Embed Size (px)
Citation preview

lable at ScienceDirect
Quaternary Science Reviews xxx (2014) 1e16
Contents lists avai
Quaternary Science Reviews
journal homepage: www.elsevier .com/locate/quascirev
An investigation of palaeodietary variability in European Pleistocenecanids
Lucy O.H. Flower*, Danielle C. SchreveCentre for Quaternary Research, Department of Geography, Royal Holloway, University of London, Egham, Surrey TW20 0EX, UK
a r t i c l e i n f o
Article history:Received 10 April 2013Received in revised form4 April 2014Accepted 12 April 2014Available online xxx
Keywords:DietWolvesCarnivoresDentitionBody sizeCanis lupusCanis mosbachensisCanis etruscusClimate
* Corresponding author.E-mail address: [email protected] (
http://dx.doi.org/10.1016/j.quascirev.2014.04.0150277-3791/� 2014 Elsevier Ltd. All rights reserved.
Please cite this article in press as: Flower, LQuaternary Science Reviews (2014), http://d
a b s t r a c t
Temporal and interspecific dietary variability were investigated in three canid taxa, Canis lupus, Canismosbachensis and Canis etruscus, across a range of British and mainland European wolf assemblages fromthe Early Pleistocene to Recent periods. Using established cranio-dental indicators to reveal dietaryspecialisations towards bone eating, flesh slicing, and non-flesh food crushing, inferences were madeconcerning the proportions of flesh to non-flesh foods in the diet, and hence the level of carnivoryadopted by each taxon. Significant temporal differences were found in the diet and frequency of toothwear of C. lupus fromMIS 3, 5a and 7 in Britain. Relative body size comparisons based on lower carnassiallength also revealed variation in body size for the Pleistocene age groups, correlating with differences indiet. Stepwise Discriminant Function Analyses revealed large-bodied MIS 5a C. lupus to be hyper-carnivorous and specialised in fast flesh slicing and to some extent bone consumption, whereas relativelysmaller-bodied MIS 3 and 7 C. lupus were both less carnivorous and more specialised in crushing non-meat foods. Modern wolves from central Sweden are smaller than those of MIS 5a and hyper-carnivorous, although with greater specialisation towards crushing of non-meat foods. Temporal varia-tions in diet were related to changes in prey diversity, competition from other carnivores, openness ofthe environment, and ultimately climate, and reflect the cranio-dental plasticity of C. lupus. In contrast,no temporal differences in diet were found in age groups of C. mosbachensis and C. etruscus, which mayrelate to more stable overall conditions in comparison to the later Pleistocene. The cranio-dental char-acteristics of the smaller-bodied mesocarnivore C. etruscus indicate adaptations to non-meat foodcrushing, whereas in the similarly small C. mosbachensis, enhanced flesh slicing capabilities and reducedcrushing abilities indicate that it was more carnivorous than C. etruscus. C. etruscus and C. mosbachensiswere both more specialised than C. lupus.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
This paper aims to investigate temporal variation in the diet ofone of the most successful Pleistocene predators, the grey wolfCanis lupus L., 1758, as well as analysing potential dietary variabilitybetween C. lupus and two other Pleistocene canids, the EarlyPleistocene Canis etruscus Forsyth-Major, 1877, and the Early toMiddle Pleistocene Canis mosbachensis Soergel, 1925. Establishingthe relative ecological roles of the different canids within achanging carnivore guild is critical for our understanding of Pleis-tocene palaeoecology and in particular, the importance of intra-and inter-specific competition. Changing dietary behaviour can betraced in different aspects of cranio-dental morphology, including
L.O.H. Flower).
.O.H., Schreve, D.C., An invesx.doi.org/10.1016/j.quascirev
the morphology of the premolars and molars, depth of themandible and degree of toothwear (Van Valkenburgh,1988a,b; VanValkenburgh, 1991; Van Valkenburgh and Koepfli, 1993). It istherefore anticipated that any morphological changes identifiedmay shed light on temporal and geographical variation in the diet ofPleistocene wolves, which may in turn be linked to changes in preydiversity, competition, environment and climate. By using theecology and subsistence behaviour of modern C. lupus as ananalogue, inferences may be drawn concerning the palaeoecologyand palaeodiet of Pleistocene C. lupus, as well as allowing com-parison with extinct canid taxa in order to establish ecologicalniches of individual species. Finally, dietary information frommodern Palaearctic wolf specimens will be compared to the Pleis-tocene dataset, in order to see whether there are differences inmorphology that might equally be explained by environmental orother parameters. The paper adopts an explicitly British perspectivefor much of the palaeodietary component, in order to maximise
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015
OLD.
JAR.
BRU.0.78MAT.
BLA.0.12
EMP0.42
REU.
00.
20.
40.
60.
81.
01.
21.
41.
61.
82.
02.
22.
4
LATE
MID
DLE
EA
RLY
PLE
ISTO
CE
NE
Aur
elia
nG
aler
ian
Late
Vill
afra
nchi
anE
pi-
Villa
franc
hian
Mid
dle
Villa
franc
hian Coste San Giacomo
Olivola
Upper Valdarno, Fonelas P-1
Untermassfeld
West Runton
Westbury-sub-Mendip Boxgrove, Sidestrand
Venta Micena
C. e
trusc
us
C. m
osba
chen
sis
C. l
upus
C. f
alco
neri
C. (
X.)
lyca
onoi
des
C. a
rnen
sis
Euro. Mam. Ages
Age Ma.
MISODP 677δ 18O to PDB (‰)
5e
7
9
11
5 4.5 4 3.5
25
63
Key Sites
Swanscombe
GraysThurrock, Polledrara diCecanibbio
Presence of Canids
A.
Mag. Polarity
Apollonia - 1
Senèze
??
3 See A.
2.6 MAT.
GAU.
91
103
Pirro Nord
*
**
BLA. 0.12
ODP 677δ 18O to PDB (‰)
Key Sites Euro. Mam. Ages
Age Ma.
Mag.polarity
0.2
0
PLE
ISTO
CE
NE
LATE
HOL.
Aur
elia
n
5 3.544.5
7
3
Banwell Bone Cave, Bosco’s Den, Steetley Quarry Cave, Stump Cross Cave, Windy Knoll, Wretton
Black Rock Quarry, Kent’s Cavern, Oreston Cave, Paviland, Pin Hole Cave, Sandford Hill, Uphill
Bleadon Cave, Hutton Cave, Ilford, Marsworth, Pontnewydd Cave
A. MIS
5a
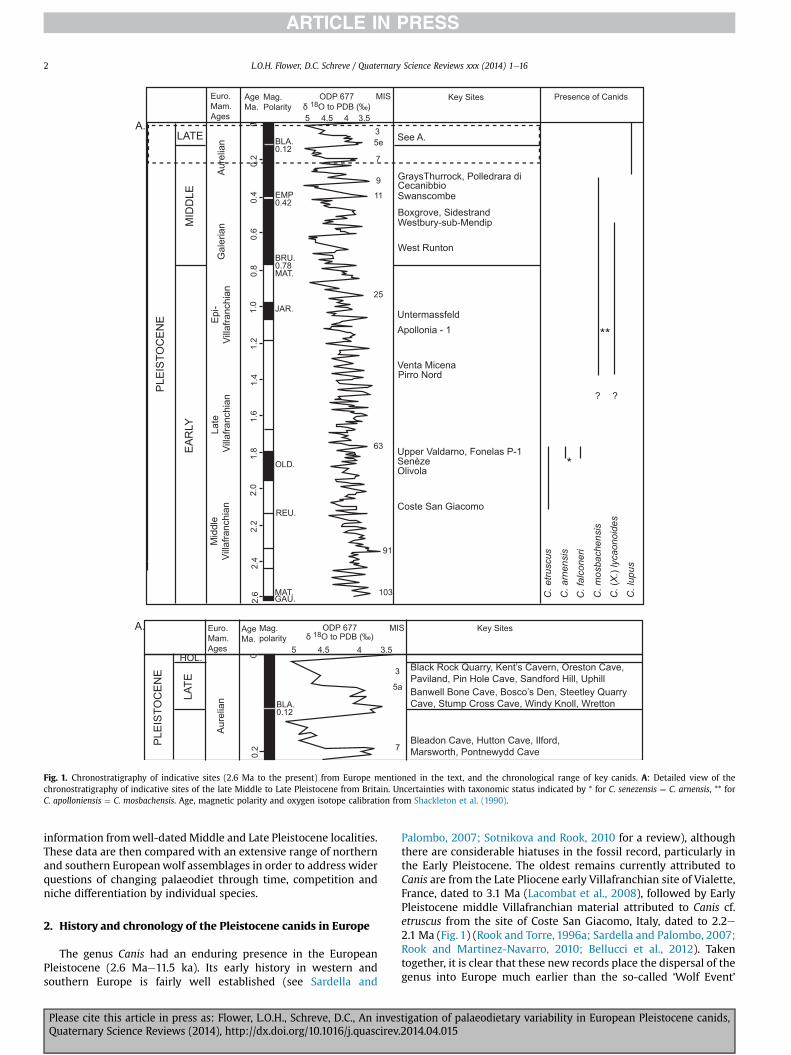
Fig. 1. Chronostratigraphy of indicative sites (2.6 Ma to the present) from Europe mentioned in the text, and the chronological range of key canids. A: Detailed view of thechronostratigraphy of indicative sites of the late Middle to Late Pleistocene from Britain. Uncertainties with taxonomic status indicated by * for C. senezensis ¼ C. arnensis, ** forC. apolloniensis ¼ C. mosbachensis. Age, magnetic polarity and oxygen isotope calibration from Shackleton et al. (1990).
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e162
information fromwell-dated Middle and Late Pleistocene localities.These data are then compared with an extensive range of northernand southern Europeanwolf assemblages in order to address widerquestions of changing palaeodiet through time, competition andniche differentiation by individual species.
2. History and chronology of the Pleistocene canids in Europe
The genus Canis had an enduring presence in the EuropeanPleistocene (2.6 Mae11.5 ka). Its early history in western andsouthern Europe is fairly well established (see Sardella and
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
Palombo, 2007; Sotnikova and Rook, 2010 for a review), althoughthere are considerable hiatuses in the fossil record, particularly inthe Early Pleistocene. The oldest remains currently attributed toCanis are from the Late Pliocene early Villafranchian site of Vialette,France, dated to 3.1 Ma (Lacombat et al., 2008), followed by EarlyPleistocene middle Villafranchian material attributed to Canis cf.etruscus from the site of Coste San Giacomo, Italy, dated to 2.2e2.1 Ma (Fig. 1) (Rook and Torre, 1996a; Sardella and Palombo, 2007;Rook and Martinez-Navarro, 2010; Bellucci et al., 2012). Takentogether, it is clear that these new records place the dispersal of thegenus into Europe much earlier than the so-called ‘Wolf Event’
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e16 3
originally recognised by Azzaroli (1983) from the late Villa-franchian typesite of the Olivola Faunal Unit at Val di Magra (Italy),which contained abundant remains of C. etruscus and was conse-quently taken as evidence for a widespread expansion of the spe-cies into Europe (Azzaroli et al., 1988).
Subsequently, other members of the genus Canis appeared inEurope, notably the coyote-like Canis arnensis Del Campana, 1913(Sardella and Palombo, 2007; Sotnikova and Rook, 2010), which co-occurred with C. etruscus and the hunting dog-like Canis falconeriForsyth-Major, 1877 (Rook, 1994) in sites of the Upper ValdarnoBasin (Tasso F.U. e late Villafranchian) (Fig. 1). However, complexissues remain regarding the systematics and phylogenetic re-lationships between the canids of the Early Pleistocene in Europe.For example Canis senezensis Martin, 1973 from Senèze (France)may be a less derived form of C. arnensis (Rook and Torre, 1996a),Canis apolloniensis Koufos and Kostopoulos, 1997 from Greece mayrepresent an early record of the small Middle Pleistocene wolf, C.mosbachensis (Garrido and Arribas, 2008a,b), and the status of bothC. falconeri and the later Canis (Xenocyon) lycaonoides Kretzoi, 1938as members of the modern African wild dog genus Lycaon (Rook,1994; Martinez-Navarro and Rook, 2003; Werdelin and Lewis,2005) remains controversial.
Nevertheless, it is clear that the previously large and diversecarnivore guild of the Early Pleistocene was in decline by the LateEarly and Middle Pleistocene, resulting in changes in the body sizestructure of the guild (Croitor and Brugal, 2010; Meloro and Clauss,2012), and the appearance of new taxa, such as C.mosbachensis andC. (X.) lycaonoides (Fig. 1) (Sotnikova and Rook, 2010) both withearliest co-occurrences at Venta Micena in Spain (Rook andMartinez-Navarro, 2010) and Pirro Nord, Italy (Petrucci et al.,2013). By the early Middle Pleistocene, an early form of theAsiatic dhole Cuon priscus Thenius, 1954 was also present insouthern Europe at Petralona Cave, Greece, (Kurtén and Poulianos,1977), and both Hundsheim (Thenius, 1954) and Mosbach (Schutt,1973) in Germany.
C. lupus first appeared in Italy during Marine Oxygen IsotopeStage (MIS) 9 at La Polledrara di Cecanibbio (Gliozzi et al., 1997)(Fig. 1). In Britain, it was the only canid species present from MIS 7onwards, its appearance linked by Turner (1995) to an increase inbody sizewhen compared to the smaller remains of C. mosbachensisfrom the early Middle Pleistocene. The oldest British record of thespecies is from Pontnewydd Cave in north Wales (Currant, 1984;Turner, 1995). Records of C. lupus are abundant in Britain and on theEuropean mainland for the Late Middle and Late Pleistocene, underboth temperate and cold-climate conditions, and the species suc-cessfully persisted into the Holocene. However, although C. lupus istoday widely distributed in the Northern hemisphere in a variety ofhabitats, it has notably restricted ranges in western and northernEurope, where remnant populations are highly fragmented (Mechand Boitani, 2010) and range is greatly restricted by humans, indirect contrast to their widespread Palaearctic Pleistocenedistribution.
It should be noted here that C. etruscus, C. mosbachensis andC. lupus are considered to form an evolutionary lineage (Kurtén,1968; Torre, 1979; Rook and Torre, 1996b; Sotnikova, 2001;Cherin et al., 2013a). However, arguments exist over the phyloge-netic position of C. mosbachensis regarding its status as a species orsubspecies in relation to C. lupus, as well as its possible closerphylogenetic relationship to C. arnensis (Garrido and Arribas,2008a; Martinez Navarro et al., 2009). Since the focus of this pa-per is to examine dietary variability rather than lineages, discussionof the evolutionary position of C. mosbachensis will not be consid-ered further and in order to avoid presumptive linkage to C. lupus,C. mosbachensis will not here be assigned to a subspecies.
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
3. Chronological and palaeoenvironmental context ofassemblages studied
Analysis of Late Middle and Late Pleistocene C. lupus palaeodietin Britainwill focus on assemblages from three climatically-distinctepisodes, the penultimate interglacial (correlated with MIS 7), theearly Devensian (¼ early Weichselian, correlated with MIS 5a) andtheMiddle Devensian (MIS 3). Thesewere deliberately selected, notonly because they contain abundant wolf material from a variety ofwell-dated depositional contexts, but also because they reflectdifferent palaeoenvironments with distinctive faunal compositions.
