Embed Size (px)
Citation preview

11 16
2 TINJAUAN PUSTAKA
2.1 Mikroalga
Mikroalga merupakan mikroorganisme fotosintetik yang dapat ditemukan di
perairan tawar dan laut. Mekanisme fotosintesis mikroalga mirip dengan
tumbuhan darat, dikarenakan kesamaan pada struktur selulosa yang
dimilikinya.Bila dibandingkan dengan organisme fotosintetik lainnya, mikroalga
paling efisien dalam menangkap dan memanfaatkan energi matahari dan CO2
untuk keperluan fotosintesis karena organisme ini mengandung klorofil serta
pigmen-pigmen lain untuk mengkonversi fotosintesis menjadi biomassa dan
akumulasi pati. Mikroalga hidup secara planktonik di perairan, namun juga dapat
hidup secara epifit dan bentik di dasar perairan yang memiliki intensitas cahaya
yang cukup (Rodjaroen et al. 2007; Gouveia 2011; Barsanti & Gualtieri 2005).
Mikroalga juga memiliki bentuk yang bervariasi seperti filamen, spiral dan bulat.
Berbagai macam morfologi mikroalga dapat dilihat pada Gambar 1.
Gambar 1 Morfologi mikroalga A: Pterosperma, B: Nephroselmis, C:Tetraselmis
D: Chlorella, E: Oocytis, F: Haematococcus, G: Pediastrum, H: Bulbochaete, I: Chaetophora dan J: Ulothrix (Leliaert et al. 2012).
Mikroalga dapat dibagi ke dalam empat kelompok utama (NREL 2003):
1) Diatom (Bacillariophyceae).
Mikroalga dalam kelompok ini mendominasi mikroalga di laut, namun
beberapa jenis diketahui hidup di air tawar. Sebanyak 100.000 jenis mikroalga
yang termasuk dalam kelompok ini. Diatom mengandung silika yang
5
terpolimerisasi dalam dinding sel. Karbon disimpan dalam bentuk minyak nabati
maupun polimer karbohidrat yang disebut chrysolaminarin.
2) Alga hijau (Chlorophyceae).
Mikroalga yang memiliki kelimpahan tinggi terutama di perairan tawar dan
hidup dalam bentuk soliter maupun koloni. Karbon disimpan dalam bentuk pati.
3) Alga hijau biru (Cyanophyceae).
Mikroalga kelompok ini memiliki struktur yang lebih menyerupai bakteri
dan berperan dalam fiksasi nitrogen. Sekitar 2000 jenis mikroalga yang termasuk
dalam kelompok ini tersebar dalam berbagai habitat.
4) Ganggang emas (Chrysophyceae).
Kelompok alga ini menyerupai diatom, namun memiliki pigmen yang lebih
rumit, dan nampak berwarna kuning, jingga atau cokelat.
Mikroalga telah sejak lama dimanfaatkan sebagai sumber bahan makanan,
terutama sebagai sumber vitamin, antioksidan, pewarna atau bahan aditif yang
aman, serta digunakan pula dalam industri farmakologi dalam skala besar. Hal ini
tidak lepas dari komposisi kimia yang terkandung dalam mikroalga, dapat dilihat
pada Tabel 1.
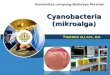
Tabel 1 Komposisi kimia protein, karbohidrat, lipid dan asam nukleat dalam %
dari bobot kering mikroalga.
Mikroalga Protein Karbohidrat Lipid As. Nukleat
Scenedesmus obliquus
Scenedesmus quandricauda
Scenedesmus dimorphus
Chlamydomonas rheinhardii
Chlorella vulgaris
Chlorella pyrenoidosa
Spirogyra sp.
