Embed Size (px)
Citation preview

Aproximació als mecanismes moleculars
implicats en la regulació de la síndrome de fugida de l’ombra en Arabidopsis
Marçal Gallemí Rovira
ADVERTIMENT. La consulta d’aquesta tesi queda condicionada a l’acceptació de les següents condicions d'ús: La difusió d’aquesta tesi per mitjà del servei TDX (www.tdx.cat) i a través del Dipòsit Digital de la UB (diposit.ub.edu) ha estat autoritzada pels titulars dels drets de propietat intel·lectual únicament per a usos privats emmarcats en activitats d’investigació i docència. No s’autoritza la seva reproducció amb finalitats de lucre ni la seva difusió i posada a disposició des d’un lloc aliè al servei TDX ni al Dipòsit Digital de la UB. No s’autoritza la presentació del seu contingut en una finestra o marc aliè a TDX o al Dipòsit Digital de la UB (framing). Aquesta reserva de drets afecta tant al resum de presentació de la tesi com als seus continguts. En la utilització o cita de parts de la tesi és obligat indicar el nom de la persona autora. ADVERTENCIA. La consulta de esta tesis queda condicionada a la aceptación de las siguientes condiciones de uso: La difusión de esta tesis por medio del servicio TDR (www.tdx.cat) y a través del Repositorio Digital de la UB (diposit.ub.edu) ha sido autorizada por los titulares de los derechos de propiedad intelectual únicamente para usos privados enmarcados en actividades de investigación y docencia. No se autoriza su reproducción con finalidades de lucro ni su difusión y puesta a disposición desde un sitio ajeno al servicio TDR o al Repositorio Digital de la UB. No se autoriza la presentación de su contenido en una ventana o marco ajeno a TDR o al Repositorio Digital de la UB (framing). Esta reserva de derechos afecta tanto al resumen de presentación de la tesis como a sus contenidos. En la utilización o cita de partes de la tesis es obligado indicar el nombre de la persona autora. WARNING. On having consulted this thesis you’re accepting the following use conditions: Spreading this thesis by the TDX (www.tdx.cat) service and by the UB Digital Repository (diposit.ub.edu) has been authorized by the titular of the intellectual property rights only for private uses placed in investigation and teaching activities. Reproduction with lucrative aims is not authorized nor its spreading and availability from a site foreign to the TDX service or to the UB Digital Repository. Introducing its content in a window or frame foreign to the TDX service or to the UB Digital Repository is not authorized (framing). Those rights affect to the presentation summary of the thesis as well as to its contents. In the using or citation of parts of the thesis it’s obliged to indicate the name of the author.

Aproximació als mecanismes moleculars implicats en la regulació de la síndrome de
fugida de l’ombra en Arabidopsis.
Marçal Gallemí Rovira2013

Centre de Recerca en Agrigenòmica Universitat de Barcelona
Departament de genètica molecular Facultat de farmàcia
Departament de Bioquímica i Biologia Molecular
Doctorat en biotecnologia
Aproximació als mecanismes moleculars
implicats en la regulació de la síndrome de
fugida de l’ombra en Arabidopsis.
Memòria presentada per Marçal Gallemí Rovira per optar al títol de doctor per la
Universitat de Barcelona
Firmat per
Director de tesis Tutor
Dr. Jaime F. Martínez-García Dr. Albert Ferrer Prats
Autor Marçal Gallemí Rovira
Barcelona, 2013


La recerca ha de ser un joc, perquè només jugant som feliços.
Ramon Margalef (el 1.984)


Agraïments
A tota la gent que d’una o altra manera hi ha contribuït, de
tot cor moltes gràcies.

Està béeeeeee, si no dic res més m’arrisco a patir accidents desafortunats, a que ningú
em presti ajuda jugant al Munchkin o ningú em vulgui canviar la pedra per ovelles al Catán. Per
tant, per no ser un marginat social (si no m’hi vaig quedar ja al renunciar al watsapp) expandiré
una mica els agraïments, però segur que no us puc agrair ni a la meitat de vosaltres la meitat
del que voldria, i això no és ni la meitat del que us mereixeu. En primer lloc voldria agrair a la
meva família, especialment pares i germà, pel seu suport, sempre fomentant que estudiés per
arribar a fer alguna cosa de profit a la vida (la tesi compta?). En segon, agrair als bons
professors (no tants com voldria) que m’han ensenyat no només coneixements, sinó a pensar i
ser crític, i que han sabut motivar el seu alumnat per seguir aprenent. En tercer, a tots els
amics, des dels companys d’institut, d’esports, de la universitat i d’aventures en general.
Perquè sense experiències com concerts de música a Girona, anar de paquet en una moto,
partits de futbol, bàsquet, tenis, frontó, tenis-taula, bàdminton, voleibol platja, sense reflexions
com perquè se la xupen els gossos, discussions absurdes sobre l’atzar de tirar els daus, sense
partides de rol, sense sopars al Milk, sense Warhammers, Blood-bowl, ni Mordheims, sense
dissabtes de cau, sense campaments, sense les travesses que ens plou, sense castells, sense
tornejos de futbol intercasteller, sense sopars d’agulles de merda, sense cantar havaneres a
classe, sense Tàrrega, sense els Gallifas, sense Aplecs, sense Quimifarres, sense festes
temàtiques a la Pensión Loli, sense la Secta, sense garrinades, sense Santes, sense callejeros,
sense les frikibiotrobades, sense la primera PCR, el primer clonatge, les primeres
transgèniques, el primer Northern, la primera qPCR, el primer Western, el primer ChIP, sense
les cançons al lab, els vídeos de tesis, els mails demanant enzims, les happy hour, les
converses de passadís i tantes altres coses que no puc posar, la vida no m’hagués fet qui sóc, i
molt probablement no estaria on sóc ara. Vosaltres, i el temps que hem compartit, m’heu fet
arribar aquí, gràcies per tots aquest moments. Com no, haig d’agrair als companys del CRAG i
especialment del LILA lab, ha estat un veritable plaer compartir feines i penúries amb tots i
totes vosaltres, molta sort. Per anar acabant, donar les infinites gràcies al director de tesi:
MOLTES GRÀCIES JAUME, has estat un gran mestre, ser el teu doctorant ha estat la feina
més fàcil del món. El penúltim agraïment va per a tots els autors citats i no citats (d’acord, haig
de reconèixer que alguns més que altres), perquè la ciència és un treball en comunitat, sense
les aportacions de la resta no es pot tirar endavant. Alguns treballs són font d’inspiració i guia
de com es pot fer bé la feina, ajudant a perseverar en moments durs, motivant a seguir per
arribar a entendre el per què dels successos observats. I per acabar, gràcies a la meva
companya de pis predilecte, la vida amb tu és més fàcil i bonica.

ÍNDEX PRÒLEG ....................................................................................................................................... 1
1. INTRODUCCIÓ ......................................................................................................................... 3
1.1. De la llavor a la planta: La llum i la fotomorfogènesis ............................................................ 3
1.2. Percepció de la llum: Els fotoreceptors i, més en concret, els fitocroms ............................... 4
1.3. Més enllà dels fitocroms: la transducció de senyal i la xarxa reguladora .............................. 8
1.4. Mecanismes moleculars dels fitocroms: la relació amor-odi amb els PIFs ......................... 10
1.5. Respostes a la llum regulades pels fitocroms: La síndrome de fugida de l’ombra (SAS) ... 15
1.6. Aspectes generals de la SAS: Quan créixer esdevé la prioritat .......................................... 17
1.7. Aspectes moleculars de la SAS: Múltiples personatges per una resposta complexa ......... 20
1.8. Relació de la SAS amb altres rutes ..................................................................................... 24
1.9. I això on va a parar? Aplicacions biotecnològiques ............................................................. 30
2. OBJECTIUS ............................................................................................................................ 33
3. RESULTATS ........................................................................................................................... 35
3.1. Capítol 1: Estudi de la relació estructura-funció d’ATHB4 ................................................... 37
3.1.1. Introducció ......................................................................................................................... 37
3.1.1.1. La família HD-Zip: Components del desenvolupament ................................................. 37
3.1.1.2. ATHB4 i la companyia autoreprimida ............................................................................ 40
3.1.2. Resultats ........................................................................................................................... 43
3.1.2.1 Disseny i obtenció de les construccions derivades d’ATHB4 ......................................... 43
3.1.2.2. Activitat biològica de les truncacions: Respostes fisiològiques ..................................... 46
3.1.2.3. Activitat biològica de les truncacions: Respostes moleculars ........................................ 48
3.1.2.4. Nous horitzons: Aprofundint en l’Nt................................................................................ 51
3.1.2.5. ATHB4 uneix realment al DNA? Seqüència d’unió i dianes directes ............................. 56
3.2. Capítol 2: Estudi del paper de les NUPs en la regulació de la SAS .................................... 62
3.2.1. Introducció ......................................................................................................................... 62
3.2.1.1. Nous components reguladors de la SAS: Les nucleoporines ........................................ 62
3.2.1.2. DRACULA2 i el seu homòleg d’animals NUP98: Una NUP, moltes funcions ............... 67
3.2.2. Resultats. .......................................................................................................................... 70
3.2.2.1. Anàlisis del mutant dra2-1: Caracterització general i fisiològica. ................................... 70
3.2.2.2. Anàlisis del mutant dra2-1: Caracterització molecular. .................................................. 74
3.2.2.3. Altres NUPs: Caracterització fisiològica i molecular. ..................................................... 78
3.2.2.4. Creuaments per canvis d’ecotip. .................................................................................... 82
3.2.2.5. Creuaments entre NUPs: Dobles mutants. .................................................................... 85
3.2.2.6. Efectes en el transport nuclear. ..................................................................................... 87

4. DISCUSSIÓ ............................................................................................................................. 93
4.1. Estudi de la relació estructura-funció d’ATHB4. .................................................................. 95
4.1.1 La família HD-Zip: una activitat biològica plena d’interaccions. ......................................... 95
4.1.2 ATHB4 uneix al DNA i actua com a repressor de la transcripció. ..................................... 97
4.1.3 Estudi de la relació estructura-funció d’ATHB4. ................................................................ 98
4.2. Regulació de les NUPs. ..................................................................................................... 101
4.2.1. DRA2: caracteritzacions fisiològiques i moleculars. ....................................................... 101
4.2.2. DRA2 i el transport a través del NPC.............................................................................. 104
4.3. ATHB4 i DRA2 en la regulació de la SAS. ......................................................................... 105
5. CONCLUSIONS. ................................................................................................................... 109
6. MATERIALS I MÈTODES. .................................................................................................... 111
6.1. Material biològic. ................................................................................................................ 111
6.1.1. Material bacterià. ............................................................................................................. 111
6.1.1.1. Soques bacterianes. .................................................................................................... 111
6.1.1.2. Condicions de cultiu de bacteris. ................................................................................. 111
6.1.1.3. Obtenció de cèl·lules competents per xoc tèrmic. ....................................................... 112
6.1.1.4. Obtenció de cèl·lules electrocompetents. .................................................................... 112
6.1.1.5. Medis de cultiu bacterians. ........................................................................................... 114
6.1.1.6. Suplements dels medis de cultiu bacterians. ............................................................... 114
6.1.2. Material vegetal. .............................................................................................................. 114
6.1.2.1. Ecotips silvestres.......................................................................................................... 114
6.1.2.2. Línies transgèniques. ................................................................................................... 114
6.1.2.3. Línies mutants de pèrdua de funció. ............................................................................ 114
6.1.2.4. Creuaments. ................................................................................................................. 115
6.1.2.5. Condicions de cultiu a l’hivernacle i al fitotró. .............................................................. 116
6.1.2.6. Condicions de cultiu in vitro. ........................................................................................ 117
6.1.2.7. Medis de cultiu de plantes. ........................................................................................... 117
6.1.2.8. Suplements dels medis de cultiu de plantes. ............................................................... 117
6.2. Plàsmids. ............................................................................................................................ 117
6.2.1. Vectors comercials. ......................................................................................................... 117
6.2.2. Construccions i transformacions en planta. .................................................................... 118
6.3. Encebadors. ...................................................................................................................... 119
6.4. Metodologies. ..................................................................................................................... 121
6.4.1. Tècniques de biologia molecular d’àcids nucleics. ......................................................... 121
6.4.1.1 Obtenció de DNA plasmídic. ......................................................................................... 121
6.4.1.2. Reaccions de modificació del DNA. ............................................................................. 122
6.4.1.3. Transformació de cèl·lules competents d’E.coli per xoc tèrmic. .................................. 122
6.4.1.4. Transformació de cèl·lules competents d’A.tumefaciens per electroporació............... 122

6.4.1.5. Reacció en cadena de la polimerasa (PCR). ............................................................... 123
6.4.1.6. Separació de DNA en gels d’agarosa .......................................................................... 124
6.4.1.7. Purificació de DNA. ...................................................................................................... 124
6.4.1.8. Seqüenciació del DNA ................................................................................................. 124
6.4.1.9. Clonatge de fragments de DNA amplificats per PCR .................................................. 125
6.4.1.10. Obtenció d’RNA total d’arabidopsis ........................................................................... 125
6.4.1.11. Quantificació d’àcids nucleics .................................................................................... 126
6.4.1.12. Retrotranscripció ........................................................................................................ 126
6.4.1.13. PCR quantitativa. ....................................................................................................... 126
6.4.1.14. Immunoprecipitació de cromatina (ChIP). .................................................................. 127
6.4.1.15. Anàlisi de l’RNA per northern-blot .............................................................................. 130
6.4.1.16. Anàlisi in situ de poliA+-RNA. ..................................................................................... 133
6.4.2. Tècniques de biologia molecular de proteïnes ............................................................... 134
6.4.2.1. Obtenció d’extractes proteics ....................................................................................... 134
6.4.2.2. Separació electroforètica de proteïnes. ....................................................................... 134
6.4.2.3. Anàlisi de proteïnes per transferència i immunodectecció. ......................................... 134
6.4.3. Tècniques bioquímiques ................................................................................................. 136
6.4.3.1. Anàlisi dels pigments isoprenoides (clorofil·les i carotenoides). .................................. 136
6.4.4. Mètodes de plantes ......................................................................................................... 136
6.4.4.1. Esterilització de llavors ................................................................................................. 136
6.4.4.2. Sembra de llavors. ....................................................................................................... 136
6.4.4.3. Obtenció de plantes transgèniques ............................................................................. 137
6.4.4.4. Anàlisi molecular de les plantes mutants ..................................................................... 138
6.4.4.5. Tractaments i assajos en planta .................................................................................. 139
6.4.4.6. Tractaments amb reguladors del creixement, hormones o dexametasona ................. 140
6.5. Eines i recursos bioinformàtics .......................................................................................... 142
7. BIBLIOGRAFIA ..................................................................................................................... 143
8. ABREVIACIONS ................................................................................................................... 157


1
PRÒLEG
La vida... que complexa i bonica pot ser!! Pensar que tot va sorgir per una mescla de
substàncies que s’han anat generant aleatòriament... Una sopa de partícules subatòmiques,
que s’ajunten per formar partícules atòmiques, que es mesclen per conformar àtoms amb
diferents percentatges d’estabilitat, que per atzar van col·lisionant i de tant en tant, es forma
alguna molècula, que entre elles poden, al seu torn, reaccionar per formar molècules més
complexes, com peces de lego que es van encaixant sense saber on arribaran. De tant en tant,
peces que es barallen i es separen, que desfan els seus lligams per tornar a començar. Potser
això és el més important a la vida, provar-ho, i tornar-ho a provar. Fins que... voilà! Surt alguna
cosa aprofitable, amb una mica de sort, potser útil o important. Aminoàcids, bases
nitrogenades, àcids orgànics... tot combinat amb l’aparent simplicitat de sals minerals o aigua.
Una combinació màgica que porta al funcionament més meravellós que podem trobar, una
evolució lenta, produïda al llarg de molt de temps, però que arriba on ni la imaginació ens
permet somiar. I tot gràcies a la principal font d’energia que ha facilitat aquestes reaccions, la
llum del Sol que ens ha donat tot el que som. Ara, encuriosits com els primers homínids,
seguim mirant al voltant per trobar-hi explicacions lògiques, per poder entendre com funciona el
misteri de la vida. Quina és la nostra història, la història de les petites molècules que es van
convertir en vida. En la meva humil posició intentaré doncs aportar el meu gra de sorra per
aquesta platja, sovint més aviat desert, amb l’únic objectiu d’acostar la humanitat al
coneixement, i que aquest ens permeti fer un món millor.
La motivació ho és tot.
Abans de començar permeteu que em disculpi per les possibles errades i que excusi
algunes informalitats de la present tesi. Fins i tot en l’àmbit més científic crec que de tant en
tant es pot fer una petita concessió a la literatura i a l’humor. Possiblement així seria més fàcil
que la societat entengui quins són els avenços científics, i és que la divulgació del resultats
segueix sent una assignatura pendent per alguns camps de la ciència. L’objectiu no és ofendre
ni fer perdre el temps, sinó mantenir el lector concentrat i atent, ser pedagògic i il·lustratiu en
les explicacions i fer amè el text. Escrit amb la millor intenció i intentant ser rigorós
científicament, espero que sigui una lectura plaent. The show must go on!


1 Introducció
3
1. INTRODUCCIÓ
1.1. De la llavor a la planta: La llum i la fotomorfogènesis.
Tot comença amb una llavor (bé, podríem parlar de si va ser abans la planta o la llavor,
però en aquest apartat ens centrarem amb la llavor). La nostra protagonista té una envoltura
rica en nutrients, una herència de la seva progenitora, que li ha de permetre arribat el moment
adequat, invertir en el negoci de la seva vida. Però aquests recursos són limitats i per tant els
ha de fer servir per tal que pugui generar aviat més ingressos. Tenint en compte que és un
organisme sèssil (que no es podrà moure de lloc), podem imaginar que és molt important tenir
control de les condicions que l’envolten abans de començar a créixer. Un error significaria la
seva mort, un intent fracassat de prosperar.
Després del fred de l’hivern és un bon moment per germinar, ja que les temperatures es
fan més moderades, augmenta la humitat i hi ha disponibilitat de nutrients. Amb aquestes
condicions favorables la llavor s’arrisca a germinar i comença a créixer la plàntula. Com que la
llum serà el seu combustible és molt important disposar d’aquest recurs en prioritat. Així la
plàntula creixerà invertint tota la seva energia inicial en allargar-se fins trobar una font de llum
suficient per satisfer les seves necessitats. Mentre no trobi aquesta llum la plàntula es
desenvolupa etiolada (o escotomorfogènica, del grec skotos que vol dir foscor). En aquest
període la plàntula presenta un color groc pàl·lid degut a la baixa quantitat de pigments
fotosintètics en els plasts, amb un hipocòtil llarg, un ganxo apical tancat i cotiledons petits i
tancats que envolten el primordi foliar no desenvolupat. No obstant, quan la plàntula trobi llum
suficient l’elongació de l’hipocòtil s’inhibirà, obrirà els seus cotiledons (els seus panells solars),
per tal de captar l’energia del sol i seguir creixent amb aquesta. D’aquesta manera es torna
verda i comença a formar les fulles, ja que augmenta l’activitat fotosintètica. En aquestes
condicions es diu que plàntula es desetiola, o que creix fotomorfogènicament (del grec photo,
que vol dir llum)(Arsovski et al. 2012).
Així és com la nostra plàntula deixa de ser heteròtrofa, mentre la plàntula obté l’energia
de les reserves de la llavor, i passa a ser autòtrofa, a l’obtenir-la de la fotosíntesi. Si tot va bé,
la plàntula tindrà llum i recursos suficients i anirà creixent, en sortiran més fulles, i establirà
l’etapa juvenil anomenada roseta. Posteriorment del centre de la roseta en sortirà la tija, que
anirà creixent i d’on sortiran les flors. D’aquesta manera arribem a la planta adulta.
Durant aquest procés la llum no és contínua, sinó que es van alternant diferents períodes
de llum i foscor (dia i nit), i es van rebent diferents qualitats i quantitats de radiacions (segons
posició del Sol, possibles ombres de vegetació veïna....). Tots aquests canvis han de ser
percebuts i interpretats per la planta per tal de respondre i adaptar-se adequadament. Cal que
sigui capaç de diferenciar la nit de l’ombra de la vegetació veïna, o un núvol passatger de la
posta de Sol, ja que l’activitat metabòlica de la planta depèn d’aquests canvis en la llum i no
seria òptim parar les màquines, parar la fotosíntesi, només pel pas d’un núvol passatger
pensant que s’ha fet de nit. Igual nosaltres no anem a dormir a les 12 del migdia encara que
sigui un dia que el cel estigui tan tapat que sembli de nit.

1 Introducció
4
Així doncs, com sent una planta aquests canvis en l’ambient? Quins canvis pot percebre
la plàntula i com reacciona per adaptar-se? Són grans preguntes, que molta gent s’ha plantejat.
Anem doncs a veure com s’ho fan.
1.2. Percepció de la llum: Els fotoreceptors i, més en concret,
els fitocroms.
Com passa sovint en la ciència, els primers fotoreceptors es van descobrir per error. Va
ser als anys 20, quan s’estaven aïllant els pigments involucrats en la fotosíntesi. No va ser fins
passats 30 anys quan es va determinar la naturalesa proteica d’alguns d’aquells pigments i es
va postular que podrien no estar involucrats en la fotosíntesis. Als anys 50, Sterling Hendricks i
Harry Borthwick investigaven aspectes d’interès agrícola, com la germinació o la floració de les
plantes, i van descriure que aquests pigments controlaven la germinació de llavors d’enciam
(Bothwick et al. 1952). Fent servir un espectrògraf dissenyat i fabricat casolanament van veure
que en les seves llavors un pols de llum vermella (R de l’anglès Red) induïa la germinació, però
aquesta inducció era inhibida exposant immediatament les llavors a un pols de llum vermella
llunyana (FR de l’anglès Far Red). Les llavors es podien tractar repetidament amb llum R i FR, i
la resposta final (germinació o no germinació) venia determinada per l’últim pols de llum,
deixant a entendre que era una resposta regulada reversiblement per la llum.
No va ser fins al 1959 que Warren Butler i Harold Siegelman van identificar i batejar com
a fitocroms aquells pigments, i fins al 1983, al laboratori de Peter Quail i Clarck Lagarias, no es
va aïllar el primer fitocrom purificat intacte. Aquest mateix laboratori, al 1985, va seqüenciar el
primer gen codificant per fitocrom. En pocs anys es van identificar altres fitocroms en plantes i a
finals dels 90 es van trobar en altres grups (gimnospermes, falgueres, molses, algues i bacteris,
entre d’altres), evidenciant que no només els organismes fotosintètics disposen d’aquest tipus
de fotoreceptor. Actualment es diferencien diversos grups de fitocroms segons la procedència
evolutiva: fitocroms de plantes, bacteriofitocroms, fitocroms de cianobacteris, de fongs i un
conjunt de molècules afins a fitocroms (Karniol et al. 2005). També a finals dels 90, i després
de la clàssica controvèrsia científica, es va establir la localització subcel·lular d’aquests
pigments, obrint diferents hipòtesis sobre els mecanismes de transducció de senyal d’aquests
fotoreceptors. Cap al 2000, també de la mà de Peter Quail i col·laboradors, es van identificar
factors que interaccionen directament amb aquests fotoreceptors per tal de regular processos
com la fotomorfogènesis.
També cap al 2000 es van identificar gens que codificaven altres fotoreceptors
moleculars en plantes (Ahmad and Cashmore 1996, Chen et al. 2004, Quail 2002, Somers and
Fujiwara 2009). Així es va comprovar que les plantes disposen de diversitat de fotoreceptors
amb diferències en estabilitat, estructura, expressió... La següent pregunta és lògica, per què
aquesta diversitat de fotoreceptors? Com actuen?
Com s’ha comentat anteriorment la planta necessita controlar no només la quantitat de
llum (o intensitat), sinó la duració, la qualitat (composició de l’espectre, relació de les diferents
longituds d’ona) i aquests són aspectes que canvien segons l’estació de l’any, la latitud, la

1 Introducció
5
proximitat d'altres plantes, factors climatològics etc. Però cada cas és diferenciable i té uns
canvis específics en l’espectre de llum, com una empremta digital. Així, per la planta, la llum és
més que una font d’energia, també és una font d’informació molt útil. Per exemple, la proximitat
de vegetació veïna es pot detectar per la reducció de llum roja (R), absorbida per la
fotosíntesis, i un augment de la llum roja llunyana (FR) reflexada per la vegetació, mentre que
els nivells de llum blava i verda es mantenen relativament constants; així mateix, l'ombra
vegetal es diferencia per una baixada en les intensitats de llum UV, blava i R, així com un
augment de la FR (Ballaré et al. 1990, Morgan and Smith 1979). Aquestes condicions són
semblants a les condicions lumíniques del vespre, on la llum del Sol es torna menys intensa,
però la durada d’un vespre és curta i ve seguida d’una nit, mentre que un ombrejat vegetal es
manté al llarg del temps durant diversos dies (Holmes and Smith 1977). Per això les plantes
han adquirit durant l’evolució un conjunt de fotoreceptors que capten les intensitats de senyal
en diverses longituds d’ona i que, a més, han establert una xarxa de regulació molt complexa
per tal de poder respondre amb precisió a cada canvi en la llum. Identificant cada situació per
les seves empremtes i responent d’una manera específica en cada cas.
Actualment, gràcies al treball de desenes de grups d’investigació, s’han identificat 5
classes de fotoreceptors en Arabidopsis thaliana (d’ara endavant simplement Arabidopsis), que
són presents en totes les plantes superiors estudiades. Arabidopsis compte amb 5 fitocroms
(phyA-phyE) amb màxims d’absorció en la llum R i FR (Franklin and Quail 2010). A més
disposa de tres classes de receptors específics de UV-A/llum blava anomenats criptocroms
(cry1-cry2), fototropines (phot1 i phot2) i zeitlupes (ZTL, FKF1 i LKP2) (Christie 2007, Demarsy
and Fankhauser 2009, Lin and Shalitin 2003, Somers and Fujiwara 2009). Hi ha un tercer
membre en la família dels criptocroms, cry3, pertanyent al grup de les ADN fotoliases. Encara
no està clar si cry3 actua realment com a fotoreceptor, però com a mínim és actiu en el rol de
reparació de l’ADN en resposta a llum UV (Pokorny et al. 2008). Les plantes superiors tenen a
més receptors UV-B amb un ampli rol en la fotomorfogènesis, tot i que de moment només un
ha estat identificat (UVR8)(Favory et al. 2009, Jenkins 2009). Finalment, un petit nombre de
respostes a la llum verda podrien estar dirigides per un altre receptor encara per identificar
(Wang and Folta 2013)(figura I.1).
Tots els fotoreceptors, excepte el UVR8, tenen una estructura bàsica semblant, basada
en una cadena polipeptídica anomenada apoproteïna unida a un grup cromòfor (Lin and Todo
2005, Rockwell et al. 2006). Les longituds d’ona d’absorció de cada receptor venen
determinades per la composició del grup cromòfor i l’apoproteïna en qüestió. El receptor URV8,
en canvi, utilitza com a cromòfor un parell de triptòfans de la mateixa cadena peptídica, residus
que faciliten la dimerització en baixes quantitats de llum UV-B i en faciliten la separació al rebre
aquesta llum (O'Hara and Jenkins 2012, Tilbrook et al. 2013). Els criptocroms i els fitocroms
controlen el creixement i desenvolupament en resposta a la qualitat, intensitat i periodicitat de
la radiació, mentre que les fototropines controlen el creixement direccional (fototropisme) i/o
moviments intracel·lulars dels cloroplasts en resposta a llum (Briggs and Christie 2002). La llum
UV-B provoca respostes com la inhibició de l’allargament de l’hipocòtil i la regulació de la

1 Introducció
6
transcripció de multitud de gens (Kim et al. 1998). Els zeitlupes estan implicats en el control del
temps de floració i del rellotge circadià (Kim et al. 2007, Song et al. 2012). No obstant, dins de
cada família, cada membre té funcions relativament diferenciades, la qual cosa proporciona un
nivell addicional d’especialització (Quail 2002).
Longitud d’ona (nm)
uv-B uv-A blau verd groc vermell vermell llunyà
400 500 600 700 800
Fo
tom
orf
og
ènes
isF
oto
sín
tesi
s
CAROTENOIDES
CLOROFIL·LESCLOROFIL·LES
CRIPTOCROMS
FOTOTROPINES FITOCROMS
(phy)
UVB
ZEITLUPS
D’entre tots els fotoreceptors, s’ha demostrat que els fitocroms tenen un rol destacat en
la germinació, promovent-la en condicions lumíniques favorables i reprimint-la en condicions
d’ombrejat o foscor (Franklin and Quail 2010), confirmant els resultats publicats per Hendricks i
Borthwick. A més, s’ha vist que per promoure la germinació els fitocroms actuen primer en la
síntesi i senyalització de les gibberel·lines (GA) (Oh et al. 2007, Piskurewicz et al. 2008),
enllaçant així la percepció de la llum amb altres vies de senyalització de la planta.
Els fitocroms són cromoproteïnes dimèriques. Com la majoria de fotoreceptors, cada
subunitat està composta per dos components: l’apoproteïna i el grup cromòfor. Tots els
fitocroms tenen el mateix cromòfor però es diferencien per l’apoproteïna. Són sintetitzats en la
seva isoforma Pr, que presenta un màxim d’absorció a la llum R (λmàx= 670 nm). Quan el
cromòfor capta llum d’aquesta longitud d’ona provoca un canvi de conformació que dóna lloc a
l’isoforma absorbent de llum FR (λmàx= 730 nm) anomenada Pfr. Aquest canvi de conformació
deixa al descobert una senyal de localització nuclear, donant lloc a la translocació cap al nucli
de la proteïna i a la seva activitat (Nagatani 2004). D’aquesta manera els fitocroms actuen com
interruptors en un fotoequilibri que depèn de les condicions de llum (una balança determinada
per la proporció de les llums R i FR que reben). Ambdues formes dels fitocroms presenten a
més un segon pic d’absorció en la zona UV-A/llum blava (Rockwell, et al. 2006). Aquestes
absorcions es corresponen amb les respostes que porten a terme, amb activitats màximes en
llums FR i algunes respostes també en UV-A/llum blava.
El repertori de fitocroms s’acostuma a classificar bàsicament en dos grups: els fotolàbils,
que són més abundants en plàntules etiolades (phyA); i els fotoestables, lògicament més
abundants en plàntules creixent en llum (phyB-E). Totes les plantes analitzades fins al moment
han complert la condició de tenir alts nivells d’expressió de PHYA i B en les llavors (Mathews
Figura I.1. Espectre de radiació de la llum i els principals fotoreceptors vegetals. Representació de l’espectre de llum i dels
fotoreceptors que capten les diferents longituds d’ona. A la part alta els fotoreceptors implicats en percebre la informació de la llum,
en la part baixa els receptors que usen la llum com a font d’energia (fotosíntesis).

1 Introducció
7
2006). Aquests dos receptors tenen funcions diferents i s’ha descrit que phyA ha evolucionat
adquirint noves activitats per tal d’adaptar la seva funció millorant l’adaptació de la planta en
condicions d’alta densitat vegetal (Mathews 2006, Rockwell, et al. 2006). De fet, phyA és el
més abundant en plàntules creixent en foscor i és degradat en presència de llum (Seo et al.
2004). A més a més phyA, a diferència de la resta de fitocroms, podria tenir algun rol específic
fins i tot quan s’acumula en el citosol, ja que mutants incapaços d’importar phyA al nucli
mostren respostes normals de gravitropisme, a diferència del que passa amb el mutant phyA
(Rosler et al. 2007). phyB és el segon fitocrom més abundant i regula la morfologia de la
plàntula en relació a la qualitat de llum: en alta proporció R:FR (quan no es troba vegetació en
la proximitat), el phyB es troba majoritàriament en la seva forma activa (Pfr) i reprimeix per
exemple l’elongació de l’hipocòtil; en baixa raó R:FR (en condicions d'alta densitat vegetal) la
isoforma majoritària és la inactiva (Pr) i això (la manca de forma activa Pfr), per exemple, des-
reprimeix l’elongació de l’hipocòtil. Més tard parlarem de la rellevància biològica d’aquesta
activitat (en l’apartat 1.6, però no siguem impacients).
Les respostes regulades pels fitocroms s’han classificat segons la quantitat d’energia de
radiació requerida per la seva resposta (Casal et al. 1998, Mancinelli 1986):
i) Respostes de molt baixa fluència de llum (VLFR, de l’anglès Very Low Fluence
Response): s’indueixen amb fluxos fotònics molt baixos (de 10-3 a 1 µmol/m2), són sensibles a
un ampli espectre de longituds d’ona (de 300 fins a 780 nm) i són respostes no reversibles.
ii) Respostes de baixa fluència de llum (LFR, de l’anglès Low Fluence Response): són
les clàssiques respostes desencadenades per llum R i revertides per llum FR, que van
permetre el descobriment dels fitocroms. Es saturen amb temps curts i baixes energies (de 1 a
103 µmol/m2), i obeeixen la llei de la reciprocitat de Bunsen-Roscoe, que diu que una resposta
ha de dependre només del total de fotons de llum rebuts, independentment de la durada de
l’exposició a la font de llum. S’indueixen amb una breu irradiació a R, que desplaça el
fotoequilibri dels fitocroms fortament, fet que es pot revertir amb una breu irradiació de FR
sempre i quan no passi molt temps entre irradiacions R-FR. Si passa massa temps, es produeix
una escapada de la reversibilitat, la magnitud de la qual és variable en funció de les respostes.
iii) Respostes d’alta irradiància de llum (HIR, de l’anglès High Irradiance Response):
Requereixen exposicions prolongades amb un flux fotònic relativament alt (>103 µmol/m2). La
magnitud de la resposta depèn de la longitud d’ona, la taxa de flux fotònic i la duració de la
irradiació. No presenten reversibilitat R-FR i no segueixen la llei de la reciprocitat. Segons es
desencadenin per una radiació R o FR es denominen R-HIR o FR-HIR, respectivament.
En contrast amb altres fitocroms, phyA és responsable exclusivament de respostes del
tipus VLFR, generades en condicions de poca llum (Shinomura et al. 1996) i de respostes a
alta irradiància de FR (FR-HIR), les quals són observades en llum contínua FR (FRc)(Nagatani
et al. 1993, Parks and Quail 1993, Whitelam et al. 1993). phyB (conjuntament amb els altres
fitocroms fotoestables) és el responsable més important de les respostes a alta irradiància de
R (R-HIR) i de la clàssica resposta reversible R:FR de baixa fluència durant la fotomorfogènesis
(Shinomura, et al. 1996). Així, els diferents fitocroms tenen algunes respostes redundants,

1 Introducció
8
altres antagonístiques i algunes sinèrgiques (Bae and Choi 2008). Aquestes dades es troben
resumides en la taula I.1.
Fitocrom Estabilitat Activitats
fotosensoresRespostes fisiològiques controlades
phyA Fotolàbil VLFRs Germinació de llavors (llum UV, visible, FRc)
FR-HIRs Desetiolació de plàntules (FRc), control del fotoperíode i inducció de
la floració (LD)
phyB Fotoestable LFRs
Germinació de llavors (Rc), Respostes de la SAS (allargament de pecíols i entrenusos, expansió dels cotilèdons, floració)
R-HIRs Desetiolació de plàntules (Rc)
phyC Fotoestable R-HIRs Desetiolació de plàntules (Rc), control del fotoperíode
phyD Fotoestable LFRs
Germinació de llavors (Rc), respostes de la SAS (allargament de pecíols i entrenusos, expansió dels cotilèdons, floració)
R-HIRs Desetiolació de plàntules (Rc)
phyE Fotoestable LFRs
Germinació de llavors (Rc), respostes de la SAS (allargament de pecíols i entrenusos, expansió dels cotilèdons, floració)
R-HIRs Desetiolació de plàntules (Rc)
VLFRs: Respostes a fluxos molt baixos R: Llum vermella LFRs: Respostes a fluxos baixos FRc: llum vermella llunyana contínua HIRs: Respostes a alta irradiància Rc: llum vermella contínua FR: Llum vermella llunyana o blanca enriquida en FR LD: Dia llarg (Long Day)
Anteriorment ja hem comentat la rellevància que té per la planta controlar les condicions
de llum, especialment perquè és la seva principal font d’energia. Tot i que moltes respostes
fisiològiques han estat identificades com regulades per llum R i/o FR, les plantes en ambients
naturals no estan exposades mai a una llum monocromàtica. Gràcies a la gran varietat de
fotoreceptors les plantes perceben un senyal lumínic complex (de composició qualitativa i
quantitativa variable), i responen d’acord a aquest, permetent-les créixer favorablement en
ampli rang de condicions lumíniques. No obstant, per la present tesi els fitocroms són els
fotoreceptors més rellevants i seran uns personatges destacats d’aquesta història, per això els
estem explicant més en detall que la resta de fotoreceptors.
Ara ja sabem el què (canvis en la llum), tenim un qui (els nostres amics informadors o
xivatos: els fotoreceptors) i el perquè (obtenir informació per adaptar el creixement a les
condicions). Ens queda un dels factors més complicats: com es passa de la detecció de la
senyal als canvis fisiològics? Hem fet alguna pinzellada a alguns factors que hi estan
involucrats, com la senyalització per hormones. Seguidament en parlarem més en detall.
1.3. Més enllà dels fitocroms: la transducció de senyal i la xarxa
reguladora.
Les plantes perceben i diferencien diverses longituds i intensitats de la llum gràcies a la
diversitat d’informadors de què disposen, però, com es coordinen els diferents receptors? És
clar que malgrat la gran quantitat de receptors i les seves diferències, han de treballar
conjuntament amb altres mecanismes de la planta per coordinar correctament el creixement i el
metabolisme. Com en tota organització hi ha d’haver alguna jerarquia i alguns factors que
coordinin les respostes i comprovin que són les més adequades a les circumstàncies. Igual que
en una fàbrica, si es vol produir més cal que es coordinin els treballadors i el que demana la
TaulaI I.1. Funcions biològiques dels fitocroms en Arabidopsis. Taula adaptada de Li et al. 2011.

1 Introducció
9
matèria primera, o sinó la producció s’estancarà per falta de recursos. En aquest aspecte la
comunitat científica ha fet (i encara està fent) molts esforços per tal d’esbrinar quins són
aquests mecanismes de control.
Posem-nos en context. La planta, mitjançant els seus fotoreceptors, percep canvis en
l’ambient, això vol dir que rep una informació (en anglès seria un input). Aquesta senyal
lumínica és processada, transformant-la en una senyal cel·lular i molecular, i acaba donant lloc
a una resposta fisiològica, normalment visible al cap d’un temps (el que seria un output).
Aquest camí és el que es coneix com a transducció de la senyal. Per entendre els mecanismes
de senyalització dels fitocroms, aquests s’han de considerar dins del context general de la
transducció de la llum.
Anàlisis transcriptòmiques van demostrar que la llum produeix canvis ràpids de
l’expressió gènica, de forma que reprogramen la planta (Devlin et al. 2003, Ma et al. 2001,
Tepperman et al. 2001). Aquesta reprogramació produeix, per exemple, la transició ràpida entre
els dos programes de desenvolupament: de l’escotomorfogènesi a la fotomorfogènesi.
Depenent de si una plàntula es desenvolupa en llum o en foscor hi ha una expressió diferencial
d’aproximadament un 20% dels gens. Els efectes de la llum s’estenen fins a la regulació gènica
de la majoria de les rutes bioquímiques situades dins d’orgànuls cel·lulars (Ma, et al. 2001). Cal
destacar que el mateix estímul lumínic pot causar efectes diferents segons l’òrgan on es rebut,
demostrant que una planta ajusta el creixement dels òrgans i dels teixits de manera específica
per tal d’adequar la seva resposta a la llum (Bou-Torrent et al. 2008, Jiao et al. 2007). Més d’un
fotoreceptor pot contribuir a una determinada resposta. Per exemple, en condicions de
laboratori es pot induir el desenvolupament fotomorfogènic de la plàntula amb llum
monocromàtica blava, R o FR, en un procés induït específicament pels criptocroms, per phyB o
per phyA, respectivament. De manera que en condicions de llum normals (on hi ha els tres
tipus de llums simultàniament), els tres fotoreceptors promouen la fotomorfogènesis
conjuntament. Per tant, a la natura hi ha d’haver punts d’integració de la senyal procedent dels
diferents fotoreceptors. Cal tenir en compte que sovint les respostes de desenvolupament
depenen d’altres factors ambientals a part de la llum (com la temperatura, la humitat, els
estressos per factors biòtics o abiòtics...), indicant que també són necessaris punts d’integració
de senyals de les diferents xarxes. Entendre la jerarquia d’aquestes xarxes formades per
factors de transcripció i identificar els elements reguladors clau en els diferents processos del
desenvolupament regulats per la llum permetria entendre, i modificar, el creixement i el
metabolisme de la planta segons les nostres necessitats.
Gràcies a la col·laboració de diversos grups s’ha vist que l’activació dels fitocroms
inicia una cascada de senyalització mitjançada per la interacció dels fitocroms amb com a
mínim un tipus de factors de transcripció, coneguts com a PIF (de l’anglès PHYTOCHROME
INTERACTING FACTOR (Castillon et al. 2007, Jiao, et al. 2007, Quail 2002). Els PIFs són
membres del nombrós club de les bHLH’s (basic Helix-Loop-Helix) i regulen diferents aspectes
del desenvolupament de la planta. D’altra banda s’han identificat gens que són ràpidament
regulats per l’acció dels fitocroms (gens PAR, de l’anglès PHYTOCHROME RAPIDLY

1 Introducció
10
REGULATED), alguns dels quals responen de manera directa per l’acció d’aquests (Roig-
Villanova et al. 2006). S’ha postulat que els PIFs podrien ser peces clau en la connexió de les
diferents senyals o estímuls que rep la planta (Leivar et al. 2012, Martínez-García et al. 2010)
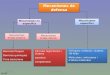
(figura I.2). La regulació del desenvolupament mitjançada per factors de transcripció aigües
avall dels fitocroms és encara tema d’estudi. Seguidament passarem a explicar els principals
avenços fets fins al moment.
nucli
phyA
PIFZ HY5
A
B
C
PIFYCCA1
LHY
PIFXCO
TOC1-L
RPT2
DOF
FT
rellotge
GDCH
CAB
RBCS
CHS
?
?
?
?
Ritmescircadians
Floració
Fotorespiració
Fotosíntesi
Biosíntesi de flavonoides
Biosíntesi de carotenoides
Allargamentde l’hipocòtil
Expansiódelscotiledons
Fototropismecitoplasma
nucli
phyXPIFX HY5
PARX
PIFY
A
CCA1
LHY
PARY
CO
TOC1
PIFZ
RPT2
DOFFT
rellotge
GDCH
CAB
RBCS
CHS
?
?
?
?
Ritmes circadians
Floració
Fotorespiració
Fotosíntesi
Biosíntesi de flavonoieds
Biosíntesi de carotenoides
Allargament de l’hipocòtil
Expansió dels cotilèdons
Fototropisme
phyY
PIF3PIFN
llum
1.4. Mecanismes moleculars dels fitocroms: la relació amor-odi
amb els PIFs.
S’ha avançat molt en el coneixement del mecanisme de transducció de senyal de llum, si
bé encara queden moltes preguntes per respondre. S’ha vist que els fotoreceptors, i
especialment els fitocroms, poden actuar de diverses maneres per regular les respostes a la
llum. Una de les primeres observacions va ser la capacitat dels fitocroms per fosforilar certs
substrats com PKS1 (de l’anglès PHYTOCHROME KINASE SUBSTRATE1) (Fankhauser and
Chory 1999), Aux/IAA (Colon-Carmona et al. 2000) i alguns criptocroms (Ahmad et al. 1998). A
més, els propis fitocroms poden ser fosforilats en diversos residus. S’ha suggerit que la
fosforilació i desfosforilació té una funció important modulant la localització subcel·lular,
l’estabilitat i/o l’afinitat a altres components, de manera dependent a les condicions de llum
(Kim et al. 2004).
Per tal de definir com són els mecanismes de transducció de la senyal dels fitocroms
s’han realitzat diversos experiments, com crivellatges de mutants en condicions de llum
determinada, o d’interacció amb els fitocroms. En assaigs de doble híbrid es van identificar
Figura I.2. Model de transducció del senyal lumínic. Es proposa que la captació de llum per part dels fitocroms (phy) dóna lloc a
unes respostes fisiològiques finals mitjançant el control de l’estabilitat dels PIFs per parts dels fitocroms. Canvis en els PIFs
impliquen canvis d’expressió de gens diana primaris, ràpidament regulats pels fitocroms (cercles blaus i grocs). Aquests gens diana
primaris codificarien factors de transcripció que regularien l’expressió d’altres gens més tardans que controlen una o més branques
de la xarxa transcripcional, tant per desenvolupament com per les respostes fisiològiques a llum. A més, hi ha diversos punts de
regulació creuada, per exemple, alguns PAR també controlen l’activitat d’alguns PIFs. Per simplificar les fletxes volen dir algun tipus
de regulació o interacció, sense detallar si positiva o negativa.

1 Introducció
11
diverses proteïnes que interaccionen físicament amb phyA i phyB. El primer element identificat
va ser PIF3, trobat en un crivellatge que utilitzava la part Ct de phyB com a esquer. Es va veure
que interacciona amb igual eficiència amb l’extrem Ct de phyA que de phyB, però que
preferentment ho fa amb la proteïna phyB intacta, essent aquesta interacció depenent de llum.
Així, PIF3 s’uneix preferentment a la forma Pfr (activa) del fitocrom, i la conversió per canvis de
llum de la forma Pfr a la forma Pr (inactiva) provoca la dissociació de PIF3 (Ni et al. 1998). És el
que es podria conèixer com la resposta “tú antes molabas”, on canvis en els fitocroms
provoquen el divorci amb els seus interactors.
Després de la identificació de PIF3 s’han trobat altres bHLHs per similitud de seqüència
aminoacídica, agrupant-se dins la subfamília 15 o grup VII de les bHLHs (Leivar and Quail
2010). De les diverses bHLHs del grup, fins a 6 membres més s’han classificat com a PIFs:
PIF1/PIL5 (Huq et al. 2004, Yamashino et al. 2003), PIF4 (Huq and Quail 2002), PIF5/PIL6
(Khanna et al. 2004, Yamashino, et al. 2003), PIF6/PIL2 (Khanna, et al. 2004, Yamashino, et al.
2003), PIF7 (Leivar et al. 2008) i PIF8. Mentre que els vuit membres restants de la família, tot i
tenir homologia de seqüència, no s’ha demostrat que puguin interaccionar amb els fitocroms:
HFR1 (LONG HYPOCOTYL IN FR) (Fairchild et al. 2000), PIL1 (PIF3-LIKE PROTEIN 1)
(Makino et al. 2002), SPT (SPATULA) (Josse et al. 2011), ALC (ALCATRAZ) (Rajani and
Sundaresan 2001), bHLH23, bHLH56, bHLH119 i bHLH127. Pràcticament tots estan implicats
en la senyalització de llum pels fitocroms, bé per ser autèntics interactors dels fitocroms (PIFs),
o bé perquè tot i no interaccionar directament amb els fotoreceptors s’ha demostrat que
heterodimeritzen amb els PIFs, modulant així la seva activitat (Castillon, et al. 2007,
Hornitschek et al. 2009). Alguns gens identificats primer com a PILs es van reanomenar
posteriorment com a PIFs al demostrar-se la seva interacció directa amb els fitocroms (Huq, et
al. 2004, Khanna, et al. 2004). S’ha demostrat la unió in vitro de PIF4, PIF5, PIF6, PIF7 i PIF8 a
la forma Pfr de phyB, mentre que PIF1 i PIF3 uneixen la forma Pfr tant de phyA com de phyB,
mitjançant dominis específics independents (Khanna, et al. 2004, Zhu et al. 2000). Tots els
PIFs són factors de transcripció que posseeixen la capacitat d’unir al DNA en forma de dímers,
generalment activant l’expressió gènica (Hornitschek et al. 2012, Oh et al. 2012).
Les interaccions entre els fitocroms i els PIFs són de gran rellevància, ja que després de
la percepció de llum els fitocroms indueixen la múltiple fosforilació dels PIFs produint la seva
degradació via proteosoma 26S (Al-Sady et al. 2008, Al-Sady et al. 2006, Bauer et al. 2004, de
Lucas et al. 2008, Lorrain et al. 2008, Nozue et al. 2007, Oh et al. 2006, Park et al. 2004, Shen
et al. 2008, Shen et al. 2007). Aquest mecanisme podríem anomenar-lo “relació d’amor-odi”, ja
que la interacció entre els dos amants desemboca en el tràgic final d’un membre de la parella,
fins i tot abans que arribi el divorci. Aquest procés sembla general en els PIFs, amb l’única
excepció de PIF7, que si bé és més estable i no es veu degradat en presència de llum, si que
és fosforil·lat, el que prevé la seva unió al DNA (Li et al. 2012). A més PIF1 i PIF3 sembla que
també tenen un mecanisme semblant de regulació independent de la seva degradació (Park et
al. 2012), suggerint que es podria tractar d'un efecte general dels fitocroms vers tots els PIFs.

1 Introducció
12
La ràpida ubiqüitinació de PIF1, PIF3, PIF4 i PIF5 en presència de llum ha suggerit que
tenen un paper conjunt en la fotomorfogènesis (Al-Sady, et al. 2006, Shen, et al. 2008).
Aquests resultats són coherents amb els alts nivells de proteïna trobats en plàntules creixent en
foscor i la seva ràpida desaparició en condicions de llum normals (alta raó R:FR, on els
fitocroms són majoritàriament actius). En el cas de PIF1 i PIF3, d’acord amb la seva capacitat
per interaccionar amb phyA, s’ha vist que també decauen en condicions de FRc, mentre que
PIF4 i PIF5 són estabilitzats en resposta a FR (Bauer, et al. 2004, Lorrain et al. 2009, Park, et
al. 2004, Shen, et al. 2007).
Estudis amb mutants i plantes sobre-expressores d’alguns PIFs han portat a demostrar
que tenen algunes activitats específiques i altres redundants. Per exemple, la germinació està
controlada prioritàriament per PIF1, juntament amb SPT i PIF6, que tenen papers molt més
limitats (Oh et al. 2004, Penfield et al. 2010, Penfield et al. 2005, Shin et al. 2009). Plantes pif1
mutants germinen fins i tot quan la majoria de fitocroms són inactius (Oh, et al. 2004, Shin, et
al. 2009) i no mostren cap canvi en nivells d’expressió de gens regulats durant la germinació al
créixer en llum Rc. S’ha vist que PIF1 és un inhibidor de la germinació que regula directament
els nivells d’ABA i GA, dues hormones essencials durant el procés de germinació (Oh et al.
2009, Piskurewicz, et al. 2008). Mitjançant la degradació de PIF1 els fitocroms actius desfan
aquesta inhibició de la germinació.
En contrast amb l’especificitat de PIF1 en la germinació, com a mínim quatre altres PIFs
controlen conjuntament el desenvolupament de la plàntula durant la fotomorfogènesis,
promovent en general el creixement (Leivar, et al. 2008, Leivar et al. 2009, Shin, et al. 2009).
PIF3 participa en diferents respostes, entre elles en el control de l’allargament de l’hipocòtil sota
llum Rc (Kim et al. 2003) i, de manera majoritària, en resposta a etilè (Zhong et al. 2012). PIF4,
en canvi, sembla estar regulant l’expansió cel·lular a l’hipocòtil i als cotiledons en resposta a Rc
(Huq and Quail 2002), i és el regulador principal en resposta a alta temperatura, promovent
l’elongació de l’hipocòtil i la floració primerenca (Franklin et al. 2011, Koini et al. 2009, Kumar et
al. 2012). Funcions similars es postulen per a PIF5 i PIF6 (Khanna, et al. 2004). PIF3, PIF4,
PIF5 i PIF7 semblen ser els reguladors principals que promouen les respostes en ombra (com
que són l’objectiu d’aquesta tesi en parlarem més en profunditat a l’apartat 1.7) (Leivar, et al.
2008, Li, et al. 2012, Lorrain, et al. 2008). El quàdruple mutant pif1pif3pif4pif5 (conegut també
com a pifq) té un fenotip fotomorfogènic quan creix en foscor, presentant alteracions moleculars
relacionades amb la desetiolació (Leivar, et al. 2008, Leivar, et al. 2009, Shin, et al. 2009). En
plàntules creixent en foscor PIF1, PIF3 i PIF5 controlen la biosíntesis de clorofil·les i,
conjuntament amb PIF4, la síntesis de carotenoides (Huq, et al. 2004, Shin, et al. 2009,
Stephenson et al. 2009, Toledo-Ortiz et al. 2010).
Així, es pot concloure que quan la planta creix en foscor, els PIFs s’acumulen i mantenen
el desenvolupament escotomorfogènic, mentre que en presència de llum, amb els fitocroms
majoritàriament actius, la inactivació i degradació dels PIFs promou el procés de desetiolació.
D’aquesta manera podríem dir que els PIFs són reguladors negatius en la transducció del
senyal lumínic, i que en general activen les respostes típiques del creixement en foscor, com

1 Introducció
13
l’elongació de l’hipocòtil o la inhibició de l'expansió dels cotiledons. No obstant, no tots els PIFs
actuen sempre negativament en la senyalització dels fitocroms. Per exemple PIF6 inhibeix
l’elongació de l’hipocòtil en llum Rc, donant lloc a un rol positiu en la senyalització en aquesta
llum (Penfield, et al. 2010).
Tot i que phyB es considera com fotoestable, cal destacar que els seus nivells
d’expressió estan lleugerament regulats pels PIFs (i per tant per acció dels propis fitocroms),
complicant sovint les interpretacions dels resultats obtinguts. Així, els nivells de phyB estan
incrementats en els mutants de PIFs, mentre que les línies sobre-expressores de PIFs tenen
nivells més baixos (Leivar, et al. 2008, Leivar and Quail 2011). Tot i que el mecanisme pel qual
els PIFs controlen els nivells de phyB encara no està clar, requereix de la interacció entre els
factors de transcripció i el fotoreceptor (Al-Sady, et al. 2008) i canvia segons les condicions de
llum. Hi ha evidències que porten a pensar que la degradació de phyB seria via ubiquitina-
proteosoma mitjançant COP1 com a E3 ligasa (Jang et al. 2010) i sembla que PIF3 tindria el
paper més rellevant en aquest mecanisme, amb aportacions de PIF1 i contribucions molt
menors de PIF4 i PIF5 (Leivar, et al. 2012).
Per complicar una mica més la trama, l’activitat dels PIFs es pot veure inhibida per altres
mecanismes més enllà de la fosforilació o la degradació, especialment en plantes desetiolades.
S’ha vist que la interacció dels PIFs amb cofactors dóna lloc a uns heterodímers que no poden
unir al DNA, de manera que aquests cofactors regulen la transcripció mitjançant la inhibició de
l’activitat dels PIFs (de Lucas, et al. 2008, Feng et al. 2008, Hornitschek, et al. 2009). PIF3 i
PIF4 heterodimeritzen amb membres de la família DELLA, sent un node de control entre
hormones i llum (Alabadi et al. 2008, de Lucas, et al. 2008, Feng, et al. 2008). L’activitat de
PIF4 i PIF5 és inhibida per HFR1, que s’acumula en altes quantitats en baixa R:FR, condició
típica de l’ombrejat o de la proximitat de vegetació veïna (Hornitschek, et al. 2009, Sessa et al.
2005). En aquest mateix sentit, alguns gens PAR com PAR1 i PAR2, essent membres de les
bHLH que han perdut el domini bàsic que facilita la unió al DNA (és a dir, contenen només un
HLH), controlen també per dimerització amb els PIFs les respostes als canvis de llum (Galstyan
et al. 2011, Hao et al. 2012). D’aquesta manera, cal considerar que tant els fitocroms com les
vàries proteïnes de la promíscua família bHLH poden formar heterodímers, la qual cosa implica
multitud de combinacions i un nivell de complexitat afegit a la regulació de factors de
transcripció per llum.
Alguns d’aquests mecanismes de control de l’activitat dels PIFs han posat de manifest
que, malgrat ser identificats com a components senyalitzadors de la llum, podrien estar regulats
i actuar de manera independent als fotoreceptors en algunes activitats. Per exemple, PIF4 i
PIF5 estan regulats també transcripcionalment pel rellotge circadià i per canvis de temperatura
(Koini, et al. 2009, Nozue, et al. 2007, Stavang et al. 2009). Així doncs, la senyalització per llum
pot influenciar les respostes a diversos estímuls, però aquests altres estímuls també poden
influenciar les respostes a la llum en una relació recíproca (Kami et al. 2010).
A part del procés de la fotomorfogènesi, hi ha també una regulació directa de l’expressió
gènica per part dels fitocroms en resposta a canvis en la llum quan la planta ja es troba

1 Introducció
14
desetiolada, més concretament, hi ha una regulació per canvis en les llums R i FR (Martínez-
García, et al. 2010). Per avalar aquest mecanisme assajos de inmunoprecipitació de la
cromatina (ChIP, de l’anglès Chromatin Immunoprecipitation), tant en ChIP-Chip com en ChIP-
seq, han permès identificar gens que són dianes directes de PIF1, PIF3, PIF4 i PIF5
(Hornitschek, et al. 2012, Oh, et al. 2012, Shin et al. 2007, Zhang et al. 2013), demostrant que
regulen l’expressió de gens involucrats en la senyalització hormonal i en la regulació de la
transcripció, especialment en respostes a canvis de llum R i FR, regulant en termes finals
diverses respostes fisiològiques i de desenvolupament. A més s’ha vist que els diversos PIFs
tenen unes 140 dianes comunes, unint-se a regions anomenades G-box del seu promotor
(CACGTG), però que cada un té moltes altres dianes específiques (Jeong and Choi 2013).
Aquesta especificitat en les dianes pot venir afectada a més per diferències en els nivells
d’expressió dels diferents PIFs segons el teixit, remarcant encara més els diferents rols
d’aquests factors. Aquestes dades recents aporten noves informacions dels successos
moleculars aigües avall dels fitocroms.
Membres de la família PIF i PIL han sigut identificats en una gran diversitat d’organismes
vegetals, tant dicotiledonis com monocotiledonis (per exemple en arròs) (Nakamura et al.
2007), i fins i tot en plantes inferiors, com la falguera Selaginella moellendorffii (e.g., NCBI
Gene ID 9657644) i la molsa Physcomitrella patens (e.g., NCBI Gene ID 5927830). L’alta
conservació dels PIFs i PILs en plantes terrestres fa pensar que aquests tenen un paper
essencial en l’adaptació a les condicions ambientals.
A part dels PIFs, la importància de la regulació per part dels fitocroms va ser posada de
manifest prèviament amb la identificació de COP1 (de l’anglès CONSTITUTIVE
PHOTOMORPHOGENIC 1). Mutants defectius d’aquest factor mostren un fenotip
fotomorfogènic en foscor, fet que porta a concloure que aquesta E3 ligasa és requerida per
mantenir la plàntula creixent escotomorfogènicament (Jiao, et al. 2007, Yi and Deng 2005).
COP1, junt amb membres de la família SPA (SUPRESSOR OF PHYA), controla l’abundància
de diversos factors regulats per llum, com HY5 (de l’anglès LONG HYPOCOTYL 5), LAF1
(LONG AFTER FR LIGHT 1), HFR1, CO (CONSTANTS), i el propi phyA (Jang et al. 2008, Jiao,
et al. 2007, Zhu et al. 2008). Consistentment aquesta llista inclou diversos factors que han de
mantenir-se en baixes concentracions en foscor i acumular-se durant la fotomorfogènesis. Tant
els fitocroms com els criptocroms en la seva forma activa inhibeixen l’activitat de COP1, però el
mecanisme encara no està del tot elucidat. Mitjançant la inhibició de COP1, i dels diversos
PIFs, els fitocroms actius produeixen canvis profunds en l’expressió gènica mediada per
aquests factors de transcripció, de manera que l’expressió dels gens és dependent de les
condicions de llum (Jiao, et al. 2007).
A més de COP1, altres factors que regulen les respostes a llum van ser trobats en
crivellatges realitzats sota llum FRc. En aquestes condicions els mutants haurien d’afectar
exclusivament en el procés de senyalització per phyA, ja que phyA és l’únic fotoreceptor que
controla les respostes FRc (Wang and Deng 2002). Així es van identificar components varis de
la senyalització per phyA com FHY1 (FAR-RED ELONGATED HYPOCOTYL 1), o el seu

1 Introducció
15
paràleg FHL (FHY1-LIKE), FHY3 i FAR1 (FAR-RED IMPAIRED RESPONSE 1). Les anàlisis
funcionals de mutants d’aquests gens han mostrat un rol de FHY1 i FHL en la localització
nuclear de la forma activa de phyA (PfrA). La captació de llum per phyA permet que passi a la
forma activa, la qual és capaç d’interaccionar amb FHL i FHY en el citoplasma. La senyal
nuclear que presenten aquests factors permet l’entrada del complex al nucli. A més els factors
FHY3 i FAR1 permeten l’acumulació de PfrA al nucli perquè activen l’expressió de FHY1 i FHL
(Hiltbrunner et al. 2006, Lin et al. 2007). Recentment s’ha vist que la fosforilació de FHY1 és el
mecanisme clau que defineix la dinàmica de senyalització de phyA sota llum R i FR en
Arabidopsis (Chen et al. 2012). Altres factors que s’han mostrat com a part important de la
senyalització de phyA són LAF1 (comentat abans com que la seva estabilitat esta regulada per
COP1) que codifica una proteïna MYB, i HFR1 (ja s'ha mencionat diverses vegades) que
codifica per una bHLH atípica (Bae and Choi 2008, Boccalandro et al. 2007). Finalment s’ha
especulat que el factor SPA1 també seria part d’aquesta via de senyalització. S’ha proposat
que el complex format entre COP1/SPA1 podria interactuar directament amb phyA per regular
la seva abundància i localització subcel·lular, actuant com un repressor de senyalització del
phyA (Saijo et al. 2008).
1.5. Respostes a la llum regulades pels fitocroms: La síndrome
de fugida de l’ombra (SAS).
Els fitocroms regulen un ampli rang de processos de desenvolupament al llarg de tot el
cicle de vida de la planta, des de la germinació de la llavor fins al temps de floració. L’aïllament
de mutants simples dels cinc fitocroms, i la posterior obtenció de mutants múltiples, ha
proporcionat informació sobre la funció dels fitocroms a través del desenvolupament vegetal.
Les principals respostes que s’ha vist que estant regulades pels fitocroms són:
- Germinació: Estudis de germinació han demostrat que mentre phyB indueix la
germinació en respostes de llum tipus LFRs i en alta raó R:FR (Li et al. 2011, Shinomura, et al.
1996), phyA la inhibeix per respostes tipus VLFR produïdes per una alta varietat d’irradiacions
(UV, visible i altes dosis de FR)(taula I.1). A més, phyA promou la germinació en llum FR
contínua en el mode HIR (Hennig et al. 2002). Tanmateix, phyD i phyE també tenen un paper
en el control de la germinació de la llavor en llum FRc (Botto et al. 1996, Dechaine et al. 2009).
També cal dir que la germinació d’una llavor està afectada per les condicions de llum en que ha
crescut la seva planta mare (Dechaine, et al. 2009).
- Desetiolació: Experiments amb els mutants deficients en diversos fitocroms han
permès caracteritzar-ne el seu rol en la fotomorfogènesis. Per exemple s’ha vist que mutants
de phyA mostren un fenotip fotomorfogènic normal quan creixen en llum blanca (W de l’anglès
White light) o Rc, però presenten un fenotip escotomorfogènic al créixer en FRc (és a dir, són
cegues a la llum FRc), confirmant que phyA és responsable de percebre i mediar vàries
respostes a la llum FR (Parks and Quail 1993, Whitelam, et al. 1993). D’altra banda el phyB és
predominant en regular les respostes de desetiolació en R contínua (Rc), ja que presenta un
fenotip parcialment escotomorfogènic al créixer en Rc. Mutants deficients en phyC mostren una

1 Introducció
16
pèrdua parcial de sensitivitat a la llum R, amb hipocòtils més llargs i cotiledons més petits que
el control silvestre (wt, de l’anglès Wild type), indicant que phyC té un rol en la regulació de la
desetiolació en R (Franklin 2003, Monte et al. 2003). Tanmateix, el phyC no actua
independentment del phyB, sinó possiblement formant heterodímers amb aquest in vivo (Clack
et al. 2009). El paper del phyD sembla minoritari, mentre que el del phyE és negligible,
demostrant un paper redundant amb phyB i phyD en la desetiolació (Devlin et al. 1998). En llum
UV-B, els fitocroms co-actuen amb fotoreceptors de llum UB-B/UV-A per regular la desetiolació
de les plàntules (Franklin and Quail 2010).
- Fototropisme: En una planta normalment la part aèria creix cap a la llum (fototropisme
positiu), mentre que les arrels creixen en direcció oposada (fototropisme negatiu). A més hi ha
alguns moviments de diferents teixits per tal de seguir el moviment del Sol al llarg del dia i
obtenir així el màxim d’hores d’insolació. En Arabidopsis el fototropisme està en general
controlat per les fototropines, però alguns fitocroms i criptocroms també poden afectar en
menor grau aquestes respostes (Jiao, et al. 2007).
- Respostes al fotoperíode: Els fitocroms són encarregats de controlar la llargada del
dia i del fotoperíode, els quals, juntament amb la temperatura, ajuden a la planta a controlar
l’estació de l’any on es troba. Aquestes senyals externes ajuden a sincronitzar el ritme del
rellotge circadià intern de la planta amb els cicles de llum/foscor del seu entorn (Kobayashi and
Weigel 2007, Mas 2008). Múltiples fotoreceptors han esta implicats en la transducció de
senyals del fotoperíode, tals com els phyA, B, D i E i els cry1 i 2 (Devlin and Kay 2000).
- Floració: La floració està regulada entre altres condicions pel fotoperíode i pel rellotge
intern. Diversos reguladors transcripcionals actuen sobre integradors de la resposta floral els
quals, al seu torn, alteren l’expressió de gens d’identitat del meristem per promoure la floració
(Turck et al. 2008). Arabidopsis mostra una floració accelerada en fotoperíodes de dia llarg (LD,
de l’anglès long-day). Anàlisis en mutants de fotoreceptors han mostrat que phyA i cry2
intervenen en la percepció d’aquests fotoperíodes llargs (recordem que són fotolàbils i només
s’acumulen en les hores de foscor). Estudis en el temps de floració mostren que mutants en
phyA mostren insensibilitat a tractaments d’allargament de la durada del dia i de trencament del
període de nit, confirmant un paper per aquest fitocrom en la percepció del fotoperíode (Reed
et al. 1994). Aquestes dades estan recolzades per la floració tardana en el mutant phyA crescut
en LD (Neff and Chory 1998). El phyB actua redundantment amb phyD i phyE reprimint la
floració en condicions de llum normal (amb elevada raó R:FR), a través de la repressió de
l’expressió de l’integrador floral FT (de l’anglès FLOWERING TIME) (Halliday et al. 2003). Per
tal de deixar de reprimir FT cal una exposició prolongada a baixes raons R:FR, assegurant que
no es dóna una transició a la floració en resposta a canvis en l’ombreig de poca durada.
Mutants deficients en phyC crescudes en dies curts mostren una lleugera acceleració en la
floració, suggerint un paper del phyC en la regulació del temps de floració en Arabidopsis
(Monte, et al. 2003).
- La síndrome de fugida de l’ombra: La síndrome de fugida de l’ombra (o SAS, de
l’anglès Shade Avoidance Syndrome) és l’estratègia que tenen moltes plantes per detectar

1 Introducció
17
aquelles plantes veïnes que podrien competir amb elles pels recursos, especialment els
lumínics, i intentar superar aquesta competència canviant el desenvolupament, per exemple
accelerant el creixement en vertical (Smith and Whitelam 1997). Una planta que creix en
condicions on la SAS és activada mostra una elongació de les tiges i els pecíols, un increment
de dominància apical i un allargament de les fulles que disminueixen la seva superfície i, si
aquest major creixement no suposa la millor captació de llum, es produeix una acceleració de
la floració. Així intenta créixer més que les plantes veïnes i obtenir millor posicionament per
captar la llum del Sol, però si no pot, es reprodueix ràpidament amb l’esperança que alguna de
les llavors trobi millor localització per créixer. Canvis en les llums R i FR percebudes pels
fitocroms són claus per aquestes respostes. Estudiar més en profunditat aquests mecanismes
és l’objectiu principal en el laboratori on s’ha realitzat la present tesi, per això es passa explicar
més àmpliament el que es coneix de la SAS, tant a nivell fisiològic com molecular.
1.6. Aspectes generals de la SAS: Quan créixer esdevé la
prioritat.
Com tots els éssers vius les plantes han evolucionat per competició amb els altres
organismes en la duresa de la selecció natural. No obstant, les plantes com a organismes
sèssils, que no es poden moure, no poden triar a quin lloc créixer, sinó que han d’adaptar-se al
lloc on neixen i competir si cal amb la vegetació veïna. Quan una planta creix envoltada d’altres
vegetals està obligada a competir pels recursos de l’ambient, i un dels més importants és la
llum. Si la planta té prou espai lliure al seu entorn, en poca densitat de massa forestal, la llum
que rep directament del Sol és relativament constant (Smith 1982, Vandenbussche et al. 2005).
Però si la planta creix amb abundant vegetació veïna hi ha diversos canvis en la llum que
percep, bàsicament canvis en la quantitat o en la qualitat. Els canvis en la quantitat de llum són
fàcilment explicables quan la planta es troba ombrejada, ja que la vegetació veïna absorbeix la
llum blava i vermella (de 400 a 700 nm) per la fotosíntesis i li impedeix rebre les mateixes
intensitats de llum del Sol que rebria normalment (Keller et al. 2011). D’altra banda, els canvis
en la qualitat de llum són bàsicament deguts a canvis en la raó R:FR, tant per la disminució de
llum R específicament absorbida per les plantes veïnes com per l’enriquiment de llum FR
reflectida per aquestes (Smith and Whitelam 1997) (figura I.3). Aquestes reduccions en la raó
R:FR es poden donar sense necessitat que es produeixi un ombrejat real, tal com ocorre per la
proximitat de vegetació veïna que eventualment podria provocar una competència pels
recursos, i desemboca en una resposta que es pot considerar un “atac preventiu”, donant lloc a
l’activació i posta en marxa de la SAS abans de l’ombrejat real. En qualsevol dels dos casos,
els canvis de llum són percebuts pels fotoreceptors i activen una sèrie de respostes comunes.
No obstant, la distinció entre aquestes dues senyals és important, ja que els fotoreceptors
implicats són diferents, en el primer cas els criptocroms i fitocroms i en el segon exclusivament
els fitocroms, i per tant els mecanismes moleculars poden ser diferents (Franklin 2008, Keller,
et al. 2011, Pierik et al. 2009). En la literatura s’intenta diferenciar els dos mecanismes parlant
de respostes de la SAS en proximitat o en ombra vegetal.

