Embed Size (px)
Citation preview

Humangenetik 17, 253--265 (1973) © by Springer-Verlag 1973
Age Determination by Immunological Techniques of the Last Common Ancestor of Man and Chimpanzee
Klausdie te r Bauer*
Faculty for Theoretical Medicine, University of Heidelberg, Germany
Received August 16, 1972
Summary. Following on from earlier work in this laboratory the age of the last common ancestor (LCA) of man and chimpanzee was determined by an immunogenetic approach. The determinant patterns of 30 different plasma-protein entities were examined for homologues in human and chimpanzee blood. Comparison of these determinants revealed the number of immunologically relevant accepted mutations separating the two species.
The proteins investigated in this way were then employed as representatives of their corre- sponding strnctural genes, since their polypeptide sequences are direct translations of the latter, i.e. actually two samples of the genomes in question were compared.
The subsequent calculation of the ages of the LCAs made use of previously estimated values for other primate groups including gorilla, orang utah, several Old World monkeys, one New World monkey and one prosimian. We used the same algorithm as in the earlier investigation for the present analysis.
The result indicates that man and chimpanzee shared a common ancestor 9.7 million years ago. This value is lower than those determined for any other species investigated in our laboratory, including gorilla and orang utah. This reaffirms the special position of the chim- panzee among other primates in relation to man, which is otherwise expressed in anatomical, physiological and psychological features. The value given is dependent on the reference point employed to introduce absolute units. The age of the eutherian common ancestor, assumed to be 70 million years, was used for the present study. Relative time scales, dividing the same time interval into 100 relative time units, are better suited to describe the results, but do not allow any comparison with morphologically derived data.
The method employed and the implications of the results obtained are discussed in some detail.
Zusammen/assung. In Fortffihrung friiherer Untersuchungen unseres Arbeitskreises wurdo das Alter des letzten gemeinsamen Vorfahren (LGV) des Menschen und des Sehimpansen mit einer immungenetischen Technik bestimmt. Die Determinantenstrukturen yon 30 Plasma- proteinen wurden jeweils fiir die Homologen der beiden Arten analysiert. Aus ihrem Vergleich ergibt sich die Zahl der immmlologiseh relevanten, akzeptierten Mutationen, in denen sieh die beiden Species unterseheiden.
Da die Polypeptidanteilc der Proteine, welche die Determinanten tragen, direkte ~ber- setzungen der zugehSrigen Strukturgene darstellen, ist die beschriebene Untersuehung zu- gleieh eine solche einer Stiehprobe der betreffcnden Genome.
Fiir die Bereehnung des Alters des LGV yon Menseh und Schimpanse warden unsere frfiher erhaltenen Resultate fiir andere Primatengruppen zum Vergleich herangezogen. Diese umfai~ten Gorilla und Orang Utan, einige Altweltaffen und je einen Neuwelt- und Halbaffen. Wir verwandten den gleichen Algorithmus wic friihcr beschrieben.
Das so erhaltene Alter fiir den LGV yon Mensch und Schimpanse betr~igt 9,7 Mi]lionen Jahre und ist damit kleiner als fiir jeden anderen bisher von uns untersuchten LGV des
* Address for reprints see at the end of this paper.

254 K. Bauer:
Menschen mit einer anderen Art; der Wert is~ auch kleiner Ms fiir die Paare Mensch/Gorilla und Mensch/Orang Utah. Dieses Ergebnis bestitigt die besondere Stellung des Schimpansen unter allen Primaten zum Menschen, wie sie aus anatomischen, physiologischen und psycho- logischen Befunden erschlossen wurde. Zugleich werden die hISglichkeiten, welche die immun- genetische Analyse in der Primatologie und Anthropologie erSffnet, erneut best~tigg.
Introduction
The reconstruction of the evolution of a protein, as it is possible by both bio- chemical and immunological means, at the same time is one of a small fraction of the genomes of the species concerned. Polypeptide chains are direct translations of structural genes; therefore the reconstruction of the underlying DNA-sequence is possible to a certain extent at least by the reverse procedure, with the limitations due to the code degeneracy. By comparative analyses of a number of homologues, common ancestor sequences, so-called nodal sequences, can be reconstructed (Day- hoff, 1969). This is again feasible at both the polypcptide and, from it, at the nucleic acid levels, i.e. the elucidation of portions of genomes from ancestor species living at different phylogenetic periods is possible by this approach. This allows a very basic investigation of evolutionary events at the molecular level (Dayhoff, 1969; Nolan and Margoliash, 1968; S6derqvist and Blomb~ek, 1971).
Unfortunately the techniques used for sequencing are rather sophisticated and require the isolation of the protein in a highly purified state. This is why only relatively few of the variety of proteins existing in higher organisms have been sequenced in a sufficiently large number of species to allow comparative analyses. Among the proteins which have been more thoroughly examined with respect to their evolution are haemoglobins and myoglobins, immunoglobulins and fibrino- peptides, corresponding to not more than a few thousand DNA base positions, certainly a very small statistical sample in relation to the whole genome. These data are highly interesting for the evolutionary history of these protein entities themselves and give a first insight into mechanisms which until recently have only been hypotheses. Moreover they are in general agreement with the taxonomic relationships of the species involved. They do not, however, open a conclusive way to the numerical investigation of the relationships between whole genomes and the organisms they code. For this purpose the fraction of genetic information elucidated this way, the statistical sample, is certainly too small. Our opinion on this subject is very clearly expressed in Dr. Buettner-Janusch's statement: "Hemoglobin i s . . . n i c e . . , to work w i t h . . , but I would like to examine 30 pro- reins before I made a taxonomic decision" (Buettner-Janusch, 1965).
