Embed Size (px)
Citation preview

5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 1/14
1
IL METABOLISMO CELLULARE
La composizione chimica della cellula è un dato certamente molto interessante, per esempio nelle
fermentazioni finalizzate alla produzione di proteine monocellulari. Ancor più interessante della
composizione è però il meccanismo con cui le sostanze costituenti la cellula sono sintetizzate, accumulate
ed eventualmente distrutte ossia il metabolismo cellulare.
Come è facile intuire il numero delle reazioni costituenti il metabolismo è elevatissimo. Per farsene
un'idea basta esaminare la composizione di uno degli organismi più semplici ( Escherichia coli) che
abbiamo visto precedentemente. Il numero dei tipi molecolari presentiè dell'ordine di 4-5.000 e per la
sintesi di ognuno di essi è necessaria almeno una reazione. Il quadro si complica ulteriormente se si
considera l'estrema varietà morfologica e quindi anche metabolica che differenzia una specie vivente
dall'altra.
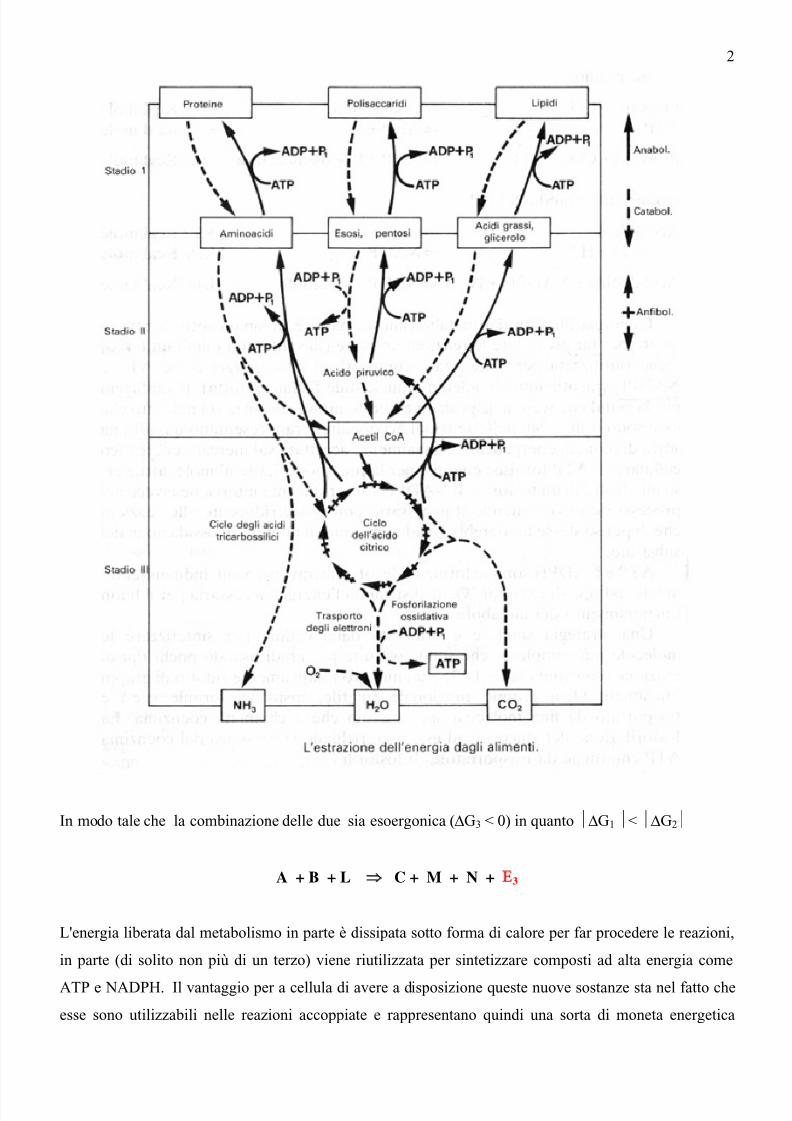
Sulla base di queste considerazioni lo studio per la conoscenza del metabolismo può apparire un'impresadisperata.Convenzionalmente il metabolismo è suddiviso in vie e cicli.
Un esempio di via è la fermentazione alcolica in cui il glucosio è trasformato in etanolo, un esempio di
ciclo è il ciclo dell'acido citrico in cui l'acido ossalacetico che reagisce con l'acetil coenzima A è
rigenerato alla fine del processo.
Si defisce catabolismo l'insieme di reazioni che conducono a una degradazione e anabolismo quelle
che portano a una sintesi.
Le prime reazioni normalmente sviluppano energia (sono cioè esoergoniche), le seconde ne assorbono e
sono endoergoniche.
