Embed Size (px)
Citation preview

BIOCHIMICA ET BIOPHYSICA ACTA 137
BBA 46104
ADENOSINE T R I P H O S P H A T E CELLULAR LEVELS IN RHODOSPIRILLUM
RUBRUM DURING TRANSITION FROM AEROBIC TO ANAEROBIC
METABOLISM
j . OELZE AND M. D. KAMEN
Department of Chemistry, University of California, San Diego, La Jolla, Calif. 92037 (U.S.A.)
(Received November 2nd, 197 o)
SUMMARY
I. Cellular ATP levels were investigated in Rhodospirillum rubrum under anaerobic and aerobic conditions in the light. All experiments were done with orga- nisms precultured in the light.
2. After a short t ime of decrease the ATP content increased under aerobic light conditions. No bacteriochlorophyll was formed in the growing cultures.
3. In cultures which had been grown in the presence of air and light the ATP level dropped upon replacement of air by nitrogen. This occurred only if the cells exhibited a low bacteriochlorophyll content, in which case pigment synthesis started.
4- A decrease of the cellular ATP content was also measurable when cells were grown anaerobically in the light. This was dependent on low bacteriochlorophyll content and new synthesis of bacterioehlorophyll.
5- Correlation of the ATP level and the various synthetic activities, e.g. bacterio- chlorophyll formation, necessary for the synthesis of the photosynthetic apparatus is discussed.
INTRODUCTION
Under anaerobic conditions in the light, or under low oxygen tension in the dark, most members of the Athiorhodaceae form the photosynthetic apparatus 1. This means that the intracellular membrane system is differentiated and extended 2-5. The whole process involves considerable metabolic act ivi ty and energy conversion.
Many investigations concerning a regulatory mechanism for the formation of photosynthetic active structures are recorded. Many authors suggest that the redox potential within a cell might possibly be a regulatory factor6, 7. Another hypothesis concerns involvement of ATP, ADP and AMP levels 8-1°. Data are available which support a correlation between the induction of synthesis of the photosynthetic membrane and the ATP level. ATP has been shown to have an inhibitory effect on tetrapyrrol synthesis TM 12 as well as on photosynthetic growth and bacteriochlorophyll synthesis 13. Recently it has been demonstrated that in Rhodopseudomonas spheroides, strain Y, a peak of cellular ATP content just precedes synthesis of bacterioehloro- phyl114.
Biochim. Biophys. Acta, 234 (1971) 137-143
I3b J. OELZE, M. D. KAMEN
In this report we describe the variation of the ATP levelin Rhodospirillum rubrum investigated under conditions which allow synthesis of the photosynthetic apparatus. It has been shown previously a that this synthesis requires the formation of several membrane components, e.g. proteins and different pigments. It seems plausible, therefore, that the high energy requirement for these different synthetic activities should influence the cellular ATP content.
MATERIALS AND METHODS
R. rubrum strain SI and FRI were cultivated under photosynthetic conditions in screw cap bottles. Every 24 h aliquots of the cultures were transferred to fresh modified Hutner medium ]5.
For the various experiments z4-I6-h-old cultures were inoculated into 2 1 cultural medium to an absorbanee of 0.03-0.04 at 12oo nm in a I-cm cuvette. All experiments were performed in a culture vessel which could be stirred continuously. The light intensity measured approximately at the center of the culture vessel, by means of a Kettering Radiometer, Model 68, was about 12.Io S ergs/cm 2 per sec before inoculation and IO.lO S ergs/em 2 per sec at tile initial time of inoculation.
