Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
www.elsevier.com/locate/ymeth
Methods 44 (2008) 229–234
Apoptosis-associated mitochondrial outer membranepermeabilization assays
Damien Arnoult *
INSERM U 542, Universite Paris-Sud, Hopital Paul Brousse, Batiment Lavoisier, 14 avenue Paul Vaillant Couturier, 94807 Villejuif cedex, France
Accepted 10 November 2007
Abstract
Following most cell death signals, pro-apoptotic Bcl-2 members as Bax and Bak are activated and oligomerize into the mitochondriaouter membrane, triggering its permeabilization and release into the cytosol of soluble apoptogenic factors such as cytochrome c involvedin caspase activation. Thus, in many studies focused on apoptosis, cytochrome c release within cells is frequently examined to assess Bax/Bak activation and mitochondrial outer membrane permeabilization. In addition, cytochrome c release can also be investigated in vitro infunctional mitochondria that have been isolated from cultured cells, offering a number of advantages. Here, protocols for measuringcytochrome c release from intact cells as well as from isolated mitochondria is detailed. Finally, assays to investigate Bax/Bak activationand olimerization are also presented.� 2007 Elsevier Inc. All rights reserved.
Keywords: Apoptosis; Mitochondria; Permeabilization; Cytochrome c; Bcl-2 family
1. Introduction
Apoptosis, is a cellular suicide program essential fordevelopment and adult tissue homeostasis in all metazoananimals. The stereotypical death throes of a cell undergo-ing apoptosis include DNA fragmentation, nuclear con-densation, cell shrinkage, plasma membrane blebbing,phosphatidylserine externalization among other events[1]. These features associated with the orderly dismantlingof the cell during apoptosis are orchestrated by the activityof a family of cysteine proteases called caspases [2].
A mitochondria-dependent step, involving mitochon-drial outer membrane permeabilization (MOMP), is associ-ated with most pro-apoptotic stimuli. This process iscontrolled by both pro- and anti-apoptotic members ofthe Bcl-2 family and leads to the release of mitochondrialapoptogenic factors such as cytochrome c, Smac/DIABLO,and Omi/HtrA2 into the cytosol [3,4]. The mechanisms bywhich the pro-apoptotic Bcl-2 family members induce the
1046-2023/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.ymeth.2007.11.003
* Fax: +33 0 1 45 59 53 43.E-mail address: [email protected]
release of these mitochondrial proteins remain controver-sial [5,6]. One model emphasizes rupture of the mitochon-drial outer membrane as a consequence of mitochondrialswelling after the opening of the permeability transitionpore (PTP). According to another model, pro-apoptoticBcl-2 proteins like Bax/Bak induce a selective process ofMOMP through the formation of channels or pores afteroligomerization, allowing the release of proteins localizedwithin the intermembrane space as cytochrome c, Smac/DIABLO, and Omi/HtrA2. Once released into the cytosol,cytochrome c, a key mitochondrial apoptogenic factor [7],triggers caspase-9 activation by binding and activating theapoptosis protease activating factor-1 (Apaf-1) in a multi-protein complex called the apoptosome [8]. Next, activatedcaspase-9 activates effector caspases such as caspase-3 thatcleave several substrates to induce the apoptotic phenotype[1,2]. The role of Smac/DIABLO and Omi/HtrA2 is moresubtle because they potentiate caspase activation by bind-ing inhibitor of apoptosis proteins (IAPs), thus blockingtheir caspase inhibitory activity [9].
Here, a number of assays for measuring apoptosis-asso-ciated MOMP are described. These include analysis of

Cytochrome c
Smac/DIABLO
Omi/HtrA2
Actin
ActD18 24-
Time (hrs)
Cyto
solic
Fractio
nH
eavyM
emb
rane
Fractio
n
STS18 24
Cytochrome c
Smac/DIABLO
Omi/HtrA2
Cox IV
Fig. 1. Analysis of the release of cytochrome c, Smac/DIABLO and Omi/HtrA2 during apoptosis using a digitonin-based subcellular fractionationtechnique. HeLa cells were treated for 18 and 24 h with 1 lM stauro-sporine (STS) or 10 lM actinomycin D (ActD), and cytosolic and heavymembrane fractions were analyzed by immunoblotting for the presence ofcytochrome c, Smac/DIABLO, and Omi/HtrA2. As loading controls,actin was used for the cytosolic fraction and the mitochondrial proteinCox IV for the heavy membrane fraction. Subcellular fractionation wasperformed as described in the text.