Vie cataboliche e vie anaboliche sono collegate da cicli come quello dell'acido gliossilico o dell'acido
citrico che possono funzionare sia per completare la degradazione di piccole molecole derivanti dalle vie
cataboliche, sia per rifornire le vie anaboliche dei necessari precursori. Cicli di questo tipo.sono chiamati
anfibolici.
Pòichè la cellula ha bisogno di energia, essa deve essere alimentata con sostanze altamente energetiche da
cui o per degradazione anaerobia, con passaggio da una struttura più complessa a una più semplice e
quindi con un aumento dell'entropia, o per ossidazione con reazioni fortemente esoergoniche, tale energia
sia ottenibile.
Le reazioni cataboliche che sono esoergoniche procedono spontaneamente. Quelle anaboliche richiedono
un apporto di energia attraverso il meccanismo delle reazioni accoppiate. Una reazione non spontanea
endoergonica (∆G1 >0):
A + L + Ε1 ⇒ N anabolismo
Si può accoppiare con una reazione fortemente esoergonica (∆G2 < 0)
B ⇒ C + M + Ε2 catabolismo
5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 2/14
2
In modo tale che la combinazione delle due sia esoergonica (∆G3 < 0) in quanto ⏐∆G1 ⏐< ⏐∆G2⏐
A + B + L ⇒ C + M + N + Ε3
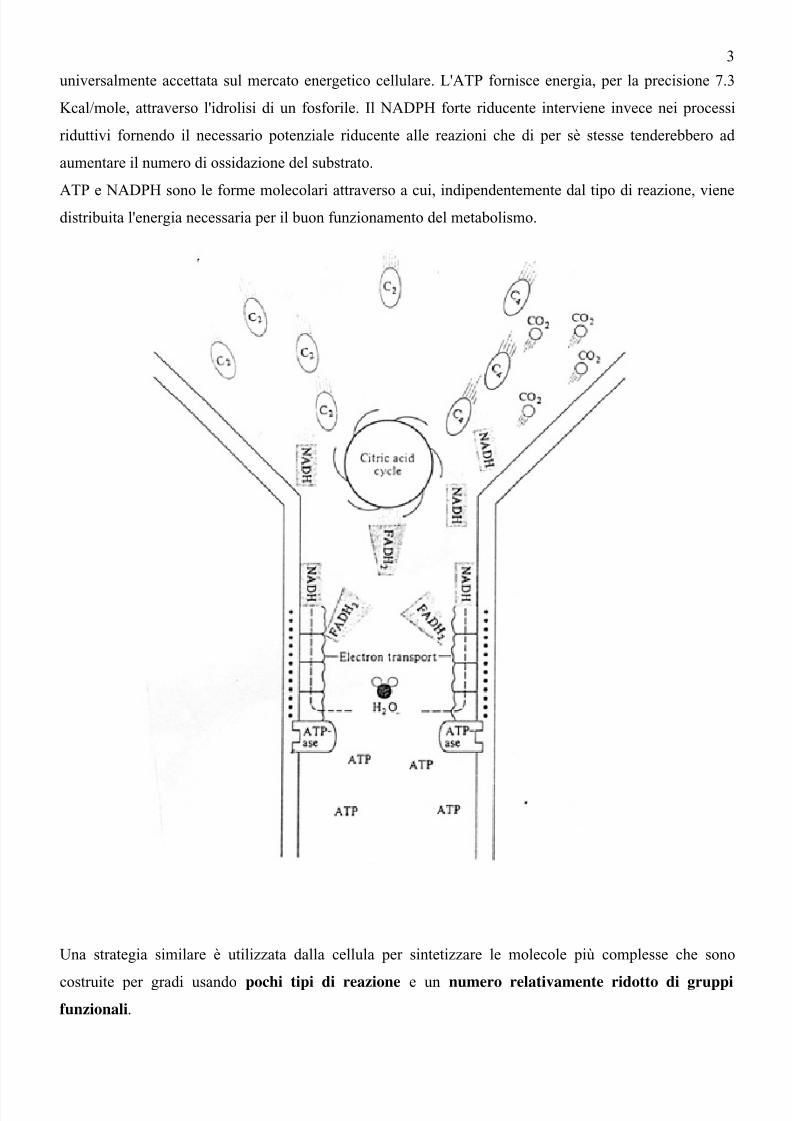
L'energia liberata dal metabolismo in parte è dissipata sotto forma di calore per far procedere le reazioni,
in parte (di solito non più di un terzo) viene riutilizzata per sintetizzare composti ad alta energia come
ATP e NADPH. Il vantaggio per a cellula di avere a disposizione queste nuove sostanze sta nel fatto che
esse sono utilizzabili nelle reazioni accoppiate e rappresentano quindi una sorta di moneta energetica
5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 3/14
3
universalmente accettata sul mercato energetico cellulare. L'ATP fornisce energia, per la precisione 7.3
Kcal/mole, attraverso l'idrolisi di un fosforile. Il NADPH forte riducente interviene invece nei processi
riduttivi fornendo il necessario potenziale riducente alle reazioni che di per sè stesse tenderebbero ad
aumentare il numero di ossidazione del substrato.