I
?.2 F
LC-
0 . 8 -
0 0 , 6 - × E
0 .4
I
o-it
3.4A- - t18
2.6 ,, .,.g'" ) l / 4 . . . . . . ~--- ' : , - - - ~--7" . . . . . . ' l ' - - - - - - i ~ , i /
o o /
E 2.2 I I I
1.0 I ]'6 0 I 2 3 4 5 6
T ime ( h )
1.2
1.0
E :).8 g e~
::).6 ~
b 2
o
0.2
0
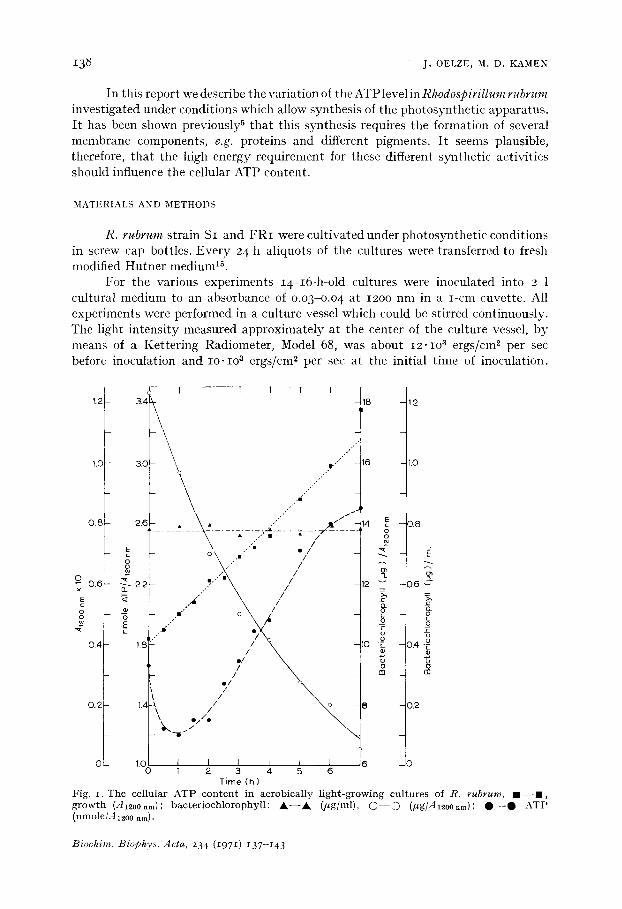
Fig. 1. The cellular AT P con t en t in aerobical ly l ight -growing cul tures of R. rubrum, m - - n , g rowth (-412OOnm): bac ter iochlorophyl l : A - - A (ffg/ml), O - - O (lt g/A12OOnm); O - - O A T P (nmole/.412o0 nm).
Biochim. Biophvs. Acta 234 (1971) 137-143
ATP LEVELS IN R . Rubrum DURING TRANSITION 139
The temperature was maintained at about 3 o°. The cultures were bubbled as indicated either with air or with purified nitrogen.
The growth of the cultures was measured in units of absorbance at 12oo nnl (ref. 14). Bacteriochlorophyll was est imated after extraction into methanol by absor- bance at 772 nm using the specific absorption coefficient of SMITH AND BEXlTEZ TM.
For ATP measurements, 5.5 ml of the culture were transferred rapidly by pressure from the culture vessel into o.5 ml of 3.6 M H2SO 4. The samples were ex- t rac ted for 3 ° min at room temperature. After this they were adjusted to pH 7.4 by the addit ion of o. 4 ml of 9 M NaOH and centrifugated for IO min at low speed to spin down the extracted cells, o.35 ml of the supernatant were used to est imate the ATP concentrat ion by the luciferin-lueiferase procedure as described by PRADET 17. The ATP-containing fluid was injected into a mixture of o.I ml diluted firefly lantern extract and o.25 ml buffer. Lyophilized firefly lanterns were obtained from Sigma Chemical Co. (Stock FLE-5o ). The light measuring appara tus was designed after tha t of HOLM-HANsEN AND BOOTH TM. The ATP values for the sample were determined by reference to a calibration curve.
I 151 34
13 k 3.0
1 I-- 2.6-
t E - - i c
o o ¢,1 O.g~ ~ 2 . 2 - o × ii -~
o I ~ _ , E i
0.7 I- ] .8~
0 . ~ -
0 . 3 _
1 I I
............. Air
. ._, _ , , /" /
/l" I
~ - i ,." I l i
I , / 0"
• / 14 \ .- / P ~,..--;" . /
i-"" o \\i ~ l I /
I . o L _ _ ~ ~ 0 1 2 3
Time (h)
I I I I |
-- 2E 11"5 . . . . . . . . . . . . . . . . . . IHt.... N2 .... I~
z
/" 24 M.3
• / ) t L,/"
~" 2C t1.1 ..
'"" iI'~i : i / ~ 16 O ~ ~
.." " ~ x" ,, ,o \ , 0
1 "~. / " i l l -e.--- I -~
',"- ~ ~'" I : / Z"
8 .0.5
i I - !