What is needed is a more rapid and simpler method of compiling comparative data on homologues of such a sample of different proteins in a sufficiently large number of species in a relatively short period of time. Such an approach has be- come feasible since several authors have introduced modern immunochemical techniques into phylogenetie research (Goodman, 1964; Goodman et al., 1970; Bauer, 1969, 1970c). Antisera, which are the analytical tools in this case, react with a variety of different surface areas on the antigen molecule, the antigenic determinants. In proteins they consist of a small number of amino acids, either forming part of the sequence (sequence determinants) or being more or less separated on the chain when unfolded, while forming one (conformation) determi-

Lass Common Ancestor of Man and Chimpanzee 255
nant in the native molecule (Crumpton, 1966). I t is evident tha t the determinant structure and amino acid sequence are colinear, because of the influence of con- formation in a somewhat modified sense, and tha t therefore determinant structure and the corresponding straetural gene are also colinear. Thus comparative analyses of the determinant patterns of homologous proteins can serve as a pa thway to the investigation of the genome itself (Bauer, 1970d).
Much simpler methods can be used to estimate to some extent the relationships among species. G. H. F. Nuttal l compiled a very large body of data on simple in vitro precipitation experiments embracing a great many species as early as 1904; he used antisera to whole blood serum from one species and measured the amount of precipitate these antisera produced with serum from other species. This type of experimentation was later continued on a small scale by Mollison (1923). While these authors demonstrated beyond doubt tha t immunological cross- reactions are strongly influenced by the taxonomic relationship between the species the antiserum is originally produced to and the one under investigation, they did not by any means introduce the approach necessary to estimate the relationship between the genomes in a more exact, especially in a numerical way.
This fact was very clearly demonstrated by Wilson et al. (1964), who observed tha t the same cross-reaction - - i.e. with the same antiserum and the same species constellation - - yielded 41, 86, and 100% of the homologous reaction in one ease and even 3, 89 and 96% in another example (human haemoglobins A 1 and S and ovalbumin from chicken and turkey), depending solely on the method used. The techniques employed by these authors included two types of complement-fixation test and quanti tat ive precipitation. I t is evident that the strength of a cross-reaction between homologues, when determined by one of these methods, is a function of the technique rather than one of the taxonomic relationship of the species involved. We therefore think that age determinations or other numerical taxonomic parameters for the last common ancestors (LCAs) as calculated e.g. by Sarich and Wilson (1967) using a complement-fixation method lack any apparent connexion with the genome and thus with the evolution of the species in question.
We have previously applied the principle shortly discussed here, namely com- parat ive determinant analysis - - which we think to be the only method presently available for deriving evolutionary parameters such as ages of LCAs from immuno- logical data - - to an investigation of primate evolution aimed at estimating the ages of the LCAs of different primate groups and man (Bauer, 1970d, 1973). This s tudy was based on a sample of 28 plasma proteins. The age values of the LCAs of man on one side and gorilla, orang utan, several Old World monkeys, one New World monkey and one prosimian were in good agreement with fossil evidence.
In the present communication we report on the investigation of chimpanzee plasma proteins undertaken to include this species in the two t ime scales of primate evolution derived previously; these two scales differ in the type of units t ime is expressed by. The methods used were essentially the same as for the other apes and the monkeys. The results once more indicate the special position of this species among primates as to its relationship with man, as the LCA's age for man and chimpanzee was found to be lower than for any other species so far examined in our laboratory, including apes and monkeys; at the same time it was still in the range indicated by morphological evidence.
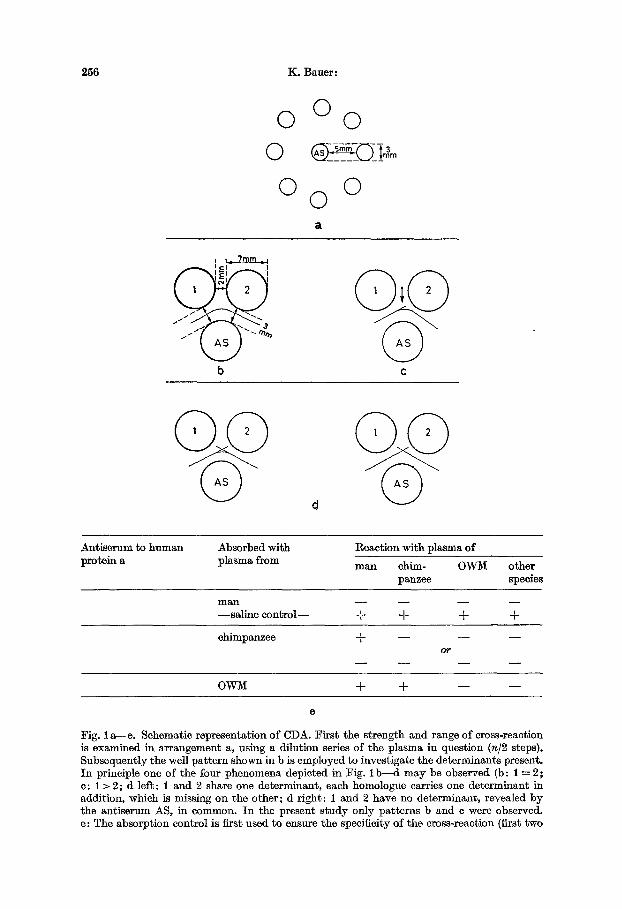
256 K. Bauer:
O O 0 0
0 0 0
', ~_ , ,~ . I
b c
Antiserum ~o h u m a n protein a
Absorbed wi th plasma from
React ion with plasma of
man chim- OW1VI o ther panzee species
m a n . . . .
