Embed Size (px)
Citation preview

A(
JI
a
ARRA
KAAGGILNP
1
stScfig
ofi1Jaeras
uCe
0h
Fisheries Research 137 (2013) 30– 40
Contents lists available at SciVerse ScienceDirect
Fisheries Research
jou rn al h om epa ge: www.elsev ier .com/ locate / f i shres
ge and growth of anglerfish (Lophius piscatorius) on the Porcupine Bankwest of Ireland) based on illicia age estimation
orge Landa ∗, Joaquín Barrado, Francisco Velasconstituto Espanol de Oceanografía, Centro Oceanográfico de Santander, Promontorio de San Martín s/n, 39080 Santander, Spain
r t i c l e i n f o
rticle history:eceived 27 January 2012eceived in revised form 28 June 2012ccepted 17 July 2012
eywords:ge estimation
a b s t r a c t
The age of white anglerfish (Lophius piscatorius) on the Porcupine Bank was estimated based on theage estimation of 979 illicia (first dorsal fin ray) obtained from a series of 9 annual groundfish surveys(2001–2009). Modifications in the methodology of illicia preparation and in the traditional biased ageestimation criterion are presented. This is the first time that a growth pattern of white anglerfish basedon illicia age estimations can be indirectly validated by tracking cohorts using the abundance indicesper age class from surveys and length-frequency analyses. Previous studies of cohort tracking using thetraditional illicia age estimation criterion showed a mismatch, suggesting a faster growth. With the new
nglerfishroundfish surveysrowth
lliciumength-frequencyortheast Atlantic Ocean
proposed criteria, abundant cohorts from 2001 to 2004, and scarce ones, from 2005 to 2007, can betracked over the time throughout several age groups. A new growth pattern and von Bertalanffy growthparameters are presented (L∞: 162.31; k: 0.088; t0: −0.894). The results are compared with previousstudies in other areas.
© 2012 Elsevier B.V. All rights reserved.
orcupine Bank. Introduction
The white anglerfish (Lophius piscatorius) is a bottom-livingpecies, found from 50 m to 1000 m depth, from the Barents Seao the Strait of Gibraltar, and in the Mediterranean and the Blackea (Whitehead et al., 1986; Dardignac, 1988). White anglerfish is aommercially important species (∼33,000 t in 2010) for Europeansheries and it is captured by a mixed trawl fishery and a targetedillnet fleet (ICES, 2011a,b).
Most growth studies on white anglerfish up to date have focusedn age estimation using hard parts, such as illicia (first dorsaln ray) (Dupouy et al., 1986; Peronnet et al., 1992; Duarte et al.,997; Quincoces et al., 1998; Landa et al., 2001; Quincoces, 2002;ónsson, 2007; Ofstad and Laurenson, 2007) or otoliths (Tsimenidisnd Ondrias, 1980; Crozier, 1989; Wright et al., 2002; Woodroffet al., 2003). Few studies have been based on analyses of tagging-ecapture specimens (Laurenson et al., 2005; Landa et al., 2008a)nd length-frequency distributions of commercial and researchurveys catches (Fulton, 1903; Dupouy et al., 1986; Jónsson, 2007).
The first growth patterns of white anglerfish in Atlantic waters
sing hard parts (illicia by Dupouy et al., 1986 and otoliths byrozier, 1989) showed similarities, but with one age class of differ-nce between them. Several European age estimation workshops∗ Corresponding author. Tel.: +34 942 291716; fax: +34 942 275072.E-mail address: [email protected] (J. Landa).
165-7836/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.fishres.2012.07.026
and hard parts exchanges have taken place to standardize an ageestimation criterion and to compare results from both hard parts(Anon, 1997, 1999; Landa et al., 2002a; Duarte et al., 2005; Landa,2012). The growth pattern is easier to distinguish in the illicium(Crozier, 1989; Woodroffe et al., 2003) and it has become thestandard structure for age estimation of white anglerfish in mostEuropean countries (Landa et al., 2008a).
The age estimation from illicia of a decadal time-series wasperformed for the southern stock (Iberian Atlantic) assessment ofwhite anglerfish, using the standardized age estimation criterionof Duarte et al. (2002). A catch-at-age by year matrix was built,but inconsistencies in cohort tracking were found (Azevedo et al.,2008). Since then no age-structured model has been used for theassessment of both northern and southern stocks of the EuropeanAtlantic southern shelf of white anglerfish (ICES, 2011a). A produc-tion model (ASPIC, Prager, 1994) has been used for the assessmentas an alternative to the age-structured models. Length-based model(CASA, Sullivan et al., 1990; Dobby, 2002) including von Bertalanffygrowth parameters is also being attempted for the stock assessmentof white anglerfish.
On the other hand, growth of white anglerfish in Atlantic watershas also been studied more recently using micro-increments inhard parts (Wright et al., 2002) and other alternative techniques
to estimate the age from hard parts, as the tagging-recapture(Laurenson et al., 2005; Landa et al., 2008a) or the analysis oflength-frequency distributions of catches (Dupouy et al., 1986;Thangstad et al., 2002; Landa, 2004; Jónsson, 2007). Landa et al.
s Research 137 (2013) 30– 40 31
(batau
LasitcbwsmKmg
abwaaepgseatalB
e(
tptaca
mit(oBds
2
(c5a2i
Table 1Number of illicia age estimates and length range of white anglerfish by annual surveyin Porcupine Bank.
Year Number Minimumlength (cm)
Maximumlength (cm)
2001 114 6 1252002 98 8 1082003 129 7 1222004 142 5 1202005 109 12 1202006 86 5 962007 97 6 1022008 81 12 107
J. Landa et al. / Fisherie
2008a) estimated new faster growth rate and growth parameters,ased on the results obtained in aforementioned growth studieslternative to the age estimates. Their results also concluded thathe growth pattern estimated by using the traditional standardizedge estimation criterion based on illicia (Duarte et al., 2002) wasnderestimated.
Velasco et al. (2008) applied the new faster growth model ofanda et al. (2008a) on the 6 years time-series of catches of whitenglerfish from annual bottom trawl surveys of Porcupine Bank,howing, for the first time, results on its abundance and distributionn that area. Velasco et al. (2008) showed that applying the tradi-ional standardized age estimation criterion on illicia to the surveyatches, a mismatch in cohort tracking over time was produced,ut when the new faster growth model was applied, some cohortsere successfully tracked. The mismatch in the cohort analysis,
uggested that the traditional age estimation criterion overesti-ated up to three of the younger age classes. When an Age Length
ey (ALK) was estimated numerically from the new faster growthodel, this mismatch disappeared, which to confirm the faster
rowth.Although the overall growth parameters of Landa et al. (2008a)
re currently suitable for the stock assessment models, it woulde also desirable to describe accurately the growth pattern ofhite anglerfish in an area (e.g. Porcupine Bank) based on the
ge estimation of hard parts of specimens from that area, as anlternative to the current approach using overall growth param-ters. A relevant advantage of the annual age estimation of hardarts is to have annual catches-at-age, which include the annualrowth variability and cohort strength, offering the possibility totudy interrelations between both. As the overall growth param-ters indicating faster growth seem to show a good fit for whitenglerfish from Porcupine Bank (Velasco et al., 2008), a fur-her step is to test if the age estimation on hard parts using
new age estimation criterion can be indirectly validated byength distribution analyses and by cohorts tracking in Porcupineank.
That new criterion should include recent modifications on agestimation methodologies, as the recommendations of Wright et al.2002) about the first supposed annual increment.
Bottom trawl surveys are nowadays one of the most impor-ant methods for assessing commercial fish stocks, because theyrovide information independent from the fishery and unlikelyo be affected by misreporting (Beare et al., 2003). Since 2001 annnual Spanish bottom trawl survey has been carried out in Por-upine Bank and therefore the temporal series of standardizedbundance indices of white anglerfish can be analysed.
The aims of the present study are: (i) to present a new age esti-ation criterion, including modifications in the methodology of
llicia preparation and in the traditional standardized age estima-ion criterion of 2002 that allows obtaining a new growth pattern;ii) to present a new growth pattern based on illicia age estimationf white anglerfish from 9 years of annual surveys in Porcupineank; (iii) to validate indirectly the new growth pattern by lengthistribution analyses and cohorts tracking in the available time-eries of Porcupine Bank survey.
. Materials and methods
Nine annual surveys, following the coordinated at the IBTSInternational Bottom Trawl Surveys) protocols (ICES, 2010a) werearried out on the Porcupine Bank from 11 to 15◦W and from
1 to 54◦N, covering a depth range of 180–800 m, between 2001nd 2009. The Porcupine Bank lies in the Northeast Atlantic, about00 km west of Ireland. Velasco et al. (2008) showed informationn detail on the Bank.
2009 123 7 111
Total 979 5 125
The survey sampling design is shown in Fig. 1, and thoroughlydescribed with the methodology used in Velasco et al. (2008) andexplained in ICES (2010b).