1 Introducció
18
Per distingir entre els efectes de la quantitat i la qualitat de llum aquests paràmetres
s’han de modificar per separat al laboratori. Per mimetitzar l’efecte de la baixada de raó R:FR el
que es fa és enriquir una font de llum blanca (W) amb una làmpada de llum FR (W+FR),
tractament que denominem ombra simulada. La majoria dels experiments de la present tesi es
refereixen a aquesta condició experimental. El mateix tipus de respostes són activades quan
les plantes que creixen en condicions de fotoperíode rebent un tractament amb llum enriquida
en FR a final del dia (EOD-FR, de l’anglès End Of Day-FR) (Fankhauser and Casal 2004). Tots
dos tractaments provoquen una reducció en la raó R:FR percebuda pels fitocroms. Aquests
tipus de respostes són reversibles amb tractaments equivalents amb llum R (EOD-FR vs. EOD-
R o llum de baixa raó R:FR vs. alta raó R:FR). Altres laboratoris estudien les respostes en
condicions d’ombrejat disminuint les intensitats de llum blava i vermella, de manera similar al
que seria l’ombra real de la vegetació. Diverses de les respostes fisiològiques són comunes en
ambdós tipus de senyal, si bé les vies moleculars semblen ser parcialment independents però
confluint en l’alteració d’activitat d’alguns PIFs (Keller, et al. 2011). L’estudi en aquestes
condicions és tema d’altres laboratoris, per tant en la present tesi ens centrarem en les
respostes a la proximitat vegetal.
SASactiva
SASinactiva
alta R:FR(SAS inactiva)
baixa R:FR(SAS activa)
(a) (b)
SASinactiva
SASactiva
Aquests tractaments que simulen la proximitat de vegetació real són una manera senzilla
i eficaç d’activar les respostes de la SAS i diferenciar els diferents orígens moleculars de les
respostes que provoquen (Fankhauser and Casal 2004). Mantenir aquesta singularitat de
senyal facilita els treballs posteriors d’anàlisis fisiològics i moleculars. S’ha vist, per exemple,
que a més dels criptocroms i els fitocroms algun altre receptor podria estar implicat en algunes
respostes relacionades amb la SAS. Experiments amb estimulació física, tocant els extrems de
les fulles de les plantes, han demostrat que aquestes fulles estimulades presenten respostes
semblants a la proximitat de vegetació veïna, fins i tot abans de percebre canvis en les
Figura I.3. La síndrome de fugida de l’ombra (SAS). (a) Esquema que representa les
condicions naturals que alteren la raó R:FR i desencadenen la SAS. La planta de l’esquerre rep
llum directa del Sol amb una alta raó R:FR i creix amb la SAS inactiva. La planta de la dreta té
vegetació veïna que absorbeix la llum R i reflexa la FR, amb una raó final R:FR més baixa, el
canvi en la llum és detectat pels fitocroms i activa la SAS. (b) Fenotips de les plàntules a 7 dies i
de plantes adultes amb las SAS inactiva (esquerre) o activa (dreta).

1 Introducció
19
condicions de llum (de Wit et al. 2012). Aquestes respostes només es donarien en plantes en
estadis prou avançats de desenvolupament, desconeixent-se encara quins serien els
mecanismes moleculars. Per tant, les respostes a ombra venen determinades per l’espècie,
l’òrgan i un factor temporal i de desenvolupament de l’organisme.
La primera resposta a ombra és la germinació, de la qual ja hem parlat anteriorment.
Hem comentat que la germinació està fortament regulada pel phyA, que imposa una segona
dormància a la llavor si hi ha una baixa raó R:FR (Smith and Whitelam 1997). Però la
germinació també ve determinada per les condicions de la planta mare, que quan detecta
aquestes condicions de llum desfavorables transmet aquesta informació a la llavor variant les
proporcions de fitocroms d’aquesta (Dechaine, et al. 2009).
En estadi de plàntula, un parell de dies de tractament en baixa raó R:FR són suficients
per iniciar canvis clarament visibles com l’elongació de l’hipocòtil i dels pecíols o l’allargament i
moviment cap amunt (hiponàstic) dels cotiledons i les fulles primàries (Smith and Whitelam
1997). L’hipocòtil és possiblement l’òrgan més estudiat en resposta a ombra, ja que és molt
fàcil de mesurar i manté una bona correlació amb el senyal lumínic. L’elongació dels pecíols
també és mesurable i manté una resposta clara amb els tractaments d’ombra durant totes les
etapes de desenvolupament de les fulles (Djakovic-Petrovic et al. 2007, Tao et al. 2008).
En etapa de roseta l’ombrejat provoca diverses respostes: s’observen menor nombre de
fulles abans de florir, un increment en la raó llargada-amplada d’aquestes i els pecíols són més
llargs. Aquest allargament dels pecíols provoca a més un moviment hiponàstic de les fulles
(Franklin 2008), per tal d’elevar-les i sobreposar-se a la vegetació veïna. Aquest moviment
també s’observa en plantes creixent en ombrejat real (amb menor llum blava) o quan les fulles
entren en contacte físic amb les fulles de la planta veïna (de Wit, et al. 2012, Keller, et al.
2011). Una altra de les respostes en aquest estadi de desenvolupament és el menor nombre de
ramificacions mitjançant la supressió del creixement de la gemma axil·lar (Finlayson 2007,
Gonzalez-Grandio et al. 2013, Smith and Jordan 1994), tant per canvis en la qualitat com la
quantitat de llum.
L’ombrejat i la proximitat vegetal durant llarg temps produeixen una floració primerenca
(Halliday et al. 1994). A més de la floració, respostes a llarga exposició d’ombrejat provoquen
alteracions com l’aclimatació fotosintètica (Boonman et al. 2009, Dietzel and Pfannschmidt
2008, Walters 2005) i canvis en el metabolisme general de la planta (Roig-Villanova 2007).
Activar la SAS requereix una inversió de recursos per part de la planta, fet que requereix
l’abolició d’altres respostes, com una reducció en els pigments de fotoprotecció (com
carotenoides,(Cagnola et al. 2012) o de defensa. Consistentment, plantes creixent en ombra
tenen respostes a estressos biòtics i abiòtics alterades.
És evident que la relació entre les respostes de la SAS i altres mecanismes és molt
important. La planta ha de ser capaç de prioritzar on invertir els seus recursos de manera més
eficient. Per exemple, en baixes temperatures algunes respostes de la SAS s'atenuen (Patel et
al. 2013). La llum R, però no les altres llums, estimulen la resistència a diversos patògens
(Ballare et al. 2012, Islam and Babadoost 2002), suggerint que la senyalització dels fitocroms

1 Introducció
20
està connectada amb els mecanismes de defensa de la planta. Plantes mutants que activen
certes respostes de la SAS constitutivament, com mutants phyAphyB, phyB o csa1
(CONSTITUTIVE SHADE AVOIDANCE 1), mostren una resistència menor a infecció per
Pseudomonas syringae (Faigon-Soverna et al. 2006, Genoud et al. 2002), suggerint un rol
d’aquests fitocroms en la defensa mediada per SA. Els tractaments amb W+FR s’ha vist que fa
plantes d’Arabidopsis i tomàquets més susceptible als herbívors (Izaguirre et al. 2006, Moreno
et al. 2009). Recentment s’ha afegit la susceptibilitat d’Arabidopsis a Botrytis cinerea, també en
pretractaments d’ombra simulada (Cerrudo et al. 2012). En un article recent (de Wit et al. 2013)
demostren que la inducció de la SAS és dominant enfront a la senyalització per defensa,
provocant una caiguda en les rutes de senyalització de JA i SA.
Així doncs, el control hormonal és clau en les respostes de la SAS. De fet, diverses
hormones s’han vist implicades en aquests mecanismes. Però això és entrar en la vessant
molecular del tema, i abans de parlar de les hormones ens cal parlar d’altres components,
altres personatges importants, de la SAS.
1.7. Aspectes moleculars de la SAS: Múltiples personatges per
una resposta complexa.
Anàlisis genètics han mostrat que en Arabidopsis, i també en moltes altres espècies,
mutants phyB tenen hipocòtils molt allargats i floració primerenca, característiques típiques de
plantes creixent en baixa raó R:FR, amb la SAS activa. Per això s’ha dit que els mutants phyB
tenen una SAS constitutiva (López-Juez et al. 1992, Somers et al. 1991). Així, phyB estaria
reprimint la SAS en condicions normals de llum (alta R:FR) i seria el principal regulador de la
SAS pel canvi de les condicions de llum. No obstant el mutant phyB encara respon a ombra,
indicant la participació d’altres fitocroms en la resposta, si bé de manera minoritària (Robson et
al. 1993). L’expressió de PHYA i PHYB està incrementada en tan sols una hora de tractament
d’ombra, i el mutant phyB manté l’augment d’expressió en PHYA, suggerint que altres fitocroms
estan col·laborant en aquesta resposta molecular. Els mutants phyA són més sensibles,
presenten respostes majors, a baixa raó R:FR, suggerint que phyA regula negativament la SAS
(té una activitat contrària a phyB) (Devlin, et al. 2003, Johnson et al. 1994). Tot i que diversos
anàlisis moleculars amb phyA i phyB porten a pensar que ambdós reprimeixen la SAS, la
diferent estabilitat, el diferent patró d’expressió i les diferències en les seves unions a PIFs, fa
que tinguin activitats quasi antagòniques. Anàlisis genètics també han demostrat que phyD i
phyE tenen una activitat similar a phyB en la regulació de la SAS. Tot i que els simples mutants
no tenen cap fenotip, dobles mutants phyBphyD o phyBphyE tenen hipocòtils més llargs i
menor resposta FR que el mutant simple phyB (Devlin, et al. 1998, Devlin et al. 1999). Així
doncs phyB sembla ser el regulador principal de la SAS en plantes desetiolades, amb
aportacions menors de phyA, phyD i phyE.
Com s’ha explicat anteriorment els fitocroms controlen les respostes de la SAS a través
del control sobre l’activitat dels PIFs. Varis PIFs semblen regular quasi totes les respostes en
ombra. PIF4 i PIF5 s’acumulen en foscor i són degradats en llum R i W, mentre que es

1 Introducció
21
reacumulen en W+FR, quan els fitocroms són inactius (Lorrain, et al. 2008). Mutants pif4 i pif5
tenen menor resposta a ombra (hipocòtils més curts), i el doble mutant pif4pif5 encara respon
menys, suggerint que són reguladors positius de la SAS conjuntament (Lorrain, et al. 2008),
dades consistents amb les obtingudes per línies sobre-expressores. A més, triples mutants
phyBpif4pif5 presenten menor fenotip elongat que el mutant simple phyB, revertint parcialment
el seu aspecte de tenir la SAS constitutiva (Hornitschek, et al. 2009). Sembla doncs que els
PIFs promouen l’allargament de l’hipocòtil, de manera equivalent al seu paper en plàntules
creixent en foscor, i que d’igual manera la seva activitat està controlada pels fitocroms.
PIF4 i PIF5 augmenten els nivells d’expressió durant els període de llum, tot i que a nivell
proteic són degradats, i disminueixen en els períodes de foscor, deixant una finestra molt petita
de poques hores abans de l’alba on es poden acumular (Nozue, et al. 2007, Yamashino, et al.
2003). Aquests canvis en els nivells d’expressió són regulats pel rellotge circadià de la planta,
que també regula l’expressió de gens com PIL1 i PIF6, amb increments a l’alba dels dos gens
(Salter et al. 2003), coincidint els pics de màxima expressió amb els pics d’elongació de
l’hipocòtil. Aquestes dades suggereixen que aquests dos PIFs (PIF4 i PIF5), conjuntament amb
PIL1, regulen les respostes a ombra simulada, o com a mínim la resposta en l’hipocòtil, de
manera conjunta amb el rellotge circadià (Lorrain, et al. 2008, Martínez-García, et al. 2010).
PIF7 té un paper bastant important en ombra, a jutjar pel fort fenotip del mutant pif7, que
mostra hipocòtils clarament més curts que el control wt en tractaments d’ombra simulada
(Leivar, et al. 2008, Li, et al. 2012). Mirant l’elongació en ombra per diferents mutants de PIFs
s’ha vist que PIF3 també podria tenir un paper, tot i que minoritari (Leivar, et al. 2012).
L’expressió de gens marcadors d’ombra en mutants de PIF4, PIF5 i PIF7 està atenuada,
de manera consistent amb els resultats fisiològics (Hornitschek, et al. 2012, Leivar, et al. 2012,
Li, et al. 2012). A més, s’ha demostrat mitjançant experiments de ChIP, que el phyB actiu
inhibeix la unió de PIF3 al DNA, i que alguns del gens regulats per PIF3 també estan implicats
en les respostes a ombra (Park, et al. 2012), confirmant que PIF3 també té un paper en
aquestes respostes.
Més enllà dels PIFs, altres gens també regulen de manera important les respostes de la
SAS. Com ja hem comentat, els gens PAR codifiquen factors o cofactors de transcripció que
regulen conjuntament amb els PIFs respostes moleculars com la síntesis o la sensibilitat
d’hormones (Roig-Villanova et al. 2007, Roig-Villanova, et al. 2006). Entre els gens PAR trobem
representants de dues grans famílies de reguladors transcriptionals: les homeodomain-leucine
zipper (HD-Zip) i les bHLH. Del primer grup, del qual en formen part ATHB2 i ATHB4, en
parlarem al capítol I; mentre que del segon grup, del qual en formen part HFR1 i PAR1, ja
n’hem comentat algunes coses en apartats anteriors.
A més d’identificar gens ràpidament regulats per l’acció dels fitocroms al laboratori s’han
identificat quins d’ells són dianes directes. Per això es van realitzar anàlisis de l'expressió
gènica de plàntules d’Arabidopsis tractades amb una hora d’ombra simulada (W+FR).
Mitjançant l’adició de cicloheximida (CHX), un inhibidor de la síntesis de proteïna, es pot
distingir quins gens mantenen els canvis d’expressió mostrant que són els primaris (figura I.4)

1 Introducció
22
(Roig-Villanova, et al. 2006). Així aquests factors PAR serien el punt d’entrada de la senyal
d’ombra a la regulació del desenvolupament i podrien actuar com a nusos de la xarxa
transcripcional que regula les diferents respostes. Entre els gens PAR que són dianes directes
de l’acció dels fitocroms trobem ATHB2, ATHB4, PAR1, PAR2 i PIL1.
(b)
ATHB4
ATHB2
PIL1
HAT2
PAR1
PHYB
RIP
0 1
Col
0 1
No
0 1
Ler
25S
(a)↑R:FR
citoplasma
nucli
gens 2rs
gens 1rs
PIF
PIF
Pfr
PfrPr
↓R:FR
PfrPr
citoplasma
nucli
FT
genes 2rs
CHX
gens 1rs
PIF
PIF
Pfr
Ja hem parlat de PIL1 i HFR1 com a reguladors de les respostes a llum. Els dos van ser
implicats en la SAS com a reguladors negatius (Galstyan, et al. 2011, Roig-Villanova, et al.
2007, Roig-Villanova, et al. 2006, Salter, et al. 2003, Sessa, et al. 2005), basant-se en estudis
amb els seus mutants, ja que mostraven hipocòtils més llargs en ombra simulada. S’ha
demostrat que HFR1, una bHLH atípica que no pot unir DNA (es a dir, una HLH), actua
genèricament com a cofactor transcripcional formant heterodímers amb autèntiques bHLHs i
evitant que aquests uneixin al DNA (Galstyan, et al. 2011); específicament, HFR1 inhibeix les
respostes de la SAS formant heterodímers amb PIF4 i PIF5, evitant que aquests uneixin al
DNA (Hornitschek, et al. 2009). Els nivells d’HFR1 estant controlats tant a nivell transcripcional
(la seva expressió augmenta en ambient amb baixa raó R:FR) com post-trancripcional (és
estable en llum, però es degrada ràpidament en foscor per l’acció de COP1) (Fairchild, et al.
2000, Hornitschek, et al. 2009, Sessa, et al. 2005). Tot i que PIL1 i HFR1 són inhibidors de
Figura I.4. Canvis transcripcionals ràpids induïts pels fitocroms després de la percepció de la proximitat vegetal. (a)
Esquema d’una cèl·lula vegetal crescuda en alta (imatge superior) o baixa raó de R:FR (inferior), indicativa de la proximitat vegetal.
En alta raó R:FR els fitocroms es troben majoritàriament actius i inhibeixen l’activitat dels PIFs, mentre que en baixa raó R:FR els
PIFs són lliures i poden activar gens que poden codificar per factors de transcripció i regular gens secundaris. La cicloheximida
(CHX) inhibeix la síntesis de proteïna de novo i per tant permet diferenciar entre els gens primaris i els secundaris. (b) Efecte de
l’ombra simulada sobre l’expressió de gens PAR. Es va fer extracció d’RNA de plàntules de 3 ecotips d’Arabidopsis (Col-0, No-0,
Ler) de 7 dies crescudes en llum blanca contínua (W) immediatament abans (0) o després d’1 h (1) de tractament amb ombra
simulada (W+FR) per estudiar el seu efecte en l’expressió dels gens indicats. Com a control de càrrega es va fer servir la sonda per
l’rRNA 25S. En vermell es mostren els gens que es van trobar com a gens primaris (1s) de l’acció de fitocroms per tractament amb
CHX. FT: factor de transcripció.

1 Introducció
23
l’elongació de l’hipocòtil (Fairchild, et al. 2000, Salter, et al. 2003, Sessa, et al. 2005), altres
membre de la subfamília 15 de les bHLH, com els PIFs i SPT, promouen la seva elongació
(Penfield, et al. 2005).
Dos altres membres de les bHLHs, PAR1 i el seu homòleg PAR2, semblen actuar de
manera similar a HFR1 controlant les respostes a llum com a cofactors (Galstyan 2010, Hao, et
al. 2012). Codifiquen proteïnes bHLH atípiques, i malgrat que semblen haver perdut la capacitat
d’unir al DNA necessiten estar al nucli per exercir la seva funció biològica (Roig-Villanova, et al.
2007). Plantes sobre-expressores de PAR1 o PAR2 mostren un fenotip nan, més fosc i menor
resposta a ombra, mentre que plantes amb aquests gens silenciats (PAR1-RNAi o par2-1)
tenen respostes incrementades (Roig-Villanova, et al. 2007), fet que porta a pensar que són
reguladors negatius de la SAS.
El fet que molts d’aquests factors, tant reguladors positius com negatius de la SAS,
tinguin nivells d’expressió incrementats en ombra porta a pensar que les respostes a llum són
complexament regulades mitjançant un control prement “accelerador i fre” per evitar una
resposta exagerada o massa prolongada. Així les respostes resultants són una suma dels
diversos reguladors, tant dels PIFs com de les bHLHs (Casal 2013, Martínez-García, et al.
2010). Cal recordar que aquests factors estan sotmesos a controls tant dels seus nivells
d’expressió com de la seva estabilitat a nivell proteic.
Més avall de tota aquesta regulació, hi ha una sèrie de gens equivalents als peons que
fan la feina, gens de creixement i desenvolupament regulats per llum i per altres estímuls, sent
gens secundaris que no responen directament per l’acció dels fitocroms. Per exemple els PRE
(PACLOBUTRAZOL-RESISTANT), també bHLHs atípiques (HLH), promouen l’elongació
cel·lular antagonísticament amb una altra HLH anomenada IBH1 (ILI1 BINDING bHLH
PROTEIN1) (Bai et al. 2012). Un marcador d’ombra com XTR7 (XYLOGLUCAN
ENDOTRANSGLYCOSYLASE 7) afecta l’estructura de la paret cel·lular (Lorrain, et al. 2009).
Alguns gens de la família dels SAUR (SMALL AUXIN-UP RNA, (Hagen and Guilfoyle 2002,
McClure and Guilfoyle 1987) s’ha vist que afecten l’expansió cel·lular (Spartz et al. 2012). Molts
d’aquests gens s’han fet servir sovint com a marcadors d’ombra, ja que els seus nivells
d’expressió es veuen afectats clarament per la SAS, però també responen a altres estímuls.
Per exemple, IBH1 interacciona amb components de brassinoesteroides (BR) com BEE1,
PRE1 va ser identificat com a regulador positiu de respostes a gibberel·lines (GA) i incrementa
la seva expressió en resposta a aquesta hormona (Lee et al. 2006), mentre que PRE6/KDR
uneix HFR1 i PRE1 interacciona amb PAR1 segrestant aquests factors de la seva interacció
amb PIF4 (Bai, et al. 2012). La família dels SAUR va ser identificada per la seva sobre-
expressió en resposta a auxines (Aux), però posteriorment han estat molt usats com a gens de
respostes primerenques a l’activitat de BRs (Sun et al. 2010, Vert et al. 2008, Yin et al. 2005).
S’ha demostrat que alguns SAUR es troben regulats conjuntament per les dues rutes
hormonals (Walcher and Nemhauser 2012).
Així doncs, hi ha una convergència de les diferents rutes en els gens de destí final,
podríem dir que molts encarregats dirigeixen als mateixos peons, que són els que fan la feina,

1 Introducció
24
els que produeixen la divisió o l’expansió cel·lular. En aquest context, la SAS és un mecanisme
que coordina el creixement conjuntament amb altres rutes importants pel desenvolupament de
la planta, entre les quals hi ha les hormonals.
1.8. Relació de la SAS amb altres rutes.
Al llarg de tota la introducció hem comentat múltiples connexions de la senyalització per
llum amb diversos mecanismes de desenvolupament de la planta. Com hem vist en l’apartat
1.5. els fitocroms intervenen en molts processos, fet molt lògic tenint en compte que són els
que reben la informació de la llum i per tant han de guiar el creixement conseqüentment. En
diversos punts hem comentat connexions amb les rutes hormonals, i és que vàries hormones
vegetals regulen la divisió i l’expansió cel·lular modulades per la fotomorfogènesi. AUXs, BRs i
GAs són generalment considerades hormones estimuladores del creixement, mentre les
citoquinines són considerades hormones inhibidores d’aquest i l’etilè té un paper complex,
regulant positivament el creixement en llum i negativament en foscor. No obstant, cap de les
rutes hormonals comparteix la totalitat de les respostes amb la SAS, és a dir, que tractaments
amb hormones o fenotips de mutants en vies hormonals no tenen mai un comportament igual al
tractament d’ombra. Aquestes fitohormones, juntament amb altres mecanismes de la planta
com el rellotge circadià o les respostes a canvis de temperatura, poden actuar en el mateix
sentit o antagonísticament en les diferents respostes a la llum. Aquestes connexions entre
senyals interns i externs formen una extensa xarxa de senyalització que permet integrar
informació de l’ambient amb el desenvolupament vegetal, de manera que els treballadors
d’aquesta xarxa saben en tot moment si poden parar per esmorzar o han de treballar
intensament per tal d’accelerar la divisió cel·lular, si han de donar un cop de mà a un company
o desplaçar-se a una altra regió, un altre teixit, per seguir amb la seva feina.
- GIBBEREL·LINEs (GAs): Durant l’explicació de l’activitat dels PIFs hem comentat que
les proteïnes de la família DELLA podien interaccionar amb PIF3 i PIF4 inhibint la seva activitat,
fent de node entre senyal lumínic i respostes a hormones. Això és degut a que les proteïnes
DELLA van ser originalment identificades com a components de la senyalització de GA, i
classificades com a repressores de respostes de creixement en ombra, estrès salí i en
tractaments de fred (Achard et al. 2008, Achard et al. 2009, Navarro et al. 2008). Moltes
d’aquestes respostes es troben compartides amb PIF4, fet que porta a pensar que com a
mínim part d’elles podria venir donada per la inactivació de PIF4 per acció de les DELLA (de
Lucas, et al. 2008, Feng, et al. 2008).
En Arabidopsis la família DELLA està codificada per cinc membres: GAI (de l’anglès GA
INSENSITIVE), RGA (REPRESSOR OF GA1), RGL1 (RGA-LIKE 1), RGL2 i RGL3, tenint tots
ells una petita seqüència aminoacídica conservada DELLA, la qual els dóna el nom (Bolle
2004). Les proteïnes DELLA es localitzen al nucli, on reprimeixen l’expressió de gens de
resposta a GAs. Tanmateix, en presència de GA les proteïnes DELLA es degraden (Achard
and Genschik 2009), donant lloc a l’activació de gens de creixement en resposta a GA, alguns
dels quals podrien estar compartits amb les respostes activades pels PIFs. L’estabilitat de les

1 Introducció
25
proteïnes DELLA i la seva degradació es veuen afectades per vàries senyals, tals com altres
hormones (auxines i etilè), la llum i condicions d’estrès d’origen biòtic i abiòtic (Achard and
Genschik 2009, Swain and Singh 2005). Aquest fet suggereix que les proteïnes DELLA poden
actuar com a integradors de senyals de varies rutes de senyalització que regulin el creixement.
Anàlisis genètics amb mutants de guany i pèrdua de funció de les DELLA han implicat a
la senyalització per GA en la regulació de les respostes de la SAS (Djakovic-Petrovic, et al.
2007). Una major estabilitat de les DELLA, com passa en el mutant gait6, dóna una resposta
menor en tractaments W+FR, suggerint el seu rol en la inhibició del creixement induït per la
proximitat vegetal. El mutant quàdruple de pèrdua de funció gairgargl1rgl2 (della4) respon
normalment a la baixa raó R:FR (Djakovic-Petrovic, et al. 2007), donant a entendre que: (1)
RGL3 té un paper destacat en respostes a ombra, de manera individual o redundant amb algun
altre membre; o (2) Les respostes de creixement de la SAS no depenen exclusivament de les
DELLA i per tant, hi ha altres reguladors del creixement en resposta a ombra independents.
Estudis en el laboratori han permès confirmar la segona hipòtesi, sense haver descartat la
primera (Cifuentes-Esquivel 2012).
Tractaments d’ombra simulada incrementen ràpidament l’expressió de gens de
biosíntesis de GAs (Cifuentes-Esquivel 2012, Hisamatsu et al. 2005). No obstant, no està clar
que els nivells de GAs actives després del tractament d’ombra simulada siguin globalment més
alts, i aquests tampoc es veuen incrementats en mutants deficients en fitocroms, de manera
que s’hipotetitza que la resposta ve determinada, com a mínim parcialment, per l’alteració de la
sensibilitat a GA en els teixits corresponents (Lopez-Juez et al. 1995, Reed et al. 1996).
En la regulació de la SAS, doncs, la llum controla el creixement de manera coordinada
amb les GAs, alterant alhora la seva síntesis i la sensibilitat local, de tal manera que en
condicions on la SAS és activa les DELLA seran més fàcilment degradades, alliberant els PIFs i
permetent-los regular els gens de creixement.
- AUXINEs (AUXs): Diversos aspectes relacionen la senyalització de les auxines amb
les respostes de la SAS. Fisiològicament les AUXs regulen múltiples aspectes del creixement
vegetal com l’expansió dels cotiledons i les fulles, l’elongació de l’hipocòtil i la tija i la
dominància apical, processos característics de la SAS.
De manera semblant a la senyalització per GAs, la inducció de creixement per les
auxines ve donada per la degradació en presència d’aquesta hormona d’una sèrie de proteïnes
que actuen com a repressores de la transcripció (les Aux/IAAs), sense que aquests repressors
uneixin al DNA (Gray et al. 2001, Ulmasov et al. 1997). Al degradar-se aquestes proteïnes
repressores, les proteïnes ARFs (AUXIN RESPONSE FACTORS), que si que poden unir al
DNA, són lliures per fer la seva funció, normalment com a activadores transcripcionals
(Ulmasov et al. 1999a, Ulmasov et al. 1999b).
L'expressió de varis gens de resposta a auxines (IAAs i SAURs, per exemple), així com
la família de transportadors d'auxines PIN (PIN3 i PIN7) canvia ràpidament després de
l'exposició a la proximitat vegetal (ombra simulada) (Devlin, et al. 2003, Nemhauser et al. 2006,
Tao, et al. 2008) o l’ombra vegetal (Carabelli et al. 2007, Sessa, et al. 2005).

1 Introducció
26
Els darrers anys, mitjançant experiments de ChIP, s’ha demostrat que PIF4, PIF5 i PIF7
modulen l’expressió de gens relacionats amb les auxines unint els promotors de gens de síntesi
i sensibilitat a auxines (Hornitschek, et al. 2012, Li, et al. 2012), produint un increment dels
nivells d’auxina activa. Cal recordar que aquests factors PIFs s’acumulen en condicions
d’ombra, quan la majoria dels fitocroms són inactius. Consistentment, s’ha demostrat que una
reducció en els nivells de phyB actiu, provocada per una baixa raó R:FR, produeix un increment
ràpid en els nivells endògens d’àcid indol-3-acètic lliure (IAA), una AUX activa en plantes. A
més, el mutant pif4pif5 té menor acumulació d’AUXs en ombra (Hornitschek, et al. 2012).
En un crivellatge on es buscaven mutants amb respostes nul·les a ombra es va veure
que aquest increment d’AUX activa implica l’acció de SAV3 (SHADE AVOIDANCE 3)/TAA1
(TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1). Els mutants sav3 tenen nivells
d’IAA reduïts i pràcticament no responen al tractament d’ombra simulada. Es va veure que
SAV3 codifica per un enzim de la biosíntesi d’auxines requerit per l’increment ràpid dels nivells
d’IAA en resposta a ombra simulada (Tao, et al. 2008), si bé Arabidopsis té altres rutes de
síntesi d’auxina dependents de triptòfan i que semblen independents de llum (Stepanova et al.
2011), i una altra ruta poc caracteritzada independent de triptòfan (Östin et al. 1999).
Altres mutants de la via d’auxines també tenen respostes a ombra disminuïdes, com els
mutants en TIR1 (TRANSPORT INHIBITOR RESPONSE 1) o AXR1 (AUXIN RESISTANT 1,
enzim que regula l’estabilitat dels transportadors d’AUX) (Pierik, et al. 2009).
A més, s’ha vist que la percepció de la llum té una gran influència en el transport
(distribució al llarg de la plàntula) i la resposta (sensibilitat) a AUXs (Halliday and Fankhauser
2003). Tractaments de plàntules wt amb un inhibidor del transport d’auxines com l’àcid
naftilftalàmic (NPA) suprimeixen la resposta de l’hipocòtil a la SAS (Sorin et al. 2009, Steindler
et al. 1999). Mutants en el transport d’auxines, com pin3-3 i axr1-12, també mostren unes
respostes atenuades en ombra (Keuskamp et al. 2010, Pierik, et al. 2009).
El transport de les auxines està sent estudiat per diversos grups. El conjunt
d’experiments porta a pensar que gran part de les auxines són sintetitzades en les fulles i
cotiledons i transportades a través de l’hipocòtil fins a les arrels. Tractaments en ombra han
demostrat alteracions en la formació d’arrels laterals (Bhalerao et al. 2002, Salisbury et al.
2007), resultats similars als obtinguts amb tractaments amb inhibidors del transport d’AUXs.
Aquests experiments suggereixen que l’exposició de plàntules a ombra simulada produeix una
reducció del transport d’AUX a través de l’hipocòtil, afavorint que les capes laterals d’aquest
teixit acumulin l’hormona provocant el seu allargament i una reducció en els nivells d’AUX que
arriba a les arrels (Morelli and Ruberti 2002). Aquests experiments connecten la percepció
d’ombra amb el transport d’AUX. De manera consistent, mitjançant una línia DR5:GUS on el
promotor s’indueix en presència d’AUX, es va veure que el tractament d’ombra simulada
provocava un canvi en la distribució de l’expressió GUS a la línia reportera (Sabatini et al. 1999,
Tao, et al. 2008). Més concretament, plàntules DR5:GUS mostren una activitat GUS
incrementada en el terç inferior de l’hipocòtil i als cotiledons, i una reducció de l’expressió a les
arrels (Roig-Villanova 2007, Salisbury, et al. 2007, Tao, et al. 2008) indicant que, després de la

1 Introducció
27
detecció de proximitat vegetal, hi ha una redistribució de les AUXs possiblement per la
modificació del seu transport.
Cal destacar que diversos estudis han posat de manifest que les AUXs i les GAs es
coordinen per regular respostes de creixement (Chapman et al. 2012, Frigerio et al. 2006,
Stavang, et al. 2009). A més, els nivells d’AUXs també es veuen controlats per rellotge circadià,
com es pot veure per les variacions circadianes en quantificacions d’IAA en inflorescències
d’Arabidopsis (Covington and Harmer 2007, Jouve et al. 1999).
En la regulació de la SAS, doncs, la llum controla el creixement de manera coordinada
amb les AUXs, incrementant la seva síntesis, modificant el seu transport i possiblement
regulant també la seva sensibilitat, tot i que de moment només es coneixen part dels
mecanismes moleculars implicats.
- BRASSINOESTEROIDEs (BRs): Els BRs són hormones esteroides vegetals que
regulen molts processos de creixement i desenvolupament, com l’elongació cel·lular, el
desenvolupament vascular o la senescència i estan involucrades en processos com les
respostes a l’estrès i la fotomorfogènesi (Li and Chory 1999). Mutants de pèrdua de funció en
les vies de síntesi o de sensibilitat a BRs causen plàntules desetiolades a la foscor, així com
fenotips pleiotròpics nans (Clouse and Sasse 1998, Li and Chory 1999), presentant rosetes
compactes de fulles verd fosc i epinàstiques, amb una elongació reduïda dels hipocòtils,
pecíols i tiges i una disminució de la dominància apical, senescència i de la fertilitat del pol·len
(Yin, et al. 2005).
A diferència de les GAs o les AUXs, la senyalització per BRs ve donada per
l’estabilització de factors de transcripció nuclears en resposta a l’hormona (Wang et al. 2002,
Yin et al. 2002). La percepció de BR produeix una inhibició de l’activitat de BIN2, un regulador
negatiu de la senyalització de BRs (Li et al. 2002), que, en baixa concentració d’hormona,
degrada factors de transcripció com BZR1 (BRASSINAZOLE RESISTANT1) i BES1/BZR2
(BRI1 EMS SUPPRESSOR1). BZR1 uneix promotors de gens de la síntesis de BRs reprimint-
ne l’activitat (He et al. 2005), de manera que estableix una auto-regulació negativa dins la
senyalització de BRs. A més, BZR1 heterodimeritza amb PIF4, regulant conjuntament gens de
resposta a hormones i llum (Oh, et al. 2012). BES1 actua sol o interaccionant amb proteïnes
bHLH conegudes com BIMs (BES1-INTERACTING MYC-LIKE) i activa la transcripció de gens
de resposta a BRs (Yin, et al. 2005).
Mutants de la síntesi de BRs, com det2-1 i dwf1, presenten respostes de la SAS
disminuïdes (Luccioni et al. 2002, Martínez-García, et al. 2010), suggerint que els BRs són
necessaris per aquestes respostes. Recentment components de la via de senyalització com els
BEEs (BR-ENHANCED EXPRESSION) i BIMs, que tenen nivells d’expressió induïts en ombra,
han estat definits com a reguladors positius de la SAS (Cifuentes-Esquivel et al. 2013). Triples
mutants bee1bee2bee3 (anomenats bee123) i bim1bim2bim3 (bim123) tenen la resposta a
ombra alterada amb hipocòtils més curts després del tractament. A més, aquests factors
interaccionen amb PAR1, suggerint que PAR1 podria regular respostes hormonals i a llum
conjuntament mitjançant aquests gens diana.

1 Introducció
28
En la regulació de la SAS, doncs, la llum controla el creixement de manera coordinada
amb els BRs, possiblement per la regulació de dianes conjuntament entre BZR1 i PIF4 i per la
inactivació dels BEEs i BIMs mitjançada per PAR1. Si bé encara no està clar com canvis en la
llum podrien afectar també la síntesis o sensibilitat a BRs.
- ETILÈ: L’etilè és una hormona molt volàtil, que les plantes usen sovint per comunicar-
se entre elles. Plantes transgèniques de tabac insensibles a l’etilè mostren una reducció de les
respostes a ombrejat real, suggerint un paper important d’aquesta hormona gasosa en la
detecció de la vegetació que es troba molt propera (Knoester et al. 1998). De manera
consistent, l’aplicació d’etilè exogen a plantes silvestres indueix respostes semblants a les de la
SAS (Pierik et al. 2004b, Pierik et al. 2004c). Curiosament, quan aquestes plantes de tabac
insensibles a l’etilè creixen en ombra simulada (baixa raó R:FR) responen de manera normal al
tractament, suggerint que l’ombra simulada pot induir respostes de la SAS independentment de
l’acció de l’etilè (Pierik, et al. 2004b). S’ha demostrat que els efectes inductors del creixement
de l’etilè venen donats en part per EIN3 (ETHYLENE-INSENSITIVE 3), que activa l’expressió
de PIF3 unint directament al seu promotor (Zhong, et al. 2012). A més, s’ha vist que algunes de
les respostes depenen de GA i AUX (Pierik et al. 2004a, Pierik, et al. 2009) i que l’etilè regula
l’estabilitat de les proteïnes DELLA (Achard and Genschik 2009, Achard, et al. 2009), de
manera que coopera amb diferents rutes hormonals per tal de controlar el creixement.
- ALTRES HORMONES (JA, SA, CK...): El paper d’altres hormones en les respostes de
la SAS és o poc rellevant o està poc estudiat. No obstant, la integració de les diverses senyals
ambientals en rutes hormonals sembla que és un fet afavorit per tal de tenir una major
coordinació entre elles, de manera que no és menyspreables que altres hormones puguin
afectar respostes típiques d’ombra.
L’àcid jasmònic (JA), descrit com a important regulador de les respostes de defensa,
retarda la degradació de les DELLA i indueix l’expressió de RGL3. RGL3, conjuntament amb
altres DELLAs, reprimeix l’activitat de les proteïnes JAZ (JA Zim-domain), que són repressores
de la senyalització de JA (Wild et al. 2012, Yang et al. 2012). A més, la baixa raó R:FR
reprimeix les respostes a patògens de manera depenent de JAZ10 (Cerrudo, et al. 2012),
rebaixant la sensibilitat a JA de manera independent a la senyalització d’àcid sal·licílic (SA),
una altra hormona que regula les respostes a infecció i que també té la senyalització
disminuïda en respostes a ombra. A més, JAZ9 interfereix en la interacció de RGA amb PIF3,
de manera que hi ha una cadena de senyal JAZ-DELLA-PIF (Yang, et al. 2012).
Si bé la majoria de respostes a ombra produeixen major creixement, en les fulles hi ha
una menor divisió cel·lular, procés que està regulat per les citoquinines (CKs). Un regulador
negatiu d’aquesta hormona, CKX6, té nivells incrementats d’expressió en tractaments d’ombra i
d’AUXs (Carabelli, et al. 2007, Werner et al. 2003), suggerint que el menor creixement de la
fulla en resposta a ombra és deguda tant per la resposta a llum directa com per l’augment
d’AUXs posterior, mitjançant una caiguda local dels nivells de les citoquinines (Alabadi, et al.
2008, Vandenbussche et al. 2007).

1 Introducció
29
Així doncs, les interaccions locals de les diferents hormones i els diferents components
de la senyalització de llum són claus per les respostes finals.
- RELACIÓ ESPAI-TEMPS: Fins ara hem donat a entendre que la majoria de reguladors
de la SAS i les seves múltiples connexions amb altres xarxes actuen conjuntament, que poden
trobar-se els components en els mateixos llocs i al mateix temps. Pràcticament tots els resultats
mostrats sobre la regulació i els mecanismes moleculars involucrats en la SAS provenen
d’experiments realitzats amb plàntules senceres, però els models que descriuen l’acció dels
fitocroms i el seu efecte en l’expressió gènica estan plantejats a nivell cel·lular, i per tant poden
ser dependents del tipus cel·lular, del teixit on ens situem. Tot i que anàlisis d’expressió
temporal i espacial porten a concloure que els fitocroms són presents en tots els teixits
analitzats (Josse et al. 2008, Montgomery 2008, Montgomery and Lagarias 2002), diverses
proves porten a pensar que la interacció de tots els components no és rellevant per a tota la
planta, sinó que alguns reguladors tenen papers específics en determinats teixits i/o només en
alguns estadis de desenvolupament.
Mentre que factors com ATHB2, ATHB4, BEEs, PAR1 i PAR2 sembla que diverses
respostes de la SAS, com l’elongació de l’hipocòtil, l’expansió dels cotiledons i les fulles
primàries, altres factors com HFR1 i BIMs regulen l’elongació de l’hipocòtil, però no semblen
tenir cap efecte en l’expansió dels cotiledons o fulles primàries (Cifuentes-Esquivel, et al. 2013,
Hornitschek, et al. 2009). A més, la sobre-expressió d’ATHB2 provoca hipocòtils més llargs,
però no altera el temps de floració. COP1 i SPA tampoc afecten el temps de floració, si bé són
dos factors essencials per les respostes d’elongació en ombra (Rolauffs et al. 2012). No
obstant, mutants defectius en SPA tenen respostes moleculars alterades mirant l’expressió de
diversos gens marcadors com XTR7, YUC2, YUC7 i YUC8, però l’efecte sobre aquests és
major que sobre altres marcadors com PIL1, ATHB2 i HFR1. Donat que tots aquests gens són
dianes de PIFs, és possible que puguin estar regulats per diferents PIFs, suggerint certa
especificitat entre els marcadors moleculars i la regulació de les diverses respostes. De manera
semblant a la floració, la ramificació sembla estar regulada independentment d’altres aspectes
de la SAS, vist que mutants en BRC1 (BRANCHED1) no afecten l’elongació de l’hipocòtil, però
si la ramificació en resposta a ombra (Gonzalez-Grandio, et al. 2013). Mutants prr5 (pseudo
response regulator5, un membre del rellotge circadià) mostren pecíols més llargs en ombra,
però un creixement no disminuït de la fulla (Takase et al. 2013). La resposta en els pecíols és
justificada mirant nivells de gens marcadors d’ombra com ATHB2 i HFR1, però el creixement
normal en la fulla és més difícil d’explicar pels canvis moleculars, ja que gens com PIF4 i PIF5
tenen nivells d’expressió més elevats com seria d’esperar després de la percepció d’ombra.
D’aquesta manera, PRR5 sembla ser requerit per la resposta fisiològica a ombra sobretot en la
fulla, mentre que la seva falta produeix un efecte invers, hipersensibilitat a ombra, en els
pecíols.
A nivell hormonal també s'ha descrit una gran importància en la localització de la senyal.
Per exemple, el desenvolupament regulat per AUXs està controlat per tres etapes: (1) el lloc de
síntesi de l’hormona (en respostes a llum normalment les fulles); (2) el transport de l’hormona a

1 Introducció
30
través dels teixits, amb la capacitat de generar un gradient (per exemple a través de l’hipocòtil)
o l’acumulació local; i (3) el nivell de percepció de l’hormona en cada teixit (Chandler et al.
2009).
Així doncs, hem vist que les respostes finals no són les mateixes en cada teixit ni per
cada període de desenvolupament de la planta. Aquestes dades porten a concloure que la SAS
és una resposta molt complexa que regula els diferents components a nivell local, actuant de
manera coordinada i mitjançant una comunicació entre els diferents teixits. Això suggereix que
la senyal d’ombra implica com a mínim dos passos: (1) una senyalització intracel·lular, on els
fitocroms inicien una cascada de senyalització; i (2) una transducció de la senyal més enllà del
teixit, de manera intercel·lular (Bou-Torrent, et al. 2008). D’aquesta manera la percepció de la
senyal lumínica (els canvis ràpids moleculars) i el lloc d’acció (els trets fisiològics alterats)
poden estar separats físicament, posant de manifest la necessitat d’entendre aquesta
comunicació a llarga distància per saber com es desenvolupa la planta durant la SAS
(Martínez-García, et al. 2010).
1.9. I això on va a parar? Aplicacions biotecnològiques.
Hem vist que els fotoreceptors regulen àmpliament el desenvolupament i el creixement
de la planta, per tant, també controlen molts dels trets d’interès en l’agricultura moderna. El
coneixement sobre la regulació molecular d’aquests trets pot portar a grans millores, tant en la
productivitat com en la qualitat del producte final.
En termes generals les respostes de la SAS resulten en un redireccionament dels
recursos energètics de la planta, invertint-los en el creixement direccional i rebaixant el tamany
dels òrgans d’emmagatzematge de la planta, que sovint són els òrgans d’interès agronòmic.
Aquestes respostes s’activen en situacions d’ombrejat real, però també de manera preventiva
per la proximitat vegetal. La primera idea és doncs eliminar o disminuir aquestes respostes a la
vegetació veïna en camps de conreu on, gràcies a les tècniques agronòmiques, les plantes no
han de patir competència per la llum (Smith 1982).
El primers intents es van fer modificant els nivells d’expressió dels fotoreceptors, però
això va donar lloc a gran diversitat de canvis, molts d’ells no desitjats. Per exemple, tot i que la
sobre-expressió de phyB en patateres transgèniques va augmentar la producció de tubercles,
la mida d’aquests era més petita (Thiele et al. 1999), i per tant no van resultar millors per
cultivar. Un altre cas va ser la sobre-expressió de phyA de civada en àlber (Populus alba), que
va donar lloc a àlbers nans amb una resposta reduïda a l’ombra simulada, possibilitant el cultiu
de més plantes en menys espai. No obstant, es va veure que aquests àlbers eren més
susceptibles a congelació degut a la pèrdua de les respostes d’EOD-FR (Olsen et al. 1997),
havent de descartar també aquesta millora genètica.
Així, millorar el coneixement dels mecanismes moleculars implicats en la SAS podria ser
de gran ajuda en programes d’obtenció de noves espècies o varietats de cultiu, permetent, per
exemple, sembrar més plantes en menys espai o obtenint noves línies que estiguin més
adaptades a les hores d’insolació del territori, a les seves temperatures o la seva humitat

1 Introducció
31
(Kebrom and Brutnell 2007). Però per això és clau identificar els gens que actuen per sota de
l’acció dels fitocroms i que afecten només a un grup de caràcters determinat, per tal de poder
modificar d’una manera més focalitzada els trets d’interès de les plantes en resposta a la llum
(Neff et al. 2000), sense alterar-ne la seva fisiologia general o les altres respostes adaptatives.


2 Objectius
33
2. OBJECTIUS L’objectiu general d'aquest treball és entendre a nivell molecular la regulació de la
síndrome de fugida de l’ombra en plantes mitjançant Arabidopsis thaliana com a organisme
model. El meu projecte es centra en caracteritzar els mecanismes moleculars d’acció d’alguns
dels factors identificats anteriorment al laboratori. Per això ens vam proposar els següents dos
objectius:
1. Estudi de la relació estructura-funció del factor de transcripció ATHB4. Anàlisis
dels mecanismes moleculars d’ATHB4 mitjançant estudis de la seva capacitat d’unió al DNA, la
identificació de dianes directes i amb la generació de plantes transgèniques per a la sobre-
expressió de formes truncades derivades d’ATHB4, per tal d'analitzar l’activitat biològica i
molecular dels diferents dominis d’aquest factor.
2. Estudi del rol de les nucleoporines (NUPs) en la regulació de la SAS. Identificació
i caracterització de mutants en gens que codifiquen autèntiques i possibles NUPs. Estudis de la
resposta a ombra simulada d’aquests mutants per avaluar la implicació de les NUPs en la
regulació de la SAS. Aproximacions per veure si tenen afectat el transport de molècules a
través de l'envolta nuclear, tant en l’export d’RNA com en l’import de proteïna.


3 Capítol 1
35
3.1. Capítol 1: Estudi de la relació estructura-funció d’ATHB4.
3.1.1. Introducció.
3.1.1.1. La família HD-Zip: Components del desenvolupament.
A la introducció general s’ha comentat la identificació de gens PAR (Roig-Villanova
2007). Entre aquests, es van analitzar gens prèviament descrits per altres autors, com
ATHB2/HAT4 (ARABIDOPSIS THALIANA HOMEOBOX-LEUCINE ZIPPER PROTEIN 2 /
HOMEOBOX PROTEIN 4), ATHB4 (Carabelli et al. 1993), HAT1/JAB i HAT2 (Sawa et al.
2002). Mitjançant el tractament amb CHX, es va demostrar que tant ATHB2 com ATHB4 eren
dianes directes de l’acció dels fitocroms, per tant eren gens de resposta primària a ombra que
valia la pena estudiar més en profunditat. A més, al mirar els seus nivells d’expressió abans i
després dels nostres tractaments d’ombra es va veure que tots ells estaven induïts, corroborant
que eren gens PAR (Sorin, et al. 2009).
Els HAT i els ATHB són factors de transcripció de la família HD-Zip. Aquesta família
només és present en plantes, si bé la regió gènica homeobox (HB, codificant pel domini
homeodomain, HD) és present en la resta de regnes. Els membres d’aquesta família es
caracteritzen per tenir el domini HD d’unió al DNA seguit d’un domini de dimerització format per
una cremallera de leucines (Leucine Zipper, Zip)(Ariel et al. 2007). S’ha descrit que, en contrast
amb factors que tenen el HB sol, la dimerització dels membres d’aquesta família a través del
Zip és necessària per tal de tenir la conformació adequada per permetre la interacció del dímer
amb el DNA (Sessa et al. 1993, Tron et al. 2001). A més, les regions N-terminal (Nt) contenen
aminoàcids de càrrega positiva que faciliten i estabilitzen de manera notable la unió al DNA,
sense ser rellevants per l’especificitat de seqüència a unir (Palena et al. 2001). La família ha
estat dividida en quatre subfamílies (classe I a IV) segons característiques com l’especificitat
d’unió al DNA (que ve determinada per alguns residus clau del HD), la composició d’introns i
exons i les similituds de seqüència per possessió d’altres dominis (Ariel, et al. 2007, Ciarbelli et
al. 2008)(figura I.5a).
La subfamília HD-Zip I inclou 17 membres i, apart dels propis HD, Zip i de la curta regió
Nt amb aminoàcids de càrrega positiva, no mostren cap més domini conegut. Membres
d'aquesta subfamília s’han implicat en control del desenvolupament per condicions ambientals,
especialment en resposta a estrès abiòtic, com estrès hídric o lumínic, en relació amb les vies
de senyalització d’etilè i àcid abscísic (ABA) (Henriksson et al. 2005, Himmmelbach et al. 2002,
Rueda et al. 2005, Valdes et al. 2012). Diversos membres de la família semblen compartir
activitat, tot i que amb algunes diferències en els patrons d’expressió, suggerint que poden ser
redundants, com a mínim parcialment. La sobre-expressió d’alguns membres produeix defectes
severs en el desenvolupament, amb fulles arrodonides i pecíols més curts (Olsson et al. 2004).
Estudis mitjançant expressió transitòria en Arabidopsis apunten que diversos membres actuen
com a activadors transcripcionals (Henriksson, et al. 2005).

3 Capítol 1
36
(b)
(a)
La subfamília III conté només 5 membres: ATHB8, PHV (PHAVOLUTA/ATHB9), PHB
(PHABULOSA/ATHB14), CNA (CORONA/ATHB15) i REV (REVOLUTA/IFL1). Els membres
d'aquest grup tenen tres dominis addicionals al HD i al Zip, tots situats en posició C-terminal al
HD-ZIP: el domini START (Steroidogenic Acute Regulatory Protein-Related Lipid Transfer), el
domini SAD (START-Adjacent Domain) i finalment la regió MEKLA. El domini START està poc
caracteritzat, especialment en plantes, però es va relacionar inicialment en la regulació,
transport i/o metabolisme de lípids (Soccio and Breslow 2003). Posteriorment, es va veure que
mutacions puntuals en aquest domini canvien l’estabilitat del seu RNA codificant, fet que va
portar a demostrar que alguns residus són dianes d’unió de microRNAs (Bao et al. 2004).
Consistentment, s’ha descrit que tots els membres de la família estan regulats
transcripcionalment i post-transcripcionalment, sent els seus RNAs codificants dianes de
diversos microRNAs (Kim et al. 2005, Mallory et al. 2004, Williams et al. 2005). Adjacent al
domini START hi ha la regió anomenada SAD, del que no es coneix gaire bé la seva funció,
però sembla ser un activador transcripcional. Finalment, més a l’extrem C-terminal (Ct), trobem
una regió amb els aminoàcids conservats MEKHLA, regió que té molta similitud amb els
dominis PAS presents en tots els regnes. Tant la regió MEKHLA com diversos PAS han estat
descrits com a sensors interns de l’estat redox de la cèl·lula, a més de respostes a llum en
plantes (Mukherjee and Burglin 2006). S’ha demostrat que la capacitat d’interacció de proteïnes
que contenen aquests dominis es veu afectada en certes circumstàncies (Chandler et al. 2007);
per exemple, s’ha vist que REV veu la seva capacitat d’interacció inhibida per aquest domini
mitjançant un impediment estèric independent de la pròpia seqüència proteica (Magnani and
Barton 2011). Així, es pot pensar que l’activitat de les proteïnes HD-Zip III pot estar regulada
tant pel domini MEKHLA, sensible a espècies reactives d’oxigen, com pel domini START,
Figura I.5. Classificació de les HD-Zip. (a) Esquema de classificació de les 4 subfamílies. (b) Estructura de la subfamília HD-Zip II.
Adaptació de Ariel et al., 2007.