- - s a l ine c o n t r o l - - + A- -4- -4-
chimpanzee + - - - - - - 05"
OWI~I + + - - - -
e
Fig. 1 a - -e . Schematic representat ion of CDA. Firs t the s t rength and range of cross-reaction is examined in ar rangement a, using a dilution series of the plasma in question (n/2 steps). Subsequently the well pa t t e rn shown in b is employed to investigate the determinants present. I n principle one of the four phenomena depicted in Fig. i b - ~ l may be observed (b: 1 ~ 2 ; c: 1 > 2; d left: 1 and 2 share one determinant , each homologue carries one de te rminant in addition, which is missing on the o ther ; d r ight : 1 and 2 have no determinant , revealed by the ant iserum AS, in common. I n the present s tudy only pat terns b and e were observed. e: The absorpt ion control is first used to ensure the specificity of the cross-reaction (first two

Last Common Ancestor of Man and Chimpanzee 257
Ma te r i a l and Methods
Plasma Samples. Human blood samples were collected from apparently healthy blood donors of a local transfusion centre. The chimpanzee (Pan troglodytes) plasma investigated was obtained from Dr. Hill 's department at the Regional Primate Research Center, Covington, La., U.S.A., as were most of our previous specimens. Data from other species were taken from our earlier publication (Bauer, 1970d, 1973). The samples were air-mailed to our laboratory and analyzed as fast as possible to avoid any possible deterioration. In some cases, especially for control purposes, the samples were frozen and kept at - - 2 0 ° until needed.
Antisera. With two exceptions only rabbit antisera were employed (Behringwerke, Mar- burg, Germany). For the batch numbers see Table 1. The reason for our mentioning and stressing these numbers is our observation in rare cases of differences in the determinant patterns an antiserum batch reacts with. Differences of this kind are probably caused by differences in the relative amount of determinant-specific antibodies in various antiserum batches (Bauer, in preparation).
Technique. We followed the methods established and described previously in detail (Bauer, 1970a, c, 1973). The investigation scheme included the following steps:
1. Determination of titre values in a standardized Ouchterlony test arrangement (Bauer, 1970a). This test - - which is in close analogy with I~uttall's type of experimentation - - was used as a screening procedure for the occurrence and strength of cross-reactions. The optimal concentrations for subsequent analyses were also estimated, especially with antisera showing prozone phenomena.
2. Comparative Ouchterlony tests were run next with all samples reacting positively. The test arrangement employed here was made more sensitive by using extra large wells. The samples to be compared were filled into neighbouring reservoirs, punched in the form of a rosette pattern into 1% agarose gels, in the centre of which the antiserum was placed. The test arrangement employed throughout is shown in Fig. lb . After incubation at 37 ° for appropriate periods of time the tests were read without previous staining. For each pair of samples - - besides control experiments these pairs consisted of plasma from man and chim- panzee in each ease - - one of the following types of reaetions was recorded: eoaleseenee, in- dicating identity of determinant patterns (Fig. I b) ; spurring, indicating that the species that "spurs" over its partner carries at least one more determinant (Fig. i c); and combinations of these (Fig. I d). Reaetion patterns of the latter type were not observed and are, moreover, not to be expected in the pair man/chimpanzee, since the antisera were specific to human deter- minants and no chimpanzee determinants not present on the human homologue as well should be expected. Therefore the only constellations observed in this series were human molecule ----- chimpanzee molecule and human sample spurs over chimpanzee sample.
3. Absorption tests, which fulfil a double function in comparative determinant analysis; they were first run as a eontrol experiment for the comparative Ouchterlony test (Fig. 1 e); secondly an antiserum to a human protein absorbed exhaustively with human plasma should no longer cross-react with any animal homologue (a positive reaetion in this ease indicating non-specific antibodies present in the antiserum). All reactions recorded in this study were specific and met this criterion.
Immunoelectrophoretic controls previously recommended from our side to add a physieo- chemical parameter (Bauer, 1969) were not made in this series because of the limited amount of plasma available.
