleorientation. A prominent role for the dynactin–dynein system in mediating these functionalattachments has been proposed4. Establishment ofspindle polarity, however, must be primed by cellularfactors partitioning asymmetrically between the bud and mother cortex, a process coupled to bud-siteselection and polarized growth characteristic of yeastcells entering S phase18. Indeed, on the one hand,several cortical components implicated in spindleorientation such as the formin Bni1, a target of thepolarizing machinery, and the actin interactorAip3/Bud619–21 are localized initially to the bud tip.The C-terminal domain of Bni1 interacts with Bud6and is essential for Bni1 function in bud-siteselection and spindle orientation22. On the otherhand, other presumed cortical elements arerestricted initially to the mother cell during spindleassembly (e.g. Num123).
‘……spindle orientation is coupled to
the establishment of a cell-polarity
axis and division plane — a
principle paramount to metazoan
development.’
Recent work has focused on identifying factorsmediating the process of microtubule-based search andcapture that directs early functional attachments withthe bud cell cortex. These studies assign a pivotal role toBim124,25 and Kar9 in this process21,26. Bim1, a yeasthomolog of human EB125 (the binding partner of theadenomatous polyposis coli, APC), can bind directly tomicrotubules and is required for the high dynamicinstability of microtubules that is characteristic of cellsbefore spindle assembly24. Dynamic instabilityconstitutes a prime feature of a microtubule-basedmechanism that relies on transient cortical interactionsin the search for the bud. Kar9 was identified initially
based on its requirement for karyogamy duringmating27. In subsequent studies, Kar9 has beenimplicated in the orientation of functional microtubuleattachments into the bud during vegetative growth27.The finding that Kar9 interacts with the tail of a type Vmyosin (Myo2) and is transported along actin filamentstowards the bud11,12 accounts for the requirement foractin filaments early in spindle orientation28,29.Accordingly, myo2 mutations or treatment with theactin-depolymerizing agent latrunculin A can preventcorrect transport of Kar9 into the bud and disruptspindle orientation11,30. The association of Kar9 withmicrotubules via Bim1, in turn, provides the basis for a postulated mechanism of microtubule capture31,32
(reviewed in Ref. 33). These findings provide amechanistic understanding of the functional linkagebetween the actin and microtubule cytoskeletons.However, the basis for segregation of one pole into thebud, and the marking of a particular pole (SPBdaughter) inthe course of spindle morphogenesis remains a mystery.The ability to designate differential fates to twoapparently identical structures is an importantproblem in embryonic patterning and one that has beenparticularly refractive to experimental manipulation.
Indeed, spindle polarity relies on temporalgeneration of astral microtubule interactions with the bud and mother cell cortex relative to spindleassembly5,6 (Fig.2). To achieve this, the temporal shift ofmicrotubule nucleation sites from the bridge of the side-by-side SPBs to the respective outer plaques must becoordinated with the program of cortical factorssequentially positioned at the bud tip and bud neckregions. Bud6 follows precisely this temporal pattern of localization20 and is in a position to provide cues forcytoplasmic microtubule–cortex interactions duringspindle assembly34 (see Fig.3 for speculative model).Moreover, Bud6 is more prominent at the bud neck inhaploid cells, paralleling the more prominent role for thebud neck in promoting microtubule–cortex attachmentsduring spindle orientation of haploids. Based on geneticanalysis, this difference has been linked to bud-siteselection control – that is, the mode of orienting celldivisions that distinguishes haploid and diploid yeastcells. Briefly, haploid cells bud adjacent to a previousbudding site (axial pattern), whereas diploid cells canbud either adjacent or opposite to the previous buddingsite (bipolar pattern)18.
Bud3, a protein essential for axial budding ofhaploids, accumulates at the bud neck part-way throughSphase35 and is necessary for efficient association ofBud6 to the neck region (Fig.3a). Deletion of BUD3 inhaploids reduces Bud6 partitioning to the bud neck anddelimits the contribution of microtubule–bud-neckinteractions during spindle morphogenesis34,concomitant with a change to a bipolar (diploid-like)budding pattern. Conversely, deletion of BNI1 displacesBud6 from the bud tip to the neck following budemergence34 and abolishes bipolar budding in diploids36,indicating a role for Bni1 in promoting Bud6 associationto the bud tip (Fig. 3a). Thus, Bud6 partitioning between
TRENDS in Cell Biology Vol.11 No.4 April 2001
http://tcb.trends.com
163Opinion

the bud tip and neck is dictated by cortical elementsrequired for cell polarity (Bni1 at the bud tip) and bud-site selection control (Bud3 at the bud neck; Fig.3a).