3 Capítol 1
37
sensible a microRNAs, de manera que l’activitat transcripcional de la família pot venir regulada
per diversos factors interns. Aquesta regulació està justificada donada la importància d’aquesta
família en el desenvolupament de la planta, sent reguladors de la formació de teixit vascular,
del meristem, de la polaritat de la fulla i participant en el transport d’auxines. S’ha vist que
membres d’aquesta família tenen algunes activitats conjuntes, de manera redundant, i altres
més específiques (Prigge et al. 2005). Més concretament, PHV i PHB són els membres que
semblen ser més redundants, mentre que REV sembla ser el més rellevant, donat que el
mutant rev simple és l’únic que presenta un fenotip clar. En conjunt, aquest tres factors regulen
la formació del meristem apical i la iniciació d'òrgans laterals (Otsuga et al. 2001) i controlen el
patró de la dominància apical al llarg del desenvolupament embrionari (Prigge, et al. 2005).
La subfamília IV conté 16 membres, que també posseeixen els dominis START i SAD,
situats a l’extrem Ct de la seqüència proteica. A més tenen un llaç enmig del domini Zip, donant
lloc a un domini conegut com Zip-Loop-Zip (ZLZ). Només quatre dels seus mutants simples
mostren algun fenotip alterat, suggerint que molts d’ells poden tenir funcions redundants i, per
tant, que els processos on intervenen poden ser vitals per la planta (Kamata et al. 2013,
Nakamura et al. 2006). El seu membre fundador i més conegut és GLABRA2 (GL2), implicat en
la formació de l’epidermis i dels tricomes (Lin and Aoyama 2012). L'expressió de quasi tots els
membres d'aquest grup està restringida a les capes cel·lulars externes, en les parts aèries de la
planta. Anàlisis amb plantes de sobre-expressió i de mutants dobles i triples han mostrat
alteracions en el desenvolupament de flors, en la diferenciació cel·lular en l'epidermis, la
formació dels tricomes i l'acumulació d’antocianines (Abe et al. 2003, Kamata, et al. 2013,
Nakamura, et al. 2006).
Finalment, la subfamília HD-Zip II, que engloba 10 gens, conté els 5 membres identificats
al laboratori com a gens PAR. Tots els membres de la subfamília tenen una regió en posició C-
terminal al HD-Zip amb alguns aminoàcids conservats, la seqüència curta CPSCE, que pot ser
responsable de la regulació de l’activitat d’aquestes proteïnes en funció de les condicions redox
de la cèl·lula. S’ha vist, en assajos in vitro fets en condicions oxidants, que aquests factors de
transcripció formen ponts Cys-Cys produint agregats multimèrics, de manera que els
macrocomplexes resultants no poden entrar al nucli ni ser funcionals (Tron et al. 2002). També
tenen una regió en posició N-terminal al HD-Zip més llarg que les altres subfamílies, amb
bastants residus conservats; especialment, es troba un motiu que conté leucines (LxLxL) que
poden estar formant un domini conegut com EAR (ERF–Associated Amphiphilic Repression
Domain, on els ERFs són factors regulats per etilè, Ethylene Response Factor). Aquesta
seqüència curta podria actuar com a repressor de la transcripció (Ciarbelli, et al. 2008, Tiwari et
al. 2004)(figura I.5.b). El fet que els membres d'aquesta subfamília posseeixin un domini
repressor, juntament al fet que uneixen al DNA, és consistent amb les nombroses dades que
apunten aquestes proteïnes com a repressores transcripcionals tant in vitro com in vivo
(Ohgishi et al. 2001, Sawa, et al. 2002, Sorin, et al. 2009, Steindler, et al. 1999, Turchi et al.
2013). Dins d’aquesta subfamília es poden separar per filogènia quatre classes de gens (α–δ).
Tots els de la classe γ (ATHB2, HAT1, HAT2) i δ (ATHB4, HAT3) han estat descrits com a

3 Capítol 1
38
induïts en baixa raó R:FR, i la reversibilitat d’aquesta inducció al tornar a l’alta R:FR ha suggerit
que HAT1, ATHB2, HAT3 i ATHB4 estant regulats a través dels fitocroms (Carabelli et al.
1996). De fet, s’ha demostrat amb experiments de ChIP que ATHB2 i alguns altres d’aquestos
gens estan directament regulat per PIF3, PIF4 i PIF5 (Hornitschek, et al. 2012, Zhang, et al.
2013). Consistentment, els nivells d’expressió d’ATHB2 són superiors en mutants phyB
(Carabelli, et al. 1996), suggerint que els fitocroms són repressors de l’expressió d’aquesta
família mitjançant la inactivació dels PIFs en condicions normals de llum (alta raó R:FR).
Més enllà del paper d’aquests factors HD-Zip II γ i δ en respostes a la llum, recentment
han estat àmpliament implicats en diversos aspectes del desenvolupament de la planta, com en
la formació del carpel (Reymond et al. 2012), la polaritat de les fulles i els cotiledons (Bou-
Torrent et al. 2012, Brandt et al. 2012) o la formació del centre apical i en l’activitat del
meristem apical (SAM, Shoot Apical Meristem), de manera conjunta amb membres de la
subfamília III (Turchi, et al. 2013). S’ha demostrat mitjançant línies mutants que ATHB4 i HAT3
són altament redundants, i que comparteixen part de l’activitat amb ATHB2, produint els dobles
mutants una pèrdua de la identitat adaxial en cotiledons i fulles (Bou-Torrent, et al. 2012,
Turchi, et al. 2013).
En termes generals doncs, la gran família HD-Zip manté un rol essencial en el
desenvolupament, on diversos membres tenen activitats que poden ser redundants, i tenint
quasi tots diversos dominis per tal de controlar-ne l’activitat. No obstant, els únics implicats de
moment en ombra són 5 membres de la subfamília II, i concretament al laboratori ens hem
centrat en el paper d’ATHB4.
3.1.1.2. ATHB4 i la companyia autoreprimida.
Els primers estudis sobre l’activitat biològica d’un membre de la subfamília HD-Zip II es
van fer sobre-expressant ATHB2 en Arabidopsis, obtenint plàntules amb un hipocòtil més llarg
amb cotiledons i fulles més petites (Schena et al. 1993). Plàntules transgèniques sobre-
expressant ATHB2 en antisentit (anti-ATHB2) van mostrar un fenotip oposat, amb pecíols i
hipocòtils més curts (Schena, et al. 1993, Steindler, et al. 1999).
Els fenotips per sobre-expressió d’ATHB2, ATHB4, HAT1, HAT2 o HAT3 són molt
semblants, mostrant plàntules amb hipocòtils i pecíols més llargs, cotiledons més petits i
epinàstics, respostes menors a ombra, i una forta alteració en el desenvolupament de les fulles,
més petites i cargolades cap amunt, en estadi de roseta. Molts d’aquests trets són típics d’un
increment en la senyalització d’auxines, fet que, junt a l’observació que alguns d’aquests
fenotips es veuen abolits quan s’aplica un inhibidor del transport d’auxina com l’NPA (Steindler,
et al. 1999), ha portat a que alguns autors proposin un model pel qual relacionen l’activitat
d’aquestes proteïnes amb la distribució d’AUXs (Morelli and Ruberti 2002). D’altra banda, la
sobre-expressió d’un sol d’aquests factors és suficient per reprimir l’expressió de la resta de
gens pertanyents a la subfamília HD-Zip II, afectant fins i tot els nivells endògens del propi
factor sobre-expressat. S’ha vist que els promotors d’aquests gens són abundants en caixes
consens on aquests mateixos factors s’uneixen, la qual cosa suggereix que aquests cinc

3 Capítol 1
39
membres formen un circuit d’autoregulació negativa, controlant els seus propis nivells
d’expressió, de manera que l’augment de l’expressió d’un dels membres implica la repressió
d’ell mateix i la de la resta de membres de la família (Ciarbelli, et al. 2008, Ohgishi, et al. 2001,
Sawa, et al. 2002, Sorin, et al. 2009, Turchi, et al. 2013). Aquest mecanisme pot ser part d’un
mecanisme de frenar la pujada d’expressió després de la percepció d’ombra, per tal d’evitar
una resposta exagerada i mantenir a la llarga uns nivells adequats d’aquests factors que tenen
un impacte tan important en el desenvolupament de la planta. No obstant, poc es coneix sobre
altres dianes d’aquests factors, i s’ignora si podrien actuar com a activadors en altres promotors
o actuen exclusivament com a repressors.
Al grup s'ha aprofundit en els mecanismes moleculars d’acció dels membres d’aquesta
subfamília HD-Zip II analitzant el paper d’ATHB4 en el control del desenvolupament i de la
SAS. Resultats previs van mostrar que la sobre-expressió constitutiva d’ATHB4 resulta en un
fenotip extremadament nan i estèril, similar a línies amb alts nivells d’expressió d’ATHB2
(Schena, et al. 1993). Donada aquesta dificultat, es van generar línies de sobre-expressió
d’ATHB4 amb activitat induïble, mitjançant el control de la localització subcel·lular de la
proteïna per fusió amb el domini GR (p35S:ATHB4-GR, línia pCS19). El receptor d’origen
animal de glucocorticoides (GR) està retingut al citosol per la seva interacció amb una proteïna
HSP90 en animals (o la seva equivalent de plantes), però en presència d’un glucocorticoide el
GR s’allibera i deixa d’estar retingut al citosol, permetent la seva localització nuclear i donant
lloc a l’activitat de la proteïna fusionada al domini GR. Les plantes no presenten
glucocorticoides, per tant l’activitat d’una proteïna nuclear fusionada al GR vindrà regulada per
l’aportació exògena de glucocorticoide, com per exemple la dexametasona (DEX).
Les plàntules transgèniques induïbles (pCS19) amb ATHB4 actiu (en presència de DEX)
mostren un fenotip nan i més fosc, semblant a la sobre-expressió d’ATHB4 no induïble, i
pràcticament no responen al tractament W+FR. D’altra banda, l’aplicació de dosis creixents
d’epibrassinòlida (EBL, un BR actiu), que indueix l’allargament de l’hipocòtil, va mostrar que les
plàntules transgèniques amb ATHB4 actiu responien més que les plàntules silvestres a la
mateixa dosi d’hormona, concloent que un increment de l’activitat d’ATHB4 confereix
hipersensibilitat als BRs (Sorin, et al. 2009). Experiments semblants amb àcid 2,4-
diclorofenoxiacètic (2,4-D, una auxina sintètica) i GA3 (una GA activa) van mostrar una
elongació de l‘hipocòtil atenuada, de manera que ATHB4 disminueix la sensibilitat a aquestes
hormones. Estudis semblants en tractaments d’ombra van permetre demostrar que ATHB4
altera la sensibilitat a aquestes hormones també en ombra, donant a entendre que és un factor
que interfereix en les respostes a ombra i en les hormonals.
La sobre-expressió d’ATHB4 també afecta l’expressió gènica en gens regulats per ombra
i per hormones. L’expressió de gens ràpidament induïts en ombra com SAUR15, SAUR68 i
HAT2, es veu reprimida per ATHB4, el que reforça el paper d’ATHB4 en la regulació de les
respostes de la SAS (Sorin, et al. 2009). Alguns d’aquests gens també responen a AUXs i BRs,
deixant entendre que ATHB4 podria estar en certa manera regulant l’acció de les hormones per
repressió de les seves dianes d’acció. A més, el fet que gens com els SAUR siguin també

3 Capítol 1
40
regulats per PAR1 porta a hipotetitzar que la senyalització per llum i per hormones conflueixen
a nivell dels gens SAUR, i que PAR1 i ATHB4 són dos dels integradors de la senyal lumínica
amb les hormonals.
Més enllà del rol en ombra s’ha descrit que mutants hat3athb4athb2 presenten cotiledons
completament radialitzats i plantes sense un meristem apical actiu (Turchi, et al. 2013), fenotips
semblants a la pèrdua de funció de les HD-Zip III, especialment de REV, PHB i PHV, que són
reguladors de l’adaxialització. Aquestes dades coincideixen amb el fenotip de fulles cargolades
cap a munt en les plantes sobre-expressores, que és similar a l’expressió ectòpica de membres
de la subfamília III en les cares abaxials (la part inferior o reversa de la fulla). A més, la
interacció entre aquestes dues subfamílies ha estat comprovada per la regulació positiva
directa que té REV sobre ATHB2, HAT2 i HAT3 in vivo. Hi ha evidències que aquestes dues
famílies reconeixen la mateixa o molt similar seqüència consens (AAT[G/C]ATT), caixa
localitzada en els promotors dels membres HD-Zip II, de manera que els nivells d’expressió
vindrien determinats a més de per la llum per canvis en la regulació dels membres de la
subfamília III (Brandt, et al. 2012). En el mateix experiment es va veure que REV controla
l’expressió de gens de síntesi d’auxina com TAA1 i YUCCA5, també unint al seus promotors i
activant-ne l’expressió. Així REV va ser involucrada en respostes a ombra, controlant tant la
síntesi d’auxina com l’expressió de diversos gens de resposta a ombra de la subfamília II de les
HD-Zip, però no s’ha pogut veure si algun mecanisme regula l’activitat d’aquest factor en
resposta a ombra.
Per complicar una mica més l’activitat d’aquests factors, s’ha vist que HAT1, ATHB2,
HAT3 i ATHB4 estan regulats positivament per SPT, membre de les bHLHs que és conegut pel
seu paper en el creixement derivat del carpel. De manera coherent, els dobles mutants
hat3athb4 presenten un desenvolupament incomplet de la regió apical en el carpel, i reduït en
el sèptum, tal com passa amb el mutant spt (Reymond, et al. 2012). Un fenotip semblant ha
estat observat també en mutants hat1, fet que ha portat a demostrar que aquest factor
interacciona genèticament amb CRC (CRABS CLAW, membre de família de factors de
transcripció YABBY que participa en el desenvolupament del meristem i del carpel)(Zuniga-
Mayo et al. 2012). Curiosament, diverses relacions entre SPT i les HD-Zip III amb les auxines
han estat publicades (Nemhauser et al. 2000, Ståldal and Sundberg 2009), tal com ha passat
amb HAT3 i ATHB4 (Sorin, et al. 2009, Turchi, et al. 2013). Recentment s’ha trobat que un gen
regulat per auxines, com BONDELOS (BDL) (un gen Aux/IAA regulat pels ARFs) està regulat
negativament per ATHB5, un membre de la subfamília I (De Smet et al. 2013). La troballa que
un membre HD-Zip reguli un gen Aux/IAA posa en relleu la connexió entre aquesta família i les
auxines. A més, assaigs in vitro porten a pensar que ATHB1, membre de les HD-Zip I, podria
unir la seqüència d’ATHB2, i recíprocament, ATHB2 podria unir la d’ATHB1 (Sessa et al. 1997,
Steindler, et al. 1999), encara que en menor eficiència que la unió a les seves pròpies
seqüències consens. Això porta a pensar que si els nivells d’expressió són prou alts, ATHB2
podria estar directament unint al promotor de BDL. Aquest mecanisme es podria estendre a

3 Capítol 1
41
altres membres de la família HD-Zip IIγ i HD-Zip IIδ, regulant potencialment altres gens
Aux/IAA, contribuint a la seva regulació espai-temporal (Carabelli et al. 2013).
Aquests resultats suggereixen que l’expressió induïda per ombra d’aquests HD-Zip II pot
promoure les respostes SAS modulant els nivells d’Aux/IAA. No obstant, és difícil de dir si
ATHB4 és un regulador positiu o negatiu d’aquestes respostes, ja que tant la sobre-expressió
com el mutant tenen respostes atenuades en termes d’elongació de l’hipocòtil en resposta a
ombra (Sorin, et al. 2009). A més, tenen un rol en el desenvolupament de manera conjunta
amb la subfamília HD-Zip III i podrien estar regulant l’expressió de gens de manera coordinada
amb membres d’aquesta subfamília o membres de la subfamília I. Donada la complexitat del
sistema serà important un avenç en l’elucidació dels mecanismes d’aquestes connexions entre
la regulació de la llum, la hormonal i el desenvolupament.
En la present tesi presentem un estudi de l’activitat dels diversos dominis d’ATHB4 i
sobre els possibles gens diana d’aquest factor.
3.1.2. Resultats.
3.1.2.1 Disseny i obtenció de les construccions derivades d’ATHB4.
Com s’ha comentat anteriorment ATHB4 és un gen PAR que està implicat en el control
de la SAS i la regulació hormonal d’aquesta, però que a més manté un rol en el
desenvolupament de la planta.
Per tal d’estudiar els diferents rols i els mecanismes moleculars d’ATHB4 es va passar a
fer-ne un estudi similar al que s’havia fet prèviament al nostre laboratori per la proteïna PAR1
(Galstyan, et al. 2011). En aquest estudi primer es diferencien els diferents dominis o regions
de la proteïna mitjançant alineaments de seqüència, tant de DNA com de proteïna, amb altres
membres de la seva família. Aquesta informació permet realitzar una sèrie de construccions
clonant els diferents dominis per separat. Posteriorment es fan diversos anàlisis amb les
construccions, entre d’altres es generen línies sobre-expressores d’aquests fragments i es
compara la seva activitat biològica amb el de la proteïna completa. A més, els resultats es
poden complementar amb estudis addicionals, tals com els de localització subcel·lular dels
fragments de proteïna i la capacitat de dimerització o d’interacció amb altres proteïnes.
Idealment, aquest tipus d’estudi permetria establir i avaluar el rol de cada domini o regió en
l’activitat de la proteïna sencera.
Està descrit que la subfamília HD-Zip II presenta una gran homologia de seqüència entre
els seus membres, especialment pels dominis característics homeodomain (HD, responsable
de la capacitat d’unió al DNA) i la cremallera de leucines (ZP, responsable de la capacitat de
dimeritzar) (Ariel, et al. 2007). Gràcies a aquestes similituds vam poder establir amb relativa
facilitat els límits d’aquests dominis funcionals de la proteïna ATHB4 per comparació amb el
que estava prèviament publicat sobre l’estructura de la proteïna ATHB2 (Ciarbelli, et al. 2008,
Ohgishi, et al. 2001) (figura C1.1). D’acord amb aquesta comparació, vam definir el domini HD,
entre els aminoàcids 143 i el 219 de la seqüència d’ATHB4, seguit pel domini ZP, entre els

3 Capítol 1
42
residus 220 i 263. Es va anomenar a la resta com a regions N-terminal (Nt, del residu 1 al 142) i
C-terminal (Ct, de l’aminoàcid 264 al 318). En aquestes dues regions hi cal destacar una
seqüència curta rellevant en cada una. En l’Nt hi trobem unes leucines inicials que formen un
motiu EAR, classificat com a domini repressor (Ciarbelli, et al. 2008, Kagale et al. 2010, Tiwari,
et al. 2004) . Està descrit que diversos membres de la família actuen com a repressors de la
transcripció (Ohgishi, et al. 2001, Steindler, et al. 1999) i que aquest motiu és potencialment el
responsable d’aquesta activitat per interacció amb proteïnes remodeladores de la cromatina
(Kagale and Rozwadowski 2011). En la regió Ct hi trobem la regió CPSCE, molt conservada
dins la subfamília II i que podria estar relacionada amb algun tipus de regulació a l’estat redox
de la planta (Ariel, et al. 2007), tot i que no hi ha cap estudi publicat al respecte.
Nt
HD
ZP
Ct
Un cop delimitats els dominis d’ATHB4, vam triar quines combinacions de regions
usaríem per fer anàlisis de l’activitat biològica. Com que la sobre-expressió constitutiva
d’ATHB4 genera un fenotip molt fort, i generalment poc fèrtil, vam decidir mantenir el sistema
induïble de la línia control pCS19, línia transgènica que s’ha descrit a la introducció i que sobre-
expressa ATHB4 sencera fusionada al domini GR (35S:ATHB4-GR), i per tant amb activitat
induïble per l’aplicació del glucocorticoide sintètic dexametasona (DEX). Tenint en compte el
paper dels dominis HD i ZP, sabent que ATHB4 pot actuar com a repressor transcripcional i
que, tal com estava descrit per ATHB2, la dimerització a través del ZP podria ser essencial
perquè el domini HD es trobés en la conformació adequada per unir al DNA, es van dissenyar
les construccions que apareixen a la figura C1.2. Es van dissenyar una sèrie d’encebadors
(MGO1-4, es disposava dels oligos pels extrems de la seqüència JO284 i CSO7) que contenien
el codó d’iniciació o eliminaven els codons de parada i/o introduïen un lloc de restricció BamHI
per facilitar el clonatge posterior en un vector binari. Els fragments desitjats es van amplificar
Figura C1.1. Alineament de seqüències de les proteïnes ATHB2 i ATHB4. Els requadres en vermell remarquen els motius
rellevants, l’EAR en l’Nt i el CPSCE del Ct. A la dreta es marquen els noms donats als diferents dominis o regions: Nt = regió N-
terminal, HD = domini homeodomain, ZP = domini leucine zipper, Ct = regió C-terminal.

3 Capítol 1
43
per PCR a partir d’un plàsmid disponible al laboratori que contenia la ORF d’ATHB4 sencera
(vector pCS19) i es van clonar en un vector comercial. Les construccions es van seqüenciar i
es van analitzar tant per PCR com per digestió per tal d’assegurar que tenien els tamanys i
característiques desitjats. Amb aquestes construccions es va passar a fer l’estudi estructura-
funció.
En una tesi anterior (Salla Martret 2012) es va estudiar la capacitat de dimerització
d’alguns d’aquests fragments derivats d’ATHB4 per assajos de doble híbrid en llevat (Y2H,
Yeast two hybrid), mostrant que ATHB4 homodimeritzava sense necessitat dels dominis Nt o
Ct. En el mateix treball es va estudiar la localització subcel·lular per tècnica de
microbombardeig en epidermis de ceba, fusionant les diferents truncacions dissenyades (figura
C1.2) a la proteïna fluorescent verda (GFP, de l’anglès Green Flourescent Protein). Es va
observar que totes les construccions amb el domini HD localitzaven específicament al nucli,
mentre que les construccions equivalents a la pMG25 i pMG26 es localitzaven tant al citosol
com al nucli, tal com passava amb el control que expressava la GFP sola (Salla Martret 2012).
Nt HD Zp GR pMG2335S
HD Zp GR pMG2435S
Zp GR pMG2535S
HD GR pMG2835S Nt
pMG29HD GR35S
GR pMG2635S Zp Ct
HD GR pMG2735S Zp Ct
Nt HD Zp GRCt pCS1935S
JO284
CSO7
MGO1 MGO2
MGO3MGO4
Per cada construcció es va digerir el vector de clonatge per les dianes BamHI (que
alliberava el fragment d’interès sencer) i es va clonar en un vector binari pCAMBIA1300
modificat (vector pMG15) que contenia el domini GR en marc de lectura amb els fragments a
clonar mitjançant el tall amb BamHI. Es va transformar Agrobacterium tumefaciens amb el
vector binari resultant i es va transformar Arabidopsis mitjançant infecció per immersió floral.
Vàries plantes transgèniques de cada construcció (generació T1) van ser seleccionades en
plaques amb higromicina, comprovant per PCR si la inserció era l’esperada i més tard
(generació T2) seleccionant per les línies amb una sola inserció.
Es van testar els nivells d’expressió de les diferents construccions per anàlisis de
Northern-blot en la generació T2, creixent durant 7 dies en llum blanca unes 300 llavors (uns 25
µL) per mostra i usant com a controls la mateixa quantitat de material vegetal per Col-0 (control
negatiu) i plàntules de la línia estable pCS19 (control positiu i referència). A partir d’aquests
anàlisis (figuraC1.3), es van seleccionar les línies que presentaven millors nivells d’expressió
per cada construcció. Així es van obtenir plantes que sobre-expressen els diferents fragments
d’ATHB4 fusionada al domini GR. Es van provar anàlisis per Western-blot, per tal de veure si
els nivells d’expressió (mRNA) es corresponien amb bons nivells de proteïna, però es va veure
Figura C1.2. Construccions derivades
d’ATHB4 (I). Esquema de les construccions
codificants per truncacions de la proteïna ATHB4 a
partir de les regions descrites a la figura RI.1. A
l’esquerre s’indica el promotor que dirigeix
l’expressió de la construcció en les plantes
transgèniques i a la dreta apareix el nom donat a
la construcció. En la primera construcció s’indica la
posició dels oligonucleòtids usats per fer els
clonatges per la resta de construccions. GR =
Receptor de glucocorticoides d'origen animal.

3 Capítol 1
44
que amb els dos anticossos comercials anti-GR usats només vam ser capaços d’identificar
bandes inespecífiques en extractes proteics totals vegetals (dades no mostrades).
3.1.2.2. Activitat biològica de les truncacions: Respostes fisiològiques.
Es va començar l’anàlisi d’activitat biològica de la proteïna mirant primer el fenotip de les
plàntules creixent en presència de DEX durant 7 dies en llum W. Com hem comentat, en
presència de DEX les plàntules de la línia que sobre-expressa la proteïna sencera (pCS19)
mostren un fenotip nan, cotiledons hiponàstics i un color més fosc característics. L’única
construcció que va mostrar un fenotip idèntic a la línia pCS19 en diverses línies va ser la
construcció pMG23, tal com es pot veure en la figura C1.4a. Diverses línies de la construcció
pMG28 també responien al tractament de DEX, presentant un fenotip molt similar a les pCS19,
però no tant fort (dades no mostrades). Respecte a les altres construccions no semblava haver-
hi cap línia amb un fenotip clarament diferent al de Col-0, si bé alguna línia puntual presentava
hipocòtils i pecíols més llargs que les plantes Col-0 (cas de la pMG27.3).
GR
29.1
29.2
29.3
28.5
28.7
25.5
25S
Co
l-0
pC
S19
23.1
23.2
24.1
24.5
24.8
24.1
2
25.1
25.2
25.3
26.1
26.2
26.3
27.3
27.4
27.5
28.1
28.2
28.4
pMG29pMG25pMG23 pMG24 pMG26 pMG27 pMG28
Wd0 d7
A continuació vam analitzar la resposta dels hipocòtils a ombra simulada (W+FR) de
línies homozigotes de cada construcció. En presència de DEX, plàntules de la línia pCS19
presentaven una disminució de la resposta dels hipocòtils a ombra simulada (Sorin, et al.
2009). En una primera aproximació, es van fer experiments creixent les plàntules tant en un
llum W com en W+FR, en presència i absència de DEX des de dia 0, tal com s’esquematitza a
la figura C1.4b. Com a línies control es van utilitzar plantes wt sense transformar (Col-0) i la
línia pCS19. Pels assajos es van sembrar dues plaques per cada tractament i, a partir del dia 2
de creixement en llum W, una de les plaques es va tractar amb llum W+FR. A dia 7 es va
mesurar la longitud de l’hipocòtil de les plàntules. Tal com es mostra a la figura C1.4c, quan
creixien en llum blanca la longitud dels hipocòtils de totes les línies analitzades estava poc
afectada pel tractament amb DEX. Sota W+FR, el control Col-0 allargava l’hipocòtil en resposta
a ombra simulada independentment del tractament en DEX. En canvi i tal com estava publicat,
les plàntules pCS19 mostraven una clara atenuació de l’elongació de l’hipocòtil de manera
dependent de l’aplicació de DEX, indicatiu de l’activitat biològica de la proteïna ATHB4 sencera
(Sorin, et al. 2009). Plàntules de les dues línies pMG23 responien de manera semblant a les de
Figura C1.3. Anàlisis d’expressió de les línies transgèniques sobre-expressant truncacions d’ATHB4. Autoradiografies
mostrant els nivells d’expressió per Northern-blot de diferents línies en la generació T2, en 3 membranes independents, creixent les
plàntules fins a dia 7 sense cap tractament. Es mostra una de les dues rèpliques biològiques, per simplificar s’ha obviat mostrar els
controls en la segona i tercera membrana. La part superior mostra l’autoradiografia de la membrana hibridada amb una sonda pel
domini GR, mentre la inferior mostra el control de càrrega hibridant amb una sonda constitutiva 25S.

3 Capítol 1
45
la línia pCS19, el que indicava que la proteïna truncada sobre-expressada en les plantes
pMG23 mostra una activitat biològica semblant al de la proteïna sencera.
A la vista dels resultats obtinguts en llum W, i per simplificar els experiments de la resta
de construccions, es va decidir créixer les plàntules de les diferents línies només en W+FR,
comparant l’elongació amb i sense DEX al medi (figura C1.5). Les anàlisis van indicar que
només les línies pMG28 tenien una activitat semblant a les pCS19, amb hipocòtils molt més
curts en DEX. Aquest resultats indiquen que el domini Nt és clau en la funció biològica
d’ATHB4 en respostes a ombra, ja que les línies que no disposen del domini Nt no semblen
tenir respostes diferents al de les plantes sense transformar (figures RI.4 i 5), mentre que les
construccions pMG23i pMG28 mostren el mateix fenotip que la línia sobre-expressora d’ATHB4
completa. Amb la mateixa comparació es pot concloure que el domini Ct no té cap activitat en
resposta a ombra, ja que la construcció pMG23 té activitat comparable a la de la línia pCS19
però li manca aquest domini. Les construccions amb els dominis HD-Zip, com la pMG24 i la
pMG27, no mostren activitat a ombra alterada, o mostren una alteració molt feble. Cal dir que
algunes construccions, com la pMG27 o la pMG26, van mostrar una lleugera disminució en la
resposta en alguna línia (com les representades en la figura), però que altres línies de la
mateixa construcció no mostraven cap activitat. Al no poder analitzar els nivells de proteïna de
les línies hem de suposar que les línies que mostren algun tipus d’alteració en resposta a DEX
és perquè expressen prou proteïna per donar lloc a aquesta, mentre que les línies que no
mostren activitat, tot i tenir nivells alts d’expressió, no podem saber si és perquè la proteïna és
poc o gens funcional o si els nivells d’expressió no es tradueixen en alts nivells de proteïna. Per
poder aclarir si algunes d’aquestes línies tenien o no activitat es va passar a mirar respostes
moleculars, possiblement més sensibles que les fisiològiques.
Lo
ng
itu
d h
ipo
còti
l(m
m)
0
1
2
3
4
5
6
7
8
Col-0 pCS19 pMG23.1.4 pMG23.2.5
0000
00
W
W+FR
d0 d2 d7
(b)(a)
(c)
W W+FR
- DEX +DEX -DEX +DEX
Col-0
pCS19
pMG23.1
pMG23.2
Col-0 pCS19 pMG23.1.4 pMG23.2.5
W+FRW
Nt HD ZppMG23
- DEX
+ DEX
3.1.2.3. Activitat biològica de les truncacions: Respostes moleculars.
Figura C1.4. Fenotip de plàntules de la construcció pMG23 en W i en tractaments d’ombra simulada. (a) Fenotips de les
plàntules a dia 7 amb i sense tractament en DEX i amb i sense tractament d’ombra per dues línies independents de la construcció
pMG23. (b) Esquema indicant les condicions de creixement: les plantes van ser crescudes des de dia 0 amb o sense DEX, es van
deixar créixer 2 dies en W i un grup es va transferir a W+FR a partir del segon dia i fins a dia 7. (c) Mesures de longitud de l’hipocòtil
en dues línies independents, homozigotes, de la construcció pMG23 i els controls wt (Col-0) i sobre-expressió d’ATHB4 sencera
(pCS19). Els símbols indiquen diferències significatives (0 P<0.05; 00 P<0.01) respecte el mateix genotip creixent en iguals condicions
de llum sense DEX (-DEX) .

3 Capítol 1
46
Tal com s’ha comentat a la introducció, ATHB4 afecta l’expressió de gens regulats per
ombra, com SAUR15 i HAT2. Treballs previs havien mostrat que plantes sobre-expressores
d’ATHB4 reprimien l’expressió de SAUR15 en tractaments amb DEX i CHX (Salla Martret 2012)
i resultats no publicats), fet que porta a pensar que aquest gen seria una diana directa
d’ATHB4. De manera semblant al que s’havia fet per SAUR15 es van fer anàlisis de microarray
comparant canvis d’expressió en mostres de la línia pCS19 respecte al seu control Col-0, en
presència i absència de DEX i en presència i absència de CHX. D’aquesta manera els gens
diferentment expressats en presència de l’inductor d’activitat i de l’inhibidor de síntesi de
proteïnes eren potencialment dianes directes de l’acció d’ATHB4, ja que tenien la seva
expressió alterada quan ATHB4 es trobava al nucli de manera independent de la síntesi de
proteïnes. Les mostres d’RNA van ser obtingudes i processades en una tesi anterior, i els
resultats crus van ser analitzats per altres membres del laboratori. Com es veu a la figura
C1.13a, el solapament entre els gens amb expressió alterada per DEX sense CHX (251 gens) i
amb CHX (532) dóna lloc a una llista de 53 gens potencialment dianes directes d’ATHB4, on
majoritàriament (52 d’aquests) presenten una repressió per l’activitat de la proteïna, suggerint
clarament que ATHB4 estaria reprimint l’expressió d’aquests gens de manera directa. Un
d’aquests era HAT2, que també està descrit com a gen de resposta a ombra. Així doncs, tant
SAUR15 com HAT2 són gens que veuen els seus nivells d’expressió incrementats en resposta
a ombra, però en canvi estan clarament reprimits per acció d’ATHB4 (figura C1.6a).
Col-0 pCS19 pMG25.5.110,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
00
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
Col-0 pCS19 pMG24.12.6
00
Lo
ng
itu
d h
ipo
còti
l(m
m)
Col-0 pCS19 pMG28.4.20,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
5,0
Col-0 pCS19 pMG29.10,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
5,0
Lo
ng
itu
d h
ipo
còti
l(m
m)
00 00
00
Col-0 pCS19 pMG26.7.40,0
1,0
2,0
3,0
4,0
6,0
5,0
7,0
00
0
Col-0 pCS19 pMG27.12.70,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
00
0
(b)
- DEX
+ DEX
W+FR
d0 d2 d7
HDpMG24 Zp ZppMG25 CtZppMG26
HD Zp CtpMG27 HDNt pMG28 pMG29
(a)
HD
Figura C1.5. Anàlisis d’activitat biològica per les construccions derivades d’ATHB4 en tractaments d’ombra simulada. (a)
Esquema indicant les condicions de creixement: les plantes van ser crescudes des de dia 0 amb o sense DEX, es van deixar créixer 2
dies en W i es va transferir a W+FR a partir del segon dia i fins a dia 7. (b) Mesures de longitud de l’hipocòtil per les diferents
construccions, es mostra un experiment i una línia representativa. Condicions experimentals com les anteriors, però només en
tractament W+FR. Els símbols indiquen diferències significatives (0 P<0.05; 00 P<0.01) respecte el mateix genotip creixent en iguals
condicions de llum sense DEX (-DEX).

3 Capítol 1
47
Vam raonar que molt probablement l’activitat de les formes truncades sobre l’allargament
de l’hipocòtil i sobre l’expressió gènica estaven relacionats. Com a assaigs complementaris
d’activitat biològica vam estudiar si l’efecte molecular de la sobre-expressió de les truncacions
era el mateix o similar al de la forma sencera, o si la deleció d’alguna regió donava lloc a
activitats moleculars alterades, tals com canvis en l’expressió de gens induïts per ombra, com
SAUR15 i HAT2. Per això es va passar a analitzar l’activitat biològica per RT-qPCR en temps
curts de resposta a ombra. Inicialment es va fer un assaig amb una línia pMG23 i els controls
Col-0 i pCS19. Per això es van sembrar 4 plaques de cada genotip, i es van créixer fins a dia 7
per agafar mostres en totes les condicions, tant en W (0h de tractament), com en W+FR (1 hora
de tractament) i amb i sense DEX des de dia 0 (figura C1.6b).
0h
+1h
SAUR15 +DEX
0,0
0,5
1,0
2,0
2,5
3,0
1,5
00
00
Col-0 pCS19 pMG23.1
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5HAT2 +DEX
Col-0 pCS19 pMG23.1
0
1
2
3
4
5
6
7SAUR15 -DEX
Col-0 pCS19 pMG23.1
Exp
ress
ióre
lati
vaE
xpre
ssió
rela
tiva
00
00
HAT2 -DEX
0
2
4
6
8
10
12
Col-0 pCS19 pMG23.1
0
(a)
HAT2SAUR15
W+FR
ATHB4
Wd0 d7
1h W+FR
GR -DEX
pCS19 pMG23.10,0
0,5
1,0
1,5
Exp
ress
ióre
lati
va
0000
(d)
0h
Nt HDpMG23
(b)
(c)
- DEX
+ DEX
Zp
Com es veu a la figura C1.6c, la línia pCS19 mostra nivells d’expressió similars al Col-0
en les mostres sense DEX, incrementant l’expressió dels gens marcadors en tractament
d’ombra. Quan aquest genotip és crescut en tractament de DEX es pot veure que els nivells
d’expressió dels gens marcadors són més baixos en llum W, però sobretot es veu que la
resposta al tractament W+FR està completament anul·lada. El mateix resultat es pot veure per
la línia pMG23 analitzada, mostrant nivells d’expressió dels gens marcadors clarament inferiors
al control wt en les mostres tractades amb DEX, especialment amb una resposta nul·la al
tractament d’ombra. Es va veure que les condicions en W eren poc informatives, ja que els
nivells bassals dels gens analitzats són prou baixos i la repressió de la seva expressió pel
tractament amb DEX és molt més clara quan aquests gens han estat induïts pel tractament
Figura C1.6. Efecte molecular de la sobre-expressió de la construcció pMG23 sobre marcadors moleculars d’activitat
d’ATHB4.
(a) HAT2 i SAUR15 són gens induïts en ombra i reprimits per l’acció d’ATHB4. (b) Esquema indicant les condicions de creixement: les
plantes van ser crescudes des de dia 0 amb o sense DEX, es van deixar créixer fins a dia 7 i es van agafar mostres abans (0h) i
després de tractament en W+FR (1h). (c) Anàlisis comparatiu de l’expressió de HAT2 i SAUR15 en plàntules de Col-0, pCS19 i pMG23
en resposta a llum W+FR, a l’esquerre el tractament sense DEX (fons blanc) i a la dreta amb DEX al medi des de dia 0 (fons blau). (d)
Nivells d’expressió del control pCS19 i de la línia analitzada per la construcció pMG23. Els símbols indiquen diferències significatives (0
P<0.05; 00 P<0.01) respecte el control wt creixent en iguals condicions.

3 Capítol 1
48
amb 1 h d’ombra simulada. Per això es va decidir comparar l’efecte de la DEX a la resta de
línies només amb tractament W+FR (1h) (figura C1.7a). A més, vam pensar que si algunes
construccions amb diverses regions demostraven no tenir activitat molecular altres
construccions més curtes, que no semblaven tenir activitat fisiològica, tampoc en tindrien a
nivell molecular i, per tant, no era necessari analitzar la resposta molecular de totes les
construccions. Així doncs, vam decidir que construccions curtes, codificants per una sola regió
(com la pMG25 i la pMG29), no serien analitzades molecularment a menys que construccions
més llargues (com la pMG24 o la pMG27) donessin lloc a alguna activitat molecular.
- DEX
+ DEX
Exp
ress
ióre
lati
va
(c)
(a) Wd0 d7
+1h W+FR
00
HAT2
Col-0 pCS19 pMG24.12
Exp
ress
ióre
lati
va
0,0
0,2
0,4
0,6
0,8
1,0
1,2SAUR15
Col-0 pCS19 pMG24.120,0
0,5
1,5
2,0
2,5
1,0
00
HAT2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Col-0 pCS19 pMG26.7
SAUR15
Col-0 pCS19 pMG26.70,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
0000
0
Col-0 pCS19 pMG27.12
SAUR15
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4HAT2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Col-0 pCS19 pMG27.12
0000
00
Col-0 pCS19 pMG28.4
SAUR15
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
0,0
0,2
0,4
0,6
0,8
1,0
1,2
00
Col-0 pCS19 pMG28.4
HAT2
0000
pCS19 pMG24.120,0
0,2
0,4
0,6
1,0
0,8
1,2
1,4
pCS19 pMG27.120,0
0,5
1,0
1,5
2,0
2,5GR
pCS19 pMG28.40,0
0,2
0,4
0,6
1,0
0,8
1,2
1,4
00 00
0000
0000
pCS19 pMG26.70,00,20,40,60,81,01,21,41,61,8
GR 0000 GRGR
HDpMG24 CtZppMG26
HD Zp CtpMG27 HDNt pMG28
- DEX
+ DEX
(b)
HAT2
Zp
Els resultats moleculars (figura C1.7b) van mostrar que les línies pMG24 i pMG27 tenien
les respostes moleculars a ombra inalterades, mentre que la línia pMG28 mostrava una clara
atenuació de la resposta quan creixia en DEX, de manera molt similar a la línia pCS19. La
construcció pMG26 no tenia cap alteració en l’expressió de HAT2, però va mostrar una lleugera
disminució de l’expressió de SAUR15. No obstant, aquesta petita variació no és suficient per
confirmar que la línia tingui cap activitat molecular. Així doncs, els resultats moleculars
confirmaren els fisiològics, posant de manifest la importància del domini Nt i la no necessitat del
Figura C1.7. Efecte de la sobre-expressió de proteïnes truncades d’ATHB4 sobre l’expressió de HAT2 i SAUR15. (a) Esquema
indicant les condicions de creixement: les plantes van ser crescudes des de dia 0 amb o sense DEX, es van deixar créixer fins a dia 7
i es van agafar mostres després de tractament en W+FR (1h). (b) Anàlisis per qPCR dels nivells d’expressió dels gens marcadors
SAUR15 i HAT2 amb una línia representativa de cada truncació. (c) Anàlisis dels nivells d’expressió de transgen amb les mateixes
mostres. Els símbols indiquen diferències significatives (0 P<0.05; 00 P<0.01) respecte el control wt creixent en iguals condicions.

3 Capítol 1
49
Ct en les respostes a ombra. Sorprenentment, malgrat estar descrit que les proteïnes de la
família HD-Zip són factors de transcripció que dimeritzen mitjançant el domini ZP i que han de
formar dímers per poder unir al DNA a través del HD, construccions que no contenen el domini
ZP, i per tant no poden dimeritzar, mostren activitat biològica (pMG28), tant en respostes
fisiològiques (més a llarg temps) com moleculars (a temps curts després de la percepció
d’ombra).
Per establir si totes les línies tenien nivells d’expressió suficients per tenir activitat
biològica es van comparar els nivells d’expressió del transgen (amb encebadors per l’etiqueta
GR) de cada línia analitzada (figura C1.6c i 7c) amb els de la línia control pCS19, veient-se que
línies amb una activitat semblant a pCS19 tenien nivells d’expressió més baixos (pMG23 i
pMG28) i altres que no mostraven activitat biològica tenien nivells d’expressió més alts que la
línia control (pMG26 i pMG27). Per tant, no eren els nivells d’expressió el que afectava a
l’activitat biològica de la proteïna.
3.1.2.4. Nous horitzons: Aprofundint en l’Nt.
A partir d’aquests resultats es va decidir estudiar més en profunditat la regió Nt. Per això
es va decidir fer noves construccions (figura C1.8). Per comprovar si la regió Nt és suficient per
mantenir, encara que sigui parcialment, l’activitat biològica la proteïna ATHB4 es va dissenyar
la construcció pMG60, sobre-expressant només aquesta regió Nt. El clonatge es va fer com
s’ha descrit per les altres construccions, usant com a motlle del vector codificant per la proteïna
ATHB4 sencera (encebadors d’amplificació 2x35S i MGO51). D’altra banda es va fer una
construcció amb una deleció parcial de l’extrem 5’ de l’Nt (pMG44) en la que faltava el motiu
EAR (encebadors d’amplificació MGO50 i CSO7). A més es van fer algunes construccions amb
mutacions puntuals en residus conservats. Les mutacions puntuals van ser introduïdes per
PCR amb encebadors que contenien els canvis de base necessaris, usant un protocol de
mutagènesis dirigida amb dues combinacions d’oligos: una amb l’encebador directe mutat i el
revers normal (el CSO7); i una segona combinació amb l’encebador revers mutat i el directe
normal (el JO284). Els productes resultants eren purificats i usats com a motlle per una segona
amplificació amb els encebadors per la seqüència sencera (JO284 i CSO7). D’aquesta manera
la segona amplificació donava lloc a la seqüència sencera usant com a motlle els productes de
la primera amplificació, productes que contenien les mutacions puntuals. Tal com s’ha explicat
anteriorment el fragment resultant va ser insertat en un vector de clonatge i es va comprovar
per PCR, digestió i finalment es va seqüenciar. Els fragments es van digerir amb BamHI, com
les primeres construccions, i es van passar al mateix vector binari pMG15 (derivat del
pCAMBIA1300) que conté en marc de lectura l’etiqueta GR.
Les mutacions puntuals generades van ser: (1) mutagènesi de la primera leucina del
motiu EAR de l’Nt (combinacions d’encebadors amb la mutació MSO36+CSO7 i
JO284+MSO37) per una alanina (Leu10-Ala), descrita com a residu clau per mantenir l’activitat
repressora d’aquest motiu (Tiwari, et al. 2004). En plantes transformades amb aquesta
construcció (pMG43) podíem preveure una caiguda de l’activitat repressora d’ATHB4 degut a la

3 Capítol 1
50
baixa interacció de proteïnes repressores amb el motiu EAR mutat; (2) mutagènesi del triptòfan
80 de l’Nt per una alanina (Trp80-Ala, construcció pMG38, encebadors amb mutacions
MGO22+CSO7 i JO284+MGO23). Aquest residu de la regió Nt es va veure que estava molt
conservat entre diversos membres de la subfamília HD-Zip II, i es coneix que el triptòfan
acostuma a ser un residu rellevant pel correcte plegament de la proteïna. Amb aquest canvi es
va pensar que es podria alterar l’activitat del domini Nt, possiblement per un mal plegament
d’aquest; i (3) una mutació puntual en el HD (pMG37, encebadors usats MGO20+CSO7 i
JO284+MGO21). Aquest canvi d’una Asn a Ala està descrit que incapacita ATHB2 per unir al
DNA, ja que aquest és un residu clau en la interacció de la proteïna amb el DNA (Sessa, et al.
1997). D’aquesta manera es podia comprovar si una modificació en la capacitat d’unir al DNA
d’ATHB4 afecta a la seva activitat en les respostes a ombra.
N HD Zp GRCt pMG4435S
Nt GR pMG6035S
Nt HD Zp GRCt pMG4335S
Nt HD Zp GRCt pMG3835S
Nt HD Zp GRCt pMG3735S
Nt HD Zp GFPCt pBJ335S
HD GFP MG6235S Nt
HD GFP MG5835S Zp Ct
A una tesi anterior (Salla Martret 2012) s’havia analitzat la localització subcel·lular de
totes les proteïnes truncades descrites a la figura C1.2. Es va observar que aquelles
construccions que contenien el domini HD es localitzaven clarament al nucli, el que suggeria
que aquest domini contenia una seqüència de localització nuclear. No disposàvem d’informació
de la regió Nt sola, que no sembla contenir cap senyal de localització subcel·lular coneguda.
Per això es va fer un experiment de microbombardeig en epiteli de ceba, equivalent als descrits
en el treball anterior, amb la regió clonada de l’Nt. El fragment Nt va ser subclonat en un vector
binari derivat de pCAMBIA1302 que permetia clonar, usant digestió amb BamHI el fragment Nt
(pMG61). Aquesta construcció permet expressar sota el control del promotor 35S el fragment
Nt fusionat amb la GFP (35S:Nt-GFP). Per tal de visualitzar amb més facilitat les cèl·lules
transformades durant el bombardeig es van co-transformar amb el vector pDsRed, que codifica
per la proteïna la proteïna vermella fluorescent (RFP). Com es mostra a la figuraC1.9, la
proteïna de fusió Nt-GFP co-localitza amb la RFP, mostrant que pot entrar al nucli i que es
localitza majoritàriament al citosol, de manera similar al que passa amb la construcció que
codifica per la proteïna GFP sola (pMS51). Aquesta localització porta a pensar que la sobre-
expressió en plantes d’aquesta regió és possible que no mostri cap activitat, posant de manifest
que potser caldria afegir-hi una senyal de localització nuclear (NSL). No obstant encara s’estan
seleccionant plantes pMG60 i no s’ha tingut temps de preparar la construcció amb la NSL
fusionada a la regió Nt.
Pel que fa a les construccions pMG37, pMG38, pMG43 i pMG44 es van transformar en
planta i es van analitzar els nivells d’expressió del transgen en la generació T2 (per northern-
Figura C1.8. Construccions derivades
d’ATHB4 (II). Esquema de les construccions
per estudiar l’activitat de la regió Nt i la
relació entre unió al DNA i activitat de la
proteïna. A l’esquerre s’indica el promotor
que dirigeix l’expressió de la construcció en
les plantes transgèniques i a la dreta apareix
el nom donat a la construcció. GFP =
Proteïna fluorescent veda. = Mutació
puntual, canvi de residu a Ala.

3 Capítol 1
51
blot o per qPCR), seleccionant les línies amb nivells d’expressió més alts pels posteriors
anàlisis (resultats no mostrats). A la generació T3, quan s’havien identificat línies homozigotes
pel transgen, es van fer assajos d’activitat biològica com els descrits anteriorment (veure figura
C1.5 i C1.10a). Com es pot veure a la figura C1.10b, només la línia pMG37 manté una activitat
biològica semblant a la línia control pCS19, amb una atenuació clara de la resposta a ombra en
presència de DEX. Les línies que sobre-expressen ATHB4 amb el domini Nt modificat (pMG38 i
pMG43) o truncat (pMG44) presenten nivells molt menors d’activitat biològica. No obstant hi ha
algunes línies que mostren una lleugera activitat, tant en la construcció pMG38 (1 línia sobre 4
analitzades, de les quals es mostren dues línies, la que té activitat i una de les que no), com
per la pMG43 (2 línies amb activitat feble sobre 4 analitzades).
pMS51 (~pC1302)
GFP RFP
W Merge
pMG61
GFP RFP
WMerge
GFP35S GFP35S Nt
RFP35SpDsRed RFP35SpDsRed
Veient aquesta variabilitat es va considerar interessant analitzar també l’activitat
molecular d’algunes d’aquestes línies, de la mateixa manera anteriorment descrita (figura
C1.7). En especial era interessant analitzar l’activitat molecular de la construcció pMG43, ja que
esperàvem que aquesta proteïna hauria de ser defectuosa en unir a proteïnes repressores i per
tant podria no mostrar activitat molecular. Els resultats amb la construcció pMG43 (figura
C1.11) mostren una repressió clara dels gens marcadors en les mostres crescudes en DEX,
confirmant l’activitat biològica del transgen i mostrant una activitat molecular fins i tot més forta
que la fisiològica. Com s’havia fet anteriorment es van analitzar també els nivells d’expressió
del transgen, mostrant nivells semblants d’expressió entre la construcció pMG43 i la línia
pCS19. Tot i que els nivells d’expressió eren similars, l’activitat de la línia pCS19 seguia sent
més forta (menor expressió dels gens), fet que ens porta a pensar que la mutació puntual en el
motiu EAR desestabilitza la interacció entre ATHB4 i les proteïnes repressores que s’uneixen al
motiu.
Tots els resultats presentats amb les diferents línies es resumeixen a la taula C1.1,
mentre que informació addicional sobre el número de línies obtingudes, analitzades i mostrades
es troba a la taula C1.2. El conjunt de resultats ens permeten dir que la regió Nt és molt
Figura C1.9. Localització subcel·lular de la regió Nt d’ATHB4. Imatges representatives de la localització subcel·lular per
bombardeig en ceba vista en el confocal. GFP = senyal de la Green Fluorecent Protein; RFP = senyal de la Red Fluorecent Protein;
W = Camp clar; Merge = Superposició de les dues fluorescències. La regleta indica 50 µm.

3 Capítol 1
52
important per l’activitat biològica de la proteïna, tot i que en algunes construccions es veu un
certa variabilitat, mostrant algunes línies activitats febles. El fet que les construccions pMG28 i
pMG37 mostrin una activitat molt similar a la línia pCS19 porta a pensar que ATHB4 està
actuant en respostes a la SAS de manera independent a la unió al DNA, ja que no sembla
necessitar el domini ZP i no l’afecten mutacions puntuals en residus descrits com a necessaris
per la unió del domini HD al DNA. Així doncs, mentre que la regió Nt és essencial per les
respostes a ombra, les regions HD o ZP semblen no ser tant rellevants, posant de manifest que
ATHB4 actua en ombra mitjançant mecanismes moleculars desconeguts.
Col-0 pCS19 pMG37.9.80,0
1,0
2,0
3,0
4,0
6,0
5,0
Lo
ng
itu
d h
ipo
còti
l(m
m)
00
00
Col-0 pCS19 pMG43.9.70,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
00
00
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
Col-0 pCS19 pMG38.14.6 pMG38.18.7
00
00
- DEX
+ DEX
W+FR
d0 d2 d7
Nt HD Zp Ct Nt HD Zp Ct Nt HD Zp Ct N HD Zp Ct
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
Col-0 pCS19
00
pMG42.2.10
00
(a)
(b)
D’altra banda, tots els resultats anteriors han fet referència a l’activitat d’ATHB4 en
respostes a ombra, però se sap que aquest factor de transcripció també té un paper en el
desenvolupament, alterant la morfologia de les fulles i cotiledons. Al treballar amb línies
induïbles, no hem vist cap fenotip en plantes adultes, ja que per l’obtenció de llavors les plantes
sempre han crescut en testos sense cap tractament. Per estudiar el paper d’algunes regions de
la proteïna en la polaritat de les fulles s’estan realitzant línies de sobre-expressió d’algunes de
les construccions anteriors sense control d’activitat (sense GR), però fusionades a la proteïna
verda fluorescent (GFP). Aquestes construccions s’han obtingut a partir del mateix insert usat
anteriorment, però fent servir com a vector binari de destí el plàsmid pCAMBIA1302, modificat
perquè l’insert quedi en fase amb la GFP per digestió amb BamHI (construccions usades en
l’estudi de localització subcel·lular). Amb aquestes noves línies de sobre-expressió es podrà
veure el fenotip en estadis més adults sense necessitat d’induir l’activitat de la proteïna. Les
dues truncacions agafades per aquest estudi han estat les equivalents a les construccions
pMG27 i pMG28 (MG58 i MG62, en la part inferior de la figura C1.8), sent la primera
construcció la més llarga que no mostra activitat i la segona la més curta que si que mostra
activitat biològica. Les línies transgèniques per aquestes construccions estan encara en procés
de selecció, però es comentarà a finals del següent apartat algun dels resultats preliminars.
Figura C1.10. Anàlisis d’activitat biològica de les construccions derivades d’ATHB4 (II) en tractaments d’ombra simulada. (a)
Esquema indicant les condicions de creixement: les plantes van ser crescudes des de dia 0 amb o sense DEX, es van deixar créixer 2
dies en W i es va transferir a W+FR a partir del segon dia i fins a dia 7. (b) Mesures de longitud de l’hipocòtil per les diferents
construccions (II), es mostra un experiment i una línia representativa. Els símbols indiquen diferències significatives (0 P<0.05; 00
P<0.01) respecte el mateix genotip creixent en iguals condicions de llum però sense DEX (-DEX).

3 Capítol 1
53
Exp
ress
ióre
lati
va
HAT2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Col-0 pCS19 pMG43.9
00
00
SAUR15
Col-0 pCS19 pMG43.90,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
GR
pCS19 pMG43.90,00,20,4
0,60,8
1,01,21,41,6
0
00
Wd0 d7
+1h W+FR- DEX
+ DEX
HAT2 SAUR15 Nt HD Zp Ct
(a)
(b)
Nt HD Zp
HD Zp
Nt HD Zp Ct
Zp
CtZp
HD Zp Ct
HDNt
Nt HD Zp Ct
Nt HD Zp Ct
Nt HD Zp Ct
N HD Zp Ct
Nt
HD
Nº total línies independents
analitzades
Fenotip nan en W+DEX
Respostes fisiològiques atenuades en
W+FR
Respostes moleculars atenuades en W+FR
Homodimerització (Y2H)
Localitzatió (microbombardeig)
pCS19 Ref. + + + + N
pMG23 3 + + + n.a. N
pMG24 3 - - - + N
pMG25 2 - - n.a. n.a. N + C
pMG26 5 - - /+a - n.a. N + C
pMG27 7 - - /+b - + N
pMG28 2 + + + n.a. N
pMG29 3 - - n.a. n.a. N
pMG37 4 + + n.a. n.a. n.a.
pMG38 4 - - /+c n.a. n.a. n.a.
pMG43 4 - - /+d + n.a. n.a.
pMG44 3 - - /+e n.a. n.a. n.a.
pMG60 n.a. n.a. n.a. n.a. n.a. N + C
Nt HD Zp
HD Zp
Nt HD Zp Ct
Zp
Zp
HD Zp Ct
HDNt
Nt HD Zp Ct
Nt HD Zp Ct
Nt HD Zp Ct
N HD Zp Ct
Nt
HD
Nº línies obtingudes
(T1)
Nº línies amb 1 inserció
(T2)
Nº línies seleccionades per expressió
(T2)
Nº total línies independents
analitzadesMostrades
Línies analitzades
en qPCR
pCS19 Ref. Ref. Ref. Ref. Ref. Ref.
pMG23 33 21 3 3 23.1 23.1
pMG24 36 7 3 3 24.12 24.12
pMG25 27 13 3 2 25.5 n.a.
pMG26 33 13 4 5 26.7 26.7
pMG27 30 10 5 7 27.12 27.3 i .12
pMG28 14 5 4 2 28.4 28.4
pMG29 22 3 3 3 29.1 n.a.
pMG37 24 8 4 4 37.9 n.a.
pMG38 21 6 4 4 38.18 i .14 n.a.
pMG43 26 6 4 4 43.9 43.9
pMG44 23 6 3 3 44.2 n.a.
pMG60 26 n.a n.a. n.a. n.a. n.a.
Ct
Figura C1.11. Efecte de la sobre-expressió de la construcció pMG43 sobre l’expressió de HAT2 i SAUR15. (a) Esquema
indicant les condicions de creixement: les plantes van ser crescudes des de dia 0 amb o sense DEX, es van deixar créixer fins a dia 7
i es van agafar mostres després de tractament en W+FR (1h). (b) Anàlisis d’expressió del marcadors moleculars HAT2 i SAUR15
d’una línia representativa per la construcció pMG43. Els símbols indiquen diferències significatives (0 P<0.05; 00 P<0.01) respecte el
control wt creixent en iguals condicions.
Taula C1.2 Resum de línies obtingudes i analitzades per les delecions d’ATHB4. Resum del nombre de línies obtingudes i
analitzades en cada generació per les diferents delecions d’ATHB4. Ref: Línia de referència, estable. = Mutació puntual, canvi de
residu a Ala.
Taula C1.1 Resum dels resultats amb les delecions d’ATHB4. Resum dels resultats obtinguts amb les delecions d’ATHB4. Ref:
Línia de referència, estable. n.a. = No analitzat; a: 3 línies sense activitat i 2 amb activitat feble; b: 5 línies sense activitat i 2 amb
activitat feble; c: 3 línies sense activitat i 1 una activitat feble; d: 2 línies sense activitat i 2 amb activitat feble; e: 3 línies sense activitat
i 1 una activitat feble; N = Localització Nuclear; C = Localització citoplasmàtica. = Mutació puntual, canvi de residu a Ala.

3 Capítol 1
54
3.1.2.5. ATHB4 uneix realment al DNA? Seqüència d’unió i dianes directes.
Tal com s’ha dit anteriorment, està demostrat que la proteïna ATHB2, molt relacionada
estructuralment amb ATHB4, uneix al DNA actuant com a repressor de la transcripció (Ohgishi,
et al. 2001, Steindler, et al. 1999). Recentment també s’ha publicat que HAT3 uneix al DNA en
experiments de ChIP actuant també com a repressor (Turchi, et al. 2013). A més, el mateix
treball posa de manifest que ATHB4 té certes activitats redundants no sols amb HAT3, sinó
també amb ATHB2, suggerint que alguns mecanismes moleculars han de ser compartits, com a
mínim respecte l’activitat relacionada amb el desenvolupament. No obstant, els nostres
resultats experimentals portarien a pensar que ATHB4 està actuant en respostes a la SAS de
manera independent a la unió al DNA, o si ho fa, seria de manera independent a l’activitat de
dimerització pel domini ZP i de la interacció amb el DNA del domini HD.
Per tal de comprovar si ATHB4 també actua com a repressor transcripcional unint al DNA
s’han realitzat diversos estudis al laboratori. Prèviament es va intentar sense èxit realitzar
assajos d’EMSA (de l’anglès Electrophoretic Mobility Shift Assay) amb proteïna ATHB4
recombinant (produïda en Escherichia coli o amb un sistema de traducció in vitro) i fragments
de DNA marcats radioactivament que contenien una seqüència consens d’unió per ATHB2
(Sessa, et al. 1993). Com que els assajos d’EMSA no van funcionar, es va passar a comprovar
la capacitat d’ATHB4 per unir al DNA mitjançant un array de proteïna (PBM, Protein Binding
Microarray), treball fet en col·laboració amb el grup del Dr. Roberto Solano (Centro Nacional de
Biotecnologia, Madrid). Aquesta tècnica, a més de comprovar in vitro la capacitat d’ATHB4 per
unir al DNA (en forma homodimèrica), permet trobar la seqüència consens on s’uneix. El que
vam fer va ser obtenir proteïna de fusió MBP-ATHB4 (MBP de l’anglès Maltose Binding
Protein), per tal que els col·laboradors poguessin purificar la proteïna mitjançant la MBP i
realitzar l’array. La proteïna es va produir en cultius d’E.coli transformats amb el vector pMG46
(amb l’insert ATHB4 en marc de lectura amb la MBP del vector pMALc2x), durant 4h d’inducció
amb IPTG. Com es pot veure en un assaig de Western-Blot amb anticòs anti-MBP extractes
dels cultius abans (T0h) i després de la inducció (T4h) mostren un augment clar de proteïna
recombinant (figuraC1.12a). Els extractes van ser enviats a Madrid per tal de fer l’array de
proteïna. De l’anàlisi informàtic d’aquest assaig s’obtenen uns índex d’enriquiment (E-score)
que reflecteixen l’afinitat d’unió de la proteïna amb les diverses seqüències de DNA presents al
microarrray (Godoy et al. 2011). Analitzant aquestes seqüències es poden identificar les
regions on la proteïna presenta més afinitat, i per tant, una seqüència consens d’unió al DNA.
En el nostre cas, ATHB4 va presentar una seqüència consens molt semblant a la descrita per
ATHB2 (Sessa, et al. 1993), amb una seqüència 7-mer palindròmica AATNATT (on N pot
representar qualsevol base), però mostrant més afinitat per les bases C o G al centre (figura
C1.12b i c). Tot i això, ampliant la seqüència consens a la 9-mer, es va poder veure que les
posicions 1 i 9 tenen preferentment la base A, mentre que altres membres de la subfamília
uneixen preferencialment a seqüències palindròmiques amb T i A respectivament. A més es va
poder veure que, si bé canvis en les bases centrals no són essencials per la unió i varien poc

3 Capítol 1
55
l’índex d’enriquiment, les posicions 2 i 6 si que són molt específiques, i canvis d’aquestes bases
rebaixen molt l’afinitat d’unió al DNA (figura C1.12d).
9‐mer7‐mer
5‘ ‐ TAAT ATTA ‐ 3’
T0 A
T0 B
T4 A
T4 B
α‐MBP
Comassie
(a) (b)
(c)
GC
MBP ATHB4
0.6
0.4
0.2
0.0
-0.2
AA
T(G
/C)A
TT
AA
T(A
/T)A
TT
AT
T(A
/T)A
AT
AT
T(G
/C)A
AT
(d)
E-
sc
ore
Així doncs, tal com està publicat per ATHB2 i HAT3, ATHB4 també uneix DNA en unes
seqüències pseudopalindròmiques semblants. Per tal de identificar gens regulats directament
per ATHB4 es va treure partit d’aquesta informació i dels anàlisis de microarray comentats
anteriorment (apartat 3.1.3), d’on havíem obtingut una llista de 53 gens potencialment regulats
per ATHB4. Tenint la seqüència d’unió consens es va fer unes anàlisis bioinformàtiques mirant
gens que contenen aquestes caixes al seu promotor i es van creuar les dades amb els resultats
del microarray de la línia pCS19 amb tractaments amb i sense DEX i amb i sense CHX. Les
anàlisis sobre les seqüències consens als promotors es van fer usant els 1000 parells de bases
(pb) anteriors a l’origen de transcripció mitjançant dues eines bioinformàtiques disponibles on-
line (Patmatch disponible a la base de dades TAIR i AthaMap, disponible a
http://www.athamap.de). Vam prendre llistes de gens amb les diferents variants de la caixa
consens, amb C o G al centre i amb A o T, i també amb la seqüència 7-mer i 9-mer. Aquestes
llistes es van solapar amb la llista de 53 gens provinent de les dades dels microarrays (figura
C1.13b). D’aquesta manera vam identificar un petit nombre de gens que potencialment han de
ser reprimits de manera directa per ATHB4 i que contenen en la seva regió promotora la caixa
consens d’unió. Usant la seqüència 9-mer (figura C1.12b) es va obtenir una llista que conté un
total de 31 gens (figura C1.13b), entre els quals es troben membres de la seva família com
HAT1, HAT2 i HAT22, però també altres gens de respostes a hormones i un transportador de
pèptids (OPT1)(Koh et al. 2002)(taula C1.3).
Amb aquesta llista de possibles gens diana directes, es va voler analitzar la unió
d’ATHB4 en les caixes trobades als seus promotors mitjançant la immunoprecipitació de
cromatina (ChIP). Els primers intents de la ChIP es van realitzar durant una estada de dos
mesos al laboratori del Dr. Christian Fankhauser (Université de Lausanne, Switzerland),
utilitzant la mateixa línia pCS19 (35S:ATHB4-GR) i un anticòs anti-GR. Aquest mateix anticòs
havia estat utilitzat amb èxit per altres autors (Kaufmann et al. 2010). En aquests experiments
es va incloure com a control positiu una línia 35S:PIF5-HA utilitzant un anticòs anti-HA, tal com
s’havia descrit (Hornitschek, et al. 2012). Al usar l’anticòs contra GR no es va aconseguir veure
mai cap enriquiment en cap dels promotors del gens mirats, si bé es va aconseguir enriquiment
Figura C1.12. Array de proteïna. (a) Anàlisis per Western-blot d’extractes de
proteïna total de cultius d’E. coli abans (T0) i després d’una inducció amb IPTG
durant 4h (T4). (b) Seqüències de consens d’ATHB4 resultat de l’array de
proteïna (PBM, Protein Binding Microarray). (c) Seqüència consens descrita per
ATHB2. (d) Representació en diagrama de caixes de puntuació d’enriquiment
(E-score).

3 Capítol 1
56
en promotors publicats com directament regulats per PIF5 en les mostres dels experiments de
ChIP corresponents. Diverses proves basades en western-blot amb anticòs anti-GR van
demostrar que anticossos contra aquest domini no són gaire eficients en plantes, ja que donen
lloc a unions inespecífiques i malgrat diversos intents es va haver de descartar l’ús d’aquest
anticòs.
- CHX (251) + CHX (532) Directes (53) Seq. Consens (18.605)(a)
(b)
Per tot l’anterior, es va passar a provar de fer la ChIP amb una línia transgènica
disponible al laboratori que expressava la proteïna ATHB4-GFP sota el promotor 35S (línia
pBJ3), permetent utilitzar anticossos anti-GFP. Aquestes línies presentaven un fenotip menys
intens que les línies que expressaven constitutivament ATHB4 sola (35S:ATHB4) (Sorin, et al.
2009), probablement perquè l’etiqueta GFP afectava a l’activitat de la proteïna. No obstant, en
el procés d’obtenció de línies independents i homozigotes es va veure que en estadi de
plàntula el fenotip era prou clar i que en molts casos les plantes adultes també presentaven
característiques de sobre-expressió d’ATHB4, com fulles més allargades i amb la làmina
cargolada cap amunt, i menor nombre de fulles, silíqües i llavors, característiques idèntiques a
la sobre-expressió d’ATHB2 o de HAT3 (Turchi, et al. 2013). En els primers estadis de
desenvolupament el mutant athb4hat3 presenta fenotips clars de polaritat, mentre que
posteriorment el fenotips es torna menys visible (Bou-Torrent, et al. 2012). A més, experiments
en ombra simulada van portar a concloure que la seva resposta era molt semblant a l’esperada,
amb un hipocòtil lleugerament més llarg en W i una resposta atenuada fortament en W+FR, de
manera semblant a la descrita per les línies pCS19 en DEX o les 35S:ATHB4. Així doncs,
basant-nos en aquests experiments, es va comprovar que l’activitat de la proteïna ATHB4-GFP
era semblant a la de la proteïna ATHB4 sola.
Figura C1.13. Diagrama de Venn per mostrar el solapament de les dades del mircorray. (a) Anàlisis de solapament de dades
entre els tractaments –CHX i +CHX per gens expressats diferencialment (BH<0.05 i FC>1.8, BH=Bonferroni-Hochberg; FC=Fold
Change) entre Col-0 i pCS19 comparant els tractaments +DEX i –DEX. Aquest grup de 53 gens estan regulats per ATHB4 en
presència de DEX i de manera independent de CHX. (b) Anàlisis de solapament del grup de 53 gens anterior amb dades
bioinformàtiques de gens que contenen la seqüència consens 9-mer (figura RI.12b) d’unió d’ATHB4 en regions de 1000pb del seu
promotor.

