Embed Size (px)
Citation preview

A. thaliana, a tiny weed whose common name is “mouse ear cress,”grows low to the ground (Fig. B.1) and produces clusters of small whiteflowers in meadows and laboratories around the globe. Like more than150,000 other species of flowering plants, including roses, daisies,tomatoes, peas, beans, and maple trees, Arabidopsis is a dicotyledo-nous angiosperm—dicotyledonous because the mature embryo car-ries two specialized seed leaves (or cotyledons), and angiosperm
because its seeds are enclosed in an ovary within a flower. Arabidopsisshares three basic characteristics with other plants: It is an autotroph,that is, a nutritionally self-sufficient organism that produces its own foodvia photosynthesis; it is nonmotile, that is, rooted to one spot; and it pro-duces new organ systems continuously throughout its life cycle (whichconsists of alternations between a diploid [2n] sporophyte generationand a haploid [n] gametophyte generation).
The impetus for studying a small flowering weed with no com-mercial potential is straightforward: Plants are the chief source offood for life on our planet, and an understanding of how they growand reproduce may make it possible to grow more nutritious, more copious crops in awide range of environments. In addition, A. thaliana is one of the premier eukaryoticexperimental model organisms. Much of what we have learned from this plant can beapplied directly to animals and other eukaryotes. Classified in the same family as mus-tard and cabbage plants, A. thaliana has several features that make it an excellent ex-perimental model. Its general strategies of growth, development, flowering, and seedproduction are the same as those of other higher plants, including many crop plants.However, it has a shorter generation time than most other plants, requiring only sixweeks for seeds to germinate and develop into mature plants that produce more seeds.Moreover, like the peas that Mendel studied, it reproduces mainly by self-fertilization,yet cross-pollination—through the removal of stamen from one plant’s flowers and theapplication of pollen from another plant’s flowers onto the stigma of the stamen-deprivedflowers—is possible. Whether self- or cross-pollinated, wild-type Arabidopsis plantsproduce a very large number of seeds—from 10,000–40,000 per plant—and these seedshave a high rate of germination. Such rapid, abundant reproduction makes it possiblefor geneticists to screen large populations of seedlings for specific phenotypes. Finally,A. thaliana grows well in the laboratory, requiring relatively little light (illuminationfrom cool, white fluorescent bulbs is sufficient) and temperatures in the rangeof Because the plant is small, researchers can grow hundreds of thousands ofthem a year in a modest-sized laboratory with no special growth facilities.
One goal of geneticists studying A. thaliana is to understand the physiology,biochemistry, growth, and development of a plant at the molecular level. They can use thisknowledge to produce new varieties of crops and other plants with increased stress toler-ance, for example, or containing more of particular parts prized for their nutritional con-tent or floral display. Ornamental plant breeders have already accomplished this secondgoal with some species. For example, the flowers of wild camellias contain stamens (malereproductive organs) and carpels (female reproductive organs) that enable them toproduce seeds; but the flowers of a cultivated variety known as Pink Perfection lack bothmale and female reproductive organs and carry many extra petals in their place (Fig. B.2).
22–26°C.
Arabidopsis thaliana: GeneticPortrait of a Model Plant ReferenceB
21
The flower of Arabidopsis thaliana.
har06584_refB_021-048 11/06/2006 03:00 PM Page 21

B.1 Genome Structure andOrganization
The genome of a model organism for genetic studies shouldbe relatively small but still share the functional genomic char-acteristics of the organisms it represents. A. thaliana fills thisbill to perfection. Its genome, composed of 125 Mb, is one ofthe smallest genomes known in the plant kingdom, notablysmaller than the genomes of most other angiosperms. Bycontrast, the genome of the angiosperm maize (Zea mays) is45 times larger. The Arabidopsis genome is about as large asthe genome of Caenorhabditis elegans (see Portrait C) and is60% the size of the Drosophila melanogaster genome (seePortrait D). The nuclear DNA of Arabidopsis is carried byfive pairs of small chromosomes with well-defined bandingpatterns. Researchers completed sequencing the wholegenome of Arabidopsis in the year 2000.
Comparing Genetic and Physical MapsGeneticists have taken advantage of different geneticstrains of Arabidopsis derived from natural isolates. Be-cause Arabidopsis is a predominantly self-pollinatingplant, each of these isolate-derived strains represents anearly pure inbred line, homozygous for a distinct set of
alleles controlling developmental and physiological charac-teristics. Common strains used in Arabidopsis researchinclude Columbia (Col) and Landsberg erecta (Ler, a mutantderivative of the Landsberg strain).
Arabidopsis strains differ from each other not only inphenotypes but also in a significant number of DNA poly-morphisms that are identifiable by RFLP, PCR, or SNPanalysis (as described in Chapter 11 of the main textbook).Different strains have different alleles at many different DNAmarker loci. Researchers can use allelic variation at DNAmarkers to follow the segregation of chromosomal regionscarrying the markers in the progeny of self-pollinated plantsderived from a cross between different ecotypes. They canalso analyze the linkage between DNA markers as well as be-tween DNA markers and morphological loci (defined by twoor more alleles conferring alternative phenotypes on the dif-ferent strains). With the resulting data, researchers can createDNA marker and phenotypic maps, and ultimately they canintegrate the two types of maps for all five chromosomes(Fig. B.3).
Little Repetitive DNA and a Tight Arrangement of GenesThe small size of the A. thaliana genome stems from the factthat it contains much less repetitive DNA than most other an-giosperms. In fact, the Arabidopsis genome contains 26,607
22 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
In our examination of A. thaliana as a model plant, we present:
• The structure and organization of the genome.• The plant’s anatomy and life cycle.• Techniques of mutational analysis, including chemical and radiation pro-
cedures, insertional mutagenesis, and the analysis of mutations to identifygene function.
• Genetic analysis applied to various aspects of development, includingembryogenesis, hormonal control systems, and responses to environmen-tal signals.
• The genetic analysis of flowering: a comprehensive example.
(a) (b)
Figure B.1 A. thaliana in the palm ofa hand.
Figure B.2 The Pink Perfection camellia as a horticulturalmutant. (a) Wild-type camellia. (b) Pink Perfection flower.
har06584_refB_021-048 11/06/2006 03:00 PM Page 22

23
1M
distance fromcentromere
(megabases)
linkage mappedDNA markers
used for cloning
overlappinggenomicclones
protein-codingloci
SM106_27..SM106_27..F20D22-5..F20D22-5..
SM31_385,7SM31_385,7NF21M12NF21M12SGCSNP148SGCSNP148
SM86_163,5SM86_163,5SGCSNP303SGCSNP303
SM35_139,0SM35_139,0
SM218_20..SM218_20..
SM82_81,0SM82_81,0
S0392S0392SM31_205,8SM31_205,8CXC750CXC750
SM175_77,1SM175_77,1
SM83_254,3SM83_254,3
SM172_87,4SM172_87,4
SM236_22..
SM131_21..SM131_21..
SM98_146,6SM98_146,6
SM254_37..SM254_37..PAP240PAP240SM234_96,3SM234_96,3
SM137_21..SM137_21..
SGCSNP392SGCSNP392
SM152_16..SM152_16..
SM195_92,5SM195_92,5
SM53_190,6SM53_190,6
AF-2AF-2SM252_32..SM252_32..
SM184_37..SM184_37..
SM126_13..SM126_13..ADHADHVE011VE011
SGCSNP142SGCSNP142
2M
3M
4M
5M
6M
7M
8M
9M
10M
11M
12M
13M
14M
15M
16M
17M
18M
19M
20M
21M
22M
23M
24M
25M
26M
27M
28M
29M
30M
Figure B.3 Physical mapping of Arabidopsis chromosome 1. Distance from the centromere is indicated at the left. DNA markersmapped to chromosome 1 by linkage analysis are shown next (with names shown for a small subset). These markers were used to probegenomic libraries to build the physical map. A contig of overlapping genomic clones is in the next column. DNA sequencing and furtheranalysis revealed all of the protein-coding loci along the chromosome. Genes on the left of the line are encoded within the DNA strandhaving a 5 end at the telomere; genes on the right are encoded on the opposite strand with a 5 end at the centromere.¿¿
har06584_refB_021-048 11/06/2006 03:00 PM Page 23

