Embed Size (px)
Citation preview

1.
2.
3.
4.
5.
6.
AN INTRODUCTORY STUDY ON THE ECOLOGY OF HYRAX (DENDROHYRAX BRUCEZ AND PROCAVZA JOHNSTONZ)
IN THE SERENGETI NATIONAL PARK
M. I. M. Turner, Serengeti National Park and R. M. Watson, Michael Grzimek Memorial Laboratory, Serengeti Research Project
SUMMARY
Dandroh~yrax W and Procawia johrtstoni populations have been studied in kopjes surrounding the senior author’s house. The two species have been described and a short historical review of the Hyracoidea is given. Some notes are made concerning “Hyraceum”. The kopjes under study are describ- ed, being typical of granite outcrops in the Serengeti region. The adaptations of both species of hyrax for the kopje habitat are des- cribed. Observations are made on the diurnal activity patterns in re- lation to climatic conditions. A list of all plants available to the animals is presented, and those eaten by either species are indicated. Comment is made on the lack of overlap in the diets of the two species, and this is suggested as be- ing partly due to lack of pressure on food resources. A discussion on the possible evolu- tion of the kopje-dwelling habit is held, which concludes that recon- vergence may have taken place as a result of forest giving way to wood- land and plains.
INTRODUCTION
The house of the senior author is sited amongst a group of kopjes or inselbergs at Seronera, which lies on the boundary between AcaciaiCommiphma woodland and open Themeda trbndra Forsk./ P m i s e t u m mezianzlm Leeke grass- lands. The rainfall over the last 12 years has averaged 75 cm, most of which falls between November and May. The dry seasons are characterised by
thunderstorms, but two or three suc- cessive months may occur without rain. Maximum shade temperatures reach just over 33OC, and minimum tempera- tures of 4OC have been recorded.
Since 1961 the senior author has made observations on the populations of tree-hyrax (Dendrohyrax brucei) and rock-hyrax (Pmavia johnstmi) living in the kopjes. In I962 a survey of all food eaten by both species was made. In 1964 work was carried out to enable the kopje, as a habitat, to be more fully described.
The rock hyrax have become excep- tionally tame, and feed on kitchen refuse from the hand. Moreover preda- tion pressure could be reasonably ex- pected to be greater at an undisturbed kopje. The provision of drinking water throughout the year is a final unnatural feature of the kopje under study.
This paper is concerned with some of the adaptations of the two species to kopje life, and makes particular refer- ence to food preferences.
Early observers of hyrax were in some confusion as to their taxonomic affinities. Biblical references to rabbits and conies were almost certainly to rock hyrax (Levit. XI. 5 and Psalm CIII. 18). The word hyrax is itself of Greek origin and means shrew in that language. The first South African settlers called i t Dasje or badger. Liinnaeus (1758) makes no mention of hyrax and one of the earliest attempts t.0 place hyrax in a systematic position was by Storr (Storr, 1780) who named i t Procawiu capensis, related to the guinea pig (Burton, 1961). Later taxonomists have made one order containing two genera for the various hjmax species - the Hyracoidea - and this order has its closest affinities with the Proboscidea.
49

EAST AFRICAN WILDLIFE JOURNAL, VOL. 3, AUGUST 1965
Figure 1 Adult rock hyrax at the edge of the kopje thicket.
The two species concerned have been identified at the British Museum of Natural History as ProcavicF johnstmi matschiei Neumann (rock hyrax) and Dendrohyrax (subgenus Heterohyrax) brucei d k n e r i Matschie (tree hyrax) (Hayman, 1964). Hayman considers Heterohyrux to be a subgenus of Den- drohyrax, since the cranial and dental distinctions between the two groups are so small. The rock hyrax is larger, and generally dark brown in colour above and slightly lighter below, with dark feet. The skull of an adult male has a maximum length of 100 mm, and the upper back molars have a transverse breadth of 8-9 mm. (Figure 1).