Wolf material from the penultimate interglacial comes fromBleadon Cave (Somerset), Crayford (Kent), Hutton Cave (Somerset),Ilford (Essex), Marsworth (Buckinghamshire), Pontnewydd CaveIntermediate Complex and Lower Breccia (Denbighshire) and Tor-newton Cave Otter Stratum (Somerset) (Fig. 2). These have beenattributed to MIS 7a, the last of three temperate substages of more-or-less equal magnitude within this interglacial (Schreve, 2001;Candy and Schreve, 2007). Assemblages of this age are assignedto the Sandy Lane Mammal Assemblage-Zone (MAZ) (Schreve,2001), characterised by the presence of steppe mammoth (Mam-muthus trogontherii) and horse (Equus ferus), with abundant lion(Panthera leo) and brown bear (Ursus arctos) and rarer spottedhyaena (Crocuta crocuta) and leopard (Panthera pardus) amongstthe large carnivores. Late MIS 7 environments were dominated byopen grasslands, with mean summer temperatures between 16 �Cand 23 �C and the mean winter temperatures between �7 �C and6 �C (Coope, 2010).
The Early Devensian group is represented by the sites of BanwellBone Cave (Somerset), Bosco’s Den (West Glamorgan), and SteetleyQuarry Cave (Nottinghamshire), Stump Cross Cave (North York-shire), Windy Knoll (Derbyshire) and Wretton (Norfolk) (Fig. 2).Both Banwell Bone Cave and Stump Cross Cave have been corre-lated with MIS 5a on the basis of Uranium-series dating by Gilmouret al. (2007) and assigned to the Banwell Bone Cave MAZ by Currantand Jacobi (2001, 2011). Sites of this age are characterised by a lowdiversity mammalian assemblage dominated by reindeer (Rangifertarandus) and bison (Bison priscus), with a very large form of brownbear (originally misidentified as polar bear on account on its greatsize) as the main predator and C. lupus the only other large carni-vore present (Currant and Jacobi, 2001; Currant, 2004). Spottedhyaena is notably absent from this fauna (Turner, 1981, 2009). Theprevailing environment was one of cold, open tundra with palae-otemperature reconstructions based on beetle assemblages typi-cally reconstructing mean summer temperatures of 7 to 11 �C andwinter temperatures of �10 to �30 �C in Britain (Maddy et al.,1998).
The Middle Devensian represents a marked shift towards abroadly more temperate part of the last cold stage, albeit onepunctuated by rapid climatic oscillations (Grootes et al., 1993). Themammalian fauna has been attributed to the Pin Hole MAZ byCurrant and Jacobi (2001, 2011) and is characterised by woollymammoth (Mammuthus primigenius), woolly rhinoceros (Coelo-donta antiquitatis), horse, giant deer (Megaloceros giganteus), asrepresented at sites such as the Kents Cavern Cave Earth (Devon)and Paviland (West Glamorgan), as well as Black Rock Quarry(Pembrokeshire), Oreston Cave (Devon), Pin Hole Cave (Derbyshire)and Sandford Hill (Somerset) (Fig. 2). Brown bear and lion arepresent but the dominant large carnivore appears to be spottedhyaena. The prevailing environment was open grassland but onethat supported exceptionally rich herbivore biomass (eg. Zimovet al., 2012). In Britain, mean summer temperatures have beenreconstructed at around 12 �C in summer and �20 �C in winter (eg.Schreve et al., 2013).
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

5
2016
18
17
14
0 100 km
23
8
6
3
7
10
1119
12
9
1
2213
4
2115
24
2526
0 400 km
A
A
2
Fig. 2. Map of northwest Europe indicating German and Italian sites used in the analysis. A: detailed map of Britain illustrating sites used in the analysis. Sites indicated in Britain: 1:Banwell Bone Cave, 2: Black Rock Quarry, 3: Bleadon Cave, 4: Bosco’s Den, 5: Boxgrove, 6: Crayford, 7: Hutton Cave, 8: Ilford, 9: Kents Cavern, 10: Marsworth, 11: Oreston Cave, 12:Paviland, 13: Pin Hole Cave, 14: Pontnewydd Cave, 15: Sandford Hill, 16: Sidestrand, 17: Steetley Quarry Cave, 18: Stump Cross Cave, 19: Tornewton Cave, 20: West Runton, 21:Westbury-sub-Mendip, 22: Windy Knoll, 23: Wretton. Sites indicated in Europe: 24: Untermassfeld, Germany, 25: Olivola (Val di Magra), Italy, 26: Upper Valdarno Basin, Italy.
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e164
For the Early Pleistocene and early Middle Pleistocene, it wasnecessary to extend the geographical range of the study to conti-nental Europe. For C. etruscus, comparisons weremade between theOlivola and Tasso F.U.s (both Late Villafranchian) from Italy. Bothtemporal groups contain similar a faunal spectrum, albeit withyounger sites in the Upper Valdarno Basin differentiated from thoseof the Olivola F.U. by the disappearance of species such as therupicaprine, Gallogoral meneghinii, and the gazelle, Gazellospiratorticornis (Gliozzi et al., 1997) and the first appearance ofC. arnensis and C. falconeri. It is worth noting however that bothF.U.s were considered by Raia et al. (2006) as part of a singlepalaeocommunity.
One of the early appearances inwestern and southern Europe ofC. mosbachensis, together with C. (X.) lycaonoides is at the Late EarlyPleistocene interglacial site of Untermassfeld in Germany(Sotnikova, 2001). This locality has been correlatedwithMIS 31 andassigned to the Epivillafranchian (Kahlke, 2000; Kahlke et al., 2011).In Britain, C. mosbachensis is first recorded in the early MiddlePleistocene Cromer Forest-bed Formation (CF-bF) at West Runton(Norfolk), as ‘small’ C. lupus by Stuart (1995), and as C. mosbachensisby Lewis et al. (2010). West Runton is assigned to the ‘CromerianComplex’ of interglacials, which can be subdivided into an oldergroup characterised by the archaic water voleMimomys savini and ayounger group with Arvicola mosbachensis (¼ cantiana). For thepresent analyses, large assemblages of C. mosbachensis from thecave site of Westbury-sub-Mendip (Somerset) and Boxgrove (WestSussex) were included, both of which may be placed within theyounger group and correlated with MIS 13 or possibly 15 (Preeceand Parfitt, 2000).
Recent C. lupus specimens were also included in the analyses,based on a large assemblage of modern wild-caught adult
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
individuals from central Sweden. Sweden has a continental climate,with large temperature ranges from cold winters to warm summers(for Uppsala: winter 1 �C to�10 �C, summer 14 �C to 18 �C) (Mobergand Bergström, 1997). Central Sweden is dominated by boreal for-est, comprised of spruce and pine, as well as birch and conifers(Arnborg, 1990) and hosts a diverse range of potential prey speciessuch as Eurasian elk (Alces alces), roe deer (Capreolus capreolus), reddeer (Cervus elaphus), reindeer, wild boar (Sus scrofa), brown hare(Lepus europaeus), mountain hare (Lepus timidus), rabbit (Orycto-lagus cuniculus), Eurasian beaver (Castor fiber) and Norwaylemming (Lemmus lemmus). As well as wolf, brown bear and lynx(Lynx lynx) are present in the large carnivore fauna.
4. Canid diet
4.1. Palaeodiet and implications for dental morphology
Both diet and body size dictate the ecological niche of a species,with factors such as prey availability, competition, environmentalconditions and climate serving as important variables in deter-mining niche attribution. Dietary differences, such as the propor-tion of flesh to non-flesh foods, can indicate the level of carnivory ofindividual species and allow classification into hypo-, meso- andhypercarnivores in ascending degree of meat consumption. Bothhypo- and mesocarnivores are relatively unspecialised, consumingsmall prey only and taking an increased amount of plant matter inthe diet (particularly hypocarnivores). The morphology of thedentition in both is characterised by reduced carnassial blades inthe m1 and P4, relative to enlarged molars (Van Valkenburgh et al.,2004). Hypercarnivores are considerably more specialised in theircranio-dental morphology, with relatively deep jaws, large canines
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e16 5
and incisors, reduced molar grinding areas and longer carnassialblades (Van Valkenburgh and Koepfli, 1993). Their diets are pre-dominantly flesh-based, although some level of bone consumptionis also common (Van Valkenburgh, 1991). Modern C. lupus is ahypercarnivore, yet in contrast to other hypercarnivorous canidssuch as the African wild dog Lycaon pictus Temminck, 1820, thelower carnassial of C. lupus lacks the full adaption to slicing,denoted by an elongation of the trigonid through the loss of abicuspid talonid ‘trenchant heel’. Rather, C. lupus has retained thesub-equal bicuspid talonid morphology, keeping it as a dual pur-pose tooth (Van Valkenburgh, 1991). Combined with the retentionof post-carnassial molars (a further contrast with wild dogs thathave often lost the m2-m3, and especially with other hyper-carnivores such as the Felidae), C. lupus is able to both crush foodsand slice flesh.
In order to make palaeodietary inferences and hence establishthe level of carnivory in Pleistocene canids, the relationship be-tween dental morphology and function needs to be examined. Ingeneral, canids share the dental formula I3/3, C1/1, P4/4, M2/3and are broadly similar in terms of overall morphology. By usingthe dental apparatus of modern C. lupus as an analogue forfunction in the Pleistocene canids, comparisons can be made onthe potential variability in diet. The cranio-dental morphology ofmodern C. lupus reflects its hypercarnivorous diet, as well as itsadaptability. The pointed canines and incisors have the dualfunction of holding prey and tearing flesh (Ewer, 1973; VanValkenburg, 2007), whilst the sectorial premolars act to bothpierce and hold prey (Van Valkenburg, 2007). The size andmorphology of the largest lower premolar (p4), the carnassials(m1, P4) and the post-carnassial molars (m2-3, M1-2) areimportant for reflecting dietary specialisation and form the focusof the analyses here. In some carnivores, changes in p4 shape havebeen related to bone-eating behaviour, based on the premise thatbone-eating specialists, such as spotted hyaenas, have enlargedand very robust premolars with a broad-based pyramidal shape,whereas flesh-eating specialists such as felids tend to havesmaller, narrower premolars (Van Valkenburgh, 1988a, 1989,1991; Werdelin, 1989). Thus for hyaenas, the specific premolaradaptation for bone-cracking enables marrow extraction, whereasin modern canids, access to marrow is gained through bone-crushing, utilising the post carnassial teeth (Werdelin, 1989).Nonetheless, despite their bone-cracking adaptations, hyaenasprocess certain bones using the wider dental complex, forexample incisors or carnassials are used to break ribs and scap-ulae (Van Valkenburgh, 1996). Wild dogs also use carnassials aswell as their usual crushing apparatus of the post-carnassialmolars to crush bone (Van Valkenburgh, 1996). It therefore ap-pears that in order to access marrow, more than one dental regioncan be used in its acquisition, depending on the type of boneexploited. The p4, with its overlapping position below the ante-rior upper carnassial and its situation in the anterior region ofmaximum bite force, may by its proximity also become involvedin bone utilisation, together with the molar crushing apparatus.However, regular involvement of teeth other than molars in thisprocess is more likely to stem from dietary stress, wherebyincreased access to marrow is necessary.
In extant canids, both hypercarnivorous and more omnivorousmembers were found to share relatively slender premolars ofsimilar size and shape, based on morphometric ratios of mea-surements by Van Valkenburgh (1991). However, notwithstandingcontroversies relating to the use of ratios in taxonomical andecological analysis (see 5.1), the same premise would also suggestthat any increase in bone-eating should be reflected by changes inthe length andwidth of p4, analysed as linear measurements ratherthan ratios.
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
The carnassial teeth (P4, m1) are specifically adapted to worktogether to cut flesh, with the paracone and metacone cusps of theP4 and the trigonid cusps of them1 all laterally flattened to enable ashearing action (Ewer, 1973). Thus, morphology of the carnassialpair is particularly informative regarding the relative proportions offlesh consumed. However, unlike the P4, the m1 is (as stated pre-viously) of dual function, incorporating both slicing (the anteriortrigonid ‘blade’) and crushing (the posterior talonid basin ‘heel’)actions (Van Valkenburg, 2007). It is therefore anticipated thatchanges in the length of the m1 trigonid or the length of the P4 willdirectly relate to the amount of flesh in the diet. Typically, flesh andbone specialists tend to have longer cutting blades in comparison tomore omnivorous species (Van Valkenburgh, 1988a; VanValkenburgh and Koepfli, 1993). A further competitive advantageof lengthening the trigonid is an increased ability to slice fleshquickly (Van Valkenburgh, 1991). Changes in carnassial width mayrelate either to strengthening, in order to reinforce this key pair ofteeth, or to facilitate increased slicing ability.
Along with the m1 talonid and m2, the upper molars have acrushing function in C. lupus (Ewer, 1973) related to the con-sumption of non-flesh foods. In hypercarnivorous species such asfelids, post-carnassial molars are very reduced, whereas theopposite is true for more omnivorous species (Van Valkenburgh,1991). The size and morphology of the molars may thereforereflect the proportion of non-meat foods in the diet. Finally,hypercarnivores such as C. lupus tend to have relatively deep jawsto cope with increased loading from killing and feeding on largeprey (Van Valkenburgh and Koepfli, 1993; Van Valkenburgh et al.,2004). Hence, study of both depth and breadth of the dentary be-tween the p3-p4 and m1-m2 can potentially reveal the size of preyable to be consumed.
4.2. Diet in modern C. lupus
The diet of modern C. lupus has been extensively documented,allowing inferences to bemade concerning potential prey choice byPleistocene C. lupus. Based on case studies from North America(Voigt et al., 1976; Fritts and Mech, 1981; Paquet, 1992; Boyd et al.,1994) and Europe (Jedrzejewski et al., 2000; Capitani et al., 2003;Kojola et al., 2004; Ansorge et al., 2006; Nowak et al., 2011), wildungulates form the main prey. In North America, white-tailed deer(Odocoileus virginianus), elk (Cervus canadensis) and moose (A.alces) are favoured, whereas red deer (C. elaphus), roe deer (C.capreolus) and wild boar (S. scrofa) are targeted in Europe, alongwith reindeer at high latitudes.
Where availability of wild ungulates is low, either permanentlyor seasonally, C. lupus is able to adapt by hunting smaller verte-brates, such as North American beaver (Castor canadensis) (Voigtet al., 1976), and in Europe, Eurasian beaver, brown hare andbirds (Jedrzejewski et al., 2000). In modern wolves, scarcity of wildungulates can also increases predation on livestock (Meriggi andLovari, 1996; Vos, 2000), as well as in certain circumstanceshousehold waste (Pullianen, 1975; Rogers and Mech, 1981).
Today, C. lupus lives and hunts in packs consisting of familymembers (Mech, 1970; Ewer, 1973), with cooperative huntingbehaviour allowing wolves to capture and kill prey much largerthan themselves (Macdonald, 1983). Typical hunting of large preyinvolves (frequently extended) pursuit and subsequent attack bytearing flesh from the hindquarters and shoulders (Mech, 1970;Ewer, 1973). Feeding commences immediately after killing prey(Mech, 1970). Areas of initial attack are often concentrated onduring feeding, with flesh on the flanks consumed quickly andrapid access to organs and viscera (Mech, 1970). Carcasses are oftenrevisited, and complete consumption of prey generally occurs(Mech, 1970; Fritts and Mech, 1981). Variables, such as snow depth
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

Table 1List of the dietary measurements used in the analyses, including measurementabbreviation and description.