2.1. Age estimation
A sub-sample of the caught white anglerfish was aged, with amaximum of five illicia for each 1 cm length class and year. Mostof the medium and large specimens caught were aged, and thesub-sample mainly affected the specimens <35 cm from years withhigh recruitments. Therefore, the age was estimated in 979 illiciaof white anglerfish ranging from 5 to 125 cm, with a mean num-ber of 109 illicia by survey (Table 1). Lengths were measured to thecentimetre below.
The traditional standardized age estimation criterion on ill-icia was based on the observation and counting of growth marks(Duarte et al., 2002). Its application produced biased ages, due tosome first growth marks were considered as annuli did not seemto be annual (Landa et al., 2008a). These authors concluded thatfurther research on optimal sectioning methods as well as mag-nification could help to improve age estimation criteria and theidentification of true annual increments (annuli).
In the present study several modifications of the traditionalstandardized methodology of preparation, observation and ageinterpretation on illicia, have been performed. An analysis has alsobeen conducted of possible shifting of those methods from the firststudy of Dupouy et al. (1986). Dupouy method was further stan-dardized at the illicia age estimation guide of Duarte et al. (2002),after several age estimation workshops (Anon, 1997, 1999; Landaet al., 2002a), and it was used in most of the growth studies usingillicia (Duarte et al., 1997; Quincoces et al., 1998; Landa et al., 2001;Ofstad and Laurenson, 2007).
The following methodological modifications have beenattempted to allow a more clear observation of the growthpattern, showing mainly the most apparent growth marks, inorder to allow the distinction of the annuli:
(i) Section thickness. After illicia were embedded in resin, sev-eral cut thicknesses of the transverse section were tested.The thinner cuts drove to the observation of more supposedannual increments, and vice versa using thicker cuts. The useof transverse sections ∼0.50–0.55 mm thick allowed the obser-vation of the clearest marked increments, probably those thatare annual. However, the use of sections thinner than 0.5 mm(∼0.4 mm) produced the observation and count of some falseannual increments as annuli, as Landa et al. (2008a) indicated.
(ii) Observation. Sections were observed using a profile projec-tor at 50×, as used initially by Dupouy et al. (1986), instead ofusing a microscope at 100× under transmitted light, which wasthe standard observation methodology overall used by Duarte
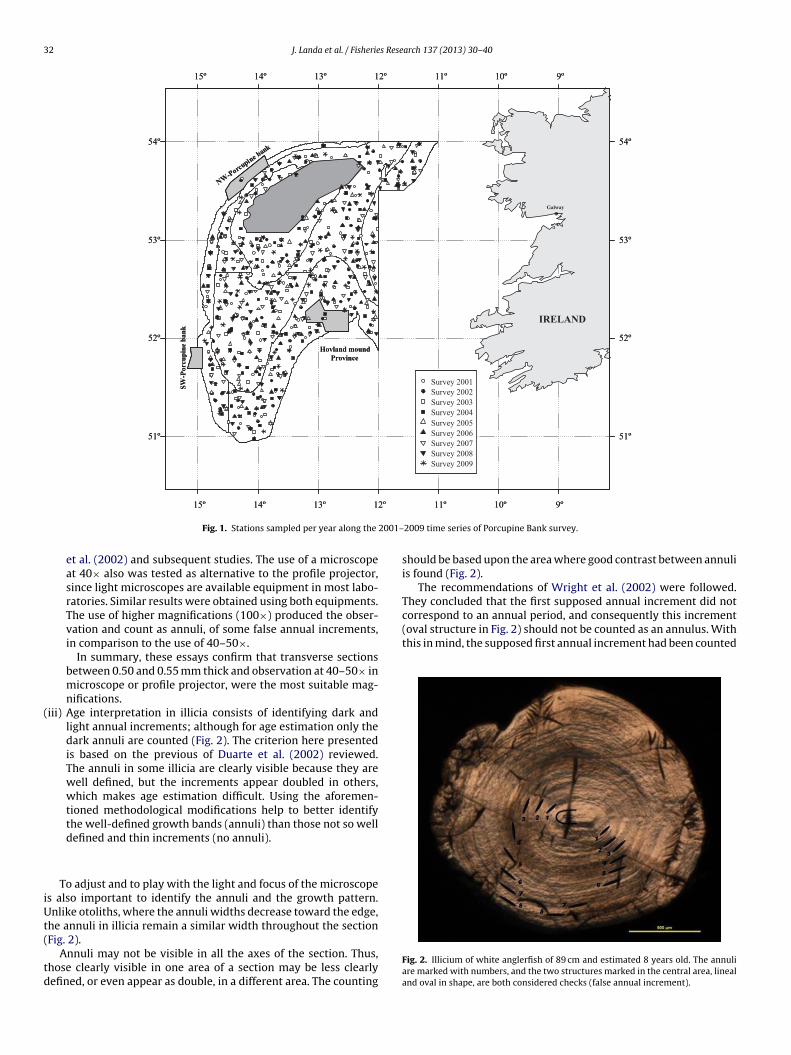
32 J. Landa et al. / Fisheries Research 137 (2013) 30– 40
Galway
IRELAND
2º
51º
52º
53º
54º
15º 14º 13º 12º 11º 10º 9º
51º
52º
53º
54º
Hovland mound
Province
NW-P
orcupin
e bank
SW
-Po
rcu
pin
e b
an
k
Survey 2001
Survey 2002
Survey 2003
Survey 2004
Survey 2005
Survey 2006
Survey 2007
Survey 2008
Survey 2009
2º
51º
52º
53º
54º
15º 14º 13º 12º 11º 10º 9º
51º
52º
53º
54º
Hovland mound
Province
NW-P
orcupin
e bank
SW
-Po
rcu
pin
e b
an
k
Survey 2001
Survey 2002
Survey 2003
Survey 2004
Survey 2005
Survey 2006
Survey 2007
Survey 2008
Survey 2009
2001–
(
iUt(
td
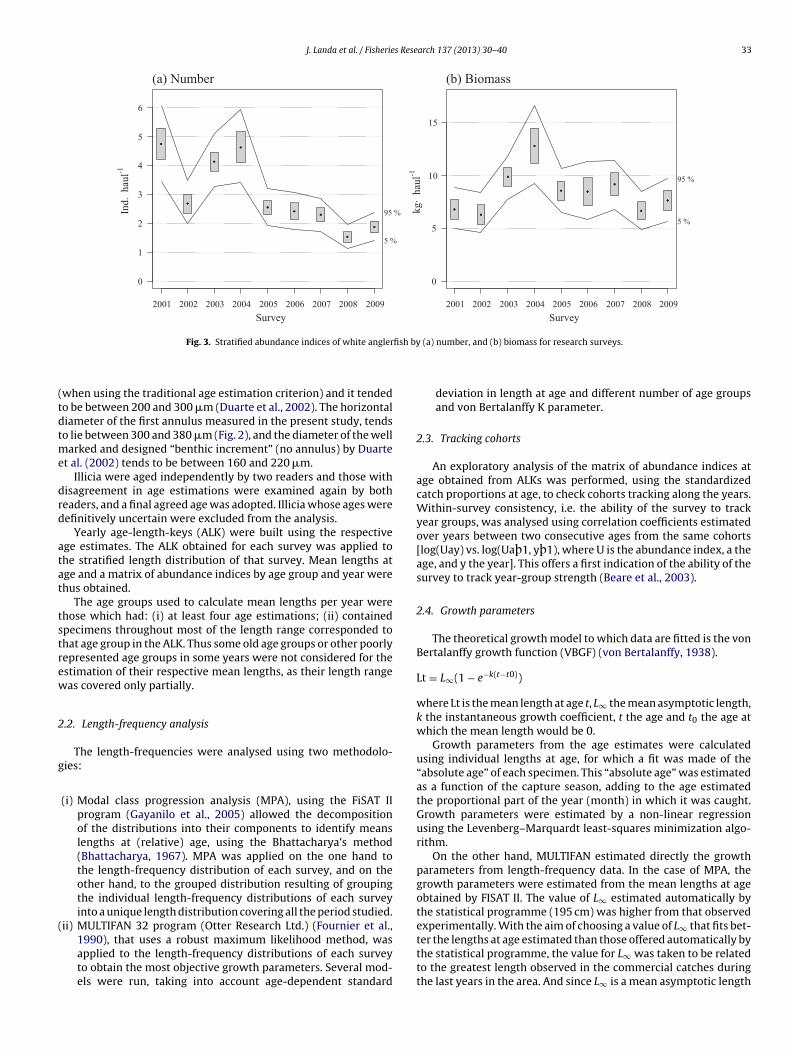
They concluded that the first supposed annual increment did notcorrespond to an annual period, and consequently this increment(oval structure in Fig. 2) should not be counted as an annulus. Withthis in mind, the supposed first annual increment had been counted
15º 14º 13º 115º 14º 13º 1
Fig. 1. Stations sampled per year along the
et al. (2002) and subsequent studies. The use of a microscopeat 40× also was tested as alternative to the profile projector,since light microscopes are available equipment in most labo-ratories. Similar results were obtained using both equipments.The use of higher magnifications (100×) produced the obser-vation and count as annuli, of some false annual increments,in comparison to the use of 40–50×.