3 Capítol 1
57
Codi AGI Nº caixes Noms del gen Codi AGI Nº caixes Noms del gen
At2g32210 8 hypothetical protein At5g52310 2COLD REGULATED 78; LOW-TEMPERATURE-INDUCED 78
At4g12470 6 azelaic acid induced 1; AZI1 At4g17870 2 PYR1 ; PYRABACTIN RESISTANCE 1
At1g14540 6 peroxidase 4 At1g23090 2 sulfate transporter 91; SULTR3
At4g17460 6 HAT1 At5g61590 2 ethylene-responsive transcription factor ERF107
At5g17860 4 calcium exchanger 7; CAX7 At5g61600 2 ethylene-responsive transcription factor ERF104
At3g14225 4 GDSL-motif lipase 4; GLIP4 At1g29430 2 SAUR-like auxin-responsive protein
At5g52760 4 copper transport family protein At1g51805 2 leucine-rich repeat protein kinase-like protein
At5g64780 4 hypothetical protein At2g30010 2 TRICHOME BIREFRINGENCE-LIKE 45; TBL45
At2g27830 4 hypothetical protein At5g07580 2 ERF/AP2 transcription factor
At5g43170 4 zinc-finger protein 3; AZF3 At4g37790 2 HAT22
At3g18830 4 polyol/monosaccharide transporter 5; ATPLT5 At4g31800 2 WRKY DNA-binding protein 18; WRKY18
At5g47370 4 HAT2 At5g55930 2 oligopeptide transporter 1; OPT1At5g24210 4 lipase class 3 family protein At2g29740 1 UDP-glucosyl transferase 71C2; UGT71C2
At1g33960 4 AVRRPT2-INDUCED GENE 1 At2g18680 1 hypothetical protein
At5g14730 4 hypothetical protein At5g64770 1 hypothetical protein
At5g61520 4 sugar transport protein 3
Així doncs, es va passar a realitzar assajos de ChIP. Es van agafar mostres de plàntules
creixent fins a dia 7 en llum W contínua. Els primers experiments han portat a veure que
ATHB4 uneix directament regions concretes (regió 2, a -1580 bp de l’ inici de traducció) del
promotor de HAT2 (figura C1.14). Les condicions en que s’ha fet la ChIP, però, encara haurien
de ser optimitzades, ja que els percentatges de recuperació són baixos i alguns controls
negatius (com regions del promotor on no hi ha seqüències consens d’unió o els encebadors
en la regió codificant) donen un cert enriquiment. Per manca de temps en el present treball no
s’han pogut optimitzar més els resultats de ChIP, però s’espera properament poder reduir el
soroll de fons i obtenir així més precisió en definir els punt d’unió en les dianes directes
d’ATHB4.
0,000
0,005
0,010
0,015
0,020
0,025
0,030Regió2
CDS
Col-0 A Col-0 B ATHB4 A ATHB4 B0,000
0,005
0,010
0,015
0,020
0,025
0,030
0,035
Col-0 A Col-0 B ATHB4 A ATHB4 B
Regió 1
Regió 2
CDS
% In
pu
t
HAT1 HAT2
MGL13 MGL29
MGL14 MGL30
Regió1 Regió2
MGO53
MGO54
CDS
MGL9
MGL10
MGL7
MGL8
MGL5
MGL6
MG25
MGL26
Regió3Regió2Regió1 Regió4
MGL1
MGL2CDS
Com s’ha comentat anteriorment, s’han transformat en planta algunes de les delecions
fusionades a GFP enlloc de GR (construccions MG58 i MG62, imatges inferiors de la figura
C1.8). Esperem poder examinar l’activitat biològica d’aquestes construccions en estadis més
adults i a més analitzar la seva capacitat d’unió al DNA de manera similar als anàlisis fets per la
línia pBJ3. Estudis preliminars del fenotip d’aquestes construccions porten a pensar que la seva
Taula C1.3 Llista de gens regulats directament per ATHB4. Llista dels 31 gens resultants del solapament entre les dades dels
microarays i les anàlisis dels promotors, provinents de la figura RI.13b. Codi AGI: Codi del gen (Arabidopsis Genome Initiative); Nº
Caixes = Número de seqüències consens 9-mer trobades en les 1000pb anteriors a l’inici de transcripció del gen en qüestió.
Figura C1.14. Unió als promotors de HAT1 i HAT2 per la línia pBJ3. Experiments preliminars de ChIP amb anti-GFP, usant
plàntules de pBJ3 crescudes durant 7 dies en llum blanca. Es mostren dues rèpliques (A i B). CDS = Seqüència codificant (control
negatiu d’enriquiment); = Seqüència consens d’unió d’ATHB4 al promotor del gen.

3 Capítol 1
58
activitat en plàntula i en resposta a ombra no es correspon amb la seva activitat en plantes en
estadi de roseta. Si la construcció pMG27 no semblava tenir cap activitat en ombra, la
construcció MG58 sembla tenir una activitat contrària a ATHB4 en estadis de roseta, amb fulles
cargolades cap avall. No obstant, aquests resultats han de ser confirmats amb més línies i en la
generació T3, en línies homozigotes, que esperem obtenir en breu.


3 Capítol 2
60
3.2. Capítol 2: Estudi del paper de les NUPs en la regulació de
la SAS.
3.2.1. Introducció.
3.2.1.1. Nous components reguladors de la SAS: Les nucleoporines.
Dins dels objectius del laboratori, es van voler identificar nous components de la
transducció de la senyal d’ombra, potencialment reguladors de la SAS. Es va partir d’informació
prèvia, on s’havia vist que PHYB era un gen PAR regulat de manera directa per l’acció dels
fitocroms (Roig-Villanova 2007). Així, es va utilitzar una línia transgènica que contenia el
promotor de PHYB de tabac fusionat al gen reporter de la luciferasa (línia PBL, PHYB:LUC, en
ecotip Ws-2, proveïda pel Dr. Andrew Millar, UK) (Bognár et al. 1999). Tractaments de només
2h amb ombra simulada van incrementar fortament l’activitat luciferasa, resultat que era
consistent amb els treballs previs esmentats.
La línia PBL es va mutagenitzar químicament per tractament amb EMS, reactiu que
introdueix mutacions puntuals aleatòries en el genoma. A partir d’aquesta població es va
establir un escrutini per tal de seleccionar aquells mutants que mostraven una activitat
luciferasa alterada en resposta al tractament d’ombra simulada, indicant així una disfunció en
l’expressió de PHYB en resposta a l’ombra simulada, probablement per l’alteració de la funció
d’algun component regulador de la SAS. El crivellatge va ser fet durant una tesi anterior en
col·laboració amb el laboratori del Dr. P. Devlin (Royal Holloway, University of London, UK).
Resultat del crivellatge va ser la identificació de mutants que anomenàrem dracula (dra), ja que
presentaven respostes atenuades a l’ombra, com si els agradés la foscor.
El primer mutant trobat, dra1, va resultar ser un nou al·lel de PHYA, fet que va servir per
comprovar que les condicions elegides eren bones per tal d’obtenir mutants amb un paper en la
regulació de la SAS aigües amunt del promotor del PHYB, en les primeres etapes de percepció
i resposta a la llum (Wang et al. 2011). El segon mutant, dra2, mostrava major activitat
luciferasa després del tractament d’ombra i presentava un fenotip en plàntula molt marcat, amb
un hipocòtil més llarg i cotiledons molt hiponàstics. Les plantes adultes també tenien severs
defectes de desenvolupament, amb floració primerenca i menor nombre de fulles, que també
eren més petites, i silíqües més petites amb baixa producció de llavors (figura I.6a i b). El fet
que algunes d’aquestes característiques eren comunes a plantes creixent en ombra feia pensar
que efectivament era un mutant implicat en la regulació de la SAS. Es va comprovar que la
mutació tenia un caràcter monogènic i recessiu, suggerint que era una pèrdua de funció. El
mapeig de la mutació, fet en col·laboració amb el grup de la Dra. M. R. Ponce (Universitat
Miguel Hernández, Campus d’Elx, Alacant), va revelar que DRA2 corresponia al gen
At1g10390, que codifica una proteïna semblant a una nucleoporina (NUP), un component del
porus nuclear. La mutació trobada a l’escrutini, anomenada dra2-1, produïa un canvi d’un codó
TGG (W) a TGA (stop) en la regió codificant (exó 10), fet que truncava el producte gènic.

3 Capítol 2
61
Aquest resultat era consistent amb el caràcter recessiu de la mutació, que suggeria que era
una pèrdua de funció. Per tal de confirmar la implicació d’aquesta mutació amb la regulació de
la SAS es va analitzar la resposta de les plàntules dra2-1 a l’ombra simulada. Es van mesurar
les longituds dels hipocòtils amb i sense tractament d’ombra simulada en plàntules de 7 dies,
veient que en llum blanca els hipocòtils del mutant dra2-1 eren clarament més llargs que els de
les plàntules silvestres (línia PBL), mentre que en ombra simulada l’elongació del mutant
estava clarament atenuada (figuraI.6c). Els resultats suggerien la implicació de les NUPs en la
regulació de la SAS i per això es van analitzar les respostes a ombra en alguns altres mutants
de NUPs descrits, mostrant que també tenien atenuades les respostes fisiològiques en ombra
(Galstyan 2010) (figuraI.6d). A més es va veure que podia haver-hi un efecte en funció de
l’ecotip, ja que mutants en el gen NUP58 (també anomenat TCU1, TRANSCURVATA 1) en
l’ecotip Ler presentaven una atenuació en la resposta que no era present en els seus al·lels de
Col-0. De manera similar el fenotip característic de l’al·lel dra2-1 (en l’ecotip Ws-2) es perdia
ràpidament al creuar aquesta mutació en l’ecotip Col-0, suggerint que el nucleoporus pot ser
diferent entre els ecotips. Aquest efecte ha estat estudiat més en profunditat en la present tesi.
nup160-4 nup96-1Col-00
1
2
3
4
5
6
7
8
Col-0 nup58-40
1
2
3
4
5
6
7
8
Ler nup58-10
1
2
3
4
5
6
7
Lo
ng
itu
d h
ipo
còti
l(m
m)
PBL dra2-1
Lo
ng
itu
d h
ipo
còti
l(m
m)
0
1
2
3
4
5
6
00
0
PBL dra2-1
dra2-1PBL
(b)(a)
PBL dra2-1
(c)
0
0
0
00
00
00
0
(d)
W
W+FR
W
W+FR
d0 d2 d7
Les NUPs són una família d’uns 30 gens en Arabidopsis, que formen un macrocomplex
cilíndric anomenat nucleoporus (NPC, Nucleopore Complex). Aquest complex regula tant
l’exportació d’RNA del nucli al citoplasma com el trànsit de proteïnes, similars o majors de
40kDa, cap a l’interior i l’exterior del nucli (Patel et al. 2007). Components de menor tamany
poden entrar i sortir per mecanismes no específics com la difusió passiva o mitjançant petits
porus com canals iònics. El NPC ha en estat àmpliament estudiat en llevat i està sent estudiat
en animals, però encara es coneix poc del nucleoporus en les plantes. Les NUPs de plantes
van ser primerament identificades per homologia de seqüència amb les de llevat i mamífers, si
Figura I.6. Anàlisis de fenotips de les NUPs. (a) Plantes adultes de 6 setmanes. Cada quadre de la regleta representa 1cm (10cm
de llarg en total). (b) Superior: Detall de les silíqües. Inferior: Aspecte de plàntules de 7 dies. (c) Mesures de longitud d’hipocòtil del
mutant dra2-1 comparat amb el seu control wt. Les plàntules van ser crescudes 2 dies en W i tractades o no amb W+FR des del
segon dia i fins a dia 7. (d) Mesures de les diferents línies mutants per NUPs analitzades en treballs anteriors al laboratori,
experiment en iguals condicions que el mostrat en l’apartat (c).

3 Capítol 2
62
bé algunes s’han identificat en diversos escrutinis genètics implicats en mecanismes com
resistència a patògens, senyalització o resposta a auxines i etilè o estrès per fred (Dong et al.
2006, Parry et al. 2006, Robles et al. 2012, Wiermer et al. 2012, Zhang and Li 2005), suggerint
que tenen un paper molt important en el desenvolupament, tal com està descrit en animals
(Franks and Hetzer 2013, Raices and D'Angelo 2012). A més, quasi tots els mutants deficients
en NUPs presenten fenotips pleiotròpics, demostrant que la regulació del tràfic mitjançada pel
NPC també podria ser molt rellevant en plantes (Parry 2012, Tamura and Hara-Nishimura
2012).
Nup62
Nup54
Nup58
Nup35
Citoplasma
Nucli
DRA2
NE
ESD4
TREX-2
Ran-GAP
WIP
WIT
NDC1
GP210
CG
1
Nu
p2
14
NUA/TPR Nup1/Nup136
Nup50
NUP98
Nup155
Nup188
Nup205
Nup88Nup93
Sec13
Sec13
Nup160/SAR1Nup96/SAR3
Nup43 Nup85Nup107 Nup133
Nup160/SAR1Nup96/SAR3
Nup43 Nup85Nup107 Nup133
NUP98
Nup93 Nup88
Les NUPs han estat classificades generalment en 3 grups segons el seu paper en el
NPC: (1) Un petit grup de NUPs amb dominis de transmembrana ancoren el NPC a l’embolcall
nuclear; (2) Unes altres tenen filaments proteics hidrofòbics, amb múltiples repeticions dels
aminoàcids FG (FXFG en vertebrats i GLFG en llevats), formant una xarxa que bloqueja
selectivament el pas a través del porus; i (3) La resta estructuren l’anell que dóna lloc al porus
central i s’uneixen per un costat a les NUPs de transmembrana i per l’altra a les NUPs amb
repeticions FG; algunes d’elles (NUP43, Sec13, Seh1 i RAE) interaccionen mitjançant unes
repeticions dels aminoàcids WD, establint interaccions hidrofòbiques.
Independentment d’aquesta classificació general, les NUPs de plantes es poden separar
en 7 grups més petits al tenir en compte la seva localització dins el macrocomplex (Tamura and
Hara-Nishimura 2012)(figura I.7):
(1) Dues úniques NUPs de transmembrana ancoren el NPC a la membrana nuclear, la
GP210 (GLYCOPROTEIN OF 210 kDa) i la NDC1 (NUCLEAR DIVISION CYCLE1).
Les NUPs amb repeticions FG es localitzen en tres zones diferents, dues perifèriques i
una central. Entre les exteriors podem trobar les citoplasmàtiques o les nucleoplasmàtiques,
Figura I.7. Esquema del nucleoporus. Esquema del nucleoporus de plantes basat en homologia amb el de llevat i animal. Hi ha
un conjunt de nucleoporines amb repeticions FG centrals (NUP62, NUP54, NUP58 i NUP35), les perifèriques citoplasmàtiques
(CG1 i NUP214) i nuclears (NUP1 i NUP50). De les altres NUPs, hi ha dos grans complexos, el de NUP107-160 i el de la NUP155,
formant l’estructura de l’anell.

3 Capítol 2
63
ambdós amb els filaments FG lliures, sense interaccionar entre ells i penjant cap a l’exterior del
NPC, permetent interaccions amb els transportadors.
(2) Dues NUPs amb dominis FG (NUP214 i CG1) formen els filaments citoplasmàtics.
(3) Al nucleoplasma, l’anomenada cistella nuclear (basket) està formada per dues NUPs
amb dominis FG (NUP136/NUP1 i NUP50) i una NUP (NUA, NUCLEAR ANCHOR), amb
dominis més rígids que els FG (dominis long coiled-coil).
(4) El grup més nombrós de NUPs FG es troben al centre del porus, en el complex de
NUP62 (NUPs-FG centrals: NUP62, NUP58, NUP54, NUP35), i una sola més exterior (NUP98).
Aquestes formen la xarxa hidrofòbica cohesiva, on els seus filaments interaccionen de manera
transitòria entre ells per tancar el porus.
Tres grups de NUPs mantenen l’estructura cilíndrica del NPC.
(5) L’anell exterior està format per un complex amb 8 NUPs (complex NUP107/160:
NUP160, NUP133, NUP107, NUP96, NUP85, NUP43, Sec13 i Seh1) i unes NUPs més
exteriors (ALADIN, RAE/GLE i Elys/HOS1).
(6) L’anell interior està format per 4 NUPs (NUP205, NUP188, NUP155, i NUP35).
(7) Finalment dues NUPs, la NUP93 i la NUP88, fan de pont entre les altres proteïnes
estructurals i les NUPs-FG, estabilitzant el macrocomplex final.
Tres complexos exteriors, que interaccionen amb el NPC però no es troben anclats al
complex de manera estable, regulen també el trànsit nuclear: (1) El complex ESD4-NUA, unit a
la part nuclear del NPC per la NUP NUA (similar a la NUP de vertebrats TPR, Translocated
Promoter Region), està relacionat amb la sumoilació i el transport de conjugats a través del
NPC (Xu et al. 2007b), presentant els mutants en TPR/NUA defectes pleiotròpics, amb floració
primerenca, defectes en la senyalització d’auxines i en l’exportació d’RNA (Jacob et al. 2007);
(2) El complex per exportació d’RNA TREX-2 (Lu et al. 2010), situat també a la part nuclear,
s’uneix al NPC a través de la NUP1 i defectes en aquest complex comporten menor exportació
d’RNA, amb alguns components que presenten redundància de funció, però mostrant que
NUP1 és essencial, ja que el seu mutant és inviable; i (3) El complex d’importació i exportació
de proteïnes i RNA’s Ran-GAP (Xu et al. 2007a), que s’uneix a la vessant citoplasmàtica de la
membrana nuclear mitjançant les proteïnes WIP-WIT; els mutants simples de RanGAP1 i
RanGAP2 no presenten fenotip, mentre que el doble mutant és inviable (Rodrigo-Peiris et al.
2011). Aquests dos últims complexos reconeixen dos tipus de proteïnes que regulen el
transport nuclear: els factors d’exportació, reconeguts pel complex TREX-2, i les carioferines,
pel complex Ran-GAP. Hi ha 17 carioferines identificades en Arabidopsis (Merkle 2011) i un
nombre indeterminat de factors exportadors d’mRNA (Meier and Brkljacic 2010). El
reconeixement de les senyals de localització nuclear (NLS, de l’anglès nuclear localization
signal) o les senyals d’exportació nuclear (NES, nuclear export signal) presents en les
proteïnes a transportar, inicia el transport actiu amb consum d’energia en un procés anomenat
cicle Ran, mitjançant la unió dels transportadors a regions receptores concretes dels filaments
FG. Diversos mutants de carioferines han estat aïllats en estudis genètics per diferents rutes,
mostrant fenotips semblants a mutants en NUPs, implicant les carioferines en mecanismes com

3 Capítol 2
64
respostes a temperatura o tolerància a la sequera (Luo et al. 2013, Wu et al. 2010). A més,
mutants en els gens homòlegs de les exportina5 i exportinaT de vertebrats (anomenats en
Arabidopsis HASTY i PAUSED respectivament)(Bollman 2003, Hunter 2003, Li 2003)
presenten fenotips pleiotròpics de desenvolupament, afectant el desenvolupament de fulles,
temps de floració i formació de silíqües, de manera similar a molts mutants en NUPs.
El primer mutant de NUPs identificat en plantes va ser nup96/mos3 (MODIFIER OF snc1-
3), aïllat en un crivellatge per identificar components de defensa a patògens (Zhang and Li
2005). Poc més tard es van identificar els mutants sar1/nup160 i sar3/nup96 (SUPPRESSOR
OF AUXIN RESISTANT), trobats en un crivellatge buscant la supressió de la resistència a
auxines causada pel mutant axr1 (AUXIN RESISTANT1)(Parry, et al. 2006). A més de
demostrar que aquests gens eren realment NUPs, també es va veure que la seva mutació
impedia la correcte exportació de poliA+-RNA del nucli, mitjançant tècniques d’hibridació in situ
amb sondes oligo-dT-fluoresceïna (que hibrida la cua poliA+ dels mRNAs). A més de l’efecte en
l’exportació d’mRNA, es va suggerir que aquestes mutacions provocaven un menor transport
de proteïnes Aux/IAA dins al nucli, fets que, conjuntament, podien resultar en una menor
resposta a les auxines, proposant així el mecanisme molecular pel qual s’havien identificat
(Parry, et al. 2006).
Curiosament, la majoria de les NUPs identificades en estudis genètics (NUP160,
NUP133, NUP96 i NUP85) corresponen a components del complex NUP107-160, amb una
funció bàsicament estructural (Tamura et al. 2010), i especialment NUP160 i NUP96 han estat
identificades en diversos crivellatges publicats, suggerint que d’alguna manera aquestes dues
NUPs deuen ser rellevants per la correcta senyalització a través del NPC, però sense ser
essencials pel desenvolupament de la planta. Tot i que l’efecte en l’exportació dels mRNAs ha
estat confirmat en altres publicacions i usat com a explicació pels defectes en les diverses rutes
de senyalització, l’efecte en la importació de proteïna no ha estat confirmat. Mutants en
NUP160 mostren defectes en la senyalització per fred, però factors importants en aquestes
respostes, com ICE1, no mostren alterada la seva localització (Dong, et al. 2006), indicant que
el defecte en el transport podria ser específic per algunes proteïnes. De fet, també està per
determinar si l’acumulació d’mRNA al nucli és un efecte inespecífic o si, per contra, només s’hi
acumulen uns transcrits determinats.
Malgrat aquesta possible especificitat sembla que algunes NUPs, com NUP85 i NUP133,
poden ser completament redundants, ja que els mutants simples no mostren cap fenotip ni
resposta alterada (Wiermer, et al. 2012), mentre que mutants en tres altres membres del mateix
complex (NUP96, NUP160 i Seh1) presenten defectes en respostes de defensa, amb
acumulació d’RNA al nucli i fenotips pleiotròpics de desenvolupament. A més, NUP96 i
NUP160 podrien ser parcialment redundants, ja que el doble mutant NUP96NUP160 mostra un
fenotip molt més sever i pràcticament estèril (Parry, et al. 2006).
Més enllà del complex NUP107-160, dues altres NUPs s’ha descrit que són essencials,
NUP88 i NUP1/NUP136. Originalment identificades en un crivellatge en respostes de defensa,
van mostrar que la seva pèrdua de funció és letal (Cheng et al. 2009, Lu, et al. 2010). Defectes

3 Capítol 2
65
en la NUP RAE presenten defectes en la divisió cel·lular i un exportació d’RNA també afectat
en Nicotiana benthamiana (Lee et al. 2009). Dos mutants en NUPs centrals han estat estudiats
més recentment: la NUP58/TCU1, on el seu mutant també té un fenotip pleiotròpic amb fulles
més petites i floració primerenca (Ferrandez-Ayela et al. 2013); i la NUP62, amb línies de
reducció de funció presentant fenotips similars als anteriors (Zhao and Meier 2011).
Pel que fa a la resta de NUPs, fa poc es va comprovar que moltes de les prèviament
identificades per homologia de seqüència codifiquen realment membres del porus nuclear
(Tamura, et al. 2010), ja que la seva expressió en fusió a GFP localitza la senyal com a
perinuclear i experiments de co-immunoprecipitació (CoIP) porten a comprovar que moltes
d’elles, entre les quals DRA2, formen part d’un sol macrocomplex. A més es va veure que
l’estructura del NPC de plantes és bastant similar a la de llevat i animals.
La idea inicial que el NPC és fix i que les NUPs són un simple component estructural ha
estat rebatut àmpliament, demostrant que el NPC té components espai-temporals i que
diverses NUPs poden localitzar-se més enllà del NPC (Griffis et al. 2002, Raices and D'Angelo
2012). Una de les NUPs que ha mostrat tenir un paper molt rellevant més enllà del NPC és la
NUP98 d’animals (NUP100 de llevat), l’equivalent al que hem anomenat DRA2 en plantes.
3.2.1.2. DRACULA2 i el seu homòleg d’animals NUP98: Una NUP, moltes funcions.
DRA2 codifica per una NUP molt semblant a la NUP98 de mamífers (Xu and Meier
2008). Tal com s’ha comentat, NUP98 és una nucleoporina de repeticions FG situada a les
parts exteriors del nucleoporus (figura I.7). Basant-se en la seqüència aminoacídica de DRA2
es va veure que el seu extrem N-terminal (Nt) manté similitud amb altres proteïnes amb
repeticions FG, com la NUP62, mentre que el seu domini C-terminal (Ct) manté similituds amb
altres NUPs com SAR1/NUP160 o SAR3/NUP96 (Parry, et al. 2006). En humans, la NUP96 és
codificada pel mateix gen que la NUP98, només que són diferents variants per un procés
d’splicing alternatiu (Sun and Guo 2008). El domini Nt de la NUP98 conté una regió d’unió amb
RAE, per regular conjuntament l’exportació d’RNA (Pritchard et al. 1999), a més, dins l’Nt hi ha
regions que permeten l’associació amb diversos transportadors com carioferines. Pel que fa al
domini Ct, en gran part és considerada una senyal d’autoproteòlisis (Rosenblum and Blobel
1999, Sun and Guo 2008), que a més ancla la NUP98 al NPC per interacció amb altres NUPs
del complex NUP107-160, permetent que el domini FG quedi penjant a les vessants exteriors
del porus on pot interaccionar amb els factors transportadors (Chatel et al. 2012).
A més de ser un component del NPC, la NUP98 està descrita com a mòbil, podent
localitzar-se també en el nucleoplasma. Dins del nucli s’han descrit unes vesícules on
s’acumula NUP98 (nuclear FG-bodies), però sense cap activitat definida (Griffis, et al. 2002).
Per això diversos grups d’investigació suggereixen que els cossos FG nuclears són resultat
d’un excés artificial de NUP98, ja que van ser observats en cultius cel·lulars de HeLa-C on
s’expressava NUP98 en nivells més alts del normal i en cultius cel·lulars que expressaven
NUP98 ectòpicament. Altres NUPs també tenen localitzacions més enllà del NPC, com
RAE/Gle1, un factor d’exportació d’RNA que també és present al citoplasma i dins el nucli (Lee,

3 Capítol 2
66
et al. 2009, Pritchard, et al. 1999), o NUP50 i NUP153, que també són presents al
nucleoplasma (Guan et al. 2000).
Està descrit que la cromatina descondensada (o eucromatina) es troba sovint
interaccionant amb el NPC, mentre que la cromatina condensada (o heterocromatina) es troba
sovint tocant l’envolta nuclear, fet que va portar a suggerir que el NPC podria actuar com a
remodelador de la cromatina. De fet, s’ha vist que la interacció de la cromatina amb el NPC
estabilitza els llaços d’eucromatina i facilita la transcripció dels gens corresponents, sent
aquesta interacció dependent de TPR i de la NUP50 (Ishii et al. 2002, Krull et al. 2010). No
obstant, la localització de NUPs al nucleoplasma ha fet pensar que poden unir a la cromatina
sense formar part del NPC (Raices and D'Angelo 2012).
NUP98 és una proteïna amb moltes activitats. Com a bona NUP és important en
l’exportació d’RNA i en l’import de proteïnes (Blevins et al. 2003, Oka et al. 2010, Terry and
Wente 2007). Però també està regulant l’expressió gènica per un mecanisme que encara no
està del tot clar, però on pot unir directament la cromatina en regions riques en caixes GA, tant
en Drosophila (Capelson et al. 2010, Kalverda et al. 2010) com en cultius de cèl·lules humanes
(Liang et al. 2013), controlant així l’expressió de gens importants pel desenvolupament. Es
postula que NUP98, NUP50 i NUP62 podrien unir els mateixos sectors de la cromatina, donat
que menors nivells d’expressió d’aquests gens porten a menor expressió de gens associats a la
regulació per NUPs, mentre que major nivells d’expressió d’aquestes NUPs porten a un
augment de l’expressió d’aquests gens marcadors, sent partícips de la regulació en
desenvolupament i del cicle cel·lular (Kalverda, et al. 2010). Ja que NUP98 no sembla tenir cap
domini d’unió al DNA, s’ha postulat que ho fa mitjançant proteïnes adaptadores que li faciliten la
interacció amb la cromatina, però encara no estan clars aquests mecanismes. NUP98 també té
un paper en la divisió cel·lular, per exemple regula la divisió mitòtica en associació amb els
microtúbuls i també influeix el temps de ruptura sent part dels mecanismes del punt de control
de la mitosis (SAC de l’anglès, Spindle Assembly Checkpoint) (Cross and Powers 2011,
Jeganathan et al. 2005). Per acabar de complicar l’activitat de NUP98, s’ha vist que, a més de
tot l’anterior, té capacitat d’estabilitzar alguns RNAs específics en cultius de cèl·lules animals,
concretament NUP98 estabilitza l’mRNA de la proteïna p21 (que s’ha relacionat en part amb la
resposta cel·lular a l’estrès) al unir-se a la seva regió 3’-UTR, suggerint que pot tenir una
activitat antitumoral (Singer et al. 2012).
Les NUPs han estat involucrades en processos de desenvolupament, però també en
diverses patologies, especialment oncològiques. Està descrit que, sobretot les NUPs
perifèriques, poden patir processos de translocació cromosòmica, donant lloc a proteïnes de
fusió entre NUPs i altres proteïnes, que poden ser factors de transcripció. Per exemple, s’ha
vist que la NUP214, normalment amb localització perinuclear, quan forma una proteïna de fusió
aberrant també es troba dins el nucli, donant lloc a defectes en l’exportació d’RNAs i de
proteïnes (Saito et al. 2004). En el cas particular de NUP98, s’han identificat més de 20
proteïnes de fusió en malalts de leucèmia mieloide, causant problemes de comportament de les
cèl·lules mare hematopoètiques, del moll de l’ós. Totes aquestes proteïnes de fusió

3 Capítol 2
67
comparteixen el domini FG (domini Nt), que podria tenir una activitat com a activador o
repressor transcripcional, fusionat a altres factors d’unió al DNA. Més de la meitat de fusions
que donen lloc a càncer són amb factors de la família homeobox (HOX, NUP98-Hox) donant
lloc a regulacions anormals en l’hematopoèsis (Bai et al. 2006, Kasper et al. 1999, Yasuhara et
al. 2009). D’altra banda, NUP98 sembla que regula específicament el transport d’una proteïna
implicada àmpliament en processos de càncer (galectin-3), mostrant-se que NUP98
interacciona amb galectin-3, afectant la senyalització mitjançada per aquest factor i controlant
l’expressió de la ß-catenina (Funasaka et al. 2013).
En el cas de la NUP98 humana, mirant dianes en cèl·lules mare embrionàries, s’ha vist
recentment que uneix a un subgrup de gens actius involucrats en el cicle cel·lular i en la
regulació de l’estructura de la cromatina i el silenciament gènic (Liang, et al. 2013). En canvi,
comparant els resultats anteriors amb línies cel·lulars humanes diferenciades, com són cèl·lules
progenitores neurals i cèl·lules de fibroblast pulmonar IMR90, NUP98 mostra unió a diferents
gens actius de funció neural i en zones silenciades de la cromatina, respectivament. Sembla
que les dianes de NUP98 són diferents segons l’estadi de desenvolupament/diferenciació de
les cèl·lules, suggerint que l’ancoratge de la cromatina al NPC pot ser rellevant durant les
primeres etapes del desenvolupament, possiblement perquè en aquestes etapes l’expressió de
gens no és gens estable i necessiten aquest anclatge al NPC per mantenir la cromatina
descondensada, especialment al llarg de cicles de divisió ràpids com en cèl·lules progenitores
neurals (Liang, et al. 2013). Per altra banda, en estats de desenvolupament més avançats, on
els teixits ja estan ben diferenciats i les expressions són més estables, la cromatina ja té les
zones actives establement obertes i no requereixen de la seva unió al NPC per mantenir la
transcripció activa (Liang, et al. 2013). Aquest fet suggereix que NUP98 a més de controlar el
transport a través del nucleoporus pot interaccionar dinàmicament amb la cromatina humana,
tant formant part el NPC com de manera independent, donant un nivell addicional de regulació
durant el desenvolupament.
El fet que NUP98 hagi estat implicada en processos cancerígens i s’hagi apuntat que
podria actuar com antitumoral ha fet que actualment estigui sent molt estudiada en animals. En
un recent treball han relacionat l’activitat de NUP98 amb les marques epigenètiques que
regulen l’anomenada “memòria transcripcional”, un mecanisme conservat en organismes uni i
pluricel·lulars (Light et al. 2013). Quan hi ha un dany al DNA, s’activen mecanismes de
reparació i protecció, expressant nous gens, i aquests canvis d’expressió es mantenen durant
diverses generacions mitjançant modificacions epigenètiques als promotors. D’aquesta manera
es pot dir que s’estableix una memòria transcripcional que perdura durant unes quantes
divisions cel·lulars. S’ha vist que el manteniment d’aquestes marques epigenètiques requereix
la interacció de NUP98/NUP100 amb els recent expressats promotors. Aquesta interacció
comporta la dimetilació de la histona H3 lisina 4 en el nucleosoma dels promotors dels gens
expressats, seguida per la iniciació del complex RNAPII que evoca la ràpida reactivació del gen
després de la divisió cel·lular. Menors nivells d’expressió de NUP98 en cèl·lules humanes o de
NUP100 en llevats porta a la pèrdua de la metilació H3K4me2 i la no iniciació del complex

3 Capítol 2
68
RNAPII en els promotors recent expressats i per tant una abolició de la reactivació dels
mateixos després de la divisió. Així, donat que les NUPs són les primeres proteïnes a contactar
amb la cromatina durant la divisió, i que a més, són requerides per la correcta descondensació
de la cromatina (Liang, et al. 2013, Walther et al. 2003), es conclou que les NUPs són
candidats a regular la transcripció en base a la ‘’memòria transcripcional’’ present durant la
mitosis (Light, et al. 2013).
Les marques epigenètiques com la metilació han estat recentment estudiades en
mecanismes importants per Arabidopsis, com el rellotge circadià (Henriques and Mas 2013,
Malapeira et al. 2012). Això fa pensar que més enllà de ser un mecanisme puntual, la regulació
del desenvolupament i la divisió cel·lular pot venir regulada fortament per aquets mecanismes i
que altres rutes, com les respostes de la SAS, també podrien estar regulades per mecanismes
semblants. El primer pas però, serà establir si diferents components del NPC afecten aquestes
respostes, per en treballs posteriors analitzar-ne els seus mecanismes de funcionament.
3.2.2. Resultats.
3.2.2.1. Anàlisis del mutant dra2-1: Caracterització general i fisiològica.
Com s’ha comentat anteriorment, en el laboratori s’havia identificat el mutant dra2-1 que
contenia una mutació en un gen codificant per la NUP98. Com altres mutants en NUPs
presentava un fenotip pleiotròpic amb floració primerenca, dominància apical i fulles i silíqües
més petites. La seva resposta a ombra estava clarament atenuada, i també presentava menor
resposta als tractaments d’auxines (Galstyan 2010). Donada la coincidència de fenotip del
mutant amb l’aspecte de plàntules silvestres creixent en medis suplementats amb auxines, es
va pensar que podia ser que el mutant tingués els nivells endògens d’aquesta hormona
incrementats. Per això es van quantificar els nivells d’auxina en aquestes plantes, en
comparació al seu control wt (línia PBL). Es van recollir uns 100mg de teixit fresc i es van
enviar per la seva quantificació al grup del Dr. Aurelio Gómez (Universitat Jaume I, Castelló).
Els resultats inicials semblen apuntar que els nivells d’IAA són semblants en el mutant respecte
al control, marcant una tendència fins i tot a ser menors en el mutant (figura C2.2a). Aquestes
quantificacions indicarien que el fenotip del mutant dra2-1 no és degut a una major quantitat
d’auxines. Per corroborar aquests resultats i per intentar disminuir l’error experimental, fet que
permetria discriminar millor les diferències, aquestes quantificacions estant sent repetides.

3 Capítol 2
69
0µM eBL
0,1µM eBL
1µM eBL
0
1
2
3
4
5
6
7
8
PBL dra2-1Col-0 dra2-4
Lo
ng
itu
d h
ipo
còti
l(m
m)
0
1
2
3
4
5
6
nup96-1 nup96-3Col-0 nup160-4
00
00
00
000
00
00
00
0
00
00
00
0
0µM GA3
5µM GA3
10µM GA3
0
1
2
3
4
5
6
7
PBL dra2-1Col-0 dra2-40,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
nup96-1 nup96-3Col-0 nup160-4
Lo
ng
itu
d h
ipo
còti
l(m
m)
00
00
0000
00 00
00
00
00
00 00
00
0000
Wd0 d7
Tractament hormonal corresponent
(a)
(b)
(c)
Dins de la caracterització de la línia mutant es va decidir mirar també les respostes
fisiològiques a altres tractaments, com les respostes a brassinoesteroides (eBL) o gibberel·lines
(GA3, en col·laboració amb altres membres del grup). Es va fer l’experiment pel mutant dra2-1 i
el seu control wt (PBL), però a més es van afegir un al·lel en fons genètic Col-0 (dra2-4) i
mutants en les NUP96 i NUP160. Es van créixer les plàntules durant 7 dies (des de l’ inici de la
germinació) en medi suplementat amb concentracions creixents de les diferents hormones i es
va mesurar la longitud de l’hipocòtil en resposta als tractaments aplicats (figura C2.1). Aquestes
hormones són inductores de creixement, pel que produeixen un allargament de l’hipocòtil en el
control wt. Comparant aquestes elongacions del wt amb el mutant dra2-1 es va veure que
mentre tenia menor resposta a BRs (al igual que s’havia descrit per a la respostes a auxines),
semblava respondre lleugerament més a GA (en ambdós casos les anàlisis de variança en
taula de dues cues, TW-ANOVA, mostren diferències significatives entre el genotip i la resposta
al tractament). A més es pot veure que l’al·lel de DRA2 en Col-0, dra2-4, mostra alteracions
molt més febles, responent gairebé sempre igual que el seu control wt, mentre que els mutants
en NUP96 i NUP160 mostren respostes atenuades en auxines i BRs tal com passa per dra2-1,
i una resposta a GAs que no arriba a ser estadísticament diferent que el wt. D’aquesta manera
veiem que els mutants en NUPs afecten no només les respostes a auxines sinó també a altres
hormones, però sembla haver-hi diferències segons l’ecotip i els al·lels, ja que no responen de
la mateixa manera ni els dos al·lels per DRA2 ni els mutants de DRA2 comparats amb els de
NUP96 i NUP160.
Figura C2.1. Respostes a hormones pels mutants dra2-1, dra2-4 i nup160-4, nup96-1 i nup96-3. (a) Esquema indicant les
condicions de creixement: les plantes van ser crescudes des de dia 0 amb el tractament d’hormona corresponent i es van deixar
créixer fins a dia 7 en llum W. (b) Mesures de longitud de l’hipocòtil en resposta a GA3. (c) Mesures de longitud de l’hipocòtil en
resposta a eBL. Esquerre: Mutants per DRA2. Dreta: Mutants en NUP160 i NUP96. El símbols indiquen diferències significatives
(0 P<0.05; 00 P<0.01) respecte el control wt creixent en igual condició.

3 Capítol 2
70
PBL
dra2-1
ng
/mg
fw
0
50
100
150
200
250
300
350
400
450
500
Clorofil·les Carotenoides
0% sucrosa
0
200
400
600
800
1000+2% Sucrosa
Clorofil·les Carotenoides0
5
10
15
20
25
30
35
ng/g
fwPBL dra2-1
IAA(a) (b)
Està descrit que el nivell de pigments fotosintètics varia en resposta a ombra (Roig-
Villanova, et al. 2007). Com que el mutant va ser identificat en un screening mesurant l’activitat
LUC, la concentració de pigments podria afectar als resultats, ja que una major acumulació de
pigments donaria lloc a una atenuació de l’activitat LUC, mentre que per contra, una menor
quantitat de pigments facilitaria quantificar més activitat luciferasa. Per això es va procedir a
quantificar els nivells de pigments, concretament els nivells de clorofil·les i carotenoides. Es van
sembrar plaques de plàntules control PBL i del mutant dra2-1 en 0.5xMS-, es van créixer 7 dies
en llum W i es van extraure i quantificar els pigments fotosintètics (Rodriguez-Concepcion et al.
2004). Els nivells de clorofil·les i carotenoides no eren significativament diferents entre els dos
genotips (figura C2.2b). No obstant, per mesurar l’activitat LUC en plàntules s’afegeix sucrosa
al medi i està descrit que la sucrosa també afecta als nivells de pigments (Dijkwel et al. 1997),
per això es va decidir mirar si els nivells de pigments estaven alterats en el mutant en presència
de sucrosa al medi. Es va recollir material com s’ha indicat anteriorment però de plàntules
creixent en medi 0.5xMS2, que conté el 2% de sucrosa, tal com s’havia fet per l’escrutini (figura
C2.2b).Tampoc es van veure diferències significatives, fet que va portar a concloure que
l’activitat luciferasa alterada del mutant era una resposta real i no un artefacte de canvis en la
concentració de pigments que podrien alterar la quantificació de la llum.
DRA2 GFP pCT935S
Nt GFP pMG5635S
dra2-1DRA2 Nt
Ct GFP pMG5535S
Arribat aquest punt, i per veure si DRA2 realment codificava una NUP, es va voler clonar
el cDNA sencer de DRA2, per tal d’expressar-lo com a proteïna de fusió amb la GFP i veure’n
la localització subcel·lular. Diversos intents van resultar infructuosos en tesis anteriors, de
manera que en el present treball vam optar per provar de fer el clonatge de dos fragments
d’aquest gen per separat i mirar-ne la localització. Com s’ha comentat a la introducció, DRA2
es pot separar en dues parts: la corresponent a les repeticions FG (que vam anomenar Nt) i la
que sembla d’incorporació al nucleoporus (anomenada Ct). Es van amplificar aquests dos
Figura C2.2. Quantificacions d’auxines i de pigments pel mutant dra2-1. (a) Quantificacions d’IAA lliure en nanograms
d’hormona per gram de teixit fresc (fw, fresh weigh). Mostres creixent durant 7 dies en W. (b) Quantificacions de pigments en
nanograms de pigments per mil·ligram de pes fresc de mostra, creixent durant 7 dies en medi 0.5xMS- (imatge de l’esquerre) o
creixent en medi 0.5xMS2 (imatge de la dreta). Les diferències entre genotips no són significatives en cap dels casos mostrats.
Figura C2.3. Construccions derivades per DRA2. Esquema de les construccions codificants per truncacions de la proteïna
DRA2. A l’esquerre s’indica el promotor que dirigeix l’expressió de la construcció en les plantes transgèniques i a la dreta
apareix el nom donat a la construcció. El primer esquema correspon al producte esperat en el mutant dra2-1. Nt = regió amb
repeticions FG N-terminal; Ct = regió C-terminal

3 Capítol 2
71
fragments per PCR utilitzant primers específics i es van clonar en un vector comercial. Els
inserts van ser introduïts en un vector binari pCAMBIA1302 que incorpora l’etiqueta GFP per fer
les proteïnes de fusió corresponents (figura C2.3). Es va estudiar la seva localització per
expressió transitòria en cèl·lules de la epidermis de porro i ceba transformades per biolística
(figura C2.4), mostrant iguals resultats en els dos tipus de teixits vegetals (es mostren els
resultats en cèl·lules de porro). Tal com s’ha explicat al capítol 1, per tal de visualitzar amb més
facilitat les cèl·lules transformades durant el bombardeig es van co-transformar amb el vector
pDsRed, que codifica per la proteïna la proteïna vermella fluorescent (RFP). A més es va afegir
uns transformació amb una proteïna nuclear com a control (construcció pBJ3, codificant per
35S:ATHB4-GFP). La proteïna Ct-GFP (pMG55) va mostrar una localització semblant a la GFP
sola (pMS51) i colocalitzava amb la proteïna RFP (pDsRed), mentre que la construcció Nt-GFP
(pMG56) mostrava una localització en vesícules al citoplasma i dins el nucli. Tal com
s’esperava, la proteïna ATHB4-GFP (pBJ3) es localitzava al nucli (figura C2.4). Finalment,
altres membres del grup van obtenir la construcció 35S:DRA2-GFP que contenia el gen sencer
per DRA2 (pCT9), però la seva sobre-expressió semblava ser poc o gens eficient, sense poder
detectar-se fluorescència (resultats no mostrats). Aquest resultats indicaven la regió Nt de
DRA2 conté alguna senyal que permet l’entrada de la proteïna al nucli, però que no és suficient
per anclar aquesta NUP al NPC. A més, sembla que un excés de DRA2 podria ser acumulat en
vesícules d’activitat desconeguda.
No s’ha pogut veure la localització típica de les NUPs com a perinuclear, no obstant, la
localització en vesícules nuclears de la proteïna Ct-GFP (pMG56) és consistent amb les dades
sobre NUP98 en cultius cel·lulars. S’estan generant plantes transgèniques sobre-expressant
aquestes dues construccions sota el promotor 35S en Col-0. L’obtenció i anàlisis d’aquestes
línies transgèniques s’ha fet en col·laboració amb una estudiant de màster (Lorenzo-Orts
2013). S’ha vist que les línies pMG55 (35S:Ct-GFP) no mostren expressió de la GFP ni fenotip,
deixant entendre que aquest fragment de la proteïna és poc estable; mentre que les línies
pMG56 (35S:Nt-GFP) mostren senyal d’activitat GFP en petites vesícules, de manera similar al
resultats de microbombardeig, i presenten fenotips bastant forts, amb defectes pleiotròpics de
desenvolupament, el que suggereix que DRA2 pot tenir una funció més enllà del nucleoporus,
dins el nucli, podent tenir efectes en la remodelació o expressió de la cromatina. Nous treballs
amb aquestes línies transgèniques poden obrir la porta a comprovar aquesta possible activitat.
Malgrat el nostre treball, i com s’ha comentat a la introducció, la demostració clara que
DRA2 estaria actuant realment com a NUP va arribar amb la publicació on DRA2 es va trobar
formant part del NPC en experiments de CoIP amb membres constituents d’aquesta estructura
(Tamura, et al. 2010).

3 Capítol 2
72
pMG56
GFP RFP
W Merge
pMS51 (~pC1302)
GFP RFP
WMerge
pBJ3
GFP RFP
WMerge
pMG55
GFP RFP
MergeW
35S GFP ATHB435S GFP
Nt35S GFP Ct35S GFP
RFP35SpDsRed RFP35SpDsRed
RFP35SpDsRed RFP35SpDsRed
3.2.2.2. Anàlisis del mutant dra2-1: Caracterització molecular.
Havíem estudiat diferents aspectes fisiològics de la línia mutant dra2-1, però encara
faltava analitzar els aspectes moleculars. Els resultats d’aquestes anàlisis indicaven que el
mutant responia menys a ombra fisiològicament; però calia saber si aquestes respostes
atenuades eren degudes a canvis moleculars en la percepció de llum o en la senyalització
posterior. Per això es van mirar nivells d’expressió de diversos gens de resposta ràpida a
ombra per anàlisis de RT-qPCR. A més, també es van quantificar els nivells d’expressió de
LUC, el gen reporter usat per l’screening, sota control del promotor de PHYB i per tant,
ràpidament activat en resposta a ombra. Primerament es va mirar la resposta molecular per
alguns gens marcadors d’ombra en tractaments d’una hora, agafant mostres abans i després
del tractament W+FR. Es a veure que gens com PIL1, HFR1 o el propi PHYB tenien una
resposta molecular atenuada en aquest mutant, amb nivells d’expressió significativament
Figura C2.4. Localització subcel·lular de les construccions per DRA2. Imatges representatives de la localització
subcel·lular vista per microbombardeig en porro i mirant al confocal per les construccions control (imatges superiors) i les de
DRA2, pMG56 i pMG55 (inferiors). GFP = senyal de la Green Fluorecent Protein; RFP = senyal de la Red Fluorecent Protein;
W = Camp clar; Merge = Superposició de les dues fluorescències. L’escala a les imatges superiors marca 100µm i a les inferior

3 Capítol 2
73
menors després del tractament en el mutant dra2-1 respecte al control wt (línia PBL).
Posteriorment ens vam plantejar si aquesta resposta atenuada era deguda a un retard en la
resposta o era una resposta consistentment més baixa al llarg del temps. Per això vam anar
més enllà i vam fer experiments de time-course, mirant les respostes a una, dues i quatre hores
de tractament W+FR. Es van analitzar diversos gens, que presenten diferents patrons
d’inducció. Com es veu a la figura C2.5, tots els gens analitzats excepte XTR7 presenten un pic
d’inducció en una hora de tractament amb W+FR en les mostres de les plàntules PBL (línia
silvestre). En les plàntules dra2-1 aquesta resposta ràpida disminueix en alguns gens (com
PHYB o PIL1), mentre que en altres sembla tenir un endarreriment en la resposta, amb pics
d’expressió a les dues hores (comportament per exemple dels gens IAA1, IAA19 i IAA29). En
altres casos (com SAUR15 i SAUR68) el mutant presenta unes respostes similars al wt (potser
una resposta lleugerament menor), però amb nivells d’expressió més alts en tots els punts. Així
doncs, l’atenuació de la resposta a ombra no és general per tots els marcadors moleculars,
sinó que afecta de manera diferent als diversos mòduls, al diferents grups de gens, de la
senyalització per llum, suggerint que la mutació dra2-1 interfereix només en alguns aspectes de
la senyalització, i que és capaç de percebre els canvis de llum correctament.
Donada la importància de les NUPs en el desenvolupament general, i més en concret
per la importància de DRA2 en les respostes a ombra, vam decidir analitzar com afectava a
l’expressió general de la planta el nostre mutant dra2-1. Per això vam analitzar el transcriptoma
de plàntules PBL i dra2-1 creixent durant 7 dies en W i agafant mostres abans (0 h) i després
d’una hora de tractament en W+FR (1 h). Es van fer les extraccions d’RNA i es van enviar al
NASC (Nottingham Arabidopsis Stock Centre, University of Nottingham, UK) per tal que fessin
l’anàlisi de qualitat dels RNA obtinguts i els chips (microarray ATH1 de Affymetrix). Els resultats
crus rebuts van ser processats per altres membres del laboratori mitjançant el programa
Rossetta Resolver (Rosetta Biosoftware, http://www.rosettabiosoftware.com) tal com s’ha
descrit per altres autors (Kaufmann, et al. 2010) i el valors P calculats es van ajustar utilitzant el
procediment Benjamini & Hochberg (BH) amb el programa R (Bioconductor multtest package
en R, http://www.bioconductor.org/packages/bioc/stable/src/contrib/html /multtest.html).

3 Capítol 2
74
Wd0 d7
+1, 2 o 4h W+FR
0h1h2h4h
Exp
ress
ióre
lati
va
PBL dra2-1
LUC
0
5
10
15
20
0
0
PHYB
PBL dra2-10,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
0
00
PIL1
0
20
40
60
80
100
120
140
PBL dra2-1
00
SAUR15
PBL dra2-10
1
2
3
4
5
6
7
8
9
10
00
0
0
IAA19
PBL dra2-10
2
4
6
8
10
12
14
16
00
XTR7
PBL dra2-10
1
2
3
4
5
6
7
8
9
10
0
IAA29
0
5
10
15
20
25
30
35
40
45
PBL dra2-1
0
0
0
PBL dra2-1
DRAL
0
5
10
15
20
25
00
DRA2
PBL dra2-10,0
0,2
0,4
0,6
0,8
1,0
1,2
00
IAA1
PBL dra2-10
2
4
6
8
10
12
0
0
0
Exp
ress
ióre
lati
vaE
xpre
ssió
rela
tiva
(a)
(b)(c)
0
1
2
3
4
5
6
7
8
9
PBL dra2-1
SAUR68
0
00
0
Aquestes anàlisis van permetre obtenir unes llistes de gens diferencialment expressats
en el mutant dra2-1 respecte al control PBL (BH<0.05 i FC=Fold Change>1,8) en W (414 gens)
i després d’una hora de W+FR (475 gens) respecte al control PBL. Estudis de classificació de
gens (GO, de l’anglès Gene Ontology) permeten veure que diverses rutes estan alterades en
ambdues condicions, especialment la ruta d’auxines, amb aproximadament una vintena de
gens de resposta a aquesta hormona afectats (aproximadament el 5% dels gens amb diferent
expressió pertanyen a la classificació de senyalització per auxines, nombre considerat
estadísticament significatiu). Tenen la seva expressió afectada gens com SAUR15, SAUR68,
IAA1, IAA2, IAA5, IAA6, IAA14, IAA19 i bastants altres SAUR-like, mostrant generalment nivells
d’expressió més alts en el mutant, resultats acordes amb les anàlisis per qPCR fetes amb
anterioritat (figura C2.5). Aquest conjunt de dades és consistent amb el fenotip de la plàntula,
que sembla tenir una senyalització per auxines constitutiva, i amb la seva resposta fisiològica a
tractaments amb auxines. Aquests resultats van ser confirmats fent anàlisis del solapament
entre els gens diferencialment expressats en el mutant en ambdós tractaments de llum (0 i 1 h
de W+FR) i llistes de gens descrits com a regulats específicament per auxines (57 gens de la
taula S13 de (Nemhauser, et al. 2006) (figura C2.6). Més enllà de la senyalització per auxines,
Figura C2.5. Efecte molecular del mutant dra2-1 sobre l’expressió de diversos gens marcadors d’ombra. (a) Esquema
indicant les condicions de creixement: es van deixar créixer les plàntules durant 7 dies i es van agafar mostres a 0h, +1, +2 i
+4h de W+FR. (b) Resultats de qPCR del time-course mirant diversos gens que responen a ombra. (c) Nivells relatius
d’expressió pel propi gen DRA2, i del seu homòleg DRAL. Es representa la mitjana dels nivells d’expressió de totes les mostres,
ja que el nivells no varien amb el tractament d’ombra. Els símbols indiquen diferències significatives (0 P<0.05; 00 P<0.01)
respecte el control wt creixent en iguals condicions.

3 Capítol 2
75
sembla que altres rutes hormonals també es veuen afectades (com les dependents d’ABA i
etilè, però semblen poc o molt poc afectades les de BRs, GAs i CKs), així com gens de
resposta a estímuls exògens tant per estrès biòtic com abiòtic. Aquests efectes encaixen amb
respostes descrites per altres mutants de NUPs, confirmant els efectes generals pleiotròpics de
mutants de components del NPC.
Específicsper auxina
dra2-1 en 0h
Específicsper auxina
dra2-1 en 1h
dra2-1 en 0h
dra2-1 en 1h
Chip-sep PIF5
Chip-sep PIF5
(a) (b)
Dins els gens de resposta a factors abiòtics s’hi inclouen gens de resposta a canvis de
llum. Mirant els canvis d’expressió només en condició d’ombra apareixen nivells d’expressió
incrementats en gens com PIF3 i SPA4 (similar a SPA1, SUPPRESSOR OF PHYA-105
MUTANT)(Rolauffs, et al. 2012). Cal comentar que PIL1 no està present en aquest microarray
d’Affymetrix, i per tant la seva falta no és deguda a una no significança dels resultats, sinó a la
manca de dades al respecte. Per tal de veure com d’afectada estava la senyalització per llum
vam fer anàlisis de solapament entre els gens alterats en el nostre mutant i dades publicades
per alguns mutants de respostes a llum, com PIF5 i PIF7. Mirant la llista de gens que podrien
estar directament regulats per PIF5 (llista de ChiP-seq amb 1218 gens, taula S1 de
(Hornitschek, et al. 2012) es va poder veure certa coincidència. A més, comparant gens
regulats diferencialment en el doble mutant pif4pif5 (taules S3 i S5 del mateix article) es podia
veure que, tot i ser anàlisis en diferents ecotips i fets en diferents laboratoris, hi havia certa
coincidència, suggerint que part del fenotip del mutant dra2-1 podria venir donat per una
senyalització defectuosa a través de PIF4 o PIF5. Es van fer anàlisis de solapament similars
amb les dades disponibles per RNA-seq en mutants de PIF7 (Li, et al. 2012), però es va veure
que el solapament era molt menor. La manca d’anàlisis de microarrays fets en les mateixes
condicions, una hora d’ombra simulada, impedeix poder comparar millor els resultats amb altres
factors de la senyalització per llum.
Mirant altres aspectes, la majoria de gens afectats en el mutant dra2-1, tant en l’anàlisi
en W com en W+FR, corresponen a proteïnes nuclears, afectant sobretot a la transcripció de
gens. El gen que més altera la seva expressió és At1g59660 (en el mutant augmenten els seus
nivells d’expressió prop de 40 vegades). Aquest gen l’hem anomenat DRA2-LIKE (DRAL), ja
Figura C2.6. Solapament de gens amb expressió afectada en dra2-1 i vies de senyalització per auxines i llum. (a)
Solapament amb la ruta d’auxines (b) Solapament amb anàlisis de PIF5.

3 Capítol 2
76
que presenta alta homologia de seqüència amb el gen DRA2. Sembla doncs que DRAL intenta
rescatar la pèrdua de DRA2, però com a mínim en l’ecotip Ws-2 no pot. També veuen
incrementats els seus nivells d’expressió dues exportines (XPO1A i B) implicades tant en
l’exportació de proteïnes com en el tràfic d’RNAs (Wu, et al. 2010), altres gens relacionats amb
la importació de proteïnes a través del NPC (com RAN1 i NTF2)(Zhao et al. 2008, Zhao et al.
2006) i l’exportació d’mRNAs (com LOS4)(Gong et al. 2005) i components del propi NPC (com
RAE, NDC1 i GLE1). A més, el propi DRA2 mostra nivells d’expressió menors en les dues
condicions. Aquestes dades donen suport a que DRA2 és realment una NUP i que la seva
mutació afecta l’expressió d’altres components del NPC, especialment els relacionats amb el
transport.
En les bases de dades disponibles (BAR, Toronto University i Genevestigator) està
descrit que cap d’aquest dos gens altera la seva expressió en ombra, si bé DRA2 és
generalment expressat en tots els teixits i estadis de desenvolupament, mentre que DRAL
només s’expressa en determinades circumstàncies, possiblement quan hi ha defectes en
transport a través del NPC. Utilitzant qPCR, vam validar que l’expressió de DRA2 i DRAL, no
canviava amb el tractament d’ombra però si entre mostres wt i dra2-1 (figura C2.5c), confirmant
que DRAL manté nivells bassals d’expressió molt baixos, però que augmenten clarament quan
hi ha algun defecte en el transport a través del NPC.
3.2.2.3. Altres NUPs: Caracterització fisiològica i molecular.
Com s’ha dit a la introducció, diferents estudis genètics en Arabidopsis han identificat
altres NUPs en mutants que mostraven alteracions en diverses rutes de senyalització de les
plantes. Fins ara, però, mai s’havien trobat mirant respostes a la llum. Era doncs interessant
mirar si els defectes en respostes a ombra eren específics per DRA2 o generals en altres
components del NPC. A més calia comprovar si altres mutants per DRA2 mostraven les
mateixes respostes que l’al·lel dra2-1 o si per algun motiu aquest al·lel presenta alguna activitat
particular. Per això es van obtenir diversos mutants d’inserció de T-DNA de la col·lecció pública
Salk per DRA2, per DRAL i per altres NUPs (figura C2.7), a més dels mutants publicats
nup160-4, nup96-1 i nup96-3, cedits pel grup del Dr. Mark Estelle (Indiana University, USA).
També es va disposar d’un mutant de la NUP58 (nup58-1) en l’ecotip Ler i dels seus quatre
al·lels en Col-0 (de nup58-2 a nup58-5), facilitats pels col·laboradors d’Elx (grup de la Dra. Mª
Rosa Ponce). Amb tots aquests mutants, comprovats per PCR i en homozigosi, es va passar a
fer-ne l’estudi fisiològic, creixent plàntules amb i sense tractament d’ombra fins a dia 7 i
mesurant-ne les longituds dels seus hipocòtils (figura C2.8a). Alguns dels al·lels es van
analitzar en la tesi anterior, com s’ha mostrat a la introducció (figura I.7d).
Els resultats van mostrar que els mutants en NUP160 i NUP96 mostraven respostes a
ombra atenuades, amb un hipocòtil més llarg en W i una menor elongació respecte al control
wt, mentre que els al·lels de DRA2 en Col-0 no presentaven cap alteració en la resposta a
ombra, ni cap fenotip apreciable, més que un hipocòtil lleugerament més llarg (es mostren 3
al·lels, però tots presentaven igual resposta). Com que els mutants d’inserció de T-DNA poden

3 Capítol 2
77
dra2-6
JO402
dra2-2
dra2-3
GO75JO403
GO74
G2340-A
(W780-STOP)
dra2-1
GO76
GO77
dra2-4 dra2-5
MGO6MGO7
GO91
GO90 GO94
Sonda de Northern
GO89GO88
dral-2
MGO9
MGO8
Sonda Northern
GO82
GO83
nup54-2nup54-1
GO81GO80MGO11
Sonda Northern
nup62-1
nup62-2
GO86
nup62-3
MGO10
GO85GO87 MGO5
DRA2/NUP98 (At1g10390)
DRAL/NUP98like (At1g59660)
NUP54 (At1g24310)
NUP62 (At2g45000)
GO78
nup58-3
GO79
nup58-4
nup58-2
nup58-5
Sonda Northern
NUP58/TCU1 (At4g37130)
C184-T
(Q184-STOP)
nup58-1
presentar diverses insercions es van realitzar alguns retrocreuaments en Col-0 per comprovar
que les línies mostraven el mateix fenotip després del creuament amb el seu propi ecotip. Tal
com era d’esperar el resultat dels creuaments (anomenats CMG5-7) va confirmar que els
mutants per DRA2 en Col-0 no tenien altres mutacions addicionals i que mantenien la mateixa
resposta després del creuament (resultats no mostrats). Els mutants en Col-0 per la NUP54 i
per la NUP58 tampoc mostraven cap alteració en la resposta a ombra, si bé semblen tenir
floracions primerenques. Els mutants en la NUP62 presentaven comportaments diferents:
mentre l’al·lel nup62-1 era inviable i la seva homozigosi donava lloc a l’avort de la llavor, un
segon al·lel (nup62-3) responia lleugerament menys a ombra, suggerint que podria
correspondre a una reducció de la funció; plantes mutants homozigotes d’un tercer al·lel
(nup62-2) eren estèrils i presentaven fenotips molt forts, amb problemes de desenvolupament
(figura C2.8b). Sembrant llavors en heterozigosi i seleccionant les plàntules mutants per fenotip
(una quarta part de la població) es va poder veure que presentaven una resposta a ombra
disminuïda, mentre que les seves germanes heterozigotes, amb aspecte wt, responien de
manera normal al tractament. A més, el genotip d’aquestes plantes es va confirmar per PCR
Figura C2.7. Mapes
genètics per diferents
NUPs. Mapes dels diferents
gens que codifiquen per
NUPs. En blanc s’indiquen els
introns i en gris els exons.
Les línies mutants estant
indicades amb triangles. Els
oligonucleòtids usats pels
genotipats i per fer les sondes
de Northern estan marcats
amb les fletxes i el nom
corresponen.

3 Capítol 2
78
utilitzant oligos específics. En resum, amb aquestes anàlisis vam veure que teníem uns
mutants de NUPs amb fenotips adults forts pleiotròpics, i respostes de l’hipocòtil a ombra
atenuades (dra2-1, nup160-4, nup96-1, nup62-2 i nup58-1), i la resta de mutants amb fenotips
molt febles i respostes a ombra pràcticament normals. A més, al·lels de mutants per NUPs en
Col-0 semblen tenir fenotips molt més febles que no pas al·lels en altres ecotips (com són dra2-
1 en Ws.2 i nup58-1 en Ler).
PBL dra2-1
Lo
ng
itu
d h
ipo
còti
l(m
m)
0
1
2
3
4
5
6
Col-0 dra2-3dra2-2 dra2-40
1
2
3
4
5
6
7
8
0
1
2
3
4
5
6
7
8
Col-0 dral-2
0
1
2
3
4
5
6
7
Col-0 nup96-1 nup96-30
1
2
3
4
5
6
7
Col-0 nup62-2Hmz
nup62-2Htz / wt
Lo
ng
itu
d h
ipo
còti
l(m
m)
0
1
2
3
4
5
6
7
8
Col-0 nup54-1 nup54-2
00
0
00 00 00
0
00
0
00
0000
00
00
00 00
00
Col-0 nup62-2 Homozigota nup62-2 Heterozigota
(c)
(a)
Col-0 nup62-2 Homozigota
0
1
2
3
4
5
6
7
Col-0 nup62-3
00
W
W+FR
(b)
W
W+FR
d0 d2 d7
Per tal de veure si els diferents al·lels eren o no mutants de pèrdua de funció es va
considerar el lloc d’inserció del T-DNA i es van analitzar els seus nivells d’expressió per anàlisis
de Northern-blot (figura C2.9). En aquest tipus d’anàlisis es pot veure no només els nivells
d’expressió sinó també el tamany aproximat del transcrit, fet que ens pot deixar veure si podria
representar també una pèrdua de funció per truncació del producte gènic. En el cas de DRA2,
només la línia mutant dra2-4 es va confirmar con a pèrdua de funció clara per la manca de
trànscrit. El fet, però, que la resta d’al·lels en aquest gen presentaven insercions que
interrompien l’ORF i mostraven fenotips similars, fa pensar que tots ells són realment de pèrdua
de funció, si bé no es pot descartar que algunes línies només corresponguin a una reducció de
funció. Per DRAL, no es va aconseguir veure senyal, ja que els nivells d’expressió d’aquest gen
són molt baixos; no obstant, una quantificació més sensible feta amb qPCR va permetre veure
Figura C2.8. Respostes fisiològiques de les línies mutants de NUPs en tractament d’ombra simulada. (a) Esquema
indicant les condicions de creixement: les plantes van ser crescudes 2 dies en W i un grup es va transferir a W+FR a partir del
segon dia i fins a dia 7. (b) Mesures de longitud d’hipocòtil de les diferents línies mutants. El símbols indiquen diferències
significatives ( 0=P<0.05; 00P<0.01) respecte el control wt creixent en igual condició. (c) Aspecte de plàntules de 7 dies i plantes
adultes, de 5 setmanes, en homozigosi i heterozigosi per l’al·lel nup62-2.