Measurement Description
p4L Maximum antero-posterior length of lower fourthpremolara
p4W Maximum medio-lateral breadth of lower fourth premolara
m1L Maximum antero-posterior length of the lower carnassiala
m1Ltrig Maximum antero-posterior length of the lower carnassialtrigonid (paraconid and protoconid)c
m1Ltal Maximum antero-posterior length of the lower carnassialtalonid basin
m1W Maximum medio-lateral breadth lower carnassiala
m2L Maximum antero-posterior length of second lower molara
m2W Maximum medio-lateral breadth of second lower molara
p3p4D Dentary depth at the p3-p4 junction of the mandibleb
p3p4B Dentary breadth at the p3-p4 junction of the mandibleb
m1m2D Dentary depth at the m1-m2 junction of the mandiblea
m1m2B Dentary breadth at the m1-m2 junction of the mandibleb
P4L Maximum antero-posterior length of upper carnassiala
P4W Maximum medio-lateral breadth of upper carnassial,including the protoconea
M1L Maximum antero-posterior buccal length of first uppermolara
M1W Maximum antero-posterior width of first upper molara
M1M2L Length of the upper molar row on the buccal sidea
a von den Driesch (1976).b Van Valkenburgh and Koepfli (1993).c Van Valkenburgh et al. (2004).
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e166
and ease of predation can affect consumption, for example fullcarcass utilisation occurs in shallow snow depths (see Boyd et al.,1994), as opposed to light carcass utilisation during deep snowdue to relative ease of predation (Mech et al., 1971).
4.3. Tooth wear and breakage
Tooth wear has previously been used to provide broad estimatesof individual age (Van Ballenberghe et al., 1975; Fritts and Mech,1981; Landon et al., 1998), on the basis that older individualshave increased incidences of highly worn teeth (Van Valkenburgh,1988b). The degree of tooth breakage is also positively correlatedwith increased individual age (Van Valkenburgh, 1988b). However,tooth wear and breakage can equally be strongly influenced by dietand correlate with the amount of tough foodstuffs in the diet(Binder et al., 2002). In a comparison of tooth wear frequencies inmodern large carnivores, Van Valkenburgh (1988b) found thattooth wear in C. lupus occurredmore rapidly, due to the inclusion ofrelativelymore bone in their diet, in comparison to L. pictus. C. lupustherefore exhibited increased rates of tooth wear related to diet,rather than increased individual age, in the sample population.Similarly, frequencies of tooth breakage in Canis dirus Leidy, 1858,from the Late Pleistocene site of Rancho La Brea (California, USA),were found to reflect variation in feeding behaviour rather thansimply differences in the age structure of the sampled populations(Binder and Van Valkenburgh, 2010).
5. Materials and methods
Material was measured from the following institutions: Britishmaterial: Bristol City Museum; British Geological Survey, Key-worth; Buckinghamshire County Museum; Manchester Museum;National Museum of Wales, Cardiff; Natural History Museum,London; The Sedgwick Museum of Earth Sciences, Cambridge;Somerset Heritage Centre, Taunton; Swansea Museum; TorquayMuseum; University Museum of Zoology Cambridge; WoollatonHall, Nottingham. German material: Forschungsstation für Quar-tärpaläontologie, Weimar. Italian material: Museo di Storia Natu-rale degli Studi di Firenze. Modern Swedish material:Naturhistoriska riksmuseet, Stockholm.
143 individuals representing three canid species (C. lupus,C. mosbachensis and C. etruscus) were examined. For the modernC. lupus group, males and females were pooled, since for thePleistocene groups, sex determination was not always possible.Individuals rather than all isolated teeth were analysed to ensurethat apparent differences in diet were as close a reflection of thepopulation as possible.
5.1. Measurements
In order to infer palaeodiet, specific dental characters wereanalysed based on their known dietary-related function (VanValkenburgh, 1988a; Van Valkenburgh, 1991; Van Valkenburghand Koepfli, 1993). Available material was measured using bothdigital and metric callipers. A list of the dietary measurements isprovided in Table 1, and shown in Fig. 3. Linear measurements weredirectly analysed rather than morphometric ratios. Although oftenused to reflect aspects of diet (Van Valkenburgh, 1988a, 1989, 1991;Van Valkenburgh and Koepfli, 1993; Van Valkenburgh et al., 2004),as well as in post-cranial material (Samuels et al., 2012; Meloroet al., 2013), the appropriateness of using ratios has been debated(e.g. Corrunccini, 1975; Atchley et al., 1976; Atchley, 1978) based ontheir failure to remove the effect of size and resultant masking ef-fects in data analyses (Albrecht, 1978). Due to these potentialproblems, direct analysis of linear measurements was chosen.
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
5.2. Tooth breakage and wear
All teeth were visually assessed for level of wear and assigned awear category as follows: Slight: no apparent wear with no bluntingof cusps; Moderate: some visible wear and blunting of cusps pre-sent, and Heavy: severewear with strongly blunted cusps (based onVan Valkenburgh, 1988b; Van Valkenburgh and Hertel, 1993).Following Van Valkenburgh (1988b) and Van Valkenburgh andHertel (1993), a tooth was counted as Broken in the analysis if itexhibited post-breakage wear from occlusal tooth contact, or con-tact with food during the animal’s lifetime.
5.3. Body size
Lower carnassial length has been used extensively in thereconstruction of carnivore body mass (e.g. Legendre and Roth,1988; Van Valkenburgh, 1990) due to its low variability, and toothsize being fixed upon eruption (Gingerich, 1974). Since the m1 iswell developed in canids as part of the functionally importantcarnassial pair, its length is considered to scale in a predictable waywith body size (Van Valkenburgh, 1990), thereby allowing basiccomparisons to be made between individuals, populations andspecies. Body size comparisons will be used here to infer relativesize differences and elucidate the relationship this may have withdiet, specifically variation in carnivory both temporally and be-tween species.
5.4. Statistical analysis
A univariate and multivariate statistical approach was used. Thedietary measurements were tested for normality using ShapiroeWilk tests, and summary statistics calculated. The relationshipbetween m1L and m1W for body mass inferences of each specieswas established using linear regression, and the significance ofslopes tested using ANCOVA. For the temporal analysis of C. lupus,the dietary measurements were analysed for variance using one-way ANOVA, with Levene’s Test employed to examine equality ofvariance, to investigate the presence of temporal differences be-tween C. lupus, C. mosbachensis and C. etruscus. Subsequent post hoc
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

M1WP4W
C.
m2W
p4Wm1W
m1Ltal
D.p3p4B
m1m2B
m2L
m1m2D
p4L
m1L
m1Ltrig
B.
p3p4D
P4L
M1-M2L
M1L
A.
Fig. 3. Cranio-dental measurements used in the analyses illustrated on the skull andmandible of C. lupus. A: Lateral view of skull, B: Lateral view of mandible, C. Ventralview of skull focussing on palate and dentition, D. Dorsal view of right mandiblefocussing on dentition. Measurement abbreviations in Table 1.
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e16 7
tests were then used for multiple comparisons if ANOVA was sig-nificant, based on Tukey HSD for those with equal variances orDunnett’s T3 for unequal variances. Tooth wear in the PleistoceneC. lupus age groups was analysed using 2-way Pearson Chi-squaretests (using 2 � 3 contingency tables) to investigate differences inthe frequency of wear within the age groups.
A cross validated stepwise Discriminant Function Analysis wasperformed on the dietary measurements shown in Table 2 toinvestigate how well the stepwise selected dietary measurementscould differentiate between both the chronological groupings ofC. lupus and the different species groups, thus indicating the dietarydifferences between both groupings. All statistical analysis wasperformed using SPSS (v. 19).
6. Results
All measurements were found to be normally distributed usingShapiroeWilk tests for normality (p > 0.05), and further analysiswas therefore conducted using parametric methods. Summary
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
statistics of the dietary measurements used in the analysis areshown in Table 2.
6.1. Body size measurements
A positive relationship between m1L and m1Wwas highlighted(Fig. 4), with the regression slopes found to be similar betweeneach species (ANCOVA F (2, 118) ¼ 0.841, p ¼ 0.434) and thus indi-cating similar rates of change in m1W and m1L in all three species.Using the lower carnassial length as a proxy for body size, it can beseen that C. lupus is relatively larger than both C. etruscus andC. mosbachensis, with the former slightly larger than the latter,albeit within the range of variation (Fig. 5). With respect to thethree chronological groupings of C. lupus, the body size indexhighlighted variable body size between groups (Fig. 5), with MIS 5acontaining the largest individuals, followed by modern C. lupus(represented by MIS 1). MIS 3 and 7 groups are both relativelysmaller than members of the other age groups, although all arewithin the high amount of size variation for each group.
6.2. Temporal variation in palaeodiet measurements betweengroups
All analysed measurements between age groups MIS 3, 5a and 7were found to be of equal variance by Levene’s tests (p > 0.05).Significant differences were found by one-way ANOVA in these agegroups in p4L, p4W, m1Ltrig, m1W, p3p4B, m1m2B (p < 0.05) andslightly significant in m1m2D (p ¼ 0.054). Post hoc tests (Table 3)revealed significant differences between MIS 3 and 5a for p4L,m1Ltrig and m1W (Tukey HSD, p < 0.05), and between MIS 5a and7 for p4W, m1Ltrig, m1W, p3p4B, m1m2B andm1m2D (Tukey HSD,p < 0.05). No significant differences were found by Tukey HSDbetween MIS 3 and 7 (p > 0.05). Therefore temporal differencesexist in the dietary measurements in C. lupus between MIS 5a andboth MIS 3 and 7.
The presence of these temporal differences in PleistoceneC. lupus raises the question of whether these differences are a ‘one-off’ or whether they are a continuing characteristic that can befound in modern C. lupus. As shown in Fig. 5, modern C. lupus (MIS1) is of broadly similar body size, based on mean lower carnassiallength, albeit more similar to the slightly larger MIS 5a individualsthan to MIS 3 or 7 specimens. Nonetheless, one-way ANOVA foundsignificant differences between the modern and Pleistocene wolfage groups in p4L, p4W, m1Ltrig, m1Ltal, m1W, m2L, m2W, p3p4B,m1m2D, m1m2B, P4W, M1L, M1W and M1M2L (p < 0.05). FurtherTukey HSD tests (Table 3) indicated significant differences betweenMIS 1 and 3 in p4L, m1Ltal, m1m2D and M1L (p < 0.05), and be-tweenMIS 1 and 5a in p4L, p4W,m1Ltrig, m1W,m1m2D, P4W,M1Land M1W (p < 0.05), as well as with MIS 1 and 7 in m1W, m2L,p3p4B, m1m2D, m1m2B, P4W, M1L and M1W (p < 0.05). Levene’stest found m2W, p3p4D and M1M2L as having unequal variance(p < 0.05) and thus Dunnett’s T3 test (Table 3) indicated m2W assignificantly different between MIS 1 and 3 only (p < 0.05), p3p4Das significantly different between MIS 1 and 7 only (p < 0.05), aswell as finding no significant differences in M1M2L between all agegroups (p > 0.05), in contrast to the one-way ANOVA result for thismeasurement. Hence, temporal differences exist between modernand Pleistocene C. lupus age groups.
The presence of such temporal differences in C. lupus suggeststhat similar differences might equally be detected in the dietarymeasurements in earlier Pleistocene canids, such as C. mosbachensisand C. etruscus. For C. mosbachensis, individuals from sites of earlyMiddle Pleistocene age in Britain were amalgamated to provide alarger group for analysis, and compared with the Late Early Pleis-tocene site of Untermassfeld. All dietary measurements between
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015
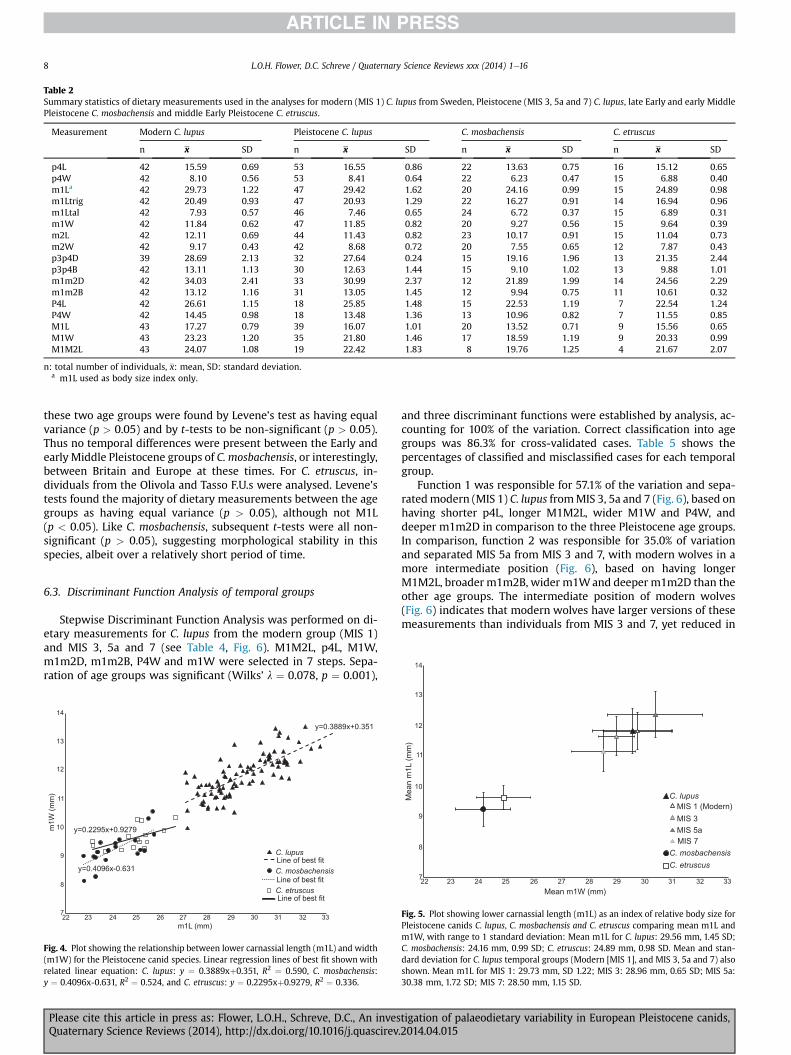
Table 2Summary statistics of dietary measurements used in the analyses for modern (MIS 1) C. lupus from Sweden, Pleistocene (MIS 3, 5a and 7) C. lupus, late Early and early MiddlePleistocene C. mosbachensis and middle Early Pleistocene C. etruscus.
Measurement Modern C. lupus Pleistocene C. lupus C. mosbachensis C. etruscus
n x SD n x SD n x SD n x SD
p4L 42 15.59 0.69 53 16.55 0.86 22 13.63 0.75 16 15.12 0.65p4W 42 8.10 0.56 53 8.41 0.64 22 6.23 0.47 15 6.88 0.40m1La 42 29.73 1.22 47 29.42 1.62 20 24.16 0.99 15 24.89 0.98m1Ltrig 42 20.49 0.93 47 20.93 1.29 22 16.27 0.91 14 16.94 0.96m1Ltal 42 7.93 0.57 46 7.46 0.65 24 6.72 0.37 15 6.89 0.31m1W 42 11.84 0.62 47 11.85 0.82 20 9.27 0.56 15 9.64 0.39m2L 42 12.11 0.69 44 11.43 0.82 23 10.17 0.91 15 11.04 0.73m2W 42 9.17 0.43 42 8.68 0.72 20 7.55 0.65 12 7.87 0.43p3p4D 39 28.69 2.13 32 27.64 0.24 15 19.16 1.96 13 21.35 2.44p3p4B 42 13.11 1.13 30 12.63 1.44 15 9.10 1.02 13 9.88 1.01m1m2D 42 34.03 2.41 33 30.99 2.37 12 21.89 1.99 14 24.56 2.29m1m2B 42 13.12 1.16 31 13.05 1.45 12 9.94 0.75 11 10.61 0.32P4L 42 26.61 1.15 18 25.85 1.48 15 22.53 1.19 7 22.54 1.24P4W 42 14.45 0.98 18 13.48 1.36 13 10.96 0.82 7 11.55 0.85M1L 43 17.27 0.79 39 16.07 1.01 20 13.52 0.71 9 15.56 0.65M1W 43 23.23 1.20 35 21.80 1.46 17 18.59 1.19 9 20.33 0.99M1M2L 43 24.07 1.08 19 22.42 1.83 8 19.76 1.25 4 21.67 2.07
n: total number of individuals, x: mean, SD: standard deviation.a m1L used as body size index only.
