Embed Size (px)
DESCRIPTION
Citation preview

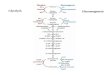
Pentose Phosphate Pathway
Three main functions:
1) Supply the cell with NADPH in order to:
a) provide reducing power for biosynthetic reactions.
b) serve as a biochemical reductant (e.g., maintain glutathione levels).
c) be utilized by the cytochrome P450 monooxygenase system.
d) as the electron source for reduction of ribo- to deoxyribonucleotides
for DNA synthesis.
2) Convert hexoses into pentoses (which are essential components of
ATP, CoA, NADP+, FAD, RNA, and DNA).
3) Enable the complete oxidative degradation of pentoses by
converting them into hexoses and trioses which can then enter
the glycolytic pathway.

The Roles of NADH and NADPH in Metabolism
There is a fundamental distinction between NADH and NADPH in
most biochemical reactions.
NADH is oxidized by the electron transport chain to generate ATP.
In contrast, NADPH functions as an electron donor (i.e., a hydride
ion donor) in biosynthetic reactions.
Recall that in the oxidation of a substrate, the nicotinamide ring of
NADP+ accepts a hydrogen ion and two electrons, which are
equivalent to a hydride ion.


Stages of the Pentose Phosphate Pathway
Stage 1: consists of the oxidative portion of the pathway in which
two oxidative reactions provide NADPH and a hexose is decarboxylated
to a pentose.
Stage 2: consists of two reversible isomerization reactions.
Stage 3: consists of the nonoxidative portion of the pathway in which
via a series of interconversions of three-, four-, five-, six-, and
seven-carbon sugars, excess pentoses are converted to hexoses
and trioses which can enter the glycolytic pathway.

Stage 1: Three reactions constitute this stage, two of which are
oxidative and generate NADPH.
Stage 1 is linked to biosynthetic reactions since NADPH and a
pentose are produced.
The reactions of stage 1 can be summarized as follows:
Glucose 6-phosphate + 2 NADP+ + H2O
ribulose 5-phosphate + 2 NADPH + 2 H+ + CO2
Thus two of the three functions of the pentose phosphate pathway are
accomplished: generation of NADPH and conversion of a hexose to a
pentose.
Glucose 6-phosphate
dehydrogenaseLactonase
6-phosphogluconate
dehydrogenase1
1
3
oxidative
decarboxylation

Stage 2 consists of two reversible isomerization reactions which convert
ribulose 5-phosphate into either ribose 5-phosphate or xylulose
5-phosphate.
Both are substrates for the Stage 3 reactions.
Ribulose 5-phosphate can also be isomerized to xylulose 5-phosphate
via the enzyme phosphopentose epimerase.
Phosphopentose isomerase
ketose
aldose

The Stage 1 + Stage 2 reactions yield 2 NADPH and 1 ribose
5-phosphate for each glucose 6-phosphate oxidized.
However, cells often need NADPH reducing power more than they
need ribose 5-phosphate for nucleotide biosynthesis.
In these cases, ribose 5-phosphate is further converted into
glyceraldehyde 3-phosphate and fructose 6-phosphate by the
enzymes transketolase and transaldolase.
These enzymes created a reversible link between the pentose
phosphate pathway and glycolysis.

Reactions of Stage 3
Stage 3 consists of non-oxidative reactions which link the pentose
phosphate pathway with glycolysis.
This stage allows:
1) excess pentoses to be converted to hexoses and trioses which can
then enter glycolysis; and
2) hexoses to be converted to pentoses, thereby allowing pentose
production without concomitant production of NADPH.
Two enzymes – transketolase and transaldolase – catalyze a series of
three reactions which convert 3 pentoses into 2 hexoses and 1 triose.
These reactions involve interconversions of 3, 4, 5, 6, and
7-carbon sugars.

Transketolase transfers a 2-carbon fragment.
Transaldolase transfers a 3-carbon fragment.
C5 + C5 C3 + C7
C7 + C3 C4 + C6
C5 + C4 C3 + C6
Net: 3 C5 2 C6 + 1 C3
transketolase
transaldolase
transketolase
Glyceraldehyde
3-phosphate
Fructose
6-phosphate

Reaction 1
Two pentoses are required: ribose 5-phosphate and xylulose
5-phosphate.
A 2 carbon fragment is transferred from the ketose to the aldose.
Catalyzed by transketolase.
Ketose
Aldose
C5 C5 C3 C7
Transketolase contains tightly bound TPP as its prosthetic group.
Wernicke Kosakoff Syndrome is an autosomal recessive disorder caused by
an alteration in transketolase which reduces its affinity for TPP.
Symptoms only develop if individual suffers from a moderate thiamine deficiency.