protein-coding genes that make up close to 40% of the totalgenome and can be grouped into functional categories by se-quence analysis (Fig. B.4). This means there is one gene forevery 4.5 kb of genome, on average—an unusually tightarrangement. In most plants, the vast majority of the genomeconsists of repetitive DNA.
Sixty-five percent of the proteins in Arabidopsis are en-coded by more than one gene; and analysis of large duplica-tions in the genome suggests that Arabidopsis derived froman ancient tetraploid. Taken together, these observations sug-gest a large extent of functional redundancy in Arabidopsis.
Like other species, the A. thaliana genome containsmultiple copies of transposable elements, as well as severalpieces of DNA that appear to have originated in the mito-chondrial and chloroplast genomes. As in animals, the cen-tromeres are hetero-chromatic regions rich in repetitiveDNA where recombination is strongly supressed.
Comparisons of the genetic and physical maps of theA. thaliana genome reveal that genetic and physical dis-tances vary widely along the plant’s chromosomes. Withineach chromosome, the frequency of recombination variesgreatly such that a centimorgan represents anywhere from30–1600 kb (or 1.6 Mb).
SummaryA. thaliana has a small genome carried by five chromo-somes. Each chromosome contains a limited amount ofrepetitive DNA outside the centromeric regions and theribosomal RNA gene clusters. Its genes, though similar instructure to the genes of other plants, are more tightlypacked and contain only small introns. As a result, a muchhigher percentage of the genome is associated with codingand genetic function than is true of most plants. The highpercentage of coding sequences and the low abundance ofdispersed repetitive DNA are features that make A. thalianaan excellent model for genetic analysis.
B.2 Anatomy and Life Cycle
To understand a plant’s growth and development at the mo-lecular level, geneticists use mutational analyses to identifygenes that influence the plant’s anatomy and life cycle. Abrief overview of the anatomy and life cycle of A. thalianaprepares the way for examining the techniques of geneticanalysis in this model plant.
Anatomy: The Basic Body PlanLike most higher plants, the adult A. thaliana is composedof only a few relatively simple organs and tissues (Fig. B.5).The aboveground shoot connected to the belowground rootsystem represents the plant’s main axis. Both root and shootcontain several cell types organized into the outer epidermallayer; the middle ground, or cortical, layer; and the innervascular cylinder (which contains the water-conductingxylem vessels and the food-conducting phloem elements).Attached to the shoot are mature leaves also composed ofthree layers: an epidermal layer, containing both hair cells,or trichomes, and small pores, or stomata (singular, stoma)that regulate gas exchange; a mesophyll layer responsiblefor photosynthesis; and vascular tissues that conduct waterand nutrients between the leaf and the stem.
The Apical Meristem Promotes Continuous Growth
Cells at the growing points of both shoot and root are or-ganized into a tissue called the apical meristem, a group ofundifferentiated cells that divide continuously. The rootapical meristem, which consists of a central zone of meri-stem cells and a peripheral zone of differentiated cells, pro-duces the cylindrical root through cell division, cellexpansion, and cell differentiation. The shoot apical meri-stem, which consists of a central zone where cell divisionsalso maintain the meristem and a peripheral zone wherecell divisions give rise to mature organs such as leaves andlateral shoots, generates the entire aboveground plant.
Vegetative Versus Reproductive Development
During vegetative development—the period of growth be-fore flowering—the shoot apical meristem initiates a spiral ofleaves in precise sequence. When the plant switches toreproductive development, the shoot apical meristem be-comes an inflorescence meristem, which, in turn, producesa series of lateral floral meristems. Each floral meristemproduces a floral primordium that gives rise to a flower.The inflorescence is the flower or group of flowers at thetip of a branch. In Arabidopsis, the flowers are arrangedin a spiral around the inflorescence stalk, as depicted inFig. B.5. Plants do not sequester a germline separate fromthe somatic tissue early in development, as is the case in
24 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
Cell growth, cell division,and DNA synthesis
Cell rescue, defense, cell death, and aging
Cellularcommunication/signal transduction
Protein destination
Intracellular transport
Cellular biogenesis
Transport facilitation
Energy
Protein synthesisIonic homeostasis
Transcription
Metabolism
Unclassified
Figure B.4 Functional analysis of Arabidopsis genes. Pro-portion of predicted Arabidopsis genes in different categories.
har06584_refB_021-048 11/06/2006 03:00 PM Page 24
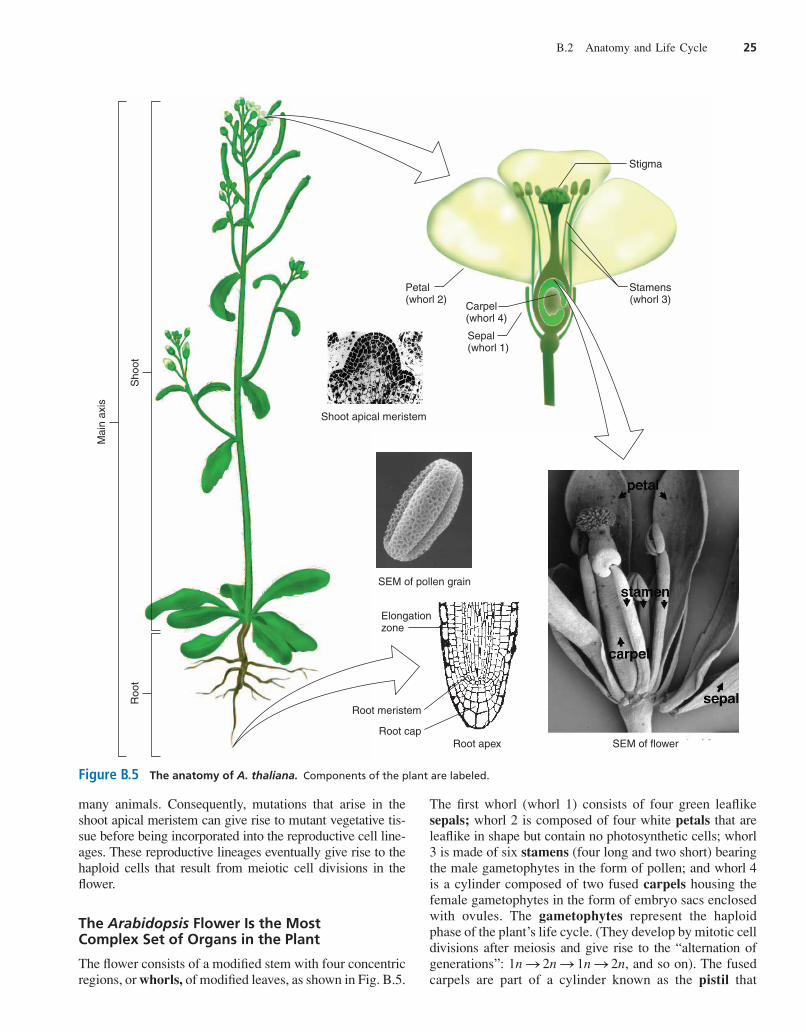
many animals. Consequently, mutations that arise in theshoot apical meristem can give rise to mutant vegetative tis-sue before being incorporated into the reproductive cell line-ages. These reproductive lineages eventually give rise to thehaploid cells that result from meiotic cell divisions in theflower.
The Arabidopsis Flower Is the Most Complex Set of Organs in the Plant
The flower consists of a modified stem with four concentricregions, or whorls, of modified leaves, as shown in Fig. B.5.
The first whorl (whorl 1) consists of four green leaflikesepals; whorl 2 is composed of four white petals that areleaflike in shape but contain no photosynthetic cells; whorl3 is made of six stamens (four long and two short) bearingthe male gametophytes in the form of pollen; and whorl 4is a cylinder composed of two fused carpels housing thefemale gametophytes in the form of embryo sacs enclosedwith ovules. The gametophytes represent the haploidphase of the plant’s life cycle. (They develop by mitotic celldivisions after meiosis and give rise to the “alternation ofgenerations”: , and so on). The fusedcarpels are part of a cylinder known as the pistil that
1n S 2n S 1n S 2n
B.2 Anatomy and Life Cycle 25
Mai
n ax
is
Roo
tS
hoot
Carpel(whorl 4)
Sepal(whorl 1)
Stamens(whorl 3)
Petal(whorl 2)
Stigma
Shoot apical meristem
SEM of pollen grain
Root apex
Root meristem
Root cap
Elongationzone
SEM of flower
Figure B.5 The anatomy of A. thaliana. Components of the plant are labeled.
har06584_refB_021-048 11/06/2006 03:00 PM Page 25
consists of pollen-receptive stigma at the top and a shortneck, or style, leading to the ovary, which houses roughly50 gamete-bearing ovules. Because the flower containsboth male and female reproductive structures, it is consid-ered a perfect flower. After fertilization, the plant sheds theorgans of the outer three whorls, and the pistil developsinto the seed-bearing fruit.
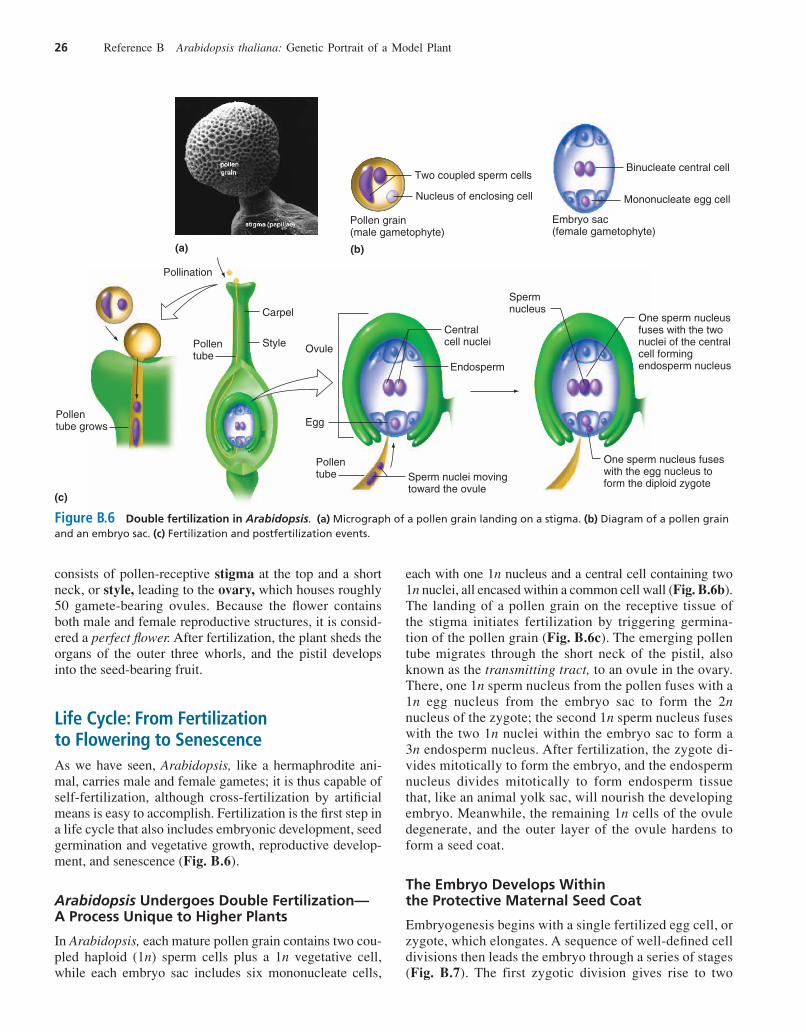
Life Cycle: From Fertilization to Flowering to SenescenceAs we have seen, Arabidopsis, like a hermaphrodite ani-mal, carries male and female gametes; it is thus capable ofself-fertilization, although cross-fertilization by artificialmeans is easy to accomplish. Fertilization is the first step ina life cycle that also includes embryonic development, seedgermination and vegetative growth, reproductive develop-ment, and senescence (Fig. B.6).
Arabidopsis Undergoes Double Fertilization—A Process Unique to Higher Plants
In Arabidopsis, each mature pollen grain contains two cou-pled haploid (1n) sperm cells plus a 1n vegetative cell,while each embryo sac includes six mononucleate cells,
each with one 1n nucleus and a central cell containing two1n nuclei, all encased within a common cell wall (Fig. B.6b).The landing of a pollen grain on the receptive tissue ofthe stigma initiates fertilization by triggering germina-tion of the pollen grain (Fig. B.6c). The emerging pollentube migrates through the short neck of the pistil, alsoknown as the transmitting tract, to an ovule in the ovary.There, one 1n sperm nucleus from the pollen fuses with a1n egg nucleus from the embryo sac to form the 2nnucleus of the zygote; the second 1n sperm nucleus fuseswith the two 1n nuclei within the embryo sac to form a3n endosperm nucleus. After fertilization, the zygote di-vides mitotically to form the embryo, and the endospermnucleus divides mitotically to form endosperm tissuethat, like an animal yolk sac, will nourish the developingembryo. Meanwhile, the remaining 1n cells of the ovuledegenerate, and the outer layer of the ovule hardens toform a seed coat.
The Embryo Develops Within the Protective Maternal Seed Coat
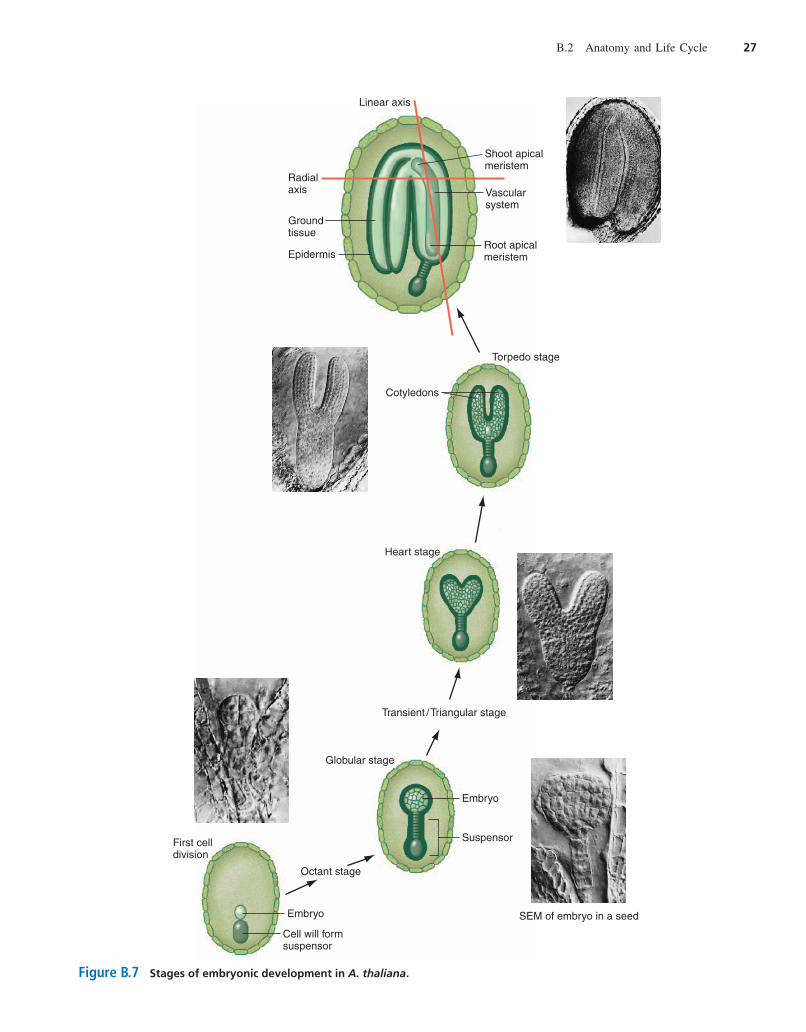
Embryogenesis begins with a single fertilized egg cell, orzygote, which elongates. A sequence of well-defined celldivisions then leads the embryo through a series of stages(Fig. B.7). The first zygotic division gives rise to two
26 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
Pollination
Pollen tube grows
Pollen tube
Carpel
Endosperm
Ovule
Central cell nuclei
Egg
Sperm nuclei moving toward the ovule
Pollentube
Sperm nucleus
One sperm nucleus fuses with the egg nucleus to form the diploid zygote
One sperm nucleus fuses with the two nuclei of the central cell forming endosperm nucleus
Style
Embryo sac(female gametophyte)
Mononucleate egg cell
Binucleate central cell
Pollen grain(male gametophyte)
Two coupled sperm cells
Nucleus of enclosing cell
(a) (b)
(c)
Figure B.6 Double fertilization in Arabidopsis. (a) Micrograph of a pollen grain landing on a stigma. (b) Diagram of a pollen grainand an embryo sac. (c) Fertilization and postfertilization events.
har06584_refB_021-048 11/06/2006 03:00 PM Page 26
B.2 Anatomy and Life Cycle 27
Embryo
Embryo
Cell will formsuspensor
First celldivision
Globular stage
Suspensor
SEM of embryo in a seed
Heart stage
Torpedo stage
Cotyledons
Linear axis
Radialaxis
Root apicalmeristem
Shoot apicalmeristem
Vascularsystem
Ground tissue
Epidermis
Transient /Triangular stage
Octant stage
Figure B.7 Stages of embryonic development in A. thaliana.
har06584_refB_021-048 11/06/2006 03:00 PM Page 27

cells, one small and one larger. The small cell constitutesthe embryo proper, while the larger cell will divide onlya few more times to produce the suspensor, a structureanalogous to the umbilical cord in mammals (whose di-visions we will not describe here). The embryo dividesfollowing a plane at right angles to the first plane of divi-sion to create a vertical wall and two side-by-side cells.The next division, which is at from the previous one,generates four cells that can be seen end-on from the top.The third embryonic division is horizontal and makes aneight-cell, or octant stage, embryo with four cells aboveand four below. Additional divisions parallel to the sur-face of the embryo result in the formation of a discreteouter cell layer that will give rise to the epidermis of themature embryo, and an innermost group of cells thatacquire an elongated shape, the first evidence of differen-tiation toward the vasculature found in the mature embryo.The embryo has now reached the globular stage. Theglobular embryo rapidly loses its globular shape andacquires a more triangular form that defines the transient
or triangular stage embryo, at which point the embryobecomes self-sufficient for growth, having used up mostof its maternally deposited reserves. Next, the embryomoves through the heart stage to the torpedo stage astwo protuberances expand and differentiate into twowell-defined, discrete cotyledons.
By the end of this progression from octant to torpedostage, cellular differentiation has established all the majortissues required for the future vegetative development ofthe plant along two main axes: the linear axis that makesup the shoot/root continuum and a radial axis moving fromthe center out to the edge of that continuum (Fig. B.7). Theshoot/root axis includes two cotyledons, which store nutri-ents in the mature seed and will differentiate into the firstrudimentary leaves after germination. The axis also in-cludes a hypocotyl, which is a short section of embryonicstem between the two cotyledons and the embryonic root,and the root and shoot meristems. The radial axis carriesthe central vascular system, a middle layer of ground tis-sues, and the outer epidermis.
As the embryo reaches maturity, growth and develop-ment cease and desiccation (water loss) occurs. When theloss of water becomes extreme enough, the embryo entersa dormant state in which virtually all metabolic activityceases. An embryo in the dormant state can withstand ad-verse environmental conditions such as winter in Madison,Wisconsin, or Buffalo, New York.
Favorable Environmental Conditions TriggerSeed Germination and Vegetative Growth
Under appropriate conditions, the Arabidopsis embryoabsorbs water and rehydrates within the seed, which en-ables growth and development to resume. The enlargingembryo bursts out of the seed coat and begins to grow
90°
along its vertical axis. At this point, environmental cuesinfluence the form the seedling will take. Both the rootand the shoot can sense gravity; the root responds bybending toward it while the shoot bends away from it.Light provides another developmental cue. If germinationoccurs in the dark, for example, under the surface of thesoil, the hypocotyl, or embryonic stem, elongates rapidly;but development of the leaflike cotyledons and activity ofthe shoot apical meristem are suppressed. The elongatinghypocotyl is hooked near its apex, an adaptation that en-ables the seedling to push its way up through the soilwhile protecting the delicate meristem. When the elongat-ing hypocotyl senses light, elongation ceases, the apicalhook opens, and the cotyledons expand and turn green aschloroplasts begin to develop and establish the potentialfor self-nourishment through photosynthesis (Fig. B.8a).This light-regulated developmental program is calledphotomorphogenesis.
With the establishment of the self-nourishing autotrophiclifestyle, the shoot apical meristem produces a compactspiral, or rosette, of leaves. Early flowering varieties ofArabidopsis may only produce 6–10 leaves before the shootapical meristem converts to reproductive development. Later-flowering varieties may produce a rosette containing dozensof leaves before flowering begins (Fig. B.8b).
Reproductive Development Begins When the Leaf-Producing Apical Meristem Switches to a Flower-Producing Meristem
Under the right conditions, the shoot meristem enlarges andswitches from producing only cells that differentiate intoleaves to producing floral meristems on its flanks. At thispoint, the shoot apical meristem becomes an inflorescence
meristem. The floral meristems arise in a spiral around theinflorescence meristem and, in turn, give rise to flower pri-mordia that differentiate into the four whorls of organs thatmake up a flower. Initiated one at a time, the flowers developsequentially. Thus a view from the apex of the inflorescencemeristem reveals the progression of floral developmentalstages spiraling away from the shoot apex. Elongation of themain stem between each flower ultimately produces the in-florescence branch containing about 35 flowers (Fig. B.9).
Both environmental cues and endogenous develop-mental signals (resulting from the genetic program)control the timing of the switch from vegetative to repro-ductive development in Arabidopsis. Day length, orphotoperiod, is a particularly important regulator offlowering. Laboratory strains, such as Columbia andLandsberg erecta, when grown under constant light or along, 16-hour day, will flower after producing fewer than10 rosette leaves. When grown in an 8-hour photoperiod,these same strains will produce over 30 leaves beforeflowering. Another significant environmental signal istemperature. If exposed to extended periods of low tem-perature during early vegetative development, normally
28 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
har06584_refB_021-048 11/06/2006 03:00 PM Page 28