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e168
these two age groups were found by Levene’s test as having equalvariance (p > 0.05) and by t-tests to be non-significant (p > 0.05).Thus no temporal differences were present between the Early andearly Middle Pleistocene groups of C. mosbachensis, or interestingly,between Britain and Europe at these times. For C. etruscus, in-dividuals from the Olivola and Tasso F.U.s were analysed. Levene’stests found the majority of dietary measurements between the agegroups as having equal variance (p > 0.05), although not M1L(p < 0.05). Like C. mosbachensis, subsequent t-tests were all non-significant (p > 0.05), suggesting morphological stability in thisspecies, albeit over a relatively short period of time.
14
6.3. Discriminant Function Analysis of temporal groups
Stepwise Discriminant Function Analysis was performed on di-etary measurements for C. lupus from the modern group (MIS 1)and MIS 3, 5a and 7 (see Table 4, Fig. 6). M1M2L, p4L, M1W,m1m2D, m1m2B, P4W and m1W were selected in 7 steps. Sepa-ration of age groups was significant (Wilks’ l ¼ 0.078, p ¼ 0.001),
m1L (mm)
m1W
(mm
)
14
13
12
11
10
9
8
722 23 24 25 26 27 28 29 30 31 32 33
C. lupus Line of best fitC. mosbachensisLine of best fitC. etruscusLine of best fit
y=0.3889x+0.351
y=0.4096x-0.631
y=0.2295x+0.9279
Fig. 4. Plot showing the relationship between lower carnassial length (m1L) and width(m1W) for the Pleistocene canid species. Linear regression lines of best fit shown withrelated linear equation: C. lupus: y ¼ 0.3889xþ0.351, R2 ¼ 0.590, C. mosbachensis:y ¼ 0.4096x-0.631, R2 ¼ 0.524, and C. etruscus: y ¼ 0.2295xþ0.9279, R2 ¼ 0.336.
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
and three discriminant functions were established by analysis, ac-counting for 100% of the variation. Correct classification into agegroups was 86.3% for cross-validated cases. Table 5 shows thepercentages of classified and misclassified cases for each temporalgroup.
Function 1 was responsible for 57.1% of the variation and sepa-ratedmodern (MIS 1) C. lupus fromMIS 3, 5a and 7 (Fig. 6), based onhaving shorter p4L, longer M1M2L, wider M1W and P4W, anddeeper m1m2D in comparison to the three Pleistocene age groups.In comparison, function 2 was responsible for 35.0% of variationand separated MIS 5a from MIS 3 and 7, with modern wolves in amore intermediate position (Fig. 6), based on having longerM1M2L, broaderm1m2B, wider m1Wand deeper m1m2D than theother age groups. The intermediate position of modern wolves(Fig. 6) indicates that modern wolves have larger versions of thesemeasurements than individuals from MIS 3 and 7, yet reduced in
Mea
n m
1L (m
m)
Mean m1W (mm)
9
10
11
12
13
7
8
22 2726252423 333231302928
C. lupus
C. etruscusC. mosbachensis
MIS 1 (Modern)MIS 3
MIS 7MIS 5a
Fig. 5. Plot showing lower carnassial length (m1L) as an index of relative body size forPleistocene canids C. lupus, C. mosbachensis and C. etruscus comparing mean m1L andm1W, with range to 1 standard deviation: Mean m1L for C. lupus: 29.56 mm, 1.45 SD;C. mosbachensis: 24.16 mm, 0.99 SD; C. etruscus: 24.89 mm, 0.98 SD. Mean and stan-dard deviation for C. lupus temporal groups (Modern [MIS 1], and MIS 3, 5a and 7) alsoshown. Mean m1L for MIS 1: 29.73 mm, SD 1.22; MIS 3: 28.96 mm, 0.65 SD; MIS 5a:30.38 mm, 1.72 SD; MIS 7: 28.50 mm, 1.15 SD.
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015
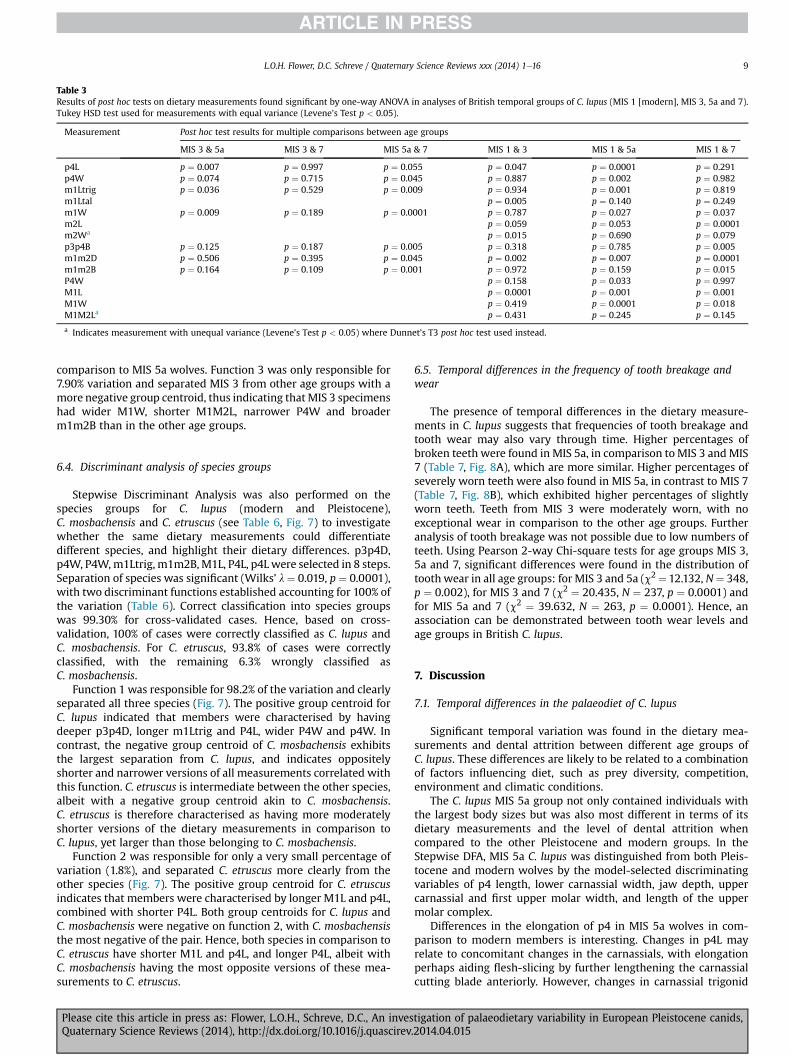
Table 3Results of post hoc tests on dietary measurements found significant by one-way ANOVA in analyses of British temporal groups of C. lupus (MIS 1 [modern], MIS 3, 5a and 7).Tukey HSD test used for measurements with equal variance (Levene’s Test p < 0.05).
Measurement Post hoc test results for multiple comparisons between age groups
MIS 3 & 5a MIS 3 & 7 MIS 5a & 7 MIS 1 & 3 MIS 1 & 5a MIS 1 & 7
p4L p ¼ 0.007 p ¼ 0.997 p ¼ 0.055 p ¼ 0.047 p ¼ 0.0001 p ¼ 0.291p4W p ¼ 0.074 p ¼ 0.715 p ¼ 0.045 p ¼ 0.887 p ¼ 0.002 p ¼ 0.982m1Ltrig p ¼ 0.036 p ¼ 0.529 p ¼ 0.009 p ¼ 0.934 p ¼ 0.001 p ¼ 0.819m1Ltal p ¼ 0.005 p ¼ 0.140 p ¼ 0.249m1W p ¼ 0.009 p ¼ 0.189 p ¼ 0.0001 p ¼ 0.787 p ¼ 0.027 p ¼ 0.037m2L p ¼ 0.059 p ¼ 0.053 p ¼ 0.0001m2Wa p ¼ 0.015 p ¼ 0.690 p ¼ 0.079p3p4B p ¼ 0.125 p ¼ 0.187 p ¼ 0.005 p ¼ 0.318 p ¼ 0.785 p ¼ 0.005m1m2D p ¼ 0.506 p ¼ 0.395 p ¼ 0.045 p ¼ 0.002 p ¼ 0.007 p ¼ 0.0001m1m2B p ¼ 0.164 p ¼ 0.109 p ¼ 0.001 p ¼ 0.972 p ¼ 0.159 p ¼ 0.015P4W p ¼ 0.158 p ¼ 0.033 p ¼ 0.997M1L p ¼ 0.0001 p ¼ 0.001 p ¼ 0.001M1W p ¼ 0.419 p ¼ 0.0001 p ¼ 0.018M1M2La p ¼ 0.431 p ¼ 0.245 p ¼ 0.145
a Indicates measurement with unequal variance (Levene’s Test p < 0.05) where Dunnet’s T3 post hoc test used instead.
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e16 9
comparison to MIS 5a wolves. Function 3 was only responsible for7.90% variation and separated MIS 3 from other age groups with amore negative group centroid, thus indicating thatMIS 3 specimenshad wider M1W, shorter M1M2L, narrower P4W and broaderm1m2B than in the other age groups.
6.4. Discriminant analysis of species groups
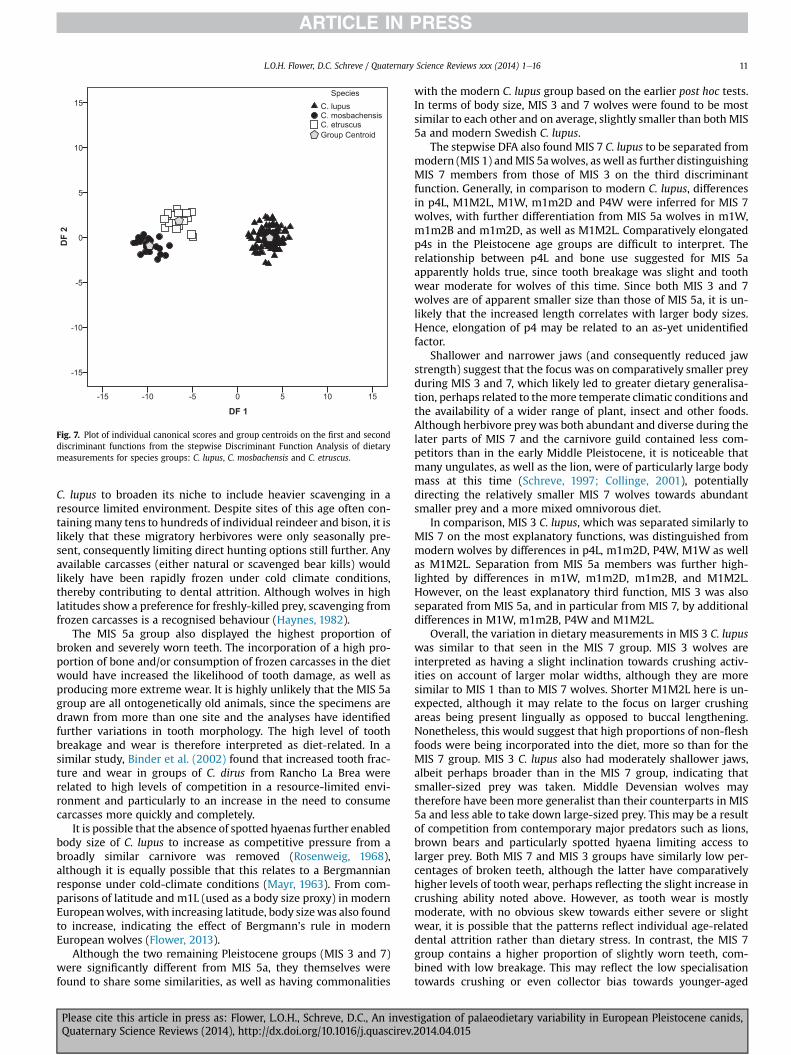
Stepwise Discriminant Analysis was also performed on thespecies groups for C. lupus (modern and Pleistocene),C. mosbachensis and C. etruscus (see Table 6, Fig. 7) to investigatewhether the same dietary measurements could differentiatedifferent species, and highlight their dietary differences. p3p4D,p4W, P4W,m1Ltrig, m1m2B,M1L, P4L, p4Lwere selected in 8 steps.Separation of species was significant (Wilks’ l ¼ 0.019, p ¼ 0.0001),with two discriminant functions established accounting for 100% ofthe variation (Table 6). Correct classification into species groupswas 99.30% for cross-validated cases. Hence, based on cross-validation, 100% of cases were correctly classified as C. lupus andC. mosbachensis. For C. etruscus, 93.8% of cases were correctlyclassified, with the remaining 6.3% wrongly classified asC. mosbachensis.
Function 1 was responsible for 98.2% of the variation and clearlyseparated all three species (Fig. 7). The positive group centroid forC. lupus indicated that members were characterised by havingdeeper p3p4D, longer m1Ltrig and P4L, wider P4W and p4W. Incontrast, the negative group centroid of C. mosbachensis exhibitsthe largest separation from C. lupus, and indicates oppositelyshorter and narrower versions of all measurements correlated withthis function. C. etruscus is intermediate between the other species,albeit with a negative group centroid akin to C. mosbachensis.C. etruscus is therefore characterised as having more moderatelyshorter versions of the dietary measurements in comparison toC. lupus, yet larger than those belonging to C. mosbachensis.
Function 2 was responsible for only a very small percentage ofvariation (1.8%), and separated C. etruscus more clearly from theother species (Fig. 7). The positive group centroid for C. etruscusindicates that members were characterised by longer M1L and p4L,combined with shorter P4L. Both group centroids for C. lupus andC. mosbachensis were negative on function 2, with C. mosbachensisthe most negative of the pair. Hence, both species in comparison toC. etruscus have shorter M1L and p4L, and longer P4L, albeit withC. mosbachensis having the most opposite versions of these mea-surements to C. etruscus.
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
6.5. Temporal differences in the frequency of tooth breakage andwear
The presence of temporal differences in the dietary measure-ments in C. lupus suggests that frequencies of tooth breakage andtooth wear may also vary through time. Higher percentages ofbroken teeth were found in MIS 5a, in comparison to MIS 3 andMIS7 (Table 7, Fig. 8A), which are more similar. Higher percentages ofseverely worn teeth were also found in MIS 5a, in contrast to MIS 7(Table 7, Fig. 8B), which exhibited higher percentages of slightlyworn teeth. Teeth from MIS 3 were moderately worn, with noexceptional wear in comparison to the other age groups. Furtheranalysis of tooth breakage was not possible due to low numbers ofteeth. Using Pearson 2-way Chi-square tests for age groups MIS 3,5a and 7, significant differences were found in the distribution oftooth wear in all age groups: for MIS 3 and 5a (c2 ¼ 12.132, N¼ 348,p ¼ 0.002), for MIS 3 and 7 (c2 ¼ 20.435, N ¼ 237, p ¼ 0.0001) andfor MIS 5a and 7 (c2 ¼ 39.632, N ¼ 263, p ¼ 0.0001). Hence, anassociation can be demonstrated between tooth wear levels andage groups in British C. lupus.