The resulting disruption of the Bud6 cortical programin bni1∆mutants leads to exaggerated microtubule–budneck interactions, which perturb spindle polarity anddynamics15,34. By contrast, bud6∆or bni1∆mutants stillsupport Kar9-dependent microtubule capture, eventhough Kar9 is mislocalized from the bud tip22,30. Moreimportantly, Kar9 does not contribute to microtubulecapture at the neck34, suggesting that not all modes ofmicrotubule capture require Kar9. Alternatively, Kar9association along actin cables11,12 raises the possibilitythat the mechanism of microtubule capture reliesadditionally on Kar9-dependent changes of microtubuledynamic parameters occurring within the budenvironment, perhaps by antagonizing Bim1 function,which in turn would contribute to microtubulepersistence and correct targeting to Bud6 cortical sites
in the bud (Fig.3b). This speculative view is inspired by a mechanism relying on control of microtubule dynamicinstability that has been proposed for the role of themicrotubule-binding protein tip1, a CLIP170-homolog in Schizosaccharomyces pombe. Tip1 appears to favorrecovery over catastrophe rates for microtubulesinteracting with the cell cortex, with the net result ofguiding microtubules to the cell ends37.
The role of motor proteins in spindle polarity and
orientation during the cell cycle
Microtubule dynamic properties are regulateddifferentially throughout the cell cycle. Relatively highmicrotubule dynamic instability characterizes G1 andsmall budded cells4,5,24 and is crucial for initialmicrotubule orientation towards the bud tip prior tospindle assembly. Stable microtubule attachments tocortical sites in the bud (persistence >3min), however,have not been observed during vegetative cell growth4,5,14
and therefore a simple tethering mechanism for astralmicrotubules is unlikely, although orientation might stillbe achieved by the sum of transient interactions with the bud cortex. An alternative mechanism wherein motor proteins couple microtubule dynamics to forcegeneration38 is more consistent with these observations.Analysis of motor mutants has been restricted toassessment of bulk microtubule length, nuclearpositioning in fixed cells, and spindle oscillations in livecells. These studies assigned antagonistic and temporallyrestricted roles to the cytoplasmic microtubule motordynein10 and the kinesin-like proteins Kip2 and Kip339,40,as well as Kar341, in regulating microtubule dynamics,mediating nuclear migration to the bud neck andfacilitating spindle translocation (reviewed in Ref.42).That motor proteins indeed effect microtubule dynamicsderives from dramatic phenotypic alterations inmicrotubule lengths in kip2∆or kip3∆38,39 mutants, withKip3 a likely protagonist of microtubule instability, andKip2 a protagonist of microtubule stability. Interestingly,plus-end dynamics of astral microtubules have beenobserved in G1 phase, mitosis and anaphase43. Increasedmicrotubule length, plus-end-directed microtubuledynamics and defects in nuclear migration in kip3∆cells can be united by invoking Kip3 as a plus-end
TRENDS in Cell Biology Vol.11 No.4 April 2001
http://tcb.trends.com
164 Opinion
TRENDS in Cell Biology
(a) (b)
1 2 3 4 5
Establishment of polarity Spindle orientation
Kar9–Bim1
Bim1?Num1
Bud6
Key:
Bni1 Bud6 Bud3 Nucleus SPB Microtubule
Fig. 3. A speculative model for establishment of polarity during spindle morphogenesis in thebudding yeast Saccharomyces cerevisiae. (a) Coordinated spindle assembly and orientation in yeast.