230 D. Arnoult / Methods 44 (2008) 229–234
cytochrome c release within intact cells as a consequence ofBax/Bak-mediated MOMP, isolation of mitochondria andan in vitro assay for cytochrome c release, and assessmentof Bax/Bak activation and oligomerization into the mito-chondria outer membrane.
2. Description of the methods
2.1. Analysis of cytochrome c release using a digitonin-based
subcellular fractionation technique
After apoptosis induction, confluent HeLa cells or mur-ine embryonic fibroblasts (MEFs) in T75 culture flask areharvested in phosphate-buffered saline (PBS; pH 7.4) con-taining 1 mM EDTA, then centrifuged at 750g for 5 min,washed in PBS, and digitonin-permeabilized for 5 min onice after resuspension of the cell pellet in 400 ll of cytosolicextraction buffer (CEB: 250 mM sucrose, 70 mM KCl,137 mM NaCl, 4.3 mM Na2HPO4, 1.4 mM KH2PO4 (pH7.2), with 300 lg/ml digitonin, and the protease inhibitorcocktail complete (Boehringer Mannheim)). For lymphoidcell lines as Jurkat or CEM, cells are resuspended in CEBcontaining 200 lg/ml digitonin at a density of 3.107/ml.After 5 min incubation on ice, plasma membrane perme-abilization of cells is confirmed by staining in a 0.2% trypanblue solution. Cells are then centrifuged at 1000g for 5 minat 4 �C. The supernatants (cytosolic fractions) are savedand the pellets solubilized in the same volume of mitochon-drial lysis buffer (MLB: 50 mM Tris, pH 7.4, 150 mMNaCl, 2 mM EDTA, 2 mM EGTA, 0.2% Triton X-100,0.3% NP-40) supplemented with protease inhibitor cocktailcomplete, followed by centrifugation at 10,000g for 10 minat 4 �C. After centrifugation, supernatants are kept as theheavy membrane fractions enriched for mitochondria. Sol-uble and heavy membrane fractions (30 and 15 lg, respec-tively) are separated by SDS–PAGE (10–20% Tricine gels;NOVEX) and transferred to nitrocellulose membranes.Importantly, we observed that the detection of cytochromec is optimal when a tricine gel is used for the immunoblot-ting. After blocking nonspecific sites for 1 h at room tem-perature with 5% nonfat milk in PBS supplemented with0.1% Tween 20, nitrocellulose membranes are incubated2 h at room temperature, or else overnight at 4 �C, withmouse anti-cytochrome c (BD Pharmingen, clone7H8.2C12) (dilution 1:2000 in PBS with 5% nonfat milksupplemented with 0.1% Tween 20). To also investigatethe release of Smac/DIABLO and Omi/HtrA2, two othermitochondrial apoptogenic factors that are co-releasedwith cytochrome c following MOMP [10], rabbit poly-clonal antibodies anti-Smac/DIABLO (ProScience)(1:1000) and anti-Omi/HtrA2 (R&D Systems) (1:1000)are used. Because the molecular masses of cytochrome c
(15 kDa), Smac/DIABLO (23 kDa), and Omi/HtrA2(36 kDa) are different, and because these antibodies donot cross-react with other proteins, the three primary anti-bodies can be mixed together. Next, immunoreactive pro-teins are visualized using horseradish peroxidase-linked
goat anti-mouse or anti-rabbit antibodies and enhancedchemiluminescence. Lastly, to confirm equal loading andtransfer of the cytosolic and heavy membrane fractions,the nitrocellulose membranes are subsequently strippedand reprobed for actin using a mouse monoclonal antibody(Sigma, clone AC40) (1:4000) or for Cox IV, a mousemonoclonal antibody (Molecular Probes, clone 10G8)(0.5 lg/ml), respectively.
