Embed Size (px)
Citation preview

A Mid-aqueductal Ependymal Organ in the Brain of the Hyrax (Procavia capensis)
W. B. ,QUAY Department o f Zoology, University of California, Berkeley, California 94720
ABSTRACT Serial sections of the brains of four adult male hyraxes revealed a hitherto undescribed ependymal organ near the middle of the midbrain aque- duct. This structural modification of the dorsal wall of the aqueduct is separate and different from both the subcommissural organ and the posterior collicular recess. It is characterized by: (1) an elaborate system of ridges and crypts, (2) numerous, crowded and distinctive ependymocytes, (3) tissue zonation, and (4) numerous vessels with sleeves of primarily longitudinally arranged dial processes. Comparisons with other ventricular organs and possible functional relations are discussed.
The ependymal lining of the vertebrate brain’s ventricular system has had a rela- tively lower level of interest in comparison with most other brain structures. However, there is increasing evidence of regional specialization and functional significance of the ependymal regions in relation to neuroembryology, regeneration and special transport and secretory activities. Regional modifications of the ependyma and the associated adjacent tissues are being shown to be more than structural peculiarities, especially in the light of experimental studies and coordinated tracer methods and electron microscopy. Results both of classical light microscopical and newer experimental and electron microscopical investigations have served to define and emphasize a specific array of such ependy- mal circumventricular organs. Most of these occur within, or at the boundaries of, the third and fourth ventricles. None have been described heretofore from the mid- region of the mammalian cerebral aque- duct (aqueduct of Sylvius).
This report describes the occurrence, structure and composition of a mid-aque- ductal ependymal organ in brains of the rock hyrax, “cony” or dassie (Procavia capensis). It is shown to be structurally differentiated and spatially separated from the nearest known circumventricular or- gans, the subcommissural organ anteriorly and the posterior collicular recess pos- teriorly. Yet, its internal structure and
J. COMP. NEUR., 142: 249-256.
composition resemble at least partially and superficially those of certain other circum- ventricular organs.
MATERIALS AND METHODS Four mature males killed within one
and one-half hours after mid-day in De- cember and in the vicinity of Vrolijkheid, Robertson, South Africa, were used. The median region of each brain was fixed in 10% formal saline and later processed by routine methods for washing, dehydration through ethanols, clearing and embedding in paraplast. Serial sections 7 p in thick- ness were cut, transversely of two of the brains and sagittally of the other two. Every fifth slide of one transverse and one sagittal series was stained by the periodic acid-Schiff (PAS) method with acid alum hematoxylin as nuclear stain (Lillie, ’54) and the intervening slides by an adapta- tion of a modified Bodian protargol method for nerve fibers. Slides from the other transverse and sagittal series were stained in rotation through the series by the fol- lowing techniques: Ehrlichs acid alum hematoxylin and eosin Y (Bensley and Bensley, ’38), chrome alum hematoxylin and phloxine (Gomori, ’41), Einarson’s gallocyanin (Lillie, ’54) and Wolbach’s Giemsa variant at 60°C (Lillie, ’54). The last three of these procedures were usually preceded by an overnight treatment in Bouin’s fluid after deparaffrnization and hydration.
249

250 W. B. QUAY
RESULTS The size and characteristics of the mid-
aqueductal organ are essentially the same in the four specimens. Hence, in the de- scriptions that follow, no distinguishing or variational comments will be made con- cerning them.