In summary, these essays confirm that transverse sectionsbetween 0.50 and 0.55 mm thick and observation at 40–50× inmicroscope or profile projector, were the most suitable mag-nifications.
iii) Age interpretation in illicia consists of identifying dark andlight annual increments; although for age estimation only thedark annuli are counted (Fig. 2). The criterion here presentedis based on the previous of Duarte et al. (2002) reviewed.The annuli in some illicia are clearly visible because they arewell defined, but the increments appear doubled in others,which makes age estimation difficult. Using the aforemen-tioned methodological modifications help to better identifythe well-defined growth bands (annuli) than those not so welldefined and thin increments (no annuli).
To adjust and to play with the light and focus of the microscopes also important to identify the annuli and the growth pattern.nlike otoliths, where the annuli widths decrease toward the edge,
he annuli in illicia remain a similar width throughout the section
Fig. 2).Annuli may not be visible in all the axes of the section. Thus,hose clearly visible in one area of a section may be less clearlyefined, or even appear as double, in a different area. The counting
11º 10º 9º11º 10º 9º
2009 time series of Porcupine Bank survey.
should be based upon the area where good contrast between annuliis found (Fig. 2).
The recommendations of Wright et al. (2002) were followed.
Fig. 2. Illicium of white anglerfish of 89 cm and estimated 8 years old. The annuliare marked with numbers, and the two structures marked in the central area, linealand oval in shape, are both considered checks (false annual increment).
J. Landa et al. / Fisheries Research 137 (2013) 30– 40 33
Survey
Ind. h
aul-1
kg
. h
aul-1
0
1
2
3
4
5
6
2001 2002 2003 2004 2005 2006 2007 2008 2009
5 %
95 %
(a) Number
Survey
0
5
10
15
2001 2002 2003 2004 2005 2006 2007 2008 2009
5 %
95 %
(b) Biomass
fish by
(tdtme
drd
atat
tstrew
2
g
(
Fig. 3. Stratified abundance indices of white angler
when using the traditional age estimation criterion) and it tendedo be between 200 and 300 �m (Duarte et al., 2002). The horizontaliameter of the first annulus measured in the present study, tendso lie between 300 and 380 �m (Fig. 2), and the diameter of the well
arked and designed “benthic increment” (no annulus) by Duartet al. (2002) tends to be between 160 and 220 �m.
Illicia were aged independently by two readers and those withisagreement in age estimations were examined again by botheaders, and a final agreed age was adopted. Illicia whose ages wereefinitively uncertain were excluded from the analysis.
Yearly age-length-keys (ALK) were built using the respectivege estimates. The ALK obtained for each survey was applied tohe stratified length distribution of that survey. Mean lengths atge and a matrix of abundance indices by age group and year werehus obtained.
The age groups used to calculate mean lengths per year werehose which had: (i) at least four age estimations; (ii) containedpecimens throughout most of the length range corresponded tohat age group in the ALK. Thus some old age groups or other poorlyepresented age groups in some years were not considered for thestimation of their respective mean lengths, as their length rangeas covered only partially.
.2. Length-frequency analysis
The length-frequencies were analysed using two methodolo-ies:
(i) Modal class progression analysis (MPA), using the FiSAT IIprogram (Gayanilo et al., 2005) allowed the decompositionof the distributions into their components to identify meanslengths at (relative) age, using the Bhattacharya’s method(Bhattacharya, 1967). MPA was applied on the one hand tothe length-frequency distribution of each survey, and on theother hand, to the grouped distribution resulting of groupingthe individual length-frequency distributions of each surveyinto a unique length distribution covering all the period studied.
ii) MULTIFAN 32 program (Otter Research Ltd.) (Fournier et al.,
1990), that uses a robust maximum likelihood method, wasapplied to the length-frequency distributions of each surveyto obtain the most objective growth parameters. Several mod-els were run, taking into account age-dependent standard(a) number, and (b) biomass for research surveys.
deviation in length at age and different number of age groupsand von Bertalanffy K parameter.
2.3. Tracking cohorts
An exploratory analysis of the matrix of abundance indices atage obtained from ALKs was performed, using the standardizedcatch proportions at age, to check cohorts tracking along the years.Within-survey consistency, i.e. the ability of the survey to trackyear groups, was analysed using correlation coefficients estimatedover years between two consecutive ages from the same cohorts[log(Uay) vs. log(Uaþ1, yþ1), where U is the abundance index, a theage, and y the year]. This offers a first indication of the ability of thesurvey to track year-group strength (Beare et al., 2003).
2.4. Growth parameters
The theoretical growth model to which data are fitted is the vonBertalanffy growth function (VBGF) (von Bertalanffy, 1938).
Lt = L∞(1 − e−k(t−t0))
where Lt is the mean length at age t, L∞ the mean asymptotic length,k the instantaneous growth coefficient, t the age and t0 the age atwhich the mean length would be 0.
Growth parameters from the age estimates were calculatedusing individual lengths at age, for which a fit was made of the“absolute age” of each specimen. This “absolute age” was estimatedas a function of the capture season, adding to the age estimatedthe proportional part of the year (month) in which it was caught.Growth parameters were estimated by a non-linear regressionusing the Levenberg–Marquardt least-squares minimization algo-rithm.
On the other hand, MULTIFAN estimated directly the growthparameters from length-frequency data. In the case of MPA, thegrowth parameters were estimated from the mean lengths at ageobtained by FISAT II. The value of L∞ estimated automatically bythe statistical programme (195 cm) was higher from that observedexperimentally. With the aim of choosing a value of L∞ that fits bet-
ter the lengths at age estimated than those offered automatically bythe statistical programme, the value for L∞ was taken to be relatedto the greatest length observed in the commercial catches duringthe last years in the area. And since L∞ is a mean asymptotic length
3 s Research 137 (2013) 30– 40
vL
2
miCtici
3
ib
vi2eteleet
2a2(ybt
3
i
3
b
icsaab2
m
p(y
3
w
0
10
20
30
40
50
60
70
80
90
100
2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010
Year
Len
gth
(cm
)_
8
7
6
5
4
3
2
1
0
4 J. Landa et al. / Fisherie
alue, that greatest length × 0.95 was selected as L∞ (Taylor, 1962;anda et al., 2002b), and its value was 161 cm.
.5. Slicing
To test if the calculated growth parameters from the age esti-ates could be useful to estimate a reliable matrix of abundance
ndices by age group and year, the slicing method (Kimura andhikuni, 1987) was applied to the stratified length compositions ofhe surveys using growth parameters. A new matrix of abundancendices at age and year was thus obtained, and the standardizedatch proportions at age were estimated to check the cohorts track-ng along the years.
. Results
The geographical distribution of the 736 hauls carried out dur-ng the nine bottom trawl surveys in Porcupine Bank performedetween 2001 and 2009 is shown in Fig. 1.
White anglerfish abundance indices recorded during the sur-eys are summarized in Fig. 3. This species was relatively abundantn 2001 and in 2004, with a drop in abundance in 2002 and since004, maintaining in relative low values during last years. The low-st values were found in 2008; however, it is worthy to point outhat during the surveys that year the gear was not working prop-rly, needing longer time to reach the ground and start trawling,arger horizontal opening and smaller vertical opening the influ-nce of those problems were difficult to estimate; however, it wasvident for most of the species in the survey; therefore, no bias inhe catchability at age can be suspected.
A similar pattern was found for biomass values except for001, when biomass was low, though the high number then wasttributable to the abundance of recruits (Fig. 3). The increase in003 and 2004 was beyond the bootstrap confidence intervals90%) and also the s.e. with respect to 2001 and 2002 (for thoseears only in number), and so was the decrease in 2005 and 2006,ut the abundances remained at similar levels at the beginning andhe end of the time series.
