Embed Size (px)
Citation preview

S
A
Ma
b
c
h
�����
a
ARR1AA
KAFFf
taamttbpt
rshF
0h
Behavioural Brain Research 237 (2013) 172– 175
Contents lists available at SciVerse ScienceDirect
Behavioural Brain Research
j ourna l ho me pa ge: www.elsev ier .com/ locate /bbr
hort communication
mygdala activation to fearful faces under attentional load
artin Mothes-Lascha,c,∗, Hans-Joachim Mentzelb, Wolfgang H.R. Miltnera, Thomas Straubec
Department of Biological and Clinical Psychology, Friedrich-Schiller-University Jena, Am Steiger 3//1, D-07743 Jena, GermanyInstitute of Diagnostic and Interventional Radiology, Friedrich-Schiller-University Jena, Bachstr. 18, D-07740 Jena, GermanyInstitute of Medical Psychology and Systems Neuroscience, University Hospital Muenster, Von-Esmarch-Str. 52, D-48149 Münster, Germany
i g h l i g h t s
We investigated the activation of the amygdala to fearful faces under high load.There was no differential amygdala activation under standard conditions (E1).We erased the eyes in half of the faces to increase the normal face saliency (E2).There was higher amygdala activation to normal but not to manipulated fearful faces.Amygdala activation depends on cognitive resources and saliency of fearful faces.
r t i c l e i n f o
rticle history:eceived 13 February 2012eceived in revised form4 September 2012ccepted 17 September 2012vailable online 24 September 2012
a b s t r a c t
While some functional imaging studies suggested an automatic activation of the amygdala to fearfulvs. neutral faces, recent studies showed the absence of amygdala activation to fearful faces under highattentional load induced by distracting tasks. The present fMRI study investigated whether this outcomecan be modulated by changing the saliency of the eyes of fearful faces. Subjects had to solve a highperceptual load task while they were presented either with normal faces (Exp. 1) or with normal faces
eywords:ttentionaceearMRI
intermixed with faces, in which pupil and iris of eyes were erased to increase the saliency of the normaleyes (Exp. 2). There was no differential amygdala activation to fearful versus neutral faces under standardconditions without any manipulation of the faces (Exp. 1). In contrast to this outcome, Experiment 2 led todifferential amygdala activation to the normal but not to the manipulated fearful vs. neutral faces. Thesefindings propose a concept of relative automaticity of the activation of the amygdala. The activationdepends on available cognitive resources and on the saliency of specific parts of fearful faces.
Several studies showed amygdala activation to fearful vs. neu-ral facial expressions under attentional load, when subject’sttention is focused on face-unrelated stimuli [1,2], suggesting anutomatic response of the amygdala to threat signals. However,ore recent studies revealed that more difficult attentional dis-
raction tasks lead to complete inhibition of differential activationo fearful vs. neutral faces in the amygdala [3–5, but see modulationy anxiety 6]. These findings suggest that fearful faces are notrocessed by the amygdala when very demanding tasks exhausthe available processing capacity.
However, it remains unclear whether this load effect is observedegardless of the saliency of faces or specific parts of fearful faces
uch as the eyes. In particular the eye region of fearful faces seems toave an important impact on the activation of the amygdala [7–10].urthermore, the amygdala is strongly involved in responses to∗ Corresponding author. Tel.: +49 3641945145; fax: +49 3641945142.E-mail address: [email protected] (M. Mothes-Lasch).
166-4328/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.bbr.2012.09.033
© 2012 Elsevier B.V. All rights reserved.
human eyes per se [11,12] and specific eye information such assize of eyes [13] or pupils [7] and gaze [14,15]. Furthermore, amyg-dala activation predicts gaze shifts to eyes of fearful faces in healthysubjects [16]. These findings suggest that the amygdala, besides itsrole in emotion, is important for attending to behaviourally relevantinformation provided by human eyes.