Once the optimal concentration of digitonin has beendetermined to permeabilize the plasma membrane of thestudied cell type, this digitonin-based subcellular fraction-ation technique is very simple to investigate cytochrome c
release during apoptosis. For example, this method wasused in Fig. 1 to study the release of cytochrome c,Smac/DIABLO and Omi/HtrA2 in HeLa cells induced toundergo apoptosis with the pro-apoptotic drugs stauro-sporine and actinomycin D. Following staurosporine oractinomycin D treatment, cytochrome c, Smac/DIABLO,and Omi/HtrA2 are found in the cytosolic fraction conse-quently to their efflux from the heavy membrane fractionenriched for mitochondria.

D. Arnoult / Methods 44 (2008) 229–234 231
2.2. Analysis of cytochrome c release using a subcellular
fractionation after mechanical rupture of the plasma
membrane
At different times after induction of apoptosis, confluentHeLa cells or MEFs in T75 culture flask are harvested withPBS containing 1 mM EDTA, then centrifuged at 750g for5 min, washed in PBS, and the cell pellet is resuspended in400 ll of ice cold isotonic mitochondrial isolation buffer(MIB: 210 mM mannitol, 70 mM sucrose, 1 mM EDTA,and 10 mM Hepes (pH 7.5), supplemented with proteaseinhibitor cocktail complete). Next, cells are broken with aDounce homogenizer (Wheaton) using the ‘‘tight’’ piston.Number of strokes depends of the used cell type. Forinstance 50–60 strokes are required for HeLa cells, 30–40strokes for MEFs. Plasma membrane rupture of cells isconfirmed by staining in a 0.2% trypan blue solution. Next,samples are transferred to Eppendorf centrifuge tubes andcentrifuged at 500g for 5 min at 4 �C to eliminate nucleiand unbroken cells. The resulting supernatant is centri-fuged at 10,000g for 30 min at 4 �C to obtain the heavymembrane fraction enriched for mitochondria. This super-natant is further centrifuged at 100,000g for 1 h at 4 �C toyield the light membrane pellet and the final soluble cyto-solic fraction. The heavy membrane fraction material isresuspended in MIB supplemented with 1% Triton X-100.Cytosolic and heavy membrane fractions are then analyzedby immunoblotting as described above. This method ofsubcellular fractionation has been used successfully in pre-vious studies [10–12].
2.3. Isolation of mitochondria and in vitro assay for
cytochrome c release
Cytochrome c release can be also investigated in vitro infunctional mitochondria that have been isolated from cul-tured cells. To purify mitochondria, we basically use cellscontaining a very dense mitochondrial network (HeLacells, MEFs etc) but this method can also be used for othercell lines. Mitochondria are isolated employing a sucrosedensity gradient centrifugation. Cells grown in confluencein 10 T175 culture flasks are harvested with PBS containing1 mM EDTA, centrifuged at 750g for 10 min, washed inPBS, and resuspended in 5 ml isotonic MIB supplementedwith protease inhibitors. Next, cells are broken on ice by 10passages through a 25G1 0.5 · 25 needle fitted on a 10 mlsyringe, and the suspension is centrifuged at 2000g in anEppendorf centrifuge at 4 �C. This procedure is repeateduntil almost all cells are broken. Supernatants from eachstep are pooled before centrifugation at 13,000g at 4 �Cfor 10 min. The resulting pellet is resuspended in 2 ml ofMIB and layered on top of a discontinuous sucrose gradi-ent consisting of 20 ml of 1.2 M sucrose in 10 mM Hepes,(pH 7.5), 1 mM EDTA, and 0.1% BSA on top of 17 mlof 1.6 M sucrose in 10 mM Hepes, (pH 7.5), 1 mM EDTA,and 0.1% BSA. Then, samples are centrifuged at27,000 rpm for 2 h at 4 �C in a Beckman SW28 rotor.
Mitochondria are recovered at the 1.6–1.2 M sucrose inter-face, washed in MIB (500 ll of the interface in 1 ml MIB),centrifuged at 13,000g at 4 �C for 10 min, and resuspendedin 100–150 ll MIB. Finally protein concentration is esti-mated. From 10 T175 of HeLa cells or MEFs in conflu-ence, with this method at least 1 mg of mitochondria ispurified in our hands.