ATP e NADPH sono le forme molecolari attraverso a cui, indipendentemente dal tipo di reazione, vienedistribuita l'energia necessaria per il buon funzionamento del metabolismo.
Una strategia similare è utilizzata dalla cellula per sintetizzare le molecole più complesse che sono
costruite per gradi usando pochi tipi di reazione e un numero relativamente ridotto di gruppi
funzionali.
5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 4/14
4
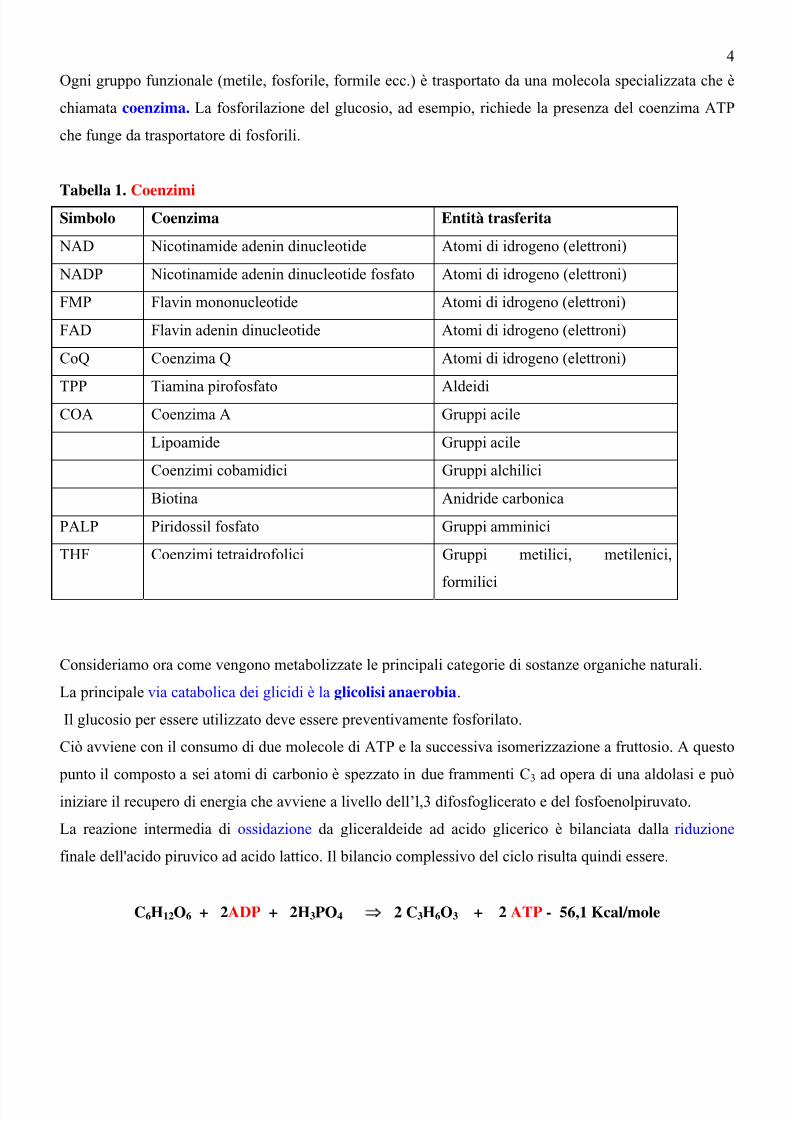
Ogni gruppo funzionale (metile, fosforile, formile ecc.) è trasportato da una molecola specializzata che è
chiamata coenzima. La fosforilazione del glucosio, ad esempio, richiede la presenza del coenzima ATP
che funge da trasportatore di fosforili.
Tabella 1. Coenzimi Simbolo Coenzima Entità trasferita
NAD Nicotinamide adenin dinucleotide Atomi di idrogeno (elettroni)
NADP Nicotinamide adenin dinucleotide fosfato Atomi di idrogeno (elettroni)
FMP Flavin mononucleotide Atomi di idrogeno (elettroni)
FAD Flavin adenin dinucleotide Atomi di idrogeno (elettroni)
CoQ Coenzima Q Atomi di idrogeno (elettroni)
TPP Tiamina pirofosfato AldeidiCOA Coenzima A Gruppi acile
Lipoamide Gruppi acile
Coenzimi cobamidici Gruppi alchilici
Biotina Anidride carbonica
PALP Piridossil fosfato Gruppi amminici
THF Coenzimi tetraidrofolici Gruppi metilici, metilenici,
formilici
Consideriamo ora come vengono metabolizzate le principali categorie di sostanze organiche naturali.
La principale via catabolica dei glicidi è la glicolisi anaerobia.