The question arises whether the lack of the differential acti-vation of the amygdala to fearful vs. neutral faces under highperceptual load can be modulated by the saliency of specific facialfeatures such as fearful eyes. If high perceptual load per se pre-vents differential activation of the amygdala to fearful vs. neutralfaces, this should be evident also under conditions that enhance therelevance of the eyes. However, when the activation of the amyg-dala is modulated by such manipulations, the amygdala activationto fearful faces under perceptual load would be a function of both
cognitive resources and the saliency of facial features.The present fMRI study investigated the effect of changing thesaliency of fearful eyes on amygdala activation to fearful vs. neutralfaces under high perceptual load. Specifically, in half of the trials

al Bra
wptwap
ctaymce
fTg8gareoho
p23saot(divewwafatatrwNdvCtd
nGrwvcrbip
M. Mothes-Lasch et al. / Behaviour
e manipulated the eyes of the faces by erasing the iris and theupil and thus completely whitening the eyes to increase the rela-ive saliency of the non-manipulated eyes. This allowed us to testhether the contextual relevance of the eyes has an effect on the
mygdala activation to fearful vs. neutral faces even under higherceptual load.
Twenty-four right-handed healthy subjects with normal ororrected-to-normal vision participated in the experiments withwelve subjects in each experiment (Exp. 1: 10 females, meange = 22.18 ± 2.36 years; Exp. 2: 7 females, mean age = 21.9 ± 2.93ears). One participant of Exp. 2 had to be excluded due to abnor-alities in brain activation. Participants provided written informed
onsent for participation in the study which was approved by thethics committee of the University of Jena.
In both experiments, a set of pictures of fearful and neutralacial expressions from eight different individuals was used [17].he pictures were digitally cropped to remove extraneous back-round information by using Adobe Photoshop software (Version.0.1, Adobe Systems Incorporated, San Jose, California). The back-round of the pictures was black. In Experiment 2, the same facess in Experiment 1 were used; however, the faces were modified byeplacing the iris and the pupil with the colour of the sclera. In bothxperiments a string of six letters (see below) was superimposedn each face using Presentation® software (Version 9.13, Neurobe-avioural Systems, Inc., Albany, California). Fig. 1 shows examplesf the stimuli used in both experiments.
Each experiment consisted of a run of 128 facial stimuliresented in randomized order. Each picture was displayed for00 ms with an inter-stimulus interval of 4300 ms. Additionally,2 null events (fixation cross) were randomly intermixed into theequence of stimuli. Regardless of picture, subjects had only tottend to the letter string that was superimposed on the centerf each face. The task was to decide whether the letter string con-ained an “X” or an “N”. Every string comprised a single target letterN or X) and five non-target letters (H, K, M, W, Z) arranged in ran-om sequence [18]. By means of this high perceptual load task,
t has been shown previously that the differential amygdala acti-ation to fearful vs. neutral faces becomes completely inhibitedven in highly anxious healthy subjects [3]. In Experiment 1, thereere two conditions of the factor emotion (fearful vs neutral faces)ith 64 trials per condition. In Experiment 2, in half of the tri-
ls the modified faces replaced the intact faces. Thus, there wereour conditions according to the factors emotion (fear vs neutral)nd modification (original vs modified) with 32 trials per condi-ion. Responses were obtained by pressing one of two buttons ofn optic fiber response box (LUMItouch, Photon Control Incorpora-ion, Burnbay, BC, Canada) with the index or the middle finger of theight hand. Presentation of stimuli as well as recording of responsesas accomplished with Presentation® software (Version 9.13,eurobehavioural Systems, Inc., Albany, California). Behaviouralata were analysed by means of repeated measures analysis ofariance (ANOVA) using SPSS software (Version 13; SPSS, Inc.,hicago, Illinois). A probability level of p < .05 was considered sta-istically significant. All data are expressed by mean ± standardeviation.