.1. Lengths at age
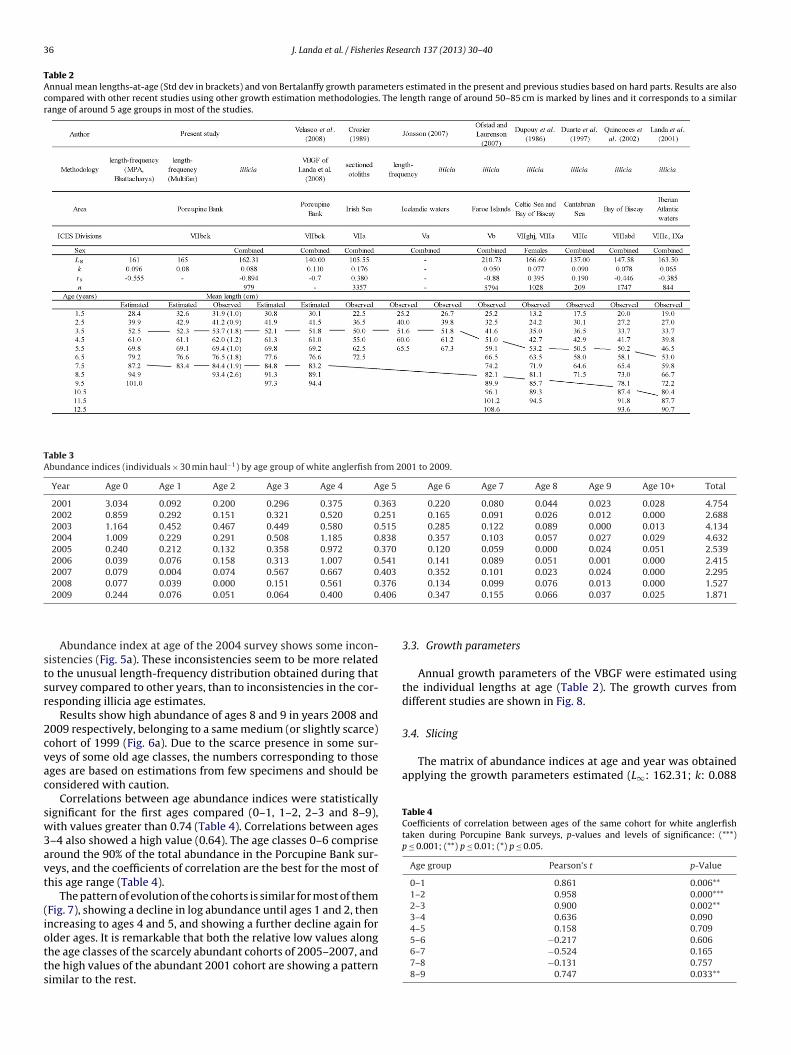
The lengths-at-age obtained from the age estimation were sim-lar to those from both length-frequency analyses (Table 2).
.1.1. Age estimationALK were compiled and the mean lengths at age were calculated
y annual survey from 2001 to 2009 (Fig. 4).Some of the age classes at some years only included a few spec-
mens, mainly belonging to the older ages. For some younger agelasses it was not possible to estimate the mean length, due to thecarce number of age estimates, i.e. age 4 in 2001; ages 5–6 in 2002;ge 0 in 2006; ages 0–1 in 2007 and ages 1–2 in 2008. Some of theseges correspond to the same cohort in different years and it coulde a first indicator of very scarce year classes (e.g., 1997, 2006 and007).
Similar mean lengths were obtained for each age group alongost of the time series (Fig. 4).Overall mean lengths at age were also estimated for the tem-
oral series as the mean of the mean lengths at age of all yearsTable 2). The standard deviations of the mean lengths at age for allears were also estimated.
.1.2. Length-frequency analysisThe growth pattern obtained by both length-frequency analyses
as similar (Table 2):
Fig. 4. Mean lengths-at-age by annual survey (2001–2009) obtained from illicia ageestimation of white anglerfish.
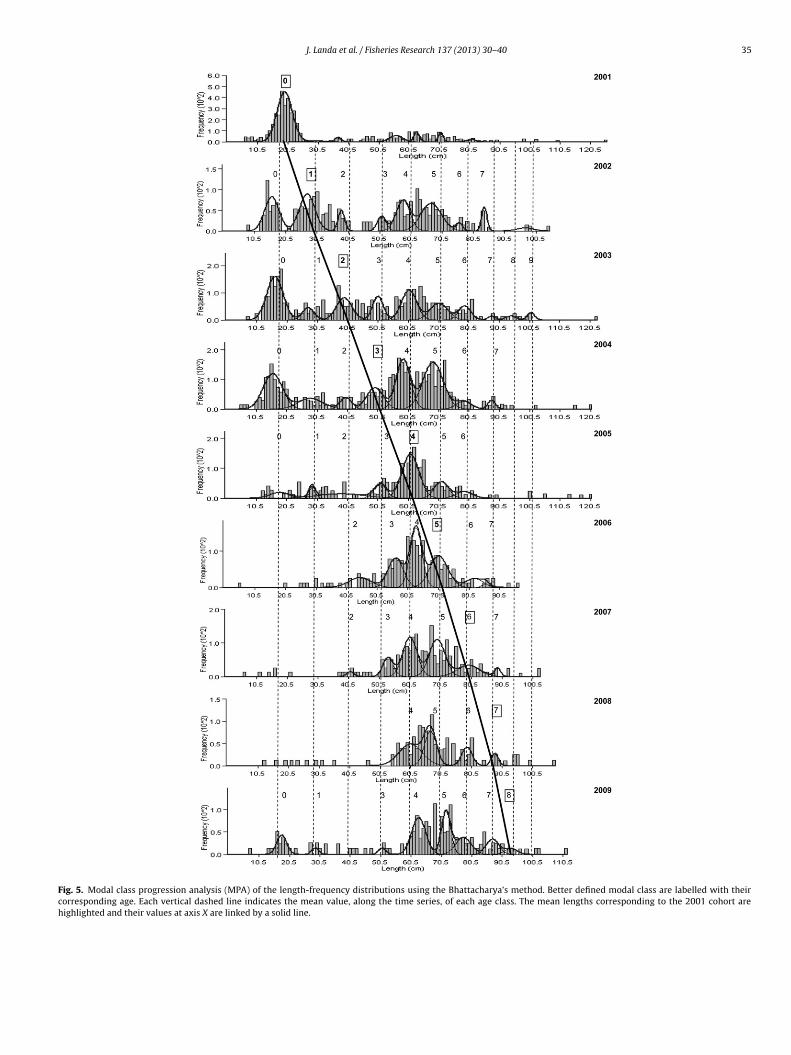
(i) MPA. Length-frequency distributions of each survey weredecomposed into their components, and their mean lengthswere estimated. Some modal classes poorly represented insome years was not able to be identified, mainly those corre-sponding to the ages 0, 1 and 2 in years 2006–2008, and thoseolder than 7 years in most of the years (Fig. 5). An overall meanlength of all surveys was estimated for each age class (Table 2).The age classes of the abundant 2001 cohort are highlighted inFig. 5. This cohort is easily tracked up to age 5 in the length-frequencies. Although age 6, belonging to that cohort, is notso clearly unimodal in 2007, there is a small and isolated clearmode in 2008 corresponding to the following age 7.
Regarding the grouped length-frequency, their mean lengthsestimated were very similar to the aforementioned overallmean lengths of all surveys.
(ii) MULTIFAN. The best fitting model included age-dependentstandard deviation in length-at-age that decreases with age.The growth parameters were chosen based on the best fits ofthe theorical modes to the length distribution and the mostrealistic L∞ values offered by the program (Table 2).
3.2. Tracking cohorts
The matrix of the abundance indices by age group and year(Table 3) was obtained after applying the ALK of each survey tothe respective stratified length distribution.
The growth pattern here estimated can be considered indi-rectly validated by tracking cohort abundance indices in the surveys(Fig. 6a), in addition to the two aforementioned length-distributionanalyses. Two main kinds of cohorts can be tracked:
(i) Abundant cohorts from 2001 to 2004. The abundance of the2003 cohort seems to be lower than those ones (Fig. 6a).
(ii) Scarce cohorts, as those of 2005–2007 and those of 1994–1998.Although the available time series covers a decade, only partof those cohorts can be tracked. The 1997 and 2006 cohortsare highlighted being the scarcest ones in the temporal series(Fig. 6a).
The presence of groups of consecutive cohorts with similar abun-dance indices is highlighted, as the aforementioned 2001–2004 or2005–2007.
J. Landa et al. / Fisheries Research 137 (2013) 30– 40 35
Fig. 5. Modal class progression analysis (MPA) of the length-frequency distributions using the Bhattacharya’s method. Better defined modal class are labelled with theircorresponding age. Each vertical dashed line indicates the mean value, along the time series, of each age class. The mean lengths corresponding to the 2001 cohort arehighlighted and their values at axis X are linked by a solid line.
36 J. Landa et al. / Fisheries Research 137 (2013) 30– 40
Table 2Annual mean lengths-at-age (Std dev in brackets) and von Bertalanffy growth parameters estimated in the present and previous studies based on hard parts. Results are alsocompared with other recent studies using other growth estimation methodologies. The length range of around 50–85 cm is marked by lines and it corresponds to a similarrange of around 5 age groups in most of the studies.
Table 3Abundance indices (individuals × 30 min haul−1) by age group of white anglerfish from 2001 to 2009.