For the use in our time scale also data obtained previously with other subhuman primates were employed. They were examined in the same way as described here. The use of these earlier data was necessary to estimate the determinant numbers on the proteins of man and Pan, which cannot be calculated from the investigation of this single pair of species.
lines); subsequently the relationship between the human and the chimpanzee homologues was examined as shown. The first alternative corresponds to Fig. I c, the second to lb . The last line indicates the test run with other species. The technique described is our standard
method for comparative phylogenetic work (Bauer, 1970d)

258 K. Bauer: Last Common Ancestor of iKan and Chimpanzee
The Principle of Comparative Determinant Analysis (CDA)
The techniques described here make it possible to state for any one pair of homologous proteins, whether the determinant number detected by the antiserum employed is equal or whether one of the homologues carries one determinant (at least) more than the other. For the current project of reconstructing the evolution of primates, especially the one of the chimpanzee, from molecular data on a quan- t i tat ive basis, we have employed the following approach:
I f an antiserum to a human protein reacts with less determinants on a homo- logue from a subhuman primate than the human molecule carries, then a mutat ion must have become accepted during the development from the LCA of man and this species to the present time. This can be assumed because the failure of the ant ibody to recognize a certain determinant must be due to a change in at least one amino acid position, which in turn is caused by a change in a t least one base in the corresponding structural gene, i.e. what is defined as a single point mutation. Evidently not all point mutat ions detectable at the amino acid level are observed in the determinant pattern, since determinants comprise a number of amino acids and since not all parts of a protein are antigenic. This is a similar phenomenon as observed between polypeptide and DIqA sequences: not all changes occurring at the nucleic acid level become apparent in the polypeptide. Therefore the number of detectable mutat ions decreases from the base level to the one of amiao acids, and from the latter to the one of determinants. On the other hand, any change in the determinant pa t te rn must be caused by one at the base level. This makes the comparison of the determinant pat terns into one of the structural genes, though the sensitivity to observe a difference is lower than in the case of direct D~TA or polypeptide sequencing.
I t is evident tha t application of the CDA principle in a single pair of species, such as man and chimpanzee, is not sufficient to estimate their position in the framework of evolution. In our s tudy we have made a comparison with data on several more subhuman primates and pr imate groups obtained earlier in this laboratory.
This way it becomes possible to calculate average acceptance rates for im- munologically detectable mutat ions for each species and to use them for further estimations, e.g. of the times at which various lines of descent diverged from each other. We have used the age of the eutherian LCA as a reference point and cal. culated the divergence times for the various primates beginning with it, as de- scribed below.
Results
The results of the determinant analysis - - in absolute numbers - - are given in Table 1. Only determinants restricted to primate homologues are taken into account .
When investigated in our standardized test system a number of human plasma proteins cross-react with non-primate molecules as well. A system of immunological evolution groups (IEG) defined after this reaction pat tern (Table 2) describes the var iety of cross-reactions of various proteins (Bauer, 1970a). I E G I I I and I E G IV proteins share determinants with one non-primate mammal at least. The values
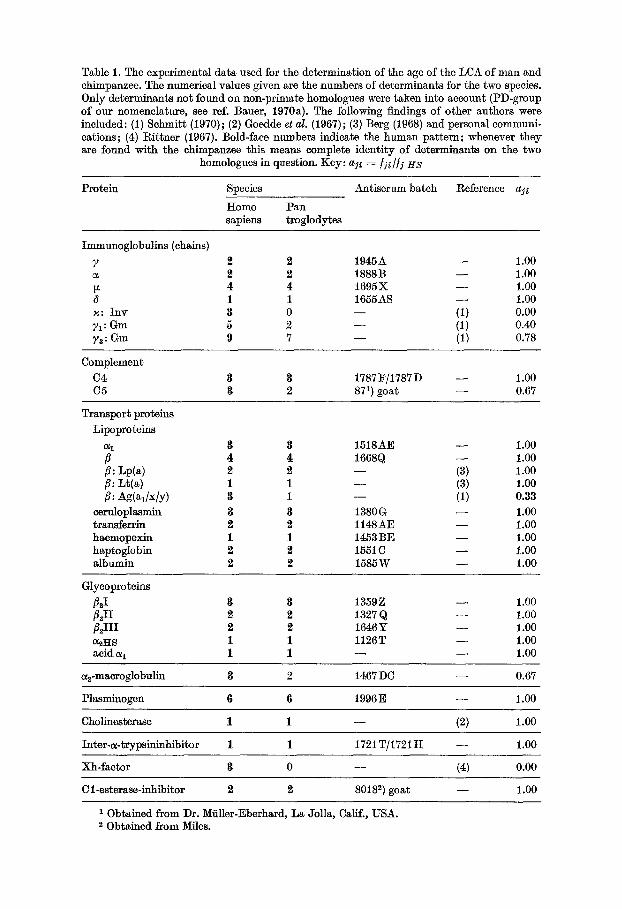