The tree hyrax is smaller, grey in colour above, almost white below with a sharp flank line of demarcation, and pale feet. The skull of an adult male has a maximum length of 84 mm, and the upper back molars have a trans- verse breadth of 6 mm. (Figure 2).
In the 18th and 19th centuries there was considerable controversy about the origin and medicinal properties of “Hyraceum” (Anon, 1850 ; Owen, 1932 ; nunberg , 1779). “Hyraceum” was used as a substitute for “Castoreum” in “hysterical complaints”, and was considered to be derived from “men- strual secretions”. The rather shiny deposits of hyraceum found in kopjes in the Serengeti are certainly formed from urine as i t drips slowly down a more or less vertical rock face from a communal urination rock above.
T’HE KOPJE A S A HABITAT
The kopjes of the western Serengeti are derived from granite and granodio- rite intrusives which have been wea- thered by wind and possibly waves into rather smooth-textured boulders, with sharp edges always present where tem- perature extremes causing uneven ex-
50

ECOLOGY OF HYRAX: TURNER AND WATSON
Figure B An adult tree hyrax on a rock in Ihe early rnorniwg.
pansions and contractions have recently exploded off a slab of rock. They pro- trude above the surrounding soils to a height of up to 40 m, presenting a beautiful and characteristic landscape. The kopjes under study (Figure 3) are 7-10 m high, and have basal circumfer- ences not exceeding 80 m. As a result of the explosive type of weathering which is continuously taking place most kopjes are characterised by large and small crevices and sheltered caves dif- fering considerably as regards climate from open ground outside the kopje.
The kopje acts as a natural water catchment system, and at the same time promotes condensation of dew and re- duces water loss through evaporation. These features in part explain the dense woody vegetation growing over and sur- rounding kopjes in the western Seren- geti. The surrounding vegetation of the kopjes being studied is open grassland with Pennisetum mezxictnncm and The- me& triundra e typical dominants.
51
Scattered Acacia tortilis (Forsk.) Hayne trees are Eound close to the kopjes, but the other woody vegetation is definitely associated with various modiiicatioins of the environment creat- ed by the kopje, a s mentioned in the previous paragraph.
Both species of hyrax are found only associated with kopjes and must there- fore be expected to be rather well adapted to the kopje habitat. A des- cription of some of these adaptations f ollo$vs.
SOME ADAPTATIONS OF THE TWO HYRAX SPECIES FOR THIE: KOPJE HABITAT
From the point of view of a small vertebrate the kopjc: habitat is remark- ably undemanding. When this habitat is compared with that presented by the surrounding open plains it becomes clear that kopjes are islands of con- stant environmental conditions sur- rounded by a sea of environmental

EAST AFRICAN WILDLIFE JOURNAL, VOL. 3, AUGUST 1965
extremes so great that only consider- able specialisation, such as burrowing, enables small vertebrates to live in it.