late-flowering varieties of Arabidopsis will flower early.This phenomenon of exposing plants to the cold early invegetative development is known as vernalization.
Since neither photoperiod stimulation nor vernaliza-tion is strictly necessary for flowering, researchers inferred
that an endogenous developmental clock is active in thisprocess. However, this endogenous program remainspoorly characterized.
Senescence: The Vegetative Plant Ages and Dies
Because plants can produce new organ systems continu-ously, they do not necessarily display the finite life spanscharacteristic of animal species. However, some plants,including Arabidopsis, do senesce and die after a singlereproductive effort. In Arabidopsis, somatic tissues suchas leaves and stems senesce a few weeks after they areinitiated at the meristem. In leaves, senescence is a ge-netically programmed process in which yellowing occursas salvage systems convert cellular constituents into mo-bile nutrients for transport out of the aging leaf (Fig. B.9).The limited longevity of individual leaves does not limitthe life span of the plant in the vegetative phase becausethe shoot apical meristem continuously produces newleaves. Rather, it is the conversion of the shoot apicalmeristem to reproductive development that delimits theproduction of new photosynthetic organs. In addition, theinflorescence meristem ceases proliferative activity aftera set number of seeds have been produced. Seeds pro-duced by self-pollination are shed from the parent plant,which undergoes senescence. Quick reproduction, theproduction of seeds by self-pollination, and the lack ofwide seed dispersal serve the plant’s strategy in naturalsettings: Arabidopsis is an ephemeral that exploits dis-turbed soils and completes its life cycle quickly to reseedthe soil near the parent plant.
B.2 Anatomy and Life Cycle 29
(a)
(b)
Figure B.8 Emerging shoot in A. thaliana. (a) The embryonicstem reaches for the surface of the soil (right seedling), where theapical hook will open, allowing the greening of the cotyledons (leftseedling). (b) In early-flowering varieties, the shoot apical meristemmay produce only a few leaves before developing flowers (left).Late-flowering varieties may produce dozens of leaves before flow-ering (right). No flowers are visible here, but the stem growing upat the center of each plant represents the primary inflorescence, orflowering stem. The late-flowering plant on the right is consider-ably older than the early-flowering plant on the left.
Figure B.9 Reproductive development and senescence inA. thaliana. Young flowering plants next to older plants under-going senescence.
har06584_refB_021-048 11/06/2006 03:00 PM Page 29

B.3 Techniques of Mutational Analysis
Researchers have used various techniques to identify muta-tions affecting many aspects of Arabidopsis anatomy anddevelopment. Some of the mutations characterized occurspontaneously in natural populations, but as described inChapter 7 of the main textbook, these spontaneous muta-tions appear at a low rate. Geneticists use mutagens toincrease their frequency. The means of inducing mutationsin Arabidopsis include exposure to mutagenic chemicals,exposure to radiation, and insertional mutagenesis.
Mutagenesis by Chemical and Irradiation ProceduresA mature seed carries a dormant embryo in which one tothree cells from the apical meristem are destined to formthe germ line of the mature plant, which will give rise tothe gametophytes. If a mutation is to be transmitted toprogeny, it has to affect one of the two alleles carried byany of these cells. The phenotype derived from a reces-sive mutation induced by seed mutagenesis in Arabidop-sis will segregate in different ratios depending on howmany gametophyte progenitor cells are present in theembryo at the time of mutagenesis. To be recovered, in-duced mutations must pass through the male and femalegametophytes, which are multicellular, genetically ac-tive, haploid structures. The gametophyte stage filters outmutations in genes that are required for gametophyte vi-ability. For this reason, most large deletions cannot bytransmitted through a plant’s gemetophytes. In contrast,deletions are often transmitted through animal gametes.This difference explains why the use of deletion muta-tion panels, a common practice in animal genetics, is notpossible in plant genetics. In practice, screens or selec-tions to identify Arabidopsis mutants generally focus oninvestigation of the M2 generation that is derived fromself-pollinated seeds harvested from plants that developfrom mutagenized seed (M1). In the M2 generation, theinduced alleles are segregating, which allows homozy-gotes to be identified and recovered.
Arabidopsis Researchers Use Two Types ofInsertional Mutagenesis: Transformation by T-DNA and Transposon TaggingStrategies for insertional mutagenesis depend on the abilityof certain classes of DNA molecules to integrate into theArabidopsis genome. If an integration disrupts a gene, itrepresents a mutation.
In the T-DNA Approach, the BacteriumAgrobacterium tumefaciens, Which Causes a Type of Benign Tumor in Plants, Is the Agent of Transformation
A. tumefaciens can transfer a piece of plasmid DNAknown as the T-DNA into the genome of wounded plantcells (Fig. B.10a).
Experimenters can modify the T-DNA to eliminateits oncogenes and insert transformational marker genesthat make the plant resistant to antibiotics (suchas kanamycin and hygromycin) or herbicides (such aschlorsulfuron). In one protocol using modified T-DNA,Arabidopsis is transformed by soaking the shoots of ma-ture three-week-old seedlings or germinating seedlingsin an Agrobacterium culture to which a strong vacuum ora weak detergent has been applied. The vacuum or deter-gent enable the bacteria to infiltrate the shoots. As thebacterium-infiltrated plants grow and eventually self-pollinate, a few gametes or zygotes are transformed byAgrobacterium, and these produce a transformed seedthat is resistant to antibiotics or herbicides. This resist-ance provides a means of selecting for transformedplants that have inserted the transgenic DNA into theirgenomes. Because the A. tumefaciens T-DNA integratesmore or less randomly into the plant genome, it has thepotential to serve as a mutagenic agent for any gene.
Initially, researchers used insertional T-DNA muta-genesis to create collections of independent transgenicArabidopsis lines, which were screened for specific mor-phological, developmental, or biochemical phenotypes.Subsequently, several large-scale projects sought to assem-ble sequence-indexed T-DNA mutant collections by deter-mining the nucleotide sequence of regions adjacent toindividual insertions. As of 2006, there were over 350,000different T-DNA insertions mapped at the nucleotide level,covering a large majority of the genes in the Arabidopsisgenome. Arabidopsis geneticists can now order from pub-lic stock centers T-DNA insertion alleles in almost anygene to begin reverse genetic analyses. Researchers havealso developed a second generation of mutagenesis toolsbased on T-DNA insertions. One variation uses a T-DNAloaded with strong transcriptional enhancer elements toamplify the expression of genes close to the insertion site.This approach, called “activation tagging,” allows the iden-tification gain-of-function alleles of redundant genes thatmay go undetected in a traditional loss-of-function muta-tion screen. Another T-DNA mutagenesis scheme uses areporter gene, such as GUS, with an incomplete promoterlacking an upstream enhancer element. GUS encodes theenzyme �-glucuronidase, which converts a colorless solu-ble compound (X-Glu) into a blue insoluble product. Cellsexpressing GUS stain blue. Because the GUS gene in themodified T-DNA has an incomplete promoter, its expres-sion depends on the insertion of the T-DNA into a plant-coding sequence; when that happens, the regulatory region
30 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
har06584_refB_021-048 11/06/2006 03:00 PM Page 30
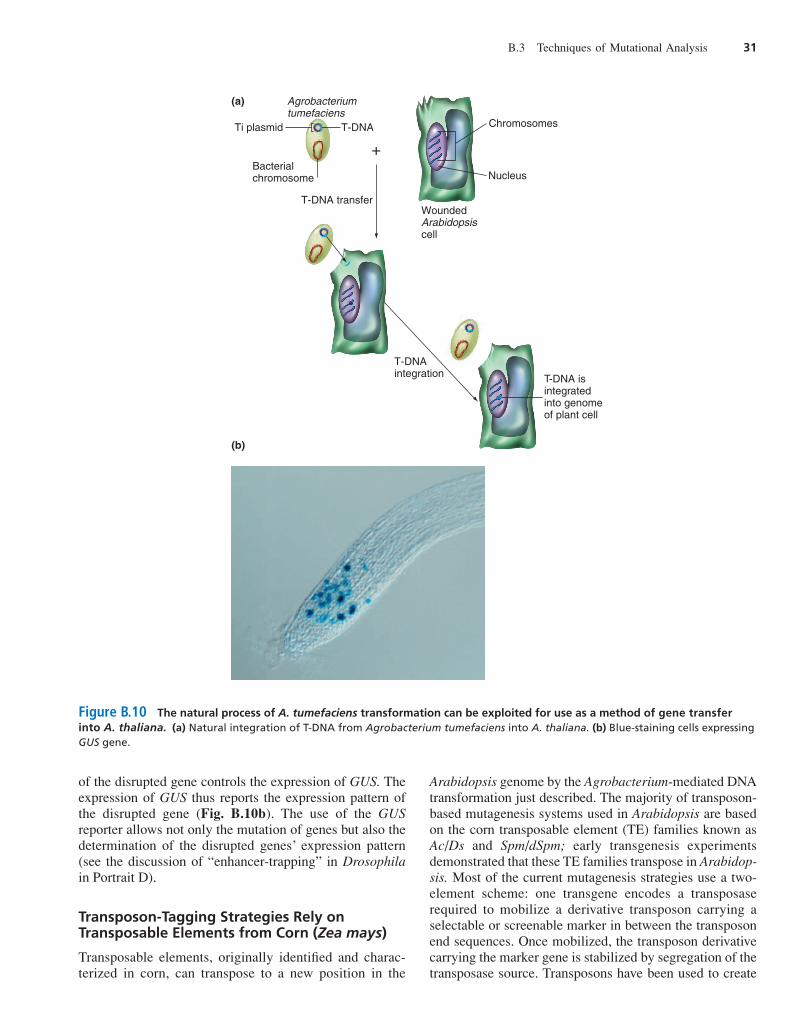
of the disrupted gene controls the expression of GUS. Theexpression of GUS thus reports the expression pattern ofthe disrupted gene (Fig. B.10b). The use of the GUSreporter allows not only the mutation of genes but also thedetermination of the disrupted genes’ expression pattern(see the discussion of “enhancer-trapping” in Drosophilain Portrait D).
Transposon-Tagging Strategies Rely onTransposable Elements from Corn (Zea mays)
Transposable elements, originally identified and charac-terized in corn, can transpose to a new position in the
Arabidopsis genome by the Agrobacterium-mediated DNAtransformation just described. The majority of transposon-based mutagenesis systems used in Arabidopsis are basedon the corn transposable element (TE) families known asAc/Ds and Spm/dSpm; early transgenesis experimentsdemonstrated that these TE families transpose in Arabidop-sis. Most of the current mutagenesis strategies use a two-element scheme: one transgene encodes a transposaserequired to mobilize a derivative transposon carrying aselectable or screenable marker in between the transposonend sequences. Once mobilized, the transposon derivativecarrying the marker gene is stabilized by segregation of thetransposase source. Transposons have been used to create
B.3 Techniques of Mutational Analysis 31
+
Agrobacteriumtumefaciens
T-DNA transfer
Bacterialchromosome
Ti plasmid
(a)
(b)
T-DNA integration
T-DNA
Wounded Arabidopsiscell
Nucleus
Chromosomes
T-DNA is integrated into genome of plant cell
Figure B.10 The natural process of A. tumefaciens transformation can be exploited for use as a method of gene transferinto A. thaliana. (a) Natural integration of T-DNA from Agrobacterium tumefaciens into A. thaliana. (b) Blue-staining cells expressingGUS gene.
har06584_refB_021-048 11/06/2006 03:00 PM Page 31
populations carrying simple insertional alleles. In addition,modified transposon elements have been engineered togenerate alleles in which the reporter gene incorporatedwithin the mobile element falls under the control of localenhancer elements (enhancer trap) or lands within a gene togenerate a transcriptional fusion (gene trap).
Insertional Mutagenesis Not Only Allows the Generation of Mutations but AlsoFacilitates Their Molecular Characterization
One advantage of insertional mutagenesis over chemicaland radiation procedures is that insertional mutagenesisproduces mutated alleles carrying a specific DNA insert(the T-DNA or the transposable element). A copy of thespecific T-DNA or transposable element that becamethe insert can therefore serve as a probe to identify andclone the gene the insert disrupts. With the use of enhanceror gene trap systems, the insertions can also indicate theexpression pattern of the tagged gene.
From Gene to Phenotype: AnalyzingMutations to Identify Gene FunctionThe genome sequencing project has identified many geneswith specific sequences, but the sequence alone is often notsufficient to attribute a specific function to a particulargene. One way to solve this problem is to mutate the geneunder study and analyze the phenotype of plants homozy-gous for the mutation. Since knocking out genes by homol-ogous recombination is not very efficient in plants,Arabidopsis researchers increasingly rely on sequence-indexed insertional mutant libraries. Another approach isthe use of gene silencing to suppress the expression ofgenes of interest.
Silencing Specific Genes with Antisense siRNA
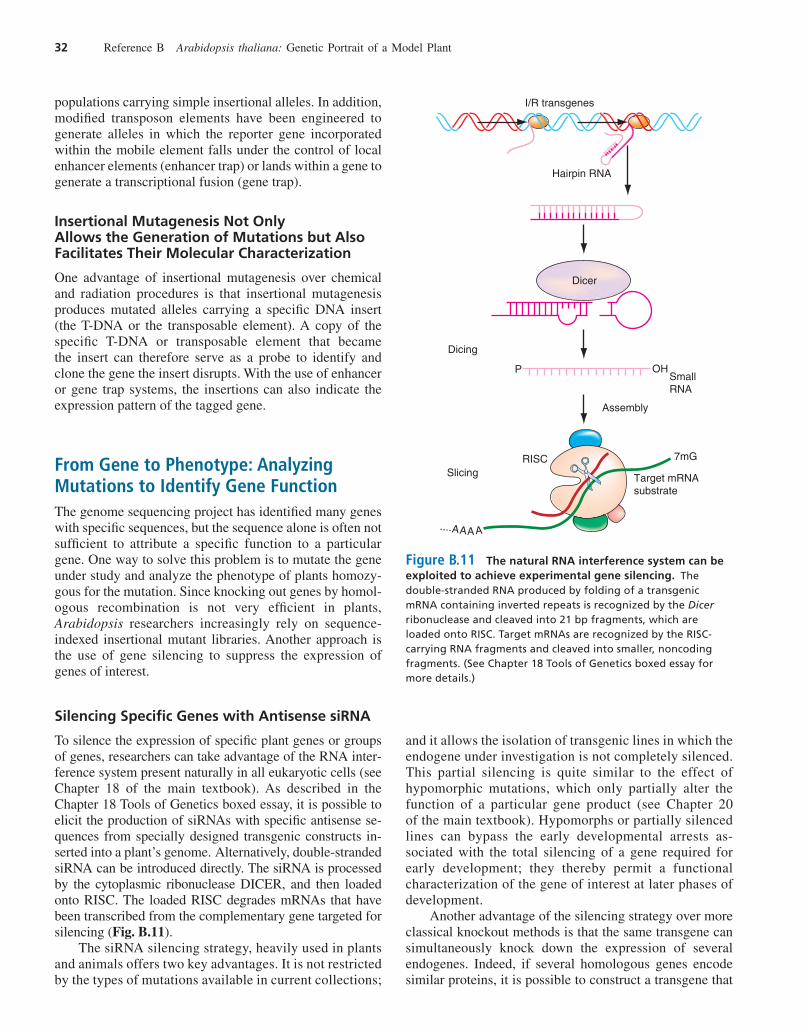
To silence the expression of specific plant genes or groupsof genes, researchers can take advantage of the RNA inter-ference system present naturally in all eukaryotic cells (seeChapter 18 of the main textbook). As described in theChapter 18 Tools of Genetics boxed essay, it is possible toelicit the production of siRNAs with specific antisense se-quences from specially designed transgenic constructs in-serted into a plant’s genome. Alternatively, double-strandedsiRNA can be introduced directly. The siRNA is processedby the cytoplasmic ribonuclease DICER, and then loadedonto RISC. The loaded RISC degrades mRNAs that havebeen transcribed from the complementary gene targeted forsilencing (Fig. B.11).
The siRNA silencing strategy, heavily used in plantsand animals offers two key advantages. It is not restrictedby the types of mutations available in current collections;
and it allows the isolation of transgenic lines in which theendogene under investigation is not completely silenced.This partial silencing is quite similar to the effect ofhypomorphic mutations, which only partially alter thefunction of a particular gene product (see Chapter 20of the main textbook). Hypomorphs or partially silencedlines can bypass the early developmental arrests as-sociated with the total silencing of a gene required forearly development; they thereby permit a functionalcharacterization of the gene of interest at later phases ofdevelopment.
Another advantage of the silencing strategy over moreclassical knockout methods is that the same transgene cansimultaneously knock down the expression of severalendogenes. Indeed, if several homologous genes encodesimilar proteins, it is possible to construct a transgene that
32 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
I/R transgenes
Hairpin RNA
OH
RISCSlicing
AAAA....
7mG
Target mRNAsubstrate
PSmallRNA
Assembly
Dicing
Dicer
Figure B.11 The natural RNA interference system can beexploited to achieve experimental gene silencing. Thedouble-stranded RNA produced by folding of a transgenicmRNA containing inverted repeats is recognized by the Dicerribonuclease and cleaved into 21 bp fragments, which areloaded onto RISC. Target mRNAs are recognized by the RISC-carrying RNA fragments and cleaved into smaller, noncodingfragments. (See Chapter 18 Tools of Genetics boxed essay formore details.)
har06584_refB_021-048 11/06/2006 09:26 PM Page 32