~ 4 JO.3 4 5 (3 7
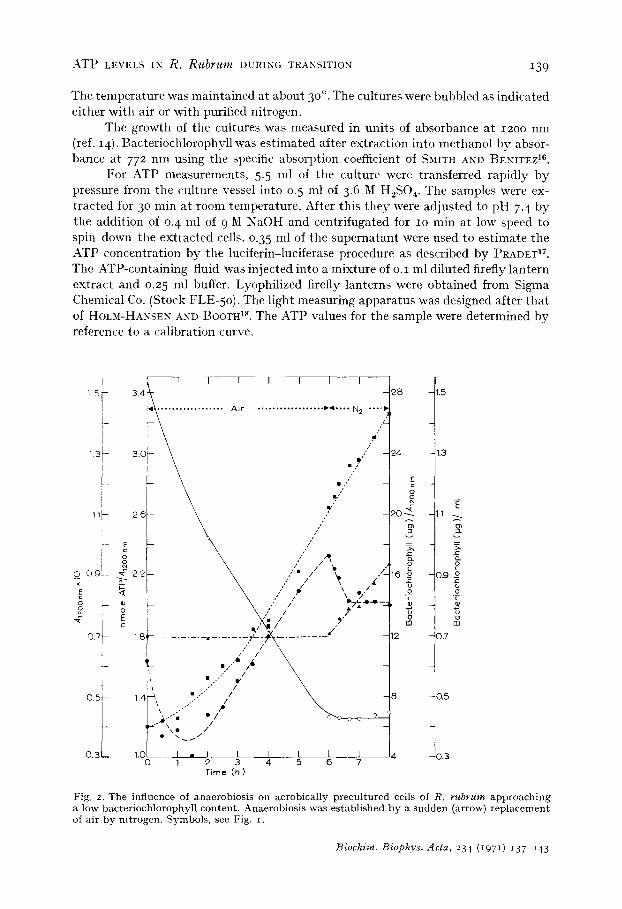
Fig. 2. The influence of anaerobiosis on aerobical ly p recu l tu red cells of R. r u b r u m app roach ing a low bacter iochlorophyl l content . Anaerobios is was es tab l i shed by a s u d d e n (arrow) r ep l acemen t of air by ni t rogen. Symbols , see Fig. i .
Bioch im . B i o p h y s . Ac ta , 234 (I97~) I37 - I43
I 4 0 J. OELZE, M. D. KAMEN
RESULTS
When anerobieally light-grown cells of R. rubrum are transferred to aerobic conditions in the light, they immediately cease synthesis of baeteriochlorophylP, 6. As growth continues under these conditions, the cellular baeteriochlorophyll content decreases. The amount of ATP per cell (A120onm) decreases only during the first few hours. Afterward it increases continuously (Fig. i).
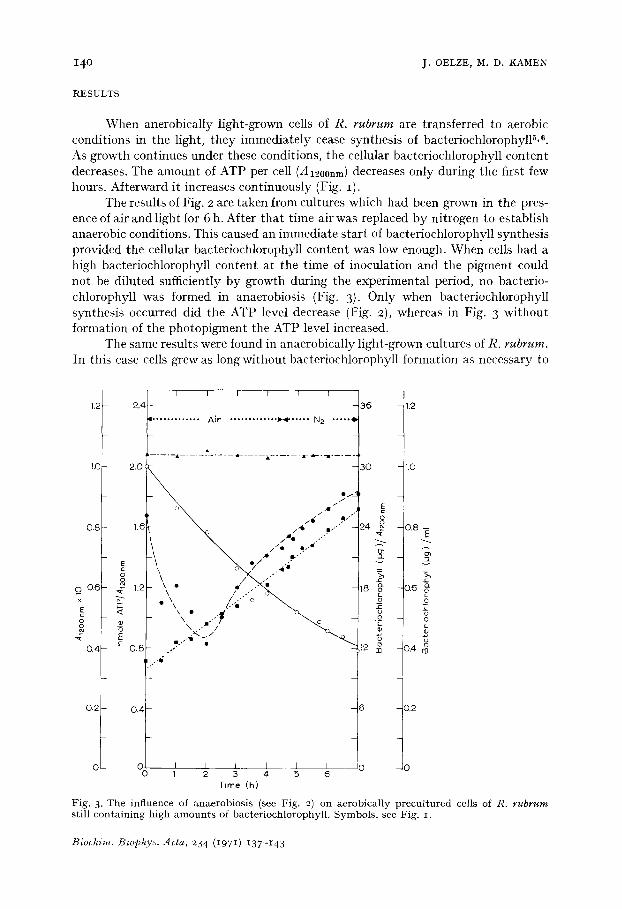
The results of Fig. 2 are taken from cultures which had been grown in the pres- ence of air and light for 6 h. After that time air was replaced by nitrogen to establish anaerobic conditions. This caused an immediate start of bacteriochlorophyll synthesis provided the cellular bacteriochlorophyll content was low enough. When cells had a high baeteriochlorophyll content at the t ime of inoculation and the pigment could not be diluted sufficiently by growth during the experimental period, no bacterio- chlorophyll was formed in anaerobiosis (Fig. 3). Only when bacteriochlorophyll synthesis occurred did the ATP level decrease (Fig. 2), whereas in Fig. 3 without formation of the photopigment the ATP level increased.