Dunaliella salina
Euglena gracilis
Prymnesium parvum
Tetraselmis maculata
Porphyridium cruentum
Spirulina platensis
Spirulina maxima
Synechoccus sp. Anabaena cylindrica
50-56
47
48-18
48
51-58
57
56-20
57
39-61
28-45
52
28-39
46-63
60-71
63 43-56
10-17
10-
21-52
17
12-17
26
33-64
32
14-18
25-33
15
40-57
48-14
13-16
15 25-30
12-14
11,9
16-40
21
14-22
12
11-21
16
14-20
22-38
23
29-14
24-9
26-7
11 14-7
3-6
3-
3-
3-
4-5
4-
4-
4-
4-
1-2
1-
1-
2-5
3-4,5
5 5-
Sumber: Becker (1994)

6
2.2 Pemanfaatan Mikroalga di Bidang Kesehatan
Mikroalga dapat dimanfaatkan sebagai bahan baku industri farmasi dan
kosmetika, karena adanya kandungan berbagai senyawa kimia yang dapat
dimanfaatkan sebagai bahan dasar untuk pengobatan dan pencegahan berbagai
macam penyakit. Yuan dan Walsh (2006) menjelaskan bahwa konsumsi alga laut
berkorelasi dengan rendahnya tingkat penderita kanker payudara di Asia Timur.
Sebagai contoh, prevalensi kasus penderita kanker payudara dalam 1 tahun per
100.000 penduduk di Jepang dan Cina masing-masing adalah 42,2 dan 13,1,
dibandingkan dengan kasus di Amerika Utara dan Eropa yang masing-masing
sebesar 125,9 dan 106,2. Teas et al. (2004) juga menjelaskan bahwa sebagian
besar kelompok masyarakat di Chad mengkonsumsi Spirulina rata-rata sebanyak
1-2 sendok makan (3-13 g) per harinya, hal ini diyakini dapat mencegah infeksi
virus HIV.
Hasil-hasil riset menjelaskan bahwa terdapat komponen aktif mikroalga
yang menunjukkan aktivitas biologis sebagai antivirus. Talyshinsky et al. (2002)
menjelaskan bahwa dekstran sulfat dan polisakarida yang dihasilkan mikroalga
berpotensi menghambat HIV tipe 1 dan 2 dengan cara menghambat induksi
sitopatogenetik dan ekspresi antigen dari virus HIV. Sulfat polisakarida yang
dihasilkan juga dapat menghambat aktivitas reversetranscriptase dan RNAse pada
proses replikasi retrovirus. Hasil riset Shih et al. (2003) menjelaskan bahwa
allophycocyanin yang dihasilkan oleh Spirulina platensis dapat menetralisir efek
sitopatik dari enterovirus pada sel manusia secara in vitro.
2.3 Hepatitis C
Hepatitis merupakan penyakit yang menyebabkan pembekakan pada hati.