Year Age 0 Age 1 Age 2 Age 3 Age 4 Age 5 Age 6 Age 7 Age 8 Age 9 Age 10+ Total
2001 3.034 0.092 0.200 0.296 0.375 0.363 0.220 0.080 0.044 0.023 0.028 4.7542002 0.859 0.292 0.151 0.321 0.520 0.251 0.165 0.091 0.026 0.012 0.000 2.6882003 1.164 0.452 0.467 0.449 0.580 0.515 0.285 0.122 0.089 0.000 0.013 4.1342004 1.009 0.229 0.291 0.508 1.185 0.838 0.357 0.103 0.057 0.027 0.029 4.6322005 0.240 0.212 0.132 0.358 0.972 0.370 0.120 0.059 0.000 0.024 0.051 2.5392006 0.039 0.076 0.158 0.313 1.007 0.541 0.141 0.089 0.051 0.001 0.000 2.4152007 0.079 0.004 0.074 0.567 0.667 0.403 0.352 0.101 0.023 0.024 0.000 2.2952008 0.077 0.039 0.000 0.151 0.561 0.376 0.134 0.099 0.076 0.013 0.000 1.527
.406
stsr
2cvac
sw3avt
(iotts
3.4. Slicing
The matrix of abundance indices at age and year was obtainedapplying the growth parameters estimated (L∞: 162.31; k: 0.088
Table 4Coefficients of correlation between ages of the same cohort for white anglerfishtaken during Porcupine Bank surveys, p-values and levels of significance: (***)p ≤ 0.001; (**) p ≤ 0.01; (*) p ≤ 0.05.
Age group Pearson’s t p-Value
0–1 0.861 0.006**1–2 0.958 0.000***2–3 0.900 0.002**3–4 0.636 0.0904–5 0.158 0.709
2009 0.244 0.076 0.051 0.064 0.400 0
Abundance index at age of the 2004 survey shows some incon-istencies (Fig. 5a). These inconsistencies seem to be more relatedo the unusual length-frequency distribution obtained during thaturvey compared to other years, than to inconsistencies in the cor-esponding illicia age estimates.
Results show high abundance of ages 8 and 9 in years 2008 and009 respectively, belonging to a same medium (or slightly scarce)ohort of 1999 (Fig. 6a). Due to the scarce presence in some sur-eys of some old age classes, the numbers corresponding to thoseges are based on estimations from few specimens and should beonsidered with caution.
Correlations between age abundance indices were statisticallyignificant for the first ages compared (0–1, 1–2, 2–3 and 8–9),ith values greater than 0.74 (Table 4). Correlations between ages
–4 also showed a high value (0.64). The age classes 0–6 compriseround the 90% of the total abundance in the Porcupine Bank sur-eys, and the coefficients of correlation are the best for the most ofhis age range (Table 4).
The pattern of evolution of the cohorts is similar for most of themFig. 7), showing a decline in log abundance until ages 1 and 2, thenncreasing to ages 4 and 5, and showing a further decline again for
lder ages. It is remarkable that both the relative low values alonghe age classes of the scarcely abundant cohorts of 2005–2007, andhe high values of the abundant 2001 cohort are showing a patternimilar to the rest.0.347 0.155 0.066 0.037 0.025 1.871
3.3. Growth parameters
Annual growth parameters of the VBGF were estimated usingthe individual lengths at age (Table 2). The growth curves fromdifferent studies are shown in Fig. 8.
5–6 −0.217 0.6066–7 −0.524 0.1657–8 −0.131 0.7578–9 0.747 0.033**
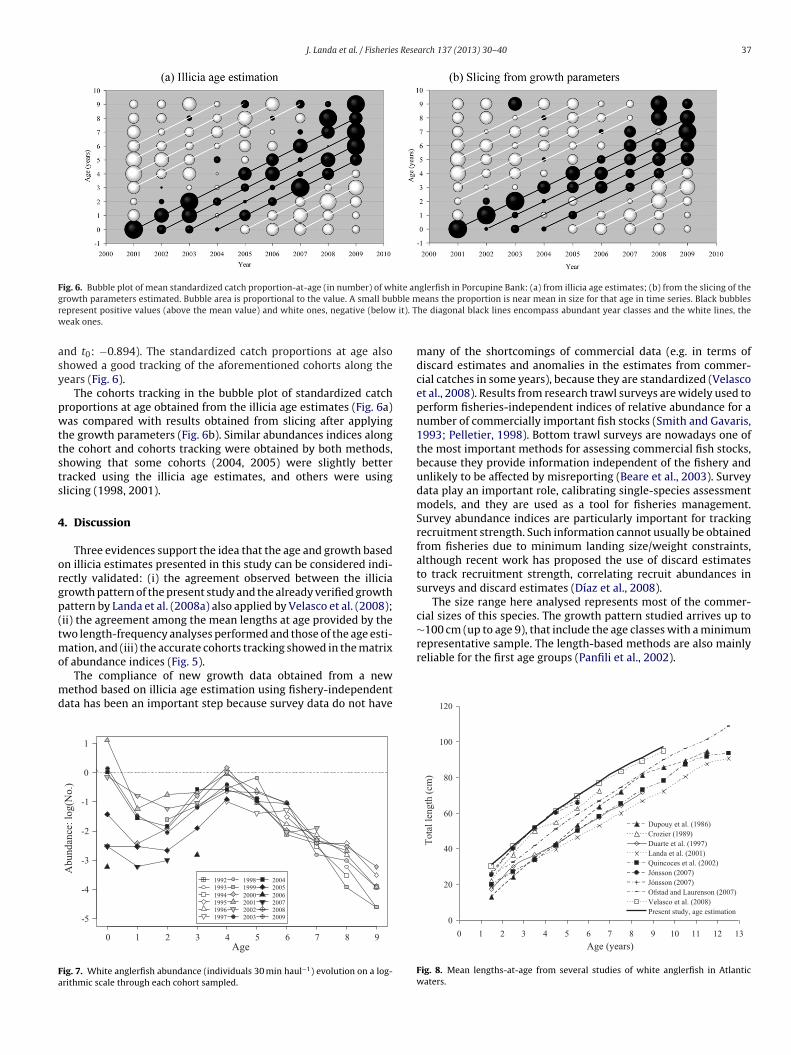
J. Landa et al. / Fisheries Research 137 (2013) 30– 40 37
Fig. 6. Bubble plot of mean standardized catch proportion-at-age (in number) of white anglerfish in Porcupine Bank: (a) from illicia age estimates; (b) from the slicing of theg ble mr it). Tw
asy
pwttsts
4
orgp(tmo
md
Fa
∼100 cm (up to age 9), that include the age classes with a minimumrepresentative sample. The length-based methods are also mainlyreliable for the first age groups (Panfili et al., 2002).
rowth parameters estimated. Bubble area is proportional to the value. A small bubepresent positive values (above the mean value) and white ones, negative (beloweak ones.
nd t0: −0.894). The standardized catch proportions at age alsohowed a good tracking of the aforementioned cohorts along theears (Fig. 6).
The cohorts tracking in the bubble plot of standardized catchroportions at age obtained from the illicia age estimates (Fig. 6a)as compared with results obtained from slicing after applying
he growth parameters (Fig. 6b). Similar abundances indices alonghe cohort and cohorts tracking were obtained by both methods,howing that some cohorts (2004, 2005) were slightly betterracked using the illicia age estimates, and others were usinglicing (1998, 2001).
. Discussion
Three evidences support the idea that the age and growth basedn illicia estimates presented in this study can be considered indi-ectly validated: (i) the agreement observed between the illiciarowth pattern of the present study and the already verified growthattern by Landa et al. (2008a) also applied by Velasco et al. (2008);ii) the agreement among the mean lengths at age provided by thewo length-frequency analyses performed and those of the age esti-
ation, and (iii) the accurate cohorts tracking showed in the matrix
f abundance indices (Fig. 5).The compliance of new growth data obtained from a newethod based on illicia age estimation using fishery-independent
ata has been an important step because survey data do not have
Age
Ab
un
dan
ce:
log
(No
.)
19921993
1994199519961997
19981999
2000200120022003
20042005
2006200720082009
0 1 2 3 4 5 6 7 8 9
-5
-4
-3
-2
-1
0
1
ig. 7. White anglerfish abundance (individuals 30 min haul−1) evolution on a log-rithmic scale through each cohort sampled.
eans the proportion is near mean in size for that age in time series. Black bubbleshe diagonal black lines encompass abundant year classes and the white lines, the
many of the shortcomings of commercial data (e.g. in terms ofdiscard estimates and anomalies in the estimates from commer-cial catches in some years), because they are standardized (Velascoet al., 2008). Results from research trawl surveys are widely used toperform fisheries-independent indices of relative abundance for anumber of commercially important fish stocks (Smith and Gavaris,1993; Pelletier, 1998). Bottom trawl surveys are nowadays one ofthe most important methods for assessing commercial fish stocks,because they provide information independent of the fishery andunlikely to be affected by misreporting (Beare et al., 2003). Surveydata play an important role, calibrating single-species assessmentmodels, and they are used as a tool for fisheries management.Survey abundance indices are particularly important for trackingrecruitment strength. Such information cannot usually be obtainedfrom fisheries due to minimum landing size/weight constraints,although recent work has proposed the use of discard estimatesto track recruitment strength, correlating recruit abundances insurveys and discard estimates (Díaz et al., 2008).