The same results were found in anaerobically light-grown cultures of R. rubrum. In this case cells grew as long without bacteriochlorophyll formation as necessary to
1.2
1.C
0.8
0 0.6
E c
0.4
I I I I I I I
2 ' 4 F ~ . . . . . . . . . . . . i r . . . . . . . . . . . . . IHI . . . . . . N 2 . . . . .
• . . . . . • . . . . . . . . . . . t . . . . k . . . . . . r~--1 . . . . LO ¢
o /,41" .,j /l( ,,~-"
1.6 "Xo / I , i- '" -
~ " ~ /~o ..-" / ~ / / e m,, "a
- \ ~ a i. '" -- ~ % e l / .,"i" =
o ° [ \ ., ..- , .2. ' • / < . . . ' ~ -
• /._.-" o \
0,8 , , " i t
F
0 I I l I _ _ I
1 2 3 4 5 Time (h)
36 - 1.2
30 - 1.0
E
24 .~ - D.8 -
i- 2
o o
12 c~ 3.4 m
0.2
I ~ ( 0 6
Fig. 3. The influence of anaerobiosis (see Fig. 2) on aerobically precultured cells of R. rubrum still containing high amounts of bacteriochlorophyll. Symbols, see Fig. I.
Biochim. Biophys. Acta, 234 (1971) 137-143
ATP LEVELS IN R. Rubrum DURING TRANSITION 141
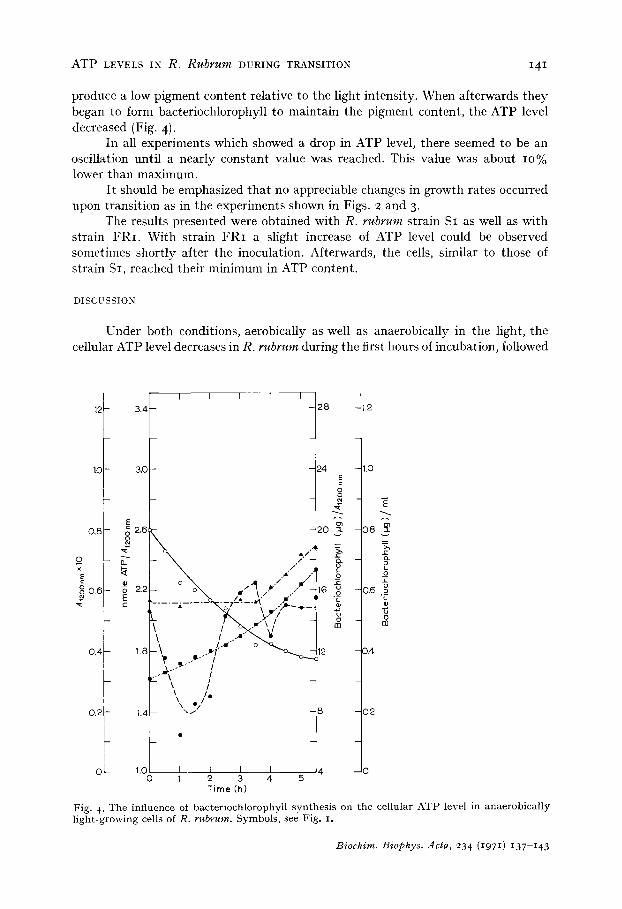
produce a low pigment content relative to the light intensity. When afterwards they began to form bacteriochlorophyll to maintain the pigment content, the ATP level decreased (Fig. 4).
