Embed Size (px)
Citation preview

Marine Pollution Bulletin 60 (2010) 1691–1700
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Assessing impacts of invasive phytoplankton: The Baltic Sea case
Irina Olenina a,b,*, Norbert Wasmund c, Susanna Hajdu d, Iveta Jurgensone e, Sławomira Gromisz f,Janina Kownacka f, Kaire Toming g, Diana Vaici�ute a, Sergej Olenin a,b
a Coastal Research and Planning Institute, Klaipeda University, Manto str. 84, Klaipeda, Lithuaniab Uni Research AS, Thormøhlensgt. 49, Bergen, Norwayc Leibniz-Institute for Baltic Sea Research, Seestr. 15, Warnemünde, Germanyd Department of Systems Ecology, Stockholm University, SE-10691 Stockholm, Swedene Latvian Institute of Aquatic Ecology, Marine Monitoring Center, Daugavgrivas str. 8, Riga, Latviaf Sea Fisheries Institute, Kołłataja 1, PL 81-332 Gdynia, Polandg Estonian Marine Institute, University of Tartu, Mäealuse str. 10a, Tallinn, Estonia
a r t i c l e i n f o a b s t r a c t
Keywords:Non-indigenous speciesCryptogenicEnvironmental impact assessmentBiological pollutionPlankton microalgae communityProrocentrum minimum
0025-326X/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.marpolbul.2010.06.046
* Corresponding author at: Coastal Research andUniversity, H. Manto str. 84, Klaipeda 92294, Lithuan+370 46 398845.
E-mail addresses: [email protected], irina.onina).
There is an increasing understanding and requirement to take into account the effects of invasive alienspecies (IAS) in environmental quality assessments. While IAS are listed amongst the most important fac-tors threatening marine biodiversity, information on their impacts remains unquantified, especially forphytoplankton species. This study attempts to assess the impacts of invasive alien phytoplankton inthe Baltic Sea during 1980–2008. A bioinvasion impact assessment method (BPL – biopollution levelindex) was applied to phytoplankton monitoring data collected from eleven sub-regions of the BalticSea. BPL takes into account abundance and distribution range of an alien species and the magnitude ofthe impact on native communities, habitats and ecosystem functioning. Of the 12 alien/cryptogenic phy-toplankton species recorded in the Baltic Sea only one (the dinoflagellate Prorocentrum minimum) wascategorized as an IAS, causing a recognizable environmental effect.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
There is an increasing need to take into account effects of inva-sive alien species (IAS), including phytoplankton, in overall envi-ronmental quality assessments (Cardoso and Free, 2008; Orendtet al., 2009). IAS are a subset of the alien (syn.: non-indigenous,non-native, exotic) species which were introduced by humans out-side of their natural range and dispersal potential. IAS can repro-duce in large numbers and rapidly extend their ranges and maycause significant impacts on native biodiversity, economic activi-ties and/or human health (for bioinvasion terminology see e.g.,Occhipinti-Ambrogi and Galil, 2004; Pyšek et al., 2009; Carlton,2009). The impacts of alien species may be interpreted as causinga decline in ecological quality due to changes in biological, chem-ical and physical properties of aquatic ecosystems (Elliott, 2003).However, there are very few methods that have been developedto assess environmental quality affected by invasive species (e.g.Olenin et al., 2007; Orendt et al., 2009.
ll rights reserved.
Planning Institute, Klaipedaia. Tel.: +370 46 398847; fax:
[email protected] (I. Ole-
In the European Alien Species Database (DAISIE, 2009; Oleninand Didziulis, 2009) some 51 phytoplankton species are indicatedas being alien to European coastal waters: 22 dinoflagellates, 20diatoms, 7 raphidophytes, 1 prymnesiophyte and 1 phaeophytespecies. This number is unlikely to be complete, because unicellu-lar plankton organisms were not reported for all European marineregions. In addition recent taxonomy revisions can result in newchanges to alien species inventories. Systematic studies of marinephytoplankton began at least a 100 years later than other investi-gations of larger biota. Still even in well studied areas (e.g. in theBaltic Sea) where there is regular monitoring every year yieldsnew additions to phytoplankton inventories (Hällfors, 2004; HEL-COM, 2009a). Phytoplankton can vary in their morphology accord-ing to their exposure to different environments; sometimes thesame species giving rise to a complex synonymy from different re-gional seas. A feature of some phytoplankton is a spontaneousappearance and this may happen without the species being re-corded before and so giving rise to the possibility that the speciesis alien. The status of several species in northern Europe has beenreviewed by Gómez (2008). However, the classification of alien,cryptogenic (species of unknown origin which can not be ascribedas being native or alien are termed cryptogenic, sensu Carlton,1996) and cosmopolitan species and the discovery of new species

1692 I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700
to science make it difficult to apportion the status of a species andthis may vary according to personal opinion.
On another hand, phytoplankton are commonly distributedwithin ships’ ballast water and many phytoplankton species havebeen spread in this way (e.g. Subba Rao et al., 1994; Gollaschet al., 1998; Olenin et al., 2000; Pertola et al., 2006). Some species,recorded from ships’ ballast water, are increasing in abundance,spreading over large areas with a consequent impact on biologicaldiversity, ecosystem functioning and socio-economic values, (e.g.Occhipinti-Ambrogi and Galil, 2004; Pyšek et al., 2009).
The list of phytoplankton recorded in the Baltic Sea consists ofmore than 2000 species (Hällfors, 2004); and is regularly updatedby the HELCOM Phytoplankton Expert Group (HELCOM, 2009a).The inventory does not indicate whether these are alien or nativeor their likely. This information, where known, can be found inthe literature presented in two regional online information sys-tems: NOBANIS (2009) and the Baltic Sea Alien Species Database(2009); additional alien species are indicated by Pertola (2006).
This paper examines the overall level of bioinvasion impactscaused by invasive alien phytoplankton in a relatively well studiedlarge marine region, the Baltic Sea. We used long-term interna-tional (Baltic Marine Program, HELCOM, 2009b) and national (Esto-nian, Latvian, Lithuanian, Polish, German and Swedish)phytoplankton monitoring data to measure the abundance and dis-tribution range of invasive alien phytoplankton species. We relatedthe abundance and distribution range to the impacts of IAS on na-tive communities, habitat and ecosystem functioning according tothe biopollution assessment method of Olenin et al. (2007). Wealso examined how routine monitoring may be used for bioinva-sion impact assessment on a scale of various sub-regions of theSea.
Fig. 1. Sub-regions of the Baltic Sea considered in this paper: I – Belt Sea area, incl. MecPolish coastal waters; V – Eastern Gotland Sea; VI – Northern Baltic, Swedish coastal wNorthern Baltic, Landsort Deep; IX – South-Eastern Baltic Sea, Lithuanian coastal wateEstonian coastal waters.
2. Materials and methods
2.1. Phytoplankton monitoring data
The research is based on joint 1980–2008 HELCOM and nationalphytoplankton monitoring program for eleven sub-regions (assess-ment units) of the Baltic Sea (Fig. 1; Table 1). Samples were ana-lyzed and species identified by phytoplankton experts, using themandatory Baltic Sea international monitoring methods (HELCOM,2009c). Data on abundance (cells L�1), relative biomass (% from to-tal phytoplankton biomass) and distribution of species at monitor-ing stations within a sub-region was used.
The following yearly periods were used in the analysis (1980–1984, 1985–1989, 1990–1994, 1995–1999, 2000–2004, 2005–2008) in each sub-region. Such periodicity usually is being usedto report the environmental status in the HELCOM area (e.g. HEL-COM, 1996, 2001). Each assessment period in a particular sub-re-gion was considered as a case study; in total there were 66 suchcase studies (i.e. 11 assessment units � 6 time periods), howeveronly 51 case studies were used for further analysis as in the restno presence of invasive phytoplankton was reported.