7. Discussion
7.1. Temporal differences in the palaeodiet of C. lupus
Significant temporal variation was found in the dietary mea-surements and dental attrition between different age groups ofC. lupus. These differences are likely to be related to a combinationof factors influencing diet, such as prey diversity, competition,environment and climatic conditions.
The C. lupus MIS 5a group not only contained individuals withthe largest body sizes but was also most different in terms of itsdietary measurements and the level of dental attrition whencompared to the other Pleistocene and modern groups. In theStepwise DFA, MIS 5a C. lupus was distinguished from both Pleis-tocene and modern wolves by the model-selected discriminatingvariables of p4 length, lower carnassial width, jaw depth, uppercarnassial and first upper molar width, and length of the uppermolar complex.
Differences in the elongation of p4 in MIS 5a wolves in com-parison to modern members is interesting. Changes in p4L mayrelate to concomitant changes in the carnassials, with elongationperhaps aiding flesh-slicing by further lengthening the carnassialcutting blade anteriorly. However, changes in carnassial trigonid
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015
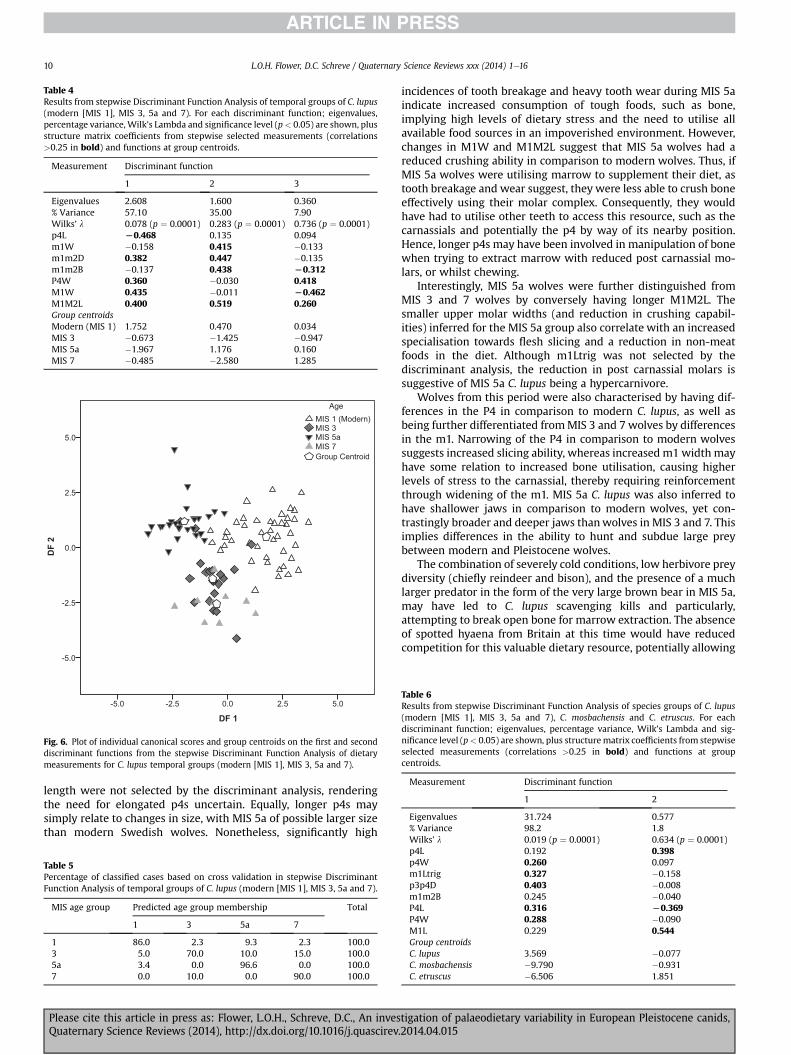
Table 4Results from stepwise Discriminant Function Analysis of temporal groups of C. lupus(modern [MIS 1], MIS 3, 5a and 7). For each discriminant function; eigenvalues,percentage variance, Wilk’s Lambda and significance level (p< 0.05) are shown, plusstructure matrix coefficients from stepwise selected measurements (correlations>0.25 in bold) and functions at group centroids.
Measurement Discriminant function
1 2 3
Eigenvalues 2.608 1.600 0.360% Variance 57.10 35.00 7.90Wilks’ l 0.078 (p ¼ 0.0001) 0.283 (p ¼ 0.0001) 0.736 (p ¼ 0.0001)p4L L0.468 0.135 0.094m1W �0.158 0.415 �0.133m1m2D 0.382 0.447 �0.135m1m2B �0.137 0.438 L0.312P4W 0.360 �0.030 0.418M1W 0.435 �0.011 L0.462M1M2L 0.400 0.519 0.260Group centroidsModern (MIS 1) 1.752 0.470 0.034MIS 3 �0.673 �1.425 �0.947MIS 5a �1.967 1.176 0.160MIS 7 �0.485 �2.580 1.285
DF 15.02.50.0-2.5-5.0
DF
2
5.0
2.5
0.0
-2.5
-5.0
Group CentroidMIS 7MIS 5aMIS 3MIS 1 (Modern)
Age
Fig. 6. Plot of individual canonical scores and group centroids on the first and seconddiscriminant functions from the stepwise Discriminant Function Analysis of dietarymeasurements for C. lupus temporal groups (modern [MIS 1], MIS 3, 5a and 7).
Table 6Results from stepwise Discriminant Function Analysis of species groups of C. lupus(modern [MIS 1], MIS 3, 5a and 7), C. mosbachensis and C. etruscus. For eachdiscriminant function; eigenvalues, percentage variance, Wilk’s Lambda and sig-nificance level (p< 0.05) are shown, plus structurematrix coefficients from stepwiseselected measurements (correlations >0.25 in bold) and functions at groupcentroids.
Measurement Discriminant function
1 2
Eigenvalues 31.724 0.577% Variance 98.2 1.8Wilks’ l 0.019 (p ¼ 0.0001) 0.634 (p ¼ 0.0001)
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e1610
length were not selected by the discriminant analysis, renderingthe need for elongated p4s uncertain. Equally, longer p4s maysimply relate to changes in size, with MIS 5a of possible larger sizethan modern Swedish wolves. Nonetheless, significantly high
Table 5Percentage of classified cases based on cross validation in stepwise DiscriminantFunction Analysis of temporal groups of C. lupus (modern [MIS 1], MIS 3, 5a and 7).
MIS age group Predicted age group membership Total
1 3 5a 7
1 86.0 2.3 9.3 2.3 100.03 5.0 70.0 10.0 15.0 100.05a 3.4 0.0 96.6 0.0 100.07 0.0 10.0 0.0 90.0 100.0
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
incidences of tooth breakage and heavy tooth wear during MIS 5aindicate increased consumption of tough foods, such as bone,implying high levels of dietary stress and the need to utilise allavailable food sources in an impoverished environment. However,changes in M1W and M1M2L suggest that MIS 5a wolves had areduced crushing ability in comparison to modern wolves. Thus, ifMIS 5a wolves were utilising marrow to supplement their diet, astooth breakage and wear suggest, they were less able to crush boneeffectively using their molar complex. Consequently, they wouldhave had to utilise other teeth to access this resource, such as thecarnassials and potentially the p4 by way of its nearby position.Hence, longer p4s may have been involved in manipulation of bonewhen trying to extract marrow with reduced post carnassial mo-lars, or whilst chewing.
Interestingly, MIS 5a wolves were further distinguished fromMIS 3 and 7 wolves by conversely having longer M1M2L. Thesmaller upper molar widths (and reduction in crushing capabil-ities) inferred for the MIS 5a group also correlate with an increasedspecialisation towards flesh slicing and a reduction in non-meatfoods in the diet. Although m1Ltrig was not selected by thediscriminant analysis, the reduction in post carnassial molars issuggestive of MIS 5a C. lupus being a hypercarnivore.
Wolves from this period were also characterised by having dif-ferences in the P4 in comparison to modern C. lupus, as well asbeing further differentiated fromMIS 3 and 7 wolves by differencesin the m1. Narrowing of the P4 in comparison to modern wolvessuggests increased slicing ability, whereas increased m1 width mayhave some relation to increased bone utilisation, causing higherlevels of stress to the carnassial, thereby requiring reinforcementthrough widening of the m1. MIS 5a C. lupus was also inferred tohave shallower jaws in comparison to modern wolves, yet con-trastingly broader and deeper jaws thanwolves in MIS 3 and 7. Thisimplies differences in the ability to hunt and subdue large preybetween modern and Pleistocene wolves.
The combination of severely cold conditions, low herbivore preydiversity (chiefly reindeer and bison), and the presence of a muchlarger predator in the form of the very large brown bear in MIS 5a,may have led to C. lupus scavenging kills and particularly,attempting to break open bone for marrow extraction. The absenceof spotted hyaena from Britain at this time would have reducedcompetition for this valuable dietary resource, potentially allowing
p4L 0.192 0.398p4W 0.260 0.097m1Ltrig 0.327 �0.158p3p4D 0.403 �0.008m1m2B 0.245 �0.040P4L 0.316 L0.369P4W 0.288 �0.090M1L 0.229 0.544Group centroidsC. lupus 3.569 �0.077C. mosbachensis �9.790 �0.931C. etruscus �6.506 1.851
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015
DF 1151050-5-10-15
DF
2
15
10
5
0
-5
-10
-15
Group CentroidC. etruscusC. mosbachensisC. lupus
Species
Fig. 7. Plot of individual canonical scores and group centroids on the first and seconddiscriminant functions from the stepwise Discriminant Function Analysis of dietarymeasurements for species groups: C. lupus, C. mosbachensis and C. etruscus.
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e16 11
C. lupus to broaden its niche to include heavier scavenging in aresource limited environment. Despite sites of this age often con-tainingmany tens to hundreds of individual reindeer and bison, it islikely that these migratory herbivores were only seasonally pre-sent, consequently limiting direct hunting options still further. Anyavailable carcasses (either natural or scavenged bear kills) wouldlikely have been rapidly frozen under cold climate conditions,thereby contributing to dental attrition. Although wolves in highlatitudes show a preference for freshly-killed prey, scavenging fromfrozen carcasses is a recognised behaviour (Haynes, 1982).
The MIS 5a group also displayed the highest proportion ofbroken and severely worn teeth. The incorporation of a high pro-portion of bone and/or consumption of frozen carcasses in the dietwould have increased the likelihood of tooth damage, as well asproducing more extreme wear. It is highly unlikely that the MIS 5agroup are all ontogenetically old animals, since the specimens aredrawn from more than one site and the analyses have identifiedfurther variations in tooth morphology. The high level of toothbreakage and wear is therefore interpreted as diet-related. In asimilar study, Binder et al. (2002) found that increased tooth frac-ture and wear in groups of C. dirus from Rancho La Brea wererelated to high levels of competition in a resource-limited envi-ronment and particularly to an increase in the need to consumecarcasses more quickly and completely.
It is possible that the absence of spotted hyaenas further enabledbody size of C. lupus to increase as competitive pressure from abroadly similar carnivore was removed (Rosenweig, 1968),although it is equally possible that this relates to a Bergmannianresponse under cold-climate conditions (Mayr, 1963). From com-parisons of latitude and m1L (used as a body size proxy) in modernEuropeanwolves, with increasing latitude, body sizewas also foundto increase, indicating the effect of Bergmann’s rule in modernEuropean wolves (Flower, 2013).
Although the two remaining Pleistocene groups (MIS 3 and 7)were significantly different from MIS 5a, they themselves werefound to share some similarities, as well as having commonalities
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
with the modern C. lupus group based on the earlier post hoc tests.In terms of body size, MIS 3 and 7 wolves were found to be mostsimilar to each other and on average, slightly smaller than both MIS5a and modern Swedish C. lupus.
The stepwise DFA also foundMIS 7 C. lupus to be separated frommodern (MIS 1) andMIS 5awolves, as well as further distinguishingMIS 7 members from those of MIS 3 on the third discriminantfunction. Generally, in comparison to modern C. lupus, differencesin p4L, M1M2L, M1W, m1m2D and P4W were inferred for MIS 7wolves, with further differentiation from MIS 5a wolves in m1W,m1m2B and m1m2D, as well as M1M2L. Comparatively elongatedp4s in the Pleistocene age groups are difficult to interpret. Therelationship between p4L and bone use suggested for MIS 5aapparently holds true, since tooth breakage was slight and toothwear moderate for wolves of this time. Since both MIS 3 and 7wolves are of apparent smaller size than those of MIS 5a, it is un-likely that the increased length correlates with larger body sizes.Hence, elongation of p4 may be related to an as-yet unidentifiedfactor.
Shallower and narrower jaws (and consequently reduced jawstrength) suggest that the focus was on comparatively smaller preyduring MIS 3 and 7, which likely led to greater dietary generalisa-tion, perhaps related to themore temperate climatic conditions andthe availability of a wider range of plant, insect and other foods.Although herbivore preywas both abundant and diverse during thelater parts of MIS 7 and the carnivore guild contained less com-petitors than in the early Middle Pleistocene, it is noticeable thatmany ungulates, as well as the lion, were of particularly large bodymass at this time (Schreve, 1997; Collinge, 2001), potentiallydirecting the relatively smaller MIS 7 wolves towards abundantsmaller prey and a more mixed omnivorous diet.
In comparison, MIS 3 C. lupus, which was separated similarly toMIS 7 on the most explanatory functions, was distinguished frommodern wolves by differences in p4L, m1m2D, P4W, M1W as wellas M1M2L. Separation from MIS 5a members was further high-lighted by differences in m1W, m1m2D, m1m2B, and M1M2L.However, on the least explanatory third function, MIS 3 was alsoseparated from MIS 5a, and in particular from MIS 7, by additionaldifferences in M1W, m1m2B, P4W and M1M2L.