3 Capítol 2
79
que els nivells d’mRNA no semblen alterats en el seu mutant (resultats no mostrats). El fet que
la línia mutant obtinguda per DRAL tingui la inserció molt a l’extrem Ct, fora de la regió
codificant, fa pensar que probablement aquesta línia no és realment una pèrdua de funció. Pel
que fa als mutants en NUP58, cap mutant presentava una pèrdua clara d’expressió, però els
tamanys diferents en els trànscrits de la NUP58 fa pensar que són productes aberrants, amb
només una part codificant per la pròpia NUP58 i que podrien no donar lloc a proteïna o fer-ho a
una no funcional. Per la NUP54, el fet que els dos al·lels tinguin les insercions en l’ORF i
presentin nivells d’expressió incrementats porta a pensar que hi ha una falta d’activitat proteica
que s’intenta compensar expressant més aquest gen, però que no es veu compensat perquè
probablement l’mRNA no doni lloc a una proteïna funcional.
dra
2-1
PB
L
Co
l-0
dra
2-2
dra
2-4
dra
2-5
dra
2-3
DRA2
25S
NUP58
25S
NUP54
25S
Co
-0
nu
p54
-2
nu
p54
-1
Co
l-0
nu
p58
-2
nu
p58
-3
nu
p58
-4
Ler
nu
p58
-1
Wd0 d7
Més enllà de les respostes fisiològiques, teníem les respostes moleculars de dra2-1 ben
caracteritzades. Vam passar a analitzar les respostes moleculars en tractaments d’una hora en
ombra simulada de diferents mutants, amb la resposta fisiològica a ombra alterada (figura
C2.10) i sense alterar (figura C2.11). Els resultats van mostrar que l’únic mutant que té
respostes moleculars a ombra clarament afectades és el dra2-1, mentre que la resta, ja sigui
pels mutants amb fenotips forts o pels febles, mostren canvis moleculars subtils i moltes
vegades no significativament diferents. Cal comentar que els mutants en NUP96 i NUP160
semblen tenir afectada la resposta de gens com IAA19 i IAA29, però al ser gens regulats també
per auxines, és difícil saber si aquests efectes són fruit d’una alteració en les respostes a
ombra o en les respostes a auxines, ja que està descrit que aquests mutants tenen la
senyalització per auxines alterada. Es pot concloure doncs que DRA2 té algun paper específic
en la senyalització molecular d’ombra. No obstant la contribució de l’ecotip ha de ser molt
significativa, ja que l’al·lel de DRA2 en Col-0 (dra2-4), malgrat tenir una tendència similar a
dra2-1, no mostra diferències significatives en les seves respostes moleculars.
També vam mirar l’efecte de les mutacions en diferents NUPs en els nivells d’expressió
de DRAL. En aquest cas, l’increment d’expressió d’aquest gen era significatiu en tots els
mutants, fins i tot en els més dèbils (com dra2-4, nup58-2 i nup96-3) però el canvi era encara
més notable en aquells al·lels considerats forts, amb un fenotip més marcat. Això ens va portar
a confirmar que, tal com pensàvem, DRAL està incrementat quan hi ha algun defecte en el
transport a través del NPC, podent-lo fer servir com a marcador d’afectació del transport
nuclear.
Figura C2.9. Anàlisis d’expressió dels diferents al·lels mutants per DRA2, NUP58 i NUP54. Autoradiografies mostrant els
nivells d’expressió per Northern-blot de diferents línies mutants de NUPs. Per simplificar es mostra una de les dues rèpliques
biològiques. La part superior mostra l’autoradiografia de la membrana hibridada amb amb una sonda per una regió específica de
cada trànscrit, mentre la inferior mostra el control de càrrega hibridant amb una sonda constitutiva 25S.

3 Capítol 2
80
PBL nup160-4 nup96-10
20
40
60
80
100
120
140
160
180PIL1
PBL nup160-4 nup96-1
IAA19
0
5
10
15
20
25
PBL nup160-4 nup96-1
IAA29
0
20
40
60
80
100
120
140
160
180
200
PBL nup160-4 nup96-1
DRAL
0
2
4
6
8
10
12
14
16
Exp
ress
ióre
lati
va
0h+1h
(a)
HFR1
Ler nup58-10
5
10
15
20
25
30
35
DRAL
Ler nup58-10123456789
00
Wd0 d7
PIL1
Ler nup58-10
50
100
150
200
250
300
0
Exp
ress
ióre
lati
va
0
00
00
0
(b)
(c)
(d)
0h +1h W+FR
(d)PIL1
Col-0 dra2-40
10
20
30
40
50
60
Exp
ress
ióre
lati
va
HFR1
0
5
10
15
20
25
Col-0 dra2-4
HFR1
Col-0 nup96-30
5
10
15
20
25
30
35
40
nup58-2
DRAL
Col-0 dra2-4
0,20,40,60,81,01,21,41,61,82,0
0,0
0DRA2
Col-0 dra2-40,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
0
DRAL
Col-0 nup96-30,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
nup58-2
00
00PIL1
Col-0 nup96-30
50
100
150
200
250
nup58-2
Exp
ress
ióre
lati
va
0h+1h
(a) Wd0 d7
0h +1h W+FR(b)
(c)(e)
3.2.2.4. Creuaments per canvis d’ecotip.
Veient la importància de l’ecotip en les respostes tant fisiològiques com moleculars es va
passar a realitzar una sèrie de creuaments per canviar d’ecotip els mutants de DRA2, tant pels
Figura C2.10. Anàlisis de les respostes molecular a ombra simulada per mutants de NUPs amb un fenotip fort. (a) Esquema
indicant les condicions de creixement: les plantes van ser crescudes durant 7 dies i es van agafar mostres abans (0h) i després
(+1h) de tractament amb W+FR. (b) i (c) Anàlisis d’expressió de diferents marcadors d’ombra per qPCR en els mutants amb fenotip
fort (en (b) per NUP160 i NUP96 i en (c) per NUP58). (d) Nivells d’expressió de DRAL en les mostres crescudes en W. Els símbols
indiquen diferències significatives ( 0P<0.05; 00P<0.01) respecte el control wt creixent en iguals condicions.
Figura C2.11. Anàlisis de les respostes moleculars a ombra simulada per mutants de NUPs amb un fenotip feble o sense
fenotip. (a) Esquema indicant les condicions de creixement: les plantes van ser crescudes durant 7 dies i es van agafar mostres
abans (0h) i després (+1h) de tractament amb W+FR. (b) i (c) Anàlisis d’expressió de gens marcadors d’ombra per qPCR en
diferents mutants de NUPs amb fenotips febles. (d) i (e) Nivells relatius d’expressió pel gen DRA2 i/o pel seu homòleg DRAL en els
mutants anteriors. Els símbols indiquen diferències significatives ( 0P<0.05; 00 P<0.01) respecte el control wt creixent en iguals

3 Capítol 2
81
al·lels de Col-0 cap a PBL (Ws-2) com el mutant dra2-1, en PBL, cap Col-0. Es van fer fins a 4
creuaments per les línies d’inserció de T-DNA (pels al·lels dra2-3 i dra2-4) cap a l’ecotip Ws-2 i
per passar el mutant dra2-1 cap a Col-0, tot i que està descrit que es necessiten fins a 6
creuaments per considerar que la mutació seleccionada es troba en un entorn amb l’ecotip
completament canviat.
0
1
2
3
4
5
6
7
Col-0 dra2-1PBL dra2-1BC3.a5
CMG34a5
0
1
2
3
4
5
6
Col-0 a5 a11 a150
1
2
3
4
5
6
Col-0 dra2-4 PBL dra2-1BC4
CMG50
Lo
ng
itu
d h
ipo
còti
l(m
m)
00
00
00
00
00
00
000
00
W
W+FR
W
W+FR
d0 d2 d7
(b)
(c)
dra2-1BC3 (CMG34)
X ♂Col-0♀dra2-1
dra2-1BC3 (CMG34)
X♀Col-0
X♀Col-0 dra2-1BC2 (CAG9)
dra2-1BC1
X♀Col-0
dra2-1BC4 (CMG50)
(a)
Com ja s’ha comentat a la introducció, tan sols un creuament del mutant original dra2-1
en Col-0 és suficient perquè aquest perdi gran part del seu fenotip, podent obtenir plantes
homozigotes dra2-1 amb hipocòtils i cotiledons molt semblants als de Col-0. Al retrocreuar dues
vegades més en Col-0, fins al que seria el tercer creuament, es van obtenir diverses línies
mutants dra2-1 (amb gran part del seu genoma pertanyen a l’ecotip Col-0). Els experiments
fisiològics amb aquestes línies mostren que encara hi ha una influència clara del fons genètic
(figura C2.12), presentant diferències clares de comportament. Mentre que la línia dra2-
1BC3.a5 (Backcross with Col-0 number 3.a5, també anomenada CMG34a5) es comporta de
manera semblant a l’al·lel dra2-4, plantes germanes dra2-1BC3.a11 i .a15 (CMG34a11 i a15)
encara retenen una resposta atenuada a ombra, amb un hipocòtil lleugerament més llarg en W.
Això porta a pensar que hi ha almenys dos factors implicats en el canvi de fenotip (a més de
DRA2), i mentre que un sembla fàcil d’eliminar (amb un sol creuament es pot perdre) un altre
component és molt més difícil (en 3 creuaments, dues línies encara el mantenen),
probablement perquè es trobi proper a la mutació dra2-1. El quart creuament amb Col-0, dra2-
1BC4 (CMG50, fet amb la línia CMG34a5) mostra la mateixa tendència que la planta pare
original, demostrant que encara que no s’hagi canviat completament el fons genètic Ws-2, si
que clarament el fenotip del mutant original dra2-1 ha estat perdut.
Figura C2.12. Respostes fisiològiques a ombra pels creuaments amb dra2-1 amb el canvi d’ecotip. (a) Representació dels
creuaments realitzats amb Col-0 i dra2-1 (ecotip Ws-2) per canviar el seu ecotip. (b) Esquema indicant les condicions de
creixement: les plantes van ser crescudes 2 dies en W i un grup es va transferir a W+FR a partir del segon dia i fins a dia 7. (c)
Mesures de longitud d’hipocòtil del mutant dra2-1 i els creuaments indicats. Els símbols indiquen diferències significatives (0
P<0.05; 00 P<0.01) respecte el control wt (Col-0) creixent en iguals condicions de llum.

3 Capítol 2
82
X ♂dra2-4♀PBL
dra2-4BP3 (CMG49)
X♀PBL
X♀PBL dra2-4BP2 (CMG31)
dra2-4BP1 (CMG10)
X♀PBL
dra2-4BP4 (CMG56)
W
W+FR
d0 d2 d7
W
W+FR(b)
(c)
(a)
0
1
2
3
4
5
6
7
8
Col-0 PBL dra2-4 dra2-1 dra2-3BP2 CMG30
dra2-4BP2 CMG31
Lo
ng
itu
d h
ipo
còti
l(m
m)
00
00
0000
00 00
0
1
2
3
4
5
dra2-1PBL a2 b1 a2 b1 0
1
2
3
4
5
6
Col-0 dra2-4
Lo
ng
itu
d h
ipo
còti
l(m
m)
00
00
00
0000
00
dra2-3BP3 (CMG48) dra2-4BP3 (CMG49) dra2-4BP3 (CMG49)
a2 b1
Per últim, vam tornar a creuar una planta dra2-1BC3 (línia CMG34a5, d’aspecte
semblant a dra2-4) amb l’ecotip Ws-2 (creuament anomenat CMG55). Un sol retrocreuament va
ser suficient per recuperar el fenotip fort de la mutació original en una proporció molt semblant
a 1/16 (unes 5 plàntules sobre 90, anàlisis fenotípic fet per triplicat i analitzat per test Χ2). Fet
que demostra que no només depèn de dra2-1 sinó com a mínim d’un factor més, propi de
l’ecotip Ws-2. Està descrit que l’ecotip Ws-2 té una mutació en el PHYD (Aukerman et al. 1997),
fet que podria ser significatiu per les respostes a llum. No obstant, es va veure (mitjançant
genotipats per PCR amb encebadors específics pel PHYD de Col-0 i de Ws-2) que les
diferències de fenotips entre dra2-1 i els seus creuaments en Col-0 eren independents del
PHYD mutat en Ws-2 i que es podia tenir la mutació dra2-1 en qualsevol fons genètic PHYD
mantenint les seves respostes i el seu fenotip.
Per tal de veure si més enllà de les respostes fisiològiques els canvis d’ecotip podien
tenir algun efecte es van fer anàlisis de qPCR mirant marcadors d’ombra en una hora de
tractament per la línia dra2-1BC3.a5 (CMG34a5), la més semblant fenotípicament a dra2-4. Els
experiments moleculars van permetre veure que malgrat la manca de fenotip, l’efecte molecular
era més fort pel creuament que per l’al·lel original de Col-0 (figura C2.14). Queda per
determinar si aquest efecte és degut a les restes d’ecotip Ws-2, o si pel contrari l’al·lel dra2-1 té
un efecte molecular específic que no tenen la resta d’al·lels.
Una altra hipòtesi podria explicar aquestes diferències, ja que seria possible que una
altra mutació puntual s’hagués produït durant la mutagènesis aleatòria amb EMS, de manera
que en realitat estaríem davant un doble mutant o que el propi transgen de la línia PBL tingués
algun efecte no caracteritzat prèviament. Per descartar aquests fets es va retrocreuar el mutant
Figura C2.13. Respostes fisiològiques a tractaments d’ombra pels creuaments amb el canvi d’ecotip. (a) Representació dels
creuaments realitzats amb PBL (ecotip Ws-2) i dra2-4 (ecotip Col-0) per canviar el seu ecotip. (b) Esquema indicant les condicions
de creixement: les plantes van ser crescudes 2 dies en W i un grup es va transferir a W+FR a partir del segon dia i fins a dia 7. (c)
Mesures de longitud d’hipocòtil de dues línies mutants DRA2 en Col-0 (dra2-3 i dra2-4). Els símbols indiquen les diferències
significatives (0P<0.05; 00P<0.01) respecte el control wt creixent en iguals condicions.

3 Capítol 2
83
dra2-1 en l’ecotip wt original Ws-2 (creuament CMG51), observant que només les plantes
homozigotes per dra2-1 tenien fenotip i que aquest era exactament igual al mutant original.
Això portava a pensar que ni el transgen de la línia PBL ni cap altra mutació aleatòria estan
implicats en els canvis fenotípics (fisiològics o moleculars) observats en dra2-1, reforçant que
altres components propis de l’ecotip són el rellevants per les diferències observades.
0
5
10
15
20
25
30
Col-0 dra2-1BC3.a5 CMG34a5
dra2-4
HFR1
Exp
ress
ióre
lati
va
0
2
4
6
8
10
12IAA19
Col-0 dra2-1BC3.a5 CMG34a5
dra2-40
1
2
3
4
5
6
7
8
9
10DRAL
Col-0 dra2-1BC3.a5 CMG34a5
dra2-4
0h+1h
00
00
0
0
00
0
Wd0 d7
0h +1h W+FR
(b)
(a)
D’altra banda es van fer els retrocreuaments recíprocs, amb els al·lels dra2-3 i dra2-4,
originals de Col-0, en PBL. Ni els segons creuaments dra2-3BP2 i dra-4BP2 (dra2-3 Backcross
with PBL number 2, també anomenats CMG30-31) ni els tercers creuaments dra2-3BP3 i dra-
4BP3 (també anomenats CMG48-49) mostren una atenuació clara de la resposta a ombra
comparant amb la línia PBL, i tampoc presenten un fenotip fort similar a dra2-1 (figura C2.13).
Aquest fet confirma la hipòtesi que el fenotip fort de dra2-1 és multifactorial.
Tot aquest conjunt de dades ens porta a pensar que el fenotip fort de dra2-1 és degut a
la pèrdua de DRA2, però també de almenys dos components genètics més de l’ecotip Ws-2, un
component llunyà al propi gen (fàcil de perdre per creuaments) i un component que deu estar
lligat a la mutació en DRA2. Per aquest motiu és difícil treure conclusions dels tests d’al·lelisme
fets entre dra2-1 i els seus altres al·lels de Col-0 (test entre ecotips diferents) i dels tests entre
dra-4BP3 (CMG49) i dra2-1 (creuament CMG57, tests d’al·lelisme entre ecotips Ws-2), aquest
últim amb un al·lel que no està completament en l’ecotip Ws-2 i al que podria mancar-li un
component lligat a la regió pròxima de DRA2 per mostrar el fenotip fort de dra2-1.
3.2.2.5. Creuaments entre NUPs: Dobles mutants.
Posant totes les dades anteriors en conjunt, havíem vist (1) que l’ecotip tenia una
influència sobre l’efecte de les mutacions al NPC, (2) que la intensitat de fenotips morfològics
es corresponia amb la magnitud d’afectació en els canvis d’expressió de DRAL, i (3) que
diverses mutacions en una NUP resultaren en fenotips febles (en Col-0). D’aquesta manera,
semblava lògic pensar que els fenotips forts eren deguts a un major defecte en el NPC i que
dobles mutants en NUPs podrien mostrar fenotips més forts que els simples mutants, resultant
semblants a dra2-1. Per això es va passar a obtenir una sèrie de dobles mutants a partir
Figura C2.14. Anàlisis molecular de respostes a ombra pel creuament CMG34, l’al·lel dra2-1 en fons genètic Col-0. (a)
Esquema indicant les condicions de creixement: les plantes van ser crescudes durant 7 dies i es van agafar mostres abans (0h) i
després (+1h) de tractament amb W+FR. (b) Anàlisis d’expressió de gens marcadors d’ombra pel mutant dra2-1BC3 (CMG34). A la
dreta nivells d’expressió de DRAL en les mostres en W. Els símbols indiquen diferències significatives (0P<0.05; 00P<0.01) respecte
el control wt creixent en iguals condicions.

3 Capítol 2
84
d’al·lels amb fenotips febles per creuaments entre ells. Alguns d’aquests creuaments es van fer
i seleccionar al nostre laboratori [nup58-2dra2-3 (CMG25); nup58-2dra2-4 (CMG20); i nup58-
2dra2-5 (CMG21)] i altres es van obtenir cedits pels nostres col·laboradors d’Elx [nup58-2
nup96-3 (CNE1); i nup58-2nup54-2 (CNE4)]. Amb aquests dobles mutants en NUPs es van fer
experiments fisiològics per avaluar l’allargament de l’hipocòtil en resposta a ombra simulada
(figura C2.15). En tots el casos l’hipocòtil del doble mutant en llum W era almenys tan llarg com
el del mutant parental més llarg (nup58-2). En ombra simulada, els hipocòtils del doble mutant
eren més curts que els de les plàntules Col-0, excepte els del doble mutant nup58-2nup54-2,
que eren tan llargs com els simples nup58-2. Per tant, com a mínim tres dels 4 dobles mutants
generats presenten menors respostes a ombra. A més, excepte el doble mutant nup58-2nup54-
2, presenten fenotips en planta adulta més forts que els simples mutants, amb alteracions de
desenvolupament similars al mutant dra2-1. Aquests resultats suporten la hipòtesi de partida de
que els al·lels forts tenen defectes majors en el NPC, mentre que els al·lels febles presenten un
NPC amb activitat pràcticament normal. A més, aquests resultats suggereixen que les
diferències entre els ecotips són probablement degudes a canvis en la composició del NPC,
presentant l’ecotip Col-0 una major flexibilitat o capacitat d’adaptació al perdre l’activitat de
NUPs concretes.
Lo
ng
itu
d h
ipo
còti
l(m
m)
0
1
2
3
4
5
6
7
8
Col-0 nup58-2 nup58-2nup54-2 CNE4
nup54-2
Lo
ng
itu
d h
ipo
còti
l(m
m)
0
1
2
3
4
5
6
7
8
Col-0 nup58-2 nup58-2nup96-3CNE1
nup96-3
00
0000
00
0
0
00
00
0000
0
0
1
2
3
4
5
6
7
8
Col-0 nup58-2 nup58-2dra2-5 CMG21
dra2-5
00
00
00
00 0
W
W+FR
d0 d2 d7
(b)
(a) W
W+FR
Col-0 nup58-2 nup58-2dra2-3CMG25
dra2-2 nup58-2dra2-4 CMG20
dra2-40
1
2
3
4
5
6
7
8
9
0000
00 00 00
00
00
00
Els resultats fisiològics mostraven similitud de resposta entre un doble mutant dra2-4
nup58-2 (CMG20) en Col-0 i el simple mutant dra2-1 en Ws-2. Faltava saber si les respostes
moleculars també es veien afectades en el mateix sentit. Per això es van fer anàlisis per qPCR
de l’expressió de PIL1 i IAA19, gens marcadors del tractament amb ombra simulada, i DRAL,
gen marcador de l’activitat del NPC, similars als descrits anteriorment (figura C2.16). Es va
veure que en les plàntules dobles mutants dra2-4nup58-2 (CMG20), els marcadors moleculars
Figura C2.15. Respostes fisiològiques a tractaments d’ombra simulada pels creuaments amb dobles mutants de NUPs. (a)
Esquema indicant les condicions de creixement: les plantes van ser crescudes 2 dies en W i un grup es va transferir a W+FR a
partir del segon dia i fins a dia 7. (b) Mesures de longitud d’hipocòtil de les diferents línies dobles mutants per NUPs. Els símbols
indiquen diferències significatives (0P<0.05; 00P<0.01) respecte el control wt creixent en iguals condicions.

3 Capítol 2
85
d’ombra analitzats no es veien alterats significativament, mentre que l’expressió de DRAL
estava més fortament induïda que en els mutants simples. Aquestes diferències, conjuntament
amb els experiments fisiològics anteriors, suggereixen que l’efecte del NPC en el
desenvolupament de la planta (visualitzat pels canvis en l’expressió de DRAL i la resposta de
l’hipocòtil a ombra simulada) estan deslligats del defectes en l’activitat de DRA2 en la regulació
de l’expressió gènica en respostes a ombra (visualitzats pels canvis en l’expressió d’alguns
marcadors moleculars induïts per ombra). Els efectes en l’expressió poden ser degudes a una
activitat anòmala de l’al·lel dra2-1 o al simple fet que un altre component del NPC, que no és la
NUP58, està afectat en l’ecotip Ws-2 respecte el Col-0.
0
2
4
6
8
10
12
14IAA19
Col-0 nup58-2 nup58-2dra2-4CMG20
dra2-40
10
20
30
40
50
60
70
80
90
100PIL1
Col-0 nup58-2 nup58-2dra2-4 CMG20
dra2-4
DRAL
0
2
4
6
8
10
12
14
16
Col-0 nup58-2 nup58-2dra2-4 CMG20
dra2-4
Exp
ress
ióre
lati
va
0h+1h
00 00
00
00
00
Wd0 d7
0h +1h W+FR(b)
(a)
3.2.2.6. Efectes en el transport nuclear.
Veient que la NUP98 d’animals s’ha descrit com una proteïna amb moltes funcions
diferents, resulta molt difícil saber la causa molecular de la deficient senyalització per ombra en
dra2-1. En una primera instància es va pensar que la mutació en DRA2 podria estar afectant el
transport de proteïnes dins al nucli, i més concretament la importació de phyB. Per això es va
fer generar el creuament CMG1, amb un al·lel mutant de DRA2 (dra2-3) i un mutant deficient
en phyB (al·lel phyB-9)(Reed, et al. 1994). Les anàlisis de resposta a ombra simulada mostrava
un fenotip fisiològic additiu pel doble mutant dra2-3phyB-9 (figura C2.17), fet que suggeria que
l’ importació de phyB no era el factor que estava afectat en el mutant. A més, les alteracions en
l’expressió gènica de la mutació dra2-1 segons les dades de microarray i de qPCR no semblen
encaixar amb les respostes esperades per un mutant deficient en la senyalització de phyB.
0
1
2
3
4
5
6
7
8
9
10
Col-0 dra2-3 dra2-3 phyB-9(CMG1)
phyB-9
Lo
ng
itu
d h
ipo
còti
l(m
m)W
W+FR
00
000
00 00
00W
W+FR
d0 d2 d7
Figura C2.16. Anàlisis molecular de l’expressió de gens marcadors d’ombra en el doble mutant nup58-2dra2-4. (a)
Esquema indicant les condicions de creixement: les plantes van ser crescudes durant 7 dies i es van agafar mostres abans (0h) i
després (+1h) de tractament amb W+FR. (b) Nivells d’expressió de gens marcadors d’ombra pel doble mutant nup58-2dra2-4
(CMG20). A la dreta nivells d’expressió de DRAL en les mostres en W. Els símbols indiquen diferències significatives (0P<0.05; 00P<0.01) respecte el control wt creixent en iguals condicions.
Figura C2.17. Anàlisis fisiològic de resposta a ombra per mirar la
interacció de DRA2 en la senyalització de PHYB. Mesures de la longitud
de l’hipocòtil del doble mutant dra2-3phyB-9, creixent 7 dies amb o sense
tractament W+FR. Els símbols indiquen diferències significatives (0P<0.05; 00P<0.01) respecte el control wt creixent en iguals condicions.

3 Capítol 2
86
Així doncs, podria ser que fos la importació d’altres proteïnes la que estigués afectada en
el mutant dra2-1. Està descrit que els mutants nup160-4 i nup96-1 podrien estar alterant la
importació de proteïnes relacionades amb la senyalització d’auxines (Parry, et al. 2006). Per tal
de veure si aquest mecanisme podria estar relacionat amb les respostes atenuades en ombra
es van generar línies transgèniques 35S:ATHB4-GR (utilitzades al capítol I) amb el nucleoporus
afectat, fent creuaments de la línia pCS19 amb el mutant nup96-1 (creuament CAG19). La línia
pCS19 acumula la proteïna ATHB4 al citosol, de manera que només quan s’hi afegeix DEX
aquesta es pot translocar al nucli. Es va pensar que si la importació de proteïnes estigués
alterada per un defecte en una NUP, la translocació al nucli de la proteïna ATHB4-GR, després
de l’addició al medi de DEX, es veuria ralentitzada i, per tant, l’expressió de gens marcadors
d’activitat d’aquesta proteïna es veuria també alentida en comparació amb la línia control
(pCS19 en fons silvestre), amb el transport nuclear no afectat. Tal com hem vist al capítol I
HAT2 i SAUR15 són bons gens marcadors d’activitat d’ATHB4, essent els seus nivells
d’expressió fortament reprimits per la translocació d’aquest factor de transcripció al nucli. En
primera instància es va provar de mirar l’expressió d’aquests gens marcadors després de
tractaments amb diferents concentracions de DEX durant 4 h (figura C2.18a). Els resultats
mostren poques diferències entre les línies transgèniques comparades, si bé sembla que la
tendència seria que la mutació nup96-1 (línia CA19) respongués lleugerament menys a les
concentracions més baixes (figura C2.18b). Es van fer anàlisis de dues cues ANOVA (amb les
dades de HAT2 i SAUR15) mostrant que els dos genotips responen diferentment al tractament.
Per tal de veure més fàcilment les diferències en la caiguda d’expressió entre les dues línies es
mostren els resultats normalitzats respecte al nivell inicial d’expressió del propi genotip (figura
C2.18c).
Wd0 d7 W -DEX
W +0,005 µM DEXW +0,05 µM DEXW +0,5 µM DEXW +5 µM DEX
wt nup96-1 (CA19)
SAUR15
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8HAT2
wt nup96-1 (CA19)0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
wt nup96-1 (CA19)0,0
0,2
0,4
0,6
0,8
1,0
1,2SAUR15
nup96-1 (CA19)wt
HAT2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Exp
ress
ióre
lati
vaE
xpre
ssió
rela
tiva
0 0
0
0
0h +4hDEX
35S:ATHB4-GR (pCS19) 35S:ATHB4-GR (pCS19)
35S:ATHB4-GR (pCS19)35S:ATHB4-GR (pCS19)
(a)
(b)
(c)
(d)
0,0
0,2
0,4
0,6
0,8
1,0
1,2GR
nup96-1 (CA19)wt
35S:ATHB4-GR (pCS19)
Figura C2.18. Anàlisis molecular d’expressió de gens marcadors de l’activitat biològica d’ATHB4 per veure l’afectació en el
tràfic nuclear. (a) Esquema indicant les condicions de creixement: es van deixar créixer les plàntules durant 7 dies i es van agafar
mostres a 0h i +4h de +DEX en diferents concentracions. (b) Nivells d’expressió dels gens marcadors pel doble mutant CA19
normalitzats respecte el control pCS19 sense tractament. (c) Nivells d’expressió de gens marcadors del doble mutant CA19
normalitzats respecte la pròpia línia sense tractament. (d) Nivell d’expressió del transgen per les dues línies. Els símbols indiquen
diferències significatives (0P<0.05; 00P<0.01) respecte el control pCS19 creixent en iguals condicions.

3 Capítol 2
87
Diferents nivells d’expressió del transgen entre les dues línies podrien explicar
diferències de resposta. Per això també es van quantificar els nivells de GR, que van resultar
ser semblants, descartant aquest efecte (figura C2.18d). Així doncs, tot i una tendència a
respondre menys en el fons nup96-1 (línia CA19), els resultats no eren prou sòlids per poder-
ho afirmar. Per això es va provar un experiment més restrictiu, usant la concentració més baixa
possible que dóna lloc a una caiguda forta dels nivells d’expressió dels gens marcadors (es va
usar 0.1µM DEX), i mirant el seu efecte a diferents temps (el que seria un time-course) (figura
C2.19) De manera similar, sembla que es manté la tendència a que la línia amb la mutació en
la NUP96 respongui lleugerament més lentament, però sense arribar a ser un resultat prou
robust. Així doncs, els nostres resultats suggereixen que NUP96 afecta lleugerament la
importació de proteïnes relacionades amb la regulació de la SAS.
Wd0 d7
0h+1h+2h+4h
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
wt nup96-1 (CA19)
SAUR15
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
wt nup96-1 (CA19)
HAT2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
wt nup96-1 (CA19)
SAUR15
0,0
0,2
0,4
0,6
0,8
1,0
1,2
wt nup96-1 (CA19)
HAT2
Exp
ress
ióre
lati
vaE
xpre
ssió
rela
tiva
0
0
00
0
00
00
00
0, +1, 2, 4h +0,1 µM DEX
35S:ATHB4-GR (pCS19) 35S:ATHB4-GR (pCS19)
35S:ATHB4-GR (pCS19) 35S:ATHB4-GR (pCS19)
(a)
(b)
(c)
A continuació, vam fer servir una aproximació com l’anteriorment explicada per analitzar
la importació de proteïnes però en el mutant dra2-1, per tal d’adreçar si DRA2 afectava
específicament l’expressió de proteïnes relacionades amb la regulació de la SAS. Per això es
van generar línies transgèniques equivalents a la pCS19 (35S:ATHB4-GR), però en l’ecotip
Ws-2 (línies pMG49). Es va comprovar que la inducció amb DEX era correcta i que la sobre-
expressió d’ATHB4 activa provocava el mateix efecte fisiològic en l’ecotip Ws-2 que en Col-0
(produint plàntules nanes, més fosques i una resposta a ombra clarament atenuada). Aquesta
línia es va creuar amb el mutant dra2-1 (creuament CMG59). Les línies CMG59 tenien un
Figura C2.19. Anàlisis molecular d’expressió de gens marcadors de l’activitat biològica d’ATHB4 per veure l’afectació en el
tràfic nuclear. (a) Esquema indicant les condicions de creixement: es van deixar créixer les plàntules durant 7 dies i es van agafar
mostres a 0, 1, 2 i +4h de +DEX a 0,1 µM. (b) Nivells d’expressió per qPCR del doble mutant CA19 normalitzats respecte el control
pCS19 sense tractament. (c) Nivells d’expressió per qPCR del doble mutant CA19 normalitzats respecte a la pròpia línia sense
tractament. Els símbols indiquen diferències significatives (0P<0.05; 00P<0.01) respecte el control pCS19 creixent en iguals
condicions.

3 Capítol 2
88
aspecte idèntic a les dra2-1 (en absència de DEX). Així es va passar a fer un experiment
equivalent a l’anterior, en una concentració baixa de DEX (0.1µM DEX) i prenent mostres en
diferents temps de tractament. De manera similar als resultats de nup96-1 en Col-0, no es va
poder veure cap diferència significativa entre la caiguda d’expressió dels gens marcadors en la
línia MG49 i el creuament CMG59, sent bàsicament resultats no concloents (resultats no
mostrats). Aquests resultats ens indiquen que dues NUPs amb un efecte important en la
regulació de les respostes de l’hipocòtil a la proximitat vegetal no pareixen afectar el transport
nuclear d’ATHB4. Per tant, l’efecte d’aquests components del NPC ha de ser degut o a la
defecte de la importació d’altres proteïnes o en el transport d’RNA’s.
sar3-1 dra2-1dra2-3 dra2-4
Col-0 Col-0 PBL
dra2-1BC3(CMG34a5)
Finalment, ha estat àmpliament descrit que diverses mutacions en components del
transport nuclear mostren alteracions en l’exportació d’RNA missatger (mRNAs). Per tal de
veure si la mutació dra2-1 també afectava l’exportació d’mRNA es van realitzar assajos in situ
per detectar els poliA+-RNAs en aquest mutant i també pels al·lels en Col-0. Tal com està
descrit, una hibridació amb una sonda d’oligo-dT marcada al seu extrem amb un grup cromòfor
(com una fluoresceïna), permet que la sonda uneixi les cues d’adenines dels mRNA acabats de
sintetitzar i visualitzar-ne la localització majoritària (és el que es coneix com WISH, Whole
amount in situ hybridization). Es van fer servir plàntules de 7 dies crescudes en condicions
normals, en W i en plaques de 0,5xMS-, i les hibridacions es van fer seguint els protocols
descrits a la bibliografia (Gong, et al. 2005, Parry, et al. 2006). Els resultats mostren clarament
que el mutant dra2-1 té un exportació d’mRNA defectiu, amb una senyal molt intensa al nucli,
mentre que els al·lels de DRA2 en Col-0 no semblen alterar el tràfic de l’RNA (figura C2.20). Tal
com està publicat, els mutants nup96-1 i nup160-4, en el fons genètic Col-0 i que tenen
respostes a ombra atenuades, també presenten una atenuació de l’exportació d’RNAs. Es va
realitzar el mateix assaig in situ amb el creuament CMG34a5, al·lel dra2-1 en Col-0 que mostra
un fenotip feble, i es va veure que tampoc mostrava l’exportació d’RNA afectat. Així doncs,
sembla que els al·lels considerats febles tenen el transport correcte o molt feblement afectat,
Figura C2.20. Anàlisis per WISH del tràfic nuclear per poliA+-RNA en diversos mutants de NUPs. Anàlisis de localització de
poliA+-RNA en els controls wt i en diferents mutants de NUPs. Imatges obtingudes en tres experiments independents amb
microscopi confocal mirant cotiledons de plàntules de 7 dies.

3 Capítol 2
89
mentre que els mutants forts presenten acumulació d’mRNAs al nucli, possiblement per una
exportació defectuosa.
De l’anàlisi del transport a través del NPC podem concloure que els fenotips observats
han de ser deguts majoritàriament a defectes en l’export d’RNA, relacionant aquest defecte
amb l’atenuació de la resposta a ombra. No obstant, l’alteració dels marcadors moleculars en
els mutants que presenten fenotips forts són diferents, ja que tal com hem vist, només dra2-1
sembla tenir les respostes moleculars a ombra atenuades. Aquest fet suggereix que cada NUP
pot estar regulant el transport d’un grup o subgrup d’RNAs o proteïnes, afectant de manera
específica diversos components de la senyalització, o que com a mínim DRA2 té un rol
específic en la regulació de la senyalització per ombra.


4 Discussió
91
4. DISCUSSIÓ
4.1. Estudi de la relació estructura-funció d’ATHB4.
La síndrome de fugida de l’ombra (SAS) regula el creixement a través de diferents
mecanismes. La seva complexa regulació, mitjançant controls hormonals tant de síntesi com de
sensibilitat, inclou a més diversos controls per tal d’evitar una resposta excessiva, en una
dualitat d’expressió tant d’activadors com repressors que dóna lloc a un control temporal i
espacial molt important. Els gens PAR identificats al laboratori han demostrat ser elements clau
en la regulació d’aquestes respostes (Roig-Villanova, et al. 2006). Concretament ATHB4 ha
resultat ser un regulador complex de la SAS i d’algunes respostes hormonals, actuant com un
integrador de la senyal ambiental (proximitat vegetal) i les vies endògenes que controlen el
creixement (hormones) (Sorin, et al. 2009); a més, i de manera redundant amb altres membres
de la seva subfamília (HD-Zip II), ATHB4 té un rol en el desenvolupament de la planta (Bou-
Torrent, et al. 2012).
Resultats previs del grup van demostrar que la sobre-expressió d’ATHB4 altera el
desenvolupament afectant l’elongació de l’hipocòtil, l’expansió dels cotiledons, el creixement de
l’arrel i el desenvolupament d’arrels laterals i la polaritat de les fulles (Bou-Torrent, et al. 2012,
Sorin, et al. 2009). Aquest fenotip és similar al descrit en les línies de sobre-expressió
constitutiva d’ATHB2 (Ohgishi, et al. 2001, Steindler, et al. 1999), HAT1 (Ciarbelli, et al. 2008) i
HAT2 (Sawa, et al. 2002). Aquestes similituds fenotípiques de les sobre-expressions, juntament
amb la redundància gènica visualitzada per la manca de fenotip dels mutants simples,
suggereixen que tots aquests factors tenen el mateix rol en el control del desenvolupament.
Aquest fet és semblant al que ocorre amb els membres de la subfamília HD-Zip III (Prigge, et
al. 2005), que també presenten certes redundàncies en activitats relacionades amb el
desenvolupament.
4.1.1 La família HD-Zip: una activitat biològica plena d’interaccions.
ATHB4 és un factor de transcripció que pertany a la gran família HD-Zip, família que
sembla ser molt important per regular el desenvolupament de la planta de manera conjunta i
coordinada, amb capacitat de resposta a diversos estímuls. Com hem vist a la introducció, totes
les subfamílies presenten certa redundància de funció entre els seus membres i diverses
subfamílies tenen dominis que regulen l’activitat de la proteïna en relació a diversos estímuls.
Aquests fets remarquen la importància de les HD-Zip en el creixement de la planta.
La percepció pels fitocroms d’una reducció en la raó R:FR pot afectar ràpidament i de
manera directa l’expressió de diversos gens de la subfamília HD-Zip II, que al seu torn
autoregulen negativament la resta de membres de la subfamília, en un mecanisme que pot ser
una manera de frenar la pujada d’expressió després de la percepció d’ombra (Ohgishi, et al.
2001), per tal d’evitar una resposta exagerada i mantenir a la llarga uns nivells adequats
d’aquests factors. No obstant, aquesta regulació negativa complica la interpretació dels

4 Discussió
92
resultats en les línies de sobre-expressió. Com a conseqüència, els fenotips similars observats
en plantes amb alta activitat d’alguns d’aquests factors pot resultar, en part, de la regulació
aigües avall de l’expressió de tota la subfamília HD-Zip II, més enllà de l’activitat pròpia d’un sol
dels factors. Així doncs podem hipotetitzar que la subfamília HD-Zip II pot actuar com a un
mòdul conjunt, especialment a jutjar per la falta de fenotips en mutants simples, però no podem
descartar especificitats de funció d’alguns membres, a jutjar per algunes diferències en
l’expressió local i temporal d’aquests factors.
De fet, és lògic pensar que, tot i aquesta autoregulació negativa àmpliament descrita,
molts membres han de tenir altres activitats transcripcionals més rellevants, perquè es fa
estrany pensar que aquests components incrementen els seus nivells d’expressió només per
després autoinhibir-se. Per tant, trobar altres dianes d’aquestes proteïnes pot obrir la porta a
entendre millor el seu paper, tant en respostes a ombra com en desenvolupament. Mirant les
possibles dianes directes d’ATHB4, hem trobat per exemple HAT1 i HAT2 (figura C1.13 i taula
C1.3). Si bé aquests gens tenen pics d’expressió màxima al principi de la percepció d’ombra,
aquests nivells es veuen atenuats en tractaments més llargs, possiblement com a resultat
d’aquesta regulació negativa de la pròpia subfamília (Bou-Torrent, et al. 2012, Ciarbelli, et al.
2008, Sorin, et al. 2009). No obstant, aquests dos membres estan àmpliament relacionats amb
altres activitats més enllà de la regulació dins la seva subfamília, com per exemple en
respostes a auxines i desenvolupament del carpel; (Sawa, et al. 2002, Zuniga-Mayo, et al.
2012). A més, hem identificat altres gens regulats directament per ATHB4, com SAUR15 o el
transportador de pèptids OPT1 (figura C1.13 i taula C1.3). Segons dades publicades, tots ells
poden ser expressats en teixits com les fulles i flors, on també s’expressa ATHB4 (Koh, et al.
2002, Turchi, et al. 2013). No obstant, els problemes tècnics amb els experiments de ChIP han
fet que encara no puguem confirmar amb rotunditat l’efecte sobre aquestes dianes.
Més enllà de la pròpia subfamília II hi ha una certa comunicació entre diferents
subfamílies HD-Zip. Per exemple, ATHB2 és un repressor transcripcional que depenent de la
seva concentració cel·lular podria unir gens diana d’altres membres de la família HD-Zip, com
per exemple d’ATHB1 o d’ATHB8 (Steindler, et al. 1999). Canvis en els nivells d’aquesta
proteïna, deguts a canvis de raó R:FR per exemple, podrien canviar els equilibris entre
diferents membres de la família HD-Zip i canviar la regulació dels seus gens diana. ATHB8 és
un gen relacionat amb auxines que s’expressa en cèl·lules provasculars i incrementa el
creixement secundari vascular. ATHB2 pot unir in vitro les mateixes caixes consens que
ATHB8, tot i que possiblement amb menor afinitat, i per tant alts nivells d’ATHB2 podrien
competir pels mateixos gens diana (Steindler, et al. 1999). Donat que les seqüències d’unió
dels diferents membres de la subfamília HD-Zip II són semblants (figura C1.12) no seria estrany
pensar que el mateix fet podria passar per ATHB4. A més, aquests factors podrien assumir
altres rols, com per exemple un paper com a cofactors transcripcionals heterodimeritzant amb
altres HD-Zip, segrestant-los de la unió als seus promotors diana o aportant una activitat
repressora al unir-se l’heterodímer al seu gen diana. Dins la família HD-Zip hi ha altres
connexions, per exemple, experiments de ChIP-seq amb una HD-Zip de la subfamília III

4 Discussió
93
involucrada en el desenvolupament de les fulles, REV, han portat a demostrar que aquest
factor uneix al promotor de diversos gens de la subfamília II, com HAT2, HAT3, ATHB2 i
ATHB4, activant-ne la seva expressió (Brandt, et al. 2012). En el mateix treball es suggereix la
possibilitat que REV podria estar unint a les mateixes seqüències en els promotors que els
propis membres de la subfamília HD-Zip II utilitzen per regular-se negativament, suggerint que
factors de la subfamília II i de la III podrien competir per la regulació dels nivells d’expressió de
la subfamília II. Així, les connexions entre membres de les HD-Zip podrien venir principalment
per una competència en les seqüències de DNA a unir, però no són descartables relacions de
interacció directe a nivell de proteïna.
En general s’ha detectat una clara redundància entre ATHB2, HAT3 i ATHB4. Les
plantes sobre-expressores d’aquests tres membres presenten fenotips molt similars i els
mutants simples tenen efectes molt febles o nuls, mentre que els dobles i triples mutants
presenten fenotips molt més forts. Les tres proteïnes afecten al transport d’auxines i al
desenvolupament del teixit vascular, canviant els patrons d’expressió de proteïnes importants
pel transport d’aquestes hormones com PIN1 (Turchi, et al. 2013). HAT3 s’uneix al promotor
d’ATHB2 i regula negativament la seva expressió. Tots aquests factors són expressats en
diversos teixits, en especial coincideixen en les vessants adaxials (superiors) dels cotiledons,
on també són expressats diversos membres de la subfamília III, fet que posa de manifest la
rellevància biològica d’aquestes connexions. Aquest joc d’interaccions fa molt complex el model
d’activitat de cada membre, ja que és molt difícil mirar-ne l’activitat individual. Això ens porta a
pensar que diferents subfamílies HD-Zip actuen com un sol mòdul regulador, controlant
diferents aspectes del desenvolupament i del creixement de la planta de manera altament
coordinada.
No seria menyspreable l’opció que el fenotip presentat per la sobre-expressió d’ATHB4
fos fruit d’una expressió ectòpica més que no pas degut a una activitat pròpia de la proteïna.
Aquest fet ha estat observat en altres sobre-expressions (per exemple per la sobre-expressió
dels phyC (Franklin 2003). No obstant, anàlisis en les bases de dades i experiments no
publicats del laboratori porten a pensar que no és el cas, ja que ATHB4 s’expressa de manera
natural en diversitat de teixits i el fet que la seva sobre-expressió produeixi canvis fisiològics en
els teixits on normalment és expressat porta a pensar que són efectes d’una activitat
augmentada, més que no pas un efecte d’expressió ectòpica. Això, sumat a la redundància de
funció amb ATHB2 i HAT3 i als fenotips conjunts de les seves sobre-expressions fa pensar que
realment tenen algun rol concret en el desenvolupament i en les respostes a ombra.
4.1.2 ATHB4 uneix al DNA i actua com a repressor de la transcripció.
L’estudi de la unió d’ATHB4 al DNA mitjançant l’ús de microarrays de proteïnes (PBM) ha
portat a identificar la seqüència consens d’unió de 7-9 pb, sent similar a altres membres de la
subfamília (figura C1.12). El residu central de les seqüències consens estava descrit com a
molt important i específic de cada proteïna de la família HD-Zip (Tron, et al. 2001), si bé en la

4 Discussió
94
subfamília II sembla que en alguns casos podria ser el mateix, mostrant que poden unir
pràcticament als mateixos llocs, tot i que les afinitats òptimes semblen variar. Les seqüències
consens 7-mer per ATHB4 i ATHB2 semblen ser idèntiques comparant el resultat del PBM amb
els resultats publicats per ATHB2, amb una especificitat d’unió a AATSATT (on S = G o C)
(Sessa, et al. 1993). Si ampliem una mica la seqüència, a la consens 9-mer, ATHB4 prefereix
als extrems una A (tant al 5’ com al 3’, AAATSATTA), mentre que ATHB2 està descrit que
prefereix una T o una C a l’extrem 5’ (T/CAATSATTA/G), amb la corresponent base
complementària al 3’ (A o G respectivament). Així, la seqüència 5’-CAATNATTG-3’ és
reconeguda per ATHB2 en assajos in vitro (Sessa, et al. 1997) i in vivo per experiments
d’expressió transitòria (Steindler, et al. 1999). Comparant aquestes seqüències d’unió dels dos
membres de la família HD-Zip II amb altres membres de la família HD-Zip veiem que són prou
similars; per exemple, ATHB1 (HD-Zip I) uneix a seqüències tipus NAATWATTN (amb W = A o
T, (Sessa, et al. 1993), i membres de les HD-Zip III uneixen a seqüències tipus
NNAATSATGNN (Brandt, et al. 2012, Turchi, et al. 2013). No seria descartable que de manera
similar al que està descrit amb ATHB2, ATHB4 també pogués competir unint dianes directes
d’altres factors de transcripció HD-Zip. A més, la possibilitat de formar complexos
heterodimèrics podria canviar lleugerament les seqüències unides in vitro i in vivo, tal com s’ha
proposat amb altres famílies de factors de transcripció (Zhang, et al. 2013).
Dades del laboratori apunten que ATHB4 actua fonamentalment com a repressor
transcripcional (microarrays comentats a l’apartat 3.1.2.3 i experiments qPCR de figuraC1.16).
Experiments preliminars de ChIP semblen confirmar aquest paper d’ATHB4 com a mínim en
membres de la seva subfamília, reafirmant aquest cicle de regulació negativa, permetent veure
que, tal com apunten altres autors, és una activitat general als diversos membres de la HD-Zip
II. L’optimització de la ChIP ha de permetre comprovar la unió d’ATHB4 a altres promotors de
gens que podrien estar regulant processos de desenvolupament més enllà de la subfamília HD-
Zip II. Serà interessant comprovar a més, si la regulació negativa de diversos gens és directa,
per una unió d’ATHB4 al seu promotor, o mitjançant altres mecanismes, com el segrest de
factors que actuen com activadors transcripcionals (actuant per tant com un cofactor
transcripcional, tal com s’ha descrit per a les bHLH atípiques HFR1 i PAR1)(Galstyan, et al.
2011) o fent de pont entre proteïnes repressores de la transcripció i factors que inicialment
podrien actuar com activadors. Aquests mecanismes, que no són excloents, no han estat mai
descrits en factors HD-Zip, però diverses dades experimentals del nostre laboratori porten a
pensar que com a mínim un d’ells podria estar succeint (figures C1.5, .7 i .10).
4.1.3 Estudi de la relació estructura-funció d’ATHB4.
En la present tesi hem analitzat la participació de les diferents regions o dominis
d’ATHB4 en les diferents activitats descrites per aquesta proteïna, com serien (1) l’efecte sobre
l’allargament de l’hipocòtil induït per l’exposició a ombra simulada i (2) la repressió
transcripcional en l’expressió de gens marcadors. Aquests fenotips s’han avaluat en plantes
transgèniques que sobre-expressen diferents formes truncades d’aquest proteïna. A més,

4 Discussió
95
aquesta informació s’ha contrastat amb les dades sobre la localització subcel·lular d’aquestes
formes i la capacitat de dimerització, obtinguda en una tesi doctoral prèvia del laboratori. Els
estudis fets amb les diferents formes truncades porten a demostrar clarament que la seva
activitat biològica depèn fortament de la regió Nt, fins ara poc estudiada. En canvi depèn molt
poc (o gens) de la regió Ct (figures C1.2-7), que conté algunes Cys que podrien ajudar a fer
més resistent la proteïna davant estrès oxidatiu. Està descrit que en la regió de dimerització,
dins el domini ZP, hi ha tres Cys que ajuden a estabilitzar la formació de dímers mitjançant
enllaços disulfur. Aquests enllaços es poden veure afectats per condicions oxidatives, de
manera que es podria desestabilitzar el dímer sota aquest tipus d’estrès. Les Cys de l’extrem
Ct no semblen ser requerides per formar els dímers (Tron, et al. 2002), però podrien ser una
cua protectora de les Cys importants, trencant-se primer els seus ponts disulfur de manera que
protegirien de l’atac oxidatiu les Cys importants, ajudant a preservar el complex actiu fins i tot
en condicions oxidatives. Així doncs, el model plantejat seria que en condicions reductores les
Cys del Ct estarien afavorint la formació de dímers i l’afinitat d’unió al DNA seria màxima,
mentre que en condicions oxidants l’activitat es podria anar perdent paulatinament a mesura
que s’anessin oxidant les Cys, i això podria canviar les afinitats d’unió al DNA fent que algunes
es perdessin i altres no. Si bé part d’aquesta hipòtesi es veu reforçada per experiments
publicats faltaria fer alguns experiments, amb les truncacions sense la regió Ct, per comprovar
si realment aquestes són menys resistents a l’estrès oxidatiu o si es produeixen canvis en
l’activitat de la proteïna o en la seva afinitat d’unió al DNA. El nostres resultats mostren que la
regió Ct no és necessària per la modulació de les respostes de la SAS, potser perquè en les
condicions assajades no hi ha estrès oxidatiu.
L’estudi de la regió Nt és molt més complex, en part perquè és més llarga i poden haver-
hi diversos motius o dominis amb rols diferents. L’única regió identificada, el motiu EAR, està
sent estudiat en un projecte en marxa, dins els objectius d’una altra tesi doctoral. En el present
treball hem avançat en l’estudi d’aquest motiu fent una mutació puntual, veient que aquesta fa
perdre part de l’activitat, sobretot en l’elongació de l’hipocòtil (construcció pMG43, figura C1.10-
11). Serà interessant analitzar com aquesta mutació impedeix la unió d’ATHB4 amb les
proteïnes repressores i si l’efecte pot ser major mutant vàries de les Leu del motiu EAR
(considerat LxLxL). La funció de la resta de la regió Nt és encara desconeguda, però sembla
que la part C-terminal d’aquest regió és insuficient per mantenir l’activitat que si és present
quan l’Nt es troba sencer (construcció pMG44, figura C1.10), reforçant la idea que el motiu EAR
és necessari per l’activitat de la proteïna en les respostes a ombra.
La variabilitat observada en algunes construccions (sobretot per línies de pMG38 i
pMG43) fa pensar que els canvis introduïts per les mutacions puntuals no són prou dramàtics
per anul·lar completament l’activitat de la proteïna, però si que causen alteracions importants,
resultant en defectes en la seva activitat. No obstant, és possible que mutacions menys
conservatives o mutacions de més d’un aminoàcid donessin lloc a resultats menys variables.
Més enllà de la regió Nt sembla que el domini HD, a més de l’activitat d’unió al DNA,
conté informació per dirigir la proteïna al nucli, ja que les construccions sense HD es localitzen

4 Discussió
96
també al citoplasma (Salla Martret 2012)(figura C1.9). El conjunt dels dominis HD i ZP són
suficients per donar lloc a la homodimerització i, experiments publicats en assajos d’expressió
transitòria in vivo, semblen mostrar que possiblement són suficients per unir al DNA (Steindler,
et al. 1999). No obstant els nostres resultats porten a pensar que, malgrat la capacitat de
dimeritzar i d’unir al DNA, el conjunt HD-Zip té poca o nul·la activitat biològica en respostes a
ombra sense la regió Nt (figura C1.2-7). A més, està descrit que el domini ZP és essencial per
unir DNA en les HD-Zip (Tron, et al. 2002), i sorprenentment la construcció pMG28, sense tenir
el domini ZP, dóna lloc a una activitat molt semblant a la proteïna sencera, suggerint que
ATHB4 potser actua en resposta a ombra de manera independent a la unió al DNA. Aquesta
hipòtesi es consistent amb l’activitat biològica en la línia pMG37, amb una mutació puntual al
HD que hauria de fer-lo incapaç d’unir al DNA, si bé és cert que no hem pogut comprovar
l’efecte d’aquesta mutació puntual sobre la capacitat d’unió al DNA de la proteïna resultant.
Està descrit que alguns HD-Zip no uneixen bé al DNA en tècniques in vitro, potser perquè
requereixen modificacions post-transcripcionals, ja que tenen punts de fosforilació que podrien
ser importants per l’activitat (Johanesson et al. 2001), això podria explicar les dificultats per fer
servir assajos d’EMSA en aquestes construccions.
Cal considerar que algunes construccions, tot i no mostrar activitat en l’allargament de
l’hipocòtil, podrien tenir algunes respostes moleculars alterades, tal com s’ha vist per la
proteïna PIF3 (Al-Sady, et al. 2008), que presenta la capacitat d’unió al DNA desacoblada de la
seva activitat en l’allargament de l’hipocòtil. Els nostres resultats mostren en general
consistència entre les alteracions fisiològiques i moleculars. Només en alguns casos
determinats, com les construccions pMG26 o pMG27 que contenen el domini ZP, es veuen
alguns fenotips febles. En aquests casos, l’activitat observada sobre l’elongació de l’hipocòtil
podria ser deguda a un efecte indirecte de nivells alts de proteïnes parcialment funcionals, que
mantenen l’habilitat per dimeritzar (via el domini ZP), de manera que podrien segrestar l’ATHB4
endogen, o altres HD-Zip, i impedir-los, com a mínim parcialment, la seva activitat. Això seria
consistent amb el fenotip de línies com la pMG27.3 que tenen característiques inverses al
guany de funció, amb cotiledons més grans i hipocòtils i pecíols més llargs.
És també sorprenent que la sobre-expressió de truncacions com la pMG28 (mancada de
domini ZP, per tant d’habilitat de dimeritzar, i per tant d’unir el DNA) i la pMG37, que conté una
mutació puntual al HD que hauria d’anul·lar la unió al DNA, tinguin una activitat com la
d’ATHB4 sencera, tant en la regulació de l’allargament de l’hipocòtil com en la regulació de
l’expressió gènica. Aquests resultats fan pensar que o bé uneixen al DNA per mecanismes no
coneguts o el paper d’ATHB4 en les respostes a ombra és independent de la unió al DNA,
suggerint que ATBH4 pot tenir un efecte sobre l’allargament de l’hipocòtil deslligat de la seva
unió al DNA. Assajos amb aquestes construccions per tal de comprovar la seva unió o no al
DNA poden ser de gran rellevància per respondre a aquesta incògnita.
Així doncs, dues opcions sembla que poden explicar el conjunt de resultats obtinguts per
les anàlisis de l’activitat biològica en les respostes a ombra: (1) ATHB4 pot unir al DNA formant
hete o homodímers mitjançant dominis de la regió Nt, de manera independent del domini ZP i

4 Discussió
97
de l’activitat d’unió al DNA del HD; o (2) en respostes a ombra, ATHB4 està actuant sense
necessitat d’unir al DNA, mitjançant interaccions amb la seva regió Nt. Futurs experiments han
de permetre confirmar una d’aquestes dues hipòtesis.
El fet que diverses construccions que sobre-expressen parts d’ATHB4 no mostrin
respostes a ombra alterades posa de manifest que l’efecte biològic és específicament degut a
la regió Nt. No obstant, caldrà comprovar si el paper d’aquestes construccions en el
desenvolupament manté el mateix patró, o si, tal com sembla en els resultats preliminars per la
construcció MG58, equivalent a la pMG27 i que mostra cert fenotip amb una alteració en la
polaritat de les fulles, les dues activitats depenen d’alguns dominis o regions diferents. Pel que
sembla per les respostes a ombra seria necessària sobretot la regió repressora Nt amb el motiu
EAR (i altres possibles activitats encara no descrites), mentre que pel desenvolupament, a més
de l’activitat repressora, serien necessàries les capacitats de dimeritzar i unir al DNA a través
dels HD i ZP.
4.2. Regulació de les NUPs.
A més de la regulació transcripcional de les respostes de la SAS, la identificació de
DRA2 ens ha permès esbrinar un nou nivell de regulació de les respostes iniciades per la
proximitat de vegetació: el transport a través del NPC. Les NUPs controlen un procés que cada
vegada més està demostrant ser molt important pels organismes eucariotes, com és el control
del tràfic de molècules a través del nucleoporus. Aquest complex regula d’una manera directa o
indirecta molts processos essencials pels organismes, ja que la translocació de proteïnes al
nucli i l’exportació d’RNAs al citosol són requerits per gairebé qualsevol resposta.
Especialment, s’està veient que la NUP98 té molts rols, no només dins el NPC, sinó també
podent estabilitzar RNAs i unir a regions encara desconegudes de la cromatina. No obstant, el
paper de l’equivalent a NUP98 en planta encara no és conegut. En la present tesi involucrem
aquest component, anomenat per nosaltres DRA2, així com altres nucleoporines, en els
processos de senyalització de llum, especialment en respostes a ombra, afectant les respostes
de la síndrome de fugida de l’ombra.
4.2.1. DRA2: caracteritzacions fisiològiques i moleculars.
La caracterització general del mutant dra2-1 ens porta a veure que, com els mutants en
altres NUPs, afecta a diverses respostes i a diverses rutes de senyalització, com les respostes
a BRs i GAs (figura C2.1, i també les dades del microarray), en el que es podrien considerar un
conjunt d’efectes pleiotròpics. No obstant, hem vist que especialment la senyalització d’auxines
sembla incrementada constitutivament, de tal manera que es produeix una menor resposta a
auxina exògena (dades del microarray)(Galstyan 2010). No obstant, els nivells reals d’auxina
no semblen ser diferents al control wt (figura C2.2a), suggerint que aquest increment en nivells
d’expressió de gens de resposta a auxines és degut a canvis en la sensibilitat, afectant sobretot
a la regulació de gens repressors de la senyalització d’auxines (IAAs).