2.2. Bioinvasion impact assessment
In order to classify the impacts of invasive alien species on na-tive species, communities, habitats and ecosystem functioning thebiopollution index (BPL) (Olenin et al., 2007) was used. Each sub-region (Fig. 1) was considered as an assessment unit. The BPL cal-culation is based on an abundance and distribution range (ADR),for each species the magnitude of bioinvasion impact was assessed.
klenburg and Kiel Bights; II – Arkona Sea; III – Bornholm Sea; IV – Southern Baltic,aters, Askö; VII – Northern Baltic, Swedish coastal waters, Himmerfjärden; VIII –
rs; X – South-Eastern Baltic, Latvian coastal waters; XI – North-Eastern Baltic Sea,

Table 1Number of sampling sites and frequency in different sub-regions (assessment units)of the Baltic Sea.
Sub-regions Phytoplanktonmonitoringsince (year)
Number ofsampling sitesper sub-region
Samplingfrequency(samples peryear)*
Belt Sea area 1979 5 7–12Arkona Sea 1979 4 5–10Bornholm Sea 1979 1 5–10Polish coastal waters 1987 116 12Lithuanian coastal
waters1981 25 4–12
Eastern Gotland Sea 1979 2 5–10Swedish coastal
waters, Askö1976 1 22–24
Swedish coastalwaters,Himmerfjärden
1977 5 8–24
Landsort Deep 1979 1 20–22Latvian coastal waters 1992 11 5–21Estonian coastal waters 1993 4 12
* Sampling frequency varies at different stations depending of the aim and type ofmonitoring
Table 2Five classes representing the abundance and distribution range of alienspecies (based on Olenin et al., 2007).
Code Description
A Low numbers in one or several localitiesB Low numbers in many localities
moderate numbers in one or several localitiesC Low numbers in all localities
moderate numbers in many localitieshigh numbers in several localities
D Moderate numbers in all localitieshigh numbers in many localities
E High numbers in all localities
I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700 1693
An abundance is ranked as ‘‘low” where a species makes up onlya small part of the relevant community: e.g. a population of analien alga forms a minor portion (few %) of the total phytoplanktoncommunity. It is ‘‘moderate” when an alien species constitutes lessthan a half of abundance of the native community; and is ‘‘high”when it exceeds half, i.e. quantitatively dominates in the invadedcommunity. The distribution scored as ‘‘one locality” when an alienphytoplankton species was found only at one sampling stationwithin an assessment unit; ‘‘several localities” (the species spreadbeyond one locality but is present in less than half of the samplingstations), ‘‘many localities” (extends to more than a half of the sta-tions) and ‘‘all localities” (found at all stations). Combination of theabundance and distribution scores gives five classes of ADR (A–E),ranking an alien species from low abundance in a few localities (A)to occurrence in high numbers in all localities (E) (Table 2).
After the ADR is estimated, it is related to the magnitude of bio-invasion impacts, ranging from no impact (0) through weak (1),moderate (2), strong (3) and massive (4). Three categories of im-pacts have been considered, namely: (1) impact on native speciesand communities (ranging from C0 to C4); (2) impact on habitats(H0 to H4); (3) impact on ecosystem functioning (E0 to E4) (for de-tails see Olenin et al., 2007).
The bioinvasion impact assessment was performed using an on-line Biopollution Assessment System (BINPAS, 2009) which trans-lates existing data on invasive species impacts into uniformbiopollution measurement units and accumulates data on bioinva-sion impacts. This system provides three confidence levels whileassessing ADR and the impacts: high (data documented by fieldstudies for the given assessment unit), medium (data documentedfor a part of the assessment unit and extrapolated to the entire sys-
tem by expert judgment) and low (expert knowledge of the speciesimpact based on data from studies made elsewhere applied; see –the online website BINPAS, 2009 for details and where all data inthis account is also presented). The overall bioinvasion impact(Biopollution level, BPL) for the assessment unit was determinedaccording to the greatest impact level for the alien species whichwas noticed during the evaluation period (Olenin et al., 2007).
3. Results
3.1. Alien and cryptogenic phytoplankton species in the Baltic Sea
During the entire monitoring period of 1980–2008 most of thealien and cryptogenic phytoplankton species were either rare(occurring in low numbers, negligible in comparison to the nativephytoplankton) or were not found in the monitoring samples (Ta-ble 3). For this reason their impact on native communities, habitatsand the Baltic Sea ecosystem functioning was not considered to besignificant.
A dictyochophyte Pseudochattonella farcimen was reported byŁotocka (2009) only from a survey performed near a pier in Sopot(Gulf of Gdansk) in the spring of 2001, though at high density(11.5 � 106 cells L�1 and biomass 927.5 lgC L�1). This species,however, never was found in monitoring samples.
The only truly invasive species, that has increased in abundanceand has spread over large areas during the study period, was thedinoflagellate Prorocentrum minimum. The assessment of bioinva-sion impact is based on the analysis of this species alone.
3.2. Assessing abundance and distribution range of P. minimum
The dinoflagellate P. minimum was reported the first time innorthern European coastal waters from the English Channel areain 1976 (Smayda, 1990). In 1979 it formed a massive bloom inthe Skagerrak area; in 1981 P. minimum reached the Kattegat andit subsequently entered the Baltic Sea: in 1982 to the Belt Sea area(Edler et al., 1982), in 1983 in Kiel fjord (Kimor et al., 1985) and in1989 in the southern part of the Baltic Sea and Gulf of Gdansk(Mackiewicz, 1995). In 1989–1993, the species was found in thecentral and northern parts of the Baltic Proper, while in 1997–1999 it extended its range to the Gulfs of Finland and of Riga (Haj-du et al., 2000; and monitoring data). Presently this species is notknown from the Gulf of Bothnia. It has become a common elementof summer-autumn phytoplankton community of the Baltic Sea.
The abundance of P. minimum has varied greatly within theassessment units (Table 4). For example, it was first recorded inthe Arkona Sea area 40 cells L�1, making up only 0.004% of the totalphytoplankton. This was based on a single cell in a 25 ml countingchamber. Whereas in Latvian coastal waters it was recorded for thefirst time at 227 � 103 cells L�1 already forming 20% of the phyto-plankton After its establishment it has shown a very high inter-an-nual variability ranging from an absence and when present from�100 cell�L�1 to blooms occurring in most samples.