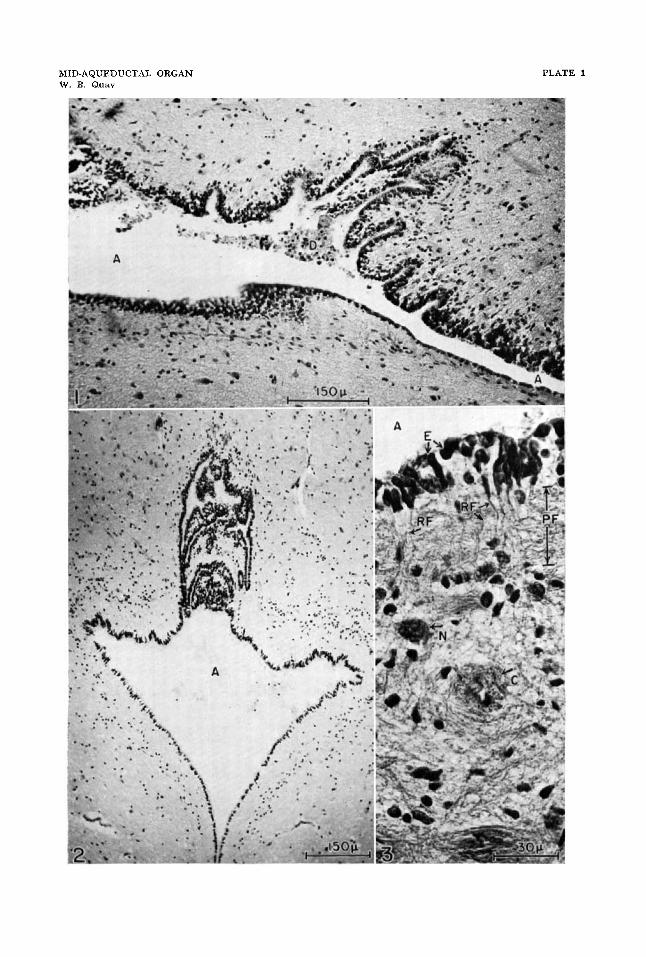
The most obvious feature of the organ is its localized system of slender transverse ridges and intervening crypts, which ex- tend about 300 p anterodorsally from the median dorsal wall of the aqueduct (figs. 1, 2). In the plane transverse to the axis of the midbrain, the organ is clearly me- dian and cannot be construed as a collicu- lar recess (fig. 2). In the sagittal plane in reference to the axis of the midbrain, the axis of the organ appears to be close to that of the paired superior colliculi. Along the longitudinal course of the aqueduct, the site of maximal dorsal projection of the organ is 4.0 mm anterior to the pos- terior base of the tectum and 2.3 mm pos- terior to the anterior limit of the tectum. It also lies 1.8 mm posterior to the pos- terior limit of the posterior commissure.
Maximum vertical and transverse inter- nal dimensions of the aqueduct are found at the level of the mid-aqueductal organ. As one progresses posteriorly along the aqueduct, the following changes are ob- served. At the posterior attenuation of the subcommissural organ the median vertical slit formed by the third ventricle has grad- ually shortened to form the median vertical slit forming the first part of the aqueduct. At the posterior end of the subcommissural organ the greatest transverse internal diam- eter of the aqueduct is about 30 p. This is its narrowest point. Posteriorly, it progres- sively widens to attain a horizontal dimen- sion in excess of 700 at the mid-aqueduc- tal organ. Further posteriorly and beyond the mid-aqueductal organ, it again narrows down to a vertical slit, about 100 in maximum diameter, before gradually wid- ening again as it passes into the fourth ventricle. The vertical extent of the aque- duct has a similar but less extreme pattern of change. Posterior to the subcommissural organ, it is about 900 p. At, and including, the crypts of the mid-acqueductal organ, it is about 1900 p. Posteriorly, it gradually shortens to 550 and then gradually
lengthens again as the fourth ventricle is approached.
Although at the region of maximum dorsal development of the mid-acqueductal organ it is composed of transverse ridges and crypts crossing the median or sagittal plane (figs. 1, 2) , anteriorly the ridges and crypts appear to be more longitudinal in orientation. It can be surmised from such sections as that in figure 2 that a U-shaped pattern of ridges and crypts may result where the longitudinal and transverse components meet in the midregion of the organ. The base of the U represents a transverse ridge or crypt, and the limbs represent bilateral longitudinal ridges or crypts extending anteriorly on either side of the sagittal plane. In both anterior and posterior directions from the organ’s mid- region, the geometry of the ependymally covered ridges is less regular, and an irreg- ular system of flattened projections or papillae more adequately characterizes their form.