Scanning was performed by a 1.5-T magnetic resonance scan-er (“Magnetom Vision Plus”; Siemens Medical Systems, Erlangen,ermany). After acquisition of a T1-weighted anatomical scan, one
un of T2*-weighted echo-planar images (EPIs) with 248 volumesas measured (TE = 50 ms, flip angle = 90◦, matrix = 64 × 64, field of
iew = 192 mm, TR = 2980 ms) in both experiments. Each volumeomprised 30 axial slices (thickness = 3 mm, gap = 1 mm, in-plane
esolution = 3 × 3 mm). The slices were acquired parallel to the lineetween anterior and posterior commissure. Furthermore, beforemaging, a shimming procedure to improve field homogeneity waserformed. The first four volumes of each run were discarded
in Research 237 (2013) 172– 175 173
from analysis to ensure that steady-state tissue magnetization wasreached.
Pre-processing and analyses of the functional data were per-formed with Brain Voyager QX software (Version 1.9; BrainInnovation, Maastricht, The Netherlands). First, all volumes wererealigned to the first volume in order to minimize artifacts becauseof head movements and a slice time correction was conducted.Further data preprocessing comprised spatial (8 mm full-widthhalf-maximum isotropic Gaussian kernel) as well as temporalsmoothing (high pass filter: 3 cycles per run; low pass filter:2800 ms; linear trend removal). The anatomical and functionalimages were coregistered and normalized to the Talairach space[19].
Statistical analyses were performed by multiple linear regres-sion of the signal time course at each voxel. The expected bloodoxygen level-dependent (BOLD) signal change for each event type(=predictor) was modelled by a hemodynamic response func-tion. Within-group statistical comparisons were conducted usinga mixed effect analysis, which considers inter-subject varianceand permits population-level inferences. In the first step, predictorestimates were generated for each individual. In the second step,predictor estimates were analyzed across subjects with repeatedmeasures ANOVA followed by planned t-tests. Analysis was con-ducted for the amygdala as region of interest (ROI), which wasdefined with help of Talairach daemon software [20] and accordingto our previous studies [10,21,22].
Statistical parametric maps resulting from the voxelwise anal-ysis were considered significant for values that survived acluster-based correction for multiple comparisons as implementedin Brain Voyager [23], which is based on a 3D extension of therandomization procedure described by Forman et al. [24]. First,voxel-level threshold was set at p < .005 (uncorrected). Thresholdedmaps were then submitted to a region of interest (ROI) – or wholebrain correction criterion that was based on the estimate of themap’s spatial smoothness and on an iterative procedure (MonteCarlo simulation) for estimating cluster-level false-positive rates.After 1000 iterations, the minimum cluster size threshold thatyielded a cluster-level false-positive rate of 5% was applied to thestatistical maps.
Analyzing differences in error rates revealed no significant effectof emotion (t = 2.19, p > .05) in Experiment 1, and no significanteffect of emotion (F = 1.22, p > .05) or modification (F(1,10) = 0.004,p > .05) as well as no significant interaction of emotion by modifi-cation (F(1,10) = 0.02, p > .05) in Experiment 2.
Analyzing differences in reaction times revealed no signif-icant effect of emotion (t = 0.76 (11), p > .05) in Experiment1. In Experiment 2, there were also no significant effects ofemotion (F(1,10) = 0.008, p > .05) or modification (F(1,10) = 1.73,p > .05) and no significant interaction of emotion by modification(F(1,10) = 0.24, p > .05).
Analysis of fMRI data revealed, in accordance with Bishop et al.[3], that there was no significant effect of emotion in the amygdalain Experiment 1 (see Fig. 2b), which supports the hypothesis thathigh attentional load inhibits the differential processing of fearfulfaces.