To study in vitro cytochrome c release in isolated mito-chondria, MOMP is induced with recombinant tBid (R&DSystems), a BH3-only member of the Bcl-2 family [13] thatrequires either Bax or Bak to trigger MOMP [14]. Hence,mitochondria (50 lg) are incubated in the presence orabsence of increased concentration of tBid (0.001, 0.01,0.1, 1, 10, and 100 nM) in 200 ll of KCl buffer (125 mMKCl, 4 mM MgCl2, 5 mM Na2HPO4, 5 mM succinate,0.5 mM EGTA, 15 mM Hepes–KOH (pH 7.4), 5 lM rote-none) for 15 min at 30 �C and then centrifuged for 5 min at13,000g at 4 �C. In time course experiments, mitochondriaare incubated with 10 nM recombinant tBid and pelletedby centrifugation after 0, 5, 10 or 15 min. Mitochondrialpellets corresponding to 5 lg proteins and the correspond-ing volume of the supernatant fractions are resolved bySDS–PAGE (10–20% Tricine gels; NOVEX) and trans-ferred to a nitrocellulose membrane. Their respective con-tents of cytochrome c, Smac/DIABLO and Omi/HtrA2are estimated by immunoblotting as described above.Equal loading of the mitochondrial pellet is verified usingan antibody against VDAC (Calbiochem, clone 31HL)(1:6000). Fig. 2 illustrates the in vitro release of cytochromec, Smac/DIABLO and Omi/HtrA2 in isolated mitochon-dria from HeLa cells incubated with tBid. Note thatMOMP can also be induced in vitro by incubating isolatedmitochondria with recombinant oligomeric Bax [15,16].
2.4. Analysis of Bax/Bak activation
Following most apoptotic signals, the pro-apoptoticproteins Bax and Bak are activated, leading to a conforma-tional change and exposure of their N-terminus [5,6,13].This conformational change can be assessed by immunoflu-orescence using specific antibodies raised against the N-ter-minal region of Bax or Bak. For the detection of the Bax orBak active form, mouse monoclonal anti-Bax (clone 6A7,Sigma) or anti-Bak (clone Ab2, Oncogene Research Prod-ucts), respectively, are used. In parallel, to confirm Baxand/or Bak activation followed by MOMP and cyto-chrome c release, cytochrome c can be visualized with asheep polyclonal antibody (Sigma).
Next is a protocol for immunofluorescence staining, asshown in Fig. 3, illustrating Bax or Bak activation andcytochrome c release during apoptosis. After staurosporinetreatment, HeLa cells cultured in Labtek chamber arewashed 3 times in PBS followed by fixation in freshly pre-pared 4% paraformaldehyde in PBS for 10 min. The fixedcells are washed three times in PBS for 10 min each fol-lowed by permeabilization in 0.15% Triton X-100 in PBSfor 15 min. The cells are then blocked for 60 min in block-

0 0.00
10.
010.
1 1 10 100
VDAC
Smac/DIABLO
Omi/HtrA2
Cytochrome c
Cytochrome c
Smac/DIABLO
Omi/HtrA2
tBid (nM) Incubation time (min)10- +
15- +
5- +
0- + tBid (10 nM)
Fig. 2. In vitro release of cytochrome c, Smac/DIABLO, and Omi/HtrA2. Mitochondria isolated from HeLa cells were incubated for 15 min at 30 �C withincreasing concentrations (in nM) of recombinant tBid (left panel). Mitochondrial and supernatant fractions were then resolved by SDS–PAGE andimmunoblotted with antibodies against the indicated proteins. In the right panel, mitochondria were incubated with (+) or without (�) 10 nM tBid at30 �C, and the mitochondrial pellet and supernatant were analyzed by immunoblotting at different time points. Equal protein loading of the mitochondrialpellet lanes was monitored using anti-VDAC antibodies.
232 D. Arnoult / Methods 44 (2008) 229–234
ing buffer (2% bovine serum albumin in PBS) followed by a2 h incubation at room temperature, or overnight at 4 �C,with mouse monoclonal anti-Bax or anti-Bak antibody(1 lg/ml in blocking buffer) and a sheep polyclonal anti-body raised against cytochrome c (1:800 in blocking buf-fer). The cells are washed three times at 10 min each inblocking buffer followed by 1 h incubation with a Alexa594 goat anti-mouse antibody (1:500) (Molecular probe)for Bax or Bak and Alexa 488 donkey anti-sheep IgG(1:500) (Molecular probe) for cytochrome c. Cells are thenwashed three times at 10 min each in PBS, followed bystaining with 1 lg/ml DAPI and examined under a ZeissLSM 510 confocal microscope. Note that Bax/Bak activa-tion in lymphoid cell lines such as Jurkat cells can beassessed by flow cytometry after immunofluorescence.