Penyakit hepatitis terdiri atas beberapa jenis, yaitu hepatitis A, B, C, D, E, F dan
G. Ketujuh hepatitis ini disebabkan oleh virus yang berbeda (WHO 2002).
Penderita hepatitis C seringkali tidak menunjukkan gejala khusus walaupun telah
bertahun-tahun terinfeksi. Gejala yang ditunjukkan sangat umum seperti lelah,
hilangnya selera makan, mual, sakit perut, urin menjadi gelap dan kulit atau mata
berwarna kuning (Solga et al. 2007). Penderita baru menyadari bahwa telah
7
terinfeksi virus hepatitis C (HCV) ketika berada pada tahap yang lebih kritis.
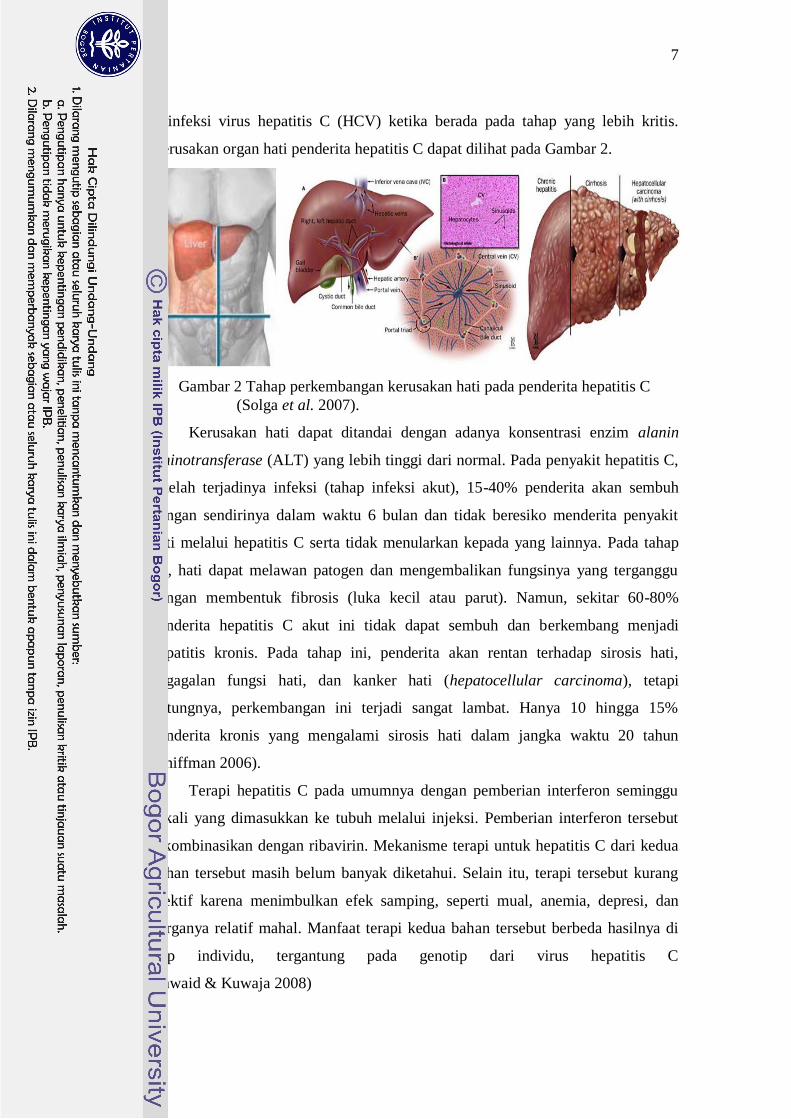
Kerusakan organ hati penderita hepatitis C dapat dilihat pada Gambar 2.
Gambar 2 Tahap perkembangan kerusakan hati pada penderita hepatitis C
(Solga et al. 2007).
Kerusakan hati dapat ditandai dengan adanya konsentrasi enzim alanin
aminotransferase (ALT) yang lebih tinggi dari normal. Pada penyakit hepatitis C,
setelah terjadinya infeksi (tahap infeksi akut), 15-40% penderita akan sembuh
dengan sendirinya dalam waktu 6 bulan dan tidak beresiko menderita penyakit
hati melalui hepatitis C serta tidak menularkan kepada yang lainnya. Pada tahap
ini, hati dapat melawan patogen dan mengembalikan fungsinya yang terganggu
dengan membentuk fibrosis (luka kecil atau parut). Namun, sekitar 60-80%
penderita hepatitis C akut ini tidak dapat sembuh dan berkembang menjadi
hepatitis kronis. Pada tahap ini, penderita akan rentan terhadap sirosis hati,
kegagalan fungsi hati, dan kanker hati (hepatocellular carcinoma), tetapi
untungnya, perkembangan ini terjadi sangat lambat. Hanya 10 hingga 15%
penderita kronis yang mengalami sirosis hati dalam jangka waktu 20 tahun
(Shiffman 2006).
Terapi hepatitis C pada umumnya dengan pemberian interferon seminggu
sekali yang dimasukkan ke tubuh melalui injeksi. Pemberian interferon tersebut
dikombinasikan dengan ribavirin. Mekanisme terapi untuk hepatitis C dari kedua
bahan tersebut masih belum banyak diketahui. Selain itu, terapi tersebut kurang
efektif karena menimbulkan efek samping, seperti mual, anemia, depresi, dan
harganya relatif mahal. Manfaat terapi kedua bahan tersebut berbeda hasilnya di