i. Considerations of Climate The interior crevices of the kopje
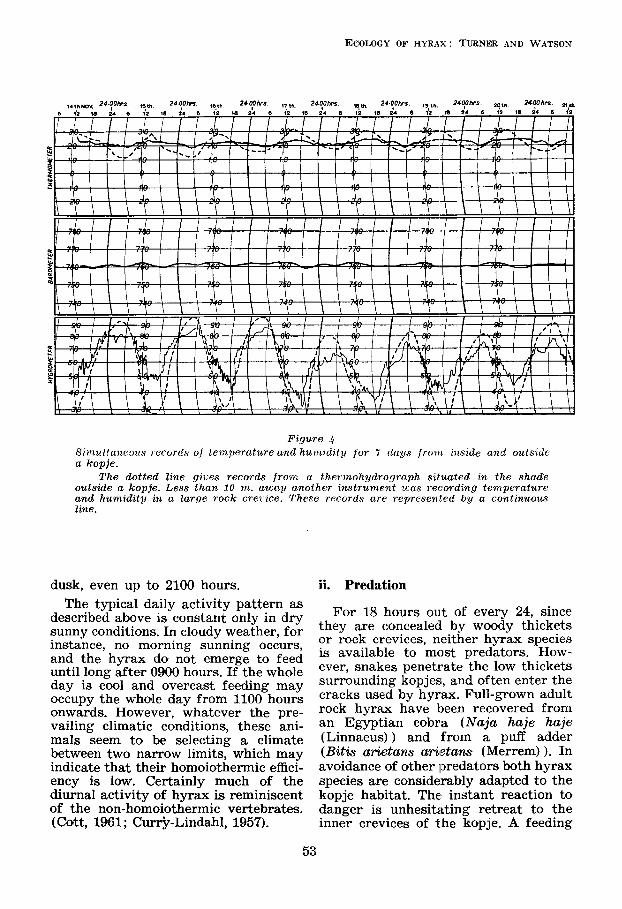
show much less violent extremes of temperature and humidity than the surrounding plains. The records from two thermo-hydrographs, one placed in a rock crevice in the heart of one of the study-kopjes, and the other placed in the shade just outside the kopje, have been superimposed to make Figure 4. Over the two-month period in which the thermohydrographs were operating the temperature inside the kopje never exceeded 25OC and never went below 17OC. The daily range was never more than 6OC. In contrast out- side the kopje the daily range was fre- quently as much as 18OC, with maxima of up to 34OC and minima down to 12°C.
Likewise the daily humidity range within the kopje only once exceeded 40 per cent., the highest humidity being recorded as 88 per cent., and the lowest as 32 per cent. Outside the kopje daily ranges of 60 per cent. were common, with maxima of 100 per cent. and minima of 12 per cent. (See Grzimek and Grzimek, 1960).
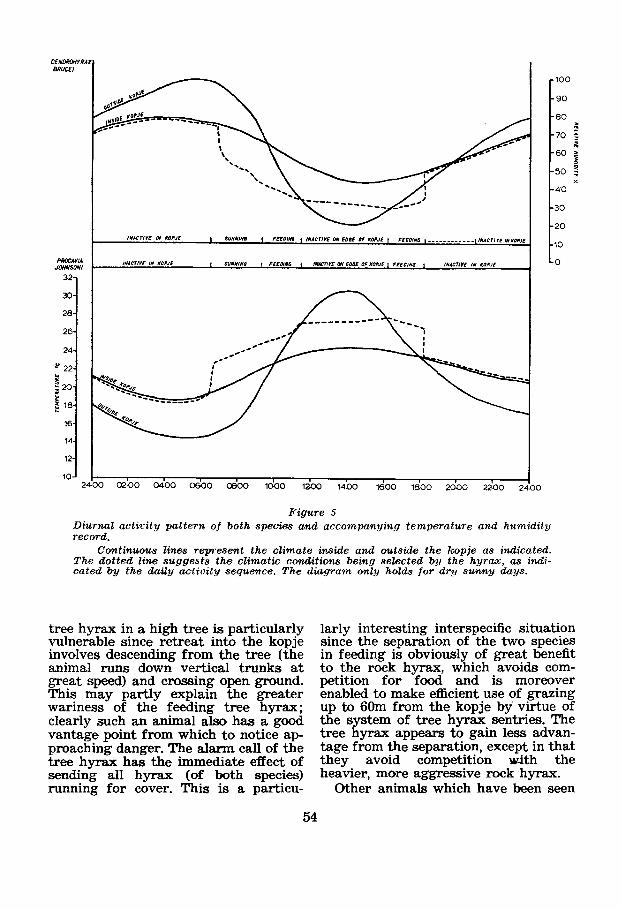
The diurnal activity pattern of both hyrax species is remarkably adapted to take advantage of the potential offered by two such different climatic environ- ments separated by no more than 1Om. This activity pattern is summarised in Figure 5. Both species show a remark- able constancy in this activity pattern ; at dawn they emerge from the crevices and climb to high rocks with an eastern aspect where they spread themselves to take the maximum radiation from the early morning sun. In the first stage of sunning all hyrax are grouped very closely together, touching other animals on all sides, with young often cIimbing on the backs of adults to burrow into the fur.