Many of the ependymal cells of the hyrax’s aqueduct can be seen to be ciliated, even with the light microscope. It is not clear, however, to what extent ciliation continues along, or into, the crypts of the mid-aqueductal organ. Part of this observa- tional difficulty can be laid to the very nar- row lumina of the crypts and part to the frequent occurrence of flocculent extracel- lular debris within, and at, the mouths of the crypts (figs. 1, 2). Within the mid- aqueductal organ, the ependymal cells are characterized by their crowded appearance, often elongate nuclei, small amount of apical (luminal) cytoplasm and frequently pronounced, radially directed cytoplasmic processes. Extending deeper within the dorsal wall of the organ are other cells which resemble tanycytes or which appear to be transitional to such a category of cell. They, also, have prominent radial cyto- plasmic processes. The small amount of apical cytoplasm and its weak chromophila in the staining techniques used here dis- tinguish the epithelial or ependymal cells of the midequeductal organ from those of subcommissural organ in the same speci- mens.
Where ridges and crypts do not dissect it too greatly, the organ sometimes shows a tissue zonation in its dorsal wall (fig. 3).

MID-AQUEDUCTAL ORGAN 25 1
Beneath a superficial nuclear zone con- taining ependymal and transitional tany- cyte perikarya there is a zone composed mostly of parallel glial fibers. This is traversed by radial cytoplasmic processes from ependymocytes and tanycytes. A third and narrow zone is characterized by its greater concentration of perikarya of glia and small neurons. Along its peripheral edge capillaries are relatively numerous and course primarily longitudinally or par- allel to the surface and the sagittal plane. The fourth zone may be considered to start with these vessels and to be relatively the thickest zone and the spongiest one in general aspect. It contains scattered glial and medium-sized neuronal perikarya in an irregular, mostly glial fibrous mesh- work. The most striking feature of this zone, however, is its system of blood ves- sels, all with thick and continuous sleeves of glial processes, most of which run longi- tudinally in relation to the particular con- tained vessel. Except for the most super- ficial vessels of this zone, the direction followed by most of them is circumferen- tial and perpendicular in relation to the axis of the midbrain. These peculiar sleeved vessels, while somewhat reminiscent of some types of pathological or senile changes, appear to be nonpathological in origin and characterize all four specimens. Although most extensive in the mid-aque- ductal organ, they can be found also in the same general tissue zone through the length of the tectum near the mid-line and the aqueduct. In no other parts of the brains of these animals were such sleeved vessels found.
The majority of the vessels within the mid-aqueductal organ and all of those in its more superficial regions are equivalent to capillaries in their calibre and inner wall structure. By the periodic acid-Schiff technique these vessels are not distinguish- able in structure from those lying in either adjacent or distant regions of midbrain neuropil. The concentration of small ves- sels per tissue volume does appear to be greater, however, within the mid-aqueduc- tal organ in comparison with such other parts of the midbrain.
Glial and ependymal cytoplasmic proc- esses in the mid-aqueductal organ were differentially stained most distinctly by
phloxine in the chrome hematoxylin and phloxine procedure following overnight treatment with Bouin's fluid. The radial processes of ependymocytes and tanycytes could often be traced to the vicinity of capillaries, neurons and more distant re- gions of tectal neuropil. However, desig- nations of the precise points of termination of such processes or fibers may not be re- liable. The periodic acid-SchifF method stained granular inclusions within some glia and perivascular cells. But, occur- rences were not distinguishable qualita- tively or quantitatively from what could be observed in some other periventricular re- gions of the brains of these animals. In other regards, the particular staining meth- ods used in this study failed to show any unusual cell contents or extracellular ma- terials within the mid-aqueductal organ. The protargol method was successful in staining nerve fibers through the brains, but within the mid-aqueductal organ these were relatively fine in diameter and appar- ently few in number. Such an appraisal is probably premature, however, without the use of electron microscopy.