The greatest abundance for P. minimum within each sub-regionof the Baltic Sea during the assessment periods is shown in the Ta-ble 5. The maximum abundance of 350 million cells L�1 was re-corded in the Gdynia Harbor basins, at the mouth of theChylonka river in the Gulf of Gdansk (Witek and Plinski, 2000; Pol-ish phytoplankton monitoring data). The abundance and distribu-tion of P. minimum was converted into standard ADR classes(Fig. 2). The highest ADR (Class E), other than that found in Polishcoastal waters during the 1995–1999 assessment period, was alsofound in the Belt Sea area (1990–1994) and also in the Lithuaniancoastal waters (in 1995–1999 and 2000–2004). The species waspresent at most sampling stations and was dominant in the phyto-
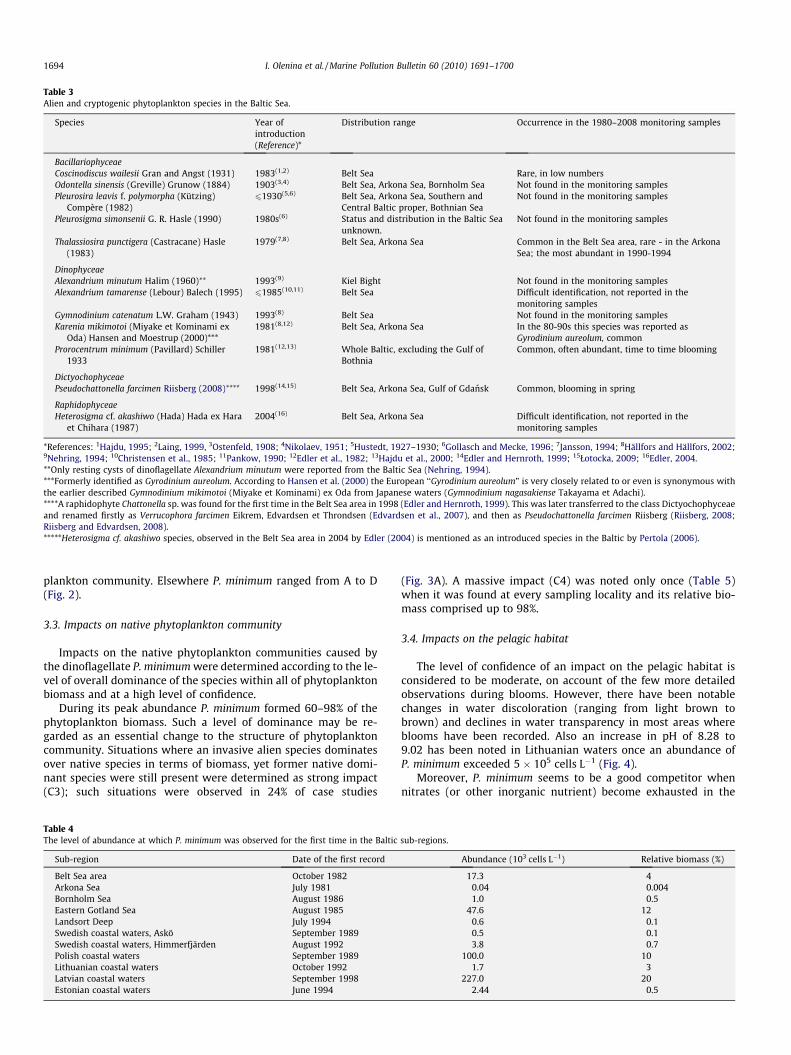
Table 3Alien and cryptogenic phytoplankton species in the Baltic Sea.
Species Year ofintroduction(Reference)*
Distribution range Occurrence in the 1980–2008 monitoring samples
BacillariophyceaeCoscinodiscus wailesii Gran and Angst (1931) 1983(1,2) Belt Sea Rare, in low numbersOdontella sinensis (Greville) Grunow (1884) 1903(3,4) Belt Sea, Arkona Sea, Bornholm Sea Not found in the monitoring samplesPleurosira leavis f. polymorpha (Kützing)
Compère (1982)61930(5,6) Belt Sea, Arkona Sea, Southern and
Central Baltic proper, Bothnian SeaNot found in the monitoring samples
Pleurosigma simonsenii G. R. Hasle (1990) 1980s(6) Status and distribution in the Baltic Seaunknown.
Not found in the monitoring samples
Thalassiosira punctigera (Castracane) Hasle(1983)
1979(7,8) Belt Sea, Arkona Sea Common in the Belt Sea area, rare - in the ArkonaSea; the most abundant in 1990-1994
DinophyceaeAlexandrium minutum Halim (1960)** 1993(9) Kiel Bight Not found in the monitoring samplesAlexandrium tamarense (Lebour) Balech (1995) 61985(10,11) Belt Sea Difficult identification, not reported in the
monitoring samplesGymnodinium catenatum L.W. Graham (1943) 1993(8) Belt Sea Not found in the monitoring samplesKarenia mikimotoi (Miyake et Kominami ex
Oda) Hansen and Moestrup (2000)***1981(8,12) Belt Sea, Arkona Sea In the 80-90s this species was reported as
Gyrodinium aureolum, commonProrocentrum minimum (Pavillard) Schiller
19331981(12,13) Whole Baltic, excluding the Gulf of
BothniaCommon, often abundant, time to time blooming
DictyochophyceaePseudochattonella farcimen Riisberg (2008)**** 1998(14,15) Belt Sea, Arkona Sea, Gulf of Gdansk Common, blooming in spring
RaphidophyceaeHeterosigma cf. akashiwo (Hada) Hada ex Hara
et Chihara (1987)2004(16) Belt Sea, Arkona Sea Difficult identification, not reported in the
monitoring samples
*References: 1Hajdu, 1995; 2Laing, 1999, 3Ostenfeld, 1908; 4Nikolaev, 1951; 5Hustedt, 1927–1930; 6Gollasch and Mecke, 1996; 7Jansson, 1994; 8Hällfors and Hällfors, 2002;9Nehring, 1994; 10Christensen et al., 1985; 11Pankow, 1990; 12Edler et al., 1982; 13Hajdu et al., 2000; 14Edler and Hernroth, 1999; 15Łotocka, 2009; 16Edler, 2004.**Only resting cysts of dinoflagellate Alexandrium minutum were reported from the Baltic Sea (Nehring, 1994).***Formerly identified as Gyrodinium aureolum. According to Hansen et al. (2000) the European ‘‘Gyrodinium aureolum” is very closely related to or even is synonymous withthe earlier described Gymnodinium mikimotoi (Miyake et Kominami) ex Oda from Japanese waters (Gymnodinium nagasakiense Takayama et Adachi).****A raphidophyte Chattonella sp. was found for the first time in the Belt Sea area in 1998 (Edler and Hernroth, 1999). This was later transferred to the class Dictyochophyceaeand renamed firstly as Verrucophora farcimen Eikrem, Edvardsen et Throndsen (Edvardsen et al., 2007), and then as Pseudochattonella farcimen Riisberg (Riisberg, 2008;Riisberg and Edvardsen, 2008).*****Heterosigma cf. akashiwo species, observed in the Belt Sea area in 2004 by Edler (2004) is mentioned as an introduced species in the Baltic by Pertola (2006).
1694 I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700
plankton community. Elsewhere P. minimum ranged from A to D(Fig. 2).
3.3. Impacts on native phytoplankton community
Impacts on the native phytoplankton communities caused bythe dinoflagellate P. minimum were determined according to the le-vel of overall dominance of the species within all of phytoplanktonbiomass and at a high level of confidence.
During its peak abundance P. minimum formed 60–98% of thephytoplankton biomass. Such a level of dominance may be re-garded as an essential change to the structure of phytoplanktoncommunity. Situations where an invasive alien species dominatesover native species in terms of biomass, yet former native domi-nant species were still present were determined as strong impact(C3); such situations were observed in 24% of case studies
Table 4The level of abundance at which P. minimum was observed for the first time in the Baltic
Sub-region Date of the first record
Belt Sea area October 1982Arkona Sea July 1981Bornholm Sea August 1986Eastern Gotland Sea August 1985Landsort Deep July 1994Swedish coastal waters, Askö September 1989Swedish coastal waters, Himmerfjärden August 1992Polish coastal waters September 1989Lithuanian coastal waters October 1992Latvian coastal waters September 1998Estonian coastal waters June 1994
(Fig. 3A). A massive impact (C4) was noted only once (Table 5)when it was found at every sampling locality and its relative bio-mass comprised up to 98%.