As the morning progresses the sunn- ing animals spread out and indulge in various grooming and stretching acti- vities. FYnally, over a period of about one hour, all the animals descend from
the sunning rocks, cautiously come out of the S a n s e z w thicket, and sttart to feed. In this activity the two species are completely separated. The tree hyrax climbs quickly into the selected tree or shrub to browse (Figure 6); the rock hyrax grazes the grassland in the immediate neighburhood of the kopje (Figure 7). Throughout most of the feeding period the animals are in the open, but rock hyrax notably seek more shaded grazing areas towards 1100 hours.
At about 1100 hours the majority of animals of both species return and rest in the rocks and undergrowth immedi- ately surrounding the kopje. They have not been observed t o return to the in- terior crevices during the day. At a b u t 1600 hours both species return to their typical feeding activity and continue feeding until a little after 1800 hours, whereupon all animals withdraw to the inner crevices of the kopje to spend the night. On moonlight nights, parti- cularly in the dry season, the tree hyrax has been observed feeding after
Figure 3 Part of the kopje under study sholuing exposed granite boulder and surrounding woody thicket.
52
ECOLOGY OF HYRAX: TURNER AND WATSON
Ffgura $ S:miiltaneoits 7 eccjrc's of temperature and hurnialty for 7 days f rom rnszde and outszdc a kopje.
T'he dotted lane gzLes records frorn n theimohydrograph sztuuted zn the shade outsade a kopje. Less than 10 m. away another znstrument was re(-ordang temperature and humzdaty an a large rock crezzce. These records are represen'ed by a continuous lane.
dusk, even up to 2100 hours. ii. Predation The typical daily activity pattern as
described above is constant only in dry sunny conditions. In cloudy weather, for instance, no morning sunning occurs, and the hyrax do not emerge to feed until long after 0900 hours. If the whole day is cool and overcast feeding may occupy the wholle day from 1100 hours onwards. However, whatever the pre- vailing climatic conditions, these ani- mals seem to be selecting a climate between two narrow limits, which ma.y indicate that their homoiothermic effici- ency is low. Certainly much of the diurnal activity of hyrax is reminiscent of the non-homoiothermic vertebrates. (Cott, 1961 ; Curjl-Lindahl, 1957).
For 18 hours out of every 24, since they are concealedl by wmdy thickets or rock crevices, neither hyrax species is available to most predators. How- ever, snakes penetrate the low thickets surrounding kopjes, and often enter the cracks used by hyrax. Full-grown adult rock hyrax have been recovered from an Egyptian cobra (Nuja haje haje (Linnaeus)) and from a puff adder (Bitis arietans ariletans (Merrem) ). In avoidance of other predators both hyrax species are considerably adapted to the kopje habitat. The, instant reaction to danger is unhesitating retreat to the inner crevices of the kopje. A feeding
53
32-
30-
28-
26-
24-
22- u
2 23- k 5 18-
16-
14-
12-
10-
-100
- 90 I 2
-70 : ::
-60 I
-50
- 40 -30
-20
-10
-0
Y
Figure 5 Diurnal acticity pattern of both species and accompanying temperature and humidity record.