Reaction 2
The products of Reaction 1 (i.e., glyceraldehyde 3-phosphate and
sedoheptulose 7-phosphate) are the substrates for Reaction 2.
A 3-carbon unit is transferred from the ketose to the aldose by the
enzyme transaldolase.
C7 C3 C4 C6

Transketolase is also utilized for the third reaction.
A 2-carbon unit is transferred from xylulose 5-phosphate (a ketose) to
erythrose 4-phosphate (an aldose).
Note: the products of this reaction, glyceraldehyde 3-phosphate and
fructose 6-phosphate, are both intermediates of the glycolytic
pathway.
C5 C4 C3 C6

The sum of the Stage 3 reactions is:
2 Xylulose 5-phosphate + ribose 5-phosphate
2 fructose 6-phosphate + glyceraldehyde 3-phosphate
If we include the Stage 2 isomerization reactions, the net reaction is:
3 Ribose 5-phosphate
2 fructose 6-phosphate + glyceraldehyde 3-phosphate
The important point is that excess ribose 5-phosphate formed by the
pentose phosphate pathway can be completely converted into
glycolytic intermediates.

The Rate Limiting Step of the Pentose Phosphate Pathway
The first reaction in the oxidative branch of the pentose phosphate
pathway catalyzed by glucose 6-phosphate dehydrogenase, is the
rate limiting step under physiological conditions.
NADPH is a potent competitive inhibitor of the enzyme. Thus, the ratio
of NADP+/NADPH regulates the pathway.
As the NADP+ level rises, the flux thru the pathway increases.
Note: The nonoxidative branch of the pathway is regulated primarily by
substrate availability.

The Flow of Glucose 6-phosphate Depends on
Physiological Need
The flow of glucose 6-phosphate depends on cellular need for NADPH,
ribose 5-phosphate, and ATP. The Pentose Phosphate Pathway can
operate in 4 different modes.
Mode 1: Much more ribose 5-phosphate than NADPH is required.
Also need:
phosphopentose isomerase
phosphopentose epimerase
For example, rapidly dividing
cells need nucleotide precursors
for DNA synthesis more than
they need NADPH.
Glycolysis 2
1
3
transketolase
transaldolase
Bypass Stage 1, feed into
glycolysis instead.
Use glycolysis +
Stage 2 and Stage 3
reactions.

Mode 2: The needs for NADPH and ribose 5-phosphate are balanced.
Oxidative Branch – Stage 1
Phosphopentose
isomerase
The stoichiometry of mode 2 is:
Glucose 6-phosphate + 2 NADP+ + H2O
ribose 5-phosphate + 2 NADPH + 2 H+ + CO2

Mode 3: Much more NADPH than ribose 5-phosphate is required;
glucose 6-phosphate is completely oxidized to CO2.
Three sets of reactions are required under these conditions.
This situation typically occurs
in adipose tissue where a high
level of NADPH is required for
fatty acid biosynthesis.
The sum of these reactions is:
Glucose 6-phosphate + 12 NADP+ + 7 H2O
6 CO2 + 12 NADPH + 12 H+ + Pi
Oxidative Branch
Phosphopentose
isomerase
transketolase
transaldolase
Gluconeogensis
(1)
(2)
(2)
(3)
(2)
Regenerate glucose 6-phosphate
from the pentose.

Mode 4: Much more NADPH than ribose 5-phosphate is required;
glucose 6-phosphate is converted into pyruvate.
ATP, NADH, and NADPH
are generated.
Five of the six carbons of
glucose 6-phosphate emerge
as pyruvate.
The stoichiometry of mode 4 is:
3 Glucose 6-phosphate + 6 NADP+ + 5 NAD+ + 5 Pi + 8 ADP
5 pyruvate + 3 CO2 + 6 NADPH + 5 NADH + 8 ATP + 2 H2O + 8 H+
Oxidative Pathway
transketolase
transaldolaseGlycolysis
(1)
(2)(3)
ATP or Biosynthetic
precursors
Phosphopentose
isomerase

The Percentage of Glucose Metabolized by the Pentose
Phosphate Pathway Varies for Different Tissues
Since, a main purpose of this pathway is to supply NADPH for reductive
syntheses, it is prominent in tissues that actively carry out the reductive
synthesis of fatty acids and/or steroids from acetyl CoA.
Liver: 5-10% of glucose is metabolized by the pentose
phosphate pathway.
Adipose Tissue: 30 – 50%
Erythrocytes: 10% (need NADPH to maintain reduced glutathione)
Thyroid gland, kidney, and brain: 3 – 5%
Muscle: activity is extremely low.