The size range here analysed represents most of the commer-cial sizes of this species. The growth pattern studied arrives up to
0
20
40
60
80
100
120
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Age (years)
Tota
l le
ngth
(cm
)_
Dupouy et al. (1986)
Crozier (1989)
Duarte et al. (1997)
Landa et al. (2001)
Quincoces et al. (2002)
Jónsson (2007)
Jónsson (2007)
Ofstad and Laurenson (2007)
Velasco et al. (2008)
Present study, age estimation
Fig. 8. Mean lengths-at-age from several studies of white anglerfish in Atlanticwaters.

3 s Rese
4
comi(msoec1apsscTrtagwoapr2fiss
tafdcsmehfieaod4t3trcnitda
4
pu
8 J. Landa et al. / Fisherie
.1. Lengths at age
Regarding the growth pattern estimated in different studies, alear similarity in the mean lengths at age is observed among mostf them (Table 2), although several methodologies for growth esti-ation were used. In a range of around 5 age groups (ages 3–7
n the present study), the species growth from ∼50 cm to ∼85 cmTable 2). A clear difference among those studies is that the afore-
entioned length range corresponds to ages 5–9 (or 6–11) in thetudy based on illicia of Dupouy et al. (1986) and in the subsequentnes where the same age estimation criterion was used (Duartet al., 1997; Quincoces et al., 1998; Landa et al., 2001) (Table 2). Thisriterion was standardised after several workshops (Anon, 1997,999). Other recent study based on illicia age estimations (Ofstadnd Laurenson, 2007) showed similar mean lengths at all ages com-ared to those of the present study, with the only difference that aupposed first false annulus seem to be counted as a true one in thattudy. This is likely the reason that the length range of 50–85 cmorresponds to ages 4–8 for Ofstad and Laurenson (2007) (Table 2).he study of Crozier (1989) based on sectioned otolith, and theecent studies of Jónsson (2007) and Velasco et al. (2008) showedhat the aforementioned length range correspond to the ages 3–7,s the present study also shows. Velasco et al. (2008) used therowth parameters previously estimated by Landa et al. (2008a),ho had previously used the available verified evidences of growth
f this species (micro-increments in hard parts, tagging-recapturend length-frequency distributions of catches) to estimate growtharameters. As the parameters estimation was based on growthates, it was not possible to estimate the t0 value (Landa et al.,008a). Velasco et al. (2008) estimated a t0 value, being the reasonor the different mean lengths at age obtained between both stud-es (Table 2). The mean lengths at age of the present study are veryimilar to those of Velasco et al. (2008), the only previous growthtudy in Porcupine Bank.
The similar mean lengths at a common age range amonghe studies based on illicia (including this work) confirms that,lthough the age estimation criterion used in former studies is dif-erent that the criterion presented here, the difference is mainlyue to the counting or not of the first increments previous to thatommon length range (<50 cm). This evidence and the similar con-tant difference in age classes among some studies from age 2 areore clearly observed in Fig. 8. Regarding the first annulus, Wright
t al. (2002) studied the illicium microincrements and verified aigh initial growth rate in the 0 age class (0.91 mm d−1) during therst 8 months of life. Landa et al. (2008a) summarised that and othervidences from the growth of white anglerfish in Atlantic waters,nd concluded that the white anglerfish could reach a mean lengthf around 27 cm at age 1, and around 39–40 cm at age 2, at the mid-le of the year. The present study showed similar lengths of 31 and1 cm for age 1 and 2 respectively, in September, in agreement tohe verified growth pattern of Landa et al. (2008a). From age 1 to, this work shows a growth rate of around 10–12 cm, decreasingo a growth rate of around 8–9 cm up to age 8. An annual growthate of 7–9 cm in the aforementioned length range of 50–85 cm isommon for all studies (Table 2). These results confirm that it isot relatively difficult to distinguish the annual increments on ill-
cia from medium ages (Landa et al., 2008a), but it is more difficulto distinguish the first annual increments. More verified and vali-ated information on age and growth is necessary to improve theccuracy in growth estimations, especially in older ages.
.2. Tracking cohorts
The scarcely abundant cohorts in recent years (2005–2008),roduced by low recruitments, can lead to a decreasing of the pop-lation abundance on Porcupine Bank in the following years, as the
arch 137 (2013) 30– 40
abundant cohorts of the years 2001–2004 disappear of the popula-tion. A decreasing trend of the population abundance and biomassis still being observed from 2004 (Fig. 3).
This study confirms the abundance increase in the intermediateage classes (mainly, 3–5), already observed by Velasco et al. (2008)and which seems related to immigration to the Bank of individualsof these ages. This fact may have influence on the low correlationvalues observed between the successive intermediate age classes(4–8) (Table 4 and Fig. 7). However, a good correlation in the abun-dances indices of the first four age classes (0–4) is observed, with aprogressive decrease in the abundance. The decreasing in the abun-dance with the age, but along of the all ages, has been observed inother more sedentary species as the megrim and four-spot megrim(Sánchez et al., 1998). Therefore, these first age classes may have amore sedentary behaviour on the bank, compared to the interme-diate ones.
Regarding the cohort tracking, if an exceptionally good or weakyear class can be followed over a long period of time throughthe age composition, this provides is a strong indication that theage-reading method is accurate (Panfili et al., 2002). The good cor-relations in the abundances indices obtained among the first agescompared support the cohort tracking observed in Fig. 6a. Thegraphical analyses (Fig. 6a) uses the standardized catch propor-tions at age and therefore avoids the problems of low correlationin the intermediate age classes due to their relative higher abun-dance. The efficient cohort tracking of the abundance indices ofwhite anglerfish (Fig. 6a and b) confirms (i) that these annual sur-veys at Porcupine Bank are extremely reliable to monitor the whiteanglerfish population abundance, and (ii) that the new illicia ageestimation criterion used can be considered valid. Velasco et al.(2008) studied the growth of a part (2001–2006) of the here ana-lysed time series, using the slicing methodology applied to thefaster growth parameters of Landa et al. (2008a). The present resultsagree with most results of Velasco et al. (2008), as, for instance, theevidence of the abundant 2001 year class that was tracked up toage 5 in 2006, and the low recruitments that began to be detectedfor 2005 and 2006. Here, using a longer time series (up to 2009),the tracking of those cohorts observed in Velasco et al. (2008) hasbeen corroborated using age estimations.
The indirect validation using cohort tracking was success-fully performed in other species, as horse mackerel or roughheadgrenadier, mainly due to an only extremely abundant cohort wasperfectly tracked (Abaunza et al., 2003; Santiago and Arrizabalaga,2005). Tracking the very scarce 2006 cohort detected for the whiteanglerfish in this study, during the next years, might also add a newevidence to corroborate the validation of the illicia age estimationcriterion (Fig. 6).
The white anglerfish in the Porcupine Bank is part of the pop-ulation managed as the northern stock of the European Atlanticsouthern shelf of this species. Velasco et al. (2008) indicated that theagreement in the detection of the same extreme good and scarcerecruitments, as those of 2001, 2005 and 2006, by several Euro-pean Atlantic bottom-trawl surveys that cover different areas ofthe stock, corroborates the consistency among these surveys. Thenew scarce recruitments from 2007 to 2009 detected in the presentstudy, and the good tracking along the cohorts of the aforemen-tioned ones in those surveys, may also confirm the consistencyamong these surveys if also a similar pattern of recruitments isobtained.
However it is important to take into account the growth plas-ticity and its influence by factors that vary geographically, astemperature or food, mainly. This study shows the growth pattern
in a restricted area of the distribution of white anglerfish. Geo-graphical movements of white anglerfish have been described forseveral areas of the European Atlantic (Pereda and Landa, 1997;Hislop et al., 2000; Arkhipov and Mylnikov, 2002; Laurenson et al.,
s Rese
2eott2app2lafmiPisstfa
4
atltiesl(LiOm
asaocaasfr(ugaQ
4
opaatte
J. Landa et al. / Fisherie
005; Landa et al., 2008b). Laurenson et al. (2005) and Landat al. (2008b) showed also interaction between Atlantic stocksf white anglerfish. Charrier et al. (2006) related the ecologicalraits of dispersion of this species with its limited genetic struc-ure observed (Crozier, 1987; O’Sullivan et al., 2005; Charrier et al.,006), although Blanco et al. (2008) also suggested that whitenglerfish may also have a more complex genetic structure of theiropulations. Those evidences of geographical movements and dis-ersion, and the high gene flow within populations (Farina et al.,004; Charrier et al., 2006) could support the idea that some bio-
ogical features, as the growth pattern of this species, could not bes different along of its geographical distribution as can be foundor other more sedentary demersal species. In some flatfishes, as
egrim and four spot megrim, significant geographical differencesn growth in the European southern shelf were found (Landa andineiro, 2000; Landa et al., 2002b). The evidences of similar growthn different Atlantic areas showed by Landa et al. (2008a) seemupport that idea. This also could indicate that it might be noto risky to try to use the growth parameters estimated here, orhose overall ones of Landa et al. (2008a), for the white anglerfishrom other areas where the age estimations are not available andge-structured models are used for the stock assessment.