tiap individu, tergantung pada genotip dari virus hepatitis C
(Jawaid & Kuwaja 2008)
8
2.4 Virus Hepatitis C
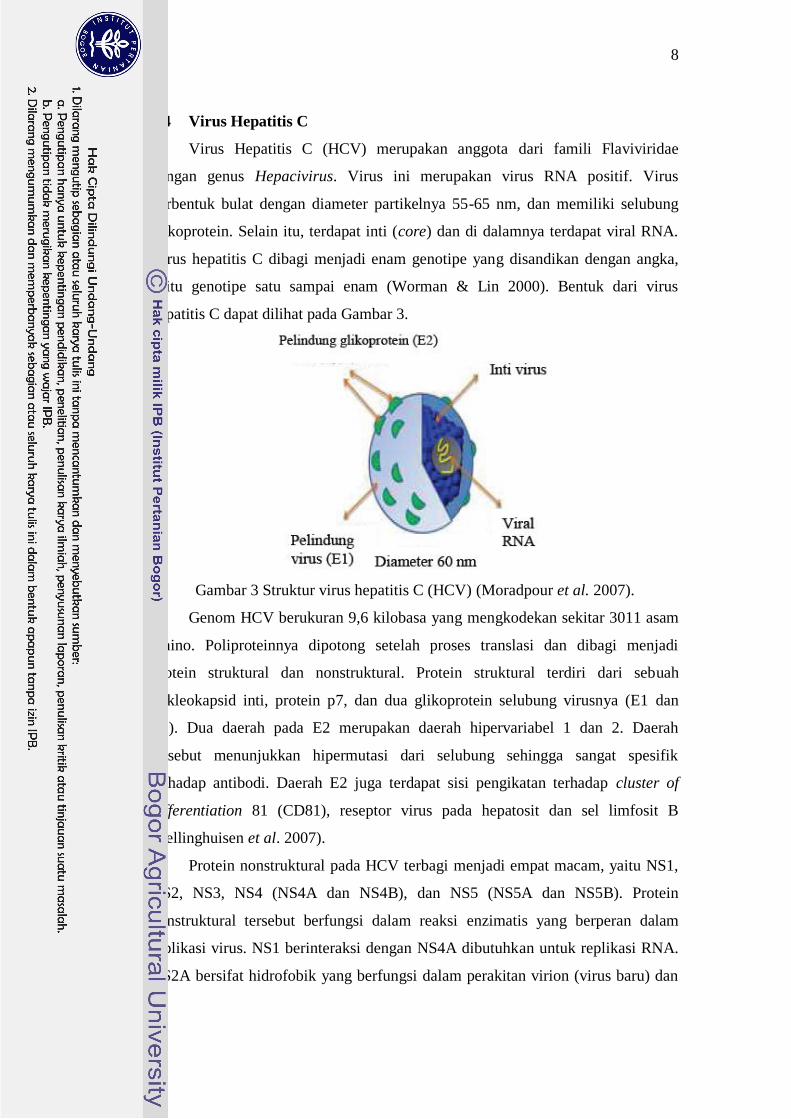
Virus Hepatitis C (HCV) merupakan anggota dari famili Flaviviridae
dengan genus Hepacivirus. Virus ini merupakan virus RNA positif. Virus
berbentuk bulat dengan diameter partikelnya 55-65 nm, dan memiliki selubung
glikoprotein. Selain itu, terdapat inti (core) dan di dalamnya terdapat viral RNA.
Virus hepatitis C dibagi menjadi enam genotipe yang disandikan dengan angka,
yaitu genotipe satu sampai enam (Worman & Lin 2000). Bentuk dari virus
hepatitis C dapat dilihat pada Gambar 3.
Gambar 3 Struktur virus hepatitis C (HCV) (Moradpour et al. 2007).
Genom HCV berukuran 9,6 kilobasa yang mengkodekan sekitar 3011 asam
amino. Poliproteinnya dipotong setelah proses translasi dan dibagi menjadi
protein struktural dan nonstruktural. Protein struktural terdiri dari sebuah
nukleokapsid inti, protein p7, dan dua glikoprotein selubung virusnya (E1 dan
E2). Dua daerah pada E2 merupakan daerah hipervariabel 1 dan 2. Daerah
tersebut menunjukkan hipermutasi dari selubung sehingga sangat spesifik
terhadap antibodi. Daerah E2 juga terdapat sisi pengikatan terhadap cluster of
differentiation 81 (CD81), reseptor virus pada hepatosit dan sel limfosit B
(Tellinghuisen et al. 2007).
Protein nonstruktural pada HCV terbagi menjadi empat macam, yaitu NS1,
NS2, NS3, NS4 (NS4A dan NS4B), dan NS5 (NS5A dan NS5B). Protein
nonstruktural tersebut berfungsi dalam reaksi enzimatis yang berperan dalam
replikasi virus. NS1 berinteraksi dengan NS4A dibutuhkan untuk replikasi RNA.
NS2A bersifat hidrofobik yang berfungsi dalam perakitan virion (virus baru) dan

9
pelepasan partikel virus. NS2B membentuk kompleks dengan NS3 berperan
sebagai kofaktor bagi serin protease dari NS3. Protein NS3 mengkodekan RNA
helikase yang berperan dalam replikasi virus. NS5A merupakan daerah yang
sensitif terhadap interferon, sedangkan NS5B berperan di dalam RNA-dependent
RNA polimerase (RdRp) (Tellinghuisen et al. 2007). Peta genomik dari HCV
dapat dilihat pada Gambar 4.