4 Discussió
98
El mutant va ser trobat mesurant-ne l’activitat LUC en tractaments de dues hores
d’ombra simulada. S’ha comprovat que les mesures no es veien afectades pels nivells de
pigments, ja que aquests són molt similars en el mutant dra2-1 respecte al seu control wt
(figura C2.2b). S’ha vist que els nivells de pigments augmenten en presència de sucrosa al
medi, mentre que les respostes a ombra es veuen atenuades (Dijkwel, et al. 1997).
La localització subcel·lular de parts de la proteïna DRA2 per microbombardeig fa pensar
que el fragment Nt resultat de la mutació dra2-1, que és equivalent a la construcció pMG56, ha
perdut la capacitat d’unir-se al NPC (figura C2.4). Aquest resultat no és sorprenent, ja que està
descrit que la NUP98 en humans obté aquest anclatge a través de la regió Ct (Sun and Guo
2008), regió prou conservada. Per tant, fragments de DRA2 sense aquesta regió Ct podrien ser
incapaços d’unir-se al NPC. No obstant, mentre que el fragment Ct assajat sembla poc estable
en planta, la regió Nt segueix sent estable i es pot localitzar formant vesícules tant dins del nucli
com al citosol (figura C2.4), de manera similar a les sobre-expressions de NUP98 en cultius de
cèl·lules humanes (Griffis, et al. 2002). A més, línies transgèniques sobre-expressant aquesta
regió Nt de DRA2 en plantes, línies pMG56, mostren fenotips clars, amb problemes de
desenvolupament, suggerint que aquesta regió és suficient per mantenir alguna activitat de la
proteïna sencera. No és descartable que aquest fenotip sigui degut a un problema en
l’emmagatzematge de la proteïna, ja que aquesta part Nt amb repeticions FG és bastant
hidrofòbica i pot causar certs efectes inesperats a les cèl·lules que la sobre-expressen, com és
el fet de veure’n la localització en vesícules també al citosol. Una manera de descartar aquest
problema seria obtenir noves línies transgèniques expressant aquesta construcció sota control
del promotor endògen de DRA2, o fusionant aquesta regió Nt a senyals de localització nuclear
(NLS), de manera que podríem esperar eliminar l’acumulació citosòlica, bé per tenir nivells
inferiors de proteïna o bé per obligar-los a localitzar-se al nucli. De fet, diverses línies
transgèniques pMG56 que no mostren activitat GFP tampoc mostren fenotip, suggerint que
nivells molt baixos d’expressió no tenen cap activitat biològica.
Per tant, pot ser que dra2-1 tingui una activitat extra que no tenen els altres al·lels
mutants per DRA2, però potser certs nivells d’expressió són necessaris per poder veure
aquesta activitat. Consistentment, al introduir aquest al·lel mutat (dra2-1) en ecotip Col-0
sembla ser similar fisiològicament a la resta de mutants de pèrdua de funció (anàlisis amb dra2-
1BC3 o CMG34a5, figura C2.12). No obstant, les respostes moleculars no són equivalents del
tot entre aquest al·lel i la resta de mutants en Col-0 (figura C2.14), suggerint que aquest al·lel
podria tenir alguna activitat molecular que la resta no tindrien i que aquesta activitat molecular
seria independent del fenotip fisiològic. És veritat que aquest creuament encara conserva part
de l’ecotip Ws-2, i que els ecotips Ws-2 i Col-0 són bastant diferents. Les plantes en Ws-2
semblen més vigoroses, creixen més i floreixen abans que les Col-0. Està descrit que tenen un
al·lel no actiu del PHYD, però el fenotip de dra2-1 és independent d’aquesta variació en el
fitocrom. Pels creuaments fets, sembla que com a mínim dos factors més intervenen en fer
notori aquest fenotip (figura C2.12; dades de segregació fenotípica en CMG55). A més, no és
possible diferenciar fenotípicament els individus heterozigots dels wt, suggerint que la mutació

4 Discussió
99
dra2-1 és realment recessiva (anàlisis de CMG51)(Galstyan 2010). Els estudis amb els
creuaments dels al·lels originals de Col-0 passats a Ws-2 (dra2-3/4BP2/3, o CMG48 i CMG49,
figura C2.13) ens porten a pensar que podria ser que hi hagués algun altre factor proper en el
genoma al gen DRA2 necessari per fer notori el fenotip, de manera que al fer els creuaments
seria molt difícil de perdre o guanyar aquest element, és a dir, que estaria lligat a DRA2.
Aquesta hipòtesi, que el fons genètic (ecotip) influenciï de manera molt important a les
respostes, s'ha descrit a la bibliografia per exemple en respostes a ombra i a la temperatura
(Patel, et al. 2013). Per tant, els resultats ens suggereixen que el fragment de DRA2 expressat
en l’al·lel dra2-1 pot tenir alguna activitat, però que el fenotip característic de la línia pot venir
donat per dos elements clau de l’ecotip Ws-2. No obstant calen més estudis per comprovar
aquestes hipòtesis i determinar l’efecte molecular del fragment Nt de DRA2 expressat en la
construcció pMG56 i en l’al·lel dra2-1. Contràriament al que hem vist per DRA2, un al·lel de
pèrdua de funció per NUP160/SAR1 trobat en ecotip Ws-2, anomenat sar1-7, sembla
comportar-se de igual manera al seus al·lels en Col-0 (Robles, et al. 2012). Així doncs, les
diferències entre els NPC d’aquests ecotips deuen ser prou específiques respecte a DRA2 o
les seves funcions.
Les anàlisis de les respostes a ombra de varis mutants en altres NUPs ens fan pensar
que en general un defecte en el NPC comporta un defecte en la senyalització per ombra, ja que
els mutants que presenten algun tipus d’alteració fenotípica responen menys al tractament,
mentre que els mutants amb aparença similar al wt, responen de manera normal a ombra
(figures C2.8). A més, sembla que diverses NUPs poden tenir papers parcialment redundants
en aquesta senyalització, ja que alguns mutants simples no presenten alteracions importants ni
en el desenvolupament ni en la senyalització de la planta (al·lels per NUP54 i NUP58 en Col-0)
(figures C2.8). No obstant, quan s’obtenen dobles mutants, aquests mostren que un major
defecte en el NPC produeix un fenotip fort (figures C2.15 i 16), amb una resposta a ombra
reduïda. D’altra banda, dos dels tres al·lels mutants en NUP62 tenen fenotips forts, suggerint
que aquesta NUP és essencial per la planta. Els fenotips d’aquestes línies mutants són
semblants als descrits a la bibliografia per plantes sobre-expressores que han silenciat el
transgen i el gen endògen (Zhao and Meier 2011), si bé en l’article els autors no veuen cap
fenotip apreciable en els tres al·lels mutants descrits en la present tesi. No entenem aquesta
diferència, però seria possible que la inviabilitat dels al·lels nup62-1 i nup62-2 no fos observada
i que, per tant, no hagin pogut aïllar mai els mutants en homozigosi.
Les dades moleculars, tant pel microarray com les diferents qPCR amb mutants en
NUPs, ens indiquen que l’expressió de DRAL és un bon indicador de l’efecte de les mutacions
sobre el NPC, mostrant que un major defecte del NPC està relacionat amb nivells més alts
d’expressió de DRAL.
Els resultats del microarray suggereixen que la mutació dra2-1 afecta a diverses rutes i
mecanismes de la planta, especialment a la senyalització per auxines, però també manté
certes coincidències amb la senyalització de llum controlada per PIF5 (figura C2.6). Com que
les anàlisis de GO per les dianes de PIF5 van concloure que hi ha un enriquiment en gens de

4 Discussió
100
resposta a auxines (Hornitschek, et al. 2012), és difícil arribar a la conclusió de si l’efecte de
DRA2 sobre la senyalització per auxines és directe o si és degut a una senyalització defectuosa
de PIFs. També es pot veure que dra2-1 afecta molt més al transport a través del NPC que no
pas a l’estructura del porus en si mateix, ja que es veuen increments els nivells d’expressió de
varis components relacionats amb el NPC, però no components de l’estructura (com seria el
complex NUP107-NUP160), sinó components de les vies de transport (importines, exportines,
altres NUPs de repeticions FG i components del complex RANGAP). Aquest augment en
l’expressió de gens implicats en el transport, tant de proteïnes com d’RNAs, pot ser que formi
part d'un mecanisme compensatori que aconsegueixi suplir parcialment la falta de DRA2, ja
que no hem vist el transport de proteïna alterat. Aquests resultats són consistents amb el fet
que NUP98 és una NUP de repeticions FG, i que per tant no forma part de l’estructura del NPC,
sinó de la malla o teixit de repeticions FG que bloquegen el porus i en regulen el pas.
4.2.2. DRA2 i el transport a través del NPC.
Els articles publicats sobre altres NUPs (Dong, et al. 2006, Parry, et al. 2006, Robles, et
al. 2012, Wiermer, et al. 2012), en especial amb mutants per NUP96 i NUP160, han explicat els
seus defectes de desenvolupament o de resposta emparant-se amb els defectes per exportar
mRNA, justificant així una menor resposta en les rutes de senyalització corresponents. En el
cas de Parry i col·laboradors (2006), a més, suggereixen un retard en la importació de proteïna,
però aquest no ha pogut ser constatat en altres treballs publicats (Dong, et al. 2006). En la
present tesi tampoc hem pogut concloure amb certesa que aquest mecanisme sigui important
per la menor resposta a ombra mostrada per aquests mutants (figures C2.18i19). Aquesta
discrepància pot ser deguda a diversos motius, que no són excloents: (1) la importació està
molt lleugerament atenuada i només en alguns mètodes i mirant molt en detall és possible
veure-ho; (2) el transport de proteïnes pot ser selectiu i específic, de manera que l’afectació
sobre la importació de proteïnes només es donaria en alguns determinats components i no en
qualsevol. En el nostre cas, si bé no ens atrevim a afirmar que la importació de la proteïna
ATHB4 estigui clarament atenuat, si que podria veure’s una tendència a jutjar pels resultats
d’expressió de SAUR15 i HAT2 en concentracions baixes de DEX (figura C2.18i19). Malgrat els
nombrosos estudis que hem fet en el present treball no hem pogut anar més enllà de confirmar
les hipòtesis publicades, podent explicar els defectes en les respostes a ombra per aquesta
deficiència en l’exportació d’alguns RNAs. Això és consistent amb mutants com los4 (low
expression of osmotically responsive genes 4) (Gong, et al. 2005), que en teoria només afecten
l’exportació d’RNA i presenten fenotips similars a mutants en NUPs. Però en el present treball a
més, hem fet una comparativa única de diferents components del NPC analitzant diversos
mutants simples i dobles d’aquests, demostrant que un major defecte en el nucleoporus dóna
lloc a l’atenuació de la resposta a ombra.
Així doncs, els nostres anàlisis ens porten a la mateixa conclusió pel mutant dra2-1 que
per les altres NUPs, una exportació deficient d’alguns mRNAs provoca un defecte en la
senyalització de les diverses rutes afectades, traduint-se aquest defecte en la senyalització en

4 Discussió
101
una menor resposta de l’hipocòtil a la proximitat vegetal. El fet que els mutants de DRA2 en
Col-0 no mostrin aquest efecte és sorprenent, ja que està descrit que NUP98, juntament amb
RAE, són part del complex d’exportació d’mRNAs (Lee, et al. 2009). L’explicació més raonable
és que DRAL, o algunes altres NUPs, puguin compensar la falta de DRA2 en Col-0. No
obstant, tampoc podem descartar que hi hagi altres funcions de la NUP98 que es vegin
afectades. Aquests resultats són consistents amb els publicats fins al moment sobre mutants
del NPC, on gairebé tots els mutants presenten floració primerenca, alteracions en sensibilitat a
auxines i defectes en l’exportació d’mRNA (Jacob, et al. 2007, Kasper, et al. 1999). Així,
sembla que diferents NUPs i proteïnes associades al NPC tenen rols similars, però amb
algunes activitats específiques independents, en el desenvolupament de la planta. A Wiemer i
col·laboradors (2012) comenten que l’exportació d’mRNA en nup160 i seh1 està afectat
lleugerament (hi ha molt poques diferències comparant dades de qPCR amb RNAs del nucli i
del citoplasma), suggerint que algunes mutacions poden produir proteïnes truncades que no
siguin pèrdues de funció completes. A més el seu estudi posa de manifest que dins un mateix
complex (el complex NUP107-160) només hi ha tres NUPs que afecten les respostes de
defensa (NUP96, NUP160 i Seh1), mentre que mutants en les 5 NUPs restants no tenen cap
alteració aparent.
D’altra banda els resultats de les hibridacions in situ (WISH) ens mostren que els efectes
en el nucleoporus en les diferents NUPs mantenen correlació entre l’atenuació de l’exportació
d’mRNA i el fenotip fisiològic en ombra. Per tant, la resposta fisiològica atenuada pot ser
deguda a efectes en l’exportació d’mRNA, mentre que les respostes moleculars atenuades han
de ser degudes a algun efecte més específic de DRA2, potser a nivell de transport de proteïnes
concretes, potser afectant el transport d’alguns mRNA concrets més importants o potser per la
seva possible implicació en l’expressió gènica, podent unir-se directament a regions de la
cromatina. Aquestes diferències moleculars dels mutants per DRA2 respecte altres mutants en
NUPs seran estudiades en futurs treballs.
4.3. ATHB4 i DRA2 en la regulació de la SAS.
A mesura que es van estudiant les respostes de la SAS es va veient la complexitat que
hi ha al seu darrere, amb una regulació a múltiples nivells. El principal mecanisme sembla ser
una estabilització d’algunes proteïnes PIF, deguda a la menor activitat dels fitocroms,
especialment de phyB. Aquests factors PIFs estarien activant tota una sèrie de factors de
creixement a través del control de síntesi i sensibilitat a hormones, com les AUXs, però a més
també estan activant una sèrie d’antagonistes per mantenir sota control aquestes respostes. El
fet que els propis PIF puguin regular l’activitat dels fitocroms i que diversos factors afectin als
propis PIFs fa que un model jeràrquic de regulació sigui molt difícil d’establir. Molts dels estudis
s’han centrat en identificar tant les dianes dels PIFs com l’activitat antagonística d’altres factors
en un estadi de plàntula i prenent com a mostres pels anàlisis plàntules senceres. Queda per
determinar, per tant, un factor temporal i espacial, ja que la rellevància biològica de moltes de
les interaccions publicades depèn fortament dels teixits on es doni aquesta interacció. Pel cas

4 Discussió
102
dels factors estudiats en la present tesi, sabem que ATHB4 és expressat durant l’embriogènesi,
en flors i en les parts adaxials de cotiledons i fulles, mentre que DRA2 sembla tenir una
expressió prou general, estesa a tots els teixits i durant els diferents estadis de
desenvolupament. No obstant, a més de la seva expressió seria important conèixer els
mecanismes que en regulen la seva activitat, mecanismes de moment desconeguts en ambdós
casos. En el present treball ens hem centrat en els efectes d’aquests factors sobre l’elongació
de l’hipocòtil, sense perdre de vista que ATHB4 actua també regulant la polaritat de les fulles i
els cotiledons.
ATHB4 sembla que regula negativament el creixement en les respostes de la SAS, fent
d’antagonista sobre dianes que els PIFs estan activant (com SAUR15 i els diversos membres
de la seva pròpia subfamília). No obstant, en contra del que semblaria, aquesta activitat és
independent de la seva funció com a factor de transcripció, ja que el resultats de la present tesi
indiquen que formes truncades sense capacitat de dimeritzar tenen la mateixa activitat que la
proteïna sencera. Postulem per tant que ATHB4 està actuant en la SAS com a cofactor, sent
possiblement un repressor de l’expressió.
La regulació de la SAS mitjançada per DRA2 és encara difícil d’explicar. Si bé hem vist
que el mutant dra2-1 té un defecte en l’exportació d’mRNA, no sabem si l'efecte és específic
sobre mRNAs concrets o afecta per igual a tot el pool cel·lular d'aquest tipus d’RNAs. Sembla
que la resposta alterada a ombra de l'hipocòtil sigui un efecte inespecífic, com els múltiples
efectes pleiotròpics de diversos mutants en NUPs. No obstant, en el present treball hem
demostrat que DRA2 manté alguna especificitat en la senyalització de llum i per AUXs.
L’aproximació per tal de veure si DRA2 podria estar participant de les respostes de la SAS
regulant la importació de factors com ATHB4 al nucli no ens ha permès veure cap efecte,
suggerint que en tot cas no seria només DRA2 l’encarregada d’aquesta importació. El mateix
resultat ha estat obtingut per la importació d’ATHB4 en altres mutants per NUPs, com la NUP96
(figura C2.18-19.). Així doncs, encara no hem pogut demostrar per quin mecanisme molecular
podria estar DRA2 afectant l’expressió gènica de marcadors d’ombra, si bé a partir de la
bibliografia i per diversos resultats suggerim que podria estar regulant-ne l’expressió de manera
directa. No obstant, queda feina pendent per demostrar aquesta hipòtesi.
Així, tant ATHB4 com DRA2 podrien estar afectant l’estructura de la cromatina per tal de
regular-ne la seva expressió, en el primer cas a través de la unió a repressors en el seu motiu
EAR i en l’altre per mecanismes encara desconeguts. Serà interessant veure els futurs avenços
en aquesta regulació, ja que s’ha publicat que l’estructura de la cromatina és una peça clau en
la regulació de la transcripció i efectes en les marques epigenètiques o en l’empaquetament
poden modificar les respostes de manera específica al teixit o al temps de desenvolupament
(Henriques and Mas 2013, Malapeira and Mas 2013).

5 Conclusions
103
5. CONCLUSIONS.
(1) ATHB4 és fonamentalment un repressor de la transcripció, com a mínim per dianes
com SAUR15, OPT1 i altres membres de la seva subfamília. ATHB4 pot actuar com a
repressor transcripcional probablement unint a seqüències consens molt similars a altres
membres de la subfamília HD-Zip II.
(2) ATHB4 forma, per tant, part d’un mòdul regulador de la SAS dins la subfamília HD-Zip
II, que pot estar, al seu torn, formant part d’una regulació conjunta del desenvolupament en
relació amb altres HD-Zip, com per exemple amb membres de la subfamília III.
(3). Les anàlisis d'estructura-funció d'ATHB4 mostren que la seva regió Ct no és
important per les respostes a ombra, mentre que la regió Nt és essencial per mantenir l’activitat
biològica d’ATHB4, especialment la seva part inicial amb el motiu EAR. A més, ATHB4 actua en
ombra de manera independent a la dimerització a través del domini ZP i, possiblement, de
manera independent a la unió al DNA pel domini HD, dades que suggereixen que ATHB4 actua
en la regulació d'aquestes respostes possiblement com a cofactor transcripcional.
(4) DRA2 és realment una NUP d’Arabidopsis, similar a la NUP98 de llevats i d’animals.
L'extrem Nt, amb moltes repeticions FG, és capaç de localitzar-se al nucli en petites vesícules
d’activitat desconeguda, de manera equivalent a la sobre-expressió de la NUP98 en mamífers,
suggerint que aquesta regió podria dur a terme alguna activitat independent del NPC dins el
nucli, tal com està descrit per la NUP98.
(5) A més de dra2-1, diversos mutants simples en altres NUPs tenen la resposta de
l'hipocòtil a ombra atenuada, així com un fenotip pleiotròpic que afecta el desenvolupament. La
combinació de mutants febles en algunes d'aquestes NUPs, sense fenotip aparent, resulta en
l’aparició del fenotip pleiotròpic i la menor resposta de l’hipocòtil en ombra, així com un
increment dels nivells d’expressió de DRAL, suggerint que aquest gen és un bon marcador de
defectes en el NPC. Per tant, el transport de proteïnes i/o RNAs a través del NPC és important
per la correcta resposta de l'hipocòtil a la proximitat vegetal.
(6) Ni la mutació dra2-1 ni la nup96-1 semblen afectar de manera rellevant a la
importació d’ATHB4 al nucli. En canvi, la mutació dra2-1 si que afecta l’exportació dels poliA+-
RNA, tal com està descrit pel mutant nup96-1 i altres components del NPC, suggerint que la
menor elongació de l’hipocòtil en ombra és un defecte general en mutants de NUPs derivat
d’aquesta exportació deficient.
(7) La mutació dra2-1 presenta, a més, un efecte molecular únic respecte altres mutants
de NUPs, amb una resposta molecular a ombra també atenuada. Si bé s’ha demostrat que
l’ecotip influencia clarament al comportament de la mutació dra2-1, queda per confirmar si la
forma truncada resultant d’aquesta mutació pot tenir alguna activitat sobre l’expressió gènica,
causant el defecte molecular, o si aquest és degut a components propis de l’ecotip Ws-2.


6 Materials i mètodes
105
6. MATERIALS I MÈTODES. A continuació es descriuen tant els materials com els protocols més utilitzats durant el
desenvolupament d’aquesta tesi doctoral. Els mètodes generals de manipulació d’àcids
nucleics i proteïnes, així com els relacionats amb el cultiu i manipulació de bacteris es van
realitzar segons els protocols descrits a Molecular cloning: A laboratory Handbook (Maniatis et
al. 1982), Current Protocols in Molecular Biology (Ausubel 1989) i Molecular cloning: A
laboratory Manual (Sambrook and Russell 2001), amb algunes petites modificacions pròpies
del laboratori.
L’aigua utilitzada generalment va ser la desionitzada (dH2O), mentre que pels
experiments de biologia molecular va ser la bidestil·lada (ddH2O, amb el destil·lador i
desionitzador d’aigua Millipore Milli-Q).
Les solucions utilitzades es van esterilitzar per calor humit (mitjançant autoclau de vapor
d’aigua a 120ºC i una atmosfera de pressió, durant 20 min, o mitjançant filtració (amb filtres de
0,45µm). El pH dels medis i solucions s’ajusta abans de sotmetre’ls als cicles d’autoclau. Per
desinfectar o tractar el material d’ús comú es van fer servir autoclaus secs.
6.1. Material biològic.
6.1.1. Material bacterià.
6.1.1.1. Soques bacterianes.
En aquest treball s’han utilitzat les següents soques bacterianes: - DH5: Genotip F- endA1 glnV44 thi-1 recA1 relA1 gyrA96 deoR nupG Φ80dlacZ∆M15
∆(lacZYA-argF)U169, hsdR17(rK- mK
+), λ–. Soca d’Escherichia coli utilitzada per als clonatges de vectors.
- TOP10: Genotip F- mcrA ∆(mrr-hsdRMS-mcrBC) φ80lacZ∆M15 ∆lacX74 nupG recA1 araD139 ∆(ara-leu)7697 galE15 galK16 rpsL(StrR) endA1 λ- . Soca d’E. coli utilitzada per als clonatges de vectors.
- BL21 pLys: Genotip F- ompT gal dcm lon hsdSB(rB- mB
-) λ(DE3) pLysS(cmR). Soca d’E. coli utilitzada per a la producció de proteïna.
- BL21 pGROE: Genotip B F- dcm ompT hsdS(rB- mB-) gal [malB+]K-12(λS). Soca d’E. coli
utilitzada per a la producció de proteïna. - C1C58/GV2260: Soca d’Agrobacterium tumefaciens utilitzada a la transformació de
plantes amb vectors binaris.
6.1.1.2. Condicions de cultiu de bacteris.
El cultiu líquid de cèl·lules d’E.coli es realitza en tubs de cultiu a 37ºC en agitació a
250 rpm, mentre que el cultiu sòlid es realitza en placa de petri a 37ºC. El cultiu líquid de
cèl·lules d’A.tumefaciens es realitza en tubs de cultiu a 28ºC en agitació a 250 rpm. El cultiu
sòlid es realitza en placa a 28ºC.
Els cultius o soques bacterianes s’emmagatzemen permanentment en stabs cel·lulars
fent servir cultius crescuts O/N amb el 25% de glicerol i guardats a -80ºC.

6 Materials i mètodes
106
6.1.1.3. Obtenció de cèl·lules competents per xoc tèrmic.
El protocol que es descriu a continuació el vam utilitzar per obtenir les cèl·lules
competents per xoc tèrmic d’E.coli. Convé que les cèl·lules de partida siguin recents i
vigoroses, per la qual cosa vam refrescar les cèl·lules fent un cultiu en placa i fent créixer
aquest cultiu en líquid.
Procediment:
1. Inocular 1 colònia aïllada de la soca utilitzada en 50 mL de medi LB.
2. Incubar en agitació (300 rpm) a 37ºC fins que la D.O.600= 0.5 ± 0.1 (2-3h).
3. Refredar el cultiu en gel durant 10 min.
4. Centrifugar el cultiu 10 min. a 4000xg a 4ºC.
5. Descartar el sobrenedant i resuspendre les cèl·lules en 15 mL de TBF1.
6. Incubar en gel durant 10 min.
7. Centrifugar 10 min. a 4000xg a 4ºC.
8. Descartar el sobrenedant i invertir el tub durant 1 min., per tal d’eliminar totalment les restes del tampó TBF1.
9. Resuspendre el sediment en 2 mL de TBF2 fred.
10. Aliquotar les cèl·lules en fraccions de 100 µL en tubs eppendorf estèrils i congelar-les
en neu carbònica. Guardar-les a -80ºC (es poden guardar durant més d’1 any).
És recomanable testar les cèl·lules competents abans d’utilitzar-les transformant 10ng de
plàsmid control i sembrant diferents dilucions per determinar-ne l’eficiència.
Solucions:
- Tampó TBF1: KAc 30 mM pH 5.8 (equilibrar amb acètic 2M), MnCl2 50 mM, CaCl2 10
mM, RbCl 100 mM, glicerol 15 % (v/v). S’esterilitza per filtració.
- Tampó TBF2: MOPS pH 7.0 10 mM (equilibrar amb NaOH 10M), CaCl2 75 mM, RbCl
10 mM, glicerol 15 % (v/v). S’esterilitza per filtració.
6.1.1.4. Obtenció de cèl·lules electrocompetents.
El protocol que es descriu a continuació el vam utilitzar per obtenir les cèl·lules
electrocompetents d’A.tumefaciens. De nou convé que les cèl·lules de partida siguin recents i
vigoroses, per la qual cosa es recomana refrescar les cèl·lules fent-les créixer en placa i
posteriorment fent un cultiu líquid.
Procediment:
1. Créixer O/N un cultiu de la soca C1C58 GV2260 d’A.tumefaciens en medi YEB amb rifampicina i carbenicil·lina, en agitació a 28ºC.
2. Inocular un cultiu de 200 mL de medi amb 2 mL (1/100 volums) del cultiu crescut O/N i incubar a 28ºC en agitació fins que la D.O.600= 0.5-0.8 (aproximadament unes 2.5-3h).
3. Refredar el cultiu en gel de 15 a 30 min.

6 Materials i mètodes
107
4. Centrifugar 20 min. a 4000xg a 4ºC i descartar el sobrenedant.
5. Resuspendre les cèl·lules en 1 volum del cultiu inicial d’aigua estèril refredada en gel. Anar en compte de no lisar les cèl·lules.
6. Centrifugar 20 min. a 4000xg a 4ºC i descartar el sobrenedant.
7. Repetir els passos 5 i 6, resuspenent les cèl·lules en ½ volum del cultiu inicial d’aigua estèril refredat en gel.
8. Resuspendre les cèl·lules en 1/50 volum del cultiu inicial en glicerol 10% refredat en gel.
9. Centrifugar 20 min. a 4000xg a 4ºC i descartar el sobrenedant.
10. Resuspendre les cèl·lules en 1/100 volums del cultiu inicial de glicerol 10% refredat en gel. La concentració de cèl·lules ha de ser 1-3 x 1010 cèl·lules/mL.
11. Fer alíquotes de 50 µL, congelar en neu carbònica i guardar a -80ºC.
6.1.1.5. Medis de cultiu bacterians.
Els medis utilitzats rutinàriament per al treball amb bacteris en aquesta tesi van ser:
- Medi LB pH 7.5: bactotriptona 100 g/L, extracte de llevat 5 g/L i NaCl 10 g/L. Ajustar el
pH amb NaOH. Per al medi sòlid s’afegeix 15 g/L d’agar. S’esterilitza en autoclau.
- Medi YEB pH 7.2: extracte de carn 5 g/L, extracte de llevat 1 g/L, peptona 5 g/L,
sacarosa 5 g/L i 2 mL/L MgSO4 1 M. Per al medi sòlid s’afegeix 15 g/L d’agar. S’esterilitza en
autoclau.
6.1.1.6. Suplements dels medis de cultiu bacterians.
Els suplements o antibiòtics van ser afegits als medis bacterians després de l’autoclau,
just abans de plaquejar o inocular els medis. A la següent taula (M.1.), s’indiquen les
concentracions de les solucions estoc i el dissolvent en el que es preparen i les concentracions
de treball.
E.coli A. tumefaciens A.thalianaAmpicil·lina (Amp) Carbenicil·lina (Cb)
100 mg/mL Etanol 50% 100 µg/mL 100 µg/mL - -
Kanamicina (Km) 50 mg/mL Aigua 25 µg/mL 25 µg/mL 25 µg/mLIPTG 23,8 mg/mL Aigua 23,8 µg/mL - -X-Gal 20 mg/mL DMF 40 µg/mL - -
Rifampicina (Rf) 50 mg/mL DMSO - 100 µg/m -Higromicina (Hyg) 50 mg/mL Aigua - - 30 µg/mLCefotaxima (Cf) 100 mg/mL Aigua - - 100 µg/mL
Concentració de treballAntibiòtic Concentració
estocDissolvent
Taula M.1. Antibiòtics utilitzats en el treball amb els bacteris E.coli i A.tumefaciens i amb plantes d’A. thaliana.
6.1.2. Material vegetal.
6.1.2.1. Ecotips silvestres.
En aquest treball s’han emprat plantes d’Arabidopsis thaliana dels ecotips Columbia-0
(Col-0), Landsberg erecta (Ler) i Wassilewskija-2 (Ws-2). La majoria de mutants i plantes sobre-

6 Materials i mètodes
108
expressores són de l’ecotip Col-0, només les que s’especifica al contrari estan en altres
ecotips.
6.1.2.2. Línies transgèniques.
Les línies transgèniques usades en aquest treball estan especificades juntament amb les
construccions, en l’apartat 6.2.2., en la Taula M.5.
6.1.2.3. Línies mutants de pèrdua de funció.
Els mutants que procedeixen de la col·lecció SALK s’han generat per inserció del T-DNA
del vector pROK2 amb el gen NPTII de resistència a Km en el genoma d’A. thaliana, en fons
genètic Col-0. Han estat generades en el laboratori del Dr. J. Ecker al SIGnAL (Salk Institute
Genomic Analisis Laboratory, http://signal.salk.edu/index.html).
Nom al·lel Nom Lab. Ecotip Codi SALK Gen Codi Atg dra2-1 CP6L Ws-2 Produïda al laboratori DRA2 At1g10390dra2-2 S62 Col-0 SALK_017077 DRA2 At1g10390
dra2-3 S63 Col-0 SALK_067219 DRA2 At1g10390
dra2-4 S83 Col-0 SALK_015016 DRA2 At1g10390dra2-5 S84 Col-0 SAIL_663_D07 DRA2 At1g10390dra2-6 S101 Col-0 SALK_103803 DRA2 At1g10390dral-2 S85 Col-0 SALK_144099 DRAL At1g59660
nup54-1 S71 Col-0 SALK_015252 NUP54 At1g24310nup54-2 S72 Col-0 SALK_106346 NUP54 At1g24310
nup58-1 tcu1-1 LerCedida per Dra. M. R.
PonceNUP58/TCU1 At1g59660
nup58-2 S64 Col-0 SALK_099638 NUP58/TCU1 At4g37130nup58-3 S65 Col-0 SALK_023793 NUP58/TCU1 At4g37130nup58-4 S66 Col-0 SAIL_655_C09 NUP58/TCU1 At4g37130nup62-1 S69 Col-0 SALK_071950 NUP62 At2g45000nup62-2 S70 Col-0 SALK_037337 NUP62 At2g45000nup62-3 S82 Col-0 SAIL_27_F01 NUP62 At2g45000nup96-1 sar3-1 Col-0 Cedida per M. Estelle SAR3/NUP96 At1g80680
nup96-3 sar3-3 Col-0 Cedida per M. Estelle SAR3/NUP96 At1g80680
nup160-4 sar1-4 Col-0 Cedida per M. Estelle SAR1/Nup160 At1g33410
phyB-9 phyB Col-0 Cedida per Dr. P. Quail PHYB At2g18790
sav3-5 S61 Col-0 SALK_022743 SAV3/TAA1 At1g70560 Taula M.2. Línies mutants.
6.1.2.4. Creuaments.
Els creuaments entre dues línies es van realitzar pol·linitzant els ovaris (♀) de flors
emasculades (flors a les que se’ls ha eliminat els estams) d’una línia amb el pol·len d’una altra
(♂). S’ha de procurar escollir els ovaris que estiguin en un estadi receptiu a la fecundació però
que no hagin estat pol·linitzats amb anterioritat (per això s’escullen flors joves, no obertes i amb
estams no desenvolupats). Si la fecundació es produeix amb èxit i es desenvolupa una silíqua,
aquesta es deixa assecar i es recullen les seves llavors (generació F1 del creuament).
L’autofecundació d’individus F1 dóna lloc a la generació F2, i successivament, l’autofecundació
d’individus F2 a la generació F3.

6 Materials i mètodes
109
Nom creuament
Planta mare Planta pare Objectiu
CA9 Col-0 dra2-1 2n Canvi d'ecotip, mutants dra2-1 en Col-0
CA19 pCS19 nup96-1 Doble mutant ATHB4-GR amb nup96-1
CMG1 phyB-9 dra2-3 Doble mutant phyB -dra2-3
CMG3 dra2-2 Col Retrocreuament (per netejar)
CMG5 Col-0 dra2-3 Retrocreuament (per netejar)
CMG6 Col-0 dra2-4 Retrocreuament (per netejar)
CMG7 Col-0 dra2-5 Retrocreuament (per netejar)
CMG8 dra2-2 PBL Canvis d'ecotip, mutants DRA2 en PBL
CMG9 dra2-3 PBL Canvis d'ecotip, mutants DRA2 en PBL
CMG10 dra2-4 PBL Canvis d'ecotip, mutants DRA2 en PBL
CMG12 dra2-5 PBL Canvis d'ecotip, mutants DRA2 en PBL
CMG13 dra2-2 dral-2 Doble mutant dra2 - dral
CMG14 dra2-3 dral-2 Doble mutant dra2 - dral
CMG15 dra2-4 dral-2 Doble mutant dra2 - dral
CMG18 dra2-4 dra2-1 Test al·lelisme (barreja ecotips)
CMG19 dra2-3 dra2-1 Test al·lelisme (barreja ecotips)
CMG20 dra2-4 nup58-2 Doble mutant dra2 - nup58
CMG21 dra2-5 nup58-2 Doble mutant dra2 - nup58
CMG22 dra2-4 nup58-4 Doble mutant dra2 - nup58
CMG23 dra2-5 nup58-4 Doble mutant dra2 - nup58
CMG24 dra2-2 nup58-2 Doble mutant dra2 - nup58
CMG25 dra2-3 nup58-2 Doble mutant dra2 - nup58
CMG30 CMG9 PBL 2n Canvi d'ecotip, mutants DRA2 en PBL
CMG31 CMG10 PBL 2n Canvi d'ecotip, mutants DRA2 en PBL
CMG32 sav3-1 nup160-4 Doble mutant sav3 - nup160
CMG34 Col-0 CA9.10S 3r Canvi d'ecotip, mutants dra2-1 en Col-0
CMG38 nup96-3 dra2-4 Doble mutant dra2 - nup96
CMG48 CMG30 PBL 3r Canvi d'ecotip, mutants DRA2 en PBL
CMG49 CMG31 PBL 3r Canvi d'ecotip, mutants DRA2 en PBL
CMG50 Col-0 CMG34 4t Canvi d'ecotip, mutants dra2-1 en Col-0
CMG51 Ws-2 dra2-1 Neteja del trangen PBL
CMG55 Ws-2 CMG34 Recuperació fenotip original dra2-1
CMG56 PBL CMG49 4t Canvi d'ecotip, mutants DRA2 en PBL
CMG57 CMG49 dra2-1 Test al·lelisme (ecotip PBL)
CMG58 MG47 dra2-1 Doble mutant PAR1-GR amb dra2-1
CMG59 MG49 dra2-1 Doble mutant ATHB4-GR amb dra2-1
CNE1 nup58-2 nup96-3 Doble mutant nup58 - nup96CNE4 nup58-2 nup54-2 Doble mutant nup58 - nup54
Taula M.3. Creuaments usats en el present treball.
6.1.2.5. Condicions de cultiu a l’hivernacle i al fitotró.
Les plantes han estat cultivades en testos amb una barreja a proporcions 3:1:1 de torba,
perlita i vermiculita respectivament. Les condicions de cultiu a l’hivernacle han estat de 22 ±
2ºC, una humitat ambiental del 55%, i 14h de llum i 10h de foscor, aproximadament. En la
càmera visitable de SD les condicions han estat de 22-24ºC, sense control d’humitat i 8h de
llum i 16h de foscor.
En ambdós llocs les plantes han estat regades per inundació de les safates que
contenen els testos 2-3 cops per setmana, segons les necessitats de les plantes. Dels 3 regs, 2
són amb aigua normal i un tercer amb una solució de fertirrigació molt diluïda (1:240).
Solucions:
- Solució de fertirrigació: Solució de Hoagli i Arnon (Arnon 1938). NO3K 8.4 mM, NH4NO3
1.2 mM, K2HPO4 1.2 mM, KH2PO4 1.2 mM, Ca(NO3)2.4H2O 2.5 mM, MgSO4.7H2O 0.7 mM,

6 Materials i mètodes
110
SO4Fe.7H2O 0.6 mM. Aquesta solució es complementa amb el quelant de ferro Kelamix 35
mg/L i amb microelements 0.4 g/L.
6.1.2.6. Condicions de cultiu in vitro.
En condicions in vitro, les plàntules es van cultivar sota llum contínua o cicles de dia llarg
(LD). Les condicions de cultiu en llum contínua s’especifiquen més endavant quan s’expliquen
els diferents tractaments. Les condicions de cultiu en LD van ser de 14h llum i 10h foscor a 22-
24ºC de Tª, i humitat no controlada
6.1.2.7. Medis de cultiu de plantes.
El medi Murashige i Skoog (medi MS) va ser utilitzat per germinar llavors i créixer les
plantes en plaques de Petri i en condicions estèrils. En general els experiments s’han realitzat
en el medi indicat com a 0,5MxS- (amb vitamines). Només en les ocasions que s’indica
especialment s’ha afegit sucrosa al 1 o 2% (p/v) al medi.
- Medi 0.5MS- pH 5.8: Murashige & Skoog medium including vitanims 2.15 g/L, MES 0.25
g/L. Ajustar el pH amb KOH. S’afegeix 8 g/L d’agar pels medis sòlids. S’esterilitza en autoclau.
6.1.2.8. Suplements dels medis de cultiu de plantes.
Els suplements que s’han afegit als medis bacterians han estat antibiòtics i hormones. A
la taula M.1 es mostren els diferents antibiòtics utilitzats, les concentracions de les solucions
estoc i el dissolvent en el que es preparen, i les concentracions de treball. Les hormones i
reguladors del creixement es detallen a l’apartat 6.4.4.6., quan es descriuen els diferents
tractaments hormonals duts a terme.
6.2. Plàsmids.
6.2.1. Vectors comercials.
Plàsmid Descripció (gen) BinariResistència
bacteriResistència
planta
pBinAr Vector binari que permet clonar un gen d’interès sota el promotor constitutiu 35S. Binari KmR KmR
pCAMBIA1302Vector binari que permet clonar un gen d’interès fusionat al gen reporter GFP sota
el promotor constitutiu 35S.Binari KmR HygR
pCAMBIA1303Vector binari que permet clonar un gen d’interès fusionat als gens reporters GUS-
GFP sota el promotor constitutiu 35S.Binari KmR HygR
pCAMBIA2300 Vector binari que permet clonar un gen d’interès sota el promotor constitutiu 35S. Binari KmR HygR
pCRII-TOPO (Invitrogen)
Vector utilitzat pel clonatge de productes de PCR pel sistema T/A cloning. AmpR, KmR
pDs-RedVector que sobre-expressa constitutivament la proteïna vermella fluorescent
(RFP), usat com a control de transformació i localització citosòlica en microbombardeig. Cedit per la Dra. Montse Pagès.
AmpR
pGEX Vector usat per produir una proteïna d’interès fusionada a la GST. AmpR
pGI0029/35S/GRVariant del vector pGreen que permet clonar un gen d’interès fusionat al receptor
de glucocorticoides (GR) sota el promotor constitutiu 35S.Binari KmR HygR
pMALc2x Vector usat per produir una proteïna d’interès fusionada a la MBP. AmpR
Taula M.4. Vectors comercials emprats en aquest treball.

6 Materials i mètodes
111
6.2.2. Construccions i transformacions en planta.
Plàsmid Descripció (gen) Vector Insert BinariEcotip
transformatpBJ3 35S: ATHB4-GFP pCAMBIA1302 pCS12 Binari Col-0
pCS19 35S: ATHB4-GRpCS14 (derivat pCAMBIA1300) pCS13 Binari Col-0
pMG15 35S:GRpCS14 (derivat pCAMBIA1300) pGreen0029:35SGR Binari
pMG16 Nt HD ZIP (ATHB4) pCRII-TOPO PCR insert JO284+MGO3pMG17 HD ZIP (ATHB4) pCRII-TOPO PCR insert MGO1+MGO3pMG18 ZIP (ATHB4) pCRII-TOPO PCR insert MGO2+MGO3 pMG19 ZIP Ct (ATHB4) pCRII-TOPO PCR insert MGO2+CSO7pMG20 HD ZIP Ct (ATHB4) pCRII-TOPO PCR insert MGO1+CSO7pMG21 Nt HD (ATHB4) pCRII-TOPO PCR insert JO284+MGO4pMG22 HD (ATHB4) pCRII-TOPO PCR insert MGO1+MGO4pMG23 35S: Nt HD ZIP-GR (ATHB4) pMG15 pMG16 Binari Col-0pMG24 35S: HD ZIP-GR (ATHB4) pMG15 pMG17 Binari Col-0pMG25 35S: ZIP-GR (ATHB4) pMG15 pMG18 Binari Col-0pMG26 35S: ZIP Ct-GR (ATHB4) pMG15 pMG19 Binari Col-0pMG27 35S: HD ZIP Ct-GR (ATHB4) pMG15 pMG20 Binari Col-0pMG28 35S: Nt HD-GR (ATHB4) pMG15 pMG21 Binari Col-0pMG29 35S: HD-GR (ATHB4) pMG15 pMG22 Binari Col-0pMG30 Sonda de Northerns amb cDNA (DRA2) pCRII-TOPO PCR amb GO76+GO77
pMG31 Sonda de Northerns amb cDNA (DRA2like) pCRII-TOPO PCR amb GO88+GO89pMG32 Sonda de Northerns amb cDNA (Nup54) pCRII-TOPO PCR amb GO80+GO83pMG33 Sonda de Northerns amb cDNA (Nup62) pCRII-TOPO PCR ambGO86+GO87
pMG34Sonda de Northerns amb gDNA (tcu1/Nup58) pCRII-TOPO PCR amb GO78+GO79
pMG35 ATHB4 amb HD mutat (Asn210Ala) pCRII-TOPO PCRs, MGO20 i MGO21pMG36 ATHB4 amb Nt mutat (Trp80Ala) pCRII-TOPO PCRs, MGO22 i MGO23pMG37 35S: ATHB4 HD mutat (Asn210Ala) pMG15 pMG35 Binari Col-0pMG38 35S: ATHB4 Nt mutat (Trp80-Ala) pMG15 pMG36 Binari Col-0pMG39 35S:ATHB4-GR pBinAr pCS13 Binari PBLpMG40 HSP18.2:GUS pTT101 pMN4 Binari PBLpMG41 ATHB4 amb Nt mutat (Leu10Ala) pCRII-TOPO PCRs, MSO36 i MSO37pMG42 N(parcial) HD ZIP Ct (ATHB4) pCRII-TOPO PCR amb MGO50+CSO7pMG43 35S: ATHB4 Nt mutat (Leu10Ala) pMG15 pMG41 Binari Col-0pMG44 35S: N(parcial) HD ZIP Ct (ATHB4) pMG15 pMG42 Binari Col-0pMG46 35S:ATHB4-MBP pMAL-c2x pCS12MG47 35S:PAR1-GR pIR33 - Binari Ws-2MG49 35S:ATHB4-GR pMG39 - Binari Ws-2MG50 HSP18.2:GUS pMG40 - Binari Col-0pMG51 pRSETA Hist:GR pRSET A pGreen0029:35SGR
pMG52Nt DRA2 (domini repeticions FG, primers 2340pb) pCRII-TOPO PCR insert CTO6+MGO36
pMG53 Ct DRA2 (últims 786pb) pCRII-TOPO PCR insert MGO37+CTO8pMG54 Nt DRA2 (domini repeticions FG) pCAMBIA1302 pMG52pMG55 Ct DRA2 pCAMBIA1302 pMG53 Binari Col-0pMG56 Nt DRA2 (domini repeticions FG) pMG54 pCT6 Binari Col-0MG58 35S: HD ZIP Ct-GFP (ATHB4) pMS56 - Binari Col-0pMG59 Nt (ATHB4) pCRII-TOPO PCR insert 2x35S+MGO51pMG60 35S: Nt-GR (ATHB4) pMG15 pMJ32 Binari Col-0pMG61 35S: Nt-GFP (ATHB4) pMS51 pMJ32 BinariMG62 35S: Nt HD-GFP (ATHB4) pMS57 - Binari Col-0pMS51 35S:GFP pCAMBIA1302 PCR amb MSO44+MSO45 Binari
Taula M.5. Construccions emprades en aquest treball i transformacions en planta.
6.3. Encebadors.

6 Materials i mètodes
112
Nom Seqüència Gen Ús2x35S 5’-CGTAAGACTGGCGAAC-3’ Promotor CaMV 35S Genotipats
35S 5'-CTTCGCAAGACCCTTCC-3' Promotor CaMV 35S Genotipats
BO33 5'-GCGGTCTATGTAGGAGAGAATGATC-3' SAUR15 (F) qPCR
BO34 5'-CCGGCACCACATATCTCTTCTT-3' SAUR15 (Rev) qPCR
BO35 5'-AAACGCAAAGAAGCTTATGAAGATG-3' SAUR68 (F) qPCR
BO36 5'-GCTGCTCTTTGTTGCCATTTC-3' SAUR68 (Rev) qPCR
BO40 5’-AAATCTCGTCTCTGTTATGCTTAAGAAG-3’ UBQ10 (F) qPCR
BO41 5’-TTTTACATGAAACGAAACATTGAACTT-3’ UBQ10 (Rev) qPCR
BO68 5'-AGAAGTGAGAGAGAAGGAATCTCCG-3' HAT2 (F) qPCR
BO69 5'-TCATCTGAGGTTCCACGTGAGTA-3' HAT2 (Rev) qPCR
BO70 5'-GGATCTAGAATGGGCGAGCTCCCAGAAGCTCG-3' GR (F) Genotipats
B071 5'-CCTACTAGTTTTTTGATGAAACAGAAGCTTTTTG-3' GR (Rev) Genotipats
BO87 5’-GGAAGCAAAACCCTTAGCATCAT-3’ PIL1 (F) qPCR
BO88 5’-TCCATATAATCTTCATCTTTTAATTTTGGTTTA-3’ PIL1 (Rev) qPCR
BO89 5’-GATGCGTAAGCTACAGCACTCGT-3’ HFR1 (F) qPCR
BO90 5’-AGAACCGAAACCTTGTCCGTCTTG-3’ HFR1 (Rev) qPCR
BO93 5'-CGGCTTGCACAGCCTCTT-3' XTR7 (F) qPCR
BO94 5'-TCGGTTGCCACTTGCAATT-3' XTR7 (Rev) qPCR
CSO7 5'-GGGGATCCGCGACCTGATTTTTGCTG-3' ATHB4 (Rev) Clonatge
CTO4 5’-GGCCATGGTTGGCTCATCTAATCCTTTTG-3’ DRA2 (F) Clonatge
CTO5 5’-GGGCTAGCAATTGTTGGGGTTTGAG-3’ DRA2 (Rev) Clonatge
CTO6 5’-GGACTAGTTCGAGTTTTGGAACGG-3’ DRA2 (F) Clonatge
CTO8 5´-GGTCTAGAAACTCCATCTTCTTCATCTTCGTCGC-3´ DRA2 (Rev) Clonatge
GO74 5’-CACTGATGACGAAGAGAG-3’ DRA2 (F) Genotipats
GO75 5’-CCATAACCGTGTCGTCCC-3’ DRA2 (Rev) Genotipats
GO76 5’-GATCTTCTGGTTTTGGGCAG-3’ DRA2 (F) Genotipats
GO77 5'-CATTGTTTGTCCAAAGGGAG-3' DRA2 (Rev) Genotipats
GO78 5’-CCAAATTTGTTAAAATGTG-3’ NUP58 (F) Genotipats
GO79 5’-ACGATATACTCCACAAAC-3’ NUP58 (Rev) Genotipats
GO80 5’-CCAATGTTCGGCACTCCG-3’ NUP54 (F) Genotipats
GO81 5’-CATCTGATACAGCTGCAGGC-3’ NUP54 (Rev) Genotipats
GO82 5’-CTTCAGAGACATTTGCAAGC-3’ NUP54 (F) Genotipats
GO83 5’-CTATGAGTCTAGTGCCATTTC-3’ NUP54 (Rev) Genotipats
GO84 5’-GATTATCAAGGAGTGGAATAC-3’ NUP62 (F) Genotipats
GO85 5’-CATTGCATCTCTAGTTGATAC-3’ NUP62 (Rev) Genotipats
GO96 5’-ATACGCCCAGTTCAACAGTGG-3’ DRA2 (F) Genotipats
GO97 5’-ATACGCCCAGTTCAACAGTGA-3’ DRA2 (Rev) Genotipats
GO104 5’-CAATGTTGTTGATGCAGCATT-3’ SAR3-1 wt (Rev) Genotipats
GO105 5’-CAATGTTGTTGATGCAGCATA-3’ SAR3-1 (mut F) Genotipats
GO106 5’-TTCACATCCTGCATCACGTC-3’ SAR3-1 (F) Genotipats
GO107 5’-GTAGAACTGGTATGTCTACGT-3’ SAR3-3 (Rev) Genotipats
GO108 5’-CTGTTTTACTAAGCTGAGATTTGG-3’ SAR3-3 (F) Genotipats
JO284 5'-AGGACAATGGGGGAAAGAGAT-3' ATHB4 (F) Clonatge
JO402 5’-GGTCGAAGAACGTGTGTCC-3’ DRA2 (F) Clonatge
JO403 5’-GGTACCAGATGACTGTCC-3’ DRA2 (Rev) Clonatge
LB3 5'-TAGCATCTGAATTTCATAACCAATCTCGATACAC-3' T-DNA left border Genotipats
LBb1 5’-CGTGGACCGCTTGCTGCAACT-3’ T-DNA left border Genotipats
MGL1 5'-GCTAATGGTGCTTGAGATCG-3’ HAT2 (F) ChIP qPCR
MGL2 5'-CGCAAGATAGACGTGAACAG-3’ HAT2 (Rev) ChIP qPCR
MGL5 5'-GCATGTGGGTCATAACAG-3’ HAT2 (F) ChIP qPCR
MGL6 5'-GTAGGCAGCAGTTAAAGG-3’ HAT2 (Rev) ChIP qPCR
MGL7 5'-CTGACCAAGTGCGTTATATC-3’ HAT2 (F) ChIP qPCR
MGL8 5'-ATCATGCTTTGCGGACTCTC-3’ HAT2 (Rev) ChIP qPCR
MGL9 5'-AATTGTCCTCTCACATGTACTG-3’ HAT2 (F) ChIP qPCRMGL10 5'-GTGATGCAAACCTCAAATTC-3’ HAT2 (Rev) ChIP qPCRMGL13 5'-TGGCCCACAACTTTGAATG-3’ HAT1 (F) ChIP qPCRMGL14 5'-CACGGATGAGAACAGATATCG-3’ HAT1 (Rev) ChIP qPCRMGL17 5'-GGGCTATTTAGCCGATCAGG-3’ bHLH129 (F) ChIP qPCRMGL18 5'-TGGGCCAACTCTGTACTC-3’ bHLH129 (Rev) ChIP qPCRMGL21 5'-CAACTCTGGCTTCTACAC-3’ bHLH129 (F) ChIP qPCRMGL22 5'-GGTCCACAAGTTGTATGC-3’ bHLH129 (Rev) ChIP qPCRMGL23 5'-CATTCATCATCGCTTCTTG-3’ bHLH129 (F) ChIP qPCRMGL24 5'-TTGGTCTACGTCATGTTTG-3’ bHLH129 (Rev) ChIP qPCRMGL25 5'-TCTTGATTAGTTACGGCATACC-3’ HAT2 (F) ChIP qPCRMGL26 5'-TTGTACCAACCACTCCAATTAG-3’ HAT2 (Rev) ChIP qPCRMGL27 5'-GCGGTTGTATGGTCAGATG-3’ HAT1 (F) ChIP qPCRMGL28 5'-GATCGCTGGTTGTGGTTAG-3’ HAT1 (Rev) ChIP qPCRMGL29 5'-CAGAAGCTAAGGCTGCT-3’ HAT1 (F) ChIP qPCR
Taula M.6. Encebadors utilitzats en aquest treball. Es mostren tant els encebadors utilitzats en la generació de
fragments per clonatge , com per ser utilitzats com a sonda, per qPCR o els comunament utilitzats per comprovar
clonatges.

6 Materials i mètodes
113
6.4. Metodologies.
6.4.1. Tècniques de biologia molecular d’àcids nucleics.
6.4.1.1 Obtenció de DNA plasmídic.
En aquest treball s’han emprat 2 mètodes d’extracció de DNA plasmídic depenent del
grau de puresa desitjat.
Minipreparació mitjançant lisi alcalina.
L’extracció de DNA plasmídic s’ha realitzat per lisi alcalina segons el mètode descrit per
Bimboin (Bimboin and Doly 1979), amb modificacions posteriors de Sambrook (Sambrook and
Russell 2001). Aquest mètode s’ha utilitzat en passos intermedis de clonatges i per a la
comprovació d’aquests mitjançant digestió o PCR.
Procediment:
1. Inocular una colònia aïllada en 3 mL de cultiu amb l’antibiòtic adient, incubar a 37ºC en agitació a 250 rpm O/N.
2. Transferir tot el cultiu a un tub eppendorf i centrifugar 1 min. a velocitat màxima ( 13000 rpm). Descartar el sobrenedant.
3. Afegir 150 µL de tampó P1. Resuspendre bé el sediment de bacteris amb l’ajuda del vòrtex.
4. Afegir 150 µL del tampó P2. Agitar suaument el tub per inversió unes 4-5 vegades. Incubar no més de 5 min. a TªA.
5. Afegir 150 µL del tampó P3 i barrejar immediatament per inversió unes 4-5 vegades. Deixar reposar 15-20 min. en gel. Centrifugar 10 min a velocitat màxima ( 13000 rpm).
6. Passar el sobrenedant a un nou tub eppendorf, afegint-hi 900 µL d’etanol 100%. Incubar 2 min. a TªA. Centrifugar 10 min. a velocitat màxima ( 13000 rpm).
7. Descartar el sobrenedant i afegir 700 µL d’etanol 70%. Resuspendre el possible pellet amb vòrtex. Centrifugar 5 min. a velocitat màxima ( 13000 rpm).
8. Decantar l’etanol i deixar assecar el DNA fins que s’evapori tot l’etanol (uns 10 min.).
Resusprendre el DNA en 50 µL d’aigua. Guardar a 4ºC.
Solucions:
- Tampó P1 pH 8.0 (guardar a 4ºC): Tris-HCl 50mM, EDTA 10 mM, RNasa A 100µg/mL. - Tampó P2 pH 8.0 (guardar a TªA): 200 mM NaOH, SDS 1%. - Tampó P3 pH 5.5 (guardar a 4ºC): KAc 3M. Ajustar el pH amb àcid acètic. - Tampó TE 1x pH 7.4: Tris-HCl pH 7.5 10mM, EDTA (pH 8.0) 1mM.
Minipreparació de DNA plasmídic d’alta qualitat
Per obtenir DNA d’alta qualitat (per exemple per seqüenciació) es va fer servir el kit
QIAprep Spin Miniprep de Qiagen, seguint les indicacions del fabricant. Per obtenir quantitats

6 Materials i mètodes
114
grans de vector necessaris pels experiments de bombardeig es va fer servir el kit comercial
QIAprep Spin Midiprep de Qiagen, seguint les indicacions del fabricant.
6.4.1.2. Reaccions de modificació del DNA.
Per a les reaccions de modificació de DNA realitzades, com ara la digestió de fragments
de DNA amb enzims de restricció, lligacions, desfosforilació d’extrems de DNA, emplenat
d’extrems protuberants, etc. s’han utilitzat diferents enzims de restricció de DNA i ligases
(Promega, Roche, etc.), seguint els protocols estàndards i les condicions aconsellades pel
fabricant en cada cas. Generalment s’ha partit d’uns 2µg de plàsmid per iniciar un procés de
digestió. En el present treball només es detallen aquells procediments que s’hagin realitzat amb
alguna modificació sobre l’original.
6.4.1.3. Transformació de cèl·lules competents d’E.coli per xoc tèrmic.
La transformació de cèl·lules d’E.coli s’utilitza per amplificar DNA plasmídic. Les cèl·lules
competents, preparades segons es descriu a l’apartat 6.1.1.3, es van transformar per xoc
tèrmic segons el següent protocol.
Procediment:
1. Descongelar en gel el tub que conté una alíquota de 100 µL de cèl·lules competents.
2. Afegir 1-5 µL (10-20 ng) de DNA i barrejar suaument.
3. Mantenir la barreja 30 min. en gel.
4. Fer un xoc tèrmic 1 min. a 42ºC.
5. Transferir el tub a gel durant 1 min.
6. Afegir 900 µL de medi LB fresc sense antibiòtics. Posar el tub a 37ºC en agitació a 250 rpm durant 1h.
7. Plaquejar dos volums, amb com a mínim un ordre de magnitud de diferència, de la transformació en dues plaques de LB sòlid amb l’antibiòtic selectiu escaient.
8. Incubar els bacteris a 37ºC O/N.
Les colònies es seleccionen per PCR utilitzant encebadors específics i/o minipreparacions seguides de digestions.
6.4.1.4. Transformació de cèl·lules competents d’A.tumefaciens per electroporació.
La transformació de cèl·lules d’A.tumefaciens s’utilitza amb plàsmids binaris, que després
s’introduiran a la planta mitjançant transformació per floral dip (apartat 6.4.4.3). Les cèl·lules
competents, preparades segons es descriu a l’apartat 6.1.1.4., es van transformar per
electroporació segons el següent protocol.
Procediment:
1. Descongelar una alíquota (50 µL) de cèl·lules electrocompetents en gel i afegir 1-2 µL de DNA (el DNA ha d’estar dissolt en una solució de baixa força iònica, com per exemple TE, o H2O). Barrejar bé i incubar en gel durant més de 5 min.

6 Materials i mètodes
115
2. Esterilitzar les cubetes d’electroporació (de 0,1 cm) rentant una vegada amb etanol 70% i una amb 100% i deixant assecar en cabina de flux laminar (5-10min.). Refredar-les en gel (5min.) i afegir-hi la barreja de cèl·lules i DNA.
3. Electroporar a 1.7 kV (el temps d’electroporació ha de ser d’uns 5,5ms). Immediatament, transferir les cèl·lules a un tub amb 1 mL de YEB fresc sense antibiòtics.
4. Incubar les cèl·lules a 28ºC en agitació durant 2h per recuperar-les de l’electroporació.
5. Plaquejar dos volums, amb com a mínim un ordre de magnitud de diferència, en medi YEB amb els antibiòtics selectius adients.
6. Incubar a 28ºC uns 2-3 dies fins l’aparició de colònies.
Comprovar les colònies per PCR utilitzant encebadors específics.
6.4.1.5. Reacció en cadena de la polimerasa (PCR).
Per tal d’amplificar fragments de DNA s’ha emprat el mètode de la reacció en cadena de
la polimerasa (polymerase chain reaction o PCR) (Saiki et al. 1989), fent servir oligonucleòtids
específics i DNA codificant (cDNA) o genòmic (gDNA) com a substrat.
Per amplificar els fragments per PCR es van utilitzar dos tipus d’enzims DNA polimerasa
termoestable: un de no comercial, obtingut al departament mitjançant tècniques de producció
de proteïna amb E. Coli, que va ser utilitzat per a la comprovació de clonatges i colònies, i un
de comercial, usat per fer els clonatges. La comercial utilitzada va ser la Platinum® Taq
(Invitrogen) que es caracteritza per tenir una baixa taxa d’error. Aquest enzim disposa dels seu
propi tampó i es va utilitzar seguint les instruccions del fabricant.
Les seqüències dels encebadors emprats s’especifiquen a l’apartat 6.3.
Amplificació per PCR
En cada cicle de PCR hi ha tres etapes que precisen de temperatures diferents i es duen a
terme en l’aparell de PCR o termociclador, el qual permet una transició molt ràpida entre una
temperatura i una altra. Les tres etapes de cada cicle són:
1. Desnaturalització del DNA en presència dels oligonucleòtids encebadors. Es realitza a
una temperatura alta, generalment 94ºC, durant 30 seg.
2. Hibridació dels encebadors a les seqüències complementàries del DNA motlle a
amplificar. Es realitza a la temperatura Tm de l’oligonucleòtid, que ve donada per la seqüència i
la llargada d’aquest. Acostuma a estar compresa entre 45 i 65ºC, prenent com a Tm general
uns 55ºC. El temps d’hibridació és d’uns 30 seg.
3. Un cop els encebadors s’han unit al DNA, actua la DNA polimerasa termoestable,
sintetitzant les cadenes de DNA complementàries en direcció 5’ a 3’. Aquesta etapa es realitza
a 72ºC per l’enzim comú i 68ºC per l’enzim comercial d’alta fidelitat. El temps depèn de la mida
del fragment a amplificar (aproximadament 1 min. per 1 kb de DNA per la polimerasa normal i
uns 2min. per 1kb per la comercial).

6 Materials i mètodes
116
Generalment s’han fet servir 30 cicles d’amplificació, amb una etapa de
desnaturalització inicial a 94ºC durant 5 minuts i una etapa final d’elongació d’uns 5 minuts a la
temperatura òptima de l’enzim (72ºC pel normal). La majoria de PCRs s’han fet en un volum
total de 25µL, amb 0,5µL d’enzim per reacció.
Solucions:
- Tampó PCR 10X: TrisCl 200 mM, KCl 500 mM.
Nota: En el cas de PCR de colònies, el DNA motllo de la PCR s’hi afegeix picant amb una
punta la colònia i submergint la punta a la barreja de PCR, si la colònia és d’E.coli. Per a
colònies d’A.tumefaciens, es pica amb una punta la colònia i es resuspèn en 10 µL d’aigua.
D’aquests, utilitzem 1 µL per fer la PCR.
La comprovació dels fragments amplificats es realitza carregant una part del volum
obtingut en un gel d’agarosa, tal com s’explica a continuació.
6.4.1.6. Separació de DNA en gels d’agarosa.
Els productes de PCR o els fragments de DNA procedents de digestions s’han separat
en gels d’agarosa/TBE amb agarosa 1% (p/v) amb el colorant d’àcids nucleics GelRed de
BIOTIUM (1:10000), en TBE 0,5x, al costat d’un marcador de pes molecular adient per a la
mida de banda esperada.
Solucions:
Tampó de càrrega 6X: glicerol 30% (v/v), xilencianol FF 0.25% (p/v), blau de bromofenol
0.25% (p/v), EDTA 0.5M pH 8.0.
Tampó TBE 10x: Tris base 0.089 M, àcid bòric 0.089 M, EDTA 20 mM pH 8.0.
6.4.1.7. Purificació de DNA.
Els fragments de DNA, tant els resolts en gels d’agarosa/TBE com els productes de PCR
d’interès, s’han purificat utilitzant el kit QIAquick Gel Extraction Kit de Qiagen, seguint les
indicacions del fabricant.
6.4.1.8. Seqüenciació del DNA..
La determinació de les seqüències de DNA purificat amb kit comercial ha estat realitzada
pel Servei de Seqüenciació del CRAG mitjançant seqüenciació automàtica, emprant el sistema
Applied Biosystems de Pharmacia (Automated Laser Fluorencence). La mitjana de lectura amb
aquest mètode va ser de 600-700 pb. Les seqüències obtingudes han estat processades
informàticament mitjançant programes bioinformàtics gratuïts a Internet, com Bioedit o BLAST
de l’NCBI, o també mitjançant el Vector NTI de Invitrogen.
6.4.1.9. Clonatge de fragments de DNA amplificats per PCR.

6 Materials i mètodes
117
Els productes de la reacció de PCR purificats van ser clonats emprant el vector pCRII-
TOPO d’Invitrogen, segons les instruccions del fabricant.
6.4.1.10. Obtenció d’RNA total d’arabidopsis.
L’obtenció d’RNA d’Arabidopsis s’ha fet en tres mètodes diferents depenent el grau de
puresa desitjat. En tots els casos l’aïllament d’RNA es fa a partir de plàntules d’arabidopsis
congelades en nitrogen líquid i emmagatzemades a -80ºC.Mètode per fenol:cloroformAquest
mètode s’ha usat per extraccions on no era necessària una alta puresa dels RNA, com per
exemple pels extractes usats per Northern-blot.
Procediment:
1. Triturar la mostra (100-300 mg) en un morter amb nitrogen líquid fins que quedi una pols fina. La mostra no s’ha de descongelar en cap moment.
2. Passar el teixit triturat a un tub eppendorf de 2 mL prèviament refredat en nitrogen líquid. Afegir 500 µL de tampó d’extracció i 500 µL de fenol:cloroform. Barrejar bé amb el vòrtex i mantenir el tub en gel fins que s’hagin triturat totes les mostres.
3. Centrifugar en microfuga 10 min. a 13000 rpm a 4ºC.
4. Transferir la fase aquosa (superior) a un eppendorf nou de 1.5 mL. Afegir 1 volum de cloroform, barrejar amb el vòrtex.
5. Centrifugar 10 min. a 13000 rpm a 4ºC.
6. Transferir la fase aquosa (superior) a un eppendorf nou de 1.5mL. Afegir 1 volum d’acetat de liti 4M i barrejar amb el vòrtex. Deixar-ho en gel O/N (almenys 4h).
7. Centrifugar 10 min. a 13000 rpm a 4ºC.
8. Eliminar el sobrenedant i resuspendre immediatament el sediment en 300 µL d’aigua estèril i 30 µL d’acetat sòdic 3M.
9. Afegir 2.5 volums d’etanol 100% per precipitar l’RNA.
10. Centrifugar 10 min. a 13000 rpm a 4ºC.
11. Eliminar el sobrenedant i afegir 500 µL d’etanol 70% (v/v).
12. Centrifugar 10 min. a 13000 rpm a 4ºC.
13. Eliminar el sobrenedant i deixar que el sediment s’assequi totalment a TªA.
14. Resuspendre el sediment en 50µL d’aigua estèril. Guardar l’RNA a -20ºC o -80ºC.
Solucions:
- Tampó d’extracció: Tris-HCl 10 mM pH7.5, NaCl 100 mM, EDTA 1 mM. S’esterilitza per autoclau i se li afegeix SDS 1% (p/v). - Acetat de liti 4M : 40.8 g d’acetat de liti en 100 mL d’aigua. S’esterilitza en autoclau. Mètode d’extracció d’RNA d’alta puresa
Pels experiments on era necessari un RNA d’alta qualitat, com per exemple per les PCR
quantitatives, es van fer les extraccions amb dos kits comercials. Inicialment es van fer amb el
sistema manual del Kit RNAeasy de Qiagen, basat en columnes i rentats per centrifugació.
Posteriorment amb el mètode semiautomàtic Maxwell Simply RNA de Promega, basat en

6 Materials i mètodes
118
purificació amb boletes magnètiques i rentats seqüencials. En qualsevol cas es va partir de
mostres biològiques d’unes 30 plàntules congelades (equivalents a uns 100mg de teixit fresc)
en nitrogen líquid i triturades en morter manualment o amb l’aparell Tissue-Lyser de Qiagen, en
aquest darrer cas afegint dues perles de vidre (4mm, casa Merck) a un tub eppendorf de 2mL
amb la mostra, mantenint en nitrogen líquid i removent dues vegades 1min amb una freqüència
de 30s-1.
6.4.1.11. Quantificació d’àcids nucleics.
La quantitat de DNA o d’RNA present en una mostra es va mesurar amb els aparells
Nanodrop del departament de Genètica Molecular del CRAG, fent tres mesures per mostra com
a mínim.
6.4.1.12. Retrotranscripció.
Aquesta tècnica s’ha utilitzat per amplificar la primera cadena de cDNAs a partir d’RNA
total. En el nostre treball s’ha emprat un encebador genèric com l’oligo(dT). Per a la síntesi de
la primera cadena de cDNA s’han utilitzat dos kits comercials, la M-MLV Reverse Transcriptase
d’Invitrogen i el Transcriptor First Strand cDNA synthesis kit de Roche, enzims que permeten la
transcripció de fragments d’RNA de fins a 7 kb en base 1ng- 5µg d’RNA. Es van seguir les
instruccions comercials partint d’uns 2µg d’RNA purificat com s’ha descrit anteriorment (volum
final de 20μL).
6.4.1.13. PCR quantitativa.
Les PCR quantitativa a temps real (qPCR) es va fer utilitzant dilucions 1:20 de cDNA
descrit a l’apartat anterior. Les reaccions es van fer en l’aparell Roche Light Cycler®480 sota el
protocol estàndard per l’enzim propi de la Roche. Es van fer servir tres rèpliques biològiques i
tres rèpliques tècniques per cada mesura i es mostra l’error estàndard calculat a partir de la
mitjana d’aquestes dades. Si no s’especifica el contrari els resultats es van normalitzar respecte
el control wt sense cap tractament i fent servir per normalitzar les mostres quantificacions del
gen UBQ10 com a housekeeping.
Les reaccions van fer-se en dues possibles quantitats segons el volum final:
10 μL Roche Light Cycler®480 SYBR Green I master MIX, 300 nM de cada
oligonucleòtid, 2 μL de dilució de cDNA, en 20 μL de reacció.
5 μL Roche Light Cycler®480 SYBR Green I master MIX, 300 nM de cada oligonucleòtid, 2 μL de dilució de cDNA, en 10 μL de reacció.
La quantificació i l’anàlisi de les dades es va fer amb el propi programa Roche Light
Cycler® 480 (versió 1.5.0SP3) i amb Microsoft Excel.
6.4.1.14. Immunoprecipitació de cromatina (ChIP).
La immunoprecipitació de cromatina és una tècnica per detectar punts d’unió d’una
proteïna en la cromatina, fent servir una immunoprecipitació de la proteïna unida al DNA.
Aquesta tècnica requereix diversos passos que han d’estar optimitzats. El més important és

6 Materials i mètodes
119
l’anticòs que es fa servir per la immunoprecipitació, però cal testejar abans passos com la
sonicació. El protocol que es va fer servir és original de Jose L. Pruneda-Paz i modificat per
Patricia Hornitschek (Hornitschek, et al. 2012).
Material vegetal:
Es van sembrar 60 mg de llavors, es van deixar créixer en llum blanca contínua en medi
0,5xMS- i sobre paper de filtre durant 7 dies. Amb aquest material es poden obtenir dues
mostres d’1g de material vegetal fresc i una tercera mostra amb menor quantitat de material.
Fixació (<1h):
En aquest primer pas el que es fa és reforçar la unió entre la proteïna i el DNA, per tal de
poder purificar la cromatina amb les proteïnes unides a ella (procés anomenat cross-link).
Abans de recollir el material és recomanable deixar l’aigua que farem servir per la fixació uns
30 min al buit per tal de remoure l’aire (farem servir 292ml per ecotip i condició).
Procediment:
1. Recollir tot el material biològic. 2. Ràpidament iniciar la fixació en 300 ml a l’1% de formaldehid al buit (292 ml H2O +8 ml
37% formaldehid). Incubar 5min, trencar el buit suament per remenar una mica la mostra, tornar a fer el buit 10min més, tornar a remenar i finalment fer 5 min més de buit.
3. Finalitzar la fixació afegint 20 ml de glicina 2M (concentració final 0.125 M) i fent el buit 5min més.
4. Rentar el material vegetal 3 vegades en aigua abundant. 5. Remoure el màxim possible d’aigua i assecar ràpidament les mostres en una mica de
paper absorbent. 6. Pesar immediatament les mostres (dividir en parts d’un gram), guardar en paper
d’alumini en nitrogen líquid i per més llarg temps a -80oC. Aquestes mostres es poden guardar mesos.
Extracció de cromatina (4-5h):
Aquest procediment aïlla primer els nuclis per després fer una lísis d’aquests i sonicar la cromatina per tal de tenir fragments del tamany desitjable per fer la IP i poder detectar els fragments purificats per qPCR. Es va procedir amb una mostra de les anteriors (per cada ecotip i condició a testejar).
Procediment:
1. Triturar fortament el teixit vegetal (1g), en un morter amb nitrogen líquid. 2. Passar el material a un tub en gel amb 30 ml de Tampó d’extracció 1, amb l’ajuda d’una
espàtula refredada en nitrogen líquid. Mantenir la mostra en gel i remenar lleugerament per homogeneïtzar la mostra en el tampó (no fer servir vòrtex, ja que podria trencar els nuclis i la cromatina).
3. Incubar en gel un parell de minuts agitant suaument de tant en tant, mentre es preparen els filtres de Miracloth.
4. Filtrar dues vegades amb el Miracloth en tubs nous en gel. 5. Centrifugar la suspensió anterior (2000xg) durant 20 min a 4oC. 6. Remoure el sobrenedant i resuspendre en 1 ml de Tampó d’extracció 2. 7. Transferir a un tub eppendorf d’1.5 ml (mantenir en gel) i centrifugar a màxima velocitat
en una microfuga (12000-14000 rpm) durant 10 min a 4oC. 8. Remoure el sobrenedant i resuspendre el pellet en 500 µl de Tampó d’extracció 3
(sempre en gel).