2.5. Assay for Bax/Bak oligomerization
During apoptosis, Bax/Bak activation is also associatedwith their oligomerization into the mitochondria outermembrane [5,6]. Their oligomerization can be investigatedby protein cross-linking. Purified mitochondria incubatedwith recombinant tBid or heavy membrane fractionsenriched in mitochondria from apoptotic cells are resus-pended in MIB, and the cross-linker disuccinimidyl suber-nate (DSS) or bis(sulfosuccinimidyl) subernate (BS3) areadded from a 10-fold stock solution to a final concentra-tion of 2 mM. After incubation for 30 min at room temper-ature, the cross-linker is quenched by the addition of 1 MTris–HCl (pH 7.5) to a final concentration of 20 mM. After
quenching, the mitochondria or heavy membrane fractionsare separated by SDS–PAGE; Bax or Bak dimers, trimersand tetramers are observed after immunoblots with Bax(rabbit polyclonal N20, Santa Cruz) (1:1000) or Bak (rab-bit polyclonal N-term, Upstate Biotechnology) (1:1000)antibodies.
3. Concluding remarks
The critical step in studying cytochrome c release inintact cells is to determine the optimal concentration ofdigitonin or the optimal number of piston strokes requiredto permeabilize the cell plasma membrane in a small vol-ume of buffer. Mitochondria are well protected within thecells, so that risks of mitochondrial outer membrane rup-ture are low. To permeabilize plasma membrane using dig-itonin treatment, the use of 50 lg/ml to 400 lg/ml ofdigitonin is recommended, depending of the studied celltype. For mechanical rupture of the plasma membranewith a Dounce homogenizer, 30–150 strokes are required,depending upon the cell size. Once permeabilization condi-tions have been optimized, it is straightforward to investi-gate cytochrome c release from the mitochondria into thecytosol by immunoblotting cytosolic and heavy membranefractions.
To purify mitochondria, it is important to begin thepurification with many cultured cells to obtain sufficientamounts of isolated mitochondria. An alternative to isola-tion of mitochondria from cells in culture is to isolate func-tional mitochondria from mouse liver or muscle tissues, as
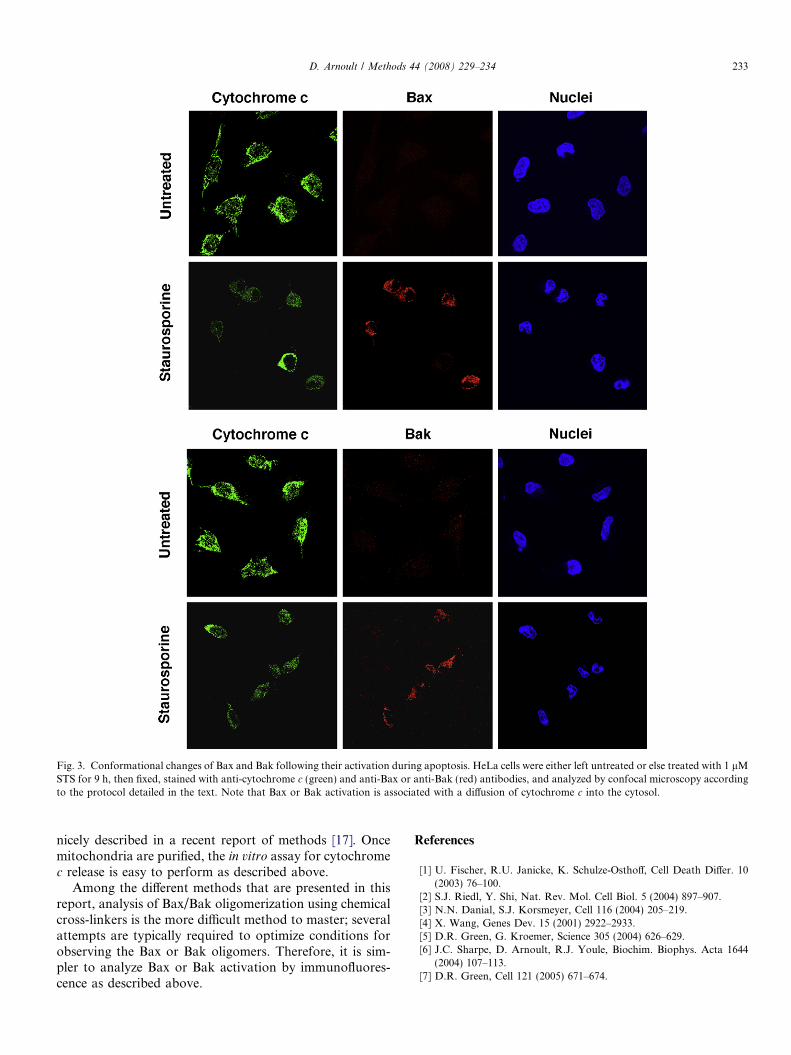