(1, 2) Timely capture of microtubules emanating from the bridge at the bud cortex (a processcontributed to by both Kar9 and Bud6) orients duplicated spindle pole bodies (SPBs). At the onset ofSPB separation, the SPBdaughter inherits these microtubule contacts (3). As a short spindle forms, asecond area of Bud6-dependent interactions appears at the bud neck. Accumulation of Bud6 at thebud neck, particularly in haploids, is promoted by the axial cortical component Bud3. Thus, de novoastral microtubules are restricted such that they interact with the bud neck region and are preventedfrom undergoing capture in the bud (4). These new interactions result in the correct fate of theSPBmother and retention of the pre-anaphase spindle at the neck (5). Accordingly, a delay inmicrotubule organization relative to SPB separation (as inferred by the kinetic behavior ofdynein–GFP labeling), in concert with the appearance of this new area for enhancedmicrotubule–cortex interactions at the neck defines the temporal window in which establishment ofcorrect spindle polarity takes place. This model explains how the Clb5-dependent kinase enforcescorrect asymmetry at a point in which two poles are already present. (b) Cartoon highlighting theasymmetric distribution of elements participating in the establishment of spindle polarity in concertwith spindle assembly. Bim1 decorates all cytoplasmic and intranuclear microtubules. For simplicity,Kar9 is represented associating with microtubules through contact with Bim1. Kar9 is delivered to thebud by a Myo2-dependent mechanism presumably involving tracking on actin cables11,12. It remainsunclear whether segregation of Kar9 to the bud compartment precedes or occurs concomitant withmicrotubule capture in wild-type, unperturbed cells. Nevertheless, microtubule-based search-and-capture in the bud always precedes spindle assembly in the unperturbed cell cycle. It is thereforeunlikely that Kar9-driven microtubule tracking via actin cables orients the spindle following assembly.Bud6 association to the bud neck occurs after initiation of spindle assembly (Ref. 34; M. Segal andK. Bloom, unpublished) and prevents microtubules that emanate from the SPBmother from entering thebud following SPB separation. Num1 initially associates with the mother cell cortex in cells with smallbuds during S phase and spindle assembly, compatible with a role in orienting the mother-bound SPBaway from neck cortical interaction sites. Num1 also appears to associate to the bud cell cortex oflarge budded cells (therefore not shown in the cartoon) late in anaphase, raising the possibility of anadditional contribution along with dynein to anaphase spindle translocation into the bud47,48.

microtubule-destabilizing protein, perhaps anchored tosites in the bud through the Bud6–Bni1 cortical scaffold.However, a direct functional link between Kip3, Kar9and the Bud6–Bni1 system has not been proven to date.
Antagonistic roles for various motors might alsocontribute by fine-tuning spindle positioning at the onsetof anaphase. The mitotic spindle is extremely dynamic atthis stage and exhibits oscillations, generally along thepolarity axis, as evidenced initially by differentialinterference microscopy studies10. The oscillatory motionof the pre-anaphase spindle, once oriented to the neck, isdependent upon the action of cytoplasmic dynein15.Because this pre-anaphase spindle does not undergolength changes, spindle dynamics at this stage mustreflect the ability of dynein to slide astral microtubulesalong the cell cortex44. One function of the kinesins Kip3and Kip2 might be to attenuate or antagonize the functionof dynein prior to the onset of anaphase. Interestingly,invitromotility studies indicate that, when conventionalkinesin and axonemal dynein are loaded on the samemicrotubule, microtubule translocation proceeds in akinesin-directed fashion – that is, the action of kinesin isdominant to that of dynein45. The processive nature ofkinesin motors might account for the ability of Kip3 tomask dynein function in pre-anaphase cells. Only upondeletion of KIP3or effectors of Kip3 is the activity ofdynein revealed in increased frequency and duration ofspindle oscillations15. Therefore, the role of Kip3 in initialspindle orientation must also be addressed in this context.Indeed, live-imaging microscopy has documented theimpact of Kip3 in the promotion of spindle disassembly46.By contrast, astral microtubule behavior during spindlemorphogenesis in kip3∆mutants remains to be evaluatedby live-imaging microscopy to refine our understanding ofthe role of Kip3 during early nuclear migration.