A radioisotopic approach can be used to assess the fraction of glucose
metabolized by the pentose phosphate pathway vs. the sum of
glycolysis, PDH, + the citric acid cycle in a given tissue.
Principle: Only the C-1 position of glucose is decarboxylated by the
pentose phosphate pathway (at the 6-phosphogluconate dehydrogenase
step).
In contrast, the C-1 and C-6 positions are decarboxylated equally when
glucose is metabolized by the glycolytic pathway, PDH, and the citric
acid cycle.
This is because in these pathways, the C-1 and C-6 from glucose end up
as C-3 in pyruvate following the triose phosphate isomerase step. Thus,
subsequent oxidation by the citric acid cycle liberates 14CO2 equally from
the original C-1 and C-6 of glucose.

Thus, 2 samples of a given tissue are prepared:
One is incubated with glucose labeled with 14C at position C-1,
whereas the other sample is incubated with glucose labeled at
position C-6. The amount of radioactive 14CO2 produced by the 2
samples is then compared.
For example, in liver:
14CO2 liberated by the sample incubated in glucose-1-14C is produced
by both the citrate acid cycle and the pentose phosphate pathway;
However, less 14CO2 is liberated by the sample incubated in
glucose-6-14C because the citric acid cycle is the only source of14CO2.
By comparing the amount of 14CO2 released with different tissues
that are labeled in the C-1 versus C-6 positions, one can determine
the proportion of glucose metabolized by one pathway versus another.

Role of Glucose 6-phosphate Dehydrogenase in the
Red Blood Cell
In the RBC glucose serves as the primary energy source. RBC’s lack
mitochondria and thus lack the enzymes of the citric acid cycle.
Therefore, glucose is metabolized exclusively by the glycolytic
pathway (90%) and the pentose phosphate pathway (10%).
The most important function of the pentose phosphate pathway in the
RBC is to maintain the tripeptide glutathione in a reduced state.
Oxidized glutathione is reduced by the enzyme glutathione reductasein a reaction which utilizes NADPH:

Functions of Reduced Glutathione
1) To serve as a sulfhydryl buffer that maintains the cysteine residues
of hemoglobin and other RBC proteins in the reduced state.
2) To maintain the iron in hemoglobin in the ferrous (i.e., reduced)
form (Fe++).
3) To detoxify by reacting with hydrogen peroxides and organic
peroxides. This reaction is catalyzed by glutathione peroxidase.
H2O2 + 2 GSH 2 H2O + GSSG
Thus reduced glutathione is essential for maintaining the normal
structure of RBCs. Cells with a lowered level of this compound
are more susceptible to hemolysis.
Certain drugs act as oxidants formation of toxic peroxides
oxidation of proteins distortion of the surface of RBCs in the
absence of glutathione, making them more susceptible to destruction.


Glucose 6-phosphate Dehydrogenase Deficiency
•Glucose 6-phosphate dehydrogenase (G6PD) deficiency is an inherited
disease characterized by hemolytic anemia caused by an inability to
detoxify oxidizing agents.
•Most common disease-producing enzyme abnormality in humans.
•Caused by a family of over 400 point mutations in the enzyme.
•Only certain mutations cause clinical symptoms.
•Most affected individuals have no symptoms until they are exposed to
certain drugs which act as oxidizing agents.
•Drug exposure can induce a hemolytic episode which in some cases
can be fatal.
•Life-span of many individuals with G6PD deficiency is shortened due to
complications arising from chronic hemolysis.

Genetic Variants of Glucose 6-phosphate Dehydrogenase
More than 400 putative variants of the G6PD have been described.
Among the properties by which the variants can be distinguished
are the following:
1) enzyme activity
2) kinetic properties
3) electrophoretic mobility
4) substrate specificity
5) enzyme stability
6) pH optimum
The severity of the disease typically correlates with the amount of
residual enzyme activity in the patient’s RBCs.