Continuous lines represent the clim.ate inside and outside the hmpje as indicated. The dotted line suggests the climatic conditions being sekcted by the hyrax, as indi- cated by the daity activity sequence. The diagram only hofda for d r y sunny days.
tree hyrax in a high tree is particularly vulnerable since retreat into the kopje involves descending from the tree (the animal runs down vertical trunks at great speed) and crossing open ground. This may partly explain the greater wariness of the feeding tree hyrax; clearly such an animal also has a good vantage point from which to notice ap- proaching danger. The alarm call of the tree hyrax has the immediate effect of sending all hyrax (of h t h species) running for cover. This is a particu-
larly interesting interspecific situation since the separation of the two species in feeding is obviously of great benefit to the rock hyrax, which avoids com- petition for food and is moreover enabled to make efficient use of grazing up to 60m from the kopje by virtue of the system of tree hyrax sentries. The tree hyrax appears to gain less advan- tage from the separation, except in that they avoid competition with the heavier, more aggressive rock hyrax.
Other animals which have been seen
54

ECOLOGY OF HYRAX: W N E R AND WATSON
to prey on hyrax during the vulnerable six hours feeding period are leopard (Panthers parcihs fusca (Meyer) ), caracal (Caracal carucul nub& (Fis- cher)) and large birds of prey, such as the martial eagle (Polemaetus belli- c o w (Daudin) 1.
iii. Fire
Grass fires sweep across the Seren- geti plains more or less every year. Some of the kopje vegetation may be burnt, but this is generally protected by the surrounding fire-resistant thicket. Fire never penetrates the inner crevices of the kopjes because they are devoid of vegetation. Once again retreat into the kopje is the hyrax’s answer to the problem.
iv. Water
As mentioned earlier, the kopje acts as a water catchment system and allows condensation in certain conditions. Nevertheless a t the height of the dry season the kopjes under study appear to have no natural water supplies and hyrax must be capable of life without surface water for at least short periods.
v. Food availability
This is dealt with in greater detail in a later section.
At this stage i t IS sufficient to remark that the shrubs growing on the kopje and the grass immediately surrounding it benefit from the physical character of the kopje in respect of catchment, condensation and shade. Many of these shrubs, important food for the tree hyrax, are confined to kopjes. These plants and small peripheral areas of grassland are growing throughout most of the year, and so present a constantly palatable food source.
Furthermore the deposits of faeces and urine within the kopje (all the hyrax defecate arid urinate in one or two places only in the kopje, which Serves to avoid fouling the smaller crevices used for sleeping) are regularly flushed through with water running off rock surfaces. This water carries back a large proportion of the fertility to the plants surrounding the kopje, and in a sense a small closed fertility cycle may be said to be operating.
E’OOD PREFERENCES
Table I lists the available food plants growing on the kopje under study and notes those plants (eaten by either hyrax species. No attempt has been made to assess the relativle importance of the food plants, but in the case of Dendro- hyrax brucei a b u t 90 per cent. of re- cordings have been of Acacia tm-tdk and Albphyltm rubifolius (Hochst.) Elngl.
Figure 6 A tree hyrax browsing A C U C ~ tortizis - their
main diet .
The immediately striking fact is the lack of overlap in diets. The possibility that the provision of water and kitchen
55

EAST AFRICAN WILDLIFE JOURNAL, VOL. 3, AUGUST 1965
TABLE 1
Listing food plants available to Procavia johnstoni and Dendrohyrax brucei, and notiny those eaten by each
Plant n a i x
PTERIDOPHYTES Adiantaceae
Actinzopterzs aiutralzs (L.f.) Link. Pellaen crrlomelanos (Sw.) Link.
AspZenzunt 7 utaefoltum (Berg.) Kze
Selagine!la wightaa Hieron var. phzllipsiann Hieron
Aspleniaceae
Selaginellaceae
ANGIOSPERMS Agavaceae
Amaranthaceae
Snnsei ierza robusta N.E. Br
-4c~iyranthes aspera L. Pupalin lnppacea (L.) JUSS.
Haevxaizthus niultzflorus Martyn
Tylophorn sp. (Greenway 10,142)
Amaryllidacea@
hsclepiadaczae
Boraginaceae Cordirx ocualis R. Br. ex DC.
Eurseraceae Comniiphora eminii Engl. Commiphora trothae Engl.