Once the spindle has oriented along the mother–budaxis, the nucleus persists at the neck for over 20min inthe unperturbed cell cycle. It is not clear what preventscontinued nuclear migration through the neck at thisjuncture. The exaggerated oscillations observed inbud6∆ mutants34 suggest that the contacts betweenmicrotubules and the bud neck cued by Bud6 might be crucial for correct pre-anaphase neck retention.Additionally, the force from depolymerizingmicrotubules, as outlined above, might not be sufficientto pull the nucleus through the neck unless Bud6 isabsent. Moreover, if cytoplasmic dynein is indeed the‘pulling’ motor, dynein function might be attenuateduntil onset of anaphase. It has been postulated thatthere are pulling forces on the spindle, but there has not been a quantitative measure of their action. Usingan ndc1mutation, which impairs the insertion of thenewly formed SPB in the nuclear envelope and preventsbipolar spindle assembly, Yeh etal.have shown thatdynein exerts the dominant pulling force. Spindletranslocation into the bud in the absence of spindleelongation proceeds at 3.6 µmmin–1, considerablygreater than the 1 µmmin–1 spindle elongationcharacteristic of the fast phase of anaphaseB10. The ‘pulling’ forces are therefore not only engaged
post-anaphase but they are restrained by spindleelongation forces or astral microtubuleassembly–disassembly at the cell cortex. Pre-anaphasenuclear retention at the neck is therefore not a ‘static’phase of the nuclear–spindle cycle. Rather, early Kip3function, in addition to providing a vector for initialspindle orientation, might antagonize cytoplasmicdynein. Spindle oscillations at and through the neck prior to the onset of anaphase reflect the balancebetween Kip3 and dynein. Upon the onset of anaphase,spindle elongation forces and limitations in microtubuledynamics prevent dynein from pulling the entirenucleus into the bud.
Concluding remarks
Coordinated spindle assembly and orientation relies onintegrating the CDK-driven program that impartscorrect inherent spindle polarity with the role of Bud6and associated cortical elements in orienting functionalcytoplasmic microtubule attachments (see Fig. 3a forproposed model). These asymmetric determinants aretightly coupled to cell patterning. Indeed, Bud6 mightprovide an interface between spindle polarity and themachinery responsible for bud-site selection thatspecifies the division plane18. The sequentialassociation of Bud6 to the bud tip and the bud neck,driven by the combination of the polarizing and bud-siteselection machinery, positions the spindle whilepreventing astral microtubule attachments from theSPBmother with the bud cell cortex. This view leads to theidea that spindle orientation is actually coupled to theestablishment of a cell-polarity axis and division plane,a principle paramount to metazoan development.
The direct contribution of motors to spindlepolarity remains an open question. Motor-drivenmicrotubule dynamic behavior is clearly crucial fororientation – yet establishment of polarity appears to be primarily regulated by the temporal control of SPB nucleation function. While the asymmetricassociation of the dynein–GFP fusion might underliepolarity, genetic analysis appears to exclude a directrole for dynein in these events6. The possible role ofother motors in imparting and/or mediating polarityremains to be assessed.
Regulation of spindle orientation duringdevelopment is exploited to specify the direction of the cleavage plane to achieve precise embryonic cellpatterning. The mechanism that governs spindleorientation shares common features with the yeastsystem as shown in recent studies in C.elegansandDrosophila, among others. The principles for poleasymmetry described for budding yeast might beextended to these systems. For example, the asymmetricinteractions of centrosomes at specialized cortical sites,involving microtubules and actin, appear to dictatespindle orientation in the two-cell-stage C.elegansembryo1. Future studies might uncover factorsanalogous to yeast Kar9 or Bud6 interplaying withcentrosome function to specify cleavage planes and, in turn, distinct developmental cell fates.