Il glucosio per essere utilizzato deve essere preventivamente fosforilato.
Ciò avviene con il consumo di due molecole di ATP e la successiva isomerizzazione a fruttosio. A questo
punto il composto a sei atomi di carbonio è spezzato in due frammenti C3
ad opera di una aldolasi e può
iniziare il recupero di energia che avviene a livello dell’l,3 difosfoglicerato e del fosfoenolpiruvato.
La reazione intermedia di ossidazione da gliceraldeide ad acido glicerico è bilanciata dalla riduzione
finale dell'acido piruvico ad acido lattico. Il bilancio complessivo del ciclo risulta quindi essere.
C6H12O6 + 2ADP + 2H3PO4 ⇒ 2 C3H6O3 + 2 ATP - 56,1 Kcal/mole

5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 5/14
5

5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 6/14
6
Questa via metabolica non richiede ossigeno ed è comune sia agli organismi aerobi che a quelli
anaerobi.
Probabilmente è una delle prime vie metaboliche elaborate dalla cellula vivente nel corso della sua
evoluzione. Il suo guadagno energetico è piuttosto basso. Si formano due molecole di ATP, pari a 15
Kcal/mole e il resto va disperso sotto forma di calore. La resa complessiva risulta quindi del 30%.La fermentazione alcolica può essere considerata una variante della glicolisi da cui differisce solo per le
due ultime reazioni. I prodotti ultimi della via metabolica sono l'alcol etilico e l'anidride carbonica.
Una via alternativa alla glicolisi è il ciclo dei pentosi che parte dal glucosio 6-fosfato e procede secondo
lo schema sotto riportato.
Ciclo dei pentosi
A differenza della via precedente che è anaerobia, il ciclo dei pentosi si sviluppa attraverso due fasi. La
prima è ossidativa e, oltre ai prodotti ossidativi, produce NADPH che serve alla cellula per le reazioni di
riduzione. Il ciclo ha una funzione fondamentale sia per questo motivo, sia per la sintesi e il catabolismo
dei pentosi che, come componenti degli acidi nucleici, sono presenti in ogni cellula. Come bilancio
complessivo della materia, il ciclo consuma 1/6 del carbonio glicidico con la formazione di CO2 e
NADPH.

5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 7/14
7
6 Glucoso-6-P + 12 NADP+ ⇒ 5 Glucoso-6-P + 6 CO2 + 12 NADPH + 12 H
++ H3PO4
La maggior parte dei microrganismi utilizzati nelle fermentazioni è di tipo aerobio e di conseguenza il
metabolismo è ossidativo e i suoi prodotti ultimi sono anidride carbonica e acqua. In questo tipo diorganismi il piruvato prodotto dalla glicolisi è dapprima decarbossilato ad acetilcoenzima A e sotto questa
forma è poi inserito nel ciclo dell'acido citrico o ciclo di Krebs.
Decarbossilazione ossidativa del piruvato
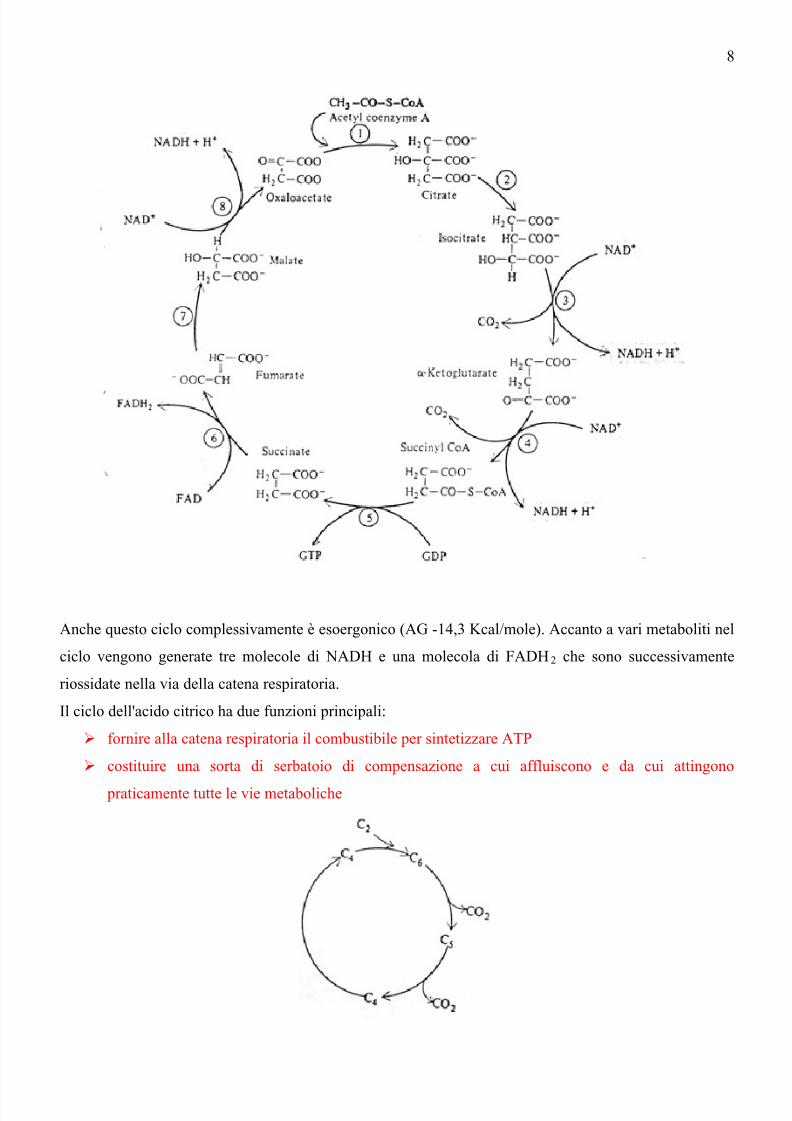
Schematicamente un'unità C2 (acetil-coenzima A) viene unita ad un'unità C4 (acido ossalacetico) per dare
l'acido citrico C6. Questo è successivamente decarbossilato ad acido α-chetoglutarico (C5) prima e ad
acido succinico poi (C4). L'acido succinico è infine ossidato ad acido ossalacetico e il ciclo riprende
dall'inizio.