Table 1. The experimental data used for the determination of the age of the LCA of man and chimpanzee. The numerical values given are the numbers of determinants for the two species. Only determinants not found on non-primate homologues were taken into account (PD-group of our nomenclature, see ref. Bauer, 19703). The following findings of other authors were included: (1) Sehmitt (1970); (2) Goedde et al. (1967); (3) Berg (1968) and personal communi- cations; (4) Rit tner (1967). Bold-face numbers indicate the human pat tern; whenever they are found with the chimpanzee this means complete identity of determinants on the two
homologues in question. Key: aj~ = / J d / 1 HS
Protein Species Antiserum batch Reference 33. ~
Homo Pan sapiens troglodytes
Immunoglobulins (chains)
7 2 2 1945A - - 1.00 a 2 2 1888B - - 1.00 tx 4 4 1695X - - 1.00 (~ 1 1 1655AS - - 1.00 ~: Inv 3 0 - - (1) 0.00 71: Gm 5 2 - - (1) 0.40 ~a: Gm 9 7 - - (1) 0.78
Complement
C4 3 3 1787~/1787D - - 1.00 C5 8 2 871) goat - - 0.67
Transport proteins
Lipoproteins
a 1 3 8 1518AE - - 1.00 fl 4 4 1668Q - - 1.00 8: up(a) 2 2 - - (3) 1.o0 8: Lt(a) 1 1 - - (3) 1.00 8: Ag(al/x/Y) 8 1 - - (1) 0.33
eeruloplasmin 3 3 1380 G - - 1.00 transferrin 2 2 1148 AE - - 1.00 haemopexin 1 1 1453 BE - - 1.00 haptoglobin 2 2 1551C - - 1.00 albumin 2 2 1585W - - 1.00
Glycoproteins
fl2I 3 3 1359Z - - 1.00 flzII 2 2 1327 Q - - 1.00 fl2III 2 2 1646¥ - - 1.00 a~vrs 1 1 1126T - - 1.00 acid 31 1 1 - - - - 1.00
a~-macroglobulin 3 2 1467 DC - - 0.67
Plasminogen 6 6 1996 E - - 1.00
Cholinesterase 1 1 - - (2) 1.00
Inter-a-trypsininhibitor 1 1 1721T/1721 H - - 1.00
Xh-faetor 8 0 - - (4) 0.00
C 1-esterase-inhibiter 2 2 80182) goat - - 1.00
1 Obtained from Dr. Mfiller-Eberhard, La Jolla, Calif., USA. 2 Obtained from Miles.

260 K. Bauer:
Table 2. The system of immunological evolution groups (IEG) of human plasma proteins. A stepwise increase in the range of cross-reactions observed is typical of this system. After
(Bauer 1971 a)
Group of species cross-reacting with human plasma protein homologue
Pongids Catarrhinians Simians Primates Eutheria ]~Iammalia
lEG
I I I a I I b I I c I I I IV
+ + ÷ + ÷ + - - + + -[- ÷ +
_ _ + ÷ + +
_ _ _ ÷ ÷ ÷
. . . . ÷ ÷
in Table 1 comprise a total of 80 determinants (on the human homologues), belonging to 30 different systems, in fulfillment of Buettner-Janusch's condition mentioned above.
A first comparison of the differences in determinant pat terns between human and chimpanzee proteins recorded in the present s tudy with our previous findings in other primates shows tha t they are in general smaller than for any other species, including the two other big apes, gorilla and orang utan.
The age of the LCA for man and chimpanzee was calculated by the method described earlier (Bauer, 1970d). For this purpose it is assumed tha t the number of accepted mutat ions - - which is expressed in the differences of determinant pat terns - - is a linear function of time. Zuckerkandl and Pauling (1965) proposed an e-function of the form
p_~-- e-at
for the probabil i ty tha t a site remains unchanged. For the relatively short period of t ime our t ime scale covers in the framework of the whole phylogencsis, we think, however, a linear relationship to be a sufficient approximation. We have therefore proposed the following formula (Bauer, 1970d) :
n l iHs
where T~ is the t ime interval in relative t ime units (RTU) from the eutherian LCA to the one of man and species i, n is the number of proteins, h~ and ]i//s the number of human determinants on the homologous proteins ] of species i and of man.
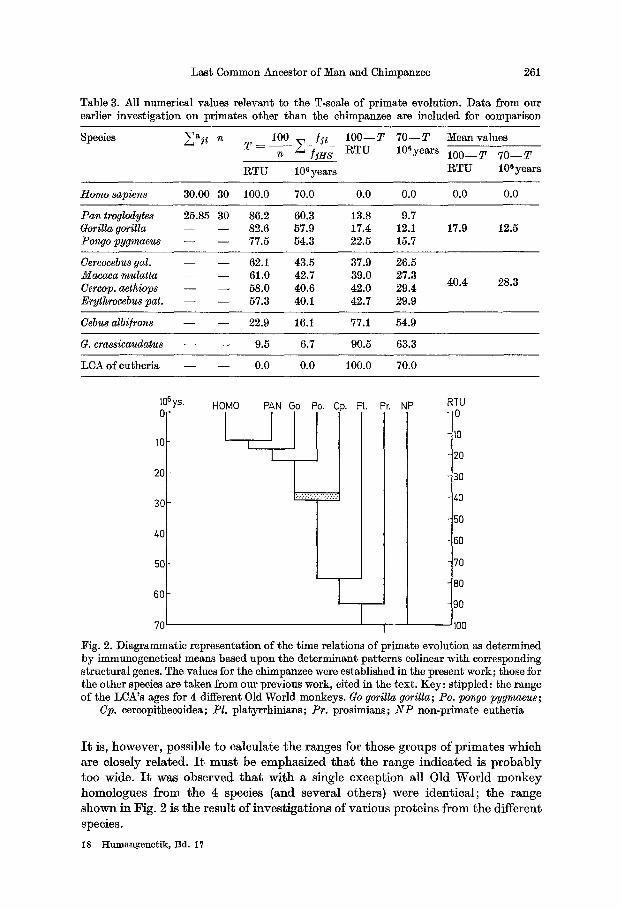
Table 3 shows the values relevant for the t ime scale. Da ta included on species other than the chimpanzee and man are taken from our previous reports (Bauer, 1970d, 1973) for reason of comparison. The two immunological t ime scales, the relative-one (RITS) and the absolute-one (AITS), differ in the units t ime is expres- sed in. In the RITS the t ime interval between the eutherian LCA and today is sub- divided into 100 RTU, for the AITS the same period is taken to be 70 million years.