TRENDS in Cell Biology Vol.11 No.4 April 2001
http://tcb.trends.com
165Opinion

References
1 Rhyu, M.S. and Knoblich, J.A. (1995) Spindleorientation and asymmetric cell fate. Cell 82,523–526
2 Byers, B. (1981) Cytology of the yeast life cycle.In The Molecular Biology of the YeastSaccharomyces: Life Cycle and Inheritance(Strathern, J.N. et al., eds), pp. 59–96, ColdSpring Harbor Laboratory Press
3 Lew, D.J. et al. (1997) Cell cycle control inSaccharomyces cerevisiae. In The Molecular andCellular Biology of the Yeast Saccharomyces(Pringle, J.R. et al., eds), pp. 607–695, Cold SpringHarbor Laboratory Press
4 Carminati, J.L. and Stearns, T. (1997)Microtubules orient the mitotic spindle in yeastthrough dynein-dependent interactions with thecell cortex. J. Cell Biol. 138, 629–641
5 Shaw, S.L. et al. (1997) Astral microtubule-basedsearching mechanism for spindle orientation andnuclear migration into the bud. J. Cell Biol. 139,985–994
6 Segal, M. et al. (2000) Coordinated spindle assemblyand orientation requires Clb5p-dependent kinase inbudding yeast. J. CellBiol.148, 441–452
7 Nasmyth, K. (1993) Control of the yeast cell cycleby the Cdc28 protein kinase. Curr. Opin. CellBiol. 5, 166–179
8 Fitch, I. et al. (1992) Characterization of four B-typecyclin genes of the budding yeast Saccharomycescerevisiae. Mol. Biol. Cell 3, 805–818
9 Haase, S.B. et al. (2001) Multi-step control ofspindle pole body duplication by cyclin-dependentkinase. Nat. Cell Biol. 3, 38–42
10 Yeh, E. et al. (1995) Spindle dynamics and cellcycle regulation of dynein in the budding yeast,Saccharomyces cerevisiae. J. Cell Biol. 130,687–700
11 Yin, P. et al. (2000) Myosin V orientates themitotic spindle in yeast. Nature 406, 1013–1015
12 Beach, D.L. et al. (2000) The role of the proteinsKar9 and Myo2 in orienting the mitotic spindle ofbudding yeast. Curr. Biol. 10, 1497–1506
13 Vallen, E.A. et al. (1992) Asymmetric mitoticsegregation of the yeast spindle pole body. Cell 69,505–515
14 Maddox, P. et al. (1999) Microtubule dynamicsfrom mating through the first zygotic division inthe budding yeast Saccharomyces cerevisiae.J. Cell Biol. 144, 977–987
15 Yeh, E. et al. (2000) Dynamic positioning ofmitotic spindles in yeast: role of microtubulemotors and cortical determinants. Mol. Biol. Cell11, 3949–3961
16 Schiebel, E. (2000) γ-Tubulin complexes: bindingto the centrosome, regulation and microtubulenucleation. Curr. Opin. Cell Biol. 12, 113–118
17 Segal, M. et al. (1998) Clb5-associated kinaseactivity is required early in the spindle pathwayfor correct preanaphase nuclear positioning inSaccharomyces cerevisiae. J. Cell Biol. 143,135–145
18 Chant, J. (1999) Cell polarity in yeast. Annu. Rev.Dev. Biol. 15, 365–391
19 Evangelista, M. et al. (1997) Bni1p, a yeast forminlinking Cdc42p and the actin cytoskeleton duringpolarized morphogenesis. Science 276, 118–122
20 Amberg, D.C. et al. (1997) Aip3p/Bud6p, a yeastactin-interacting protein that is involved inmorphogenesis and the selection of bipolarbudding sites. Mol. Biol. Cell 8, 729–753
21 Heil-Chapdelaine, R.A. et al. (1999) Formin’ theconnection between microtubules and the cellcortex. J. Cell Biol. 144, 809–811
22 Lee, L. et al. (1999) Control of mitotic spindleposition by the Saccharomyces cerevisiae forminBni1p. J. Cell Biol. 144, 947–961
23 Farkasovsky, M. and Kuntzel, H. (1995) YeastNum1p associates with the mother cell cortexduring S/G2 phase and affects microtubularfunctions. J. Cell Biol. 131, 1003–1014
24 Tirnauer, J.S. et al. (1999) Yeast Bim1p promotesthe G1-specific dynamics of microtubules. J. CellBiol. 145, 993–1007
25 Tirnauer, J.S. and Bierer, B.E. (2000) EB1proteins regulate microtubule dynamics, cellpolarity, and chromosome stability. J. Cell Biol.149, 761–766
26 Bloom, K. (2000) It’s a kar9ochore to capturemicrotubules. Nat. Cell Biol. 6, 96–98
27 Miller, R.K. and Rose, M.D. (1998) Kar9p is a novelcortical protein required for cytoplasmic microtubuleorientation in yeast. J. Cell Biol.140, 377–390
28 Theesfeld, C.L. et al. (1999) The role of actin inspindle orientation changes during theSaccharomyces cerevisiae cell cycle. J. Cell Biol.146, 1019–1032