expresses an inverted duplication of the DNA fragmentconserved between these genes. When introduced intoplants, such a transgene will silence all genes that encodemRNAs sharing nucleotide similarity with it. This elimi-nates the need to isolate numerous mutations in each one ofthese genes, and to develop a complex scheme of crossesaimed at bringing together in the same plant multiple mu-tations in members of the gene family under investigation.This strategy makes it possible to determine the phenotypeassociated with the disappearance or reduction of a specificprotein function that is encoded by multiple, functionallyredundant genes.
Insertional mutagenesis and gene-silencing strategiesfor moving from gene to phenotype have found wide usenow that the Arabidopsis genome has been completely se-quenced. Indeed, geneticists currently not only identify thegenes involved in specific functions, they also use theirknowledge of gene sequences and mutation-induced phe-notypes to define what specific genes do.
B.4 The Genetic Analysis ofDevelopment in Arabidopsis
Although geneticists only partially understand the mech-anisms of development in plants and animals, they doknow that instructions for the formation of a body planand the elaboration of specific functions are ultimatelyencoded in the DNA. Mutations can thus change theposition, structure, or function of an organ or alter anindividual’s orderly passage through the life cycle. Wenow look at the use of genetic analysis in the study ofembryogenesis, hormone control systems, and responsesto environmental signals.
The Genetic Analysis of EmbryogenesisMutations that are lethal to the embryo or that arrest em-bryonic development have provided a basis for understand-ing embryonic development in Arabidopsis. Screenings formutations resulting in developmental arrest at a specificstage of embryogenesis have led to the identification ofmore than 500 embryo-lethal and embryo-defective muta-tions. A significant portion of the embryo-lethal mutantsare arrested at the transient stage. Many of them carrymutations in housekeeping genes.
Researchers Have Found Very Few Maternal-Effect Mutations in Arabidopsis
Such mutations occur in maternal genes whose proteinproducts are deposited in the egg during oogenesis.
Maternal-effect mutations disrupt embryogenesis in allprogeny of a mutant mother, regardless of the zygote’sgenotype.
The small number of maternal-effect mutations con-stitutes a major difference between plants and certainkinds of animals, such as Drosophila, that develop fromlarge eggs. (In Drosophila, numerous maternal-effect mu-tations alter the early stages of embryogenesis, as de-scribed in Portrait D.) A major reason for the nearabsence of maternal effects in Arabidopsis is that theplant’s embryonic development does not depend on alarge cytoplasm filled with maternal products (as happensin Drosophila). Instead, it depends from a very early stageon a cytoplasm newly produced from the zygote’s owngenome (as in mammals).
Also unlike what happens in Drosophila, in Arabidopsis,very few zygotic mutations (that is, mutations in the zy-gote’s own genes) affect embryogenesis before the globularstage. At least part of this difference between Arabidopsisand Drosophila may derive from the fact that in plantsmany zygotic genes are expressed very early in embryo-genesis (as they are in mammals), and these zygotic genesmay encode protein products that are functionally redun-dant with those of maternal origin. In addition, many plantgenes are members of multigene families in which eachmember encodes a protein whose function duplicates thatof some of the other members. Hence, as in mammals, thescarcity of maternal-effect and early zygotic mutations inplants may simply reflect a large amount of functionalredundancy between maternally deposited and zygoticallyderived products.
Screenings for Mutations That ArrestDevelopment at a Specific Stage ofEmbryogenesis Enable the Identification of Important Regulatory Processes in Plant Development
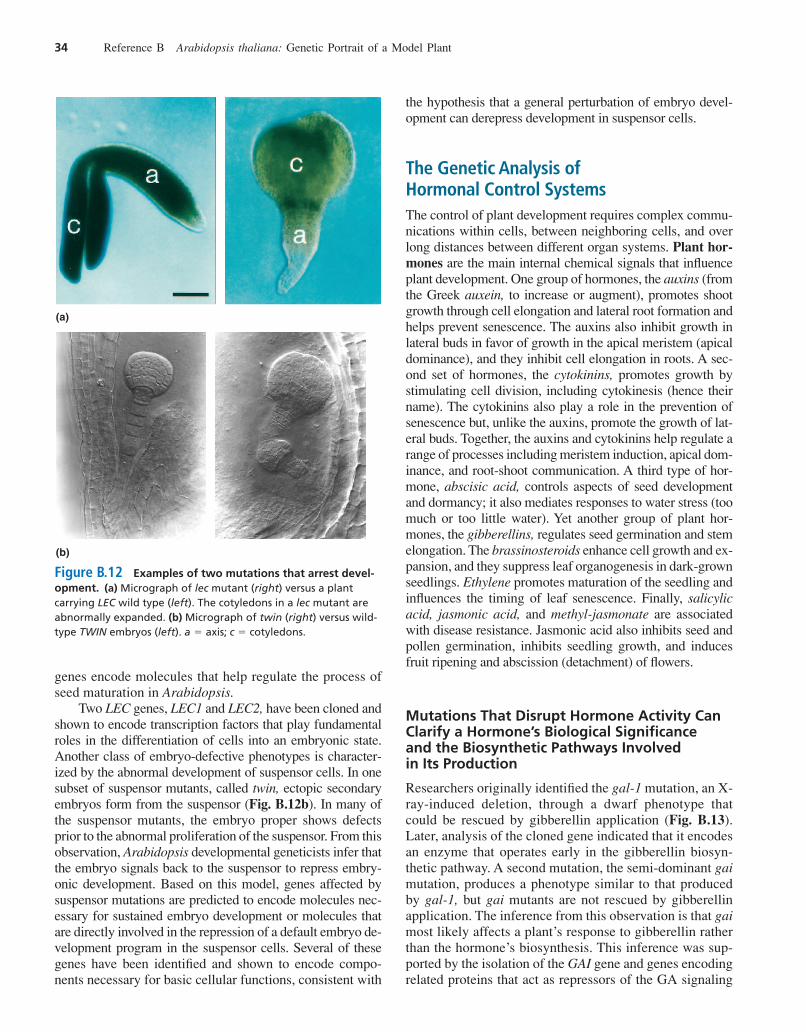
A large number of mutations affecting Arabidopsis em-bryo development have been isolated. We will highlightjust two of these mutations to give a flavor of range of phe-notypes and mechanisms involved. One type of embryo-defective mutation known as leafy cotyledon (lec) resultsin embryos with cotyledons resembling mature leavesrather than wild-type cotyledons (Fig. B.12a). As de-scribed earlier, cotyledons are embryonic structures thatfirst appear late in the globular stage; leaves, by contrast,are products of the apical meristem that appear only aftergermination. Unlike cotyledons, leaves have trichomes—small, hairlike projections from the epidermis—and a leaf-specific pattern of vascularization. In addition to causingthe conversion of cotyledons into leaves, lec mutationsproduce seeds unable to germinate because they cannotwithstand the desiccation that accompanies seed matura-tion. These observations suggest that the wild-type LEC
B.4 The Genetic Analysis of Development in Arabidopsis 33
har06584_refB_021-048 11/06/2006 03:00 PM Page 33
genes encode molecules that help regulate the process ofseed maturation in Arabidopsis.
Two LEC genes, LEC1 and LEC2, have been cloned andshown to encode transcription factors that play fundamentalroles in the differentiation of cells into an embryonic state.Another class of embryo-defective phenotypes is character-ized by the abnormal development of suspensor cells. In onesubset of suspensor mutants, called twin, ectopic secondaryembryos form from the suspensor (Fig. B.12b). In many ofthe suspensor mutants, the embryo proper shows defectsprior to the abnormal proliferation of the suspensor. From thisobservation, Arabidopsis developmental geneticists infer thatthe embryo signals back to the suspensor to repress embry-onic development. Based on this model, genes affected bysuspensor mutations are predicted to encode molecules nec-essary for sustained embryo development or molecules thatare directly involved in the repression of a default embryo de-velopment program in the suspensor cells. Several of thesegenes have been identified and shown to encode compo-nents necessary for basic cellular functions, consistent with
the hypothesis that a general perturbation of embryo devel-opment can derepress development in suspensor cells.
The Genetic Analysis of Hormonal Control SystemsThe control of plant development requires complex commu-nications within cells, between neighboring cells, and overlong distances between different organ systems. Plant hor-
mones are the main internal chemical signals that influenceplant development. One group of hormones, the auxins (fromthe Greek auxein, to increase or augment), promotes shootgrowth through cell elongation and lateral root formation andhelps prevent senescence. The auxins also inhibit growth inlateral buds in favor of growth in the apical meristem (apicaldominance), and they inhibit cell elongation in roots. A sec-ond set of hormones, the cytokinins, promotes growth bystimulating cell division, including cytokinesis (hence theirname). The cytokinins also play a role in the prevention ofsenescence but, unlike the auxins, promote the growth of lat-eral buds. Together, the auxins and cytokinins help regulate arange of processes including meristem induction, apical dom-inance, and root-shoot communication. A third type of hor-mone, abscisic acid, controls aspects of seed developmentand dormancy; it also mediates responses to water stress (toomuch or too little water). Yet another group of plant hor-mones, the gibberellins, regulates seed germination and stemelongation. The brassinosteroids enhance cell growth and ex-pansion, and they suppress leaf organogenesis in dark-grownseedlings. Ethylene promotes maturation of the seedling andinfluences the timing of leaf senescence. Finally, salicylicacid, jasmonic acid, and methyl-jasmonate are associatedwith disease resistance. Jasmonic acid also inhibits seed andpollen germination, inhibits seedling growth, and inducesfruit ripening and abscission (detachment) of flowers.
Mutations That Disrupt Hormone Activity CanClarify a Hormone’s Biological Significance and the Biosynthetic Pathways Involved in Its Production
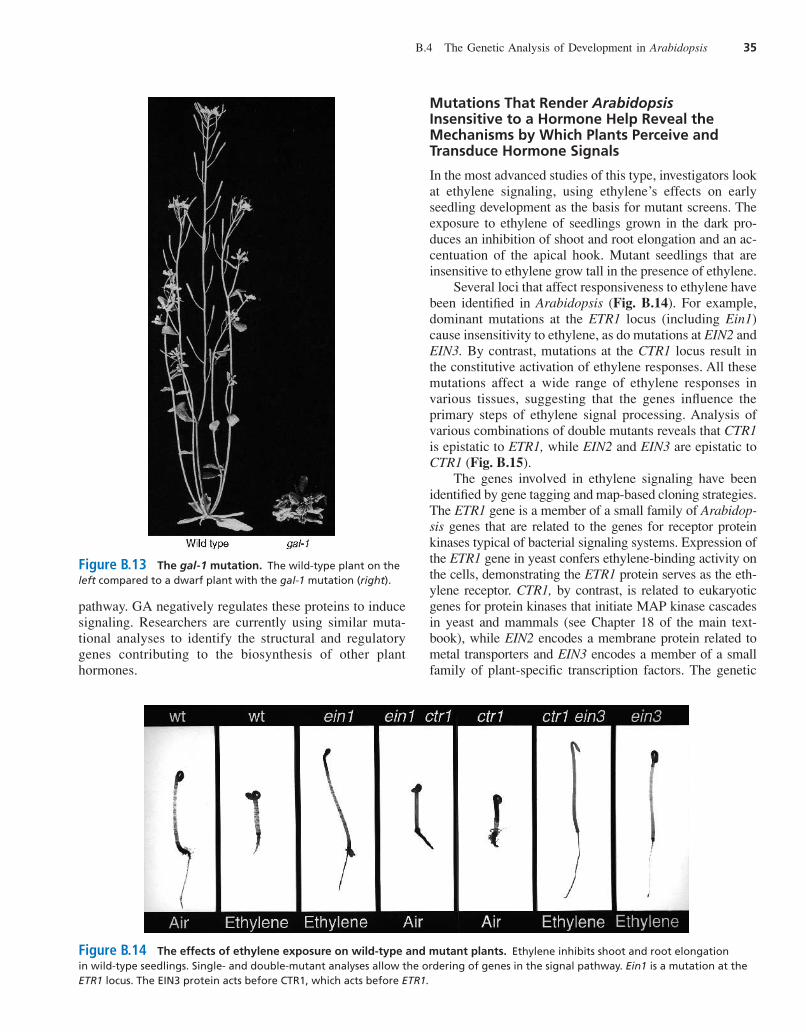
Researchers originally identified the gal-1 mutation, an X-ray-induced deletion, through a dwarf phenotype thatcould be rescued by gibberellin application (Fig. B.13).Later, analysis of the cloned gene indicated that it encodesan enzyme that operates early in the gibberellin biosyn-thetic pathway. A second mutation, the semi-dominant gaimutation, produces a phenotype similar to that producedby gal-1, but gai mutants are not rescued by gibberellinapplication. The inference from this observation is that gaimost likely affects a plant’s response to gibberellin ratherthan the hormone’s biosynthesis. This inference was sup-ported by the isolation of the GAI gene and genes encodingrelated proteins that act as repressors of the GA signaling
34 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
Figure B.12 Examples of two mutations that arrest devel-opment. (a) Micrograph of lec mutant (right) versus a plantcarrying LEC wild type (left). The cotyledons in a lec mutant areabnormally expanded. (b) Micrograph of twin (right) versus wild-type TWIN embryos (left). a � axis; c � cotyledons.
(a)
(b)
har06584_refB_021-048 11/06/2006 03:00 PM Page 34
pathway. GA negatively regulates these proteins to inducesignaling. Researchers are currently using similar muta-tional analyses to identify the structural and regulatorygenes contributing to the biosynthesis of other planthormones.
Mutations That Render ArabidopsisInsensitive to a Hormone Help Reveal theMechanisms by Which Plants Perceive andTransduce Hormone Signals
In the most advanced studies of this type, investigators lookat ethylene signaling, using ethylene’s effects on earlyseedling development as the basis for mutant screens. Theexposure to ethylene of seedlings grown in the dark pro-duces an inhibition of shoot and root elongation and an ac-centuation of the apical hook. Mutant seedlings that areinsensitive to ethylene grow tall in the presence of ethylene.
Several loci that affect responsiveness to ethylene havebeen identified in Arabidopsis (Fig. B.14). For example,dominant mutations at the ETR1 locus (including Ein1)cause insensitivity to ethylene, as do mutations at EIN2 andEIN3. By contrast, mutations at the CTR1 locus result inthe constitutive activation of ethylene responses. All thesemutations affect a wide range of ethylene responses invarious tissues, suggesting that the genes influence theprimary steps of ethylene signal processing. Analysis ofvarious combinations of double mutants reveals that CTR1is epistatic to ETR1, while EIN2 and EIN3 are epistatic toCTR1 (Fig. B.15).
The genes involved in ethylene signaling have beenidentified by gene tagging and map-based cloning strategies.The ETR1 gene is a member of a small family of Arabidop-sis genes that are related to the genes for receptor proteinkinases typical of bacterial signaling systems. Expression ofthe ETR1 gene in yeast confers ethylene-binding activity onthe cells, demonstrating the ETR1 protein serves as the eth-ylene receptor. CTR1, by contrast, is related to eukaryoticgenes for protein kinases that initiate MAP kinase cascadesin yeast and mammals (see Chapter 18 of the main text-book), while EIN2 encodes a membrane protein related tometal transporters and EIN3 encodes a member of a smallfamily of plant-specific transcription factors. The genetic
B.4 The Genetic Analysis of Development in Arabidopsis 35
Figure B.13 The gal-1 mutation. The wild-type plant on theleft compared to a dwarf plant with the gal-1 mutation (right).
Figure B.14 The effects of ethylene exposure on wild-type and mutant plants. Ethylene inhibits shoot and root elongationin wild-type seedlings. Single- and double-mutant analyses allow the ordering of genes in the signal pathway. Ein1 is a mutation at theETR1 locus. The EIN3 protein acts before CTR1, which acts before ETR1.
har06584_refB_021-048 11/06/2006 03:00 PM Page 35