Fig. 2 gives a graphic representation of the t ime scale as a pedigree. For the LCA of catarrhinians the range of results for the 4 species of Old World monkeys, our calculation is based on, are indicated (stippled). I t is not possible to calculate variance from the algorithm as long as only a single set of data can be employed.
Last Common Ancestor of Man and Chimpanzee 261
Table 3. All numerical values relevant to the T-scale of primate evolution. Data from our earlier investigation on primates other than the chimpanzee are included for comparison
n 1 0 0 _ ]1~ Species ~aj~ T = T ) ' 1 0 0 - - T 7 0 - - T Mean values RTU 106years 1 0 0 - - T 7 0 - - T
RTU 10 ~ years R T U 10 s years
Homo sapiens 30.00 30 100.0 70.0 0.0 0.0 0.0 0.0
P a n troglodytes 25.85 30 86.2 60.3 13.8 9.7 Gorilla gorilla - - - - 82.6 57.9 17.4 12.1 17.9 12.5 Pon9o pygmaeus - - - - 77.5 54.3 22.5 15.7
Cercocebus gal. - - - - 62.1 43.5 37.9 26.5 ~lacaca mulat ta - - - - 61.0 42.7 39.0 27.3
40.4 28.3 Cercop. aethiops - - - - 58.0 40.6 42.0 29.4 Erythrocebus pat. - - - - 57.3 40.1 42.7 29.9
Cebus albi]rons - - - - 22.9 16.1 77.1 54.9
G. erassicaudatus - - - - 9.5 6.7 90.5 63.3
LCA of eutheria - - - - 0.0 0.0 100.0 70.0
10SY s" HOMO PAN Go Po. Cp. PI. Pr, NP ~TU C 0
l0 lC
20
2C 30
3C ~0
5O
4C 50
5C 70
BO 6C
90
7C I 1oo
Fig. 2. Diagrammatic representation of the time relations of primate evolution as determined by immunogenetical means based upon the determinant patterns colinear with corresponding structural genes. The values for the chimpanzee were established in the present work; those for the other species are taken from our previous work, cited in the text. Key: stippled: the range of the LCA's ages for 4 different 01d World monkeys. Go gorilla gorilla; Po. pongo pygmaeus;
Cp. eercopithecoidea; Pl. platyrrhinians; Pr. prosimians; N P non-primate eutheria
I t is, h o w e v e r , poss ib le to ca l cu la t e t h e r anges for t hose g r o u p s o f p r i m a t e s w h i c h a re c lose ly r e l a t ed . I t m u s t be e m p h a s i z e d t h a t t h e r a n g e i n d i c a t e d is p r o b a b l y t o o wide . I t was o b s e r v e d t h a t w i t h a s ingle e x c e p t i o n a l l Old W o r l d m o n k e y h o m o l o g u c s f r o m t h e 4 species (and seve ra l o thers ) were i d e n t i c a l ; t h e r a n g e
s h o w n in F ig . 2 is t h e r e su l t o f i n v e s t i g a t i o n s o f v a r i o u s p r o t e i n s f r o m t h e d i f fe ren t species.
18 Humangenetik, Bd. 17

262 K. Bauer:
Discussion
The rate at which mutations in the determinant pattern detectable by im- munological means occur is assumed here to be a linear function of time elapsed. This condition may not be fulfilled in single proteins. But it should apply when larger numbers of proteins are investigated:
This way both randomness and differences in evolutionary rates for the various proteins should average out (Zuekerkandl and Pauling, 1965). I f a reason- able number of proteins is taken into account the corresponding portion of genetic information can be taken as a sufficiently large statistical sample to draw con- clusions on the whole genomes and thus to compare the whole genomes in question and with them the organisms they code.
Even if the sample is large enough it may still not fulfill the conditions imposed by a statistician, because proteins included in the investigation are not selected strictly at random, as they should be; but with the pragmatic touch of examining those proteins which are easily available, such as plasma proteins, and for which antisera can be obtained. At the present time we have no other data at our dis- posal but those compiled in Table 1. We are therefore not able to select a sample but from them, a procedure necessarily making it rather small. However, we think this fact not to be too restrictory, since plasma proteins include such a wide variety of molecular types, differing in physicochemieal, biochemical, functional and many other aspects, tha t the deviations from randomness are certainly not as grave as appears at first sight.