Gambar 4 Peta genomik HCV (Anzola dan Burgos 2003).
2.5 Ribonucleid Acid (RNA) Helikase
Helikase berasal dari kata “helix” yang berarti struktur pasangan DNA
“double helix” dan “ase” yang berarti enzim, sehingga helikase berarti enzim yang
memisahkan pasangan rantai DNA (DNA helikase) atau RNA (RNA helikase).
Helikase pertama kali ditemukan dalam proses replikasi DNA bakteri
Eschericia coli. RNA helikase ditemukan pada bakteri, khamir, dan virus. Pada
virus hepatitis C, enzim ini dikodekan oleh protein NS3 RNA helikase
(Kadare & Haenni 1997).
Mekanisme kerja RNA helikase HCV secara umum adalah pertama-tama
helikase akan berikatan pada ujung 3’ RNA utas ganda. Tahap kedua, ATP akan
berikatan pada sisi aktif RNA helikase dan dihidrolisis pada gugus fosfat terluar
menghasilkan ADP dan fosfat anorganik (Pi). Pada proses hidrolisis ATP ini
mengeluarkan energi yang cukup besar dan digunakan untuk memisahkan RNA
utas ganda menjadi utas tunggal. Pemisahan RNA utas ganda dilakukan dengan
pemutusan ikatan hidrogen yang mengikat kedua utas tersebut

10
(Utama et al. 2005). Mekanisme kerja RNA helikase HCV dapat dilihat pada
Gambar 5.
Gambar 5 Mekanisme kerja RNA helikase HCV (Hairany 2010).
Berdasarkan mekanisme kerja tersebut, selain memiliki aktivitas untuk
memisahkan utas ganda RNA, RNA helikase juga memiliki aktivitas untuk
menghidrolisis ATP (ATPase) dan aktivitas pengikatan RNA (RNA-binding).
Ketiga aktivitas ini saling berpengaruh satu dengan lainnya. Oleh karena itu,
helikase menjadi target yang potensial untuk penemuan obat antivirus. Obat
antivirus ini dapat dikembangkan dengan suatu senyawa yang dapat menghambat
(inhibitor) aktivitas helikase.
2.6 Polisakarida
Polisakarida, atau bisa disebut “glikan”, terdiri dari monosakarida dan
turunannya. Polisakarida terbagi menjadi homopolisakarida dan
heteropolisakarida. Homopolisakarida atau homoglikan merupakan polisakarida
yang penyusunnya hanya terdiri dari satu jenis monosakarida, sedangkan
penyusun heteropolisakarida lebih dari satu jenis monosakarida. Komponen
umum polisakarida adalah glukosa, fruktosa, galaktosa, mannosa, arabinosa dan
xylosa. Beberapa turunan monosakarida yang terdapat pada polisakarida adalah
gula amino (glukosamin dan galaktosamin) dan asam gula sederhana (glukuronat
dan asam iduronat). Penyebutan homopolisakarida dapat berdasarkan unit gula

11
penyusunnya, sehingga glukosa homopolisakarida dapat disebut “glukan”, sama
halnya dengan mannosa homopolisakarida yang dapat disebut “mannan”
(d’Ayala et al. 2008).
Polisakarida telah digunakan sebagai pengental, flokulan dan minyak
pelumas. Beberapa polisakarida dari mikroalga berpotensi sebagai antivirus
(Huleihel et al. 2001). Salah satu jenis mikroalga merah, Porphyridium cruentum
merupakan salah satu penghasil polisakarida ekstraseluler dalam jumlah besar.
Sel-sel mikroalga dibungkus oleh polisakarida sulfat dalam bentuk gel. Selama
pertumbuhan dalam media cair, viskositas medium meningkat karena pengeluaran
polisakarida dari permukaan sel ke dalam media (polisakarida larut air). Kapsul
polisakarida paling tipis selama fase pertumbuhan dan tebal selama fase stasioner
(Arad & Richmond 2004). Menurut Laurienzo (2010) bahwa mikroalga diketahui
memanfaatkan polisakarida yang disintesisnya untuk bertahan hidup pada kondisi
lingkungan yang ekstrim. Letak polisakarida pada sel mikroalga dapat dilihat pada
Gambar 6.