In Experiment 2, there was no significant effect of emotion ormodification in the amygdala. However, we found a significantinteraction of emotion by modification (coordinates of peak voxel[x, y, z] = −27, 1, −20; size = 243 mm3; F(1,10) = 28.13, p < .05 [cor-rected], see Fig. 2). This effect was based on increased activationto original fearful vs. neutral faces (t (10) = 5.4, p < .0005), whilethere was no significant effect for the manipulated fearful vs. neu-
tral faces (Fig. 2). Thus, the manipulation of the eye region in halfof the trials led to activation to fearful versus neutral faces in theother trials even under high perceptual load. Additionally, we con-ducted an explorative analysis of the time series of the difference of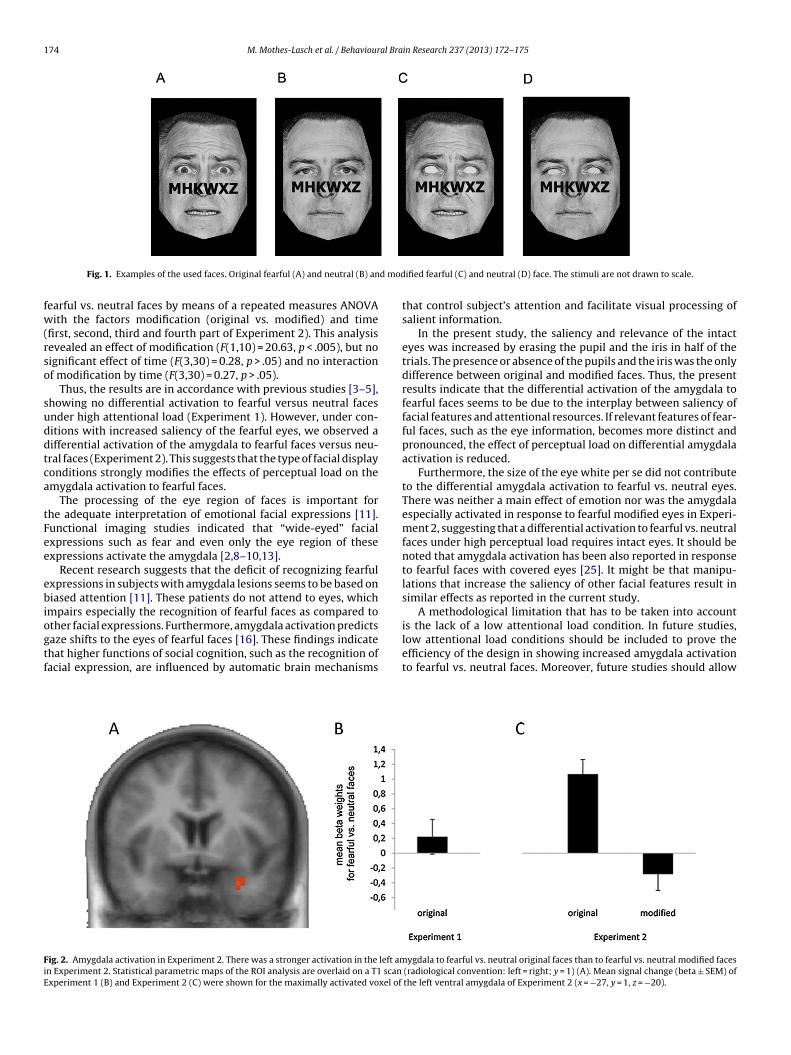
174 M. Mothes-Lasch et al. / Behavioural Brain Research 237 (2013) 172– 175
d mod
fw(rso
suddtca
tFee
ebiogtf
FiE
Fig. 1. Examples of the used faces. Original fearful (A) and neutral (B) an
earful vs. neutral faces by means of a repeated measures ANOVAith the factors modification (original vs. modified) and time
first, second, third and fourth part of Experiment 2). This analysisevealed an effect of modification (F(1,10) = 20.63, p < .005), but noignificant effect of time (F(3,30) = 0.28, p > .05) and no interactionf modification by time (F(3,30) = 0.27, p > .05).
Thus, the results are in accordance with previous studies [3–5],howing no differential activation to fearful versus neutral facesnder high attentional load (Experiment 1). However, under con-itions with increased saliency of the fearful eyes, we observed aifferential activation of the amygdala to fearful faces versus neu-ral faces (Experiment 2). This suggests that the type of facial displayonditions strongly modifies the effects of perceptual load on themygdala activation to fearful faces.
The processing of the eye region of faces is important forhe adequate interpretation of emotional facial expressions [11].unctional imaging studies indicated that “wide-eyed” facialxpressions such as fear and even only the eye region of thesexpressions activate the amygdala [2,8–10,13].