The method used by us (CDA), based upon the comparison of determinant patterns colinear with the corresponding structural genes, is certainly superior to techniques which measure the amount of precipitate formed or the complement fixation resulting. Not only are the data of Wilson and his group (1964), mentioned earlier in this communication, a clear and convincing piece of evidence for the total dependence of results with this group of methods on the exact experimental conditions used. Moreover, these other methods fail to detect differences in anti- genie structures in a numerical way which can be used to calculate and derive numerical parameters. Also a theoretical understanding of the interaction between single determinants and antibodies bound to them and of the components of the complement system is still far from being complete. Complement fixation tests are quite an indirect method of measuring antigen-antibody binding, making the detection system more complex without apparent advantage over precipitation measurements. The latter, however, is seemingly dependent on the levels of the various determinant-specific antibodies present in the antiserum which take part in the formation of the reaction product measured. Determinants carried by a protein are to a certain degree exchangeable without any effect on the precipitation reaction; since in principle already two determinants per molecule are sufficient to give maximum precipitation, provided that optimal concentrations of the corresponding antibodies are present in the reaction medium.
There is apparently no connexion between the quanti ty of precipitate or the strength of complement fixation and the determinant patterns which are eolinear with the strnetural genes that can be used for a quantitative approach. We do not see any other way of connecting antigen-antibody reactions with the genome but

Last Common Ancestor of Man and Chimpanzee 263
via the determinant pattern; while we do not exclude other technical methods, the two older techniques are certainly unsnitable for this purpose for the reasons described. Such a connexion must in our opinion be considered a prerequisite of any investigation of evolutionary events based on immunological or other data. For these reasons we feel unable to accept the theoretical concept of the immuno- logically determined time values for the LCAs of man and several other primates as employed by Sarich and Wilson (1967). Their results have been criticized by several authors for various reasons, as e.g. by Simons (1969), because they failed to fit into the pat tern of estabhshcd palaeontological evidence and by Read and Lestrfl (1970) for mathematical reasons. One of the main criticisms they met was that their values for the ages of LCAs of man and various apes and monkeys differed widely from those obtained by fossil evidence.
Our own published data, calculated as described from the investigation of the determinant patterns of at least 15 different plasma proteins per species were largely in accordance with classical values; this is true for all primate groups examined (Bauer, 1970d, 1973), and especially in the case of the two more dis- puted LCAs, those of man and gorilla and man and orang utan. Our present result for the chimpanzee fits into this same pattern. I t is actually at the lower limit set by authors such as Pilbeam (personal communication), who put the LCA for man and pongids in the region of between 10 and 20 million years of age.
The method we have employed also has several limitations; the first is the fact that only human determinants, i.e. determinants found on human molecules, are included in the estimation of ages of LCAs. This is due to the fact that antisera to human plasma proteins were used and arc actually the only antisera of this type available for such a variety of different proteins. In principle it is possible to use any other of the primate species as the immunogen donor; however, this would involve the isolation of all the proteins to be examined in these species, the pro- duction of antisera and then the same procedure as described here for antisera to human proteins. We doubt whether the results would be very different ff experi- mentation were performed in tMs way, since the overall distribution of mutations detected tMs way should be independent of the species from which the immunogen is derived. We feel, however, that investigations of this type might provide a possibility for control, especially as the present time-scale values are calculated in a way not allowing estimation of standard deviations. The preparatory work necessary for such a series of parallel analyses is, however, rather complicated and the major advantage of immunological methods in evolutionary research, i.e. the availability of a large body of data in a relatively short period of time, is partly lost.
As to the absolute numerical values given in the /kITS of primate evolution, we have previously stressed that its accuracy is in principle inferior to that of the RITS, because geological estimations are combined with data obtained by in- vestigations of molecular structures. Thus uncertainties not inhaerent in the im- munogenetie method used may enter and influence the AITS. We think relative time scales used in our own and in other studies of this kind, including biochemical ones, to be better estimations of the real time relationships.
In the present case we have arbitrarily set the age of the eutherian LCA as being 70 million years. I f the actual age is, say, 75 million years the RITS remains
18"

264 K. Bauer:
unchanged, while the AITS values mus t be recalculated for every single species included. This means t h a t the R I T S reflects the results of a comparat ive investi- gat ion a t the molecular level in a more accurate and direct way, while AITS, measuring the LCA ages in absolute units, incorporate uncertainties beyond the control of either the immunogenet icis t or the biochemist. However, as soon as a reference point is chosen , /k ITS provide a direct comparison between morpho- logical and immunogenet ic results.
I f the value of 9.7 million years for the age of the LCA of m a n and chimpanzee is accepted as a reasonable approximat ion this result a t the same t ime once more stresses the special position of this species in relation to man among the great apes. A number of other features of the chimpanzee have already pu t i t into the closest neighbourhood to m a n among all animals. These properties include anatomical , physiological and psychological factors (Gallup, 1969) as well as others, such as the susceptibility to poliomyelitis which is shared by man and Pan. I t therefore appears only reasonable t h a t the age of the LCA of the two species in this investi- gat ion turns out to be lower than those calculated for man and gorilla or man and orang u t a h : the m a n y properties common to man and chimpanzee find their ex- planat ion in the sharing of a larger pa r t of the genome than for any other species belonging to pongids and hominids.
Moreover we th ink it most remarkable t h a t two approaches as different as the morphological one and one based upon genetic informat ion in all cases lead to very similar results as to the relationships between the various primates. This fact makes CDA a very interesting tool for evolut ionary research.