Capparis fascicularis DC. Capparidaceae
ssp. elaeagnoides (Gilg) De Wolf
dlnerua triphyZZa A. Rich. var. johannis (Volkens & Gilg) De Wolf
Celastraceae Cassine aethiopica Thuilb.
Notonia gregorii S. Moore Notonia petraea (Muschl.) R. E. Fries
Grassula pentandra (Royle ex Edgw.)
KaZanchoe marmorata Bak.
Compositae
Crassulaceac
Schoenland
Cucurbitaceae Momordica trifo'liolata Hook. f .
Zuphorbiaceae Acalypha fruticosa Forsk. Phyllanthus sepialis Muell. Arg.
Plant form Observed Okserved eaten by eaten by Pr ocav ia johnstoni brucei
Dendr o h yr ax
herb
herb herb
herb
climbing shrub
shrub
tree tree
climbing shrub
shrub
tree
herb herb
herb
herb
climbing herb
shrub shrub
*
56

ECOLOGY O F HYR.4X: TURNER AND WATSON
Plant name
Gramineae Andropogon sp. (Greenway 9,900 & 9,975) Aristida adscensimis L. Brmhiaria eruciformis (Sw.) Griseb. Cenchrus ciliaris L. Chluris roxburghiana Schult. Chloris pycnothrix Trin. Cynodm dactylon (L.) Pers. Cynodon plectostachyus (K. Schum.) Pilger Dactyloctenium aeggptium (L.) Beauv. Digitaria macroblephara (Hack.) Stapf En,neapogon elegans (Nees) Stapf Eragrostis aspera (Jacq.) Nees Eragrostis cilianewis (All.) Lutati Eragrostis tenuifolia Hochst. ex A. Rich. Harpach,m schimperi Hochst. ex A. Rich. Heteropogm contortus (L.) Beauv. ex
Hyparrhenia dissoluta (Nees ex Steud.)
Hyparrhenia filipendula (Hochst. ex Steud.) Stapf
Loudetia arund&mea (Hochst. ex A. Rich)
Microchloa kunthii Desv. Panicum maximum Jacq. Pennisetum stramineum Peter Rhynchelytrum repens (Willd.) C. E. Hubb. Sporobolus pyromi.dalis Beauv. Themeda triandra Forsk. Trachypogon spicatus (L.f.) Ktze. Tragus berteronianus Schult.
Hoslundia opposita Vahl
Iboza sp. (Greenway 10,350) Plectranthus Jlaccidus Guerke Pbctranthus prostratus Guerke
A b s macrosiphon Bak. Aloe seczcndiflora Engl. Aloe volkensii Ehgl.
Hibiscus sp. (Greenway 9,926)
Turrwa fischeri Guerke
Acacia tortilia (Forsk.) Hayne
Albizia hurveyi Fourn.
Ficus sonderi Miq.
Rhynchosia chrysadenia Taub.
Roem. & Schult.
C. E. Hubb.
Steud. var. hensii (De Wild.) C. E. Hubb.
Labiatae
Liliaceae
Malvaceae
Meliaceae
Mimosaceae
ssp. spbocarpa (Hochst. ex A. Rich.) Brenan
Moraceae
Papilionaceae
herb herb herb herb herb herb herb herb herb herb herb herb herb herb herb herb
Plant form Observed Observed eaten by eaten by Procnvia Dendrohyrax johnstoni brucei
*
* * * * *
* * herb
herb
herb
herb herb herb herb herb herb herb herb
shrub
shrub herb herb
herb herb tree
shrub
tree
tree
tree
tree
climbing herb
* *
* *
*fruit only
* *
* *seeds *
only
57

EAST AFRICAN WILDLIFE JOURNAL, VOL. 3, AUGUST 1965
Plant name
Plumbaginaceae
Portulacnceae
Plurnbago zeylanica L.
Talinun? cnffrzCm Eckl. & Zeyh.
Ziziphirs mucronata Willd.