Fig. 3. Conformational changes of Bax and Bak following their activation during apoptosis. HeLa cells were either left untreated or else treated with 1 lMSTS for 9 h, then fixed, stained with anti-cytochrome c (green) and anti-Bax or anti-Bak (red) antibodies, and analyzed by confocal microscopy accordingto the protocol detailed in the text. Note that Bax or Bak activation is associated with a diffusion of cytochrome c into the cytosol.
D. Arnoult / Methods 44 (2008) 229–234 233
nicely described in a recent report of methods [17]. Oncemitochondria are purified, the in vitro assay for cytochromec release is easy to perform as described above.
Among the different methods that are presented in thisreport, analysis of Bax/Bak oligomerization using chemicalcross-linkers is the more difficult method to master; severalattempts are typically required to optimize conditions forobserving the Bax or Bak oligomers. Therefore, it is sim-pler to analyze Bax or Bak activation by immunofluores-cence as described above.
References
[1] U. Fischer, R.U. Janicke, K. Schulze-Osthoff, Cell Death Differ. 10(2003) 76–100.
[2] S.J. Riedl, Y. Shi, Nat. Rev. Mol. Cell Biol. 5 (2004) 897–907.[3] N.N. Danial, S.J. Korsmeyer, Cell 116 (2004) 205–219.[4] X. Wang, Genes Dev. 15 (2001) 2922–2933.[5] D.R. Green, G. Kroemer, Science 305 (2004) 626–629.[6] J.C. Sharpe, D. Arnoult, R.J. Youle, Biochim. Biophys. Acta 1644
(2004) 107–113.[7] D.R. Green, Cell 121 (2005) 671–674.

234 D. Arnoult / Methods 44 (2008) 229–234
[8] S.J. Riedl, G.S. Salvesen, Nat. Rev. Mol. Cell Biol. 8 (2007) 405–413.[9] D.L. Vaux, J. Silke, Biochem. Biophys. Res. Commun. 304 (2003)
499–504.[10] D. Arnoult, B. Gaume, M. Karbowski, J.C. Sharpe, F. Cecconi, R.J.
Youle, EMBO J. 22 (2003) 4385–4399.[11] D. Arnoult, A. Grodet, Y.J. Lee, J. Estaquier, C. Blackstone, J. Biol.
Chem. 280 (2005) 35742–35750.[12] D. Arnoult, N. Rismanchi, et al., Curr. Biol. 15 (2005) 2112–2118.
[13] S.N. Willis, J.M. Adams, Curr. Opin. Cell Biol. 17 (2005) 617–625.
[14] M.C. Wei, W.X. Zong, et al., Science 292 (2001) 727–730.[15] B. Antonsson, S. Montessuit, S. Lauper, R. Eskes, J.C. Martinou,
Biochem. J. 345 (2) (2000) 271–278.[16] D. Arnoult, P. Parone, J.C. Martinou, B. Antonsson, J. Estaquier,
J.C. Ameisen, J. Cell. Biol. 159 (2002) 923–929.[17] C. Frezza, S. Cipolat, L. Scorrano, Nat. Protoc. 2 (2007) 287–295.





