In all experiments which showed a drop in ATP level, there seemed to be an oscillation until a nearly constant value was reached. This value was about lO% lower than maximum.
It should be emphasized that no appreciable changes in growth rates occurred upon transition as in the experiments shown in Figs. 2 and 3.
The results presented were obtained with R. rubrum strain SI as well as with strain FRI. With strain FRI a slight increase of ATP level could be observed sometimes shortly after the inoculation. Afterwards, the cells, similar to those of strain SI, reached their minimum in ATP content.
DISCUSSION
Under both conditions, aerobically as well as anaerobically in the light, the cellular ATP level decreases in R. rubrum during the first hours of incubation, followed
i2 -
10-
0 0 ' 8 t
o o.6
0'4 I
0,2~
at
e o
I I I I I 3.4 - 2;
3.0 2'~
2.6 X ,2( A / _
,,d / / / , I
/ ,if,/ 22 ""~ ~'" - - ~ • r ~ \ r ~/' ,,"
, / A " t ¢
. . . . . ;. . . . . . . . 7"-'--"*~ ./ ~ - - . . . . . , . . . - .
L F",. )'" I" -
1 8 ~..~- • _ . . y : l
, , . - " , " /
1,4 -- \~\\\-'°/I/° - 8
1.0 I I i I I 4 0 1 2 3 4 5
Time (h)
1.2
1.0
o.8
o~
0.6 "i
.4
.2
J 0
F i g . 4. T h e i n f l u e n c e o f b a c t e r i o c h l o r o p h y l l s y n t h e s i s o n t h e c e l l u l a r A T P l e v e l in a n a e r o b i c a l l y l i g h t - g r o w i n g c e l l s of R. r u b r u m . S y m b o l s , s e e F i g . z.
Biochim. Biophys. Mcta, 234 (1971) 137-143

142 J. OELZE, M. D. KAMEN
by an increase. In addition, preliminary experiments indicate that even the total amount of the adenine nucleotides decreases during this period. Similar results have been reported with R. spheroides, strain Y (ref. 14).
SCH6N TM has reported that in R. rubrum the ATP level is higher for aerobic light grown cultures than under anaerobic conditions. This might account for the lowering of ATP content when aerated cultures are bubbled through suddenly with nitrogen in the light (Fig. 2). However, the ATP decrease is observed only for cultures with low bacteriochlorophyll content (Fig. 2) and not for cultures which still contain high amounts of bacteriochlorophyll (Fig. 3). So, in this respect, our results are in agreement with data reported previously ~9,2° concerning an ATP level independent of anaerobiosis or aerobiosis in the light. This implies that the decrease of the ATP level in Fig. 2 should be attributed to changes other than those involving the energy- generating systems photosynthesis and respiration (see discussion below).
In Fig. 4 it is demonstrated that in anaerobically light-grown cells bacterio- chlorophyll is formed only when the cells have reached a critical low pigment content. Down to this value the ATP level increases, as seen in all the other figures. In l:ig. 2, as in Fig. 4, the ATP level drops when pigment synthesis begins. These results and the strong dependency of decrease in ATP level on conditions which induce bacterio- chlorophyll synthesis suggest a correlation between both bacteriochlorophyll synthesis and cellular ATP value.
It has been reported that in R. spheroides strain Y the ATP level exhibits a peak with a maximum just before bacteriochlorophyll synthesis begins under anaerobic light conditions 1~. When the first pigment synthesis becomes measurable, the value for ATP is low again. This suggests that the amount of cellular ATP has a "trigger" function in initiating pigment synthesis. However, no such effect is found in cultures of R. rubrum which exhibit their highest ATP levels precisely when bacteriochlorophyll synthesis starts. The ATP level continues to decrease within the first hour as the bacteriochlorophyll concentration increases (Fig. 2). (The sharp drop which is noted eventually is not what would be expected from the lowering of bacteriochlorophyll content. Rather one would expect a slow decrease.) It appears that the need for ATP for bacteriochlorophyll synthesis masks any transient which may be present. In fact, it is remarkable to find a transient peak in ATP cellular level when it is remem- bered that the assay of ATP levels involves the total extractable ATP rather than a portion of perhaps small magnitude which may be active at the site of pigment synthesis. Demonstration of "trigger" effects is probably possible only under special conditions when the major portion of ATP is not being utilized in a great variety of ways consequent on metabolic needs. The present results indicate that baeterio- chlorophyll synthesis as well as concomitant formation of the photosynthetic membrane draws heavily on ATP levels which in turn leads to a low ATP steady state level when bacteriochlorophyll levels reach a high value.
