Embed Size (px)
Citation preview

Adenosine Triphosphate Content of Marine BacteriaAuthor(s): Robert D. Hamilton and Osmund Holm-HansenSource: Limnology and Oceanography, Vol. 12, No. 2 (Apr., 1967), pp. 319-324Published by: American Society of Limnology and OceanographyStable URL: http://www.jstor.org/stable/2833045 .
Accessed: 17/06/2014 02:14
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
American Society of Limnology and Oceanography is collaborating with JSTOR to digitize, preserve andextend access to Limnology and Oceanography.
http://www.jstor.org
This content downloaded from 194.29.185.209 on Tue, 17 Jun 2014 02:14:08 AMAll use subject to JSTOR Terms and Conditions

ADENOSINE TRIPHOSPHATE CONTENT OF MARINE BACTERIA'
Robert D. Hamilton and Osmund Holm-Hansen Institute of Marine Resources, University of California, La Jolla 92037
ABSTRACT The adenosine triphosphate (ATP) content of seven selected cultures of marine bacteria,
representing five genera, has been determined during growth in chemostats and during the various growth phases in batch cultures. The range of ATP content in chemostat- grown cells was 0.5-6.5 X 10' /g ATP/cell, or 0.3 to 1.1% of the cell carbon. The ATP content of senescent cells in batch cultures and in starved cells was generally about one- fifth the concentration found in logarithmically growing cells. The average content of AT? in all the bacteria examined was 0.4% of cell carbon. It is concluded that ATP data can provide a useful estimate of heterotrophic biomass in the oceans.
INTRODUCTION Dissolved organic compounds and par-
ticulate, nonliving, organic matter account for over 99% of the total organic carbon in the oceans. Little is known about the role of this vast reservoir of material in marine community metabolism. One reason is the lack of methods for assaying the standing stock of heterotrophic microorganisms as well as their in situ activities.
Isolation plating techniques, extinction dilution, and direct microscopic counting are commonly used to enumerate marine bacteria. It is generally conceded that these methods produce erroneous esti- mates of the numbers of living bacteria in a water sample. The plating and extinction dilution techniques are selective because of the chemical composition of the media and because of inherent physical parameters such as temperature and pressure. In addi- tion, extinction dilution may be biologically selective in that one form may easily over- grow another. Therefore, these two ap- proaches are believed to produce low esti- mates of standing bacterial stocks. Direct counting, on the other hand, is believed to yield high estimates because of the diffi-
IThis research was supported in part by U.S. Atomic Energy Conmission Contract AT( 11-1)- 34, Project 108 and, in part, by U.S. Public Health Service Grant No. 1-ROl ES-00035-03. We are indebted to Miss Hester Kobayashi and Miss Kay Austin for their technical assistance during the course of this investigation. We also thank Dr. J. D. H. Strickland for his comments and sugges- tions and Dr. R. R. Colwell for providing certain of the cultures.
culty in distinguishing bacteria from bac- teria-sized, inert particles. Moreover, this method gives no estimate of viable cells and is therefore inapplicable to any prob- lem involving in situ activities.
Recently a method was developed for the quantitative assay of adenosine triphos- phate (ATP) in seawater samples (Holm- Hansen and Booth 1966). This method is based on the firefly luciferin-luciferase re- action as described by Strehler and Mc- Elroy (1957). The ATP content of a water sample may be related to its heterotrophic population if one assumes that no ATP is associated with nonliving material and if one takes samples at a depth which pre- cludes the occurrence of autotrophic plants. Macroscopic forms may be removed from samples by filtration through a net of 35-pt mesh size.
To relate ATP concentrations in the ocean to microbial biomass, it is necessary to know the approximate ATP content of the mi- crobial species present. As it is impossible to make a taxonomic analysis of the micro- biota in each water sample, the accuracy of such biomass estimations would depend upon the inter-generic constancy of ATP concentration. In addition, such estimates would depend upon the range of ATP con- centrations to be found in cells of the same species in different physiological conditions. In this paper, we report the results of an investigation into these variations in the ATP content of marine bacteria selected as representative of the genera that most commonly occur in seawater samples.
319
This content downloaded from 194.29.185.209 on Tue, 17 Jun 2014 02:14:08 AMAll use subject to JSTOR Terms and Conditions

320 ROBERT D. HAMILTON AND OSMUND HOLM-HANSEN
MATERIALS AND METHODS
The bacteria used in these experiments were all isolated from the marine environ- ment and required media with the ionic composition of seawater for growth.