DISCUSSION
Examination both of serial sections of the brains of mammals of other Orders (Marsupialia, Insectivora, Chiroptera, Ro- dentia and Primates) in the author's col- lection and of published descriptions and figures of mammalian midbrain structure (Huber et al., '43; and others) has failed to reveal anything comparable to what is designated here as the mid-aqueductal organ. Although common and widespread in parts of Africa and Asia Minor, hyraxes or conies have been studied anatomically or physiologically very little. The living members of this group form a single family (Procaviidae) and Order (Hyracoidea). The members of this Order, the hyracoids, are distinct and separate anatomically as a group as far back as the Lower Oligocene. They are often considered to be structur- ally the most generalized of the so-called subungulates (Romer, '66), but their spe- cific relations or phylogenetic affinities are uncertain (Simpson, '45). A distant rela- tion to the Proboscidea (elephants and allies) has been claimed occasionally. Pub- lished descriptions of midbrain structure

252 W. B. QUAY
in elephants (Prgcechdl, ’25; and others) have failed to note or illustrate any aque- ductal peculiarities.
Superficial structural similarities may be seen between the hyrax mid-aqueductal organ and the dorsal walls of the tectal recesses in some lower vertebrates, as for example in some amphibians and reptiles (Paul, ’67; Campos-Ortega, ’65). This may be derived, however, from a surviving and primitive pattern of numerous radiating ependymal cells and tanycytes with alter- nating nuclear and fibrous circumferential zones through which the radiating cellular processes extend.
The mid-aqueductal organ is cytologi- cally and structurally different and topo- graphically separated from the subcommis- sural organ. This includes a recess within the subcommissural organ and often desig- nated as the mesocoelic recess (Dendy and Nicholls, ’10; Rakic, ’65). On cytological, structural and topographical grounds, the mid-aqueductal organ is also distinct and different from the structure called by Wis- lock and Leduc (’54) the posterior collicu- lar recess. This structure, also called “ven- triculus corporis quadrigemini posterioris” (Palmgren, ’21), has been illustrated, often adventitiously, especially in works on ro- dent neuroanatomy. A trace of it may be found in the hyrax brains.
The mid-aqueductal organ bears a closer cytological and structural resemblance to some of the circumventricular organs of the third and fourth ventricles; namely, the subfornical organ (or intercolumnar tubercle), the paraventricular organs, the infundibular recess and the area postrema. In all of these, in at least some species, there is : (1 ) cellular zonation, (2) epen- dymal and tanycyte modifications, (3 ) a bridging by these cells of the tissues be- tween the ventricular and vascular sys- tems, and ( 4 ) a greater vascularity (Hofer, ’59; Schimrigk, ’66; Leonhardt, ’66; Koella and Sutin, ’67; and others). The extent to which these similarities indicate in more exacting terms common features in cellu- lar specialization and functional meaning remains to be determined. Nevertheless, in the mid-aqueductal organ the increased ventricular surface area formed by epen- dymal ridges and crypts and the abundance of ependymal and tanycyte links from ven-
tricle to vessels are suggestive of an enhanced transport system between cere- brospinal fluid and blood. The common finding of masses of luminal degenerating cellular and flocculent debris attached to the organ (fig. 1) may be suggestive, also, of some sort of special activity. Pyknotic nucleated cells within these masses clearly antedate the death of the animal, but the exact sources of these cells and whether their location in the slides represents truly the relations in vivo cannot be decided with certainty.
ACKNOWLEDGMENTS
I am grateful to Bob Millar, senior officer, Predator Control Research Farm, Department of Nature Conservation, Pro- vincial Administration of the Cape of Good Hope, for the specimens; to Victor Gin and Stephen Chun for laboratory assistance; to James Hendel and Emily Reid for profes- sional assistance with the illustrations; and to Helen Sherry for bibliographic and typist services.
LITERATURE CITED Bensley, R. R., and S . H. Bensley 1938 Hand- book of Histological and Cytological Technique. Univ. of Chicago Press, Chicago, 162 pp.
Campos-Ortega, J. A. 1965 Aportaciones a la organizacih de las cubiertas ventriculares del enckfalo de 10s reptiles, referidas a 10s llama- dos “6rganos circumventriculares.” Anales de Anat., 14: 171-216.