Recent research suggests that the deficit of recognizing fearfulxpressions in subjects with amygdala lesions seems to be based oniased attention [11]. These patients do not attend to eyes, which
mpairs especially the recognition of fearful faces as compared to
ther facial expressions. Furthermore, amygdala activation predictsaze shifts to the eyes of fearful faces [16]. These findings indicatehat higher functions of social cognition, such as the recognition ofacial expression, are influenced by automatic brain mechanismsig. 2. Amygdala activation in Experiment 2. There was a stronger activation in the left amn Experiment 2. Statistical parametric maps of the ROI analysis are overlaid on a T1 scanxperiment 1 (B) and Experiment 2 (C) were shown for the maximally activated voxel of
ified fearful (C) and neutral (D) face. The stimuli are not drawn to scale.
that control subject’s attention and facilitate visual processing ofsalient information.
In the present study, the saliency and relevance of the intacteyes was increased by erasing the pupil and the iris in half of thetrials. The presence or absence of the pupils and the iris was the onlydifference between original and modified faces. Thus, the presentresults indicate that the differential activation of the amygdala tofearful faces seems to be due to the interplay between saliency offacial features and attentional resources. If relevant features of fear-ful faces, such as the eye information, becomes more distinct andpronounced, the effect of perceptual load on differential amygdalaactivation is reduced.
Furthermore, the size of the eye white per se did not contributeto the differential amygdala activation to fearful vs. neutral eyes.There was neither a main effect of emotion nor was the amygdalaespecially activated in response to fearful modified eyes in Experi-ment 2, suggesting that a differential activation to fearful vs. neutralfaces under high perceptual load requires intact eyes. It should benoted that amygdala activation has been also reported in responseto fearful faces with covered eyes [25]. It might be that manipu-lations that increase the saliency of other facial features result insimilar effects as reported in the current study.
A methodological limitation that has to be taken into account
is the lack of a low attentional load condition. In future studies,low attentional load conditions should be included to prove theefficiency of the design in showing increased amygdala activationto fearful vs. neutral faces. Moreover, future studies should allowygdala to fearful vs. neutral original faces than to fearful vs. neutral modified faces (radiological convention: left = right; y = 1) (A). Mean signal change (beta ± SEM) ofthe left ventral amygdala of Experiment 2 (x = −27, y = 1, z = −20).

al Bra
temrt
shaoo
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
M. Mothes-Lasch et al. / Behaviour
he examination of the time series of the activation as well as theffects of the order of the faces during the very first trials. Further-ore, the collection of clear indicators of saliency like unexpected
ecognition memory tests or as provided by the measurement ofhe EEG could promote the understanding of the recent findings.
Taken together, our findings reveal the importance of thealiency of fearful eyes for the activation of the amygdala underigh attentional load. The findings suggest a concept of relativeutomaticity of amygdala activation to fearful faces. The activationf the amygdala depends on both available cognitive resources andn the saliency of specific parts of fearful faces.
eferences
[1] Anderson AK, Christoff K, Panitz D, De Rosa E, Gabrieli JD. Neural correlatesof the automatic processing of threat facial signals. Journal of Neuroscience2003;23:5627–33.
[2] Vuilleumier P, Armony JL, Driver J, Dolan RJ. Effects of attention and emotionon face processing in the human brain: an event-related fMRI study. Neuron2001;30:829–41.
[3] Bishop SJ, Jenkins R, Lawrence AD. Neural processing of fearful faces: effectsof anxiety are gated by perceptual capacity limitations. Cerebral Cortex2007;17:1595–603.
[4] Pessoa L, McKenna M, Gutierrez E, Ungerleider LG. Neural processing of emo-tional faces requires attention. Proceedings of the National Academy of Sciencesof the United States of America 2002;99:11458–63.
[5] Silvert L, Lepsien J, Fragopanagos N, Goolsby B, Kiss M, Taylor JG, et al. Influenceof attentional demands on the processing of emotional facial expressions in theamygdala. Neuroimage 2007;38:357–66.