Chromobacterium marinum is a pelagic isolate obtained by Dr. C. E. ZoBell and identified in our laboratory (Hamilton and Austin 1967). The Pseudomonas spp. were isolated in our laboratory, using a chemostat enrichment technique, from coastal sea- water (Hamilton, Morgan, and Strickland 1966), while the Serratia sp. was isolated from the same environment by conventional techniques. The latter isolate was identified by Dr. R. R. Colwell, who also supplied us with the Vibrio sp. and an estuarine isolate, the Micrococcus sp. All cultures were maintained in chemostats during the experimental period. These cultures pro- vided physiologically reproducible cells for routine ATP assay as well as inocula for batch culture experiments. Dilution rates varied slightly between isolates but were all close to 0.2/hr.
All media were prepared with artificial seawater (Lyman and Fleming 1940). The medium for the Pseudomonas spp. con- tained 1.0 mg glucose, 1.0 g (NH4)2SO4, and 0.01 g KH2PO4/liter. The glucose content of batch cultures was occasion- ally raised to 100 mg glucose/liter when large cell yields were required. The Vibrio sp., Serratia sp., and Micrococcus sp. were grown on 1.0% Bacto-peptone; C. marinum was grown in a medium consisting of 0.5% Bacto-peptone, 0.1% Bacto-yeast extract, 0.1% Bacto-casitone, and 0.01% KNO3.
Batch cultures were grown in approxi- mately 1 liter of medium in Fernbach flasks on a reciprocating shaker at 20C. Samples were withdrawn aseptically, and the cell number and ATP content were determined as outlined below.
The cells of a logarithmic growth phase culture of Pseudomonas sp. (GL-7) were collected by centrifugation at 5C at 27,000 x g for 30 min. The cells were washed once in artificial seawater and centrifuged. The washed cells were added to 1 liter of artifi-
cial seawater containing the usual amounts of nitrogen and phosphate but with no added carbon source. They were added in sufficient numbers to approximate their concentration in the original batch culture. These starved cells were incubated in the same manner as the usual batch culture and their ATP content followed with time.
Viable cells in chemostat or batch cul- tures were determined by a drop-plate tech- nique. However, in the case of the Micro- coccus sp. direct counting procedures had to be used in addition to drop plates be- cause the cells grew in clumps that could not be dispersed.
Cell carbon was analyzed after the method of Menzel and Vaccaro (1964) as modified by Holm-Hansen et al. (1966). Dry weight determinations were also made. Late log-phase cultures were used for both determinations. The cells were centrifuged from their culture media at 5C. They were washed once with artificial seawater, cen- trifuged, and resuspended in a small vol- ume of distilled water. Aliquots were taken for the carbon analyses while other aliquots were dried (16 hr at 70C) on tared planchets and weighed on a Cahn electro-balance.