5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 8/14
8
Anche questo ciclo complessivamente è esoergonico (AG -14,3 Kcal/mole). Accanto a vari metaboliti nel
ciclo vengono generate tre molecole di NADH e una molecola di FADH2 che sono successivamente
riossidate nella via della catena respiratoria.
Il ciclo dell'acido citrico ha due funzioni principali:
fornire alla catena respiratoria il combustibile per sintetizzare ATP
costituire una sorta di serbatoio di compensazione a cui affluiscono e da cui attingono
praticamente tutte le vie metaboliche
5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 9/14
9
Attraverso a questo ciclo è quindi possibile l'interconversione ad esempio di un eccesso di glicidi in altri
metaboliti come ad esempio i lipidi.
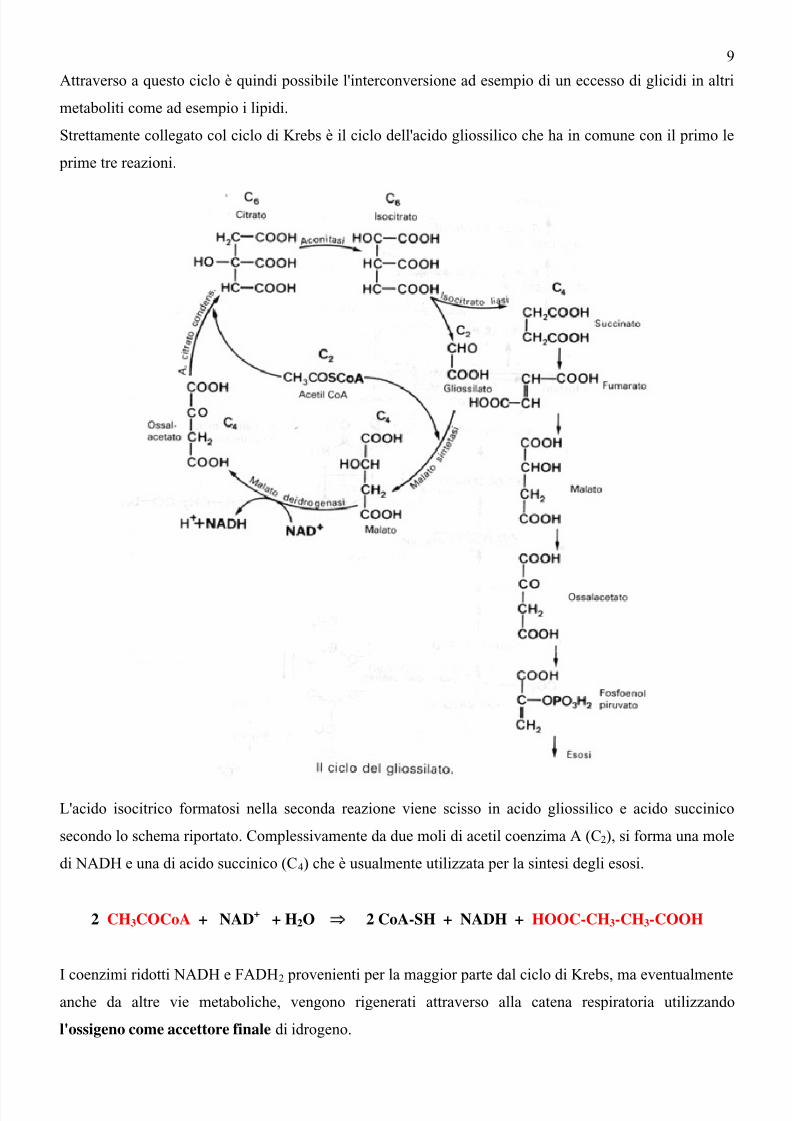
Strettamente collegato col ciclo di Krebs è il ciclo dell'acido gliossilico che ha in comune con il primo le
prime tre reazioni.