[6] Cornwell BR, Alvarez RP, Lissek S, Kaplan R, Ernst M, Grillon C. Anxiety overridesthe blocking effects of high perceptual load on amygdala reactivity to threat-related distractors. Neuropsychologia 2011;49:1363–8.
[7] Demos KE, Kelley WM, Ryan SL, Davis FC, Whalen PJ. Human amygdala sensi-tivity to the pupil size of others. Cerebral Cortex 2008;18(12):2729–34.
[8] Whalen PJ, Kagan J, Cook RG, Davis FC, Kim H, Polis S, et al. Human amygdalaresponsivity to masked fearful eye whites. Science 2004;306:2061.
[9] Morris JS, deBonis M, Dolan RJ. Human amygdala responses to fearful eyes.Neuroimage 2002;17:214–22.
10] Straube T, Dietrich C, Mothes-Lasch M, Mentzel HJ, Miltner WH. The volatil-ity of the amygdala response to masked fearful eyes. Human Brain Mapping2010;31:1601–8.
[
in Research 237 (2013) 172– 175 175
11] Adolphs R, Gosselin F, Buchanan TW, Tranel D, Schyns P, Damasio AR. Amechanism for impaired fear recognition after amygdala damage. Nature2005;433:68–72.
12] Spezio ML, Huang PY, Castelli F, Adolphs R. Amygdala damage impairseye contact during conversations with real people. Journal of Neuroscience2007;27:3994–7.
13] Hardee JE, Thompson JC, Puce A. The left amygdala knows fear: laterality in theamygdala response to fearful eyes. Social Cognitive and Affective Neuroscience2008;3:47–54.
14] Akiyama T, Kato M, Muramatsu T, Umeda S, Saito F, Kashima H. Unilateral amyg-dala lesions hamper attentional orienting triggered by gaze direction. CerebralCortex 2007;17:2593–600.
15] Straube T, Langohr B, Schmidt S, Mentzel HJ, Miltner WH. Increased amygdalaactivation to averted versus direct gaze in humans is independent of valenceof facial expression. Neuroimage 2010;49:2680–6.
16] Gamer M, Büchel C. Amygdala activation predicts gaze toward fearful eyes.Journal of Neuroscience 2009;29:9123–6.
17] Ekman P, Friesen WV. Pictures of facial affect. Palo Alto, CA: Consulting Psy-chologists Press; 1976.
18] Lavie N. Perceptual load as a necessary condition for selective attention.Journal of Experimental Psychology: Human Perception and Performance1995;21:451–68.
19] Talairach J, Tournoux P. Co-planar stereotaxic atlas of the human brain.Stuttgart: Thieme; 1988.
20] Lancaster JL, Woldorff MG, Parsons LM, Liotti M, Freitas ES, Rainey L, et al.Automated Talairach Atlas labels for functional brain mapping. Human BrainMapping 2000;10:120–31.
21] Straube T, Kolassa IT, Glauer M, Mentzel HJ, Miltner WHR. Effect of taskconditions on brain responses to threatening faces in social phobics: an event-related functional magnetic resonance imaging study. Biological Psychiatry2004;56:921–30.
22] Mothes-Lasch M, Mentzel HJ, Miltner WH, Straube T. Visual attention mod-ulates brain activation to angry voices. Journal of Neuroscience 2011;31:9594–8.
23] Goebel R, Esposito F, Formisano E. Analysis of functional image analysis contest(FIAC) data with brainvoyager QX: from single-subject to cortically alignedgroup general linear model analysis and self-organizing group independentcomponent analysis. Human Brain Mapping 2006;27:392–401.
24] Forman SD, Cohen JD, Fitzgerald M, Eddy WF, Mintun MA, Noll DC. Improvedassessment of significant activation in functional magnetic resonance imag-
ing (fMRI): use of a cluster-size threshold. Magnetic Resonance in Medicine1995;33:636–47.25] Asghar AU, Chiu YC, Hallam G, Liu S, Mole H, Wright H, et al. Anamygdala response to fearful faces with covered eyes. Neuropsychologia2008;46:2364–70.