3.4. Impacts on the pelagic habitat
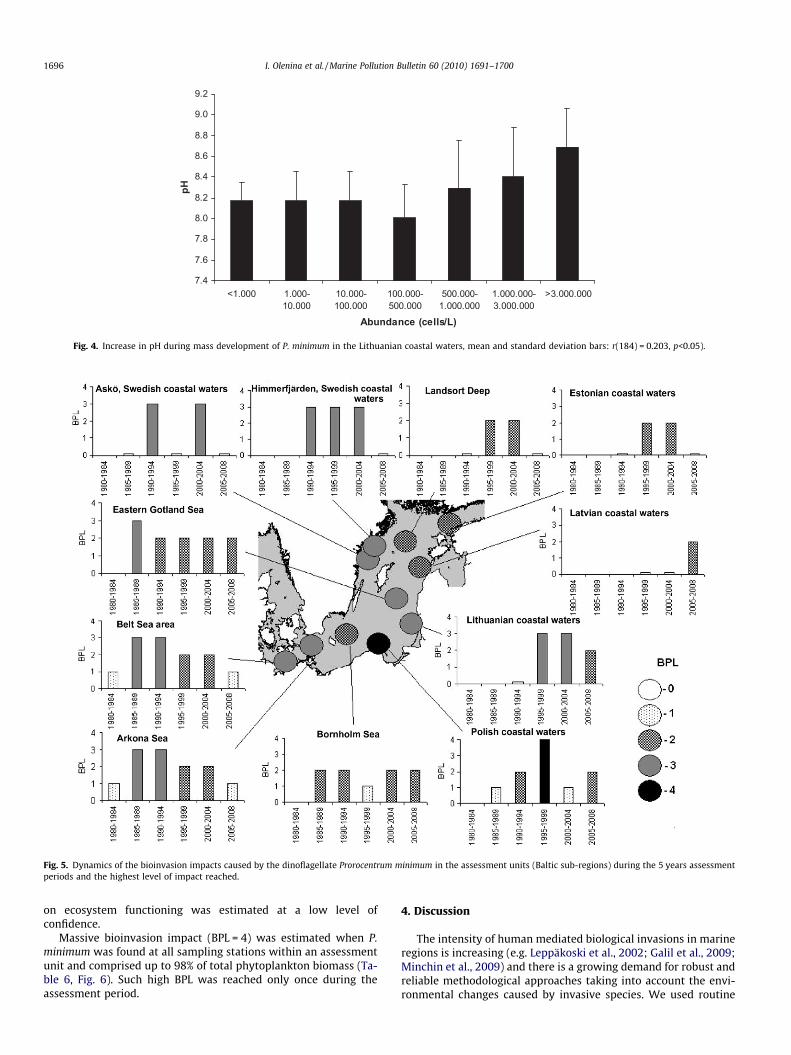
The level of confidence of an impact on the pelagic habitat isconsidered to be moderate, on account of the few more detailedobservations during blooms. However, there have been notablechanges in water discoloration (ranging from light brown tobrown) and declines in water transparency in most areas whereblooms have been recorded. Also an increase in pH of 8.28 to9.02 has been noted in Lithuanian waters once an abundance ofP. minimum exceeded 5 � 105 cells L�1 (Fig. 4).
Moreover, P. minimum seems to be a good competitor whennitrates (or other inorganic nutrient) become exhausted in the
sub-regions.
Abundance (103 cells L�1) Relative biomass (%)
17.3 40.04 0.0041.0 0.5
47.6 120.6 0.10.5 0.13.8 0.7
100.0 101.7 3
227.0 202.44 0.5
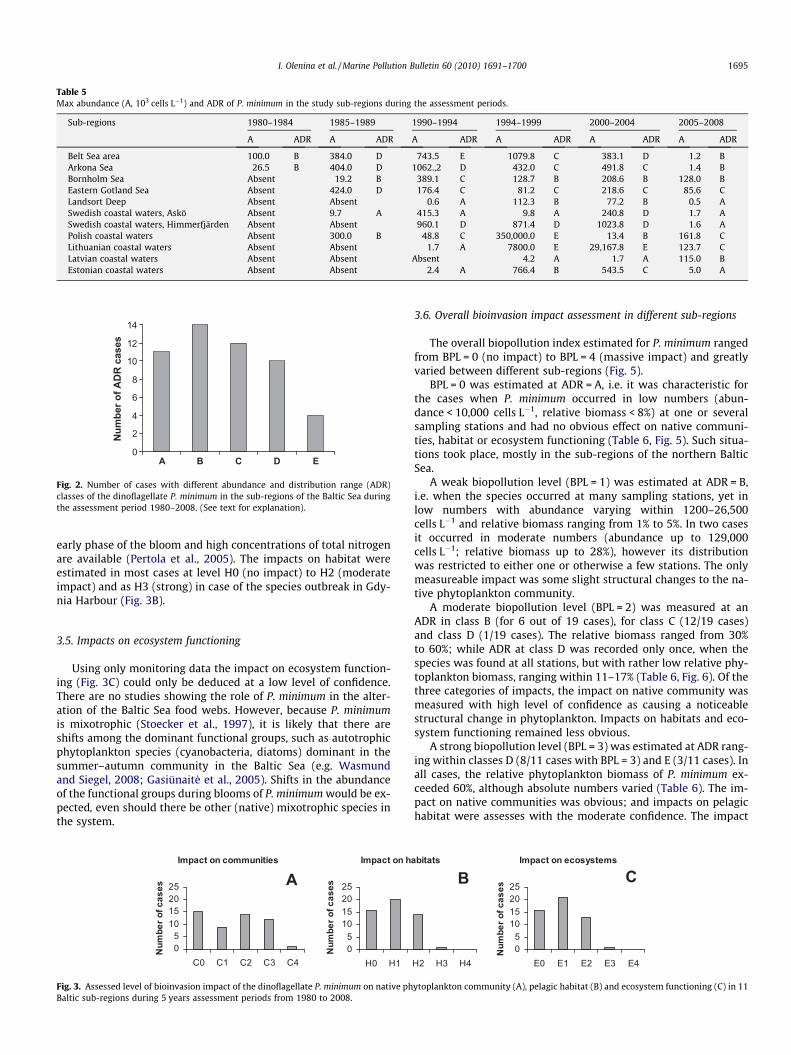
Table 5Max abundance (A, 103 cells L�1) and ADR of P. minimum in the study sub-regions during the assessment periods.
Sub-regions 1980–1984 1985–1989 1990–1994 1994–1999 2000–2004 2005–2008
A ADR A ADR A ADR A ADR A ADR A ADR
Belt Sea area 100.0 B 384.0 D 743.5 E 1079.8 C 383.1 D 1.2 BArkona Sea 26.5 B 404.0 D 1062.,2 D 432.0 C 491.8 C 1.4 BBornholm Sea Absent 19.2 B 389.1 C 128.7 B 208.6 B 128.0 BEastern Gotland Sea Absent 424.0 D 176.4 C 81.2 C 218.6 C 85.6 CLandsort Deep Absent Absent 0.6 A 112.3 B 77.2 B 0.5 ASwedish coastal waters, Askö Absent 9.7 A 415.3 A 9.8 A 240.8 D 1.7 ASwedish coastal waters, Himmerfjärden Absent Absent 960.1 D 871.4 D 1023.8 D 1.6 APolish coastal waters Absent 300.0 B 48.8 C 350,000.0 E 13.4 B 161.8 CLithuanian coastal waters Absent Absent 1.7 A 7800.0 E 29,167.8 E 123.7 CLatvian coastal waters Absent Absent Absent 4.2 A 1.7 A 115.0 BEstonian coastal waters Absent Absent 2.4 A 766.4 B 543.5 C 5.0 A
0
2
4
6
8
10
12
14
A B C D E
Num
ber o
f AD
R c
ases
Fig. 2. Number of cases with different abundance and distribution range (ADR)classes of the dinoflagellate P. minimum in the sub-regions of the Baltic Sea duringthe assessment period 1980–2008. (See text for explanation).
I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700 1695
early phase of the bloom and high concentrations of total nitrogenare available (Pertola et al., 2005). The impacts on habitat wereestimated in most cases at level H0 (no impact) to H2 (moderateimpact) and as H3 (strong) in case of the species outbreak in Gdy-nia Harbour (Fig. 3B).