L'acido isocitrico formatosi nella seconda reazione viene scisso in acido gliossilico e acido succinico
secondo lo schema riportato. Complessivamente da due moli di acetil coenzima A (C2), si forma una mole
di NADH e una di acido succinico (C4) che è usualmente utilizzata per la sintesi degli esosi.
2 CH3COCoA + NAD+
+ H2O ⇒ 2 CoA-SH + NADH + HOOC-CH3-CH3-COOH
I coenzimi ridotti NADH e FADH2 provenienti per la maggior parte dal ciclo di Krebs, ma eventualmenteanche da altre vie metaboliche, vengono rigenerati attraverso alla catena respiratoria utilizzando
l'ossigeno come accettore finale di idrogeno.

5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 10/14
10
Essi trasportano ioni idrogeno ed elettroni e viene prodotta energia quando si combinano con l’ossigeno
per dare acqua.
4 H+
+ 4 e-
+ O2 → H2O
L'energia liberata è in parte recuperata sotto forma di ATP nelle fosforilazioni ossidative. La catena
respiratoria e la fosforilazione ossidativa sono catalizzate da enzimi di membrana che negli organismi
eucarioti è la membrana del mitocondrio, in quelli procarioti la membrana cellulare. Questa semplice
reazione avviene in molti stadi. Gli enzimi sono sistemati in ordine di affinità crescente per gli elettroni,
così questi scorrono attraverso il sistema enzimatico.
La maggior parte del fabbisogno energetico della cellula è fornito da queste reazioni.
Le vie metaboliche finora descritte portano alla demolizione dei glicidi.
In condizioni favorevoli questi composti possono anche essere sintetizzati a partire da composti più
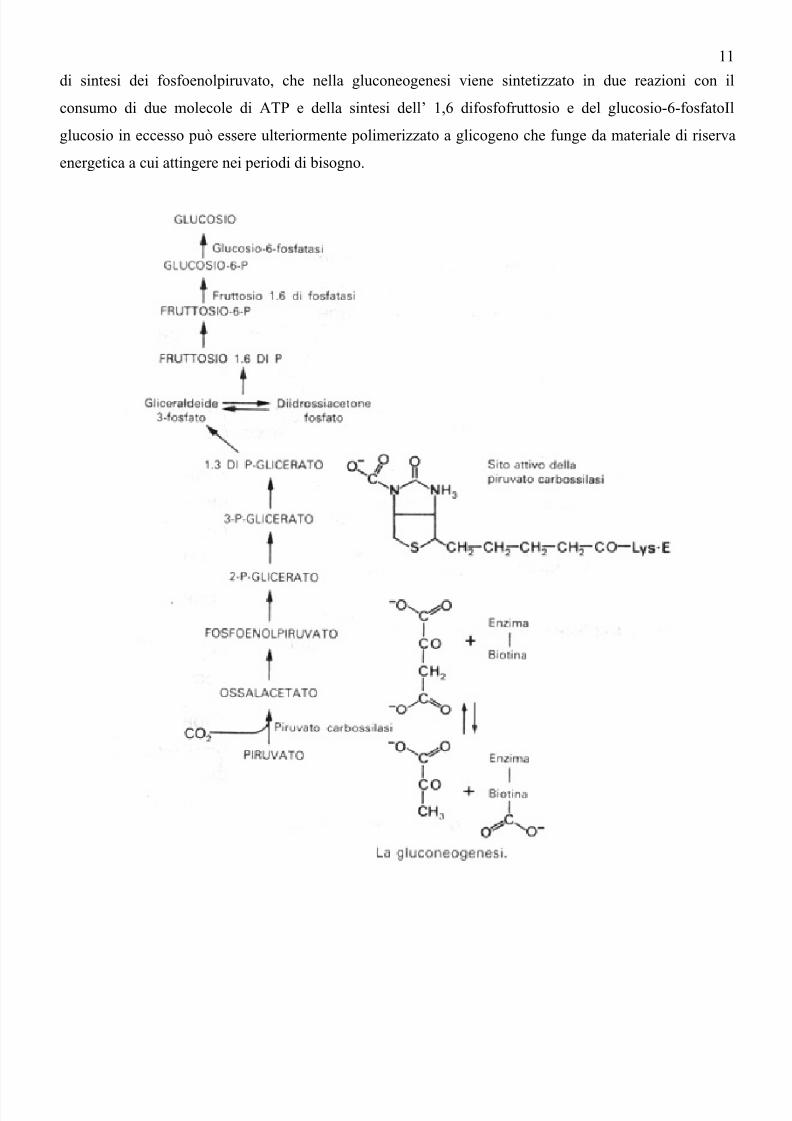
semplici. Una via molto importante è la gluconeogenesi che per le reazioni meno esoergoniche ripercorre
in senso inverso la glicolisi, discostandosene per le reazioni in cui questa via produce ATP e cioè quella
5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 11/14
11
di sintesi dei fosfoenolpiruvato, che nella gluconeogenesi viene sintetizzato in due reazioni con il
consumo di due molecole di ATP e della sintesi dell’ 1,6 difosfofruttosio e del glucosio-6-fosfatoIl
glucosio in eccesso può essere ulteriormente polimerizzato a glicogeno che funge da materiale di riserva
energetica a cui attingere nei periodi di bisogno.