Overall, the variation in dietary measurements in MIS 3 C. lupuswas similar to that seen in the MIS 7 group. MIS 3 wolves areinterpreted as having a slight inclination towards crushing activ-ities on account of larger molar widths, although they are moresimilar to MIS 1 than to MIS 7 wolves. Shorter M1M2L here is un-expected, although it may relate to the focus on larger crushingareas being present lingually as opposed to buccal lengthening.Nonetheless, this would suggest that high proportions of non-fleshfoods were being incorporated into the diet, more so than for theMIS 7 group. MIS 3 C. lupus also had moderately shallower jaws,albeit perhaps broader than in the MIS 7 group, indicating thatsmaller-sized prey was taken. Middle Devensian wolves maytherefore have been more generalist than their counterparts in MIS5a and less able to take down large-sized prey. This may be a resultof competition from contemporary major predators such as lions,brown bears and particularly spotted hyaena limiting access tolarger prey. Both MIS 7 and MIS 3 groups have similarly low per-centages of broken teeth, although the latter have comparativelyhigher levels of tooth wear, perhaps reflecting the slight increase incrushing ability noted above. However, as tooth wear is mostlymoderate, with no obvious skew towards either severe or slightwear, it is possible that the patterns reflect individual age-relateddental attrition rather than dietary stress. In contrast, the MIS 7group contains a higher proportion of slightly worn teeth, com-bined with low breakage. This may reflect the low specialisationtowards crushing or even collector bias towards younger-aged
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015
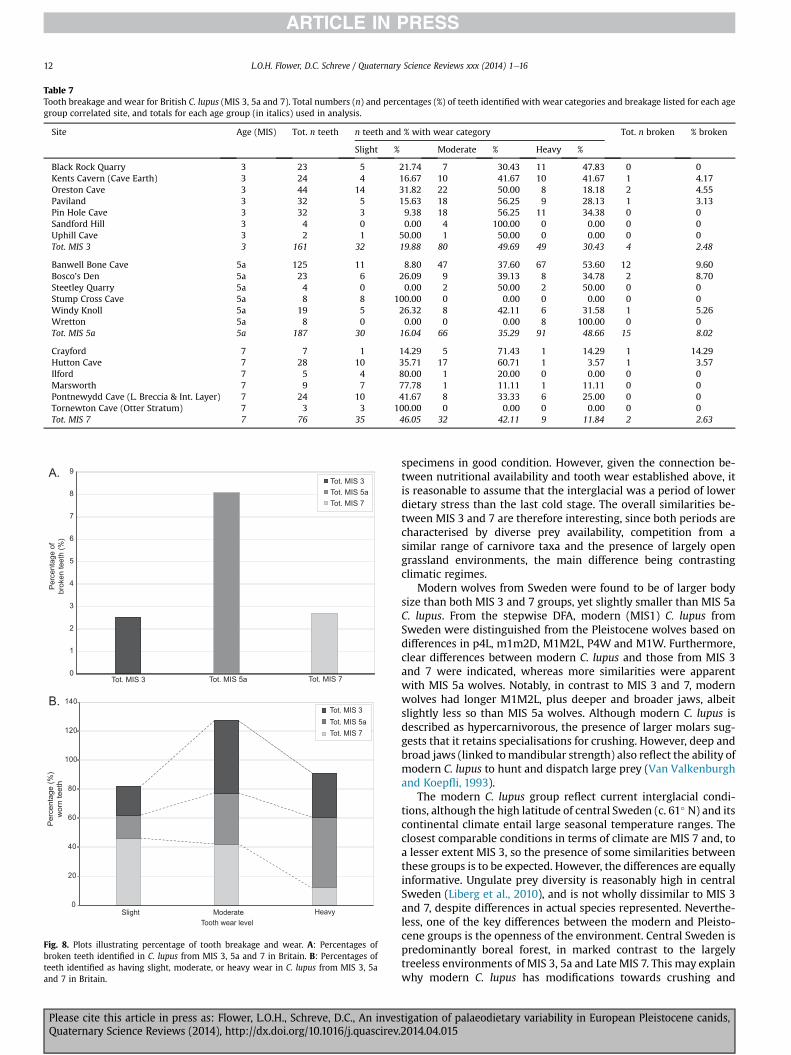
Table 7Tooth breakage and wear for British C. lupus (MIS 3, 5a and 7). Total numbers (n) and percentages (%) of teeth identified with wear categories and breakage listed for each agegroup correlated site, and totals for each age group (in italics) used in analysis.
Site Age (MIS) Tot. n teeth n teeth and % with wear category Tot. n broken % broken
Slight % Moderate % Heavy %
Black Rock Quarry 3 23 5 21.74 7 30.43 11 47.83 0 0Kents Cavern (Cave Earth) 3 24 4 16.67 10 41.67 10 41.67 1 4.17Oreston Cave 3 44 14 31.82 22 50.00 8 18.18 2 4.55Paviland 3 32 5 15.63 18 56.25 9 28.13 1 3.13Pin Hole Cave 3 32 3 9.38 18 56.25 11 34.38 0 0Sandford Hill 3 4 0 0.00 4 100.00 0 0.00 0 0Uphill Cave 3 2 1 50.00 1 50.00 0 0.00 0 0Tot. MIS 3 3 161 32 19.88 80 49.69 49 30.43 4 2.48
Banwell Bone Cave 5a 125 11 8.80 47 37.60 67 53.60 12 9.60Bosco’s Den 5a 23 6 26.09 9 39.13 8 34.78 2 8.70Steetley Quarry 5a 4 0 0.00 2 50.00 2 50.00 0 0Stump Cross Cave 5a 8 8 100.00 0 0.00 0 0.00 0 0Windy Knoll 5a 19 5 26.32 8 42.11 6 31.58 1 5.26Wretton 5a 8 0 0.00 0 0.00 8 100.00 0 0Tot. MIS 5a 5a 187 30 16.04 66 35.29 91 48.66 15 8.02
Crayford 7 7 1 14.29 5 71.43 1 14.29 1 14.29Hutton Cave 7 28 10 35.71 17 60.71 1 3.57 1 3.57Ilford 7 5 4 80.00 1 20.00 0 0.00 0 0Marsworth 7 9 7 77.78 1 11.11 1 11.11 0 0Pontnewydd Cave (L. Breccia & Int. Layer) 7 24 10 41.67 8 33.33 6 25.00 0 0Tornewton Cave (Otter Stratum) 7 3 3 100.00 0 0.00 0 0.00 0 0Tot. MIS 7 7 76 35 46.05 32 42.11 9 11.84 2 2.63
0
20
40
60
80
100
120
140
Tooth wear levelSlight Moderate Heavy
Per
cent
age
(%)
wor
n te
eth
Tot. MIS 3Tot. MIS 5aTot. MIS 7
B.
0
1
2
3
4
5
6
7
8
9
Per
cent
age
of
brok
en te
eth
(%)
Tot. MIS 3 Tot. MIS 5a Tot. MIS 7
Tot. MIS 3Tot. MIS 5aTot. MIS 7
A.
Fig. 8. Plots illustrating percentage of tooth breakage and wear. A: Percentages ofbroken teeth identified in C. lupus from MIS 3, 5a and 7 in Britain. B: Percentages ofteeth identified as having slight, moderate, or heavy wear in C. lupus from MIS 3, 5aand 7 in Britain.
L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e1612
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
specimens in good condition. However, given the connection be-tween nutritional availability and tooth wear established above, itis reasonable to assume that the interglacial was a period of lowerdietary stress than the last cold stage. The overall similarities be-tween MIS 3 and 7 are therefore interesting, since both periods arecharacterised by diverse prey availability, competition from asimilar range of carnivore taxa and the presence of largely opengrassland environments, the main difference being contrastingclimatic regimes.
Modern wolves from Sweden were found to be of larger bodysize than both MIS 3 and 7 groups, yet slightly smaller than MIS 5aC. lupus. From the stepwise DFA, modern (MIS1) C. lupus fromSweden were distinguished from the Pleistocene wolves based ondifferences in p4L, m1m2D, M1M2L, P4W and M1W. Furthermore,clear differences between modern C. lupus and those from MIS 3and 7 were indicated, whereas more similarities were apparentwith MIS 5a wolves. Notably, in contrast to MIS 3 and 7, modernwolves had longer M1M2L, plus deeper and broader jaws, albeitslightly less so than MIS 5a wolves. Although modern C. lupus isdescribed as hypercarnivorous, the presence of larger molars sug-gests that it retains specialisations for crushing. However, deep andbroad jaws (linked tomandibular strength) also reflect the ability ofmodern C. lupus to hunt and dispatch large prey (Van Valkenburghand Koepfli, 1993).
The modern C. lupus group reflect current interglacial condi-tions, although the high latitude of central Sweden (c. 61� N) and itscontinental climate entail large seasonal temperature ranges. Theclosest comparable conditions in terms of climate are MIS 7 and, toa lesser extent MIS 3, so the presence of some similarities betweenthese groups is to be expected. However, the differences are equallyinformative. Ungulate prey diversity is reasonably high in centralSweden (Liberg et al., 2010), and is not wholly dissimilar to MIS 3and 7, despite differences in actual species represented. Neverthe-less, one of the key differences between the modern and Pleisto-cene groups is the openness of the environment. Central Sweden ispredominantly boreal forest, in marked contrast to the largelytreeless environments of MIS 3, 5a and Late MIS 7. This may explainwhy modern C. lupus has modifications towards crushing and
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e16 13
deeper jaws, since the pursuit of prey is more difficult in closedhabitats and there are increased hiding places for small to mediumprey, making hunting a more labour-intensive exercise for lessnutritional reward. In boreal forest across northern Europe, themajority of ungulates are of large body size (from c.200 kg in adultred deer to c.800 kg in elk; Nowak, 1991) and may be difficult tofind in the forest, solitary or only seasonally available. Therefore,despite the difficulties and dangers in subduing such animals, op-portunities to take such large prey must be seized, hence therequirement for much stronger jaws in modern Swedish wolves incomparison to those of MIS 3 and 7. A final key difference is thatwith the exception of brown bear, wolf is now the largest predatorin the Palaearctic and the only one equipped for pursuit hunting.The dietary flexibility of the wolf has undoubtedly been one of thereasons for its past success and indeed, its ability to cope in mar-ginal environments as a result of modern human persecution.
Along with the dietary differences, temporal differences are alsoapparent with body size, with the more hypercarnivorous wolvesfrom modern Sweden and potentially MIS 5a being slightly larger,and the more generalist MIS 3 and MIS 7 populations being smalleron average. These findings therefore do not support the presence inBritain in MIS 3 of the large sized Canis lupus maximus (Boudadi-Maligne, 2012), which is present in southwest France at this time.
A more complex scenario is therefore present than envisaged byTurner (1981), whereby body mass changes were related primarilyto climatic forcing. It is possible that the relatively larger body sizesencountered in the modern Swedish population and MIS 5a areexplicable by Bergmann’s Rule, reflecting both the high latitudeorigin of the modern wolves from Sweden and the intense coldclimatic conditions of MIS 5a in Britain. However, as seen above,dietary variability may also be influential. Additionally, within-group variation was present in each chronological grouping. Formodern C. lupus, this variation is a function of sexual size dimor-phism. Such dimorphism may also be present in the Pleistocenegroups, as originally suggested by Turner (1981), based on thedifferences in tooth dimensions between Ipswichian and Deven-sian C. lupus. However, it is worth noting that sexual size dimor-phism in wolves is considered to be slight in comparison to othercarnivores such as felids and mustelids (Ewer, 1973; Dayan et al.,1992). Comparable variation in body size has also been noted inPleistocene subspecies of C. lupus in France, correlatedwith climaticand environmental change (Brugal and Boudadi-Maligne, 2011).
7.2. Comparison with the palaeodiets of other Pleistocene canids
The intraspecific variability of C. lupus seems to be uniquewithin the Pleistocene canids analysed here. NeitherC. mosbachensis nor C. etruscus varied between assemblages,despite competition from a richer carnivore guild including othercanids. This indicates comparative morphological stability in thesetaxa and inferred dietary constancy in contrast to C. lupus. The midEarly Pleistocene was a period of climatic stability, pre-dating theMid-Pleistocene Revolution and the switch to eccentricity-dominated Milankovitch cycles. Although both the Olivola andTasso F.U.s are considered to be close in time in the Early Pleisto-cene (Raia et al., 2006), the lack of variation in C. etruscus betweenthese faunal units is all the more surprising, given the increase incarnivore competition over this period. C. etruscus went fromapparently being a lone canid during the Olivola F.U., to beingjoined by the smaller C. arnensis and the larger C. falconeri duringthe Tasso F.U. Although C. etruscus, as well as C. arnensis andC. falconeri, was considered by Cherin et al. (2013b) as having a dietincluding >70% meat (following Van Valkenburgh’s (1988a, 1989)carnivore dietary categories), the precise details of individualniche differentiation between these canids remain obscure, with
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
the role of C. etruscus apparently stable and not affected by thearrival of other canid taxa.
Similar temporal stability is apparent with C. mosbachensis. Itshould be noted that although the C. mosbachensis sampleconsidered here covers temperate-climate episodes only, it is bothgeographically and chronologically extensive, ranging from the LateEarly Pleistocene of Germany to the early Middle Pleistocene ofBritain and covering around half amillion years. At this time, Britainremained connected by a landbridge to mainland Europe, therebyallowing free movement of species and enhancing the overallsimilarity of British and continental taxa (Stuart, 1995). C. (X.)lycaonoides also co-occurred with C. mosbachensis in both theUntermassfeld and Westbury-sub-Mendip assemblages, althoughthis did not apparently lead to morphological change in the smallerspecies; the presence of a larger canid may thus have effectivelyconstrained C. mosbachensiswithin its dietary niche. Although boththe Late Early and early Middle Pleistocene contained a morediverse carnivore guild, the presence of diverse and apparentlyabundant prey may have alleviated competitive pressure througheffective resource partitioning in a productive environment.
Stepwise DFA of the species groups was more revealing. Clearseparation and very strong group membership to each speciesgroup was found (99.30% for cross validated cases). C. lupus (thelargest species based on m1 length) had overall greater measure-ments in comparison to C. etruscus and C. mosbachensis and wasmost clearly separated from the latter. The most discriminatingdietary measurements selected by the model for C. lupus were p4width, jaw depth (at the p3-p4 junction), upper carnassial size, andlower carnassial blade length. In comparison to C. etruscus andC. mosbachensis, C. lupus also has more intermediate sized M1lengths and p4 lengths in comparison to C. etruscus, and moremoderate upper carnassial lengths than in C. mosbachensis, perhapssignifying a relative lack of specialisation in comparison to theother canids.
C. etruscus and C. mosbachensis were more similar to each otherthan to C. lupus, both in their relative smaller body size and thedimensions of the dietary indicators, with C. mosbachensis thesmallest, as well as most differentiated by the stepwise DFA.Bearing this size difference in mind, disparities in upper molarlength, p4 length and upper carnassial lengths become particularlyimportant in discriminating between these species. It is suggestedthat C. etruscusmay have been more of a mesocarnivore than eitherC. lupus or C. mosbachensis, its enhanced crushing capabilities andshorter/narrower carnassials blades reflecting a higher proportionof non-flesh foods in the diet and thus a wider dietary breadth andbehavioural flexibility. This correlates well with a previous studysuggesting C. etruscus had a relatively omnivorous diet (Croitor andBrugal, 2010). The combination of modest size and shallow jawsalso indicate that smaller sized prey was the focus, especially aslarger carnivore competitors may have excluded C. etruscus fromtackling larger prey anyway. However, this is in contrast to Meloro(2011), who inferred that based on mandibular shape analysis,C. etruscus was a consumer of both large and small prey.
The highest degree of separation from C. lupus was shown byC. mosbachensis. Not only was the latter the smallest in body size ofall three taxa but its inferred reduced blades on narrower carnas-sials, reduced upper molars and shallower jaws imply a lesscarnivorous lifestyle than C. lupus and an ability to take only smallprey. A similar conclusion was reached by Meloro (2011), based onmandibular shape analysis. However, C. mosbachensis was furtherdifferentiated from C. lupus and C. etruscus by its reduced crushingcapability and potential increased slicing ability indicated byelongated upper carnassials. This implies both a more carnivorousmode than C. etruscus and more dietary specialisation than C. lupus.This contradicts the view that C. mosbachensis was an omnivorous
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e1614
canid (Palmqvist et al., 1999). By developing faster slicing ability (asevidenced by changes in the upper carnassials), any potential di-etary inflexibility resulting from reduced crushing mechanismsmay have been counteracted, thereby equipping C. mosbachensiswith a competitive stress coping mechanism.