6 Materials i mètodes
120
9. En un tub eppendorf nou afegir 500 µl de Tampó d’extracció 3. 10. Deixar el pellet resuspès sobre aquesta capa amb cura, es formen dues capes ben
separades. 11. Centrifugar durant 1 h a velocitat màxima (12000-14000 rpm) a 4oC. Això ens dona lloc
a 3 capes diferents, amb els nuclis formant el pellet inferior. 12. Remoure el sobrenedant i resuspendre els nuclis en 500 µl de Tampó de lísis de nucli
(sempre mantenint en gel). Es pot fer una petita centrifugació a 100 rpm uns 30s per reduir les bombolles i millorar la sonicació posterior.
13. Sonicar per obtenir fragments de 400-700pb. Es va fer servir un biorupter (posició High), i es van fer dues rondes de 10 min (30sec ON 30 Sec OFF).
14. Centrifugar la mostra durant 10 min a màxima velocitat a 4oC. 15. Transferir el sobrenedant a un nou tub i tornar a centrifugar 10 min a 4oC. 16. Transferir el sobrenedant a nous tubs eppendorf especials (low binding tubes) en
alíquotes de 150 µl i congelar en nitrogen líquid. S’obtenen 3 mostres, que es poden fer servir com a rèpliques, poden ser guardades a -80ºC.
Immunoprecipitació (IP):
En aquest pas primer es preparen les perles (que poden ser d’agarosa o sefarosa) que contenen una proteïna que uneix fortament a l’anticòs primari, el que permet precipitar les perles amb la proteïna d’interès (unida al DNA). Es van fer servir Sepharose-beads amb proteïna A (GE Healthcare). Totes les centrifugacions amb les perles es fan durant 1min a 2000rpm i a 4ºC, mantenint les mostres en gel durant el procés. Posteriorment es fan la incubació amb l’anticòs, on es pot variar el temps i la quantitat d’anticòs usat, i els rentats, per acabar revertint el procés de cross-link i amb la purificació del DNA obtingut.
Procediment:
1. Rentar les perles de sefarosa (2x40µl /mostra) amb 1 ml de tampó de ChIP. 2. Afegir 40µl de les perles rentades (però encara no bloquejades) al tub on farem la
immunoprecipitació, amb els 150 µl d’extracció de cromatina anterior, incubar uns 10min a 4ºC en agitació (millor per rotació). Centrifugar i descartar les perles. Amb aquest pas eliminem interaccions inespecífiques del nostre extracte amb les perles.
3. Mentre fem la incubació anterior bloquegem unes altres perles amb 1 ml de tampó de ChIP + 10µL (1 mg/ml) BSA i 10 µl (10µg/ml) de DNA d’esperma de salmó sonicat. Incubar també uns 10min a 4ºC en agitació (millor per rotació). En aquest pas eliminem les interaccions inespecífiques de les perles amb el DNA.
4. Rentar les perles dos cops amb 1 ml de tampó de ChIP. Aquestes perles estan equilibrades i bloquejades, llestes per la IP.
5. Afegir 1350 µl de tampó de ChIP als 150 µl de cromatina (en el tub especial low binding) i separar d’aquesta manera:
a. 120 µl com a Input (guardar a -20°C). Ha de ser un 10% de la IP. b. 1200 µl per la IP c. La resta es guarda com a control de sonicació.
6. Afegim 40 µl de les perles bloquejades i 1µg de l’anticòs (anti-GFP de Invitrogen en el nostre cas).
7. Incubar a 4°C en rotació durant 4h. Molt sovint es fa o/v.
8. Centrifugar i quedar-se amb els sobrenedant.
Rentats de 5-10 min a 4°C en agitació.
9. 2x rentats amb tampó de rentat Low salt
10. 1x tampó High salt

6 Materials i mètodes
121
11. 2x tampó LiCl.
12. Finalment 1x TE. Hem acabat els rentats.
13. Afegir 250µl de tampó NaHCO3+SDS, incubar 15min a 50ºC agitant vigorosament. Centrifugar i passar el sobrenedant a un nou tub.
14. Resuspendre les perles de nou en 250µl de tampó NaHCO3+SDS, incubar 15min a 55-60ºC remenant també vigorosament. Centrifugar i ajuntar els dos sobrenedants (total 500µL). En aquests passos hem desfet la interacció de la proteïna A, de manera que hem eluït la nostra proteïna i el seu DNA lligat.
Pels últims passos afegir també les mostres corresponents als Input (afegir a l’Input 500µL de tampó NaCO3+SDS)
15. Overnight a 65°C (revertir el cross-link).
16. Tractament amb RNase, afegint 2µl d’enzim incubant 1-2h, a TªA.
17. Degradació de la proteïna amb 2µl Proteinasa K (Roche) i incubar durant 2h a 45°C.
18. Purificació de DNA amb kit comercial (com està explicat anteriorment, kit de Qiagen).
19. Resuspendre el DNA en 60 µl H2O (o tampó EB del kit).
20. Fer dilucions 1/3 del DNA per usar en qPCR.
qPCR dirigida:
Es va seguir el mateix protocol que per qPCR normal, sobre 10µl de reacció es van fer
servir 2µl de mostra (diluïda 1/3), amb les combinacions d’oligonucleòtids adequades i fent en
la mateixa placa les mostres i els Inputs corresponents.
Tampons:
- Tampó d’extracció 1: 0.4M sucrosa, 10 mM Tris-HCl pH8.0, 0.035% 2-mercaptoetanol (BME), 5mM benzamidina, 1x Inhibidors de proteases Roche (complet sense EDTA). - Tampó d’extracció 2: 0.25M sucrosa, 10 mM Tris-HCl pH8.0, 10mM MgCl2, 1% Tritó X-100, 0.035% BME, 5mM benzamidina, 1x Inhibidors de proteases Roche. - Tampó d’extracció 3: 1.7M sucrosa, 10 mM Tris-HCl pH8.0, 2mM MgCl2, 0.15% Tritó X-100, 0.035% BME, 5mM benzamidina, 1x Inhibidors de proteases Roche. - Tampó de lísis de nuclis: 50 mM Tris-HCl pH8.0, 10mM EDTA, 1% SDS, 5mM benzamidina, 1x Inhibidors de proteases Roche. - Tampó de ChIP: 16.7 mM Tris-HCl pH8.0, 1.2mM EDTA, 1.1% Tritó X-100, 167mM NaCl, 5mM benzamidina, 1x Inhibidors de proteases Roche. - Tampó Low Salt: 20 mM Tris-HCl pH8.0, 2mM EDTA, 0.5% Tritó X-100, 0.2% SDS, 150mM NaCl. - Tampó High Salt: 20 mM Tris-HCl pH8.0, 2mM EDTA, 0.5% Tritó X-100, 0.2% SDS, 500mM NaCl. - Tampó TE: 10 mM Tris-HCl pH8.0, 1mM EDTA. - Tampó NaHCO3 +SDS: 50 mM NaHCO3, 1% SDS. - Tampó de LICl: 0.25 LiCl, 1%NP40, 1% desoxicolat de sodi, 1mM EDTA, 10mM Tris-HCl.
6.4.1.15. Anàlisi de l’RNA per northern-blot.
Preparació de les mostres
Les mostres es preparen afegint 16 µL del tampó de càrrega per RNA a 10 µg d’RNA
total en un volum de 10 µL. Les mostres es desnaturalitzen 15 min. a 65ºC i es mantenen en

6 Materials i mètodes
122
gel fins al moment de carregar-les, quan els afegim 4 µL de tampó de càrrega 6x (apartat
6.4.1.6) per facilitar la seva càrrega en el gel.
Preparació del gel d’agarosa al 1.2% i electroforesi
El gel es prepara fonent 1.8 g d’agarosa en 105 mL d’aigua i 15 mL de MEN 10X. Es
deixa refredar fins aproximadament 65ºC i s’afegeixen 30 mL de formaldehid al 37.5%. Es
barreja bé i s’aboca al portagels. El tampó d’electroforesi és MEN 1X, que s’afegeix fins al nivell
del gel, sense arribar a cobrir-lo. Es carreguen les mostres i s’acaben de cobrir els pous amb
tampó MEN 1X, i es corre l’electroforesi a 100V durant 1h en campana de gasos.
Transferència a membrana de niló
Després de l’electroforesi el gel d’RNA es renta amb aigua durant 10 min. per tal
d’eliminar l’excés de formaldehid, i es procedeix a transferir l’RNA a una membrana de niló per
capil·laritat i en presència d’un tampó d’elevada força iònica (SSC 10X).
1. Col·locar el gel invertit sobre un paper Whatman 3mm prèviament mullat amb el tampó, utilitzat com pont per a l’absorció per capil·laritat del tampó de transferència SSC 10X. Les bombolles que puguin quedar entre el gel i el paper Whatman s’eliminen amb l’ajut d’una vareta de vidre o d’una pipeta.
2. Humitejar la membrana de niló de la mida del gel en aigua i col·locar-la sobre el gel, eliminant les bombolles de la manera descrita anteriorment.
3. Humitejar un paper Whatman 3mm de la mida del gel, col·locar-lo sobre la membrana de niló i eliminar les bombolles.
4. Afegir uns quants papers Whatman 3mm secs i una pila de papers absorbents.
5. Col·locar un vidre al damunt i un pes per a fer pressió sobre el gel. Deixar transferint O/N a TªA.
6. Retirar la membrana de niló i fixar l’RNA a la membrana amb un aparell stratalinker (Stratagene) que uneix covalentment l’RNA a la membrana mitjançant radiació UV.
Solucions: - Tampó de càrrega per RNA: formamida desionitzada, formaldehid 37.5% i tampó MEN
10X (30:9:7, v:v:v). - Tampó SSC 20X (filtració): NaCl 3M, C6H5Na3O7.2H2O (citrat trisòdic) 0.3M. - Tampó MEN 10X: MOPS 200mM, CH3COONa (acetat de liti) 50 mM, EDTA 10 mM.
Tinció de la membrana per comprovar la transferència de l’RNA
Donat que el gel d’RNA no conté bromur d’etidi, la manera que tenim de visualitzar l’RNA a
la membrana i comprovar així l’eficiència de la transferència i la qualitat dels RNA és tenyir-la
amb una solució de blau de metilè (Wilkinson et al. 1991).
Procediment:
1. Incubar en agitació durant uns minuts la membrana de niló després de la transferència amb la quantitat suficient de solució de blau de metilè com per cobrir-la totalment.
2. Quan la membrana es veu tenyida, retirar la solució de blau de metilè i fer uns quants rentats amb aigua destil·lada, fins que s’elimina l’excés de solució i només resten tenyits els RNAs.
3. Si es desitja guardar un registre de la tinció, es pot escanejar la membrana o simplement fer-ne una fotocòpia.

6 Materials i mètodes
123
4. Rentar la membrana amb la solució de rentat fins que desaparegui totalment la tinció.
Solucions:
- Solució de blau de metilè: blau de metilè 0.03% (p/v), NaAc 0.3M, en aigua.
- Solució de rentat: SSC 1% (v/v), SDS 1% (v/v).
Obtenció de sondes de DNA i marcatge radioactiu
Les sondes per a les hibridacions northern-blot es van generar amplificant per PCR DNA
genòmic de l’ecotip Col-0 amb encebadors específics per a cada gen. Els productes de PCR es
van clonar en el vector pCRII-TOPO. La identitat dels inserts es va confirmar per seqüenciació i
posteriorment s’aïllaren per digestió amb enzims de restricció o per PCR amb els encebadors
específics. Aquests productes són els que es van marcar radioactivament amplificant amb
l’encebador indicat a la següent taula.
Gen Plàsmid motlle Encebador per marcatgeDRA2 pMG30 GO77
DRAL pMG31 GO89
NUP54 pMG32 GO83
NUP58 pMG34 GO79
GR pGI0029/35S/GR BO71
25SCedida pel Dr.F. Cantón (Universidad de Málaga).
Random priming
Taula M.7. Sondes radioactives utilitzades en aquest treball.
Per la sonda 25S es va fer servir el kit comercial Random Primed DNA Labeling Kit de
Boehringer i s’han seguit les indicacions del fabricant.
Es van purificar totes les sondes eliminant els nucleòtids no incorporats en una
microcolumna de Sephadex G-50 (ProbeQuant G-50, Amersham Biosciences).
Hibridació de membranes northern-blot amb sondes de DNA
Procediment:
1. Prehibridar la membrana durant >30 min. en un volum de 10 mL de tampó Church dins d’un cilindre d’hibridació a 65ºC.
2. Desnaturalitzar la sonda radioactiva durant 3 min. a 95ºC just abans d’utilitzar-la. Posar-la immediatament en gel.
3. Afegir la sonda radioactiva al tampó Church.
4. Hibridar la membrana a 65ºC un mínim de 5h (es recomana deixar-la O/N).
5. Fer 3 rentats de 20 min. amb tampó de rentat a 65ºC.
6. Segellar la membrana dins una bossa de plàstic i exposar sobre una pantalla phosphor screen (GR HealthCare).
El marcatge es visualitzen utilitzant l’escàner Storm820 (GE HealthCare), i les intensitats
de banda es quantifiquen utilitzant el software Quantity One (BioRad).
Solucions:

6 Materials i mètodes
124
- Tampó Church d’hibridació i prehibridació: SDS 7% (p/v), EDTA 1 mM, tampó fosfat 0.125 M pH 7.2.
- Tampó de rentat: SDS 2% (p/v), EDTA 2 mM, tampó fosfat 40 mM pH 7.2.
Deshibridació de membranes
En el cas de que una membrana s’hagi d’hibridar amb més d’una sonda, la membrana
es deshibrida mitjançant 2 rentats amb el tampó de deshibridació durant 20 min. a 42ºC.
Solucions:
- Tampó de deshibridació: SSC 0.1% (p/v), SDS 0.1% (p/v).
6.4.1.16. Anàlisi in situ de poliA+-RNA.
Aquest protocol és una adaptació del de Dong (Gong, et al. 2005) amb algunes
adaptacions.
1. Unes quatre 4 plàntules en igual estadi de desenvolupament (7 dies de creixement) són
agafades pels controls i per mostra a analitzar i es posen en vials de vidre amb 5 mL de
Còctel de fixació, una mescla 1:1 de Tampó de fixació:heptà (agitat vigorosament).
Agitar els vials suaument 30min a TªA.
2. Deshidratar amb rentats 2 x 5 min en metanol absolut (uns 1.5-2mL, en aquest pas
extraiem els pigments com les clorofil·les).
3. Rentats 3 x 5 min en etanol absolut (aquest material es pot guardar uns dies en etanol).
4. Pemeabilització incubant 30 min en 1:1 etanol:histo-clear (l’Histo-clear és un substitut
menys tòxic del xilè, usat originalment per desestabilitzar la paret cel·lular i les
membranes).
5. Rentats 2 x 5 min amb etanol absolut.
6. Rentats 2 x 5 min amb metanol absolut.
7. Un rentat de 5 min amb 1:1 metanol:Tampó de fixació sense 5% formaldehid.
8. Les mostres són postfixades en Tampó de fixació durant 30 min a TªA.
9. Rentats 2 x Tampó de fixació sense 5% formaldehid.
10. Un rentat amb 1mL de Tampó d’hibridació “Perfect Hyb Plus hybridization buffer”
(Sigma-Aldrich; H-7033).
11. Fem una prehibridació afegint a cada vial 2 mL del tampó d’hibridació comercial i
incubant durant 1 h a uns 50ºC.
12. Finalment, hibridem afegint 15 pmol de sonda (45-mer oligo(dT), uns 3µL de l’estoc
5µM, marcat amb una molècula de fluoresceïna a l’extrem 5’, sonda comprada a
Sigma-Aldrich Company) i incubem a 50ºC en foscor durant més de 8 h (o/v).
13. Després de la hibridació es poden fer uns rentats per eliminar la sonda restant, amb 1 x
30 min de 2xSSC+0.1% SDS a 50ºC en foscor.
14. Un rentat 1x 5 min amb 0,5x SSC+0.1% SDS a 50ºC en foscor.

6 Materials i mètodes
125
Aquestes mostres són muntades amb aigua sobre un portaobjectes i cobertes amb un
cobreobjectes, i observades immediatament al confocal (Leica TCS SP) utilitzant una línea
d’excitació d’argó a 488 nm (Leica Microsystems, Heidelberg, Alemanya) i filtres d’emissió
de 522/DF35. Aproximadament 14-16 seccions òptiques en passos d’1,5 µm van ser
agafades i projectades amb el software propi del confocal. Els experiments es van repetir
com a mínim dues vegades observant com a mínim 3 de les mostres.
Tampons:
- 1xSSC: 0.15M NaCl i 0.015M citrat de sodi.
- Tampó de fixació: 120 mM NaCl, 10 mM Tampó fosfat a pH 7.2, 2.7 mM KCl, 40 mM
EGTA, 0.1% Tween 20, 10% DMSO i, excepte en els passos indicats, 5% formaldehid.
6.4.2. Tècniques de biologia molecular de proteïnes.
6.4.2.1. Obtenció d’extractes proteics.
Per a l’extracció de proteïnes en mostres vegetals es va utilitzar un mètode
desnaturalitzant (Al-Sady, et al. 2006):
1. Pesar uns 200 mg de mostra, congelar en N2(L) i triturar (amb morter o Tissue-lyser).
2. Afegir immediatament 600 µL de tampó d’extracció de proteïnes.
3. Bullir (a 95ºC) durant 2 min. (Es recomana foradar el tub perquè no s’obri de cop amb l’augment de la pressió interna).
4. Centrifugar 10 min. a 13000 rpm.
5. Recuperar el sobrenedant en un nou tub, afegir-hi igual volum de tampó de càrrega per proteïnes.
6. Congelar immediatament en nitrogen líquid.
7. Emmagatzemar les mostres a -80ºC.
Solucions:
- Tampó d’extracció de proteïnes: Tris-HCl 50 mM pH 7.5, NaCl 150 mM, NP40-inicial 0,5%, EDTA 1 mM, DTT 3mM, i inhibidors de proteases (aprotinina 2 µg/L, leupeptina 3 µg/L, pepstatina 1 µg/L i PMSF 2 mM).
- Tampó 2xTM per carregar proteïnes: Tris 125 mM pH 6.8, bromofenol 0.04%, SDS 4% (p/v), glicerol 20% (v/v). Just abans d’usar afegir ß-mercaptoetanol 10%, i si és per fer extraccions de proteïna en E.coli afegir inhibidors de proteases (aprotinina 2 µg/L, leupeptina 3 µg/L, pepstatina 1 µg/L i PMSF 2 mM).

6 Materials i mètodes
126
6.4.2.2. Separació electroforètica de proteïnes.
Les proteïnes es van separar mitjançant electroforesi desnaturalitzant per proteïnes
(SDS-PAGE) seguint el protocol descrit a Sambrook (Sambrook and Russell 2001) fent servir
tancs d’electroforèsis de BioRad, seguint les instruccions del fabricant.
6.4.2.3. Anàlisi de proteïnes per transferència i immunodectecció.
Transferència de proteïnes (Western-blot)
A partir d’un gel SDS-PAGE s’ha fet la transferència de les proteïnes a membranes de
nitrocel·lulosa o PVDF seguint el mètode humit descrit per (Towbin et al. 1979). Es va fer servir
l’aparell Mini Trans-Blot Electrophoretic Transfer Cell de BioRad seguint les instruccions del
fabricant. S’utilitzaren 750 mL de tampó de transferència, i les condicions de transferència van
ser 1.5h a 4ºC, amb el voltatge fixat a 100 V.
Solucions: - Tampó de transferència (a 4ºC): 6.07g Tris Base, 28.83g glicina, 200 mL de metanol
(10%) i 1800 mL de H2O.
Immunodetecció quimioluminiscent
La detecció de les proteïnes es va fer per quimioluminescència utilitzant un anticòs
primari específic contra la proteïna a detectar i un anticòs secundari anti-anticòs primari
conjugat amb una peroxidasa (Amersham Pharmacia), que trenca el reactiu quimioluminescent
ECL Plus (ECL Plus West Blotting detection system, Amersham Pharmacia).
Procediment:
1. Bloquejar amb TBST 1X + 2 % (p/v) de llet en pols durant 1h a TªA en agitació.
2. Retirar el bloqueig i afegir l’anticòs primari en tampó TBST 1X amb 0,5 % de llet en pols.
3. Incubar amb l’anticòs primari O/N a 4ºC en agitació.
4. Rentar 3 x 5 min. amb TBST amb 0,5 % de llet en pols a TªA en agitació.
5. Incubar 30 min. amb l’anticòs secundari (anti-anticòs primari) en tampó TBST 1X amb 0,5 % de llet en pols a TªA en agitació.
6. Rentar 3 x 5 min. amb TBST amb 0,5 % de llet en pols a TªA en agitació.
7. Reacció ECL Plus i revelat segons les instruccions del fabricant (imatges de luminescència en LAS4000 de Fujifilm).
Solucions: - Tampó TBST 10X: 200 mL Tris pH7.5 1M, 300 mL NaCl 5M, 10 mL Tween 20 en 1L de
H2O.
6.4.2.4. Anticossos utilitzats
Anticossos primaris:
- -GR (Santa Cruz i Affinity BioReagents): No han funcionat mai bé.
- -MBP (produït pel Dr. Juan José López-Moya): Dilució de treball 1:500

6 Materials i mètodes
127
- -GFP (Invitrogen): Dilució de treball 1:5000
- -Hist HRP (Pierce): Dilució de treball 1:5000
Anticossos secundaris:
- -mousse HRP (Dako). Dilució de treball 1:5000.
- -Rabbit HRP (Pierce): Dilució de treball 1:4000
6.4.3. Tècniques bioquímiques.
6.4.3.1. Anàlisi dels pigments isoprenoides (clorofil·les i carotenoides).
Aquestes anàlisis han estat dutes a terme seguint un protocol facilitat pel laboratori del Dr.
M. Rodríguez Concepción seguint un mètode d’extracció de pigments descrit per Fraser (Fraser
et al. 2000). Es va partir d’uns 30mg de teixit fresc per mostra i es van usar 4 mostres
biològiques, fent dues lectures de les absorbàncies en un espectròmetre SpectraMax M3
(bioNova Científica). Es mostra la mitjana de les 4 mostres amb l’error estàndard associat.
6.4.4. Mètodes de plantes.
6.4.4.1. Esterilització de llavors.
Les llavors d’arabidopsis van ser esterilitzades en tots aquells experiments que
implicaven sembrar-les per al cultiu en placa.
Procediment:
1. Remullar les llavors en solució d’imbibició (v/v) durant almenys 20 min. A partir d’aquest moment es treballa en condicions d’esterilitat sota campana de flux laminar utilitzant solucions i materials estèrils.
2. Retirar la solució i afegir solució d’esterilització. Barrejar bé i mantenir les llavors en aquesta solució no més de 10 min.
3. Retirar la solució i afegir aigua estèril. Barrejar bé i deixar que les llavors sedimentin al fons del tub.
4. Repetir el pas número 3, 4 cops més, amb un total de 5 rentats.

6 Materials i mètodes
128
Solucions: - Solució d’imbibició: Tween 20 0.1 % (v/v). - Solució d’esterilització: Tween 20 0.1 %, lleixiu 10 % (v/v).
6.4.4.2. Sembra de llavors.
Depenent del tipus d’experiment que vulguem dur a terme, les llavors es sembren de la
següent manera:
Sembra de llavors en grup sobre el medi de cultiu
Unes 1000-2000 llavors es sembren en grup directament sobre el medi de cultiu quan es
tracta de llavors de la generació T1, procedents de plantes transformades amb agrobacteri,
d’entre les quals es volen seleccionar aquelles que continguin el transgen.
Procediment:
1. Retirar l’aigua de l’últim rentat de l’esterilització.
2. Resuspendre les llavors en 2.5 mL d’una solució estèril d’agarosa 0.1 % (p/v), que permet que les llavors s’escampin per la superfície del medi sense que s’agreguin.
3. Abocar les llavors en la solució d’agarosa sobre el medi sòlid 0.5MS- amb l’antibiòtic de selecció corresponent i l’antibiòtic cefotaxima per eliminar les possibles restes d’agrobacteri, escampar-les per tota la placa i deixar-les assecar sota campana amb les plaques obertes.
Sembra de llavors una a una sobre el medi de cultiu
Les llavors es sembren d’una en una directament sobre el medi de cultiu quan es tracta de
facilitar el seu recompte (per exemple, en els estudis de segregació d’un transgen) o per als
experiments fisiològics, en una densitat de 100 llavors per placa.
Procediment:
1. Resuspendre les llavors en l’aigua de l’últim rentat de l’esterilització.
2. Pipetejar les llavors amb una pipeta P1000 i deixar-les anar una a una sobre el medi sòlid corresponent contactant suaument la punta de la pipeta amb el medi.
Sembra de llavors en grup sobre paper de filtre
Les llavors es sembren en grup sobre paper de filtre (60g/m2, prèviament autoclavat) quan
es volen dur a terme anàlisis d’RNA, de proteïnes o bé realitzar algun tipus de tractament que
impliqui transferir les plàntules a una determinada solució. La germinació de les llavors sobre
paper de filtre permet que aquestes absorbeixin tots els nutrients del medi alhora que fa
possible la collita ràpida del material biològic sense restes de medi de cultiu, facilitant la
posterior extracció d’RNA o proteïnes, o bé facilitant la transferència de plàntules d’una placa a
una altra.
Procediment:
1. Resuspendre les llavors en l’aigua de l’últim rentat de l’esterilització mitjançant pipeteig.
2. Abocar les llavors sobre el paper de filtre situat sobre el medi sòlid o sembrar-les una a una amb la P1000. Deixar assecar lleugerament les plaques per tal d’eliminar el possible excés d’aigua.
Independentment del sistema de sembra utilitzat, un cop sembrades les llavors les plaques
es segellen amb cinta porosa micropore i s’emmagatzemen en foscor a 4ºC entre 3 i 5 dies.
Aquest procés, conegut com estratificació, permet el trencament de la dormància de les llavors,

6 Materials i mètodes
129
assegurant la germinació sincronitzada de totes les llavors quan les posem en les condicions
de llum i temperatura òptimes pel creixement.
Quan sembrem les llavors directament en terra per cultivar-les en testos a l’hivernacle,
aquestes també es sotmeten al mateix període d’estratificació comentat anteriorment.
6.4.4.3. Obtenció de plantes transgèniques.
Transformació de plantes d’arabidopsis
Plantes d’arabidopsis de l’ecotip silvestre Col-0 (o Ws-2 en els casos indicats) es van
transformar per floral dipping segons el mètode descrit per Clough (Clough et al. 1999), amb un
cultiu d’A.tumefaciens portador de la construcció que en cada cas es volia introduir a la planta
(apartat 6.4.1.4.).
Procediment:
1. Cultivar les plantes d’arabidopsis a transformar en testos a l’hivernacle. En testos d’11.5 cm de diàmetre amb la barreja de substrat coberta per una reixeta, es sembren unes 8-10 llavors, i es seleccionaran unes 6-7 plantes, densitat que permetrà un bon creixement.
2. Quan les plantes han desenvolupat inflorescències de 5-7 cm (aproximadament 6 setmanes després de la sembra), tallar les inflorescències a ran de roseta (tallar la inflorescència principal de la planta fa que es formin noves tiges florals laterals, d’aquesta manera augmentem el número de tiges florals susceptibles a ser transformades). Després de 9-10 dies les noves tiges florals estan en un estadi òptim per a la transformació.
3. Dos dies abans de la transformació, inocular 2 mL de medi YEB amb els antibiòtics adequats amb la soca de l’agrobacteri portadora de la construcció a transformar. Incubar O/N a 28ºC en agitació.
4. Inocular un cultiu d’1 L de medi YEB amb els antibiòtics adequats amb 1 mL del precultiu. Incubar O/N a 28ºC en agitació.
5. Centrifugar el cultiu 10 min. a 4000xg. Descartar el sobrenedant.
6. Resuspendre les cèl·lules en 300 mL de sacarosa 5 % (p/v).
7. Just abans de transformar les plantes afegir 0.02 % (v/v) de Silvet al cultiu d’agrobacteri.
8. Abocar el cultiu en un recipient adequat i submergir-hi les inflorescències de les plantes durant 5 min.
9. Deixar els testos en posició horitzontal en una safata sobre paper de filtre, que absorbirà l’excés d’agrobacteri, i cobrir-la amb un plàstic.
10. Mantenir la safata durant 2-4 dies al fitotró amb baixa intensitat de llum.
11. Passat aquest temps, retirar el plàstic que cobreix la safata, el paper de filtre i posar les plantes en posició vertical per a ser cultivades normalment a l’hivernacle.
6.4.4.4. Anàlisi molecular de les plantes mutants.
Obtenció ràpida de DNA genòmic de planta
Per a l’extracció del DNA genòmic de les plantes hem seguit el protocol descrit per Edwards (Edwards et al. 1991).
Procediment:

6 Materials i mètodes
130
1. Macerar el teixit (una fulla jove en general) en un tub eppendorf durant no més de 15 seg. amb un èmbol de plàstic (pellet pestle polypropylene, SIGMA-Aldrich). Si el nombre de mostres és alt es poden fer amb l’aparell Tissue-Lyser de Qiagen, afegint dues boletes de vidre al tub, posant en nitrogen líquid per congelar el teixit i removent dues vegades 1min. amb una freqüència de 30s-1.
2. Afegir 400 µL de tampó d’extracció i acabar de macerar el teixit.
3. Barrejar amb el vòrtex i centrifugar 1 min. a 13000 rpm.
4. Descartar el sediment i barrejar el sobrenedant ( 400 µl) amb 400 µL d’isopropanol.
5. Incubar a TªA durant 2 min i centrifugar 5 min. a 13000 rpm.
6. Descartar el sobrenedant i rentar el sediment amb 700 µL d’etanol 70%.
7. Centrifugar 5 min. a 13000 rpm i descartar el sobrenedant.
8. Resuspendre el sediment en 50 µL d’aigua destil·lada i autoclavada. Guardar a -20ºC.
Solucions: - Tampó d’extracció: Tris-HCl 200 mM pH 7.5, NaCl 250 mM, EDTA 25 mM i SDS 0.5%.
Comprovació del genotip per PCR
La tècnica de la PCR s’utilitza per detectar ràpidament la presència d’un transgen
introduït a la planta. Aquesta tècnica també s’usa per genotipar les plantes mutants procedents
de col·leccions per inserció d’un T-DNA (SALK) , utilitzant oligonucleòtids de l’element
d’inserció de cada línia i de la regió genòmica flanquejant al seu lloc d’inserció.
La reacció de PCR utilitza com a motlle 5 µL de DNA procedent de l’extracció ràpida de
DNA genòmic de l’apartat anterior i una parella d’encebadors adequada.
6.4.4.5. Tractaments i assajos en planta.
Les plàntules destinades als tractament de llum o hormonals han estat cultivades en
placa en una cambra de cultiu in vitro I-36VL (Percival Scientific Inc, Perry, IA, USA) a 22ºC i
humitat no controlada on la llum Wc és generada per 4 tubs fluorescents verticals cool-white
(80 µmol/m2s de radiació fotosintèticament activa).
Tractaments d’ombra simulada
Els tractaments d’ombra simulada han consistit en enriquir W (alta raó R:FR, 3.2-4.5)
amb FR (λ=727 nm) generada per una làmpada LED (QB1310CS-670-735 LED, Quantum
Devices Inc, Barneveld, WI, USA), provocant una baixa raó R:FR (0.05). La taxa de fluència
s’ha mesurat utilitzant un espectròmetre EPP2000 (StelalrNet Inc., Tampa, FL, USA). El
disseny del tractament dependrà de la finalitat amb que es fa: anàlisi del fenotip molecular o
anàlisi del fenotip fisiològic.
Tractament d’ombra simulada per les anàlisis del fenotip molecular (RNA)
Es van sembrar llavors de les línies d’interès en placa sobre paper de filtre. Al setè dia
(7d) les plantes s’han sotmès a W+FR. S’han recollit mostres immediatament abans i després

6 Materials i mètodes
131
de les hores (h) de tractament indicades en cada cas, congelant-les immediatament en nitrogen
líquid per a l’extracció de l’RNA.
Tractament d’ombra simulada per les anàlisis de fenotip fisiològic
Aquest tractament d’ombra va consistir en sembrar llavors de les línies d’interès en placa
i després de l’estratificació germinar les plantes 2d en Wc. Al segon dia, un grup de plantes
s’han mantingut sota Wc mentre que un altre grup de plantes s’han sotmès a W+FR. Cinc dies
després, és a dir, als 7 dies d’edat de les plàntules, s’han pres imatges de les plàntules per
analitzar el seu fenotip (per exemple medició de l’hipocòtil descrit a l’apartat següent).
Mesura dels fenotips fisiològics
Les anàlisis de fenotip de les plàntules es van realitzar sobre imatges digitals utilitzant el
programa ImageJ (National Institute of Health ImageJ software. Bethesda, MD, USA.
http://rsb.info.nih.gov/ij). Per a les mesures dels hipocòtils, aquests es van col·locar plans sobre
plaques d’agar per poder prendre les imatges. Es van analitzar un mínim de 15 plàntules per a
cada tractament, i cada experiment es va repetir almenys 2 cops. En aquest tipus d’anàlisis es
representa la mitjana de les mesures amb les barres d’error que indiquen l’error estàndard de
les mitjanes.
6.4.4.6. Tractaments amb reguladors del creixement, hormones o dexametasona.
Els tractaments realitzats han estat de 2 tipus:
a) De llarga durada. Quan les plàntules són cultivades en presència d’una determinat
agent, aquest és afegit al medi abans de plaquejar-lo.
b) De curta durada. Els tractaments curts (d’unes poques) es realitzen col·locant les
plàntules crescudes sobre paper de filtre en una placa que conté 4 mL de la solució en aigua
de l’agent amb què les vulguem tractar a la concentració corresponent. A continuació es
mostren els diferents agents utilitzats en aquest treball:
AgentConcentració
estoc DissolventGA3 0.1 M Etanol (100%)
Epibrassinòlida (eBL) 5 mM Etanol (100%)
2,4-D 50 mM Etanol (100%)DEX 5 mM Etanol 50%
Àcid 2,4 diclorofenòxiacàtic. Auxina sintètica.Dexametasona. Corticoide sintètic.
DescripcióÀcid gibberèl.lic (GA).
Anàleg sintètic dels BRs.
Taula M.8. Reguladors de creixement i altres agents utilitzats en aquest treball.
6.4.4.7. Localització subcel·lular per bombardeig.
Per a l’estudi de la localització subcel·lular es van bombardejar cèl·lules de ceba i porro
mitjançant un aparell de bombardeig per pressió d’heli PDS1000/He de Bio-Rad. Es van repetir
els bombardejos dues vegades com a mínim. En aquest procediment el que es fa és dipositar
vectors binaris que codifiquen per les proteïnes a estudiar en partícules d’or, que són
disparades a un epiteli vegetal, fet que permet a algunes partícules entrar dins les cèl·lules i
deixar expressar el vector. Com a control de transformació i per localitzar més fàcilment les

6 Materials i mètodes
132
cèl·lules transformades es va fer servir una cotransformació del vector d’interès amb el plàsmid
pDsRed (RFP), permetent localitzar les cèl·lules per fluorescència vermella.
Preparació de microprojectils
Tot el procediment es duu a terme en campana de flux laminar.
1. Rentar 60mg de partícules d’or (1μm diàmetre) amb 1ml d’etanol absolut (qualitat HPLC).
2. Agitar amb el vòrtex a velocitat màxima durant 10min.
3. Centrifugar 1min a 10000rpm i eliminar el sobrenedant.
4. Rentar amb 1ml de glicerol estèril 50%. Vòrtex a màxima velocitat 30s.
5. Centrifugar 1min a 10000rpm i eliminar el sobrenedant.
6. Repetir el rentat amb glicerol dues vegades més.
7. Aliquotar en eppendorfs en volums de 40μl, que serà el que farem servir pels trets de biolística. Es poden preparar diverses alíquotes i guardar-les durant mesos a -20ºC.
Preparació de les plaques amb el teixit vegetal
Tallem la ceba o el porro i agafem parts tendres de teixit (capes bastant interiors).
Retirem amb unes pinces esterilitzades (rentades amb etanol i flamejades) una capa fina
d’epiteli tendre i la dipositem en una placa amb medi MS1. És recomanable posar diversos
trossets d’epiteli al voltant del centre de la placa. Cal preparar com a mínim una placa per
construcció a bombardejar. Es van deixar incubar les plaques en foscor, a 22ºC, durant unes 4h
abans de procedir amb el bombardeig.
Precipitació del DNA
Aquest procediment es farà per cada placa a bombardejar.
1. Es parteix d’una alíquota de 40 μl de partícules d’or com la descrita prèviament, es vortegen a velocitat màxima 5 min.
2. Afegir-hi 10 μl de DNA (4 μgr de cada plàsmid i la resta d’aigua) i es torna a agitar 5min.
3. Afegir-hi 100 μl d’aigua estèril i tornar a agitar 5min.
4. Afegir 150 μl de Ca2Cl/espermidina (125 μl Ca2Cl 2.5M + 25μl espermidina 0.1M) i agitar 5min.
5. Sedimentar les partícules en gel en posició vertical 15min.
6. Centrifugar 30s i descartar el sobrenedant amb la pipeta.
7. Afegir 500 μl d’etanol absolut (HPLC) i vòrtex 20s.
8. Repetir els passos 5-7.
9. Resuspendre en 16 μl d’etanol absolut (HPLC) i deixar-ho en gel.
10. Sonicar 10s i vortejar 10s, dues vegades consecutives amb cada mostra.
11. Cada alíquota ens serveix per bombardejar dues vegades, el que cobreix una placa. Mantenir les mostres en gel fins al moment del bombardeig.

6 Materials i mètodes
133
Bombardeig amb PDS-1000 He System
1. Encendre la cabina de flux laminar. Esterilitzar l’aparell i les peces amb etanol absolut. En plaques de Petri esterilitzar les membranes portadores i les reixetes de parada amb etanol 100% i deixar-les assecar a la campana.
2. Encendre la bomba de buit. Obrir la clau de l’heli i ajustar a la pressió necessària pel disc de ruptura.
3. Afegir 8 μl de mostra (remenant bé amb la pipeta) sobre la membrana portadora sense tocar-la (en excés), i deixar assecar uns 5min.
4. Posar la reixeta de parada en el suport disparador, posar la membrana portadora sobre el suport i col·locar el suport de manera que la superfície amb la mostra miri cap a la reixeta.
5. Posar el disc de ruptura en el seu suport i l’enroscar-lo en l’extrem del cilindre de gas fins que quedi fix.
6. Col·locar el suport disparador al nivell més proper a la base.
7. Posar la placa amb el teixit vegetal i tancar la càmera.
8. Accionar la bomba de buit fins que el manòmetre marqui 27 (0.1 atm).
9. Accionar el botó de tret (fire) fins a sentir la ruptura del disc, immediatament deixar de pressionar i desfer el buit. Girar la placa i repetir el tret amb una nova membrana i reixeta.
10. Incubar les plaques a fosques a 22ºC de 18 a 48h abans de mirar-les al microscopi.
Transcorregudes 16-20h des del bombardeig, les cèl·lules de ceba i porro es van col·locar sobre un portaobjectes amb aigua i es van cobrir amb un cobreobjectes. La fluorescència GFP es va analitzar en un microscopi confocal Leica TCS SP utilitzant una línea d’excitació d’argó a 488 nm (Leica Microsystems, Heidelberg, Alemanya). Es van examinar un mínim de 2 mostres independents per a cada construcció.
Solucions:
- Espermidina 0.1M “free base”: Abans d’usar s’ha de tractar l’espermidina 5min a 65ºC. És recomanable fer servir alíquotes fresques cada vegada.
6.5. Eines i recursos bioinformàtics.
Durant aquesta tesi s’han utilitzat les següents webs de consulta, les seves eines
bioinformàtiques i els programes esmentats:
- Pubmed: http://www.ncbi.nlm.nih.gov/pubmed/
- The Arabidopsis Information Resource (TAIR): http://www.arabidopsis.org/
- Salk Institute Genomic Analysis Laboratory (SIGnAL): http://signal.salk.edu/
- The Bio-Analytic Resource for Plant Biology (BAR): http://bar.utoronto.ca
- Genevestigator: https://www.genevestigator.ethz.ch/ (Zimmermann et al. 2004).
- Venny: http://bioinfogp.cnb.csic.es/tools/venny/index.html (Oliveros 2007)
- DAVID: http://david.abcc.ncifcrf.gov/ (Huang da et al. 2009)
- Vector NTI 10 (Invitrogen)
- Image J (free software)
- GraphPad Prism 4 (GraphPad)


7 Bibliografia
135
7. BIBLIOGRAFIA. Abe, M., Katsumata, H., Komeda, Y. and Takahashi, T. (2003) Regulation of shoot epidermal cell
differentiation by a pair of homeodomain proteins in Arabidopsis. Development, 130, 635-643. Achard, P. and Genschik, P. (2009) Releasing the brakes of plant growth: how GAs shutdown DELLA
proteins. Journal of experimental botany, 60, 1085-1092. Achard, P., Gong, F., Cheminant, S., Alioua, M., Hedden, P. and Genschik, P. (2008) The cold-
inducible CBF1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism. The Plant cell, 20, 2117-2129.
Achard, P., Gusti, A., Cheminant, S., Alioua, M., Dhondt, S., Coppens, F., Beemster, G.T. and Genschik, P. (2009) Gibberellin signaling controls cell proliferation rate in Arabidopsis. Current biology : CB, 19, 1188-1193.
Ahmad, M. and Cashmore, A. (1996) Seeing blue: the discovery of cryptochrome. Plant molecular biology, 30, 851-861.
Ahmad, M., Jarillo, J.A., Smirnova, O. and Cashmore, A.R. (1998) The CRY1 blue light photoreceptor of Arabidopsis interacts with phytochrome A in vitro. Molecular cell, 1, 939-948.
Al-Sady, B., Kikis, E.A., Monte, E. and Quail, P.H. (2008) Mechanistic duality of transcription factor function in phytochrome signaling. Proceedings of the National Academy of Sciences of the United States of America, 105, 2232-2237.
Al-Sady, B., Ni, W., Kircher, S., Schafer, E. and Quail, P.H. (2006) Photoactivated phytochrome induces rapid PIF3 phosphorylation prior to proteasome-mediated degradation. Molecular cell, 23, 439-446.
Alabadi, D., Gallego-Bartolome, J., Orlando, L., Garcia-Carcel, L., Rubio, V., et al. (2008) Gibberellins modulate light signaling pathways to prevent Arabidopsis seedling de-etiolation in darkness. The Plant journal : for cell and molecular biology, 53, 324-335.
Ariel, F.D., Manavella, P.A., Dezar, C.A. and Chan, R.L. (2007) The true story of the HD-Zip family. Trends in plant science, 12, 419-426.
Arnon, D.I. (1938) Microelements in Culture-Solution Experiments with Higher Plants. American journal of botany, 25, 322-325.
Arsovski, A., Galstyan, A., Guseman, J.M. and L.Nemhauser, J. (2012) Photomorphogenesis. Arabidopsis Book.
Aukerman, M.J., Hirschfeld, M., Wester, L., Weaver, M., Clack, T., Amasino, R.M. and Sharrock, R.A. (1997) A deletion in the PHYD gene of the Arabidopsis Wassilewskija ecotype defines a role for phytochrome D in red/far-red light sensing. The Plant cell, 9, 1317-1326.
Ausubel, F. (1989) Current Protocols in Molecular Biology. Brooklyn, NY: Greene Publishing Association. Bae, G. and Choi, G. (2008) Decoding of light signals by plant phytochromes and their interacting
proteins. Annual review of plant biology, 59, 281-311. Bai, M.Y., Fan, M., Oh, E. and Wang, Z.Y. (2012) A triple helix-loop-helix/basic helix-loop-helix cascade
controls cell elongation downstream of multiple hormonal and environmental signaling pathways in Arabidopsis. The Plant cell, 24, 4917-4929.
Bai, X.T., Gu, B.W., Yin, T., Niu, C., Xi, X.D., Zhang, J., Chen, Z. and Chen, S.J. (2006) Trans-repressive effect of NUP98-PMX1 on PMX1-regulated c-FOS gene through recruitment of histone deacetylase 1 by FG repeats. Cancer research, 66, 4584-4590.
Ballare, C.L., Mazza, C.A., Austin, A.T. and Pierik, R. (2012) Canopy light and plant health. Plant physiology, 160, 145-155.
Ballaré, C.L., Scopel, A.L. and Sáncez, R.A. (1990) Far-Red radiation reflected from adjacent leaves: An early signal of competition in plant canopies. Science, 247, 329-332.
Bao, N., Lye, K.W. and Barton, M.K. (2004) MicroRNA binding sites in Arabidopsis class III HD-ZIP mRNAs are required for methylation of the template chromosome. Developmental cell, 7, 653-662.
Bauer, D., Viczian, A., Kircher, S., Nobis, T., Nitschke, R., et al. (2004) Constitutive photomorphogenesis 1 and multiple photoreceptors control degradation of phytochrome interacting factor 3, a transcription factor required for light signaling in Arabidopsis. The Plant cell, 16, 1433-1445.
Bhalerao, R.P., Eklof, J., Ljung, K., Marchant, A., Bennett, M. and Sandberg, G. (2002) Shoot-derived auxin is essential for early lateral root emergence in Arabidopsis seedlings. The Plant journal : for cell and molecular biology, 29, 325-332.
Bimboin, H. and Doly, J. (1979) A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res, 7, 1613-1623.
Blevins, M.B., Smith, A.M., Phillips, E.M. and Powers, M.A. (2003) Complex formation among the RNA export proteins Nup98, Rae1/Gle2, and TAP. The Journal of biological chemistry, 278, 20979-20988.
Boccalandro, H., Casal, J. and Serna, L. (2007) Secret message at the plant surface. Plant signaling & behavior, 2, 373-375.

7 Bibliografia
136
Bognár, L.K., Hal, A., Ádám, É., Thain, S.C., Nagy, F. and Millar, A.J. (1999) The circadian clock controls the expression pattern of the circadian input photoreceptor, phytochrome B. Proceedings of the National Academy of Sciences of the United States of America, 96, 14652-14657.
Bolle, C. (2004) The role of GRAS proteins in plant signal transduction and development. Planta, 218, 683-692.
Bollman, K.M. (2003) HASTY, the Arabidopsis ortholog of exportin 5/MSN5, regulates phase change and morphogenesis. Development, 130, 1493-1504.
Boonman, A., Prinsen, E., Voesenek, L.A. and Pons, T.L. (2009) Redundant roles of photoreceptors and cytokinins in regulating photosynthetic acclimation to canopy density. Journal of experimental botany, 60, 1179-1190.
Bothwick, H.A., Hendricks, S.B., Parker, M.W., Toole, E.H. and Toole, V.K. (1952) A reversible photoreaction controlling seed germination. Proceedings of the National Academy of Sciences of the United States of America, 38, 662-666.
Botto, J.F., Sanchez, R.A., Whitelam, G.C. and Casal, J.J. (1996) Phytochrome A Mediates the Promotion of Seed Germination by Very Low Fluences of Light and Canopy Shade Light in Arabidopsis. Plant physiology, 110, 439-444.
Bou-Torrent, J., Roig-Villanova, I. and Martinez-Garcia, J.F. (2008) Light signaling: back to space. Trends in plant science, 13, 108-114.
Bou-Torrent, J., Salla-Martret, M., Brandt, R., Musielak, T., Palauqui, J.C., Martinez-Garcia, J.F. and Wenkel, S. (2012) ATHB4 and HAT3, two class II HD-ZIP transcription factors, control leaf development in Arabidopsis. Plant signaling & behavior, 7, 1382-1387.
Brandt, R., Salla-Martret, M., Bou-Torrent, J., Musielak, T., Stahl, M., et al. (2012) Genome-wide binding-site analysis of REVOLUTA reveals a link between leaf patterning and light-mediated growth responses. The Plant journal : for cell and molecular biology, 72, 31-42.
Briggs, W.R. and Christie, J.M. (2002) Phototropins 1 and 2: versatile plant blue-light receptors. Trends in plant science, 7, 204-210.
Cagnola, J.I., Ploschuk, E., Benech-Arnold, T., Finlayson, S.A. and Casal, J.J. (2012) Stem transcriptome reveals mechanisms to reduce the energetic cost of shade-avoidance responses in tomato. Plant physiology, 160, 1110-1119.
Capelson, M., Liang, Y., Schulte, R., Mair, W., Wagner, U. and Hetzer, M.W. (2010) Chromatin-bound nuclear pore components regulate gene expression in higher eukaryotes. Cell, 140, 372-383.
Carabelli, M., Morelli, G., Whitelam, G. and Ruberti, I. (1996) Twilight-zone and canopy shade induction of the Athb-2 homeobox gene in green plants. Proceedings of the National Academy of Sciences of the United States of America, 93, 3530-3535.
Carabelli, M., Possenti, M., Sessa, G., Ciolfi, A., Sassi, M., Morelli, G. and Ruberti, I. (2007) Canopy shade causes a rapid and transient arrest in leaf development through auxin-induced cytokinin oxidase activity. Genes & development, 21, 1863-1868.
Carabelli, M., Sessa, G., Baima, S., Morelli, G. and Ruberti, I. (1993) The Arabidopsis Athb-2 and -4 genes are strongly induced by far-red-rich light. The plant Journal, 4, 469-479.
Carabelli, M., V, R., G, M. and I., R. (2013) Homeodomain-Leucine Zipper II family of transcription factors to the limelight: Central regulators of plant development. Plant signaling & behavior, 8.
Casal, J.J. (2013) Photoreceptor signaling networks in plant responses to shade. Annual review of plant biology, 64, 403-427.
Casal, J.J., Sánchez, R.A. and Botto, J.F. (1998) Modes of action of phytochromes. Jounal of Experimental Botany, 49, 127-138.
Castillon, A., Shen, H. and Huq, E. (2007) Phytochrome Interacting Factors: central players in phytochrome-mediated light signaling networks. Trends in plant science, 12, 514-521.
Cerrudo, I., Keller, M.M., Cargnel, M.D., Demkura, P.V., de Wit, M., Patitucci, M.S., Pierik, R., Pieterse, C.M. and Ballare, C.L. (2012) Low red/far-red ratios reduce Arabidopsis resistance to Botrytis cinerea and jasmonate responses via a COI1-JAZ10-dependent, salicylic acid-independent mechanism. Plant physiology, 158, 2042-2052.
Ciarbelli, A.R., Ciolfi, A., Salvucci, S., Ruzza, V., Possenti, M., et al. (2008) The Arabidopsis homeodomain-leucine zipper II gene family: diversity and redundancy. Plant molecular biology, 68, 465-478.
Cifuentes-Esquivel, N. (2012) Análisis de la interconexión entre rutas transcripcionales hormonales y del síndrome de huida de la sombra en Arabidopsis. In Facultat de biologia: Universitat de Barcelona.
Cifuentes-Esquivel, N., Bou-Torrent, J., Galstyan, A., Gallemi, M., Sessa, G., Salla Martret, M., Roig-Villanova, I., Ruberti, I. and Martinez-Garcia, J.F. (2013) The bHLH proteins BEE and BIM positively modulate the shade avoidance syndrome in Arabidopsis seedlings. The Plant journal : for cell and molecular biology.
Clack, T., Shokry, A., Moffet, M., Liu, P., Faul, M. and Sharrock, R.A. (2009) Obligate heterodimerization of Arabidopsis phytochromes C and E and interaction with the PIF3 basic helix-loop-helix transcription factor. The Plant cell, 21, 786-799.
Clough, R.C., Jordan-Beebe, E.T., Lohman, K.N., Marita, J.M., Walker, J.M., Gatz, C. and Vierstra, R.D. (1999) Sequences within both the N- and C-terminal domains of phytochrome A are required for PFR ubiquitination and degradation. Plant J, 17, 155-167.

7 Bibliografia
137
Clouse, S.D. and Sasse, J.M. (1998) BRASSINOSTEROIDS: Essential Regulators of Plant Growth and Development. Annu Rev Plant Physiol Plant Mol Biol, 49, 427-451.
Colon-Carmona, A., Chen, D.L., Yeh, K.C. and Abel, S. (2000) Aux/IAA proteins are phosphorylated by phytochrome in vitro. Plant physiology, 124, 1728-1738.
Covington, M.F. and Harmer, S.L. (2007) The circadian clock regulates auxin signaling and responses in Arabidopsis. PLoS biology, 5, e222.
Cross, M.K. and Powers, M.A. (2011) Nup98 regulates bipolar spindle assembly through association with microtubules and opposition of MCAK. Molecular biology of the cell, 22, 661-672.
Chandler, J.W., Cole, M., Flier, A., Grewe, B. and Werr, W. (2007) The AP2 transcription factors DORNROSCHEN and DORNROSCHEN-LIKE redundantly control Arabidopsis embryo patterning via interaction with PHAVOLUTA. Development, 134, 1653-1662.
Chandler, J.W., Cole, M., Flier, A. and Werr, W. (2009) BIM1, a bHLH protein involved in brassinosteroid signalling, controls Arabidopsis embryonic patterning via interaction with DORNROSCHEN and DORNROSCHEN-LIKE. Plant molecular biology, 69, 57-68.
Chapman, E.J., Greenham, K., Castillejo, C., Sartor, R., Bialy, A., Sun, T.P. and Estelle, M. (2012) Hypocotyl transcriptome reveals auxin regulation of growth-promoting genes through GA-dependent and -independent pathways. PloS one, 7, e36210.
Chatel, G., Desai, S.H., Mattheyses, A.L., Powers, M.A. and Fahrenkrog, B. (2012) Domain topology of nucleoporin Nup98 within the nuclear pore complex. Journal of structural biology, 177, 81-89.
Chen, F., Shi, X., Chen, L., Dai, M., Zhou, Z., et al. (2012) Phosphorylation of FAR-RED ELONGATED HYPOCOTYL1 is a key mechanism defining signaling dynamics of phytochrome A under red and far-red light in Arabidopsis. The Plant cell, 24, 1907-1920.
Chen, M., Chory, J. and Fankhauser, C. (2004) Light signal transduction in higher plants. Annu Rev Genet, 38, 87-117.
Cheng, Y.T., Germain, H., Wiermer, M., Bi, D., Xu, F., et al. (2009) Nuclear pore complex component MOS7/Nup88 is required for innate immunity and nuclear accumulation of defense regulators in Arabidopsis. The Plant cell, 21, 2503-2516.
Christie, J.M. (1998) Arabidopsis NPH1: A Flavoprotein with the Properties of a Photoreceptor for Phototropism. Science, 282, 1698-1701.
Christie, J.M. (2007) Phototropin blue-light receptors. Annual review of plant biology, 58, 21-45. de Lucas, M., Daviere, J.M., Rodriguez-Falcon, M., Pontin, M., Iglesias-Pedraz, J.M., et al. (2008) A
molecular framework for light and gibberellin control of cell elongation. Nature, 451, 480-484. De Smet, I., Lau, S., Ehrismann, J.S., Axiotis, I., Kolb, M., Kientz, M., Weijers, D. and Jurgens, G.
(2013) Transcriptional repression of BODENLOS by HD-ZIP transcription factor HB5 in Arabidopsis thaliana. Journal of experimental botany, 64, 3009-3019.
de Wit, M., Kegge, W., Evers, J.B., Vergeer-van Eijk, M.H., Gankema, P., Voesenek, L.A. and Pierik, R. (2012) Plant neighbor detection through touching leaf tips precedes phytochrome signals. Proceedings of the National Academy of Sciences of the United States of America, 109, 14705-14710.
de Wit, M., Spoel, S.H., Sanchez-Perez, G.F., Gommers, C.M., Pieterse, C.M., Voesenek, L.A. and Pierik, R. (2013) Perception of low red:far-red ratio compromises both salicylic acid- and jasmonic acid-dependent pathogen defences in Arabidopsis. The Plant journal : for cell and molecular biology, 75, 90-103.
Dechaine, J.M., Gardner, G. and Weinig, C. (2009) Phytochromes differentially regulate seed germination responses to light quality and temperature cues during seed maturation. Plant, cell & environment, 32, 1297-1309.
Demarsy, E. and Fankhauser, C. (2009) Higher plants use LOV to perceive blue light. Current opinion in plant biology, 12, 69-74.
Devlin, P.F. and Kay, S.A. (2000) Cryptochromes are required for phytochrome signalling to circadian clock but not for rythmicity. The Plant cell, 12, 2499-2509.
Devlin, P.F., Patel, S.R. and Whitelam, G.C. (1998) Phytochrome E influences internode elongation and flowering time in Arabidopsis. The Plant cell, 10, 1479-1487.
Devlin, P.F., Robson, P.R.H., Patel, S.R., Goosey, L., Sharrock, R.A. and Whitelam, G.C. (1999) Phytochrome D acts in the shade-avoidance syndrome in Arabidopsis bt controlling elongation growth and flowering time. Plant physiology, 119, 909-915.
Devlin, P.F., Yanovsky, M.J. and Kay, S.A. (2003) A genomic analysis of the shade avoidance response in Arabidopsis. Plant physiology, 133, 1617-1629.
Dietzel, L. and Pfannschmidt, T. (2008) Photosynthetic acclimation to light gradients in plants stands comes out of shade. Plant signaling & behavior, 3:12, 116-118.
Dijkwel, P.P., Huijser, C., Weisbeek, P.J., Chua, N.-H. and Smeekens, S.C.M. (1997) Sucrose control of phytochrome A signalling in Arabidopsis. The Plant cell, 9, 583-595.
Djakovic-Petrovic, T., de Wit, M., Voesenek, L.A. and Pierik, R. (2007) DELLA protein function in growth responses to canopy signals. The Plant journal : for cell and molecular biology, 51, 117-126.
Dong, C.H., Hu, X., Tang, W., Zheng, X., Kim, Y.S., Lee, B.H. and Zhu, J.K. (2006) A putative Arabidopsis nucleoporin, AtNUP160, is critical for RNA export and required for plant tolerance to cold stress. Molecular and cellular biology, 26, 9533-9543.

7 Bibliografia
138
Edwards, K., Johnstone, C. and Thompson, C. (1991) A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res, 19, 1349.
Faigon-Soverna, A., Harmon, F.G., Storani, L., Karayekov, E., Staneloni, R.J., et al. (2006) A constitutive shade-avoidance mutant implicates TIR-NBS-LRR proteins in Arabidopsis photomorphogenic development. The Plant cell, 18, 2919-2928.
Fairchild, C.D., Schumaker, M.A. and Quail, P.H. (2000) HFR1 encodes an atypical bHLH protein that acts in phytochrome A signal transduction. Genes & development, 14, 2377-2391.
Fankhauser, C. and Casal, J.J. (2004) Phenotypic characterization of a photomorphogenic mutant. The Plant journal : for cell and molecular biology, 39, 747-760.
Fankhauser, C. and Chory, J. (1999) Light receptor kinases in plants! Current biology : CB, 9, R123-126. Favory, J.J., Stec, A., Gruber, H., Rizzini, L., Oravecz, A., et al. (2009) Interaction of COP1 and UVR8
regulates UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis. The EMBO journal, 28, 591-601.
Feng, S., Martinez, C., Gusmaroli, G., Wang, Y., Zhou, J., et al. (2008) Coordinated regulation of Arabidopsis thaliana development by light and gibberellins. Nature, 451, 475-479.
Ferrandez-Ayela, A., Alonso-Peral, M.M., Sanchez-Garcia, A.B., Micol-Ponce, R., Perez-Perez, J.M., Micol, J.L. and Ponce, M.R. (2013) Arabidopsis TRANSCURVATA1 Encodes NUP58, a Component of the Nucleopore Central Channel. PloS one, 8, e67661.
Finlayson, S.A. (2007) Arabidopsis Teosinte Branched1-like 1 regulates axillary bud outgrowth and is homologous to monocot Teosinte Branched1. Plant & cell physiology, 48, 667-677.
Franklin, K.A. (2003) Mutant Analyses Define Multiple Roles for Phytochrome C in Arabidopsis Photomorphogenesis. The Plant cell, 15, 1981-1989.
Franklin, K.A. (2008) Shade avoidance. The New phytologist, 179, 930-944. Franklin, K.A., Lee, S.H., Patel, D., Kumar, S.V., Spartz, A.K., et al. (2011) Phytochrome-interacting
factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proceedings of the National Academy of Sciences of the United States of America, 108, 20231-20235.
Franklin, K.A. and Quail, P.H. (2010) Phytochrome functions in Arabidopsis development. Journal of experimental botany, 61, 11-24.
Franks, T.M. and Hetzer, M.W. (2013) The role of Nup98 in transcription regulation in healthy and diseased cells. Trends in cell biology, 23, 112-117.
Fraser, P.D., Pinto, M.E.S., Holloway, S.E. and Bramley, P.M. (2000) Applications og high-performance liquid chromatography with photodiode array detection to the metabolic profiling of plant isoprenoids. The plant Journal, 24, 551-558.
Frigerio, M., Alabadi, D., Perez-Gomez, J., Garcia-Carcel, L., Phillips, A.L., Hedden, P. and Blazquez, M.A. (2006) Transcriptional regulation of gibberellin metabolism genes by auxin signaling in Arabidopsis. Plant physiology, 142, 553-563.
Funasaka, T., Balan, V., Raz, A. and Wong, R.W. (2013) Nucleoporin Nup98 mediates galectin-3 nuclear-cytoplasmic trafficking. Biochemical and biophysical research communications, 434, 155-161.
Galstyan, A. (2010) Identification and characterization of shade avoidance components in Arabidopsis thaliana. In Facultat de biologia: Universitat de Barcelona.
Galstyan, A., Cifuentes-Esquivel, N., Bou-Torrent, J. and Martinez-Garcia, J.F. (2011) The shade avoidance syndrome in Arabidopsis: a fundamental role for atypical basic helix-loop-helix proteins as transcriptional cofactors. The Plant journal : for cell and molecular biology, 66, 258-267.
Genoud, T., Buchala, A.J., Chua, N.-H. and Métraux, J.-P. (2002) Phytochrome signalling modulates the SA-perceptive pathway in Arabidopsis. The plant Journal, 31, 87-95.
Godoy, M., Franco-Zorrilla, J.M., Perez-Perez, J., Oliveros, J.C., Lorenzo, O. and Solano, R. (2011) Improved protein-binding microarrays for the identification of DNA-binding specificities of transcription factors. The Plant journal : for cell and molecular biology, 66, 700-711.
Gong, Z., Dong, C.H., Lee, H., Zhu, J., Xiong, L., Gong, D., Stevenson, B. and Zhu, J.K. (2005) A DEAD box RNA helicase is essential for mRNA export and important for development and stress responses in Arabidopsis. The Plant cell, 17, 256-267.
Gonzalez-Grandio, E., Poza-Carrion, C., Sorzano, C.O. and Cubas, P. (2013) BRANCHED1 Promotes Axillary Bud Dormancy in Response to Shade in Arabidopsis. The Plant cell, 25, 834-850.
Gray, W.M., Kepinski, S., Rouse, D., Leyser, O. and Estelle, M. (2001) Auxin regulates SCF TIR1-dependent degradation of AUX/IAA proteins. Nature, 414, 271-276.
Griffis, E.R., Altan, N., Lippincott-Schwartz, J. and Powers, M.A. (2002) Nup98 is a mobile nucleoporin with transcription-dependent dynamics. Molecular biology of the cell, 13, 1282-1297.
Guan, T., Kehlenbach, R.H., Schirmer, E.C., Kehlenbach, A., Fan, F., Clurman, B.E., Arnheim, N. and Gerace, L. (2000) Nup50, a Nucleoplasmically Oriented Nucleoporin with a Role in Nuclear Protein Export. Molecular and cellular biology, 20, 5619-5630.
Hagen, G. and Guilfoyle, T. (2002) Auxin-responsive gene expression: genes, promoters and regulatory factors. Plant molecular biology, 49, 373-385.
Halliday, K.J. and Fankhauser, C. (2003) Phytochrome-hormonal signalling networks. The New phytologist, 157, 449-463.