For determination of ATP, samples (0.1- 1.0 ml) of the bacterial cultures were filtered through 25-mm HA Millipore? fil- ters (pore size, 0.45 It) as soon as possible after removal from the culture vessels. The filter was quickly placed in a small beaker and the cells killed by the addition of 3-4 ml of boiling Tris buffer (0.02 M, pH 7.75). After heating at lOOC for 5 min, to allow for complete extraction of ATP, the super- natant liquid was transferred to a test tube, and the filter again extracted with 1 ml of Tris buffer. The extracts were combined, diluted to 5.0 ml with Tris buffer, and frozen at -20C until the time of analysis. In some cases the filtration was omitted, and 0.1-ml samples were pipetted directly into 5.0 ml of boiling Tris buffer. As the concentration of salts in the nutrient me- dium interfered with the luciferase reaction, the final volume of extract had to be at least 25 times that of the sample con- taining the bacterial cells. Comparable re-
This content downloaded from 194.29.185.209 on Tue, 17 Jun 2014 02:14:08 AMAll use subject to JSTOR Terms and Conditions
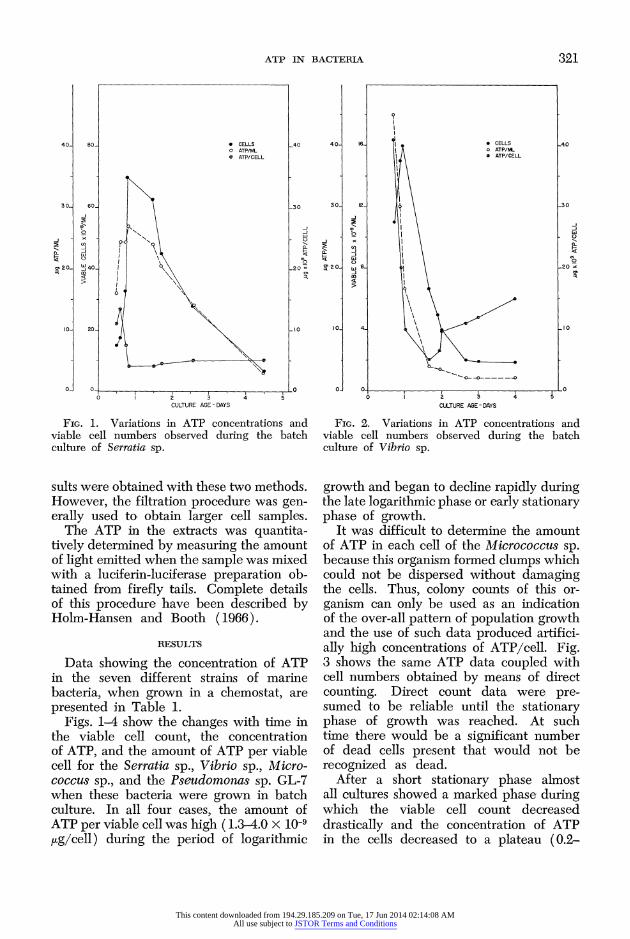
ATP IN BACTERIA 321
40_ 80_ * CELLS 40 0 ATP/ML * ATP/CELL
3o0 60| 30
R G 4 { K 2
120- 40 I 20_
I , '0 , , .
0- 00 -0
0 1 2 3 4 5 CULTURE AGE -DAYS
FIG. 1. Variations in ATP concentrations and viable cell numbers observed during the batch culture of Serratia sp.
sults were obtained with these two methods. However, the filtration procedure was gen- erally used to obtain larger cell samples.
The ATP in the extracts was quantita- tively determined by measuring the amount of light emitted when the sample was mixed with a luciferin-luciferase preparation ob- tained from firefly tails. Complete details of this procedure have been described by Holm-Hansen and Booth (1966).
RESULTS
Data showing the concentration of ATP in the seven different strains of marine bacteria, when grown in a chemostat, are presented in Table 1.
Figs. 1-4 show the changes with time in the viable cell count, the concentration of ATP, and the amount of ATP per viable cell for the Serratia sp., Vibrio sp., Micro- coccus sp., and the Pseudomonas sp. GL-7 when these bacteria were grown in batch culture. In all four cases, the amount of ATP per viable cell was high (1.3-4.0 x 10-9 jug/cell) during the period of logarithmic
40 16 o CELLS 40 *ATP/ML
* ATP/CELL
30 O 12 1 230
I -~~~~~~~~~~~~~~~~~~j o w
cutr ofViro p
012 - S_ I2
10- 4- 10
growth~~ l an ea o eln apdydrn
0 0 0 0 I 2 3' 4' 5
CULXTURE AGE-DAYS
FiG. 2. Variations in ATP concentrations and viable cell numbers observed during tbe batch culture of Vibrio sp.
growth and began to decline rapidly during the late logarithmic phase or early stationary phase of growth.
It was difficult to determine the amount of ATP in each cell of the Micrococcus sp. because this organism formed clumps which could not be dispersed without damaging the cells. Thus, colony counts of this or- ganism can only be used as an indication of the over-all pattern of population growth and the use of such data produced artifici- ally high concentrations of ATP/cell. Fig. 3 shows the same ATP data coupled with cell numbers obtained by means of direct counting. Direct count data were pre- sumed to be reliable until the stationary phase of growth was reached. At such time there would be a significant number of dead cells present that would not be recognized as dead.