Gambar 6 Polisakarida Porphyridium cruentum (Arad & Richmond 2004).
Prosedur isolasi polisakarida dari mikroorganisme tergantung pada letak
polisakarida terikat pada dinding sel atau diekskresikan oleh sel sebagai pelindung
atau pengotor. Isolasi dapat dilakukan dengan ekstraksi dari biomassa sel. Namun,
pada masa ini isolasi polisakarida dilakukan dengan sentrifugasi maupun filtrasi
untuk memisahkan produk dari sel (Giavasis & Bilianderis 2006).

12
2.7 Kromatografi
Kromatografi merupakan suatu proses migrasi diferensial, komponen-
komponen senyawa yang dibawa oleh fasa gerak, dan ditahan secara selektif oleh
fasa diam. Peristiwa tersebut terjadi di dalam kolom kromatografi. Adanya
peristiwa yang komplek pada metode kromatografi, menjadikan kromatografi
dapat digunakan untuk menganalisis senyawa sampai sedetail mungkin. Prinsip
kromatografi adalah penggunaan dua fase yang berbeda yaitu fasa tetap dan fasa
bergerak. Proses pemisahan tergantung pada gerakan relatif dari dua fasa tersebut
(Al Baarri 2003). Penelitian ini menggunakan 4 teknik kromatografi, yaitu
kromatografi gel filtrasi, kromatografi penukar ion (ion-exchange), kromatografi
lapis tipis dan kromatografi cair kinerja tinggi.
2.7.1 Kromatografi gel filtrasi
Kromatografi gel filtrasiatau sering disebut filtrasi gel merupakan salah satu
metode yang digunakan untuk memisahkan senyawa menurut ukuran dan bentuk.
Sampel kemudian dimasukan pada ujung atas kolom dan elusi dilakukan dengan
memberikan larutan bufer melalui kolom. Larutan bufer ini memiliki prinsip tidak
boleh lebih polar dibandingkan dengan fase diam atau yang disebut juga kolom.
Besar molekul akan terbagi menjadi 3 bagian yang ditunjukkan oleh berbagai
warna pada Gambar 7.
Gambar 7 Kromatografi gel filtrasi (Koolman 2005).
Molekul yang berukuran besar tidak mampu menembus matriks dari kolom
sehingga akan melewati kolom lebih dahulu. Bobot molekul menengah dan bobot

13
molekul kecil akan tertahan oleh kolom lebih lama (Koolman 2005). Batas
pemisahan dari sebuah ukuran merupakan indikasi bobot molekul untuk tipe
polimer (Hagel 1998).
Keuntungan dari metode ini adalah dapat memisahkan dengan baik molekul
besar dari molekul kecil serta dapat menggunakan berbagai pelarut tanpa harus
mengganggu proses pemisahan. Penggunaan kromatografi gel filtrasi ini akan
didapatkan pemisahan yang baik, sensitifitas yang baik, dan waktu yang
diperlukan untuk pemisahan cepat. Selain itu tidak ada sampel yang tertinggal
karena pelarut tidak berinteraksi dengan fase diam (Skoog 2006). Namun
Kehilangan molekul dapat terjadi selama proses pemurnian dengan menggunakan
teknik kromatografi gel filtrasi karena autolisis (Scopes 1987).
Prinsip dasar kromatografi gel filtrasi adalah partikel dengan ukuran yang
berbeda akan dielusi melalui fase stasioner pada tingkat yang berbeda. Hal ini
menyebabkan pemisahan partikel berdasarkan ukuran. Setiap kolom memiliki
jangkauan berat molekul yang dapat dipisahkan. Molekul besar tidak dapat
terjebak dalam matriks fase diam sehingga akan terlebih dahulu terlewati kolom.
Bobot molekul menengah dan kecil terjebak dalam matriks sehingga akan lebih
lama untuk terlewati fase diam (Skoog 2006).