5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 12/14
12
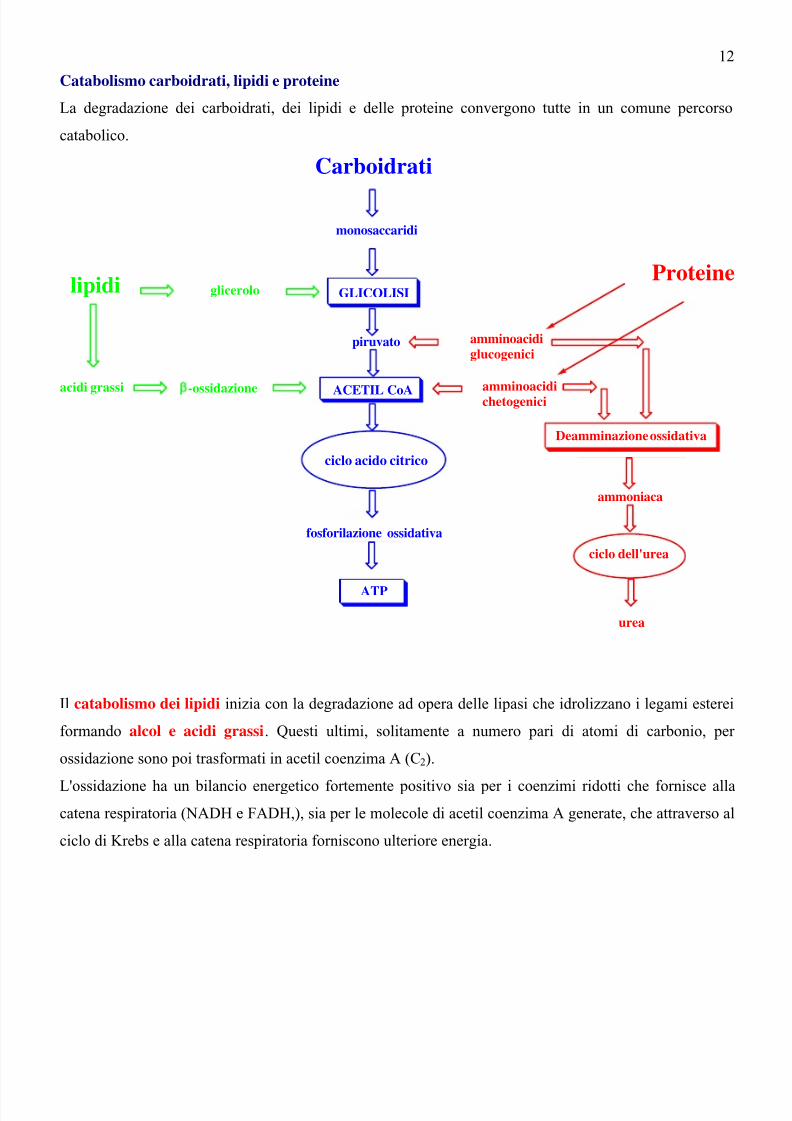
Catabolismo carboidrati, lipidi e proteine
La degradazione dei carboidrati, dei lipidi e delle proteine convergono tutte in un comune percorso
catabolico.
Carboidrati
monosaccaridi
GLICOLISI
piruvato
ACETIL CoA
ciclo acido citrico
fosforilazione ossidativa
ATP
glicerolo
acidi grassi β-ossidazione
lipidiProteine
amminoacidiglucogenici
amminoacidichetogenici
Deamminazione ossidativa
ammoniaca
ciclo dell'urea
urea
Il catabolismo dei lipidi inizia con la degradazione ad opera delle lipasi che idrolizzano i legami esterei
formando alcol e acidi grassi. Questi ultimi, solitamente a numero pari di atomi di carbonio, per
ossidazione sono poi trasformati in acetil coenzima A (C2).