It is important to note that the clear separation of the canids bythe stepwise DFA is suggestive of differences in body size influ-encing the analysis. Aside from the associated problems of usingratios in analyses (see earlier), removal of size-related effects indietary analyses is difficult. Nonetheless, as predator size correlateswith diet and ecology of the species, it is perhaps more informativeto consider diet and body size together but to retain a caveat in anydietary inferences.
8. Conclusions
In summary, it can be seen that body size, species diversity, levelof competition, openness of the environment and climatic condi-tions were important factors in influencing canid diet. Of the threecanids studied, C. lupus is the only one to show variation in terms oflevel of carnivory and body size (based on m1 length). During MIS5a and when faced with low prey availability, severe climaticconditions and dietary stress, C. lupus is thought to have adopted amore hypercarnivorous mode than in previous periods, includingincreased scavenging and bone consumption (facilitated by theabsence from Britain of spotted hyaena). The development of anefficient flesh-slicing apparatus, together with strong jaws, point toa hypercarnivorous diet, doubtless exacerbated by the limitedavailability of non-meat foods at this time. In contrast, wolves fromboth MIS 3 and MIS 7 demonstrate a more omnivorous diet, withincreased crushing capabilities when compared to the MIS 5agroup. Both periods are characterised by open environments andhigh prey availability, so differences relating to tooth wear mayrelate to the contrasting climatic conditions present. Although MIS3 is within the last cold stage, it represents a period of climaticamelioration compared to the Early Devensian of MIS 5a, withsummer temperatures around 4 �C below the lower end of the MIS7 summer temperature range but much colder winters. With moreabundant prey in both periods and the presence of a wider range ofnon-meat foods when compared with MIS 5a, wolves in MIS 7 andMIS 3 were able to adopt a more generalist diet. Nevertheless, theirshallower jaws indicate that they were able to tackle only small-medium prey and were probably outcompeted by (particularly)lions and spotted hyaena.
Although more comparable in terms of climate and prey di-versity with MIS 3 and 7, the modern (MIS 1) C. lupus sample fromSweden has many differences with these two groups and is regar-ded as beingmore hypercarnivorous. The study populations inhabita forested rather than open environment, which may lead to dif-ficulties and greater energy expenditure capturing smaller prey,thereby necessitating exploitation of large prey. As the only largepursuit predator in its environment, the modern Swedish wolfaccordingly possesses strong and deep jaws to subdue andconsume large prey. Its reduced ability to slice flesh quickly prob-ably reflects minimal intraspecific competition. Nevertheless, theapparent specialisation towards crushing indicates that underbroadly temperate-climate conditions, a large range of non-meatfoods is also incorporated in the diet.
The differing capabilities to crush, slice meat and eat bone un-derscore the flexibility of the cranio-dental morphology of C. lupusand explain its success as a species during the Quaternary. Theevidence from discrete groups from coeval sites indicates that suchdietary changes are not reflective of local ecomorphs but ratherreveal species-wide dietary shifts in response to particular climaticand ecological parameters. Cranio-dental plasticity within C. lupus
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
has also been observed in the Late Pleistocene of North America,with Eastern Beringianwolves more specialised towards killing andconsuming relatively large prey and/or habitual scavenging incomparison to contemporary wolves from Rancho La Brea and theirextant counterparts (Leonard et al., 2007). This highlights thesuccess of C. lupus as a species in adapting to different environ-mental conditions.
In comparison to the flexibly hypercarnivorous C. lupus,C. etruscuswas found to be a mesocarnivore, whilst C. mosbachensiswas less carnivorous than C. lupus, but also less omnivorous thanC. etruscus. The main differences between the three canids are notonly apparent through examination of the cranio-dental di-mensions but are also correlated with variations in body size. Thus,the hypercarnivorous C. lupus is relatively larger than the meso-carnivore C. etruscus and the relatively more carnivorousC. mosbachensis. However, although similar in body size,C. mosbachensis and C. etruscus adopted subtly different dietarystrategies, possibly relating to the presence of different carnivorecompetitors within similarly large and diverse carnivore guilds.Overall, both C. etruscus and C. mosbachensis are remarkably con-stant in their morphology, both temporally and geographically.
The within-group body size variation for C. lupus requiresfurther explanation and may be related either to sexual sizedimorphism or to Bergmann’s Rule. The British sites used in thisstudy are individually well-dated with multiple relative dates and/or absolute geochronology and are therefore taken as representa-tive of particular climatic stages. That said, they may not be pre-cisely contemporary within individual marine Oxygen IsotopeStages (a particular issue given the rapid climatic oscillations of MIS3 and more widely, the standard errors implicit in different radio-metric methods) and minor temporal and regional differences maybe present. An important element for future study relates to theperiodic isolation of Britain from the European mainland, followingthe creation of the Strait of Dover during the Late Middle Pleisto-cene. The imposition of a biogeographical barrier during periods ofhigh sea level has the potential to enhance temporal variation inboth diet and body size of British wolf populations as a result oftheir isolation. Comparison of British Pleistocene C. lupus withspecimens from well-dated sites on the European mainland mightreveal whether the temporal differences observed extended over amuch larger geographical area or whether they were peculiar toBritain.
In conclusion, the survival of C. lupus during the Pleistocene andits continued success in modern times can be attributed largely toits plastic cranio-dental morphology. The ability to alter its diet andmodify its dental specialisation to cope with changing ecologicalconditions is a reflection of the baseline flexibility of this mostadaptable of carnivores.
Acknowledgements
The authors would like to thank two anonymous reviewers fortheir comments, which greatly improved the manuscript. The au-thors acknowledge Andy Currant for general discussion andgranting access to material at the NHM, and Simon Blockley forstatistical guidance. The authors would also like to thank thefollowing who generously allowed access to material and providedhelp with specimens: Roger Vaughan (Bristol City Museum), PaulShepherd (British Geological Survey, Keyworth), Mike Palmer(Buckinghamshire County Museum), David Gelsthorpe (Man-chester Museum), Elizabeth Walker (National Museum of Wales,Cardiff), Matthew Riley (The Sedgwick Museum of Earth Sciences,Cambridge), Dennis Parsons (Somerset Heritage Centre, Taunton),Nicholas Riall (Swansea Museum), Barry Chandler and JennyHumphries (Torquay Museum), Matthew Lowe (University
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e16 15
Museum of Zoology Cambridge), Neil Turner (Woollaton Hall,Nottingham), Ralf-Dietrich Kahlke (Forschungsstation für Quar-tärpaläontologie, Weimar), Lorenzo Rook and Elisabetta Cioppi(Museo di Storia Naturale degli Studi di Firenze), Lars Werdelin andDaniela Kalthoff (Naturhistoriska riksmuseet, Stockholm).
Funding for this research was provided by: Reid ResearchScholarship, RHUL (for Ph.D. funding); QRA New Research WorkersAward (for visiting museum collections in southwest Britain);SYNTHESYS (SE-TAF-1784) financed by the European Community-Research Infrastructure Action under FP7 ‘Capacities’ specific pro-gramme (for visiting the modern C. lupus collection at Natur-historiska Riksmuseet, Stockholm). This paper is dedicated to thememory of Alan Turner, carnivore supremo and friend.
References
Albrecht, G.H., 1978. Some comments on the use of ratios. Syst. Zool. 27 (1), 67e71.Ansorge, H., Kluth, G., Hahne, S., 2006. Feeding ecology of wolves Canis lupus
returning to Germany. Acta Theriol. 51 (1), 99e106.Arnborg, T., 1990. Forest types of northern Sweden, introduction to and translation
of ‘Det nordsvenska skogstypsschemat’. Vegatatio 90, 1e13.Atchley, W.R., Gaskins, C.T., Anderson, D., 1976. Statistical properties of ratios. I.
Empirical results. Syst. Zool. 25, 137e148.Atchley, W.R., 1978. Ratios, regression intercepts and the scaling of data. Syst. Zool.
27 (1), 78e83.Azzaroli, A., 1983. Quaternary mammals and the ‘End Villafranchian’ dispersal event
e a turning point in the history of Eurasia. Palaeogeogr. Palaeoclim. Palaeoecol.44, 117e139.
Azzaroli, A., De Giuli, C., Ficcarelli, G., Torre, D., 1988. Late Pliocene to Early Mid-Pleistocene mammals in Eurasia: faunal succession and dispersal events.Palaeogeogr. Palaeoclim. Palaeoecol. 66, 77e100.
Bellucci, L., Mazzini, I., Scardia, G., Bruni, L., Parenti, F., Giacomo Segre, A., SegreNaldini, E., Sardella, R., 2012. The site of Coste San Giacomo (Early Pleistocene,central Italy): Palaeoenvironmental analysis and biochronological overview.Quat. Int. 267, 30e39.
Binder, W.J., Thompson, E.N., Van Valkenburgh, B., 2002. Temporal variation intooth fracture among Rancho La Brea dire wolves. J. Vertebr. Paleontol. 22 (2),423e428.
Binder, W.J., Van Valkenburgh, B., 2010. A comparison of tooth wear and breakage inRancho La Brea sabertooth cats and dire wolves across time. J. Vertebr. Pale-ontol. 30 (1), 255e261.
Boudadi-Maligne, M., 2012. Une nouvelle sous-espèce de loup (Canis lupus maximusnov. Subsp.) dans le Pléistocène supérieur d’Europe occidentale. C. R. Palevol. 11,475e484.
Boyd, D.K., Ream, R.R., Pletscher, D.H., Fairchild, M.W., 1994. Prey taken by colo-nising wolves and hunters in the glacier National Park area. J. Wildl. Manag. 58(2), 289e295.
Brugal, J.-P., Boudadi-Maligne, M., 2011. Quaternary small to large canids in Europe:taxonomic status and biochronological contribution. Quat. Int. 243 (1), 171e182.
Candy, I., Schreve, D., 2007. Land-sea correlation of Middle Pleistocene temperatesub-stages using high-precision uranium-series dating of tufa deposits fromsouthern England. Quat. Sci. Rev. 26, 1223e1235.
Capitani, C., Bertelli, I., Varuzza, P., Scandura, M., Apollonio, M., 2003. A comparativeanalysis of wolf (Canis lupus) diet in three different Italian ecosystems. Mamm.Biol. 69 (1), 1e10.
Cherin, M., Bertè, D.F., Rook, L., Sardella, R., 2013a. Re-defining Canis etruscus(Canidae, Mammalia): a new look into the evolutionary history of Early Pleis-tocene dogs resulting from the outstanding fossil record from Pantalla (Perugia,central Italy). J. Mamm. Evol. http://dx.doi.org/10.1007/s10914-013-9227-4.
Cherin, M., Berté, D.F., Sardella, R., Rook, L., 2013b. Canis etruscus (Canidae, Mam-malia) and its role in the faunal assemblage from Pantalla (Perugia, centralItaly): comparison with the Late Villafranchian large carnivore guild of Italy.Boll. Soc. Paleontol. Ital. 52 (1), 11e18.
Collinge, S.E., 2001. Body Size and Community Structure in British PleistoceneMammals (Ph.D thesis). University College London, U.K.
Coope, G.R., 2010. Coleopteran faunas as indicators of interglacial climates in centraland southern England. Quat. Sci. Rev. 29, 1507e1514.
Corrunccini, R.S., 1975. Multivariate analysis in biological anthropology: someconsiderations. J. Hum. Evol. 4, 1e19.
Croitor, R., Brugal, J.-P., 2010. Ecological and evolutionary dynamics of the carnivorecommunity in Europe during the last 3 million years. Quat. Int. 212, 98e108.
Currant, A.P., 1984. The Mammalian remains. In: Green, H.S. (Ed.), PontnewyddCave. A Lower Palaeolithic Hominid Site in Wales, The First Report. NationalMuseum Wales Quaternary Studies Monograph, 1, pp. 177e181.
Currant, A., Jacobi, R., 2001. A formal mammalian biostratigraphy for the LatePleistocene of Britain. Quat. Sci. Rev. 20, 1707e1716.
Currant, A., 2004. The Quaternary mammal collections at the Somerset CountyMuseum, Taunton. In: Schreve, D.C. (Ed.), The Quaternary Mammals of Southernand Eastern England Field Guide. Quaternary Research Association, pp. 101e109.
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
Currant, A.P., Jacobi, R.M., 2011. The mammalian faunas of the British Late Pleisto-cene. In: Ashton, N.M., Lewis, S.G., Stringer, C.B. (Eds.), The Ancient HumanOccupation of Britain. Dev. Quat. Sci., vol. 14. Elsevier, pp. 165e180.
Dayan, T., Simberloff, D., Tchernov, E., Yom-Tov, Y., 1992. Canine carnassials: char-acter displacement in the wolves, jackals and foxes of Israel. Biol. J. Linn. Soc. 45,315e331.
Ewer, R.F., 1973. The Carnivores. Weidenfeld and Nicolson, London, U.K.Flower, L.O.H., 2013. Canid evolution and palaeoecology in the Pleistocene of
western Europe, with particular reference to the wolf Canis lupus L. 1758 (Un-published Ph.D. thesis). University of London.
Fritts, S.H., Mech, L.D., 1981. Dynamics, movements, and feeding ecology of a newlyprotected wolf population in northwestern Minnesota. Wildl. Monogr. 80, 3e79.
Garrido, G., Arribas, A., 2008a. Canis accitanus nov. sp., a new small dog (Canidae,Carnivora, Mammalia) from Fonelas P-1 Plio-Pleistocene site (Gaudix basin,Granada, Spain). Geobios 41, 751e761.
Garrido, G., Arribas, A., 2008b. Reflections on the carnivores of the upper Villa-franchiam represented in the fossil record of the Fonelas P-1 site. In: Arribas, A.(Ed.), Vertebrados del Plioceno superior terminal en el suroeste de Europa:Fonelas P-1 y el Proyecto Fonelas. Cuadernos del Museo Geominero, vol. 10.Instituto Geológico y Minero de Espana, Madrid, pp. 85e146.
Gilmour, M., Currant, A., Jacobi, R., Stringer, C., 2007. Recent TIMS dating resultsfrom British Late Pleistocene vertebrate faunal localities: context and inter-pretation. J. Quat. Sci. 22 (8), 793e800.
Gingerich, P.D., 1974. Size variability of the teeth in living mammals and thediagnosis of closely related sympatric fossil species. J. Palaeontol. 48 (5), 895e903.
Gliozzi, E., Abbazzi, L., Argenti, A., Azzaroli, A., Caloi, L., Capasso Barbato, L., diStefano, G., Esu, D., Ficcarelli, G., Girotti, O., Kotsakis, T., Masini, F., Mazza, P.,Mezzabotta, C., Palombo, M.R., Petronio, C., Rook, L., Sala, B., Sardella, R.,Zanalda, E., Torre, D., 1997. Biochronology of selected mammals. Molluscs andostracodes from the Middle Pliocene to the Late Pleistocene in Italy. The state ofthe art. Riv. Ital. Paleontol. Stratigr. 103 (3), 369e388.
Grootes, P.M., Stulver, M., White, J.W.C., Johnsen, S., Jouzel, J., 1993. Comparison ofoxygen isotope records from the GISP2 and GRIP Greenland ice cores. Nature366, 552e554.
Haynes, G., 1982. Utilization and skeletal disturbances of North American preycarcasses. Arctic 35 (2), 266e281.
Jedrzejewski, W., Jedrzejewska, B., Okarma, H., Schmidt, K., Zub, K., Musiani, M.,2000. Prey selection and predation by wolves in Bia1owieza Primeval Forest,Poland. J. Mamm. 81 (1), 197e212.