3.5. Impacts on ecosystem functioning
Using only monitoring data the impact on ecosystem function-ing (Fig. 3C) could only be deduced at a low level of confidence.There are no studies showing the role of P. minimum in the alter-ation of the Baltic Sea food webs. However, because P. minimumis mixotrophic (Stoecker et al., 1997), it is likely that there areshifts among the dominant functional groups, such as autotrophicphytoplankton species (cyanobacteria, diatoms) dominant in thesummer–autumn community in the Baltic Sea (e.g. Wasmundand Siegel, 2008; Gasi�unaite et al., 2005). Shifts in the abundanceof the functional groups during blooms of P. minimum would be ex-pected, even should there be other (native) mixotrophic species inthe system.
Impact on communities
0510152025
C0 C1 C2 C3 C4
Num
ber o
f cas
es AImpact on ha
0510152025
H0 H1
Num
ber o
f cas
es
Fig. 3. Assessed level of bioinvasion impact of the dinoflagellate P. minimum on native phBaltic sub-regions during 5 years assessment periods from 1980 to 2008.
3.6. Overall bioinvasion impact assessment in different sub-regions
The overall biopollution index estimated for P. minimum rangedfrom BPL = 0 (no impact) to BPL = 4 (massive impact) and greatlyvaried between different sub-regions (Fig. 5).
BPL = 0 was estimated at ADR = A, i.e. it was characteristic forthe cases when P. minimum occurred in low numbers (abun-dance < 10,000 cells L�1, relative biomass < 8%) at one or severalsampling stations and had no obvious effect on native communi-ties, habitat or ecosystem functioning (Table 6, Fig. 5). Such situa-tions took place, mostly in the sub-regions of the northern BalticSea.
A weak biopollution level (BPL = 1) was estimated at ADR = B,i.e. when the species occurred at many sampling stations, yet inlow numbers with abundance varying within 1200–26,500cells L�1 and relative biomass ranging from 1% to 5%. In two casesit occurred in moderate numbers (abundance up to 129,000cells L�1; relative biomass up to 28%), however its distributionwas restricted to either one or otherwise a few stations. The onlymeasureable impact was some slight structural changes to the na-tive phytoplankton community.
A moderate biopollution level (BPL = 2) was measured at anADR in class B (for 6 out of 19 cases), for class C (12/19 cases)and class D (1/19 cases). The relative biomass ranged from 30%to 60%; while ADR at class D was recorded only once, when thespecies was found at all stations, but with rather low relative phy-toplankton biomass, ranging within 11–17% (Table 6, Fig. 6). Of thethree categories of impacts, the impact on native community wasmeasured with high level of confidence as causing a noticeablestructural change in phytoplankton. Impacts on habitats and eco-system functioning remained less obvious.
A strong biopollution level (BPL = 3) was estimated at ADR rang-ing within classes D (8/11 cases with BPL = 3) and E (3/11 cases). Inall cases, the relative phytoplankton biomass of P. minimum ex-ceeded 60%, although absolute numbers varied (Table 6). The im-pact on native communities was obvious; and impacts on pelagichabitat were assesses with the moderate confidence. The impact
bitats
H2 H3 H4
BImpact on ecosystems
0510152025
E0 E1 E2 E3 E4
Num
ber o
f cas
es
C
ytoplankton community (A), pelagic habitat (B) and ecosystem functioning (C) in 11
7.4
7.6
7.8
8.0
8.2
8.4
8.6
8.8
9.0
9.2
<1.000 1.000-10.000
10.000-100.000
100.000-500.000
500.000-1.000.000
1.000.000-3.000.000
>3.000.000
Abundance (cells/L)
pH
Fig. 4. Increase in pH during mass development of P. minimum in the Lithuanian coastal waters, mean and standard deviation bars: r(184) = 0.203, p<0.05).
Fig. 5. Dynamics of the bioinvasion impacts caused by the dinoflagellate Prorocentrum minimum in the assessment units (Baltic sub-regions) during the 5 years assessmentperiods and the highest level of impact reached.
1696 I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700
on ecosystem functioning was estimated at a low level ofconfidence.
Massive bioinvasion impact (BPL = 4) was estimated when P.minimum was found at all sampling stations within an assessmentunit and comprised up to 98% of total phytoplankton biomass (Ta-ble 6, Fig. 6). Such high BPL was reached only once during theassessment period.
4. Discussion
The intensity of human mediated biological invasions in marineregions is increasing (e.g. Leppäkoski et al., 2002; Galil et al., 2009;Minchin et al., 2009) and there is a growing demand for robust andreliable methodological approaches taking into account the envi-ronmental changes caused by invasive species. We used routine
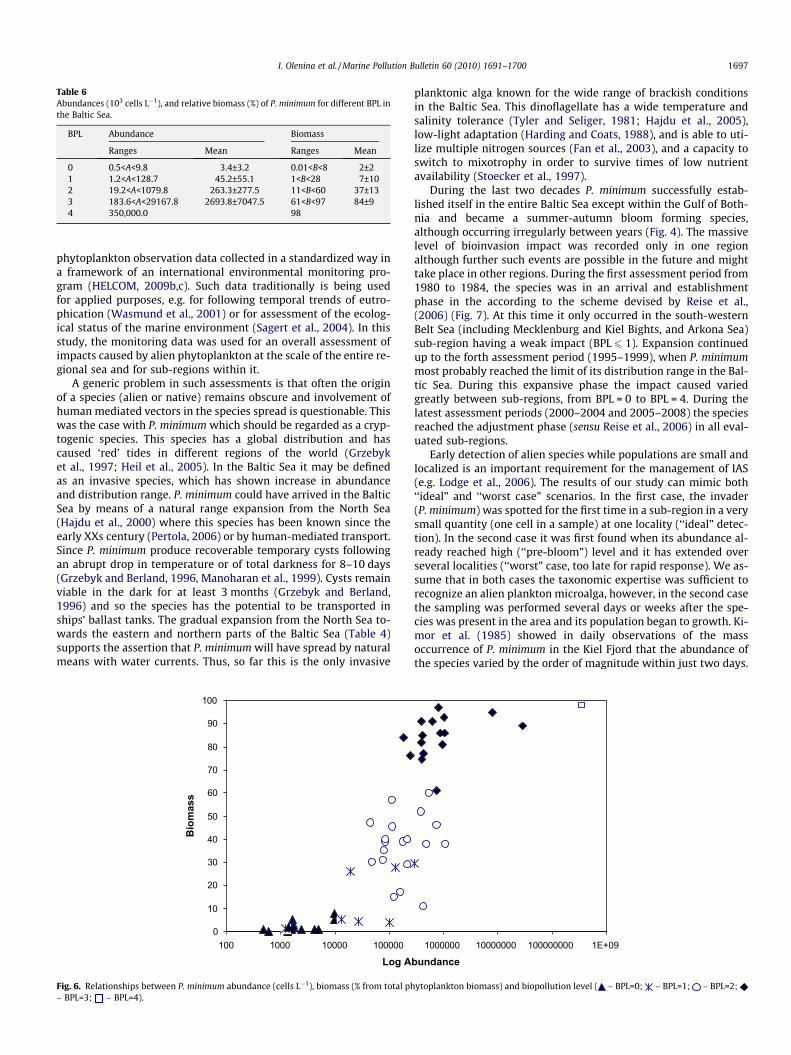
Table 6Abundances (103 cells L�1), and relative biomass (%) of P. minimum for different BPL inthe Baltic Sea.