2.7.2 Kromatografi ion-exchange
Kromatografi penukaran ion (ion-exchange chromatography) biasa
digunakan untuk pemurnian materi biologis. Purwadaria (1999) menjelaskan
bahwa pada sistem kromatografi ini, molekul senyawa dipisahkan berdasarkan
perbedaan afinitas terhadap penukar ion. Afinitas molekul dengan penukar ion
dapat dilepaskan dengan mengubah kadar garam atau pH larutan eluen. Selain itu
sistem pengaturan perubahan kadar garam atau pH eluen baik dengan gradasi
linier ataupun gradasi bertingkat dapat pula mempengaruhi jumlah molekul yang
terpisah.
Metode ini dapat dilakukan dalam dua tipe, yaitu dalam kolom maupun
ruang datar (planar). Terdapat dua tipe penukaran ion, yaitu penukaran kation
(cation exchange) dan penukaran anion (anion exchange). Pada penukaran kation,
fase stasioner bermuatan negatif sedangkan pada penukaran anion, fase stasioner
bermuatan positif, dapat dilihat pada Gambar 8.

14
Gambar 8 Kromatografi ion-exchange (Harper 2005).
Muatan-muatan molekul akan memiliki sifat ketika muatan molekul yang
sama dengan kolom, maka molekul tersebut akan terelusi, namun muatan pada
molekul tidak sama dengan kolom, maka molekul tersebut akan membentuk
ikatan ionik dengan kolom (Carrier 1997). Prinsip dasar yang digunakan adalah
molekul dengan muatan positif bersih pada pH tertentu akan berikatan dengan
gugus fungsional bermuatan negatif seperti carboxylates atau sulfat (penukar
kation). Demikian pula, molekul bermuatan negatif bersih berikatan dengan
molekul bermuatan positif pada gugus fungsional, biasanya tersier atau kuaterner
amina (penukar anion).
2.7.3 Kromatografi lapis tipis
Kromatografi Lapis Tipis (KLT) dikembangkan oleh Izmailoff dan
Schraiber pada tahun 1938. KLT merupakan bentuk kromatografi planar, selain
kromatografi kertas dan elektroforesis. Berbeda dengan kromatografi kolom yang
mana fase diamnya diisikan atau dikemas di dalamnya, pada kromatografi lapis
tipis, fase diamnya berupa lapisan yang seragam (uniform) pada permukaan
bidang datar yang didukung oleh lempeng kaca, pelat aluminium atau pelat plastik
(Gandjar & Rohman 2007). Teknik ini biasa digunakan untuk memisahkan
komponen dari suatu campuran senyawa organik alam, sintetis, dan campuran
kompleks anorganik. Fase gerak yang digunakan tergantung dari senyawa yang
ingin dipisahkan (Harjadi 1976).
Pemisahan komponen melalui berbagai tahap. Pertama dilakukan pemisahan
sampel dengan penotolan pada plat silika yang telah didesain. Plat silika pada
bagian bawah diberi sebuah garis untuk menandakan posisi awal penotolan.

15
Selanjutnya dibuat pula sebuah garis akhir menggunakan pensil. Jarak antara garis
awal dengan garis akhir biasanya 5 cm. Plat yang telah ditotol dengan sampel
dimasukkan ke dalam bejana pengembang yang telah terdapat eluen hasil proses
penjenuhan yang dilakukan selama 20 menit. Penjenuhan berfungsi agar eluen
lebih efektif dalam memisahkan komponen tersebut. Eluen akan memisahkan
komponen hingga garis akhir yang telah didesain. Semakin dekat kepolaran antara
sampel dengan eluen maka sampel akan semakin terbawa oleh fase gerak tersebut
(Wilson & Walker 1994). Hal tersebut dapat dilihat pada Gambar 9.
Gambar 9 Kromatografi lapis tipis (Tissue 1996).
Tahapan selanjutnya adalah visualisasi atau deteksi. Deteksi atau visualisasi
sampel yang tidak berwarna dapat menggunakan dua cara, yaitu penyinaran
dengan sinar UV (254 nm dan 356 nm) dan pereaksi kimia (ninhidrin, FeSO4,
Dragendroff, dan anilin). Pada saat disinari dengan sinar UV, komponen yang
terpisahkan akan terlihat seperti spot atau bidang kecil yang berwarna gelap.
Deteksi komponen juga dapat dilakukan dengan menempatkan kromatogram pada
bejana tertutup yang telah dijenuhkan dengan kristal iod. Uap kristal iod bereaksi
dengan komponen yang terpisahkan dan terlihat seperti bercak-bercak kecoklatan.