36 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
CTR1 EIN2EIN3EIL1EIL2
ERF1 Responses
ETR1ETR2ERS1ERS2EIN4
C2H4
ETR-FAMILY
Two-componentregulators
CTR1
RAF-likekinase
Map-kinasecascade EIN2
Nramp metaltransporter
EIN3/EIL
transcriptionfactors
ERF1
EREBPtranscriptionfactor
A A A
Responses
A A A
Figure B.15 The genetic interactions and biochemical identities of components of the ethylene signal transduction pathway.Progressing from left to right, ethylene is thought to regulate negatively a family of membrane-associated receptors. The histidine-kinase transmitter domains of members of the receptor family interact with the regulatory domain of the Raf-like kinase CTR1. Thereceptor/CTR1 complex negatively regulates a membrane protein (EIN2) related to a superfamily of metal transporters. The cytoplasmicC-terminal domain of EIN2 positively signals downstream to the EIN3 family of transcription factors. ERF1 is rapidly induced in responseto ethylene and is capable of activating a subset of ethylene responses when expressed.
analysis of this pathway has identified components of a com-plex signaling pathway that leads from signal perception atthe membrane to a transcriptional cascade in the nucleus.
Arabidopsis mutants that are insensitive to other planthormones have also been isolated; the gai mutant discussedearlier is an example. The isolation of genes affected in thevarious mutants demonstrates that protein phosphatases,farnesyl transferases, transcription factors, and the ubiqui-tin protein degradation pathway play a role in plant hor-mone signaling, while genes for membrane-basedpermeases and transporters contribute to hormone trans-port. The fact that many of the genes identified are homol-ogous to genes in yeast and mammals is evidence that thebasic information-processing systems arose very early inthe evolution of eukaryotes. On the other hand, receptorsfor ethylene, cytokinins, and red light (the phytochromes)are related to bacterial sensory receptors. These systemswere likely inherited from the bacterial endosymbiont an-cestor of the chloroplast.
The Genetic Analysis ofPhotomorphogenesis—The Regulation of Growth and Development in Responseto Lighting CuesDuring growth and development, plants continuously altertheir responses to changing environmental signals. One ofthe most dramatic series of developmental changes takes
place when the seedling grows up through the soil andencounters light. Collectively known as photomorphogene-sis, these changes include the opening of the apical hookand the generation of chloroplasts. Two classes of photore-ceptors—phytochromes and cryptochromes—mediate thechanges. Receptors in the phytochrome family sense lightprimarily in the red and far-red wavelengths. Receptorsin the cryptochrome family perceive blue wavelengthsof light.
Mutational Analyses Have Helped Characterize the Photoreceptor Molecules by Which Plants Receive Light Signals
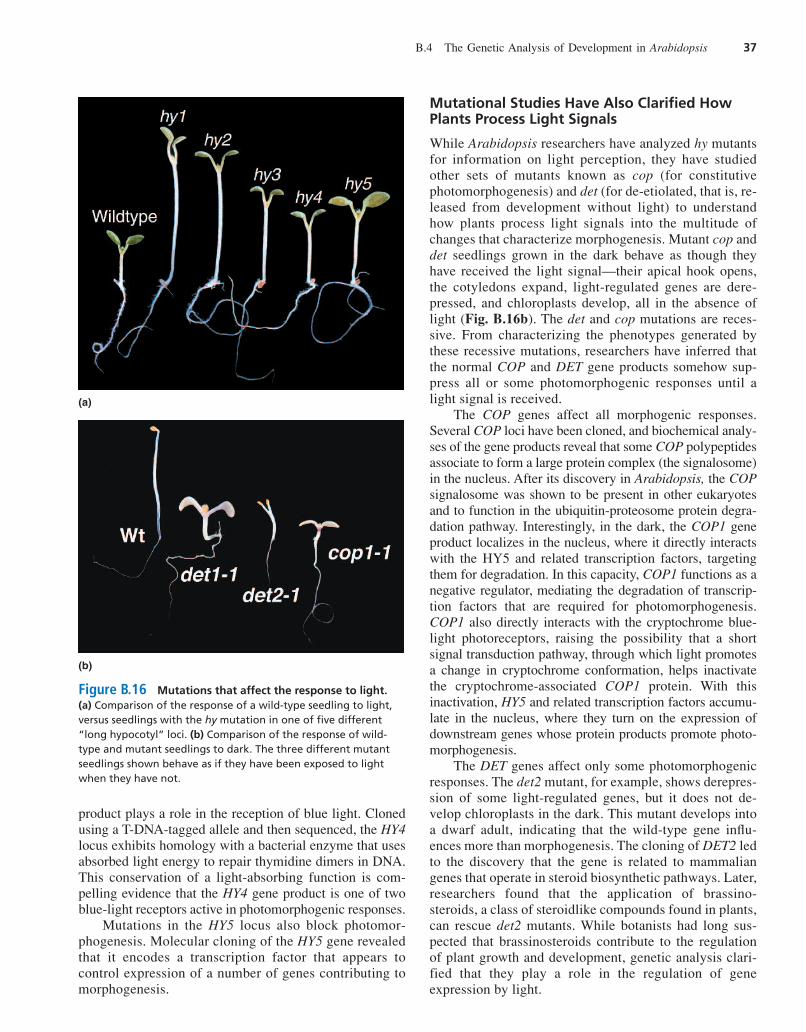
White light inhibits the elongation of the hypocotyl, orseedling stem, in Arabidopsis. A class of photomor-phogenic mutants referred to as hy (for long hypocotyl)show reduced sensitivity to white light (Fig. B.16a). Link-age map locations for two of the hy mutations, hy3 and hy8,coincide with the locations of phyA and phyB members ofthe phytochrome gene family. The hy3 mutant has an al-tered response to red wavelengths of light, while the hy8mutant has an altered response to far-red wavelengths oflight, indicating that these two genes have different roles inmediating photomorphogenesis. Two other mutations, hy1and hy2, block the biosynthesis of the tetrapyrolle pros-thetic group associated with all phytochromes.
The hy4 mutant in Arabidopsis shows a decreased sen-sitivity to blue light, suggesting that the normal HY4 gene
har06584_refB_021-048 11/06/2006 09:26 PM Page 36
product plays a role in the reception of blue light. Clonedusing a T-DNA-tagged allele and then sequenced, the HY4locus exhibits homology with a bacterial enzyme that usesabsorbed light energy to repair thymidine dimers in DNA.This conservation of a light-absorbing function is com-pelling evidence that the HY4 gene product is one of twoblue-light receptors active in photomorphogenic responses.
Mutations in the HY5 locus also block photomor-phogenesis. Molecular cloning of the HY5 gene revealedthat it encodes a transcription factor that appears tocontrol expression of a number of genes contributing tomorphogenesis.
Mutational Studies Have Also Clarified HowPlants Process Light Signals
While Arabidopsis researchers have analyzed hy mutantsfor information on light perception, they have studiedother sets of mutants known as cop (for constitutivephotomorphogenesis) and det (for de-etiolated, that is, re-leased from development without light) to understandhow plants process light signals into the multitude ofchanges that characterize morphogenesis. Mutant cop anddet seedlings grown in the dark behave as though theyhave received the light signal—their apical hook opens,the cotyledons expand, light-regulated genes are dere-pressed, and chloroplasts develop, all in the absence oflight (Fig. B.16b). The det and cop mutations are reces-sive. From characterizing the phenotypes generated bythese recessive mutations, researchers have inferred thatthe normal COP and DET gene products somehow sup-press all or some photomorphogenic responses until alight signal is received.
The COP genes affect all morphogenic responses.Several COP loci have been cloned, and biochemical analy-ses of the gene products reveal that some COP polypeptidesassociate to form a large protein complex (the signalosome)in the nucleus. After its discovery in Arabidopsis, the COPsignalosome was shown to be present in other eukaryotesand to function in the ubiquitin-proteosome protein degra-dation pathway. Interestingly, in the dark, the COP1 geneproduct localizes in the nucleus, where it directly interactswith the HY5 and related transcription factors, targetingthem for degradation. In this capacity, COP1 functions as anegative regulator, mediating the degradation of transcrip-tion factors that are required for photomorphogenesis.COP1 also directly interacts with the cryptochrome blue-light photoreceptors, raising the possibility that a shortsignal transduction pathway, through which light promotesa change in cryptochrome conformation, helps inactivatethe cryptochrome-associated COP1 protein. With thisinactivation, HY5 and related transcription factors accumu-late in the nucleus, where they turn on the expression ofdownstream genes whose protein products promote photo-morphogenesis.
The DET genes affect only some photomorphogenicresponses. The det2 mutant, for example, shows derepres-sion of some light-regulated genes, but it does not de-velop chloroplasts in the dark. This mutant develops intoa dwarf adult, indicating that the wild-type gene influ-ences more than morphogenesis. The cloning of DET2 ledto the discovery that the gene is related to mammaliangenes that operate in steroid biosynthetic pathways. Later,researchers found that the application of brassino-steroids, a class of steroidlike compounds found in plants,can rescue det2 mutants. While botanists had long sus-pected that brassinosteroids contribute to the regulationof plant growth and development, genetic analysis clari-fied that they play a role in the regulation of geneexpression by light.
B.4 The Genetic Analysis of Development in Arabidopsis 37
(a)
(b)
Figure B.16 Mutations that affect the response to light.(a) Comparison of the response of a wild-type seedling to light,versus seedlings with the hy mutation in one of five different“long hypocotyl” loci. (b) Comparison of the response of wild-type and mutant seedlings to dark. The three different mutantseedlings shown behave as if they have been exposed to lightwhen they have not.
har06584_refB_021-048 11/06/2006 03:00 PM Page 37