BPL Abundance Biomass
Ranges Mean Ranges Mean
0 0.5<A<9.8 3.4±3.2 0.01<B<8 2±21 1.2<A<128.7 45.2±55.1 1<B<28 7±102 19.2<A<1079.8 263.3±277.5 11<B<60 37±133 183.6<A<29167.8 2693.8±7047.5 61<B<97 84±94 350,000.0 98
I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700 1697
phytoplankton observation data collected in a standardized way ina framework of an international environmental monitoring pro-gram (HELCOM, 2009b,c). Such data traditionally is being usedfor applied purposes, e.g. for following temporal trends of eutro-phication (Wasmund et al., 2001) or for assessment of the ecolog-ical status of the marine environment (Sagert et al., 2004). In thisstudy, the monitoring data was used for an overall assessment ofimpacts caused by alien phytoplankton at the scale of the entire re-gional sea and for sub-regions within it.
A generic problem in such assessments is that often the originof a species (alien or native) remains obscure and involvement ofhuman mediated vectors in the species spread is questionable. Thiswas the case with P. minimum which should be regarded as a cryp-togenic species. This species has a global distribution and hascaused ‘red’ tides in different regions of the world (Grzebyket al., 1997; Heil et al., 2005). In the Baltic Sea it may be definedas an invasive species, which has shown increase in abundanceand distribution range. P. minimum could have arrived in the BalticSea by means of a natural range expansion from the North Sea(Hajdu et al., 2000) where this species has been known since theearly XXs century (Pertola, 2006) or by human-mediated transport.Since P. minimum produce recoverable temporary cysts followingan abrupt drop in temperature or of total darkness for 8–10 days(Grzebyk and Berland, 1996, Manoharan et al., 1999). Cysts remainviable in the dark for at least 3 months (Grzebyk and Berland,1996) and so the species has the potential to be transported inships’ ballast tanks. The gradual expansion from the North Sea to-wards the eastern and northern parts of the Baltic Sea (Table 4)supports the assertion that P. minimum will have spread by naturalmeans with water currents. Thus, so far this is the only invasive
0
10
20
30
40
50
60
70
80
90
100
100 1000 10000 100000
Log A
Bio
mas
s
Fig. 6. Relationships between P. minimum abundance (cells L�1), biomass (% from total ph– BPL=3; – BPL=4).
planktonic alga known for the wide range of brackish conditionsin the Baltic Sea. This dinoflagellate has a wide temperature andsalinity tolerance (Tyler and Seliger, 1981; Hajdu et al., 2005),low-light adaptation (Harding and Coats, 1988), and is able to uti-lize multiple nitrogen sources (Fan et al., 2003), and a capacity toswitch to mixotrophy in order to survive times of low nutrientavailability (Stoecker et al., 1997).
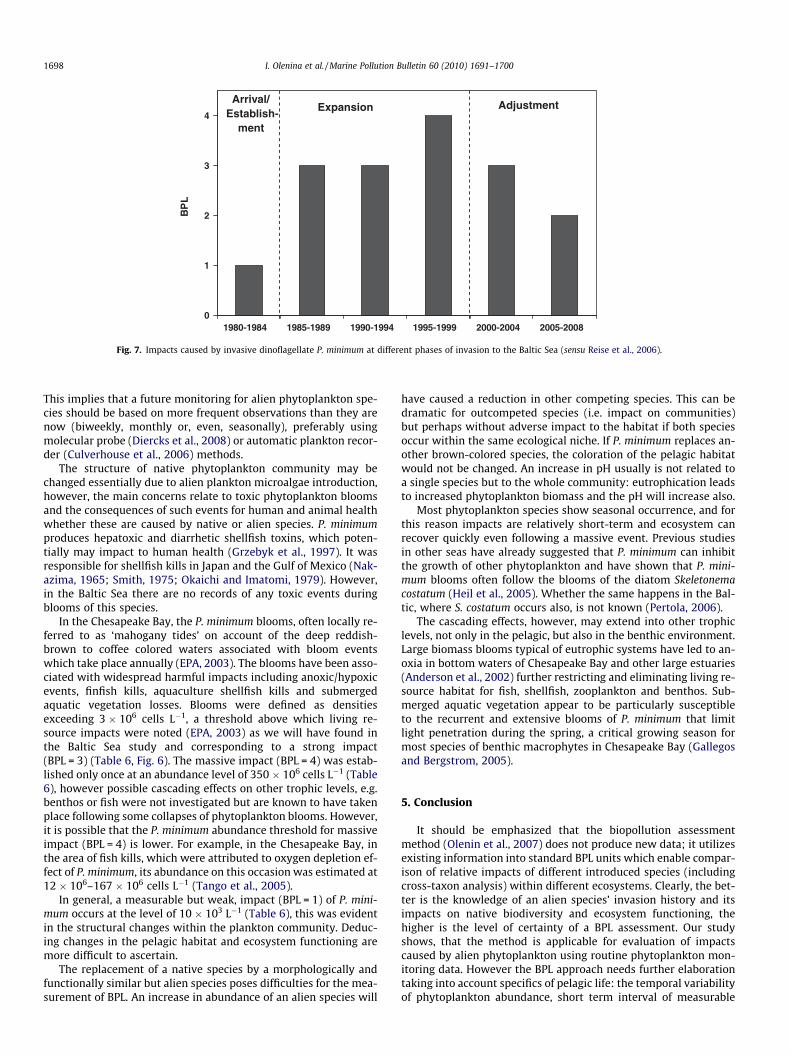
During the last two decades P. minimum successfully estab-lished itself in the entire Baltic Sea except within the Gulf of Both-nia and became a summer-autumn bloom forming species,although occurring irregularly between years (Fig. 4). The massivelevel of bioinvasion impact was recorded only in one regionalthough further such events are possible in the future and mighttake place in other regions. During the first assessment period from1980 to 1984, the species was in an arrival and establishmentphase in the according to the scheme devised by Reise et al.,(2006) (Fig. 7). At this time it only occurred in the south-westernBelt Sea (including Mecklenburg and Kiel Bights, and Arkona Sea)sub-region having a weak impact (BPL 6 1). Expansion continuedup to the forth assessment period (1995–1999), when P. minimummost probably reached the limit of its distribution range in the Bal-tic Sea. During this expansive phase the impact caused variedgreatly between sub-regions, from BPL = 0 to BPL = 4. During thelatest assessment periods (2000–2004 and 2005–2008) the speciesreached the adjustment phase (sensu Reise et al., 2006) in all eval-uated sub-regions.
Early detection of alien species while populations are small andlocalized is an important requirement for the management of IAS(e.g. Lodge et al., 2006). The results of our study can mimic both‘‘ideal” and ‘‘worst case” scenarios. In the first case, the invader(P. minimum) was spotted for the first time in a sub-region in a verysmall quantity (one cell in a sample) at one locality (‘‘ideal” detec-tion). In the second case it was first found when its abundance al-ready reached high (‘‘pre-bloom”) level and it has extended overseveral localities (‘‘worst” case, too late for rapid response). We as-sume that in both cases the taxonomic expertise was sufficient torecognize an alien plankton microalga, however, in the second casethe sampling was performed several days or weeks after the spe-cies was present in the area and its population began to growth. Ki-mor et al. (1985) showed in daily observations of the massoccurrence of P. minimum in the Kiel Fjord that the abundance ofthe species varied by the order of magnitude within just two days.
1000000 10000000 100000000 1E+09
bundance
ytoplankton biomass) and biopollution level ( – BPL=0; – BPL=1; – BPL=2;
0
1
2
3
4
1980-1984 1985-1989 1990-1994 1995-1999 2000-2004 2005-2008
BP
L
Arrival/ Establish-
ment
Expansion Adjustment
Fig. 7. Impacts caused by invasive dinoflagellate P. minimum at different phases of invasion to the Baltic Sea (sensu Reise et al., 2006).