Pai'etta hochstetteri Brem. Tarenwa grai.eolens ( S . Moore) Brem.
Rhaninaceae
Ruhiaceae
Rutaceae Teclea nobilis Del. Teclea simplicifolia (Engl.) Verdoorn Teclea trichocarpa (Engl.) Engl.
AZZophyZus rubifoZius (Hochst.) Engl. Pappen capensis (Spreng.) Eckl. & Zeyh.
Craterostignia plnntaginewm. Hochst,
Grewia f a l l a z K. Schuin.
Obetia p inmt i f i da Bak.
Rhoicissus rer.oilii Planch.
Sapindaceae
Scrophulariaceae
Tiliaceae
Urticaceae
Vitaceac
refuse is obscuring the true position cannot be overlooked, although such observations as have been made on other kopjes do not differ from the observations on our own kop je. The rock hyrax is an almost exclusive grazer, but also eats the seeds of Acacia tortilk and some Commiphsra trothae Engl. ; the tree hyrax is an exclusive browser. The two A b e species eaten by the rock hyrax were introduced to the kopje by the senior author's wife. In respect of feeding therefore the two species of hyrax co-habiting many kopjes in the Serengeti are completely separated, whereas in many other activities they are indistinguishable. The significance of this finding is considered in the final section. At this stage it is fruitful to point out that the larger herbivores of the Serengeti plains whose ranges over- lap have diets of a more or less over-
Plant form Observed Observed eaten by eaten by Procavia Dendrohyrax johns tmi brucei
shrub
herb
tree
shrub shrub
tree tree tree
shrub tree
herb
shrub
shrub
climbing shrub
lapping nature (Watson and Kerfoot, 1964 ; Om, 1964) although successional considerations to some extent obscure the picture.
However, one of us (R.M.W.) has evidence from field observations that the degree of this overlap in diet is far greater in the dry season than in the wet. From this comparison we can ten- tatively suggest that the degree of overlap in diet reflects the pressure on food resources at that time. And so. considering the kopje ecosystem, i t is possible that population levels for each species are dictated by crevice avail- ability rather than food availability, and that the grazing/browsing prea- sure never reaches a high level. The authors are at present investigating the population ecology of the two species in a single group of kopjes.
58

DISCUSSION ON THE EVOLUTION OF THE KOPJE-DWELLING HABIT IN ROCK AND
TREE HYRAX
Returning to the food preferences of the two species it is of interest to note that Procavia johnstoni grazes a fairly wide selection of grasses but does not browse any of the plants which can be considered characteristic of kopjes. Similarly Dendrohyrm k c e i , although browsing shrubs typically associated with kopjes, relies on Acacia tortilk for the bulk of its food. Kopjes otherwise suitable for habitation by both species of hyrax are found to hold only Pro- cavia johnstmi if no Acacia tortilis grows nearby.
And so an additional explanation of the extreme separation of diets i s offer- ed: we consider Hyracoidea to have been originally forest-dwelling, as is the present-day Dendrohyrax aalidus. At some stage, -possibly as the forest started t o give way to woodland, a form
took to grazing and lost some forest- dwelling features. As the forests con- tinued to recede, this woodland hyrax, already separated from the forest forms in diet and habits, became confined to the small islands of forest and wood- land associated with kopjes. The change involved from liviing in hollow tree trunks to living in rock crevices would not seem to be grext.
At the same time arboreal forms, ini- tially quite different in habits from the ground-living hyrax, became confined to kopjes and developed the same adapta- tions for kopje life, So the arboreal and ground-living hyrax reconverged. But no convergence of diet or feeding habits took place because space considerations inside the kopje never allowed popula- tions to put pressure on the food re- sources.
The authors consider the separation of diets and feeding habits to be so pro- nounced as to be unlikely to have arisen from food competition in the past. As
Figure 7 A group of rock hyra& grazing at the edge of the kopje. Part of the Sanstxieria thicket
can be seen.