B.5 The Genetic Analysis of Flowering:A Comprehensive Example
One of the major life events in the development of an an-giosperm is the switch to reproductive growth that culmi-nates in flowering. During the switch, the reprogrammingof the apical meristem from a leaf-producing to a flower-producing tissue gives rise to an astonishing series ofevents: The meristem no longer generates mature leavesin a specific spiral pattern. Instead, it becomes an inflo-rescence meristem (IM) that produces smaller (cauline)leaves, an elongated stem, and many side shoots. The
meristems of the side shoots develop into floral meristems(FMs) that produce a single flower of specific design. TheIM is indeterminate because it produces FMs indefinitely.By contrast, the FMs are determinate because they nor-mally produce a fixed number of floral organs in a precisepattern of concentric rings, and in the process, they losethe potential for meristematic activity as cells of themeristem become incorporated into the maturing pistil(Fig. B.17).
Arabidopsis accomplishes the reprogramming of themeristem through regulation of a complex network ofintricately overlapping genetic pathways in response toenvironmental cues. Some of these determine the bodyplan of the flower; others specify the identity of the floralmeristem; still others control the timing of floral meristemformation.
38 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
Stigma
Style
Gynoecium (ovary)
Anther
Petal Petal
Lateral stamen
Developing petal
Filament and anther = stamen
Scar of removed sepal
Anther of another developing stamen (there are six)
Remaining sepal (other three have been removed)
Sepal primordium (others have been removed
to reveal inner organs)
Developingstamen
Petalprimordium
(a)
Developinggynoecium(ovary)
The single remaining sepal primordium
One of the six stamen primordium
Ovary primordium
Petal primordium
Sepal primordium (1 of 4)
Late-stage flower primordiumshowing early steps in sepal formation
Shoot apical meristem
Early-stage flower primordium
(b)
(c)
(d)
Figure B.17 Differentiation in whorls of floral organs. This series of SEMs shows the earliest stage of development of theA. thaliana flower. (a) Sepals are just beginning to form. Three of the four developing sepals have been removed to (b) reveal the otherorgan primordia. Petal and stamen formation are noticeable in (c). The structure of the stamen and anthers become more defined in(d). Again, three outer sepals have been removed to show the developing flower parts; mature flower carrying four sepals, six stamen,and one gynoecium.
har06584_refB_021-048 11/06/2006 03:00 PM Page 38

How Genes Determine the Body Plan of a FlowerThe floral primordium is an outgrowth of undifferenti-ated cells near the tip of an inflorescence meristem. Ascells in the primordium divide, differentiate, and elon-gate according to genetic instructions, a flower emerges.A wild-type Arabidopsis flower, we have seen, carriesfour green sepals in the outside whorl, four white petalsjust inside the sepals, six stamens in the next-to-centerwhorl, and a cylinder of two fused carpels in the center(review Fig. B.5). Cascades of genetic signals alert cellsto their position within the developing flower and therebydirect the proper growth and differentiation of the organsthat make up a flower. Mutants can have an abnormalarrangement of organs.
Analyses of Homeotic Mutations Reveal That in Arabidopsis, Three Types of Single-GeneProducts Influence Floral Pattern Formation
A homeotic gene is one that plays a role in determininga tissue’s identity during development. A homeotic
mutation causes cells to misinterpret their position inthe blueprint and become normal organs in inappropriatepositions. Homeotic mutations thus alter the overall bodyplan. In one study, researchers looked at mutants with anabnormal order and selection of floral organs. They ob-tained some of the mutants for their study from T-DNAmutagenized laboratory populations, and they used themutagen ethyl methane sulfonate to generate more.Phenotypic screens of a large number of mutants showedthey fell into three classes (Fig. B.18a). Class A dis-played carpels instead of sepals in the first whorl, andstamens instead of petals in the second whorl, for anoverall radial pattern of carpels, stamens, stamens,carpels. In controlled crosses, the ratio of normal to ab-normal phenotypes indicated that one gene was responsi-ble for this abnormal radial pattern. Researchers namedthe gene carrying the mutations that caused this aberrantpattern APETALA2 (AP2) for the lack of petals arising asa result of the mutation. Class B mutants carried sepals inthe first and second whorls, and carpels in the third andfourth whorls. Mutations in either APETALA3 (AP3) orPISTILLATA (PI) caused this deviation from the normalpattern (in botanical terminology, pistillate flowers lackstamen). Class C mutants had an abnormal radial pattern ofsepals, petals, petals, sepals. Mutations in the AGAMOUS(AG) gene, named for the lack of gamete-forming tissuesresulting from the mutations, determined these mutantphenotypes. Loss-of-function ag mutations also lead tothe production of additional whorls of structures at thecenter of the flower, indicating that the wild-typeAGAMOUS gene is required for the determinate nature offloral meristems.
The Radial Pattern Model Suggests How Three Classes of Genes Could Determinethe Identity of Floral Organs
For the sake of simplicity, consider that class A mutants aremissing class A gene activity, B mutants are missing B geneactivity, and C mutants are missing C gene activity. The ra-dial pattern model makes three sets of assumptions aboutthese three classes of genes (Fig. B.18b):
1. A genes are active early in the emergence of the first andsecond whorls; B genes are active early in the defini-tion of the second and third whorls; and the C gene isactive early in the differentiation of the third and fourthwhorls.
2. The gene products of these three types of genes, aloneand in combination, determine the identity of the floralorgans as they develop from the flower primordium.The A product determines sepals, A and B together de-termine petals, the combination of B plus C determinesstamens, and the C product alone determines carpels.
3. The activities of A and C are mutually exclusive; that is,in areas where A is active, C is repressed, and in areaswhere C is active, A is repressed. Moreover, if a muta-tion incapacitates A, C is abnormally active and viceversa.
The model provides an elegant explanation of the mu-tational evidence. To confirm its predictions, researcherscarried out further genetic investigations. In one line of ex-periments, they bred double and triple mutants and pre-dicted what they would look like. For example, they bredplants with mutations in both AP2 (the A gene) and AP3 (aB gene) and predicted that with only normal C activity inthe flower primordium, the emerging flower would consistof all carpels. The resulting flowers confirmed the predic-tion. Similarly, they eliminated both B and C activity bybreeding plants with mutations in the AP3 and AG genes,and as predicted, the resulting mutants bore flowers con-sisting of only sepals (Fig. B.18c). In triple mutants lackingA, B, and C gene activity, the flower organs differentiatedas leaves (Fig. B.18d). Parallel studies in snapdragons sup-port the ABC radial pattern model of angiosperm flowerdevelopment.
The cloning and sequencing of the homeotic genes forfloral identity have further validated the radial patternmodel. All of the genes produce DNA-binding transcrip-tion factors. In situ hybridizations using DNA fragmentscomplementary to A-, B-, and C-type mRNAs have moni-tored the activity of the normal genes and shown that the Band C genes (AP3, PI, and AG) are expressed specificallyin the whorls predicted by the model (Fig. B.18e). The Agene, AP2, however, is expressed in all four whorls of theflower although its product affects organ identity only inthe first and second whorls. In ap2 mutants, AG RNA ap-pears in all four whorls of the flower, as predicted by themodel. In these ap2 mutants, however, only the first and
B.5 The Genetic Analysis of Flowering: A Comprehensive Example 39
har06584_refB_021-048 11/06/2006 03:00 PM Page 39
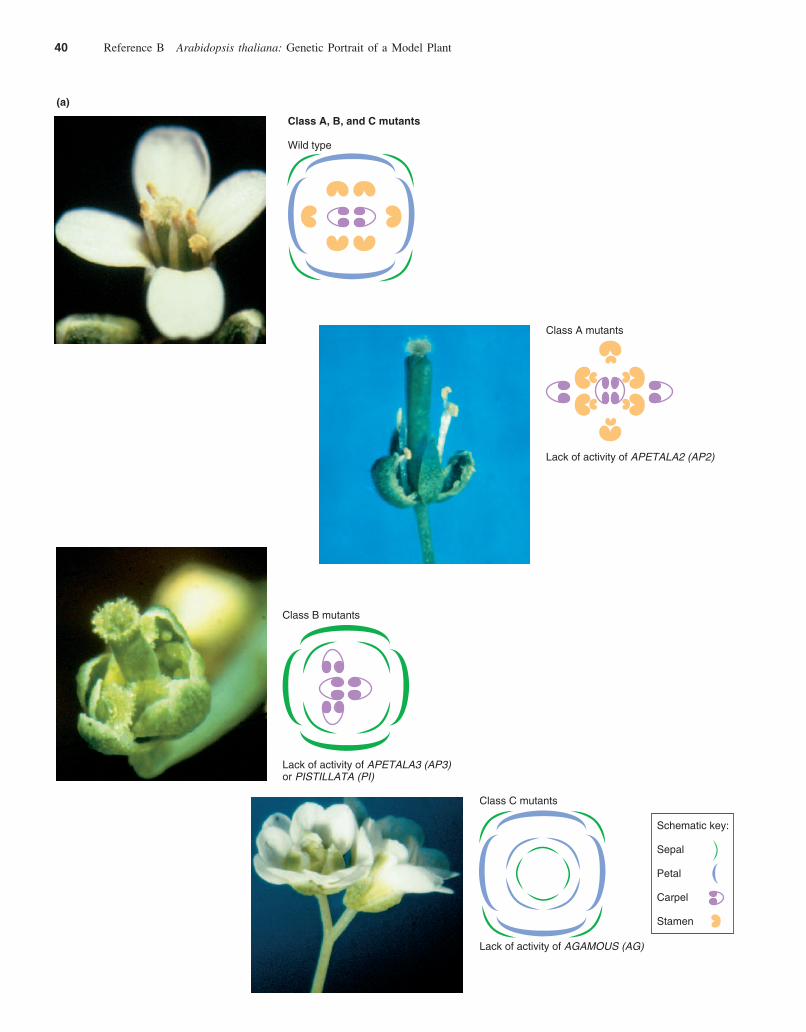
40 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
Class C mutants
Lack of activity of AGAMOUS (AG)
Schematic key:
Sepal
Petal
Carpel
Stamen
(a)
Class B mutants
Lack of activity of APETALA3 (AP3)or PISTILLATA (PI)
Class A mutants
Lack of activity of APETALA2 (AP2)
Class A, B, and C mutants
Wild type
har06584_refB_021-048 11/06/2006 03:00 PM Page 40