1698 I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700
This implies that a future monitoring for alien phytoplankton spe-cies should be based on more frequent observations than they arenow (biweekly, monthly or, even, seasonally), preferably usingmolecular probe (Diercks et al., 2008) or automatic plankton recor-der (Culverhouse et al., 2006) methods.
The structure of native phytoplankton community may bechanged essentially due to alien plankton microalgae introduction,however, the main concerns relate to toxic phytoplankton bloomsand the consequences of such events for human and animal healthwhether these are caused by native or alien species. P. minimumproduces hepatoxic and diarrhetic shellfish toxins, which poten-tially may impact to human health (Grzebyk et al., 1997). It wasresponsible for shellfish kills in Japan and the Gulf of Mexico (Nak-azima, 1965; Smith, 1975; Okaichi and Imatomi, 1979). However,in the Baltic Sea there are no records of any toxic events duringblooms of this species.
In the Chesapeake Bay, the P. minimum blooms, often locally re-ferred to as ‘mahogany tides’ on account of the deep reddish-brown to coffee colored waters associated with bloom eventswhich take place annually (EPA, 2003). The blooms have been asso-ciated with widespread harmful impacts including anoxic/hypoxicevents, finfish kills, aquaculture shellfish kills and submergedaquatic vegetation losses. Blooms were defined as densitiesexceeding 3 � 106 cells L�1, a threshold above which living re-source impacts were noted (EPA, 2003) as we will have found inthe Baltic Sea study and corresponding to a strong impact(BPL = 3) (Table 6, Fig. 6). The massive impact (BPL = 4) was estab-lished only once at an abundance level of 350 � 106 cells L�1 (Table6), however possible cascading effects on other trophic levels, e.g.benthos or fish were not investigated but are known to have takenplace following some collapses of phytoplankton blooms. However,it is possible that the P. minimum abundance threshold for massiveimpact (BPL = 4) is lower. For example, in the Chesapeake Bay, inthe area of fish kills, which were attributed to oxygen depletion ef-fect of P. minimum, its abundance on this occasion was estimated at12 � 106–167 � 106 cells L�1 (Tango et al., 2005).
In general, a measurable but weak, impact (BPL = 1) of P. mini-mum occurs at the level of 10 � 103 L�1 (Table 6), this was evidentin the structural changes within the plankton community. Deduc-ing changes in the pelagic habitat and ecosystem functioning aremore difficult to ascertain.
The replacement of a native species by a morphologically andfunctionally similar but alien species poses difficulties for the mea-surement of BPL. An increase in abundance of an alien species will
have caused a reduction in other competing species. This can bedramatic for outcompeted species (i.e. impact on communities)but perhaps without adverse impact to the habitat if both speciesoccur within the same ecological niche. If P. minimum replaces an-other brown-colored species, the coloration of the pelagic habitatwould not be changed. An increase in pH usually is not related toa single species but to the whole community: eutrophication leadsto increased phytoplankton biomass and the pH will increase also.
Most phytoplankton species show seasonal occurrence, and forthis reason impacts are relatively short-term and ecosystem canrecover quickly even following a massive event. Previous studiesin other seas have already suggested that P. minimum can inhibitthe growth of other phytoplankton and have shown that P. mini-mum blooms often follow the blooms of the diatom Skeletonemacostatum (Heil et al., 2005). Whether the same happens in the Bal-tic, where S. costatum occurs also, is not known (Pertola, 2006).
The cascading effects, however, may extend into other trophiclevels, not only in the pelagic, but also in the benthic environment.Large biomass blooms typical of eutrophic systems have led to an-oxia in bottom waters of Chesapeake Bay and other large estuaries(Anderson et al., 2002) further restricting and eliminating living re-source habitat for fish, shellfish, zooplankton and benthos. Sub-merged aquatic vegetation appear to be particularly susceptibleto the recurrent and extensive blooms of P. minimum that limitlight penetration during the spring, a critical growing season formost species of benthic macrophytes in Chesapeake Bay (Gallegosand Bergstrom, 2005).
5. Conclusion
It should be emphasized that the biopollution assessmentmethod (Olenin et al., 2007) does not produce new data; it utilizesexisting information into standard BPL units which enable compar-ison of relative impacts of different introduced species (includingcross-taxon analysis) within different ecosystems. Clearly, the bet-ter is the knowledge of an alien species’ invasion history and itsimpacts on native biodiversity and ecosystem functioning, thehigher is the level of certainty of a BPL assessment. Our studyshows, that the method is applicable for evaluation of impactscaused by alien phytoplankton using routine phytoplankton mon-itoring data. However the BPL approach needs further elaborationtaking into account specifics of pelagic life: the temporal variabilityof phytoplankton abundance, short term interval of measurable

I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700 1699
impacts (which may be easily overlooked in existing monitoringpractice), possible shifts in other trophic levels and cascading ef-fects in pelagic and benthic habitats.
Acknowledgements
We would like to thank Dan Minchin of Marine Organism Inves-tigation, Ireland for valuable comments on earlier version of themanuscript. Great thanks to Aleksas Naršcius and Viktoras Didziu-lis of Coastal Research and Planning Institute, Klaipeda Universityfor development of the online BPL assessment system. This studywas supported (for I.O., D.V. and S.O.) by the EU FP7 project MEECE(Marine Ecosystem Evolution in a Changing Environment) and theLithuanian State Science and Studies Foundation project BINLIT(Biological invasions in Lithuanian ecosystems under the climatechange: causes impacts and projections); and by the Swedish EPÁsMarine Monitoring Program (for S.H.).
References
Anderson, D.M., Glibert, P.M., Burkholder, J.M., 2002. Harmful algae blooms andeutrophication: nutrient sources, composition, and consequences. Estuaries 25,704–726.
Baltic Sea Alien Species Database, 2009. In: Olenin, S., Leppäkoski, E., Daunys, D.(Eds.), http://www.ku.lt/nemo/alien_species_search.html (accessed online2009-08-18).
BINPAS, 2009. Biopollution assessment system. Available from: (http://www.corpi.ku.lt/biopollution/) (assessed online 2009-11-22).
Cardoso, A.C., Free, G., 2008. Incorporating invasive alien species into ecologicalassessment in the context of the Water Framework Directive. Aquatic Invasions3 (4), 361–366.
Carlton, J.T., 1996. Biological invasions and cryptogenic species. Ecology 77, 1653–1655.
Carlton, J.T., 2009. Deep Invasion Ecology and the Assembly of Communities inHistorical Time. Ecological Studies, 204 Biological Invasions in MarineEcosystems Springer: 13-56.
Christensen, T., Koch, C., Thomsen, H.A., 1985. Distribution of algae in Danish saltand brackish waters. – University of Copenhagen, 64pp.
Culverhouse, P.F., Williams, R., Benfield, M., Flood, P.R., Sell, A.F., Mazzocchi, M.G.,Buttino, I., Sieracki, M., 2006. Automatic image analysis of plankton: futureperspectives. Mar. Ecol. Prog. Ser. Vol. 312, 297–309.
DAISIE, 2009. DAISIE. Handbook of Alien Species in Europe. Invading Nature –Springer Series In Invasion Ecology. Vol. 3, Springer, 399pp.
Diercks, S., Metfies, K., Medlin, L.K., 2008. Molecular probe sets for the detection oftoxic algae for use in sandwich hybridization formats. J. Plankton Res. 30 (4),439–448.
Edler, L., Hernroth, B., 1999. Planktonåret 1998–Havsmiljön. Juni 1999, 5–6.Edler, L., 2004. Algal situation in Swedish marine waters 10–15 May 2004, SMHI
Oceanic Services. – AlgAware 4: 1–4. Avialable from: http://www.smhi.se/oceanografi/oce_info_data/reports/alg/algsit04_4.pdf.