29 Heil-Chapdelaine, R.A. et al. (2000) Dynein-dependent movements of the mitotic spindle inSaccharomyces cerevisiae do not requirefilamentous actin. Mol. Biol. Cell 11, 863–872
30 Miller, R.K. et al. (1999) The cortical localizationof the microtubule orientation protein, Kar9p, isdependent upon actin and proteins required forpolarization. J. Cell Biol. 144, 963–975
31 Korinek, W.S. et al. (2000) Molecular linkageunderlying microtubule orientation towardcortical sites in yeast. Science 287, 2257–2259
32 Lee, L. et al. (2000) Positioning of the mitoticspindle by a cortical microtubule capturemechanism. Science 287, 2260–2262
33 Schuyler, S.C. and Pellman, D. (2001) Search,capture and signal: games microtubules andcentrosomes play. J. Cell Sci. 114, 247–255
34 Segal, M. et al. (2000) Bud6 directs sequentialmicrotubule interactions with the bud tip and budneck during spindle morphogenesis inSaccharomyces cerevisiae. Mol. Cell. Biol.11,3689–3702
35 Chant, J. et al. (1995) Role of Bud3p in producingthe axial budding pattern of yeast. J. Cell Biol.129, 767–778
36 Zahner, J.E. et al. (1996) Genetic analysis of thebipolar pattern of bud site selection in the yeastSaccharomyces cerevisiae. Mol. Cell. Biol. 16,1857–1870
37 Brunner, D. and Nurse, P. (2000) CLIP170-liketip1p spatially organizes microtubular dynamicsin fission yeast. Cell 102, 695–704
38 Lombillo, V.A. et al. (1995) Minus-end-directedmotion of kinesin-coated microspheres driven bymicrotubule depolymerization. Nature 373,161–164
39 Cottingham, F.R. and Hoyt, M.A. (1997) Mitoticspindle positioning in Saccharomyces cerevisiae isaccomplished by antagonistically acting microtubulemotor proteins. J. Cell Biol.138, 1041–1053
40 DeZwaan. T.M. et al. (1997) Kinesin-related KIP3of Saccharomyces cerevisiae is required for adistinct step in nuclear migration. J. Cell Biol.138, 1023–1040
41 Saunders, W.S. et al. (1997) The Saccharomycescerevisiae kinesin-related motor Kar3p acts atpreanaphase spindle poles to limit the numberand length of cytoplasmic microtubules. J. CellBiol. 137, 417–431
42 Hunter, A.W. and Wordeman, L. (2000) How motorproteins influence microtubule polymerizationdynamics. J. Cell Sci. 113, 4379–4389
43 Maddox, P.S. (2000) The polarity and dynamics ofmicrotubule assembly in the budding yeastSaccharomyces cerevisiae. Nat. Cell Biol. 2, 36–41
44 Adames, N.R. and Cooper, J.A. (2000)Microtubule interactions with the cell cortexcausing nuclear movements in Saccharomycescerevisiae. J. Cell Biol. 149, 863–874
45 Vale, R.D. et al. (1992) Directional instability ofmicrotubule transport in the presence of kinesinand dynein, two opposite polarity motor proteins.J. Cell Biol. 119, 1589–1596
46 Straight, A.F. et al. (1998) Time-lapse microscopyreveals unique roles for kinesins during anaphasein budding yeast. J. Cell Biol. 143, 687–694
47 Heil-Chapdelaine, R.A. et al. (2000) The corticalprotein num1p is essential for dynein-dependentinteractions of microtubules with the cortex.J. Cell Biol. 151, 1337–1344
48 Farkasovsky, M. and Kuntzel, H. (2001) CorticalNum1p interacts with the dynein intermediatechain Pac11p and cytoplasmic microtubules inbudding yeast. J. Cell Biol. 152, 251–262
TRENDS in Cell Biology Vol.11 No.4 April 2001
http://tcb.trends.com 0962-8924/01/$ – see front matter © 2001 Elsevier Science Ltd. All rights reserved. PII: S0962-8924(01)01958-4
166 Opinion
ErratumThe legend to Fig. 2 of the article 'Timing is everything: regulation of mitotic exit and cytokinesis by the MEN and SIN'
by Dannel McCollum and Kathleen L. Gould (Trends Cell Biol. 11, 89–95, 2001) contains a production error. The final part of the legend should read:
Following cyclin-dependent kinase (CDK) inactivation in anaphase, Cdc7p recruits the Sid1p–Cdc14p kinase complex to the SPB12. Sid1p–Cdc14p could then promote activation of the SPB-localized Sid2p–Mob1p kinase complex,
which then translocates to the cell-division site to trigger initiation of cell division7,8,13.
We apologize to the authors and our readers for the error.