B.5 The Genetic Analysis of Flowering: A Comprehensive Example 41
(d) (e)
Figure B.18 Homeotic mutants and flower formation in Arabidopsis. (a) Class A, B, and C mutants. (b) Diagram showing howthe activities of gene types A, B, and C can produce the four types of floral organs: sepals (se), petals (pe), stamens (st), and carpels (ca).The spatial and temporal expression of each gene type is represented by rings containing the three primary colors at the bottom of thediagram. Following this color-coded representation forward, A (blue) expression alone gives rise to sepals, while C (red) expressionalone gives rise to carpels. A and B expression gives rise to petals (blue � yellow � green), while C and B expression gives rise tostamens (red � yellow � orange). (c) ap2 / ap3, ap2 / ag, and ap3 / ag double mutants. (d) ap2 / ap3 / ag triple mutant. (e) Hybridizationprobe visualizing AP3 activity in the flower primordium.
second whorls have an altered phenotype. The explanationis as follows: AP1 (one of the A-function genes) is oneverywhere in the early flower primordium as is AP2. AGturns on in whorls 3 and 4, repressing AP1 and thus ex-cluding A function from whorls 3 and 4. In ap2 mutants,AG comes on everywhere and represses all AP1 activity, sothere is C function in all whorls, but no A function. Thus inap2 mutants, you expect to see only whorl 1 and whorl2 phenotypes.
The data discussed here indicate that the A, B, andC genes are necessary for radial patterning in flowers. How-ever, it appears that they are not sufficient to determineflower organ identity. For instance, the expression ofC genes in leaves does not convert them into carpels.Subsequent genetic analysis of flower patterning inArabidopsis has demonstrated that three functionallyredundant genes (SEPALLATA1, 2, and 3, abbreviated sep1,2, and 3) are essential for the activity of the B- and C-type
genes. Indeed, triple sep1 sep2 sep3 mutants form flowersthat carry an indeterminate number of whorls. Interestingly,single sep1, sep2, or sep3 mutants, or double mutants carry-ing different combinations of these genes, have wild-typeflowers, indicating that the three genes are functionallyredundant. Molecular analyses of these genes have providedan explanation for these striking observations. Such analy-ses show that sep1, 2, and 3 encode putative transcriptionfactors that are highly similar to each other, as well as toAG. These three proteins can bind directly to type B andC proteins, forming heteropolymers that are likely toregulate the expression of different sets of target genes.
In short, the A, B, C model explains one part of a com-plicated genetic process—the interpretation of cellular posi-tion within the flower primordium—but leaves unansweredthe question of what establishes the positions. As we seenext, the differentiation of floral organs, in addition to the ap-propriate expression of the A, B, C, and sep genes, requires
A
B
se sepe pest stca
B
C C A
(b) (c)
har06584_refB_021-048 11/06/2006 09:26 PM Page 41

the activity of many other interacting genes at the propertime and place in an immense hierarchy of genetic control.
Earlier-Acting Genes Specify the Identity of the Floral MeristemThe floral meristem produces the flower primordium thatdifferentiates into four whorls of organs, two of which con-tain the gametes. But what tells cells in the side shoots ofan inflorescence meristem to become a floral meristem? Toanswer this question, researchers analyzed loss-of-functionsingle and double mutants (Fig. B.19a and b) and over-expression transgenic plants. They constructed the transgenicplants by first fusing a strong constitutive promoter called35S with the DNA of the cloned gene under study and theninserting the promoter/gene fusion product into otherwise
wild-type Arabidopsis plants. Table B.1 describes six ofthe floral meristem development genes uncovered by theseloss-of-function and overexpression strategies. By compar-ing the effects of each gene’s activity with that of the oth-ers, the researchers were able to suggest how they interact.
Model of the Progressive Specification of the Floral Fate
The first sign of floral development at the molecular level isexpression of the LEAFY (LFY) gene. Activation of LFYmarks the position of a future floral meristem and takesplace before a visible FM appears. With inadequate LFYfunction, the meristem remains an inflorescence meristem.Next comes the expression APETALA1 (AP1) and CAULI-FLOWER (CAL), whose activities overlap in newly formedmeristems. The orderly expression of LFY, AP1, and CAL
42 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
(a)
(b)
(c)
Figure B.19 Identifying genes that influence development of the floral meristem. (a) Single mutants Ify-6 (left), ap1–1 (middle),and tfl-2 (right). (b) Double mutant ap1–1 / cal-1. (c) Hybridization probes visualize the activity of AG RNA in the center of the flowerprimordium.
har06584_refB_021-048 11/06/2006 03:00 PM Page 42

promotes emergence of an unbranched flowering structurewith four whorls of organs. Loss-of-function mutations inany of these genes prevents the shoot from flowering. In-terestingly, mutations in the AP2 gene enhance the effectsof lfy and ap1 mutations.
Expression of AG occurs at the center of the floralmeristem where its product represses AP1 expression andfurther commits the meristem to the determinate productionof flowers (Fig. B.19c). A lack of AG activity can cause thedeveloping FM to revert to an IM. With the expression ofLFY, AP1, and AG, cells in the lateral, floral meristems be-come increasingly committed to reproductive development.
Wild-type Arabidopsis plants produce only lateralflowers because the TFL1 gene prevents the expression ofFM-producing genes in the center of the IM. As a result theindeterminate inflorescence meristem continues to growand produce new side shoots even as older lateral shootsare activating the genetic program for flowering.
Some Genes Control the Timing of FM Formation and FloweringLaboratory-grown, wild-type Arabidopsis plants adjust theirflowering time in response to the amount of light they receive.Those that receive 16 hours of light a day begin to flowerthree weeks after germination, while those that receive only8–10 hours of light do not initiate flowering until at least sixweeks after germination. Moreover, the shorter days increasethe duration of all phases of development, causing the pro-duction of more leaves in the rosette and more flowers on theinflorescence. Exposure of hydrated seeds (which have takenup water and are ready to germinate) or young plants to lowtemperatures for three to six weeks after germination acceler-ates the flowering of wild-type plants.
As Table B.2 shows, two types of mutations influenceflowering time: those that delay it, and those that precipi-
tate it. Researchers identified many of the mutations thatdisrupt normal flowering by screening laboratory popula-tions for their flowering responses. They also found thatsome mutations identified on the basis of other phenotypesaffect flowering. For example, phyB mutants, which havean altered response to far-red wavelengths of light, andcop1 mutants, which grow in the dark as if they havereceived a light signal, both flower early.
To understand the function of genes that affect floweringtime, researchers have analyzed single mutants, double mu-tants, and transgenic plants containing extra copies of a par-ticular gene. Among their findings is the observation thatcop1, phyB double mutants show the Cop phenotype of shorthypocotyls (rather than the phyB phenotype of expandedhypocotyls). This suggests that COP1 acts downstream ofPHYB. Since both mutations accelerate flowering, the nor-mal gene products may be part of a pathway required torepress flowering, and several overlapping pathways mostlikely lead to flowering. Another finding is that environmen-tal influences on flowering time affect the interactionsamong floral meristem identity genes. The lfy mutants, forexample, show a stronger phenotype under short days thanunder long days; and there is more COP1 RNA in youngseedlings grown under long days than under short days, sug-gesting that the COP1 gene helps promote flowering.
Hundreds of Mutational Analyses HaveGenerated a Preliminary Model to Guide Future Research on Flowering
Since there are probably hundreds of genes that contributeto the control of floral development, the genetic analysis offlowering is a work in progress. Nevertheless, certainobservations enable us to build a preliminary genetic modelof the process. It appears that at least four pathways pro-mote flowering in Arabidopsis: a pathway regulated by daylength (photoperiod-dependent pathway), an autonomous
B.5 The Genetic Analysis of Flowering: A Comprehensive Example 43
TABLE B.1 Six of the Arabidopsis Genes That Control Meristem Identity
Loss of Function (Mutant Phenotypes Gain of Function (Phenotypes Arabidopsis Gene in Meristem Identity) of Transgenic Plants)
LEAFY (LFY ) Conversion of flower to shoot or shoot-like Conversion of lateral shoots to flower; structures; floral reversion terminal flower; early flowering
APETALA1 (AP1) Conversion of flower to shoot; branched flowers Conversion of lateral shoots to flower; terminal flower; early flowering
CAULIFLOWER (CAL) Enhances ap1 phenotypes Not tested
APETALA2 (AP2) Enhances ap1 and Ify phenotypes Not tested
AGAMOUS (AG) Indeterminate flowers; axillary buds; floral Conversion of lateral shoots to flower; reversion terminal flower; early flowering; suppression
of late flowering mutants
TERMINAL FLOWER (TFL1) Terminal flower; early flowering Not tested
Reference: TIG January 1998 Vol. 14 No. 1, “To Be, Or Not To Be, A Flower—Control of Floral Meristem Identity” by H. Ma.
har06584_refB_021-048 11/06/2006 03:00 PM Page 43
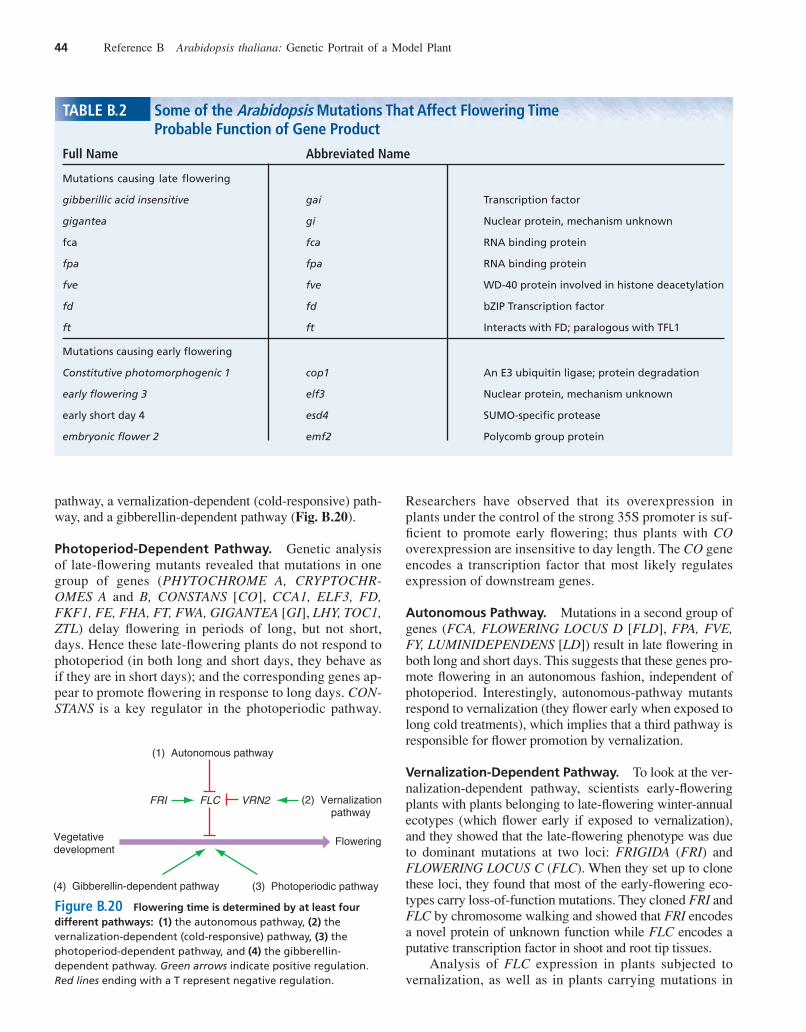
pathway, a vernalization-dependent (cold-responsive) path-way, and a gibberellin-dependent pathway (Fig. B.20).
Photoperiod-Dependent Pathway. Genetic analysisof late-flowering mutants revealed that mutations in onegroup of genes (PHYTOCHROME A, CRYPTOCHR-OMES A and B, CONSTANS [CO], CCA1, ELF3, FD,FKF1, FE, FHA, FT, FWA, GIGANTEA [GI], LHY, TOC1,ZTL) delay flowering in periods of long, but not short,days. Hence these late-flowering plants do not respond tophotoperiod (in both long and short days, they behave asif they are in short days); and the corresponding genes ap-pear to promote flowering in response to long days. CON-STANS is a key regulator in the photoperiodic pathway.
Researchers have observed that its overexpression inplants under the control of the strong 35S promoter is suf-ficient to promote early flowering; thus plants with COoverexpression are insensitive to day length. The CO geneencodes a transcription factor that most likely regulatesexpression of downstream genes.
Autonomous Pathway. Mutations in a second group ofgenes (FCA, FLOWERING LOCUS D [FLD], FPA, FVE,FY, LUMINIDEPENDENS [LD]) result in late flowering inboth long and short days. This suggests that these genes pro-mote flowering in an autonomous fashion, independent ofphotoperiod. Interestingly, autonomous-pathway mutantsrespond to vernalization (they flower early when exposed tolong cold treatments), which implies that a third pathway isresponsible for flower promotion by vernalization.
Vernalization-Dependent Pathway. To look at the ver-nalization-dependent pathway, scientists early-floweringplants with plants belonging to late-flowering winter-annualecotypes (which flower early if exposed to vernalization),and they showed that the late-flowering phenotype was dueto dominant mutations at two loci: FRIGIDA (FRI) andFLOWERING LOCUS C (FLC). When they set up to clonethese loci, they found that most of the early-flowering eco-types carry loss-of-function mutations. They cloned FRI andFLC by chromosome walking and showed that FRI encodesa novel protein of unknown function while FLC encodes aputative transcription factor in shoot and root tip tissues.
Analysis of FLC expression in plants subjected tovernalization, as well as in plants carrying mutations in
44 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
TABLE B.2 Some of the Arabidopsis Mutations That Affect Flowering Time Probable Function of Gene Product
Full Name Abbreviated Name
Mutations causing late flowering
gibberillic acid insensitive gai Transcription factor
gigantea gi Nuclear protein, mechanism unknown
fca fca RNA binding protein
fpa fpa RNA binding protein
fve fve WD-40 protein involved in histone deacetylation
fd fd bZIP Transcription factor
ft ft Interacts with FD; paralogous with TFL1
Mutations causing early flowering
Constitutive photomorphogenic 1 cop1 An E3 ubiquitin ligase; protein degradation
early flowering 3 elf3 Nuclear protein, mechanism unknown
early short day 4 esd4 SUMO-specific protease
embryonic flower 2 emf2 Polycomb group protein
(1) Autonomous pathway
FRI FLC VRN2 (2) Vernalization pathway
Flowering
(3) Photoperiodic pathway(4) Gibberellin-dependent pathway
Vegetativedevelopment
Figure B.20 Flowering time is determined by at least fourdifferent pathways: (1) the autonomous pathway, (2) thevernalization-dependent (cold-responsive) pathway, (3) thephotoperiod-dependent pathway, and (4) the gibberellin-dependent pathway. Green arrows indicate positive regulation.Red lines ending with a T represent negative regulation.
har06584_refB_021-048 11/06/2006 03:00 PM Page 44