Our results show that the correlation of low ATP levels with high bacterio- chlorophyll levels characteristic of steady-state conditions in all photosynthetic bacteria holds for R. r~brum but that a regulatory function of ATP level in formation of pigment is not evident under conditions of transition from aerobic to anaerobic conditions in this organism.
Biochim. Bioph.vs. ~4cta, 234 (1971) I37-I43

ATP LEVELS IN R. Rubrum DURING TRANSITION 143
ACKNOWLEDGMENTS
\Ve gratefully acknowledge grants-in-aid from the National Institutes of Health (HD-oI262) and the National Science Foundation (GB-7o33X). J.O. holds a fellowship from the Deutsche Forsehungsgemeinschaft.
R E F E R E N C E S
i J. LASCELLES, in A. H. ROSE AND J. F. WILKINSON, Advances in Microbial Physiology, Acade- mic Press, New York, 1968, p. i.
2 A. GORCHEIN, A. NEUBERGER AND G. H. TAIT, Proe. Roy. Soc. London Ser. B, 17o (1968) 319. 3 J. YA.XrASHITA AND M. D. KAMEN, Biochem. Biophys. Res. Commun., 34 (1969) 418. 4 J. OELZE AND G. DREWS, Biochim. Biophys. Acta, 173 (1969) 448. 5 J. OELZE AND G. DREWS, Bioehim. Biophys. Aeta, 203 (197 o) 189. 6 G. COHEN-BAZlRE, VV. R. SISTROM AND R. Y. STANIER, J. Cellular Comp. Physiol., 49 (1957) 25. 7 \V. R. SISTROM, in H. GEST, A. San Pietro and L. P. VERNON, Bacterial Photosynthesis, Antioch
Press, Yellow Springs, Ohio, 1963, p. 53- 8 G. COHEN-BAZlRE AND ~,¥. R. SISTROM, in L. P. VERNON AND G. R. SEELY, The Chlorophylls,
Academic Press, New York, 1966, p. 313 . 9 G. A. SOJKA AND ]7I. GEST, Proc. Natl. Acad. Sci. U.S., 61 (1968) 1486.
IO G. SCH6N, Arch. Mikrobiol., 66 (I969) 348. I t A. GAJDOS AND M. GAJDOS-T6R6K, Nature, 199 (1965) lO93. 12 A. GAJDOS AND ~'V[. GAJDOS-TOROK, Nature, 207 (1965) 640. 13 G. A. SOJKA AND H. GEST, Bacteriol. Proc., 197 o, p. 64. 14 M. FANICA-GAIGNIER, J. CLEMENT-•ETRAL AND M. D. KAMEN, Biochim. Biophys. Acta, 226
(1971 ) 135. 15 H. DE KLERK, R. G. BARTSCH AND M. D. KAMEN, Biochim. Biophys. Acta, 97 (I965) 275. 16 J. H. C. SMITH AND A. BENITEZ, in K. PEACH AND M. V. TRACEY, Moderne 2~lethoden der
Pflanzenanalyse, Vol. 4, Springer, Berlin, 1955, p. 179. 17 A. PRADET, Physiol. Vdgdtale, 5 (1967) 209. 18 O. HOLM-HANSEN AND C. R. BOOTH, Limnol. Oceanog., i i (1966) 51o. 19 J. RAMIREZ AND L. SMITH, Biochim. Biophys. Acta, 153 (1968) 466. 20 1:. WELSCH AND L. SMITH, Biochemistry, 8 (1969) 3403 .
Bioehim. Biophys. Acta, 234 (1971) 137-143





