After a short stationary phase almost all cultures showed a marked phase during which the viable cell count decreased drastically and the concentration of ATP in the cells decreased to a plateau (0.2-
This content downloaded from 194.29.185.209 on Tue, 17 Jun 2014 02:14:08 AMAll use subject to JSTOR Terms and Conditions
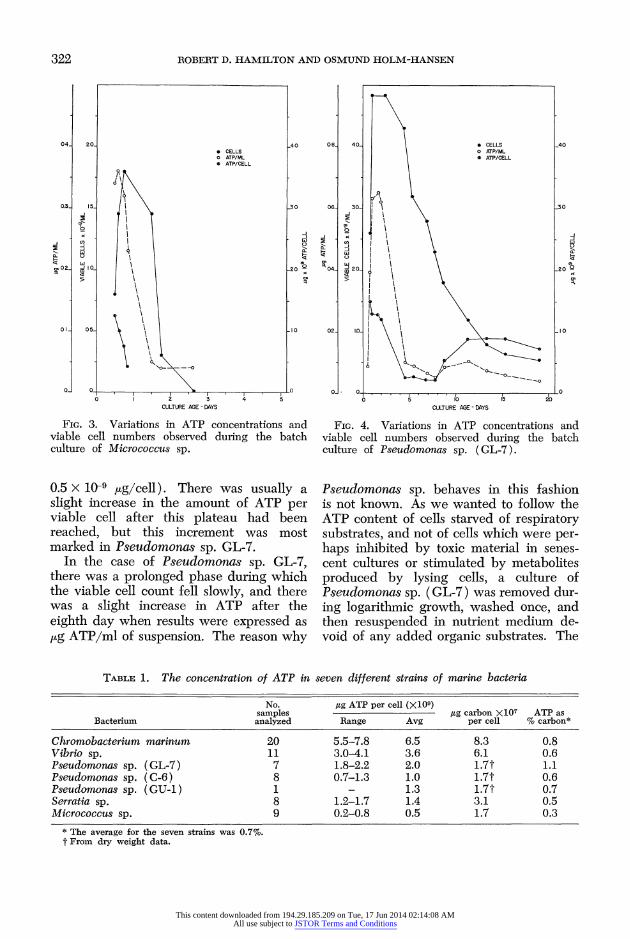
322 ROBERT D. HAMILTON AND OSMUND HOLM-HANSEN
04_ 20_ _40 * CELLS o ATP/ML * ATP/CELL
'9
03- 15 I 30
o02 V0 20 >
01- 05- ,10
0 1 2 3 4 5 CULTURE AGE -DAYS
FIG. 3. Variations in ATP concentrations and viable cell numbers observed during the batch culture of Micrococcus sp.
0.5 x 10-9 jig/cell). There was usually a slight increase in the amount of ATP per viable cell after this plateau had been reached, but this increment was most marked in Pseudomonas sp. GL-7.
In the case of Pseudomonas sp. GL-7, there was a prolonged phase during which the viable cell count fell slowly, and there was a slight increase in ATP after the eighth day when results were expressed as ,ug ATP/ml of suspension. The reason why
08_ 40 | CELLS _40 o ATP/ML * ATP/CELL
0 I _
'IO~~~~~~~~~~~~~~~~~~I
0- 0 0 0 5' o10 15 20
CULTURE AGE - DYS
FIG. 4. Variations in ATP concentrations and viable cell numbers observed during the batch culture of Pseudomonas sp. (GL-7).
Pseudomonas sp. behaves in this fashion is not known. As we wanted to follow the ATP content of cells starved of respiratory substrates, and not of cells which were per- haps inhibited by toxic material in senes- cent cultures or stimulated by metabolites produced by lysing cells, a culture of Pseudomonas sp. (GL-7) was removed dur- ing logarithmic growth, washed once, and then resuspended in nutrient medium de- void of any added organic substrates. The
TABLE 1. The concentration of ATP in seven different strains of marine bacteria
No. ,ug ATP per cell (X109) samples ,ug carbon X107 ATP as
Bacterium analyzed Range Avg per cell % carbon*
Chromobacterium marinum 20 5.5-7.8 6.5 8.3 0.8 Vibrio sp. 11 3.0-4.1 3.6 6.1 0.6 Pseudomonas sp. (GL-7) 7 1.8-2.2 2.0 1.7t 1.1 Pseudomonas sp. (C-6) 8 0.7-1.3 1.0 1.7t 0.6 Pseudomonas sp. (GU-1) 1 _ 1.3 1.7t 0.7 Serratia sp. 8 1.2-1.7 1.4 3.1 0.5 Micrococcus sp. 9 0.2-0.8 0.5 1.7 0.3
* The average for the seven strains was 0.7%. t From dry weight data.