FRI or in some of the genes belonging to the autonomouspathway, increased researchers’ understanding of themode of action of vernalization pathway genes. FLC ex-pression appears to decrease to undetectable levels in frimutants and autonomous-pathway mutants, suggestingthat FRI helps promote FLC expression. Furthermore,vernalization suppresses expression of FLC, and this FLCsuppression is maintained over the entire life cycle of theplant. It is not, however, transmitted to the next genera-tion. Thus, FLC expression correlates with an early-flowering phenotype, suggesting that FLC is a keynegative regulator of the transition between the vegetativeand reproductive phases in Arabidopsis (Fig. B.20). Totest this hypothesis, researchers created plants that over-express FLC under the control of the strong 35S pro-moter. These plants flowered late and could not beinduced to flower early by mutations in FRI and in the au-tonomous pathway, nor by vernalization. Conversely,loss-of-function mutations at FLC resulted in an early-flowering phenotype, as discussed previously.
It is interesting that, upon vernalization, FLC is si-lenced not only during cold treatment but also during theentire life cycle of the plant. This means that the plantmaintains a memory of the cold treatment, but, as we saw,that memory is not transmitted to the next generation. Tounderstand this long-term memorization of vernalization,scientists isolated mutations that eliminate a plant’s re-sponsiveness to vernalization. One such mutation, namedvrn2, gave rise to a plant that still responded to vernaliza-tion by decreasing FLC expression but that did not main-tain FLC suppression after vernalization ended. Instead,the gene became active again, and the plant flowered latedespite having been subjected to vernalization. When theseresearchers cloned the VRN2 gene by chromosome walk-ing, they found that it encodes a regulatory protein with ho-mology to animal proteins of the Polycomb-complex.Polycomb-complex proteins suppress the expression ofmultiple developmental genes by modifying the chromatinstructure at their periphery.
Gibberellin-Dependent Pathway. Gibberellins alsopromote flowering, through a fourth pathway. Mutations
that affect gibberellin biosynthesis or response often resultin later-flowering phenotypes (Fig. B.20).
Genetic analyses of flowering in Arabidopsis have un-covered a number of mechanisms that help control the tran-sition from vegetative to reproductive development inplants. Some of the key regulators identified to date alsoappear to contribute to the control of flowering in otherplants, including crops. For instance, FRI and FLC ho-mologs seem to confer a winter-annual habit (in which coldtreatment gives rise to an early-flowering phenotype) or abiennial habit (in which cold treatment is required for flow-ering) on cultivars of many different types of domesticatedplants. Further research will lead to a better understandingof flowering time in plants. It will also aid in the design ofstrategies for engineering different flowering habits incrops and horticultural species.
To Flesh out the Multiple Pathway Model, It Will Be Necessary to Identify More of the Genes That Influence Flowering
There are several reasons why some of these genes havenot yet been identified: The genome is not yet saturatedwith mutations; some genes have redundant functions; andmutations in regulatory genes controlling earlier stages offloral development do not necessarily cause unique floralphenotypes. To find these hard-to-identify genes that influ-ence flowering, researchers can look for new mutants, tryto isolate suppressors and enhancers of existing mutations,and devise protocols that test for the late function ofearly-acting genes (see Chapter 20 of the main textbook).
In addition to identifying more of the genes active inthe pathways that promote flowering, researchers willcontinue to analyze the mechanisms by which these geneswork. To this end they will seek to demonstrate the acti-vation of transcription by encoded proteins and to dis-cover the target genes of the transcription factors.Because the food consumed by people around the globeconsists largely of grains, fruits, and other flower prod-ucts, an understanding of flowering at the molecular levelmay increase the chances of feeding an ever-expandinghuman population.
Connections 45
Connections
Plants are fundamentally different from animals at the levelof the whole organism. In contrast to animals, plants con-tinue to produce new organ systems throughout their lifecycles, are autotrophic, and do not move about. Despitethese differences, the cellular machinery and genetic mech-anisms of plants and animals are surprisingly similar. Infact, many of the genes involved in basic metabolic func-tions and information processing show a striking degree ofhomology between plants and animals. For example, the
AGAMOUS gene specifies a DNA-binding protein thatmost likely regulates downstream genes. A human relativeof the AGAMOUS gene makes a DNA-binding protein,known as serum response factor, that regulates the c-fosoncogene.
The remaining genetic portraits (C, D, and E) look atgenetic mechanisms that propel development in threemodel animals: the nematode C. elegans, the fruit flyDrosophila, and the house mouse Mus musculus.
har06584_refB_021-048 11/06/2006 03:00 PM Page 45

46 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
1. Arabidopsis makes an excellent model organism forgenetic analysis because of its diploid nature, its ca-pacity for prolific reproduction, and the relativelysmall size of its genome. The genome is carried in fivechromosomes that contain little repetitive DNA and atight arrangement of genes with a few small introns.
2. Arabidopsis switches from vegetative to reproductivedevelopment when the leaf-producing apical meri-stem becomes a flower-producing meristem. Afterflowering, it undergoes double fertilization to producean embryo and an endosperm that develops within aprotective maternal seed coat. In response to properconditions, including favorable light and temperature,the seed germinates and vegetative growth beginsagain. The vegetative plant eventually ages and dies.
3. Researchers use several techniques of genetic analy-sis in Arabidopsis to generate mutations and charac-terize them at the molecular level. Mutagenesis bytreatment with chemicals or X-rays produces a vari-ety of mutations for study. Insertional mutagenesis byA. tumefaciens and transposons produces mutatedalleles carrying a specific DNA insert that can serveas a probe for identifying and cloning the gene of in-terest. Researchers have also assembled large collec-tions of sequence-indexed insertion mutants. Theability to generate and obtain mutations makes it pos-sible to move from the identification of a gene to thephenotype it produces. It is also possible to determinegene function with the use of novel RNA-based si-lencing techniques, which do not depend on therecovery of mutations in the gene of interest.
4. The genetic analysis of mutations that affect embryo-genesis has revealed that in one respect, Arabidopsisis more similar to mammals than to arthropods suchas flies. Like mammals and unlike flies, plants havevery few maternal-effects mutations and few zygoticmutations before the globular stage. Mutationalanalyses have also helped identify some of the regu-latory processes involved in embryogenesis. Futureresearch will try to pin down the molecular mecha-nisms that control embryogenesis.
5. The genetic analysis of hormonal control systems hasclarified the biological significance of hormones andthe biosynthetic pathways that produce them. It hasalso shown how Arabidopsis perceives and trans-duces hormone signals. Some of the genes involvedin hormone production and hormone responses arehomologous to those found in yeast and mammals.
6. The genetic analysis of growth and development inresponse to light conditions, known as photomorpho-genesis, has clarified how plants receive and processlight signals. Some of the genes that contribute tophotomorphogenesis are related to mammalian genesthat operate in steroid biosynthetic pathways.
7. From the genetic analysis of flowering, researchershave proposed that three types of genes determine theidentity of floral organs. These genes operate in themiddle of a regulatory cascade in which earlier-acting genes specify the identity of the floral meri-stem, while other genes control the timing of bothfloral meristem formation and flowering.
Essential Concepts
Solved Problems
I. The St strain of Arabidopsis switches from vegeta-tive to reproductive (flower) development late in theseason compared to the Limburg-5 strain, whichflowers early. How could you determine if floweringtime is based on the alleles of a single or multiplegenes? Describe the results you would expect for thetwo different cases.
Answer
This problem requires an understanding of single- andmultigene inheritance. The two different strains couldbe crossed and the hybrid selfed to produce an F2 gen-eration. If the phenotype is due to a single gene, youwould expect discrete phenotypes in the F2 generation—
either two or three phenotypes, depending on whetherthere was complete or incomplete dominance. If thetrait is determined by several genes, there would be aspectrum of phenotypes in the F2 generation rangingfrom early to late flowering.
II. HY5 is a gene that encodes a positive regulator of geneexpression in response to light in Arabidopsis.a. What do you predict would happen to the expres-
sion of light-regulated genes in a loss-of-functionhy5 mutant?
b. To examine the action of the Hy5 protein at themolecular level, the expression of promoter-GUS reporter gene fusions was monitored. Oneof the fusions contained a light response element
har06584_refB_021-048 11/06/2006 03:00 PM Page 46

Problems 47
(G box) and the other lacked the light responseelement. Results are shown here. (A � indicatesan increase in expression.)
Promoter elements Light-activated GUS expression
HY5� hy5�
No light � Light No light � LightG box � � � �No G box � � � �
What would you conclude about the action of Hy5?c. If you wanted to determine if HY5 regulates
expression in nonphotosynthetic as well as photo-synthetic tissues of the plant, how would you designthe experiment?
Answer
This question requires an understanding of generegulation.a. Loss of function of a positive regulator would re-
sult in no expression of genes in response to light.
b. Both the G box in the promoter and the HY5 geneproduct are needed to get increased gene expres-sion in response to light. (It is reasonable to hy-pothesize that HY5 interacts directly or indirectlywith the G box sequence.)
c. There are several ways in which you could exam-ine the tissue specificity of light-regulated, HY5-dependent transcription. In all cases, the wildtype (HY5) and hy5 mutant should be comparedwith and without the light stimulation. RNA canbe isolated from photosynthetic and nonphoto-synthetic tissues in the two strains, with andwithout light, and hybridized using a probe froma light-regulated, HY5-dependent gene; or DNAfrom a HY5-dependent gene can be hybridized tofixed tissues in the different plants (in situ hy-bridization) or a reporter gene linked to a pro-moter of an HY5-regulated gene could be used tomonitor expression in tissues.
Problems
B-1 For each of the terms in the left column, choose thebest matching phrase in the right column.
a. apical meristem 1. light-regulated developmental program
b. photomorphogenesis 2. variety of plant
c. whorls 3. growing point of a plant containingundifferentiated cells
d. inflorescence 4. concentric portions of a flower
e. strain 5. group of flowers at the tip of a branch
B-2 Why are the following considered to be advantages forthe geneticist or molecular biologist using Arabidopsisas a model plant system compared to corn or peas?a. generation time of six weeksb. 10,000–40,000 seeds per plantc. genome size similar to that of C. elegans
B-3 Arabidopsis has a small genome yet carries out thesame functions as plants such as tobacco and pea,which have genomes 50–100 times larger. How canArabidopsis accomplish similar physiology with solittle DNA?
B-4 Different strains produce different numbers of seeds(seed sets).a. If you wanted to determine whether this trait was
due to a single gene or multiple genes, how wouldyou test this?
b. If the trait was due to multiple genes, what char-acteristic of the different strains would you use tolocate genes involved?
B-5 What is the ploidy of each of the following tissues,what nuclei combined to form them, and how are they
related genetically to the plant on which they arefound?a. endospermb. zygotec. embryo sac cells
B-6 After germinating mutagenized seed, there may besome mutant phenotypes in the first generation. How-ever, these are not usually the plants that a geneticistwill choose to study. Why not?
B-7 Match the type of mutagenic agent with the best use(assume there is only one match for each agent).
a. promoterless 1. generate a collection of plants with GUS-containing vector different loss-of-function mutations
b. EMS 2. study expression of genes
c. kanr Ti 3. generate temperature-sensitive mutants
B-8 An alternative to the use of sequence-indexed mu-tant libraries is an RNA-based silencing protocol fordetermining gene function. Explain how the pheno-type associated with a loss-of-function mutation at aparticular gene can be ascertained even when no mu-tations in that gene are actually available to theresearcher.
B-9 A sterile mutant was identified in which there was arandom instead of directed growth of pollen tubes to-ward the ovary. The mutant was discovered after mu-tagenesis of seeds and self-fertilization of the progenyplants. It was not known therefore whether the sterilitydefect was a male or female defect. How could you de-termine that?
har06584_refB_021-048 11/06/2006 03:00 PM Page 47

48 Reference B Arabidopsis thaliana: Genetic Portrait of a Model Plant
B-10 How could you find out if an APETALA homolog ex-ists in petunias, snapdragons, or potatoes?
B-11 The gravitropic response in plants includes upwardcurve of the hypocotyl and flowering stalk and down-ward growth of the root.a. It has been postulated that this response is medi-
ated by auxin. What is a genetic experiment totest this hypothesis?
b. Auxin-regulated mRNAs have been identified inArabidopsis. How would you determine the dis-tribution of these auxin-regulated mRNAs in thepart of the plant responding to gravity?
B-12 Ethylene affects plant growth and development inmany ways including inhibition of stem and rootelongation, stimulation of seed germination, andripening of fruits. Ethylene biosynthesis occurs in theplant at several different developmental times. Mu-tants that are defective in ethylene stimulation mightbe defective in the biosynthesis of ethylene by theplant or in the signaling pathway in cells that receiveand process the signal.a. The ctr1 mutant has a defect in the signaling path-
way resulting in a constitutive ethylene response.If you treat this mutant with an inhibitor of ethyl-ene biosynthesis, what effect do you expect?
b. Ethylene overproduction mutants eto1, eto2, andeto3 are sensitive to inhibitors of ethylenebiosynthesis. Would you conclude that the ETOgenes are biosynthetic or signaling mutants?
c. No loss-of-function mutations have been iso-lated in the ETR1 gene. This could indicate thatthe gene is required for growth or that there isredundancy of this gene. How could you look forredundancy of this gene in the genome?
d. A series of bases in the promoter region of sev-eral ethylene responsive genes have been identi-fied that are essential for proper DNA binding tospecific transcription factors in the ethylene-responsive element-binding protein (EREBP)family. The APETALA2 gene product has apolypeptide domain that is homologous to apolypeptide domain found in the EREBPs. Whywould these proteins show evidence of homol-ogy in one of their domains?
e. Genetic engineering of ETR1 and/or CTR1 genesmay prevent or modify the ethylene responseand could be very helpful in regulating the ripen-ing of fruits and vegetables. If you need to con-trol expression of the genes, what would youwant to be part of the DNA construct that you in-jected into the plant?
har06584_refB_021-048 11/06/2006 03:00 PM Page 48





