.3. Growth parameters
Zivkov et al. (1999) considered that, in several studies, the vari-tions on growth parameters do not correspond with those ofhe growth rates. These authors recommended the comparison ofengths at age of a common range of ages instead the comparison ofhe growth parameters, as a more simple and precise method. Thiss indeed that has been performed and discussed in this work. Nev-rtheless growth parameters were also estimated in the presenttudy and compared with those of previous ones, showing a simi-ar slope for most of them. The mean asymptotic length estimatedL∞) should be close to the maximum length observed. Values of∞ between 140 and 170 cm are estimated in most of the stud-es for white anglerfish, however in other studies (Crozier, 1989;fstad and Laurenson, 2007) its value is not so close to the observedaximum length of this species.On the other hand when the growth parameters are compared
mong studies, it should take into account the inverse relation-hip between L∞ and k. Moreover, although similar mean lengthst age can be estimated in some studies (Table 2); different valuesf the growth parameters can be calculated. For those reasons theomparison of the growth parameters among studies is not easy,nd therefore the comparison performed among mean lengths atge can be considered more suitable. Nevertheless, some conclu-ion can be obtained. Thus, considering only the studies with theaster growth pattern, values of L∞ of around 140 and 160 cm cor-espond to values of k of around 0.11 and 0.08–0.09, respectivelypresent study; Velasco et al., 2008; Landa et al., 2008a). These k val-es are higher than those estimated in other studies with slowerrowth pattern (0.09, 0.08 and 0.07 for L∞ values of around 140, 150nd 160 cm, respectively) (Duarte et al., 1997; Landa et al., 2001;uincoces, 2002).
.4. Slicing
Regarding the better tracking of some cohorts using the slicingf growth parameters compared to the illicia age estimations, it isossible that the scarce illicia available in some age groups did notllow obtain an adequate representation of age estimates in those
ge groups. However, these lacks might have been overcome usinghe growth parameters. Annual ALKs are necessary to perform forhe stock assessment due mainly to the variation in the strength ofach cohort. However the aforementioned evidence could indicatesarch 137 (2013) 30– 40 39
that annual variations in the growth pattern of this species are notso evident as to have a preference of using the annual age estima-tions compared to the use of the growth parameters. It seems thatboth approaches could be considered acceptable. However it couldbe interesting to compare if similar results are obtained in otherareas when both methodologies are applied.
Acknowledgements
We are grateful to Enrique Rodríguez-Marín, Lucía Canas andCelso Farina for their collaboration. We thank the crews of RV “Viz-conde de Eza” and the scientific teams of IEO, the Marine Institute,and AZTI that made the sampling surveys possible. We thank Cov-adonga Orejas, Wolf Isbert, three anonymous reviewers and theeditor for the revision of the manuscript and their helpful sugges-tions. The Spanish surveys on Porcupine Bank are supported bythe Instituto Espanol de Oceanografía and by the Data CollectionFramework for the EU Common Fisheries Policy.
References
Abaunza, P., Gordo, L., Karlou-Riga, C., Murta, A., Eltink, A., Santamaria, M.T.G., Zimm-ermann, C., Hammer, C., Lucio, P., Iversen, S.A., Molloy, J., Gallo, E., 2003. Growthand reproduction of horse mackerel, Trachurus trachurus (Carangidae). Rev. FishBiol. Fish. 13 (1), 27–61.
Anon, 1997. International ageing workshop on European monkfish. Int. Doc. IFRE-MER, 34pp.
Anon., 1999. Third International Ageing Workshop on European Anglerfish (Lisbon,March 8–12, 1999). Int. Doc. IPIMAR, 106pp.
Arkhipov, A., Mylnikov, N., 2002. On the pelagic phase of the life cycle of the commonangler (Lophius piscatorius Linne) around seamounts of the Rockall-Hatton area.ICES Document CM 2002/M: 33.
Azevedo, M., Cardador, F., Costas, G., Duarte, R., Farina, A.C., Landa, J., Sampedro,M.P., 2008. Final Report: Improving the quality of southern anglerfish stocksassessment (ABA), (UE DG FISH/2004/03-22).
Bhattacharya, C.G., 1967. A simple method of resolution of a distribution into Gauss-ian components. Biometrics 23, 115–135.
Beare, D., Castro, J., Cotter, J., van Keeken, O., Kell, L., Laurec, A., Mahé, J.C., et al., 2003.Final report: evaluation of research surveys in relation to management advice,(EVARES) UE DG FISH/2001/02).
Blanco, G., Borrell, Y.J., Cagigas, M.E., Vazquez, E., Prado, J.A.S., 2008. Microsatellites-based genetic analysis of the Lophiidae fish in Europe. Mar. Freshwater Res. 59(10), 865–875.
Charrier, G., Chenel, T., Durand, J.D., Girard, M., Quiniou, L., Laroche, J., 2006. Dis-crepancies in phylogeographical patterns of two European anglerfishes (Lophiusbudegassa and Lophius piscatorius). Mol. Phylogenet. Evol. 38, 742–754.
Crozier, W.W., 1987. Biochemical genetic variation and population structure inangler-fish Lophius piscatorius L. from the Irish Sea and west of Scotland. J. Exp.Mar. Biol. Ecol. 106, 125–136.
Crozier, W.W., 1989. Age and growth of angler-fish (Lophius piscatorius L.) in theNorth Irish Sea. Fish. Res. 7, 267–278.
Dardignac, J., 1988. Les pêcheries du Golfe de Gascogne. Bilan des connaissances.Rapports Scientifiques et Techniques de l’Ifremer 9, 111–130.
Díaz, P., Santos, J., Velasco, F., Serrano, A., Pérez, N., 2008. Anglerfish discard estimatesand patterns in Spanish Northeast Atlantic trawl fisheries. ICES J. Mar. Sci. 65,1350–1361.
Dobby, H., 2002. A length-based assessment of anglerfish in Division VIa: develop-ments in growth modelling. Working Document for the Working Group on theAssessment of Northern Shelf Demersal Stocks.
Duarte, R., Azevedo, M., Pereda, P., 1997. Study on the growth of southern black andwhite monkfish stocks. ICES J. Mar. Sci. 54, 866–874.
Duarte, R., Landa, J., Quincoces, I., Dupouy, H., Bilbao, E., Dimeet, J., Marc al, A., et al.,2002. Anglerfish Ageing Guide. In: Report of the 4th International Ageing Work-shop on European Anglerfish, 40pp.
Duarte, R., Landa, J., Morgado, C., Marc al, A., Warne, S., Barcala, E., Bilbao, E., et al.,2005. Report of the Anglerfish Illicia/Otoliths Ageing Workshop. IPIMAR, Lisbon,47pp.
Dupouy, H., Pajot, R., Kergoat, B., 1986. Etude de la croissance des baudroies Lophiuspiscatorius et L. budegassa, de L’Atlantique Nord-Est obtenue a’ partir de l’illicium.Revue des Travaux de l’Institut des Pêches Maritimes 48, 107–131.
Farina, A.C., Duarte, R., Landa, J., Quincoces, I., Sánchez, J.A., 2004. Multiple stockidentification approaches of anglerfish (Lophius piscatorius and L. budegassa) inwestern and southern European waters. ICES Document CM 2004/EE: 25.
Fournier, D.A., Sibert, J.R., Majkowsky, J., Hampton, J., 1990. MULTIFAN: a likelihood-based method for estimating growth parameters and age composition from
multiple length-frequency data sets, illustrated using data for Southern BluefinTuna (Thunnus maccoyii). Can. J. Fish. Aquat. Sci. 47, 301–317.Fulton, T.W., 1903. The Distribution, Growth, and Food of the Angler (Lophius pis-catorius). Fishery Board for Scotland, pp. 186–199 (Part III. Twenty-first AnnualReport).

4 s Rese
G
H
I
I
I
I
J
K
L
L
L
L
L
L
L
L
L
O
O
P
0 J. Landa et al. / Fisherie
ayanilo Jr., F.C., Sparre, P., Pauly, D., 2005. FAO-ICLARM Stock Assessment Tools II(FiSAT II). Revised version. User’s guide. FAO Computerized Information Series(Fisheries), No. 8, Revised version. FAO, Rome, p. 168.
islop, J.R.G., Holst, J.C., Skagen, D., 2000. Near-surface captures of post-juvenileanglerfish in the north-east Atlantic – an unresolved mystery. J. Fish Biol. 57,1083–1087.