Acknowledgements. The author gratefully acknowledges the generous supply of chimpanzee plasma as well as of samples of other primate plasmas provided for our previous studies by Dr. Hill from the Delta Regional Primate Research Center, Covington, La., U.S.A. This con- tribution is part of a project sponsored by a grant (Ba 356/3) from Deutsche Forschungs- gemeinschaft, Bonn-Bad Godesberg, Germany. The author thanks Miss U. Reinle and Mrs. R. Rauschnabel for excellent technical assistance. During part of the experimental series this paper is based upon the author made use of laboratory facilities at ~he Heidelberg University Serological Institute.
References
Bauer, K. : Heterologe Reaktionen yon S~ugetierseren mit monospezifischen Antihumanseren. Ein Beitrag zur Evolution der Serumprofeine. Humangenetik 7, 76--90 (1969).
Bauer, K. : Cross-reactions between human and animal plasma proteins. I. The immunological evolution groups (IEG) I and II. Humangenetik 8, 325---329 (1970a).
Bauer, K. : Cross-reactions between human and animal plasma proteins. II. The influence of structural and functional factors. Humangenetik 10, 1--7 (1970b).
Bauer, K. : The antigenic determinants of several human plasma proteins. Their determinations from the investigation of the cross-reactions between human and other mammalian plasmas. Int. J. Prof. Res. 2, 137--145 (1970c).
Bauer, K.: An immunological time scale for primate evolution consisfent with fossil evidence. Humangenetik 10, 344---350 (1970d).
Bauer, K.: Cross-reactions between human and animal plasma proteins. IV. l~on-eutherian mammals. Humangenetik 13, 49--58 (1971a).
Bauer, K. : Cross-reactions of four kangaroo plasma proteins with their human homologues. Int. J. Prof. Res. 8, 233--235 (1971b).

Last Common Ancestor of Man and Chimpanzee 265
Bauer, K.: Immunologische Untersuchungcn fiber die molekulare Evolution menschlicher Plasmaproteine in ihrer Beziehnng zur Stammesgeschichte der Primaten. Fortschritte der allgemeinen und klinischen Humangenetik. Stuttgart: Thieme (Accepted for publication, 1973).
Berg, K.: The Lp system. Series haematol. 1, 111--136 (1968). Buettner-Janusch, H. : Comment in the discussion of Part I I I of (V. Bryson and H. J. Vogel,
eds.) Evolving genes and proteins. New York: Academic Press 1965. Crumpton, M. J. : The molecular basis of the serological specifity of proteins, with particular
reference to sperm-wtmle myoglobin. Prec. 2nd FEBS Meeting 1, 61--80 (1966). Dayhoff, M. O. : Atlas of protein sequence and structure, Vol. 4. Silver Spring (Md.) : National
Biomedical Research Foundation 1969. Gallup, G. G. : Chimpanzees: Self-recognition. Science 167, 86 (1970). Goedde, H.W. , Doenicke, A., Altland, K.: Pseudocholinesterasen. Berlin-Heidelberg-New
York: Springer 1967. Goodman, M.: The specificity of proteins and the process of primate evolution. Protides of
the biological fluids, p. 70. Amsterdam: Elsevier 1964. Goodman, M., Moore, G. W., Farris, W., Poulik, E. : In: The chimpanzee, Vo]. 2, p. 318. Basel-
New York: Karger 1970. Mollison, T.: In: Abderhaldens Handbuch der biologischen Arbeitsmethoden, Abt. 9, p. 553.
Berlin-Wien: Urban und Schwarzenberg 1923. Nolan, C., Margoliash, E. : Comparative aspects of primary structures of proteins. Ann. Rev.
Biochem. 87, 727--790 (1968). Nuttall, G. H. F. : Blood immunity and blood relationship. Cambridge: Cambridge University
Press 1904. Pilbeam, D. : Personal communication 1970. Read, D. W., Lestril, P. E. : Hominid phylogeny and immunology: a critical appraisal. Science
168, 578--580 (1970). Rittner, C. : Inter- and intra-species variations withiu the Xh antigen. Vex Sang. (Basel) 18,
29--31 (1967). Sarich, V. M., Wilson, A.C.: Immunological time scale of hominid evolution. Science 158,
1200--1202 (1967). Schmitt, J. : Blur- und Serumgruppen bei Affen. Humangenetik 8, 261--280 (1970). Simons, E. L. : The origins and radiation of the primates. Ann. N.Y. Acad. Sci. 167, 319--331
(1969). S6derquist, T., Blomb~ck, B. : Fibrinogen structure and evolution. Naturwissenschaften 58,
16--23 (1971). Wilson, A. C., Kaplan, N. O., Levine, L., Pesce, A., Reichlin, M., Allison, W. S.: Evolution
of lactic dehydrogenases. Fed. Prec. 28, 1258--1264 (1964). Zuckerkandl, E., Pauling, L.: Evolutionary divergence and convergence in proteins. In
(V. Bryson and H. J. Vogel, eds.) : Evolving genes and proteins, p. 97. hTew York: Academic Press 1965.
Priv.-Doz. Dr. K. Bauer D-6900 Heidelberg 1 Rohrbacher Stral]e 8/III/3 Federal Republic of Germany