Dendy, A,, and G. E. Nicholls 1910 On the occurrence of a rnesocoelic recess in the human brain, and its relation to the sub-commissural organ of lower vertebrates; with special refer- ence to the distribution of Reissner’s fibre i n the vertebrate series and its possible function. Proc. Roy. SOC. London, Ser. B, 82: 515-529.
Gomori, G. 1941 Observations with differential stains on human islets of Langerhans. Am. J. Path., 17: 395-406.
Hofer, H. 1959 Zur Morphologie der circum- ventrikularen Organe des Zwischenhirnes der Saugetiere. 2001. Anz., Verh. Deut. 2001. Ges., Suppl. 22: 202-251.
Huber, G. C., E. C. Crosby, R. T. Woodburne, L. A. Gillilan, J. 0. Brown and B. Tamthai 1943 The mammalian midbrain and isthmus regions. I. The nuclear pattern. J. Comp. Neur., 78:
Koella, W. P., and J. Sutin 1967 Extra-blood- brain-barrier brain structures. Intern. Rev. Neurobiol., 10: 31-55.
Leonhardt, H. 1966 Uber ependymale Tanycy- ten des 111. Ventrikels beim Kaninchen in elek- tronenmikroskopischer Betrachtung. Z. Zell- forsch., 74: 1-11.
129-534.

MID-AQUEDUCTAL ORGAN 253
Lillie, R. D. 1954 Histopathologic Technic and Practical Histochemistry. Blakiston Co., New York, x + 501 pp.
Palmgren, A. 1921 Embryological and mor- phological studies on the midbrain and cere- bellum of vertebrates. Acta Zool., 2: 1-94.
Paul, E. 1967 Uber die Typen der Ependym- zellen und ihre regionale Verteilung bei Rana temporaria L. Mit Bernerkungen iiber die Tany- cytenglia. 2. Zellforsch., 80: 461-487.
Pr&cechtgl, A. 1925 Some notes upon the finer anatomy of the brain stem and basal ganglia of Elephas indicus. Proc. Kon. Akad. Wetensch. te Amsterdam, Sec. Sci., 28: 81-93.
Rakic, P. 1965 Mesocoelic recess in the human brain. Neurology, 15: 708-715.
Romer, A. S. 1966 Vertebrate Paleontology. Third edition. Univ. of Chicago Press, Chicago, x + 468 pp.
Schimrigk, K. 1966 Uber die Wandstruktur der Seitenventrikel und des dritten Ventrikels beim Menschen. Z. Zellforsch., 70: 1-20.
The principles of classifica- tion and a classification of mammals. Bull. Am. Mus. Nat. Hist., 85: i-xvi+ 1-350.
Wislocki, G . B., and E. H. Leduc 1954 The cytology of the subcommissural organ, Reiss- ner’s fiber, periventricular glial cells and pos- terior collicular recess of the rat’s brain. J. Cornp. Neur., 101: 283-309.
Simpson, G. G. 1945

PLATE 1
EXPLANATION O F FIGURES
1 Sagittal section of mid-aqueductal organ stained with chrome hema- toxylin and phloxine. Anterior is toward the right in the figure.
2 Transverse section of mid-aqueductal organ stained with PAS and acid alum hematoxylin.
3 Small vertically cut region from the dorsal wall of a sagittally sec- tioned mid-aqueductal organ stained with chrome hematoxylin and phloxine.
Abbreviations: A, aqueduct of Sylvius; C, capillary with sleeve of longi- tudinal neuroglial fibers; D, cellular debris attached to, and associated with, the aqueductal organ; E, perikarya of ependymal cells; N, neuron perikaryon; PF, zone of parallel neuroglial fibers; RF, radial ependymal and tanycyte processes.
254
MID-AQUEDUCTAL ORGAN W. B. Quay
PLATE 1



















![Cerebrospinal Fluid Research BioMed Central...[4,5]. Analogous to these observations in laboratory ani-mals, Sarnat reported presence of ependymal denudation in human hydrocephalic](https://img.pdfslide.net/doc/110x75/60bd59d7dc03be6c0f35bf24/cerebrospinal-fluid-research-biomed-central-45-analogous-to-these-observations.jpg)