59

stated earlier, seasonal increase in the pressure on food resources in the case of the large herbivores of the Serengeti results in increased overlap in diet. And even in the wet season plains game have very overlapping diets, at least in terms of plant species eaten. Competition for food among the plains game has pro- bably resulted in no separation of diet of the sort seen between the two hyrax species, but rather a separation between growth stages and components of the pIants eaten. This kind of separation of diet in response to competition appears to be very efficient, and therefore i t is difficult to assign the separate diets and feeding habits of the two hyrax species to competition.
It is interesting to reflect on why Procawia johnstoni did not develop the burrowing habit, which would have given it potential to extend its range enormously. However, as long as climb- ing (even of a limited nature up rock faces) is part of an animal’s repertoire, burrowing activity seems to be pre- cluded, for only a small proportion of animals indulge in both activities.
ACKNOWLEDGEMENTS We are extremely grateful to Dr. P. J.
Greenway, O.B.E., for identifying all the plants mentioned in this paper. Mrs. K. Turner has helped in various record- ing activities, and is responsible for the tameness of the animals. The hyrax have been identified by Mr. R. W. Hay- man of the British Museum (Nat. Hist.), and his description is used. Capt. C. R. S. Pitman has given us valuable advice, and interesting historical infor- mation has come from A. Cruickshank. The thermohydrographs used have been supplied through Professor B. Grzimek.
Dr. P. E. Glover have given valuable advice during the preparation of this paper. The senior author acknowledges the permission of MI-. J. S. Owen, O.B.E., Director of Tanzania National Parks, to present this paper; the junior author acknowledges the permission of Dr. P. E. Glover and F. A. 0. t o present this paper.
REFERENCES ANON (1850). Animal Substances. Pkwrn. J . ,
Sept : 827-831. BURTON, M. (1961). Rock hyrax. Lmd. I l l .
News, Sept. 9th, p. 420. COTT, H. B. (1961). Scientific results of an in-
quiry into the ecology and economic s ta tus of the Nile crocodile (Croco&ylzcs niZoticu.9) in Uganda and Northern Rhodesia. T r a m .
CURRY-LINDAHL, K. (1957). Behaviour of the tropical rock lizard A g a m a cyanogaster (Ruppell) in hot environments. Ann. SOC. xool. Belg., 87 : 45-74.
GRZIMEK, M. and GRZIMFX, B. (1960). A study of the game of the Serengeti p:ains. 2. Saugetierk., 25 : 1-61.
HAYMAN, R. W. (1964). Letter dated 29th Oct. 1964 to C. R. S . Pitman. Br. Mus. (Nat. Hist.) Lond.
LINNAEUS, C. V. (1758). Systema Naturae. I. Uppsala.
ORR, D. J. C. (1964). Grazing succession of game animals in Ngorongoro Crater. Tan- gnnyika Game Department, Dar 68 Salaam. Typescript, 30 pp.
OWEN, R. (1832). Proc. 2001. SOC. Lond., quoted in ANON (1850).
SALE, 3. (196.0). The Hyracoidea. A review of the systematic position and biology of the hyrax. J . E. A f r . nnt. Hist . SOC., 23(5) :
THUNBERC (1779). Travels in Europe, Asia and Africa between the years 1770 and 1779. Quoted in ANON. (1850).
WATSON, R. M. and KEKFOOT, 0. (1964). A Short note on the intensitv of eTazinP of the
~ 0 0 1 . SOC. L m d . , 29(4) : 211-356.
185-188.
Serengeti plains by ilains->ame.- 2. Suu- getjerk., 29(5) :337-320.
Authors’ address: M. I. M. Turner, Serengeti National Park, and R. M. Watson, Serengeti Research Project, “Michael Grzimek Memorial Laboratory”, P. 0. Bos 3134, Arusha, Tanzania.
(Received for publication January, 1965)
60





