This content downloaded from 194.29.185.209 on Tue, 17 Jun 2014 02:14:08 AMAll use subject to JSTOR Terms and Conditions
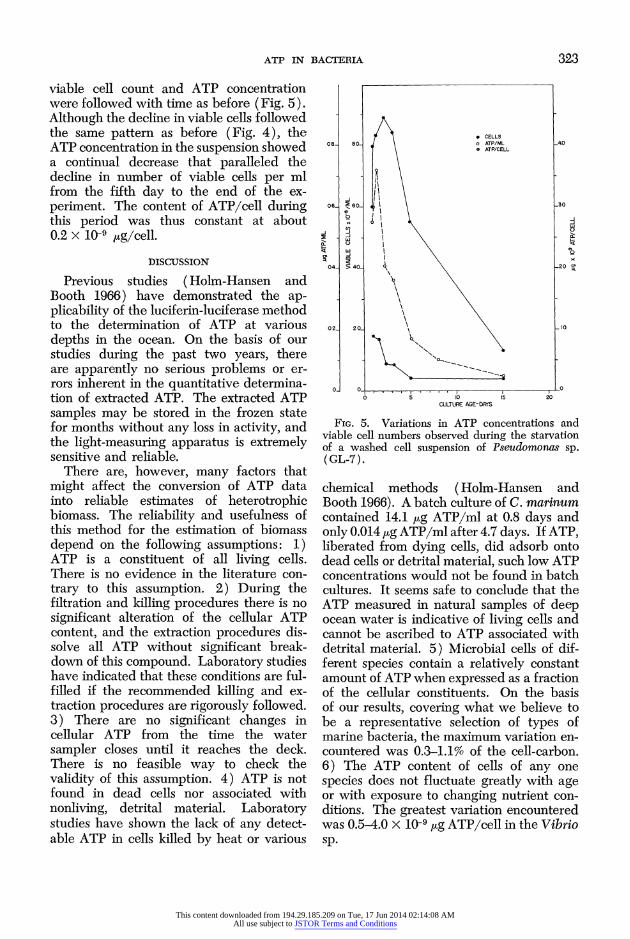
ATP IN BACTERIA 323
viable cell count and ATP concentration were followed with time as before (Fig. 5). Although the decline in viable cells followed the same pattern as before (Fig. 4), the ATP concentration in the suspension showed a continual decrease that paralleled the decline in number of viable cells per ml from the fifth day to the end of the ex- periment. The content of ATP/cell during this period was thus constant at about 0.2 x 10-9 kg/cell.
DISCUSSION
Previous studies (Holm-Hansen and Booth 1966) have demonstrated the ap- plicability of the luciferin-luciferase method to the determination of ATP at various depths in the ocean. On the basis of our studies during the past two years, there are apparently no serious problems or er- rors inherent in the quantitative determina- tion of extracted ATP. The extracted ATP samples may be stored in the frozen state for months without any loss in activity, and the light-measuring apparatus is extremely sensitive and reliable.
There are, however, many factors that might affect the conversion of ATP data into reliable estimates of heterotrophic biomass. The reliability and usefulness of this method for the estimation of biomass depend on the following assumptions: 1) ATP is a constituent of all living cells. There is no evidence in the literature con- trary to this assumption. 2) During the filtration and killing procedures there is no significant alteration of the cellular ATP content, and the extraction procedures dis- solve all ATP without significant break- down of this compound. Laboratory studies have indicated that these conditions are ful- filled if the recommended killing and ex- traction procedures are rigorously followed. 3) There are no significant changes in cellular ATP from the time the water sampler closes until it reaches the deck. There is no feasible way to check the validity of this assumption. 4) ATP is not found in dead cells nor associated with nonliving, detrital material. Laboratory studies have shown the lack of any detect- able ATP in cells killed by heat or various
* CELLS 08- 80 o ATP/ML 40
\ ATP/CELL
_1 06 60- 630
~w w
0 5 10 15 20 CULTURE AGE-DAYS
FIG. 5. Variations in ATP concentrations anld viable cell numbers observed during the starvation of a washed cell suspension of Pseuctononass sp. ( GL-7).
chemical methods ( Hoim-Hansen and Booth 1966). A batch culture of C. marinum contained 14.1 ,ug ATP/ml at 0.8 days and only 0.014 ,ug ATP/ml after 4.7 days. If ATP, liberated from dying cells, did adsorb onto dead cells or detrital material, such low ATP concentrations would not be found in batch cultures. It seems safe to conclude that the ATP measured in natural samples of deep ocean water is indicative of living cells and cannot be ascribed to ATP associated with detrital material. 5) Microbial cells of dif- ferent species contain a relatively constant amount of ATP when expressed as a fraction of the cellular constituents. On the! basis of our results, covering what we believe to be a representative selection of types of marine bacteria, the maximum variation en- countered was 0.3-1.1% of the cell-carbon. 6) The ATP content of cells of any one species does not fluctuate greatly with age or with exposure to changing nutrient con- ditions. The greatest variation encountered was 0.5-4.0 x 10-9 ,ug ATP/cell in the Vibrio sp.