Kahlke, R.-D., 2000. The Early Pleistocene (Epivillafranchian) faunal site of Unter-massfeld (Thuringia, Central Germany) synthesis of new results. Eraul 92, 123e138.
Kahlke, R.-D., Garcia, N., Kostopoulos, D.S., Lacombat, F., Lister, A.M., Mazza, P.P.A.,Spassov, N., Titov, V.V., 2011. Western Palaearctic palaeoenvironmental condi-tions during the Early and early Middle Pleistocene inferred from largemammal communities, and implications for hominin dispersal in Europe. Quat.Sci. Rev. 30, 1368e1395.
Kojola, I., Huitu, O., Toppinen, K., Heikura, K., Heikkinen, S., Ronkainen, S., 2004.Predation on European wild forest reindeer (Rangifer tarrandus) by wolves(Canis lupus) in Finland. J. Zool. Lond. 263, 229e235.
Koufos, G.D., Kostopoulos, D.S., 1997. New carnivore material from the Plio-Pleistocene of Macedonia (Greece) with the description of a new canid.Münch. Geowiss. Abh. A 34, 33e63.
Kurtén, B., 1968. Pleistocene Mammals of Europe. Weidenfeld and Nicholson,London.
Kurtén, B., Poulianos, A.N., 1977. New stratigraphic and faunal material from Pet-ralona Cave with special reference to the Carnivora. Anthropos. Athens 4, 47e130.
Lacombat, F., Abbazzi, L., Ferretti, M.P., Martinez-Navarro, B., Moulle, P.-E.,Palombo, M.-R., Rook, L., Turner, A., Valli, A.M.-F., 2008. New data on the earlyVillafranchian fauna from Vialette (Haute-Loire, France) based on the collectionof the Crozatier Museum (Le Puy-en-Velay, Haute-Loire, France). Quat. Int. 179,64e71.
Landon, D.B., Waite, C.A., Peterson, R.O., Mech, L.D., 1998. Evaluation of age deter-mination techniques for Gray wolves. J. Wildl. Manag. 62 (2), 674e682.
Legendre, S., Roth, C., 1988. Correlation of carnassial tooth size and body weight inrecent carnivores (Mammalia). Hist. Biol. 1, 85e98.
Leonard, J.A., Vilà, C., Fox-Dobbs, K., Koch, P.L., Wayne, R.K., Van Valkenburgh, B.,2007. Megafaunal extinctions and the disappearance of a specialised wolfecomorph. Curr. Biol. 17, 1146e1150.
Lewis, M., Pacher, M., Turner, A., 2010. The larger Carnivora of the West Runtonfreshwater bed. Quat. Int. 228 (1e2), 116e135.
Liberg, O., Bergström, R., Kindberg, J., 2010. Ungulates and their management inSweden. In: Apollonio, M., Andersen, R., Putman, R. (Eds.), European Ungulatesand Their Management in the 21st century. Cambridge University Press, Cam-bridge, U.K.
Macdonald, D.W., 1983. The ecology of carnivore social behaviour. Nature 301, 379e384.
Maddy, D., Lewis, S.G., Scaife, R.G., Bowen, D.Q., Coope, G.R., Green, C.P., Hardaker, T.,Keen, D.H., Rees-Jones, J., Parfitt, S., Scott, K., 1998. The Upper Pleistocene de-posits at Cassington, near Oxford, England. J. Quat. Sci. 13 (3), 205e231.
Martinez-Navarro, B., Rook, L., 2003. Gradual evolution in the African hunting doglineage. Systematic implications. C. R. Palevol. 2, 695e702.
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015

L.O.H. Flower, D.C. Schreve / Quaternary Science Reviews xxx (2014) 1e1616
Martinez-Navarro, B., Belmaker, M., Bar-Yosef, O., 2009. The large carnivores from’Ubeidiya (early Pleistocene, Israel): biochronological and biogeographical im-plications. J. Hum. Evol. 56 (5), 514e524.
Mayr, E., 1963. Animal Species and Evolution. Harvard University Press, Cambridge,U.S.A.
Mech, L.D., 1970. The Wolf: the Ecology and Behaviour of an Endangered Species.The Natural History Press, New York.
Mech, L.D., Frenzel Jr., L.D., Karns, P.D., 1971. The effect of snow conditions on thevulnerability of white-tailed deer to wolf predation. In: Mech, L.D.,Frenzel Jr., L.D. (Eds.), Ecological Studies of the Timber Wolf in NortheasternMinnesota. Forest Service: U.S. Department of Agriculture, pp. 51e59.
Mech, L.D., Boitani, L., (IUCN SSC wolf specialist group), 2010. Canis lupus. In: IUCN2012. IUCN Red List of Threatened Species. Version 2012.2. www.iucnredlist.org. Downloaded on 03 April 2013.
Meloro, C., Clauss, M., 2012. Predator-prey fluctuations in the Plio-Pleistocene.Palaios 27, 90e96.
Meloro, C., 2011. Feeding habits of Plio-Pleistocene large carnivores as revealed bythe mandibular geometry. J. Vertebr. Paleontol. 31 (2), 428e446.
Meloro, C., Elton, S., Louys, J., Bishop, L.C., Ditchfield, P., 2013. Cats in the forest:predicting habitat adaptations from humerus morphometry in extant and fossilFelidae (Carnivora). Paleobiology 39 (3), 323e344.
Meriggi, A., Lovari, S., 1996. A review of wolf predation in southern Europe: does thewolf prefer wild prey to livestock? J. Appl. Ecol. 33 (6), 1561e1571.
Moberg, A., Bergström, H., 1997. Homogenization of Swedish temperature data. PartIII: the long temperature records from Uppsala and Stockholm. Int. J. Climatol.17, 667e699.
Nowak, R., 1991. Walker’s Mammals of the World. Johns Hopkins University Press,Baltimore.
Nowak, S., Mys1ajek, R.W., K1osi�nska, A., Gabry�s, G., 2011. Diet and prey selection ofwolves (Canis lupus) recolonising Western and Central Poland. Mamm. Biol. 76,709e715.
Palmqvist, P., Arribas, A., Martinez-Navarro, B., 1999. Ecomorphological study oflarge canids from the lower Pleistocene of southeastern Spain. Lethaia 32, 75e88.
Paquet, P.C., 1992. Prey use strategies of sympatric wolves and coyotes in RidingMountain National Park, Manitoba. J. Mammol. 73 (2), 337e343.
Petrucci, M., Cipullo, A., Martìnez Navarro, B., Rook, L., Sardella, R., 2013. The lateVillafranchian (Early Pleistocene) carnivores (Carnivora, Mammalia) from PirroNord (Italy). Palaeontogr. Abt. a Paläozool. Stratigr. 298, 113e145.
Preece, R.C., Parfitt, S.A., 2000. The Cromer forest-bed formation: new thoughts onan old problem. In: Lewis, S.G., Whiteman, C.A., Preece, R.C. (Eds.), The Qua-ternary of Norfolk and Suffolk Field Guide. Quaternary Research Association,pp. 1e29.
Pullianen, E., 1975. Wolf ecology in Northern Europe. In: Fox, M.W. (Ed.), The WildCanids. Van Nostrand Reinhold Co, pp. 292e299.
Raia, P., Piras, P., Kotsakis, T., 2006. Detection of Plio-Quaternary large mammalcommunities of Italy. An integration of fossil faunas, biochronology and simi-larity. Quat. Sci. Rev. 25, 846e854.
Rogers, L.L., Mech, L.D., 1981. Interactions of wolves and black bears in northeasternMinnesota. J. Mammol. 62 (2), 434e436.
Rook, L., 1994. The Plio-Pleistoceen old world Canis (Xenocyon) ex. gr. falconeri. Boll.Soc. Paleontol. Ital. 33 (1), 71e82.
Rook, L., Torre, D., 1996a. The wolf-event in western Europe and the beginning ofthe late Villafranchian. N. Jahrb. Geol. Palaeont. Mh. 8, 495e501.
Rook, L., Torre, D., 1996b. The latest Villafranchian e early Galerian small dogs of theMediterranean area. Acta Zool. Crac. 39 (1), 427e434.
Rook, L., Martinez-Navarro, B., 2010. Villafranchian: the long story of a Plio-Pleistocene European large mammal biochronological unit. Quat. Int. 219 (1e2), 134e144.
Rosenweig, M.L., 1968. The strategy of body size in mammalian carnivores. Am.Midl. Nat. 80 (2), 299e315.
Samuels, J.X., Meachen, J.A., Sakai, S.A., 2012. Postcranial morphology and the lo-comotor habits of living and extinct carnivorans. J. Morphol. 274, 121e146.
Sardella, R., Palombo, M.R., 2007. The Pliocene-Pleistocene boundary: which sig-nificance for the so called ‘Wolf Event’? Evidences from Western Europe.Quaternaire 18 (1), 65e71.
Shackleton, N.J., Berger, A., Peltier, W.R., 1990. An alternative astronomical calibra-tion of the lower Pleistocene timescale based on ODP Site 677. Trans. R. Soc.Edinb. Earth Sci. 81, 251e261.
Schreve, D.C., 1997. Mammalian Biostratigraphy of the Later Middle Pleistocene inBritain (Ph.D thesis). University College London, UK.
Schreve, D., 2001. Differentiation of the British late Middle Pleistocene interglacials:the evidence from mammalian biostratigraphy. Quat. Sci. Rev. 20, 1693e1705.
Please cite this article in press as: Flower, L.O.H., Schreve, D.C., An invesQuaternary Science Reviews (2014), http://dx.doi.org/10.1016/j.quascirev
Schreve, D., Howard, A., Currant, A., Brooks, S., Buteux, S., Coope, G.R., Crocker, B.,Field, M., Greenwood, M., Greig, J., Toms, P., 2013. A Middle Devensian woollyrhinoceros from Whitemoor Haye Quarry, Staffordshire (UK): palae-oenvironmental context and significance. J. Quat. Sci. 28 (2), 118e130.
Schutt, G., 1973. Revision der Cuon und Xenocyon-Funde (Canidae, Mammalia) ausden altpleistozänen Mosbacher Sanden (Wiesbaden, Hessen). Mainz. Natur-wiss. Arch. 12, 49e77.
Sotnikova, M., 2001. Remains of Canidae from the Lower Pleistocene site ofUntermassfeld. In: Kahlke, R.-D. (Ed.), Das Plestozan von Untermassfled beiMeiningen (Thuringen). Teil 2, vol. 40(2). Römisch-germanisches Zentralmu-seum, pp. 607e632. Monographien.
Sotnikova, M., Rook, L., 2010. Dispersal of the Canini (Mammalian, Canidae: Caninae)across Eurasia during the LateMiocene to Early Pleistocene. Quat. Int. 212, 86e97.
Stuart, A.J., 1995. Insularity and Quaternary vertebrate faunas in Britain and Ireland.In: Preece, R.C. (Ed.), Island Britain: a Quaternary Perspective. Geological SocietySpecial Publication, vol. 96, pp. 111e125.
Thenius, E., 1954. Die Caniden (mammalia) aus dem Altquartär von Hundsheim(Niederösterreich) nebst Bemerkungen sur Stammesgeschichte der GattungCuon. N. Jahrb. Geol. Paläontologie, Abh. 99, 230e286.
Torre, D., 1979. The Ruscinian and the Villafranchian dogs of Europe. Boll. Soc.Paleontol. Ital. 18 (2), 162e165.
Turner, A., 1981. Aspects of the Palaeoecology of Large Predators, Including Man,During the British Upper Pleistocene, With Particular Emphasis on Predator-prey Relationships (Ph.D. thesis). University of Sheffield, UK.
Turner, A., 1995. Evidence for Pleistocene contact between the British Isles and theEuropean continent based on distributions of larger carnivores. In: Preece, R.C.(Ed.), Island Britian: a Quaternary Perspective. Geol. Soc. Special Publication,vol. 96, pp. 141e149.
Turner, A., 2009. The evolution of the guild of large Carnivora of the British Islesduring the Middle and Late Pleistocene. J. Quat. Sci. 24 (8), 991e1005.
Van Ballenberghe, V., Erickson, A.W., Byman, D., 1975. Ecology of the timber wolf inNortheastern Minnesota. Wildl. Monogr. 43, 3e43.
Van Valkenburgh, B., 1988a. Trophic diversity in past and present guilds of largepredatory mammals. Paleobiology 14 (2), 155e173.
Van Valkenburgh, B., 1988b. Incidence of tooth breakage among large, predatorymammals. Am. Nat. 131 (2), 291e302.
Van Valkenburgh, B., 1989. Carnivore dental adaptations and diet: a study of trophicdiversity within guilds. In: Gittleman, J.L. (Ed.), Carnivore Behaviour, Ecologyand Evolution. Cornell University Press, pp. 410e436.
Van Valkenburgh, B., 1990. Skeletal and dental predictors of body mass in carni-vores. In: Damuth, J., MacFadden, B.J. (Eds.), Body Size in Mammalian Paleobi-ology: Estimation and Biological Implications. Cambridge University Press,pp. 181e205.
Van Valkenburgh, B., 1991. Iterative evolution of hypercarnivory in canids (Mam-malia: Carnivora): evolutionary interactions among sympatric predators.Paleobiology 17 (4), 340e362.
Van Valkenburgh, B., 1996. Feeding behaviour in free-ranging, large African carni-vores. J. Mammol. 71 (1), 240e254.
Van Valkenburgh, B., Hertel, F., 1993. Tough times at La Brea: tooth breakage in largecarnivores of the Late Pleistocene. Science 261 (5120), 456e459.
Van Valkenburgh, B., Koepfli, K.-P., 1993. Cranial and dental adaptations to preda-tion in canids. In: Dunstone, N., Gorman, M.L. (Eds.), Mammals as Predators, vol.65. Symp. Zool. Soc. Lond, pp. 15e37.
Van Valkenburgh, B., Wang, X., Damuth, J., 2004. Cope’s Rule, hypercarnivory, andextinction in North American Canids. Science 306 (101), 101e104.
Van Valkenburg, B., 2007. Déjà vu: the evolution of feeding morphologies in theCarnivora. In: Annual Meeting of the Society for Integrative and ComparativeBiology, Phoenix, Arizona, USA. Integrative and Comparative Biology. OxfordUniversity Press, pp. 1e17.
Voigt, D.R., Kolenosky, G.B., Pimlott, D.H., 1976. Changes in summer foods of wolvesin central Ontario. J. Wildl. Manag. 40 (4), 663e668.
von den Driesch, A., 1976. A Guide to the Measurement of Animal Bones fromArchaeological Sites. Peabody Museum of Archaeology and Ethnology, HarvardUniversity, Cambridge, U.S.A.
Vos, J., 2000. Food habits and livestock depredation of two Iberian wolf packs (Canislupus signatus) in the north of Portugal. J. Zool. Lond. 251, 457e462.
Werdelin, L., 1989. Constraint and adaptation in the bone-cracking canid Osteo-borus (Mammalia: Canidae). Paleobiology 15 (4), 387e401.
Werdelin, L., Lewis, M.E., 2005. Plio-Pleistocene carnivora of eastern Africa: speciesrichness and turnover patterns. Zool. J. Linn. Soc. 144, 121e144.
Zimov, S.A., Zimov, N.S., Tikhonov, A.N., Chapin III, F.S., 2012. Mammoth steppe: ahigh-productivity phenomenon. Quat. Sci. Rev. 57, 26e45.
tigation of palaeodietary variability in European Pleistocene canids,.2014.04.015





