CES, 2010a. Report of the International Bottom Trawl Survey Working Group(IBTSWG), 22–26 March 2010 Lisbon, Portugal. ICES CM 2010/SSGESST:06,261pp.
CES, 2010b. Addendum 2: IBTS manual on the Western and Southern areas 2010.Revision III. Agreed during the meeting of the International Bottom Trawl SurveyWorking Group, 22–26 March 2010, Lisbon, Portugal, 58pp.
CES, 2011a. Report of the Working Group on the Assessment of Southern Shelfstocks of Hake, Monk and Megrim (WGHMM), 5–11 May 2011, Copenhagen,Denmark, ICES CM 2011/ACOM:11, 625pp.
CES, 2011b. Report of the Working Group for Celtic Seas Ecoregion (WGCSE), 11–19May 2011, Copenhagen, Denmark. ICES CM 2011/ACOM:12, 1573pp.
ónsson, E., 2007. Verification of anglerfish (Lophius piscatorius) age estimationthrough comparison of length modes of age read fish (illicia) to length modes oflarge year-classes appearing in the Icelandic stock, ICES CM 2007/K: 03.
imura, D.K., Chikuni, S., 1987. Mixtures of empirical distributions—an iterativeapplication of the age-length key. Biometrics 43 (1), 23–35.
anda, J., 2004. Attempts of growth validation in Lophius piscatorius. In: Duarte, R.,Landa, J., Morgado, C., Marc al, A., Warne, S., Barcala, E., Bilbao, E., et al. (Eds.),Report of the Anglerfish Illicia/Otoliths Ageing Workshop. IPIMAR, Lisbon, pp.32–37, 47pp.
anda, J. 2012. Report of the Anglerfish (Lophius piscatorius) illicia and otolithsexchange 2011, 61 pp. http://groupnet.ices.dk/AgeForum/ExchangesReports
anda, J., Duarte, R., Quincoces, I., 2008a. Growth of white anglerfish (Lophius pisca-torius) tagged in the Northeast Atlantic, and a review of age studies on anglerfish.ICES J. Mar. Sci. 65, 72–80.
anda, J., Duarte, R., Quincoces, I., Dupouy, H., Bilbao, E., Dimeet, J., Lucio, P., Marc al,A., McCormick, H., Ni Chonchuir, G., 2002a. Report of the 4th International AgeingWorkshop on European Anglerfish , IPIMAR, Lisbon, 133pp.
anda, J., Pérez, N., Pineiro, C., 2002b. Growth patterns of the four spot megrim(Lepidorhombus boscii) in the northeast Atlantic. Fish. Res. 55, 141–152.
anda, J., Pereda, P., Duarte, R., Azevedo, M., 2001. Growth of anglerfish (Lophiuspiscatorius and L. budegassa) in Atlantic Iberian waters. Fish. Res. 51, 363–376.
anda, J., Pineiro, C., 2000. Megrim (Lepidorhombus whiffiagonis) growth in the North-eastern Atlantic based on back-calculation of otolith rings. ICES J. Mar. Sci. 57,1077–1090.
anda, J., Quincoces, I., Duarte, R., Farina, A.C., Dupouy, H., 2008b. Movements ofblack and white anglerfish (Lophius budegassa and L. piscatorius) in the NortheastAtlantic. Fish. Res. 94 (1), 1–12.
aurenson, C.H., Johnson, A., Priede, I.G., 2005. Movements and growth of monkfishLophius piscatorius tagged at the Shetland Islands, northeastern Atlantic. Fish.Res. 71, 185–195.
fstad, L.H., Laurenson, C., 2007. Biology of anglerfish Lophius piscatorius in Faroesewaters, ICES CM 2007/K: 07.
’Sullivan, M., Wright, P.J., Verspoor, E., Knox, D., Piertney, S., 2005. Absence of spatialand temporal genetic differentiation at microsatellite loci in northeast Atlanticanglerfish (Lophius piscatorius), ICES Document CM 2005/T: 18.
anfili, J., de Pontual, H., Troadec, H., Wright, P.J., 2002. Manual of Fish Sclerochronol-ogy, IFREMER-IRD co-edition. Brest, 464pp.
arch 137 (2013) 30– 40
Pelletier, D., 1998. Intercalibration of research survey vessels in fisheries: a reviewand an application. Can. J. Fish. Aquat. Sci. 55, 2672–2690.
Pereda, P., Landa, J., 1997. Recuperación de dos ejemplares de rape blanco Lophiuspiscatorius Linnaeus, 1758 en el stock norte (divisiones VIII a y b del CIEM)tras ser marcados en el stock sur (división VIII c). Boletín Instituto Espanol deOceanografía 13, 15–24.
Peronnet, I., Dupouy, H., Rivoalen, J.J., Kergoat, B., 1992. Methods of ageing basedon caudal fin-rays for megrim and on sections of illicium for anglerfishes(Lophius piscatorius and L. budegassa). Colloquium National ORSTOM/INRA,Boudy, France, pp 4–6 (1991 March) (Lepidorhombus wiffiagonis).
Prager, M.H., 1994. A suite of extension to a non-equilibrium surplus-productionmodel. Fish. Bull. 92, 374–389.
Quincoces, I., 2002. Crecimiento y reproducción de las especies Lophius budegassaSpinola 1807, y Lophius piscatorius Linneo 1758, del Golfo de Vizcaya. PhD the-sis, Departamento de Zoología y Dinámica Celular Animal, Universidad del PaísVasco, Spain, 258pp.
Quincoces, I., Santurtún, M., Lucio, P., 1998a. Biological aspects of white anglerfish(Lophius piscatorius) in the Bay of Biscay (ICES Divison VIIIa, b, d) in 1996–1997.ICES Document CM 1998/O: 48.
Sánchez, F., Pérez, N., Landa, J., 1998. Distribution and abundance of megrim (Lepi-dorhombus boscii and Lepidorhombus whiffiagonis) in the northern Spanish shelf.ICES J. Mar. Sci. 55, 494–514.
Santiago, J., Arrizabalaga, H., 2005. An integrated growth study for North Atlanticalbacore (Thunnus alalunga Bonn 1788). ICES J. Mar. Sci. 62, 740–749.
Smith, S.J., Gavaris, S., 1993. Improving the precision of abundance estimates ofeastern Scotian Shelf Atlantic cod from bottom trawl surveys. N. Am. J. Fish.Manage. 13, 35–47.
Sullivan, P.J., Lai, H.-L., Gallucci, V.F., 1990. A catch-at-length analysis that incorpo-rates a stochastic model of growth. Can. J. Fish. Aquat. Sci. 47, 184–198.
Taylor, C.C., 1962. Growth equation with metabolic parameters. J. Cons. Int. Explor.Mer. 23, 366–370.
Thangstad, T., Dyb, J.E., Jonsson, E., Chevonne, L., Ofstad, L.H., Reeves, S.A., 2002.Anglerfish (Lophius spp.) in Nordic European Waters. Status of Current Knowl-edge an Ongoing Research. Institute of Marine Research, Bergen, Norway.
Tsimenidis, N.C., Ondrias, J.C., 1980. Growth studies on the angler-fishes Lophiuspiscatorius L., 1758 and Lophius budegassa Spinola, 1807 in Greek waters. Tha-lassographica 3, 63–94.
Velasco, F., Landa, J., Barrado, J., Blanco, M., 2008. Distribution, abundance, andgrowth of anglerfish (Lophius piscatorius) on the Porcupine Bank (west ofIreland). ICES J. Mar. Sci. 65 (7), 1316–1325.
von Bertalanffy, L., 1938. A quantitative theory of organic growth (inquiries ongrowth laws II). Hum. Biol. 10, 181–213.
Whitehead, P.J., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonesse, E., 1986. Fishes ofthe North-eastern Atlantic and the Mediterranean. UNESCO, Paris, 1362pp.
Woodroffe, D.A., Wright, P.J., Gordon, J.D.M., 2003. Verification of annual incrementformation in the white anglerfish, Lophius piscatorius using the illicia and sagittaotoliths. Fish. Res. 60, 345–356.
Wright, P.J., Woodroffe, D.A., Gibb, F.M., Gordon, J.D.M., 2002. Verification ofthe first annulus formation in the illicia and otoliths of white angler-
fish, Lophius piscatorius using otolith microstructure. ICES J. Mar. Sci. 59,587–593.Zivkov, M.T., Trichkova, T.A., Raikova-Petrova, G.N., 1999. Biological reasons for theunsuitability of growth parameters and indices for comparing fish growth. Envi-ron. Biol. Fish. 54 (1), 67–76.





