L'ossidazione ha un bilancio energetico fortemente positivo sia per i coenzimi ridotti che fornisce alla
catena respiratoria (NADH e FADH,), sia per le molecole di acetil coenzima A generate, che attraverso al
ciclo di Krebs e alla catena respiratoria forniscono ulteriore energia.
5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 13/14
13
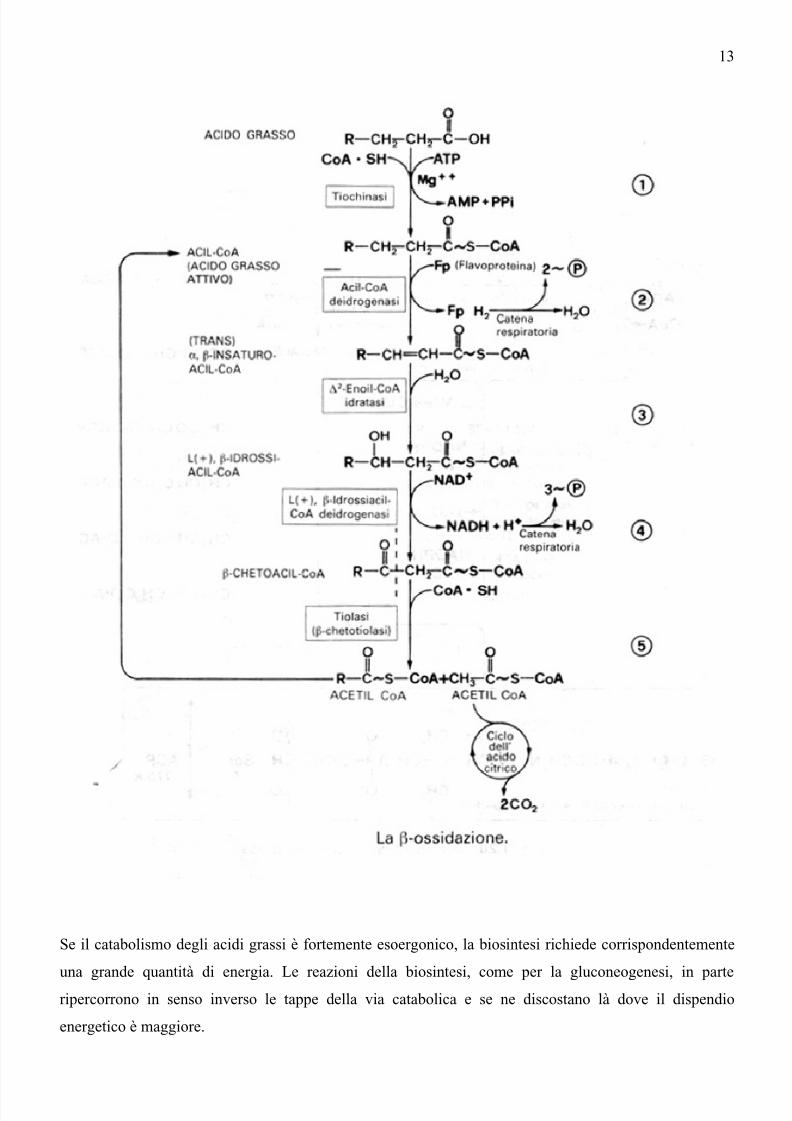
Se il catabolismo degli acidi grassi è fortemente esoergonico, la biosintesi richiede corrispondentemente
una grande quantità di energia. Le reazioni della biosintesi, come per la gluconeogenesi, in parteripercorrono in senso inverso le tappe della via catabolica e se ne discostano là dove il dispendio
energetico è maggiore.

5/15/2018 A Metabolismo Cellulare - slidepdf.com
http://slidepdf.com/reader/full/a-metabolismo-cellulare 14/14
14
Il catabolismo dei protidi procede attraverso tre fasi. Nella prima il polimero è idrolizzato nei singoli
amminoacidi ad opera di una serie di proteasi con diversa specificità. Gli amminoacidi derivati
dall’idrolisi possono essere metabolizzati lungo due vie principali: la deamminazione, che porta
all’eliminazione dell’azoto mediante il ciclo dell’urea, e il metabolismo della parte carbonica, che entra
nel ciclo della’acido citrico.
La più importante via riguarda il metabolismo dell’azoto che passa per la transamminazione con l'acido
α-chetoglutarico e la successiva deamminazione ossidativa dell'acido glutammico così formatosi che
porta a ione ammonio e rigenera α-chetoglutarato.
Lo ione ammonio generato può poi essere allontanato tal quale (organismi ammoniotelici) oppure dopo
essere trasformato in. urea (organismi ureotelici) o in acido urico (organismi uricotelici). Lo scheletro
carbonioso rimasto dopo la desamminazione se è il caso viene opportunamente modificato ed introdotto
nelle vie metaboliche già esaminate.





