Aplikasi dari teknik ini dapat digunakan untuk analisis kuantitatif
(membandingkan retardation factor (Rf) senyawa murni dengan komponen, pola
sidik jari, dan menentukan jumlah komponen) dan preparatif (untuk memperoleh
senyawa murni). Nilai Rf yang akan dihasilkan dari suatu senyawa bernilai sama
meskipun jarak plat yang digunakan berbeda (Wilson & Walker 1994).
2.7.4 Kromatografi cair kinerja tinggi
Kromatografi cair kinerja tinggi atau High Performance Liquid
Chromatography (HPLC) merupakan suatu metode yang sensitif dan akurat untuk
penentuan kuantitatif serta baik untuk pemisahan senyawa yang tidak mudah

16
menguap. Pemisahan dengan HPLC mempunyai beberapa keuntungan
dibandingkan dengan metode konvensional seperti waktu analisis yang cepat,
biaya yang rendah dan kemungkinan untuk menganalisis sampel yang tidak stabil
(Nurhamidah 2005). Komponen penyusun HPLC secara skematik dapat dilihat
pada Gambar 9.
Gambar 10 Skematik komponen HPLC (LC Resources Inc. 2001)
Mardiana dan Ramdani (2008) menjelaskan komponen HPLC yang terdiri
dari :
1) Tandon (Reservoir)
Reservoir terbuat dari gelas atau stainless stell. Jumlahnya bisa satu, dua
atau lebih. Reservoir yang baik disertai degassing system yang berfungsi untuk
menghilangkan gas-gas yang terlarut dalam solven. Gas terlarut tersebut antara
lain adalah oksigen. Degassing dilakukan dengan mengalirkan gas inert dengan
kelarutan yang sangat kecil.
2) Pompa
Fungsi pompa adalah untuk memompa fase gerak (solvent) ke dalam kolom
dengan aliran yang konstan dan reproducible.
3) Katup injektor
Bagian ini merupakan tempat dimana sampel diinjeksikan untuk selanjutnya
dibawa oleh fase gerak ke dalam kolom.
4) Kolom
Kolom merupakan jantung kromatografi. Berhasil atau tidaknya suatu
analisis tergantung pada pemilihan kolom dan kondisi percobaan yang sesuai.

17
Kolom dapat dibagi menjadi dua kelompok yaitu kolom analitik dan kolom
preparatif.
5) Detektor
Suatu detektor dibutuhkan untuk mendeteksi adanya komponen sampel di
dalam kolom (analisis kualitatif) dan menghitung kadarnya (analisis kuantitatif).
6) Recorder
Hasil pembacaan detektor kemudian diolah oleh suatu processor kemudian
dikirim ke recorder. Recorder akan membuat suatu tampilan kromatogram. Untuk
HPLC dilengkapi seperangkat software yang dapat menghitung luas kromatogram
dan bahkan sekaligus menghitung kadarnya.
2.8 Uji Kolorimetri ATPase
Penentuan aktivitas penghambatan RNA helikase HCV menggunakan uji
kolorimetri ATPase (Utama et al. 2000). Pengujian ini mengukur besar
penghambatan terhadap RNA helikase pada salah satu aktivitas enzimatiknya,
yaitu ATPase (RNA-stimulated ATPase). Penghambatan terhadap aktivitas
ATPase, secara tidak langsung juga menghambat aktivitas RNA helikase secara
keseluruhan, karena helikase membutuhkan energi yang dihasilkan dari hidrolisis
ATP untuk memisahkan untai ganda RNA (Hairany 2010).
Prinsip ujinya adalah pengukuran fosfat bebas/anorganik (Pi) yang
terbentuk dari hidrolisis ATP oleh RNA helikase. Fosfat bebas akan membentuk
kompleks warna dengan amonium molibdat membentuk fosfomolibdat.
Fosfomolibdat akan bereaksi dengan enzim RNA helikase sehingga protein akan
mengendap dan menimbulkan kekeruhan. Polivinil alkohol akan melarutkan
kembali protein yang mengendap sehingga tidak terjadi kekeruhan. Warna yang
terbentuk sebanding dengan konsentrasi fosfat bebas yang dihasilkan dari
hidrolisis ATP. Penghentian reaksi warna dengan penambahan Na-sitrat yang
dapat mencegah pembentukan warna yang berlebih (Chan et al. 1986).