This content downloaded from 194.29.185.209 on Tue, 17 Jun 2014 02:14:08 AMAll use subject to JSTOR Terms and Conditions

324 ROBERT D. HAMILTON AND OSMUND HOLM-HANSEN
The chemostat-grown cultures and the senescent batch cultures probably represent the extremes of cellular condition one might expect to find in the sea. We have, there- fore, averaged a representative number of observations from both types of cells and obtained a figure for the average ATP con- tent of these bacteria. This figure is 1.5 x 10-9 ug ATP/cell. Taking an average of the cell carbon data reported in Table 1 for the five genera and making the assump- tion that a cell would have the same carbon content whether exponentially growing or senescent, an average of the ATP/cell data may be expressed as percentage cell car- bon. The figure so obtained is 0.4%.
While the ATP/cell value might be used to convert ATP yield data into the number of heterotrophic bacteria present, it would seem more reasonable to use the average ATP concentration of the cells expressed as percentage carbon. This avoids the im- plication that all the heterotrophs in the ocean are bacteria.
Some investigators (see Strickland 1965; Fournier 1966) have reported the occur- rence of microscopic, nonbacterial hetero- trophs below the euphotic zone. These forms may be a significant fraction of the total heterotrophic biomass of deep ocean waters, but such forms are poorly de- scribed in terms of their distribution and ecology. It would therefore be difficult, even if cultures were available, to establish an estimate of the ATP content of a repre- sentative selection of these forms. We can say, however, that most algae investigated to date (Coombs et al. 1967a, 1967b; Holm-Hansen and Booth 1966; Syrett 1958) have had an ATP content (on a dry weight or carbon basis) in about the same range as that reported here.
REFERENCES COOMBS, J., P. J. HALICKI, 0. HOLM-HANSEN, AND
B. E. VOLCANI. 1967a. Studies on the bio- chemistry and fine structure of silica shell formation in diatoms. Changes in concentra- tion of nucleoside triphosphate in silicon-starva- tion synchrony of Navicula pelliculosa (Br6b.) Hilse. Exptl. Cell Res., (in press).
- AND . 1967b. Studies on the biochemistry and fine struc- ture of silica shell formation in diatoms. Changes in concentration of nucleoside tri- phosphate during synchronized division of Cylindrotheca fusiformis Reimann and Lewin. Exptl. Cell Res., (fin press).
FouRNIER, R. 0. 1966. North Atlantic deep sea fertility. Science, 153: 1250-1252.
HAMILTON, R. D., AND K. E. AUSTIN. 1967. Physiological and cultural characteristics of Chromobacterium marinzum sp. n. Antonie van Leeuwenhoek, J. Microbiol. Serol., (in press).
, K. M. MORGAN, AND J. D. H. STRICKLAND. 1966. The glucose uptake kinetics of some marine bacteria. Can. J. Microbiol., 12: 995-1003.
HoLM-HANsEN, O., AND C. R. BOOTH. 1966. The measurement of adenosine triphosphate in the ocean and its ecological significance. Limnol. Oceanog., 11: 510-519.
, J. COOMBS, B. E. VOLCANI, AND P. M. WILLIAMS. 1966. Quantitative micro-deter- mination of lipid carbon in microorganisms. Anal. Biochem., (in press).
LYMAN, J., AND R. H. FLEMING. 1940. Com- position of sea water. J. Marine Res., 3: 134-149.
MENZEL, D. W., AND R. F. VACCARO. 1964. The measurement of dissolved organic and particulate carbon in seawater. Limnol. Oceanog., 9: 138-142.
STREHLER, B. L., AND W. D. McELRoY. 1957. Assay of adenosine triphosphate, p. 871-8,73. In S. P. Colowick and N. 0. Kaplan [eds.], Methods in enzymology, v. 3. Academic, New York.
STRICKLAND, J. D. H. 1965. Phytoplankton and marine primary production. Ann. Rev. Micro- biol., 19: 127-162.
SYRETr, P. 1958. Respiration rate and internal adenosine triphosphate concentration in Chlo- rella. Arch. Biochem. Biophys., 75: 117- 124.
This content downloaded from 194.29.185.209 on Tue, 17 Jun 2014 02:14:08 AMAll use subject to JSTOR Terms and Conditions





