•G6PD A- is the prototype of the moderate (class III) form of the disease.
•The A- variant occurs with high frequency amongst the African-American
population (gene frequency of the A- allele ~ 11%) and causes a
susceptibility to drug-induced hemolytic anemia.
•The enzyme encoded by the A- allele is kinetically normal, but displays
a substantially reduced half-life (i.e., 13 days versus 62 days).
Consequence: about 3 days after an oxidizing drug is administered to an
A- individual, there is a pronounced hemolytic episode. Older RBCs
which have little functional enzyme and hence are deficient in NADPH
and reduced glutathione are destroyed.
After ~ 1 week recovery begins since most of the remaining (i.e., younger
RBCs), as well as newly produced RBCs, have relatively normal enzyme
levels.
Provides protection against the drug-induced hemolysis.
Glucose 6-phosphate Dehydrogenase Deficiency

•The high frequency of the A- variant suggests that the deficiency may
be advantageous under certain environmental conditions.
•In fact it confers partial protection against malaria (since the causative
parasite requires reduced glutathione and pentose phosphate pathway
products).
•Illustrates the interplay between heredity and environment.
The G6PD Mediterranean variant is the prototype of a more severe
(class I) deficiency.
•Enzyme shows normal stability, but barely detectable activity in RBCs.
•A broader spectrum of drugs causes hemolysis and the hemolysis
is not as self limiting as in individuals with the A- variant.

Molecular Biology of Glucose 6-phosphate Dehydrogenase
•The cloning of the G6PD gene has enabled identification of mutations
that cause G6PD deficiency. All are point mutations in the coding
region of the gene.
•Mutations causing nonspherocytic hemolytic anemia: cluster near the
the NADP+ binding site.
•Mutations causing milder forms of the disease cluster near the glucose
6-phosphate binding site.

Organ Integration of Carbohydrate Metabolism
Liver: essential for providing fuel (i.e., glucose) to the brain, muscle,
and other peripheral organs.
The liver extracts Glucose from the blood.
Glycogen
Release glucose
when needed.
Pentose Phosphate
Pathway
NADPH for biosynthesis;
pentoses, hexoses.
Glycolysis
Mainly as a source
of biosynthetic
intermediates; also
to make some ATP
Note:
1) If there is insufficient glucose in the blood, the liver will break down its
glycogen and/or synthesize glucose via gluconeogenesis to increase
blood glucose supplied to other tissues.
2) Liver mainly uses α-keto acids derived from amino acid catabolism to supply
its own ATP needs.

Brain: Glucose is virtually the sole fuel source for the human brain,
except during times of starvation. In the resting state, the brain
accounts for 60% of glucose utilization.
•Glucose is obtained either from the diet or the liver.
•Brain does not contain significant stores of glycogen.
•Brain does not carry out gluconeogenesis since it has no glucose
6-phosphatase.
Muscle: In contrast to brain, muscle has a large store of glycogen
(i.e., ~ 75% of total body glycogen).
•Its major fuels are glucose, fatty acids, and ketone bodies.
•Like brain, muscle lacks glucose 6-phosphatase, and so it does not
carry out gluconeogenesis or export glucose.
•Instead muscle retains glucose, its preferred fuel, for bursts of activity.

Metabolic Interchanges Between Muscle and Liver

•During active contraction, glycolysis >> citric acid cycle. Therefore,
pyruvate is reduced to lactate which then flows to liver where it is
converted into glucose (Cori Cycle).
•Muscle also produces much alanine (by transamination of pyruvate).
This alanine can also be converted into glucose by the liver.
Adipose Tissue: Triacylglycerols stored in adipose tissue provide
an enormous reservoir of metabolic fuel.
•A principal function of this tissue is to synthesize triacylglycerols from
fatty acyl CoA derivatives and glycerol 3-phosphate.
•Thus, adipose cells need to metabolize glucose via glycolysis in order
to provide sufficient glycerol 3-phosphate (which originates from
dihydroxyacetone phosphate) for triacylglycerol synthesis.
•Also, adipose cells need to carry out some pentose phosphate pathway
in order to supply sufficient NADPH for synthesis.

Metabolic Relationships in the Well-Fed State