7 Bibliografia
139
Halliday, K.J., Koorneef, M. and Whitelam, G. (1994) Phytocrhome B and at least one other phytochrome mediate the accelerated flowering response of Arabidopsis thaliana L. to Low Red/Far-Red ratio. Plant physiology, 104, 1311-1313-1315.
Halliday, K.J., Salter, M.G., Thingnaes, E. and Whitelam, G.C. (2003) Phytochrome control of flowering is temperature sensitive and correlates with expression of the floral integrator FT. The Plant journal : for cell and molecular biology, 33, 875-885.
Hao, Y., Oh, E., Choi, G., Liang, Z. and Wang, Z.Y. (2012) Interactions between HLH and bHLH factors modulate light-regulated plant development. Molecular plant, 5, 688-697.
He, J.X., Gendron, J.M., Sun, Y., Gampala, S.S., Gendron, N., Sun, C.Q. and Wang, Z.Y. (2005) BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science, 307, 1634-1638.
Hennig, L., Stoddart, W.M., Dieterle, M., Whitelam, G.C. and Schafer, E. (2002) Phytochrome E Controls Light-Induced Germination of Arabidopsis. Plant physiology, 128, 194-200.
Henriksson, E., Olsson, A.S., Johannesson, H., Johansson, H., Hanson, J., Engstrom, P. and Soderman, E. (2005) Homeodomain leucine zipper class I genes in Arabidopsis. Expression patterns and phylogenetic relationships. Plant physiology, 139, 509-518.
Henriques, R. and Mas, P. (2013) Chromatin remodeling and alternative splicing: Pre- and post-transcriptional regulation of the Arabidopsis circadian clock. Seminars in cell & developmental biology, 24, 399-406.
Hiltbrunner, A., Tscheuschler, A., Viczian, A., Kunkel, T., Kircher, S. and Schafer, E. (2006) FHY1 and FHL act together to mediate nuclear accumulation of the phytochrome A photoreceptor. Plant & cell physiology, 47, 1023-1034.
Himmmelbach, A., Hoffmann, T., Leube, M., Höhener, B. and Grill, E. (2002) Homeodomain protein ATHB6 is a target of the protein phosphatase ABI1 and regulates hormone responses in Arabidopsis. The EMBO journal, 21, 3029-3038.
Hisamatsu, T., King, R.W., Helliwell, C.A. and Koshioka, M. (2005) The involvement of gibberellin 20-oxidase genes in phytochrome-regulated petiole elongation of Arabidopsis. Plant physiology, 138, 1106-1116.
Holmes, M.G. and Smith, H. (1977) The function of phytochrome in the natural environment. Photochem. Photobiol., 25, 533-545.
Hornitschek, P., Kohnen, M.V., Lorrain, S., Rougemont, J., Ljung, K., et al. (2012) Phytochrome interacting factors 4 and 5 control seedling growth in changing light conditions by directly controlling auxin signaling. The Plant journal : for cell and molecular biology, 71, 699-711.
Hornitschek, P., Lorrain, S., Zoete, V., Michielin, O. and Fankhauser, C. (2009) Inhibition of the shade avoidance response by formation of non-DNA binding bHLH heterodimers. The EMBO journal, 28, 3893-3902.
Huang da, W., Sherman, B.T. and Lempicki, R.A. (2009) Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res, 37, 1-13.
Hunter, C.A. (2003) PAUSED Encodes the Arabidopsis Exportin-t Ortholog. Plant physiology, 132, 2135-2143.
Huq, E., Al-Sady, B., Hudson, M., Kim, C., Apel, K. and Quail, P.H. (2004) Phytochrome-interacting factor 1 is a critical bHLH regulator of chlorophyll biosynthesis. Science, 305, 1937-1941.
Huq, E. and Quail, P.H. (2002) PIF4, a phytochrome-interacting bHLH factor, functions as a negative regulator of phytochrome B signaling in Arabidopsis. The EMBO journal, 21, 2441-2450.
Ishii, K., Arib, G., Lin, C., Houwe, G.V. and Laemmli, U.K. (2002) Chromatin Boundaries in Budding Yeast: The Nuclear Pore Connection. Cell, 109, 551-562.
Islam, S.Z. and Babadoost, M. (2002) effect of Red light treatment of seedlings of pepper, pumpkin, and tomato on occurrence of Phytophthora damping-off. Plant Pathology, 37, 678-681.
Izaguirre, M.M., Mazza, C.A., Biondini, M., Baldwin, I.T. and Ballare, C.L. (2006) Remote sensing of future competitors: impacts on plant defenses. Proceedings of the National Academy of Sciences of the United States of America, 103, 7170-7174.
Jacob, Y., Mongkolsiriwatana, C., Veley, K.M., Kim, S.Y. and Michaels, S.D. (2007) The nuclear pore protein AtTPR is required for RNA homeostasis, flowering time, and auxin signaling. Plant physiology, 144, 1383-1390.
Jang, I.C., Henriques, R., Seo, H.S., Nagatani, A. and Chua, N.H. (2010) Arabidopsis PHYTOCHROME INTERACTING FACTOR proteins promote phytochrome B polyubiquitination by COP1 E3 ligase in the nucleus. The Plant cell, 22, 2370-2383.
Jang, S., Marchal, V., Panigraphi, K.C., Wenkel, S., Spoppe, W., Deng, X.-W., Valverde, F. and Coupland, G. (2008) Arabidopsis COP1 shapes the temporal pattern of CO conferring a photoperiodic flowrering response. The EMBO journal, 27, 1277-1288.
Jeganathan, K.B., Malureanu, L. and van Deursen, J.M. (2005) The Rae1-Nup98 complex prevents aneuploidy by inhibiting securin degradation. Nature, 438, 1036-1039.
Jenkins, G.I. (2009) Signal transduction in responses to UV-B radiation. Annual review of plant biology, 60, 407-431.
Jeong, J. and Choi, G. (2013) Phytochrome-interacting factors have both shared and distinct biological roles. Molecules and cells, 35, 371-380.

7 Bibliografia
140
Jiao, Y., Lau, O.S. and Deng, X.W. (2007) Light-regulated transcriptional networks in higher plants. Nature reviews. Genetics, 8, 217-230.
Johanesson, H., Wang, Y. and Engström, P. (2001) DNA-binding and dimerization preferences of Arabidopsis homeodomain-leucine zipper transcription factors in vitro. Plant molecular biology, 45, 63-73.
Johnson, E., Bradley, M., Harberd, N.P. and Whitelam, G.C. (1994) Photoresponses of light-grown phyA mutants of Arabidopsis. Plant physiology, 105, 141-149.
Josse, E.M., Foreman, J. and Halliday, K.J. (2008) Paths through the phytochrome network. Plant, cell & environment, 31, 667-678.
Josse, E.M., Gan, Y., Bou-Torrent, J., Stewart, K.L., Gilday, A.D., et al. (2011) A DELLA in disguise: SPATULA restrains the growth of the developing Arabidopsis seedling. The Plant cell, 23, 1337-1351.
Jouve, L., Gaspar, T., Kevers, C., Greppin, H. and Agosti, R.D. (1999) Involvement of indole-3-acetic acid in the circadian growth of the first internode of Arabidopsis. Planta, 209, 136-142.
Kagale, S., Links, M.G. and Rozwadowski, K. (2010) Genome-wide analysis of ethylene-responsive element binding factor-associated amphiphilic repression motif-containing transcriptional regulators in Arabidopsis. Plant physiology, 152, 1109-1134.
Kagale, S. and Rozwadowski, K. (2011) EAR motif-mediated transcriptional repression in plants: An underlying mechanism for epigenetic regulation of gene expression. Epigenetics, 6, 141-146.
Kalverda, B., Pickersgill, H., Shloma, V.V. and Fornerod, M. (2010) Nucleoporins directly stimulate expression of developmental and cell-cycle genes inside the nucleoplasm. Cell, 140, 360-371.
Kamata, N., Okada, H., Komeda, Y. and Takahashi, T. (2013) Mutations in epidermis-specific HD-ZIP IV genes affect floral organ identity in Arabidopsis thaliana. The Plant journal : for cell and molecular biology, 75, 430-440.
Kami, C., Lorrain, S., Hornitschek, P. and Fankhauser, C. (2010) Light-Regulated Plant Growth and Development. 91, 29-66.
Karniol, B., Wagner, J.R., Walker, J.M. and Vierstra, R.D. (2005) Phylogenetic analysis of the phytochrome superfamily reveals distinct microbial subfamilies of photoreceptors. The Biochemical journal, 392, 103-116.
Kasper, L.H., K., B.P., A., S.C., E., P.C., L., C.M. and A., v.D.J.M. (1999) CREB binding protein interacts with Nucleoporin-specific FG repeats that activate transcriptions and mediate NUP98-HOXA9 oncogenity. Molecular and cellular biology, 19, 767-776.
Kaufmann, K., Wellmer, F., Muino, J.M., Ferrier, T., Wuest, S.E., et al. (2010) Orchestration of floral initiation by APETALA1. Science, 328, 85-89.
Kebrom, T.H. and Brutnell, T.P. (2007) The molecular analysis of the shade avoidance syndrome in the grasses has begun. Journal of experimental botany, 58, 3079-3089.
Keller, M.M., Jaillais, Y., Pedmale, U.V., Moreno, J.E., Chory, J. and Ballare, C.L. (2011) Cryptochrome 1 and phytochrome B control shade-avoidance responses in Arabidopsis via partially independent hormonal cascades. The Plant journal : for cell and molecular biology, 67, 195-207.
Keuskamp, D.H., Pollmann, S., Voesenek, L.A., Peeters, A.J. and Pierik, R. (2010) Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during competition. Proceedings of the National Academy of Sciences of the United States of America, 107, 22740-22744.
Khanna, R., Huq, E., Kikis, E.A., Al-Sady, B., Lanzatella, C. and Quail, P.H. (2004) A novel molecular recognition motif necessary for targeting photoactivated phytochrome signaling to specific basic helix-loop-helix transcription factors. The Plant cell, 16, 3033-3044.
Kim, B.C., Tennessen, D.J. and Last, R.L. (1998) UV-B-induced photomorphogenesis in Arabidopsis thaliana. The plant Journal, 15, 667-674.
Kim, J., Jung, J.H., Reyes, J.L., Kim, Y.S., Kim, S.Y., et al. (2005) microRNA-directed cleavage of ATHB15 mRNA regulates vascular development in Arabidopsis inflorescence stems. The Plant journal : for cell and molecular biology, 42, 84-94.
Kim, J., Yi, H., Choi, G., Shin, B. and Song, P.S. (2003) Functional characterization of phytochrome interacting factor 3 in phytochrome-mediated light signal transduction. The Plant cell, 15, 2399-2407.
Kim, J.I., Shen, Y., Han, Y.J., Park, J.E., Kirchenbauer, D., Soh, M.S., Nagy, F., Schafer, E. and Song, P.S. (2004) Phytochrome phosphorylation modulates light signaling by influencing the protein-protein interaction. The Plant cell, 16, 2629-2640.
Kim, W.Y., Fujiwara, S., Suh, S.S., Kim, J., Kim, Y., et al. (2007) ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature, 449, 356-360.
Knoester, M., Loon, L.C.v., Heuvel, J.v.d., Henning, J., Bol, J.F. and Linthorst, H.J.M. (1998) Ethylene-insensitive tobacco lacks nonhost resistance against siol-borne fungi. Proceedings of the National Academy of Sciences of the United States of America, 95, 1933-1937.
Kobayashi, Y. and Weigel, D. (2007) Move on up, it's time for change--mobile signals controlling photoperiod-dependent flowering. Genes & development, 21, 2371-2384.
Koh, S., Wiles, A.M., Sharp, J.S., Naider, F.R., Becker, J.M. and Stacey, G. (2002) An Oligopeptide Transporter Gene Family in Arabidopsis. Plant physiology, 128, 21-29.

7 Bibliografia
141
Koini, M.A., Alvey, L., Allen, T., Tilley, C.A., Harberd, N.P., Whitelam, G.C. and Franklin, K.A. (2009) High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Current biology : CB, 19, 408-413.
Krull, S., Dorries, J., Boysen, B., Reidenbach, S., Magnius, L., Norder, H., Thyberg, J. and Cordes, V.C. (2010) Protein Tpr is required for establishing nuclear pore-associated zones of heterochromatin exclusion. The EMBO journal, 29, 1659-1673.
Kumar, S.V., Lucyshyn, D., Jaeger, K.E., Alos, E., Alvey, E., Harberd, N.P. and Wigge, P.A. (2012) Transcription factor PIF4 controls the thermosensory activation of flowering. Nature, 484, 242-245.
Lee, J.Y., Lee, H.S., Wi, S.J., Park, K.Y., Schmit, A.C. and Pai, H.S. (2009) Dual functions of Nicotiana benthamiana Rae1 in interphase and mitosis. The Plant journal : for cell and molecular biology, 59, 278-291.
Lee, S., Lee, S., Yang, K.Y., Kim, Y.M., Park, S.Y., Kim, S.Y. and Soh, M.S. (2006) Overexpression of PRE1 and its homologous genes activates Gibberellin-dependent responses in Arabidopsis thaliana. Plant & cell physiology, 47, 591-600.
Leivar, P., Monte, E., Al-Sady, B., Carle, C., Storer, A., Alonso, J.M., Ecker, J.R. and Quail, P.H. (2008) The Arabidopsis phytochrome-interacting factor PIF7, together with PIF3 and PIF4, regulates responses to prolonged red light by modulating phyB levels. The Plant cell, 20, 337-352.
Leivar, P., Monte, E., Cohn, M.M. and Quail, P.H. (2012) Phytochrome signaling in green Arabidopsis seedlings: impact assessment of a mutually negative phyB-PIF feedback loop. Molecular plant, 5, 734-749.
Leivar, P. and Quail, P.H. (2010) PIFs: pivotal components in a cellular signaling hub. Trends in plant science.
Leivar, P. and Quail, P.H. (2011) PIFs: pivotal components in a cellular signaling hub. Trends in plant science, 16, 19-28.
Leivar, P., Tepperman, J.M., Monte, E., Calderon, R.H., Liu, T.L. and Quail, P.H. (2009) Definition of early transcriptional circuitry involved in light-induced reversal of PIF-imposed repression of photomorphogenesis in young Arabidopsis seedlings. The Plant cell, 21, 3535-3553.
Li, J. (2003) PAUSED, a Putative Exportin-t, Acts Pleiotropically in Arabidopsis Development But Is Dispensable for Viability. Plant physiology, 132, 1913-1924.
Li, J. and Chory, J. (1999) Brassinosteroid action in plants. Journal of experimental botany, 50, 275-282. Li, J., Li, G., Wang, H. and Deng, X.W. (2011) Phytochrome Signaling Mechanisms. The Arabidopsis
Book, 26. Li, J., Nam, K.H., Vafeados, D. and Chory, J. (2002) BIN2, a New Brassinosteroid-Insensitive Locus in Arabidopsis. Plant physiology, 127, 14-22. Li, L., Ljung, K., Breton, G., Schmitz, R.J., Pruneda-Paz, J., et al. (2012) Linking photoreceptor
excitation to changes in plant architecture. Genes & development, 26, 785-790. Liang, Y., Franks, T.M., Marchetto, M.C., Gage, F.H. and Hetzer, M.W. (2013) Dynamic association of
NUP98 with the human genome. PLoS genetics, 9, e1003308. Light, W.H., Freaney, J., Sood, V., Thompson, A., D'Urso, A., Horvath, C.M. and Brickner, J.H. (2013)
A conserved role for human Nup98 in altering chromatin structure and promoting epigenetic transcriptional memory. PLoS biology, 11, e1001524.
Lin, C. and Shalitin, D. (2003) Cryptochrome structure and signal transduction. Annual review of plant biology, 54, 469-496.
Lin, C. and Todo, T. (2005) The cryptochromes. Genome biology, 6, 220. Lin, Q. and Aoyama, T. (2012) Pathways for epidermal cell differentiation via the homeobox gene
GLABRA2: update on the roles of the classic regulator. Journal of integrative plant biology, 54, 729-737.
Lin, R., Ding, L., Casola, C., Ripoll, D.R., Feschotte, C. and Wang, H. (2007) Transposase-derived transcription factors regulate light signaling in Arabidopsis. Science, 318, 1302-1305.
Lopez-Juez, E., Kobayashi, M., Sakurai, A., Kamiya, Y. and Kendrick, R.E. (1995) Phytochrome, Gibberellins, and Hypocotyl Growth (A Study Using the Cucumber (Cucumis sativus L.) long hypocotyl Mutant). Plant physiology, 107, 131-140.
López-Juez, E., Nagatani, A., Tomizawa, K.-l., Deak, M., Kern, R., Kendrick, R.E. and Furuya, M. (1992) The Cucumber Long Hypocotyl Mutant Lacks a Light-Stable
PHY B-like Phytochrome. The Plant cell, 4, 241-251. Lorenzo-Orts, L. (2013) Las nucleoporinas poseen un papel específico y particular en la regulación de la
expresión génica de Arabidopsis thaliana. In Facultat de biociències: Universitat Autònoma de Barcelona.
Lorrain, S., Allen, T., Duek, P.D., Whitelam, G.C. and Fankhauser, C. (2008) Phytochrome-mediated inhibition of shade avoidance involves degradation of growth-promoting bHLH transcription factors. The Plant journal : for cell and molecular biology, 53, 312-323.
Lorrain, S., Trevisan, M., Pradervand, S. and Fankhauser, C. (2009) Phytochrome interacting factors 4 and 5 redundantly limit seedling de-etiolation in continuous far-red light. The Plant journal : for cell and molecular biology, 60, 449-461.

7 Bibliografia
142
Lu, Q., Tang, X., Tian, G., Wang, F., Liu, K., et al. (2010) Arabidopsis homolog of the yeast TREX-2 mRNA export complex: components and anchoring nucleoporin. The Plant journal : for cell and molecular biology, 61, 259-270.
Luccioni, L.G., Oliverio, K.A., Yanovsky, M.J., Boccalandro, H.E. and Casal, J.J. (2002) Brassinosteroid Mutants Uncover Fine Tuning of Phytochrome Signaling. Plant physiology, 128, 173-181.
Luo, Y., Wang, Z., Ji, H., Fang, H., Wang, S., Tian, L. and Li, X. (2013) An Arabidopsis homolog of importin beta1 is required for ABA response and drought tolerance. The Plant journal : for cell and molecular biology, 75, 377-389.
Ma, L., Li, J., Qu, L., Hager, J., Chen, Z., Zhao, H. and Deng, X.W. (2001) Light control of Arabidopsis development entails coordinated regulation of genome expression and cellular pathways. The Plant cell, 13, 2589-2607.
Magnani, E. and Barton, M.K. (2011) A per-ARNT-sim-like sensor domain uniquely regulates the activity of the homeodomain leucine zipper transcription factor REVOLUTA in Arabidopsis. The Plant cell, 23, 567-582.
Makino, S., Matsushika, A., Kojima, M., Yamashino, T. and Mizuno, T. (2002) The APRR1/TOC1 Quintet Implicated in Circadian Rhythms of Arabidopsis thaliana: I. Characterization with APRR1-Overexpressing Plants. Plant & cell physiology, 43, 58-69.
Malapeira, J., Khaitova, L.C. and Mas, P. (2012) Ordered changes in histone modifications at the core of the Arabidopsis circadian clock. Proceedings of the National Academy of Sciences of the United States of America, 109, 21540-21545.
Malapeira, J. and Mas, P. (2013) A chromatin-dependent mechanism regulates gene expression at the core of the Arabidopsis circadian clock. Plant signaling & behavior, 8, e24079.
Mallory, A.C., Reinhart, B.J., Jones-Rhoades, M.W., Tang, G., Zamore, P.D., Barton, M.K. and Bartel, D.P. (2004) MicroRNA control of PHABULOSA in leaf development: importance of pairing to the microRNA 5' region. The EMBO journal, 23, 3356-3364.
Mancinelli, A. (1986) Comparison of Spectral Properties of Phytochromes from Different Preparations. Plant physiology, 82, 956-961.
Maniatis, T., Fritsch, E.F. and Sambrook, J. (1982) Molecular cloning: A laboratory manual Cold Spring Harbor Laboratory press.
Martínez-García, J.F., Galstyan, A., Salla-Martret, M., Cifuentes-Esquivel, N., Gallemí, M. and Bou-Torrent, J. (2010) Regulatory Components of Shade Avoidance Syndrome. 53, 65-116.
Mas, P. (2008) Circadian clock function in Arabidopsis thaliana: time beyond transcription. Trends in cell biology, 18, 273-281.
Mathews, S. (2006) Phytochrome-mediated development in land plants: red light sensing evolves to meet the challenges of changing light environments. Molecular ecology, 15, 3483-3503.
McClure, B.A. and Guilfoyle, T. (1987) Characterization of a class of small auxin-inducible soybean polyadenlated RNAs. Plant molecular biology, 9, 611-623.
Meier, I. and Brkljacic, J. (2010) The Arabidopsis Nuclear Pore and Nuclear Envelope. The Arabidopsis Book, 24.
Merkle, T. (2011) Nucleo-cytoplasmic transport of proteins and RNA in plants. Plant cell reports, 30, 153-176.
Monte, E., Alonso, J.M., Ecker, J.R., Zhang, Y., Li, X., Young, J., Austin-Phillips, S. and Quail, P.H. (2003) Isolation and characterization of phyC mutants in Arabidopsis reveals complex crosstalk between phytochrome signaling pathways. The Plant cell, 15, 1962-1980.
Montgomery, B.L. (2008) Right place, right time: Spatiotemporal light regulation of plant growth and development. Plant signaling & behavior, 3, 1053-1060.
Montgomery, B.L. and Lagarias, J.C. (2002) Phytochrome ancestry: sensors of bilins and light. Trends in plant science, 7, 357-366.
Morelli, G. and Ruberti, I. (2002) Light and shade in the photocontrol of Arabidopsis growth. Trends in plant science, 7, 399-404.
Moreno, J.E., Tao, Y., Chory, J. and Ballare, C.L. (2009) Ecological modulation of plant defense via phytochrome control of jasmonate sensitivity. Proceedings of the National Academy of Sciences of the United States of America, 106, 4935-4940.
Morgan, D.C. and Smith, H. (1979) A systematic relationship between phytochrome-controlled development and species habitat for plants grown in simulated natural radiation. Planta, 145, 253-259.
Mukherjee, K. and Burglin, T.R. (2006) MEKHLA, a novel domain with similarity to PAS domains, is fused to plant homeodomain-leucine zipper III proteins. Plant physiology, 140, 1142-1150.
Nagatani, A. (2004) Light-regulated nuclear localization of phytochromes. Current opinion in plant biology, 7, 708-711.
Nagatani, A., Reed, J.W. and Chory, J. (1993) lsolation and lnitial Characterization of Arabidopsis Mutants That Are Deficient in Phytochrome A. Plant physiology, 102, 269-277.
Nakamura, M., Katsumata, H., Abe, M., Yabe, N., Komeda, Y., Yamamoto, K.T. and Takahashi, T. (2006) Characterization of the class IV homeodomain-Leucine Zipper gene family in Arabidopsis. Plant physiology, 141, 1363-1375.

7 Bibliografia
143
Nakamura, Y., Kato, T., Yamashino, T., Murakami, M. and Mizuno, T. (2007) Characterization of a Set of Phytochrome-Interacting Factor-Like bHLH Proteins in Oryza sativa. Bioscience, Biotechnology, and Biochemistry, 71, 1183-1191.
Navarro, L., Bari, R., Achard, P., Lison, P., Nemri, A., Harberd, N.P. and Jones, J.D. (2008) DELLAs control plant immune responses by modulating the balance of jasmonic acid and salicylic acid signaling. Current biology : CB, 18, 650-655.
Neff, M.M. and Chory, J. (1998) Genetic Interactions between Phytochrome A, Phytochrome B, and Cryptochrome 1 during Arabidopsis Development. Plant physiology, 118, 27-36.
Neff, M.M., Fankhauser, C. and Chory, J. (2000) Light: an indicator of time and place. Genes&Development, 14, 257-271.
Nemhauser, J.L., Feldman, L.J. and Zambryski, P.C. (2000) Auxin and ETTIN in Arabidopsis gynoecium morphogenesis. Development, 127, 3877-3888.
Nemhauser, J.L., Hong, F. and Chory, J. (2006) Different plant hormones regulate similar processes through largely nonoverlapping transcriptional responses. Cell, 126, 467-475.
Ni, M., Tepperman, J.M. and Quail, P.H. (1998) PIF3, a phytochrome-interacting factor necessary for normal photoinduced signal transduction, is a novel basic helix-loop-helix protein. Cell, 95, 657-667.
Nozue, K., Covington, M.F., Duek, P.D., Lorrain, S., Fankhauser, C., Harmer, S.L. and Maloof, J.N. (2007) Rhythmic growth explained by coincidence between internal and external cues. Nature, 448, 358-361.
O'Hara, A. and Jenkins, G.I. (2012) In vivo function of tryptophans in the Arabidopsis UV-B photoreceptor UVR8. The Plant cell, 24, 3755-3766.
Oh, E., Kang, H., Yamaguchi, S., Park, J., Lee, D., Kamiya, Y. and Choi, G. (2009) Genome-wide analysis of genes targeted by PHYTOCHROME INTERACTING FACTOR 3-LIKE5 during seed germination in Arabidopsis. The Plant cell, 21, 403-419.
Oh, E., Kim, J., Park, E., Kim, J.I., Kang, C. and Choi, G. (2004) PIL5, a phytochrome-interacting basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis thaliana. The Plant cell, 16, 3045-3058.
Oh, E., Yamaguchi, S., Hu, J., Yusuke, J., Jung, B., et al. (2007) PIL5, a phytochrome-interacting bHLH protein, regulates gibberellin responsiveness by binding directly to the GAI and RGA promoters in Arabidopsis seeds. The Plant cell, 19, 1192-1208.
Oh, E., Yamaguchi, S., Kamiya, Y., Bae, G., Chung, W.I. and Choi, G. (2006) Light activates the degradation of PIL5 protein to promote seed germination through gibberellin in Arabidopsis. The Plant journal : for cell and molecular biology, 47, 124-139.
Oh, E., Zhu, J.Y. and Wang, Z.Y. (2012) Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses. Nature cell biology, 14, 802-809.
Ohgishi, M., Oka, A., Morelli, G., Ruberti, I. and Aoyama, T. (2001) Negative autoregulation of the Arabidopsis homeobox gene ATHB-2. The Plant journal : for cell and molecular biology, 25, 389-398.
Oka, M., Asally, M., Yasuda, Y., Ogawa, Y., Tachibana, T. and Yoneda, Y. (2010) The mobile FG nucleoporin Nup98 is a cofactor for Crm1-dependent protein export. Molecular biology of the cell, 21, 1885-1896.
Oliveros, J.C. (2007) VENNY. An interactive tool for comparing lists with Venn Diagrams. http://bioinfogp.cnb.csic.es/tools/venny/index.html.
Olsen, J.E., Juntilla, O., Nilsen, J., Eriksson, M.E., Martinussen, I., Olsson, O., Sandberg, G. and Moritz, T. (1997) Ectiopic expression of oat phytochrome A in hybrid aspen changes critical daylength for growth and prevents cold acclimatization. The plant Journal, 12, 1339-1350.
Olsson, A.S.B., Engström, P. and Söderman, E. (2004) The homeobox genes ATHB12 and ATHB7 encode potential regulators of growth in response to water deficit in Arabidopsis. Plant molecular biology, 55, 663-677.
Östin, A., IIlic, N. and Cohen, J.D. (1999) An in Vitro System from Maize Seedlings for Tryptophan-Independent Indole-3-Acetic Acid Biosynthesis1. Plant physiology, 119, 173-178.
Otsuga, D., DeGuzman, B., Prigge, M.J., Drews, G.N. and Clarck, S.E. (2001) REVOLUTA regulates meristem initiation at lateral positions. The plant Journal, 25, 223-236.
Palena, C.M., Tron, A.E., Bertoncini, C.W., Gonzalez, D.H. and Chan, R.L. (2001) Positively charged residues at the N-terminal arm of the homeodomain are required for efficient DNA binding by Homeodomain-leucine Zipper Proteins. J. Mol. Biol., 308, 39-47.
Park, E., Kim, J., Lee, Y., Shin, J., Oh, E., Chung, W.-I., Liu, J.R. and Choi, G. (2004) Degradation of Phytochrome Interacting Factor 3 in Phytochrome-Mediated Light Signaling. Plant & cell physiology, 136, 968-975.
Park, E., Park, J., Kim, J., Nagatani, A., Lagarias, J.C. and Choi, G. (2012) Phytochrome B inhibits binding of phytochrome-interacting factors to their target promoters. The Plant journal : for cell and molecular biology, 72, 537-546.
Parks, B.M. and Quail, P.H. (1993) hy8, a New Class of Arabidopsis Long Hypocotyl Mutants Deficient in Functional Phytochrome A. The Plant cell, 5, 39-48.
Parry, G. (2012) Assessing the function of the plant nuclear pore complex and the search for specificity. Journal of experimental botany, 13.

7 Bibliografia
144
Parry, G., Ward, S., Cernac, A., Dharmasiri, S. and Estelle, M. (2006) The Arabidopsis SUPPRESSOR OF AUXIN RESISTANCE proteins are nucleoporins with an important role in hormone signaling and development. The Plant cell, 18, 1590-1603.
Patel, D., Basu, M., Hayes, S., Majlath, I., Hetherington, F.M., Tschaplinski, T.J. and Franklin, K.A. (2013) Temperature-dependent shade avoidance involves the receptor-like kinase ERECTA. The Plant journal : for cell and molecular biology, 73, 980-992.
Patel, S.S., Belmont, B.J., Sante, J.M. and Rexach, M.F. (2007) Natively unfolded nucleoporins gate protein diffusion across the nuclear pore complex. Cell, 129, 83-96.
Penfield, S., Josse, E.M. and Halliday, K.J. (2010) A role for an alternative splice variant of PIF6 in the control of Arabidopsis primary seed dormancy. Plant molecular biology, 73, 89-95.
Penfield, S., Josse, E.M., Kannangara, R., Gilday, A.D., Halliday, K.J. and Graham, I.A. (2005) Cold and light control seed germination through the bHLH transcription factor SPATULA. Current biology : CB, 15, 1998-2006.
Pierik, R., Cuppens, M.L., Voesenek, L.A. and Visser, E.J. (2004a) Interactions between ethylene and gibberellins in phytochrome-mediated shade avoidance responses in tobacco. Plant physiology, 136, 2928-2936.
Pierik, R., Djakovic-Petrovic, T., Keuskamp, D.H., de Wit, M. and Voesenek, L.A. (2009) Auxin and ethylene regulate elongation responses to neighbor proximity signals independent of gibberellin and della proteins in Arabidopsis. Plant physiology, 149, 1701-1712.
Pierik, R., Voesenek, L.A., de Kroon, H. and Visser, E.J. (2004b) Density-induced plant size reduction and size inequalities in ethylene-sensing and ethylene-insensitive tobacco. Plant Biol (Stuttg), 6, 201-205.
Pierik, R., Whitelam, G.C., Voesenek, L.A., de Kroon, H. and Visser, E.J. (2004c) Canopy studies on ethylene-insensitive tobacco identify ethylene as a novel element in blue light and plant-plant signalling. The Plant journal : for cell and molecular biology, 38, 310-319.
Piskurewicz, U., Jikumaru, Y., Kinoshita, N., Nambara, E., Kamiya, Y. and Lopez-Molina, L. (2008) The gibberellic acid signaling repressor RGL2 inhibits Arabidopsis seed germination by stimulating abscisic acid synthesis and ABI5 activity. The Plant cell, 20, 2729-2745.
Pokorny, R., Klar, T., Hennecke, U., Carell, T., Batschauer, A. and Essen, L.O. (2008) Recognition and repair of UV lesions in loop structures of duplex DNA by DASH-type cryptochrome. Proceedings of the National Academy of Sciences of the United States of America, 105, 21023-21027.
Prigge, M.J., Otsuga, D., Alonso, J.M., Ecker, J.R., Drews, G.N. and Clark, S.E. (2005) Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. The Plant cell, 17, 61-76.
Pritchard, C.E.J., Fornerod, M., Kasper, L.H. and Deursen, J.M.A.v. (1999) RAE1 Is a Shuttling mRNA Export Factor That Binds to a GLEBS-like NUP98 Motif at the Nuclear Pore Complex through Multiple Domains. Journal of cell biology, 145, 237-253.
Quail, P.H. (2002) Photosensory perception and signalling in plant cells: new paradigms? Current opinion in cell biology, 14, 180-188.
Raices, M. and D'Angelo, M.A. (2012) Nuclear pore complex composition: a new regulator of tissue-specific and developmental functions. Nature reviews. Molecular cell biology, 13, 687-699.
Rajani, S. and Sundaresan, V. (2001) The Arabidopsis myc/bHLH gene ALCATRAZ enables cell separation in fruit dehiscence. Current biology : CB, 11, 1914-1922.
Reed, J.W., Foster, K.R., Morgan, P.W. and Chory, J. (1996) Phytochrome B affects responsiveness to gibberellins in Arabidopsis. Plant physiology, 112, 337-342.
Reed, J.W., Nagatani, A., Elich, T.D., Fagan, M. and Chory, J. (1994) Phytochrome A and Phytochrome B Have Overlapping but Distinct Functions in Arabidopsis Development. Plant physiology, 104, 1139-1149.
Reymond, M.C., Brunoud, G., Chauvet, A., Martinez-Garcia, J.F., Martin-Magniette, M.L., Moneger, F. and Scutt, C.P. (2012) A light-regulated genetic module was recruited to carpel development in Arabidopsis following a structural change to SPATULA. The Plant cell, 24, 2812-2825.
Robles, L.M., Deslauriers, S.D., Alvarez, A.A. and Larsen, P.B. (2012) A loss-of-function mutation in the nucleoporin AtNUP160 indicates that normal auxin signalling is required for a proper ethylene response in Arabidopsis. Journal of experimental botany, 63, 2231-2241.
Robson, P., Whitelam, G.C. and Smith, H. (1993) Selected Components of the Shade-Avoidance Syndrome Are Displayed in a Normal Manner in Mutants of Arabidopsis thaliana and Brassica rapa Deficient in Phytochrome B. Plant physiology, 102, 1179-1184.
Rockwell, N.C., Su, Y.S. and Lagarias, J.C. (2006) Phytochrome structure and signaling mechanisms. Annual review of plant biology, 57, 837-858.
Rodrigo-Peiris, T., Xu, X.M., Zhao, Q., Wang, H.J. and Meier, I. (2011) RanGAP is required for post-meiotic mitosis in female gametophyte development in Arabidopsis thaliana. Journal of experimental botany, 62, 2705-2714.
Rodriguez-Concepcion, M., Fores, O., Martinez-Garcia, J.F., Gonzalez, V., Phillips, M.A., Ferrer, A. and Boronat, A. (2004) Distinct light-mediated pathways regulate the biosynthesis and exchange of isoprenoid precursors during Arabidopsis seedling development. The Plant cell, 16, 144-156.
Roig-Villanova, I. (2007) Control molecular i fisiològic de la Síndrome de fugida de l'ombra en Arabidopsis thaliana. In Facultat de biologia. Barcelona: Universitat de Barcelona.

7 Bibliografia
145
Roig-Villanova, I., Bou-Torrent, J., Galstyan, A., Carretero-Paulet, L., Portoles, S., Rodriguez-Concepcion, M. and Martinez-Garcia, J.F. (2007) Interaction of shade avoidance and auxin responses: a role for two novel atypical bHLH proteins. The EMBO journal, 26, 4756-4767.
Roig-Villanova, I., Bou, J., Sorin, C., Devlin, P.F. and Martinez-Garcia, J.F. (2006) Identification of primary target genes of phytochrome signaling. Early transcriptional control during shade avoidance responses in Arabidopsis. Plant physiology, 141, 85-96.
Rolauffs, S., Fackendahl, P., Sahm, J., Fiene, G. and Hoecker, U. (2012) Arabidopsis COP1 and SPA genes are essential for plant elongation but not for acceleration of flowering time in response to a low red light to far-red light ratio. Plant physiology, 160, 2015-2027.
Rosenblum, J.S. and Blobel, G. (1999) Autoproteolysis in nucleoporin biogenesis. Proceedings of the National Academy of Sciences of the United States of America, 96, 11370-11375.
Rosler, J., Klein, I. and Zeidler, M. (2007) Arabidopsis fhl/fhy1 double mutant reveals a distinct cytoplasmic action of phytochrome A. Proceedings of the National Academy of Sciences of the United States of America, 104, 10737-10742.
Rueda, E.C., Dezar, C.A., Gonzalez, D.H. and Chan, R.L. (2005) Hahb-10, a sunflower homeobox-leucine zipper gene, is regulated by light quality and quantity, and promotes early flowering when expressed in Arabidopsis. Plant & cell physiology, 46, 1954-1963.
Sabatini, S., Beis, D., Wolkenfelt, H., Murfett, J., Guilfoyle, T., et al. (1999) An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. Cell, 99, 463-472.
Saijo, Y., Zhu, D., Li, J., Rubio, V., Zhou, Z., Shen, Y., Hoecker, U., Wang, H. and Deng, X.W. (2008) Arabidopsis COP1/SPA1 complex and FHY1/FHY3 associate with distinct phosphorylated forms of phytochrome A in balancing light signaling. Molecular cell, 31, 607-613.
Saiki, R.K., Walsh, P.S., Levenson, C.H. and Erlich, H.A. (1989) Genetic analysis of amplified DNA with immobilized sequence-specific oligonucleotide probes. Proceedings of the National Academy of Sciences of the United States of America, 86, 6230-6234.
Saito, S., Miyaji-Yamaguchi, M. and Nagata, K. (2004) Aberrant intracellular localization of SET-CAN fusion protein, associated with a leukemia, disorganizes nuclear export. International journal of cancer. Journal international du cancer, 111, 501-507.
Salisbury, F.J., Hall, A., Grierson, C.S. and Halliday, K.J. (2007) Phytochrome coordinates Arabidopsis shoot and root development. The Plant journal : for cell and molecular biology, 50, 429-438.
Salter, M.G., Franklin, K.A. and Whitelam, G.C. (2003) Gating of the rapid shade-avoidance response by the circadian clock in plants. Nature, 426, 680-683.
Salla Martret, M. (2012) Control hormonal i genètic de la síndrome de fugida de l'ombra en Arabidopsis thaliana. In Facultat de farmàcia: Universitat de Barcelona.
Sambrook, J. and Russell, D.W. (2001) Molecular cloning: a laboratory manual: Cold Spring Harbor Laboratory press.
Sawa, S., Ohgishi, M., Goda, H., Higuchi, K., Shimada, Y., Yoshida, S. and Koshiba, T. (2002) The HAT2 gene, a member of the HD-Zip gene family, isolated as an auxin inducible gene by DNA microarray screening, affects auxin response in Arabidopsis. The Plant journal : for cell and molecular biology, 32, 1011-1022.
Schena, M., Lloyd, A.M. and Davis, R.W. (1993) The HAT4 gene of Arabidopsis encodes a developmental regulator. Genes & development, 7, 367-379.
Seo, H.S., Watanabe, E., Tokutomi, S., Nagatani, A. and Chua, N.H. (2004) Photoreceptor ubiquitination by COP1 E3 ligase desensitizes phytochrome A signaling. Genes & development, 18, 617-622.
Sessa, G., Carabelli, M., Sassi, M., Ciolfi, A., Possenti, M., Mittempergher, F., Becker, J., Morelli, G. and Ruberti, I. (2005) A dynamic balance between gene activation and repression regulates the shade avoidance response in Arabidopsis. Genes & development, 19, 2811-2815.
Sessa, G., Morelli, G. and Ruberti, I. (1993) The Athb-1 and -2 HD-Zip domains homodimeriza forming complexes of different binding specifcities. The EMBO journal, 12, 3507-3017.
Sessa, G., Morelli, G. and Ruberti, I. (1997) DNA-binding Specificity of the Homeodomain-leucine Zipper Domain. J. Mol. Biol., 274, 303-309.
Shen, H., Zhu, L., Castillon, A., Majee, M., Downie, B. and Huq, E. (2008) Light-induced phosphorylation and degradation of the negative regulator PHYTOCHROME-INTERACTING FACTOR1 from Arabidopsis depend upon its direct physical interactions with photoactivated phytochromes. The Plant cell, 20, 1586-1602.
Shen, Y., Khanna, R., Carle, C.M. and Quail, P.H. (2007) Phytochrome induces rapid PIF5 phosphorylation and degradation in response to red-light activation. Plant physiology, 145, 1043-1051.
Shin, J., Kim, K., Kang, H., Zulfugarov, I.S., Bae, G., Lee, C.H., Lee, D. and Choi, G. (2009) Phytochromes promote seedling light responses by inhibiting four negatively-acting phytochrome-interacting factors. Proceedings of the National Academy of Sciences of the United States of America, 106, 7660-7665.
Shin, J., Park, E. and Choi, G. (2007) PIF3 regulates anthocyanin biosynthesis in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis. The Plant journal : for cell and molecular biology, 49, 981-994.

7 Bibliografia
146
Shinomura, T., Nagatani, A., Hanzawa, H., Kubota, M., Watanabe, M. and Furuya, M. (1996) Action spectra for phytochrome A- and B-specific photoinduction of seed germination in Arabidopsis thaliana. Proceedings of the National Academy of Sciences of the United States of America, 93, 8129-8133.
Singer, S., Zhao, R., Barsotti, A.M., Ouwehand, A., Fazollahi, M., et al. (2012) Nuclear pore component Nup98 is a potential tumor suppressor and regulates posttranscriptional expression of select p53 target genes. Molecular cell, 48, 799-810.
Smith, H. (1982) Light quality, photo-perception and plant strategy. Ann. Rev. Plant Physiol., 33, 481-518. Smith, H. and Whitelam, G. (1997) The shade avoidance syndrome: multiple responses mediated by
multiple phytochromes. Plant, Cell and Environment, 20, 840-844. Smith, J.E. and Jordan, P.W. (1994) Stand Density Effects on Branching in an Annual Legume (Senna
obtusifolia). Annals of botany, 74, 25. Soccio, R.E. and Breslow, J.L. (2003) StAR-related lipid transfer (START) proteins: mediators of
intracellular lipid metabolism. The Journal of biological chemistry, 278, 22183-22186. Somers, D.E., A.Sharrock, R., Tepperman, J.M. and Quail, P.H. (1991) The hy3 Long Hypocotyl Mutant
of Arabidopsis is Deficient in Phytochrome B. The Plant cell, 3, 1263-1274. Somers, D.E. and Fujiwara, S. (2009) Thinking outside the F-box: novel ligands for novel receptors.
Trends in plant science, 14, 206-213. Song, Y.H., Smith, R.W., To, B.J., Millar, A.J. and Imaizumi, T. (2012) FKF1 conveys timing information
for CONSTANS stabilization in photoperiodic flowering. Science, 336, 1045-1049. Sorin, C., Salla-Martret, M., Bou-Torrent, J., Roig-Villanova, I. and Martinez-Garcia, J.F. (2009)
ATHB4, a regulator of shade avoidance, modulates hormone response in Arabidopsis seedlings. The Plant journal : for cell and molecular biology, 59, 266-277.
Spartz, A.K., Lee, S.H., Wenger, J.P., Gonzalez, N., Itoh, H., et al. (2012) The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. The Plant journal : for cell and molecular biology, 70, 978-990.
Ståldal, V. and Sundberg, E. (2009) The role of auxin in style development and apical-basal patterning of the Arabidopsis thaliana gynoecium. Plant signaling & behavior, 4, 83 - 85.
Stavang, J.A., Gallego-Bartolome, J., Gomez, M.D., Yoshida, S., Asami, T., Olsen, J.E., Garcia-Martinez, J.L., Alabadi, D. and Blazquez, M.A. (2009) Hormonal regulation of temperature-induced growth in Arabidopsis. The Plant journal : for cell and molecular biology, 60, 589-601.
Steindler, C., Matteucci, A., Sessa, G., Weimar, T., Ohgishi, M., Aoyama, T., Morelli, G. and Ruberti, I. (1999) Shade avoidance responses are mediated by the ATHB-2 HD-zip protein, a negative regulator of gene expression. Development, 126, 4235-4245.
Stepanova, A.N., Yun, J., Robles, L.M., Novak, O., He, W., Guo, H., Ljung, K. and Alonso, J.M. (2011) The Arabidopsis YUCCA1 flavin monooxygenase functions in the indole-3-pyruvic acid branch of auxin biosynthesis. The Plant cell, 23, 3961-3973.
Stephenson, P.G., Fankhauser, C. and Terry, M.J. (2009) PIF3 is a repressor of chloroplast development. Proceedings of the National Academy of Sciences of the United States of America, 106, 7654-7659.
Sun, Y., Fan, X.Y., Cao, D.M., Tang, W., He, K., et al. (2010) Integration of brassinosteroid signal transduction with the transcription network for plant growth regulation in Arabidopsis. Developmental cell, 19, 765-777.
Sun, Y. and Guo, H.C. (2008) Structural constraints on autoprocessing of the human nucleoporin Nup98. Protein science : a publication of the Protein Society, 17, 494-505.
Swain, S.M. and Singh, D.P. (2005) Tall tales from sly dwarves: novel functions of gibberellins in plant development. Trends in plant science, 10, 123-129.
Takase, M., Mizoguchi, T., Kozuka, T. and Tsukaya, H. (2013) The unique function of the Arabidopsis circadian clock gene PRR5 in the regulation of shade avoidance response. Plant signaling & behavior, 8, e23534.
Tamura, K., Fukao, Y., Iwamoto, M., Haraguchi, T. and Hara-Nishimura, I. (2010) Identification and characterization of nuclear pore complex components in Arabidopsis thaliana. The Plant cell, 22, 4084-4097.
Tamura, K. and Hara-Nishimura, I. (2012) The molecular architecture of the plant nuclear por complex. Journal of experimental botany, 10.
Tao, Y., Ferrer, J.L., Ljung, K., Pojer, F., Hong, F., et al. (2008) Rapid synthesis of auxin via a new tryptophan-dependent pathway is required for shade avoidance in plants. Cell, 133, 164-176.
Tepperman, J.M., Zhu, T., Chang, H.S., Wang, X. and Quail, P.H. (2001) Multiple transcription-factor genes are early targets of phytochrome A signaling. Proceedings of the National Academy of Sciences of the United States of America, 98, 9437-9442.
Terry, L.J. and Wente, S.R. (2007) Nuclear mRNA export requires specific FG nucleoporins for translocation through the nuclear pore complex. The Journal of cell biology, 178, 1121-1132.
Thiele, A., Herold, M., Lenk, I., Quail, P.H. and Gatz, C. (1999) Heterologous Expression of Arabidopsis Phytochrome B in Transgenic Potato Influences Photosynthetic Performance and Tuber Development. Plant physiology, 120, 73-81.
Tilbrook, K., Arongaus, A.B., Binkert, M., Heijde, M., Yin, R. and Ulm, R. (2013) The UVR8 UV-B Photoreceptor: Perception, Signaling and Response. Arabidopsis Book, 11, e0164.

7 Bibliografia
147
Tiwari, S.B., Hagen, G. and Guilfoyle, T.J. (2004) Aux/IAA proteins contain a potent transcriptional repression domain. The Plant cell, 16, 533-543.
Toledo-Ortiz, G., Huq, E. and Rodríguez-Concepción, M. (2010) Direct regulation of phytoene synthase gene expression and carotenoid biosynthesis by phytochrome-interacting factors. Proceedings of the National Academy of Sciences of the United States of America, 107, 11626-11631.
Towbin, H., Staehelin, T. and Gordon, J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A, 76, 4350-4354.
Tron, A.E., Bertoncini, C.W., Chan, R.L. and Gonzalez, D.H. (2002) Redox regulation of plant homeodomain transcription factors. The Journal of biological chemistry, 277, 34800-34807.
Tron, A.E., Bertoncini, C.W., Palena, C.M., Chan, R.L. and Gonzalez, D.H. (2001) Combinatorial interactions of two amino acids with a single base pair define target site specificity in plant dimeric homeodomain proteins. Nucleic Acids Res, 29, 4866-4872.
Turck, F., Fornara, F. and Coupland, G. (2008) Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annual review of plant biology, 59, 573-594.
Turchi, L., Carabelli, M., Ruzza, V., Possenti, M., Sassi, M., et al. (2013) Arabidopsis HD-Zip II transcription factors control apical embryo development and meristem function. Development, 140, 2118-2129.
Ulmasov, T., Hagen, G. and Guilfoyle, T. (1999a) Activation and repression of transcription by auxin-response factors. Proceedings of the National Academy of Sciences of the United States of America, 96, 5844-5849.
Ulmasov, T., Hagen, G. and Guilfoyle, T.J. (1999b) Dimerization and DNA binding of auxin response factors. The plant Journal, 19, 309-319.
Ulmasov, T., Murfett, J., Hagen, G. and Guilfoyle, T.J. (1997) Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. The Plant cell, 9, 1963-1971.
Valdes, A.E., Overnas, E., Johansson, H., Rada-Iglesias, A. and Engstrom, P. (2012) The homeodomain-leucine zipper (HD-Zip) class I transcription factors ATHB7 and ATHB12 modulate abscisic acid signalling by regulating protein phosphatase 2C and abscisic acid receptor gene activities. Plant molecular biology, 80, 405-418.
Vandenbussche, F., Habricot, Y., Condiff, A.S., Maldiney, R., Van der Straeten, D. and Ahmad, M. (2007) HY5 is a point of convergence between cryptochrome and cytokinin signalling pathways in Arabidopsis thaliana. The Plant journal : for cell and molecular biology, 49, 428-441.
Vandenbussche, F., Pierik, R., Millenaar, F.F., Voesenek, L.A. and Van Der Straeten, D. (2005) Reaching out of the shade. Current opinion in plant biology, 8, 462-468.
Vert, G., Walcher, C.L., Chory, J. and Nemhauser, J.L. (2008) Integration of auxin and brassinosteroid pathways by Auxin Response Factor 2. Proceedings of the National Academy of Sciences of the United States of America, 105, 9829-9834.
Walcher, C.L. and Nemhauser, J.L. (2012) Bipartite promoter element required for auxin response. Plant physiology, 158, 273-282.
Walters, R.G. (2005) Towards an understanding of photosynthetic acclimation. Journal of experimental botany, 56, 435-447.
Walther, T.C., Alves, A., Pickersgil, H., Loïodice, I., Hetzer, M., et al. (2003) The Conserved Nup107-160 Complex Is Critical for Nuclear Pore Complex Assembly. Cell, 113, 195-206.
Wang, H. and Deng, X.W. (2002) Arabidopsis FHY3 defines a key phytochome A signaling component directly interacting with its homologous FAR1. The EMBO journal, 21, 1339-1349.
Wang, X., Roig-Villanova, I., Khan, S., Shanahan, H., Quail, P.H., Martinez-Garcia, J.F. and Devlin, P.F. (2011) A novel high-throughput in vivo molecular screen for shade avoidance mutants identifies a novel phyA mutation. Journal of experimental botany, 62, 2973-2987.
Wang, Y. and Folta, K.M. (2013) Contributions of green light to plant growth and development. American journal of botany, 100, 70-78.
Wang, Z.-Y., Gendron, J., Chen, M., Fujioka, S., Nakano, T., et al. (2002) Nuclear-Localized BZR1 Mediates Brassinosteroid-Induced Growth and Feedback Suppression of Brassinosteroid Biosynthesis. Developmental cell, 2, 505-513.
Werner, T., Motyka, V., Laucou, V., Smets, R., Van Onckelen, H. and Schmulling, T. (2003) Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. The Plant cell, 15, 2532-2550.
Whitelam, G.C., Johnson, E., Peng, J., Carol, P., Anderson, M.L., Cowl, J.S. and Harberdb, N.P. (1993) Phytochrome A Null Mutants of Arabidopsis Display a Wild-Type Phenotype in White Light. The Plant cell, 5, 757-768.
Wiermer, M., Cheng, Y.T., Imkampe, J., Li, M., Wang, D., Lipka, V. and Li, X. (2012) Putative members of the Arabidopsis Nup107-160 nuclear pore sub-complex contribute to pathogen defense. The Plant journal : for cell and molecular biology, 70, 796-808.
Wild, M., Daviere, J.M., Cheminant, S., Regnault, T., Baumberger, N., Heintz, D., Baltz, R., Genschik, P. and Achard, P. (2012) The Arabidopsis DELLA RGA-LIKE3 is a direct target of MYC2 and modulates jasmonate signaling responses. The Plant cell, 24, 3307-3319.

7 Bibliografia
148
Wilkinson, M., Doskow, J. and Lindsey, S. (1991) RNA blots: staining procedures and optimization of conditions. Nucleic Acids Res, 19, 679.
Williams, L., Grigg, S.P., Xie, M., Christensen, S. and Fletcher, J.C. (2005) Regulation of Arabidopsis shoot apical meristem and lateral organ formation by microRNA miR166g and its AtHD-ZIP target genes. Development, 132, 3657-3668.
Wu, S.J., Wang, L.C., Yeh, C.H. and Lu, C.A. (2010) Isolation and characterization of the Arabidopsis heat-intolerant 2 (hit2) mutant reveal the essential role of the nuclear export receptor EXPORTIN1A (XPO1A) in plant heat tolerance. The New phytologist, 186, 833-842.
Xu, X.M. and Meier, I. (2008) The nuclear pore comes to the fore. Trends in plant science, 13, 20-27. Xu, X.M., Meulia, T. and Meier, I. (2007a) Anchorage of plant RanGAP to the nuclear envelope involves
novel nuclear-pore-associated proteins. Current biology : CB, 17, 1157-1163. Xu, X.M., Rose, A., Muthuswamy, S., Jeong, S.Y., Venkatakrishnan, S., Zhao, Q. and Meier, I. (2007b)
NUCLEAR PORE ANCHOR, the Arabidopsis homolog of Tpr/Mlp1/Mlp2/megator, is involved in mRNA export and SUMO homeostasis and affects diverse aspects of plant development. The Plant cell, 19, 1537-1548.
Yamashino, T., Matsushika, A., Fujimori, T., Sato, S., Kato, T., Tabata, S. and Mizuno, T. (2003) A link between circadian-controlled bHLH factors and the APRR1/TOC1 quintet in Arabidopsis thaliana. Plant & cell physiology, 44, 619-629.
Yang, D.-L., Yao, J., Mei, C.-S., Tong, X.-H., Zeng, L.-J., et al. (2012) Plant hormone jasmonate priorizes defense over growth by interfering with gibberillin signaling cascade. Proceedings of the National Academy of Sciences of the United States of America, E1, 192-200.
Yasuhara, N., Oka, M. and Yoneda, Y. (2009) The role of the nuclear transport system in cell differentiation. Seminars in cell & developmental biology, 20, 590-599.
Yi, C. and Deng, X.W. (2005) COP1 - from plant photomorphogenesis to mammalian tumorigenesis. Trends in cell biology, 15, 618-625.
Yin, Y., Vafeados, D., Tao, Y., Yoshida, S., Asami, T. and Chory, J. (2005) A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell, 120, 249-259.
Yin, Y., Wang, Z.Y., Mora-Garcia, S., Li, J., Yoshida, S., Asami, T. and Chory, J. (2002) BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell, 109, 181-191.
Zhang, Y. and Li, X. (2005) A putative nucleoporin 96 Is required for both basal defense and constitutive resistance responses mediated by suppressor of npr1-1,constitutive 1. The Plant cell, 17, 1306-1316.
Zhang, Y., Mayba, O., Pfeiffer, A., Shi, H., Tepperman, J.M., Speed, T.P. and Quail, P.H. (2013) A quartet of PIF bHLH factors provides a transcriptionally centered signalling hub that regulates seedling morphogenesis through differencial genes in Arabidopsis. PLoS genetics, 9, 20.
Zhao, Q., Brkljacic, J. and Meier, I. (2008) Two distinct interacting classes of nuclear envelope-associated coiled-coil proteins are required for the tissue-specific nuclear envelope targeting of Arabidopsis RanGAP. The Plant cell, 20, 1639-1651.
Zhao, Q., Leung, S., Corbett, A.H. and Meier, I. (2006) Identification and characterization of the Arabidopsis orthologs of nuclear transport factor 2, the nuclear import factor of ran. Plant physiology, 140, 869-878.
Zhao, Q. and Meier, I. (2011) Identification and characterization of the Arabidopsis FG-repeat nucleoporin Nup62. Plant signaling & behavior, 6, 330-334.
Zhong, S., Shi, H., Xue, C., Wang, L., Xi, Y., Li, J., Quail, P.H., Deng, X.W. and Guo, H. (2012) A molecular framework of light-controlled phytohormone action in Arabidopsis. Current biology : CB, 22, 1530-1535.
Zhu, D., Maier, A., Lee, J.H., Laubinger, S., Saijo, Y., Wang, H., Qu, L.J., Hoecker, U. and Deng, X.W. (2008) Biochemical characterization of Arabidopsis complexes containing CONSTITUTIVELY PHOTOMORPHOGENIC1 and SUPPRESSOR OF PHYA proteins in light control of plant development. The Plant cell, 20, 2307-2323.
Zhu, Y., Tepperman, J.M., Fairchild, C.D. and Quail, P.H. (2000) Phytochrome B binds with greater apparent affinity than phytochrome A to the basic helix-loop-helix factor PIF3 in a reaction requiring the PAS domain of PIF3. Proceedings of the National Academy of Sciences of the United States of America, 97, 13419-13424.
Zimmermann, P., Hirsch-Hoffmann, M., Hennig, L. and Gruissem, W. (2004) GENEVESTIGATOR. Arabidopsis microarray database and analysis toolbox. Plant physiology, 136, 2621-2632.
Zuniga-Mayo, V.M., Marsch-Martinez, N. and de Folter, S. (2012) JAIBA, a class-II HD-ZIP transcription factor involved in the regulation of meristematic activity, and important for correct gynoecium and fruit development in Arabidopsis. The Plant journal : for cell and molecular biology, 71, 314-326.

8 Abreviacions
149
8. ABREVIACIONS 2,4-D Àcid 2,4-diclorfenoxiacètic KmR Resistència a kanamicina
A Adenina LD Dia llarg
µg Micrograms LFRs Respostes de baixa fluència
µL Microlitre LUC Luciferasa
µM Micromolar M Molar
ABA Àcid abscísic MBP Domini d'unió de la maltosa
Amp Ampicil·lina MetOH Metanol
AmpR Resistència a ampicil·lina mg Mil·ligrams
APB Motiu d’unió a phyB min. Minuts
AUX Auxina mL Mil·lilitres
bHLH Dominis Basic helix-loop-helix mM Milimolar
BRs Brassinosteroides MOPS Àcid 2-(N-morfolin) etansulfònic
BRZ Brassinazol mRNA RNA missatger
C Citosina mV Milivolts
CaMV Virus del mosaic de la coliflor NASC Nottingham Arabidopsis Stock Centre
Cb Carbenicil·lina NE Envoltura nuclear
cDNA DNA complementari NES Senyal d'exportació nuclear
ChIP Immunoprecipitació de cromatina ng Nanograms
CHX Cicloheximida NLS Senyal de localització nuclear
CK Citoquinina nm Nanòmetres
cm Centímetres NPA Àcid naftiltalàmic
cry Fotoreceptor criptocrom NPC Complex del nucleoporus
Ct Carboxi terminal Nt Amino terminal
d Dies Nup Nucleoporina
D.O. Densitat òptica O/N Tota la nit
DEX Dexametasona PAR Factors ràpidament regulats per acció dels fitocroms
DMF N-N-dimetilformamida pb Parells de bases
DNA Àcid desoxiribonucleic PCR Reacció en cadena de la Polimerasa
dNTP Desoxiribonucleòtid Pfr Isoforma activa dels fitocroms, absorbent de llum roja llunyana
EBL Epibrassinòlida phot Fotoreceptor fototropina
EMS Etil meta-sulfonat PHY Fotoreceptor fitocrom
EMSA Assaig de retard en mobilitat electroforètica
PIF Proteïnes interactores dels fitocroms
EOD-FR Llum roja llunyana al final del dia PMSF Fluorur de fenilmetan-sulfonil
EOD-R Llum roja al final del dia Pr Isoforma inactiva dels fitocroms, absorbent de llum roja
EtOH Etanol R Llum roja
FR Llum roja llunyana R:FR Raó entre llum roja i roja llunyana
FRc Llum roja llunyana contínua Rc Llum roja contínua
FT Factor de transcripció RFP Proteïna vermella fluorescent
g Força centrífuga relativa (RCF) RNA Àcid ribonucleic
g Grams RNAi Àcid ribonucleic d’interferència
G Guanina rpm Revolucions per minut
GA3 Àcid gibberèl·lic RT-PCR Retrotranscripció per PCR
GAs Gibberel·lines s Segons
GFP Proteïna verda fluorescent SA Àcid salicílic
GR Receptor de glucocorticoides SAS Síndrome de la fugida de l’ombra
h Hores SAUR Small Auxin Up-RNA
HD Domini homeodomain SD Dia curt
HD-Zip Dominis homeodomain-leucine-zipper Tª Temperatura
HIRs Respostes d’alta irradiància TªA Temperatura ambient
HLH Dominis Helix-loop-helix T-DNA DNA transferit
Hyg Higromicina UV-A Llum ultravioleta A

8 Abreviacions
150
HygR Resistència a la higromicina UV-B Llum ultravioleta B
IAA Àcid Indolacètic (auxina). V Volts
IP Immunoprecipitació v/v Relació volum/volum
IPTG Isopropil s-D-tiogalactòsid VLFRs Respostes de molt baixa fluència
JA Àcid jasmònic W Llum blanca
kb Kilobase W+FR Ombra simulada
kDa Kilodalton ZP Cremallera de Leucines
Km Kanamicina λ Longitud d’ona
En seqüències aminoacídiques:
F Phe, fenilalanina S Ser, serina Y Tyr, tirosina K Lys, lisina W Trp, triptòfan
L Leu, leucina P Pro, prolina H His, histidina D Asp, àcid aspàrtic R Arg, arginina I Ile, isoleucina T Thr, treonina Q Gln, glutamina E Glu, àcid glutàmic G Gly, glicina M Met, metionina A Ala, alanina N Asn, asparagina C Cys, cisteïna V Val, valina
Portada: Imatges de l’autor obtingudes al llarg de la present tesi.
Contraportada: Imatges de l’autor obtingudes al llarg de la present tesi i dibuix cedit per Víctor Rodríguez.
Φ

Nup62
Nup54
Nup58 NDC1
GP210
NUP98
Nup155
Nup188
Nup205
Sec13Nup160/SAR1Nup96/SAR3
Nup43 Nup85Nup107 Nup133
Nup160/SAR1
Nup93 Nup88
Nup35
ESD4
p
Nup88Nup93
Sec13
Nup160/SAR1Nup96/SAR3
Nup43 Nup85Nup107 Nup133
NUP98
Play is the highest form of research.Albert Eisntein