Edler, L., Ærtebjerg, G., Granéli, E., 1982. Exceptional plankton blooms in theentrance to the Baltic Sea–the Kattegat and Belt Sea area.–International Councilfor the Exploration of the sea. ICES C.M. L 20, 1–6.
Edvardsen, B., Eikrem, W., Shalchian,-Tabrizi, K., Riisberg, I., Johnsen, G., Naustvoll,L., Throndsen, J., 2007. Verrucophora farcimen gen. et sp. nov.(Dictyochophyceae, Heterokonta) – a bloom-forming ichthyotoxic flagellatefrom the Skagerrak, Norway. J. Phycol. 43, 1054–1070.
Elliott, M., 2003. Biological pollutants and biological pollution–an increasing causefor concern. Mar. Pollut. Bull. 46, 275–280.
EPA, 2003. Ambient water quality criteria for dissolved oxygen, water clarity andchlorophyll a for the Chesapeake Bay and its tidal tributaries. US EPA Region IIIChesapeake Bay Program Office, Annapolis, MD. EPA 903-R-03-002, 231pp.
Fan, C., Glibert, P.M., Burkholder, J.M., 2003. Characterization of the affinity fornitrogen, uptake kinetics, and environmental relationships for Prorocentrumminimum in natural blooms and laboratory cultures. Harmful Algae 2, 283–299.
Galil, B.S., Gollasch, S., Minchin, D., Olenin, S., 2009. Alien Marine Biota of Europe. In:DAISIE. Handbook of Alien Species in Europe. Invading Nature – Springer SeriesIn Invasion Ecology. Vol. 3, Springer, pp. 93-104.
Gallegos, C.L., and Bergstrom, P.W., 2005. Effects of a Prorocentrum minimum bloomon light availability for and potential impacts on submersed aquatic vegetationin upper Chesapeake Bay. – Harmful Algae 4: 553–574.
Gasi�unaite, Z.R., Cardoso, A.C., Heiskanen, A.-S., Henriksen, P., Kauppila, P., Olenina,I., Pilkaityte, R., Purina, I., Razinkovas, A., Sagert, S., Schubert, H., Wasmund, N.,2005. Seasonality of coastal phytoplankton in the Baltic Sea: influence ofsalinity and eutrophication. Estuarine. Coastal Shelf Sci. 65 (1–2), 239–252.
Gollasch, S., Mecke, R., 1996. Eingeschleppte Organismen. In: Lozan, J.L., Lampe, R.,Matthaus, W., Rachor, E., Rumohr, H., Westernhagen, H.v. (Eds.), Warnsignaleaus der Ostsee. Parey Buchverlag, Berlin, pp. 146–150.
Gollasch, S., Dammer, M., Lenz, J., Andres, H.G., 1998. Non-indigenous organismsintroduced via ships traffic into German waters. In Carlton (Ed.), Ballast Water:Ecological and Fisheries Implications. ICES Coop. Res. Rep. No. 224, 50-64.
Gómez, F., 2008. Phytoplankton invasions: comments on the validity of categorizingthe non-indigenous dinoflagellates and diatoms in European Seas. Mar. Pollut.Bull. 56, 620–628.
Grzebyk, A., Berland, B., 1996. Influence of temperature, salinity and irradiance ofgrowth of Prorocentrum minimum (Dinophyceae) from the Mediterranean Sea. J.Plankton Res. 18, 1837–1849.
Grzebyk, A., Denardou, B.B., Pouchus, Y.F., 1997. Evidence of a new toxin in the red-tide dinoflagellate Prorocentrum minimum. J. Plankton Res. 19 (8), 1111–1124.
Hajdu, S., 1995. Coscinodiscus wailesii Gran & Angst 1931. – In: Snoeijs, P. &Potapova, M. (Eds.), Intercalibration and Distribution of Diatom Species in theBaltic Sea. – The Baltic Marine Biologists Publication 16c, Vol. 3, 36pp.
Hajdu, S., Edler, L., Olenina, I., Witek, B., 2000. Spreading and Establishment of thePotentially Toxic Dinoflagellate Prorocentrum minimum in the Baltic Sea. J. Int.Rev. Hydrobiol. 85 (5–6), 561–575.
Hajdu, S., Pertola, S., Kuosa, H., 2005. Prorocentrum minimum (Dinophyceae) in theBaltic Sea: morphology, occurrence – a review. Harmful Algae 4, 471–480.
Hällfors, G., Hällfors, S., 2002. New centric diatom in the Baltic Sea. – FinnishInstitute of Marine Research, Annual Report 2001, 21pp.
Hällfors, G., 2004. Checklist of Baltic Sea Phytoplankton Species (including someheterotrophic protistan groups). HELCOM Baltic Sea Environment ProceedingsNo. 95, Helsinki Commission, Helsinki, Finland.
Hansen, G., Daugbjerg, N., Henriksen, P., 2000. Comparative study of Gymnodiniummikimotoi and Gymnodinium aureolum, comb. nov. (=Gyrodinium aureolum)based on morphology, pigment composition, and molecular data. J. Phycol. 36,394–410.
Harding Jr., L.W., Coats, D.W., 1988. Photosynthetic physiology of Prorocentrummarine-leboure (Dinophyceae) during its subpicnocline transport in ChesapeakeBay. J. Phycol. 24, 77–89.
Heil, C.A., Glibert, P.M., Fan, C., 2005. Prorocentrum minimum (Pavillard) Schiller Areview of a harmful algal bloom species of growing worldwide importance.Harmful Algae 4, 449–470.
HELCOM, 1996. Third Periodic Assessment of the State of the Marine Environmentof the Baltic Sea, 1989-1993; Background document. Baltic Sea EnvironmentProceedings, No. 64B. Helsinki Commission, Helsinki, Finland.
HELCOM, 2001. Fourth Periodic Assessment of the. State of the Environment of theBaltic Marine Area, 1994–1998. Baltic Sea Environment Proceedings No. 82A,Helsinki Commission, Helsinki, Finland.
HELCOM, 2009a. Phytoplankton Expert Group Biovolume file. Available from:http://www.ices.dk/env/repfor/PEG_BVOL2009.xls (download 2009-09-07).
HELCOM, 2009b. Phytoplankton sampling frequency. Available from: http://www.helcom.fi/groups/monas/CombineManual/PartA/en_GB/Fig6/ (download2009-09-07).
HELCOM, 2009c. Guidelines concerning phytoplankton species composition,abundance and biomass. Available from: http://www.helcom.fi/groups/monas/CombineManual/AnnexesC/en_GB/annex6/ (download 2009-09-25).
Hustedt, F., 1927–1930. Die Kieselalgen Deutschlands, Österreichs und der Schweizmit Berücksichtigung der übrigen Länder Europas sowie der angrenzendenMeeresgebiete. 1. Teil. – Akademische Verlagsgesellschaft m. b. H. Leipzig, pp.851–855.
Jansson, K., 1994. Alien species in the marine environment. Introductions to theBaltic Sea and the Swedish West coast – Swedish Environmental ProtectionAgency. Report 4357, 1–67.
Kimor, B., Moigis, A.G., Dohms, V., Stienen, C., 1985. A case of mass occurrenceof Prorocentrum minimum in the Kiel Fjord. Mar. Ecol. Prog. Ser. 27, 209–215.
Laing, I., 1999. Coscinodiscus wailesii (Gran & Angst, 1931). In: Exotics across theocean. Case histories on introduced species: their general biology, distribution,range expansion and impact. University of Kiel, Germany, Department ofFishery Biology, Institute for Marine Science, 1–4.
Leppäkoski, E., Gollasch, S., Gruszka, P., Ojaveer, H., Olenin, S., Panov, V., 2002. TheBaltic – a sea of invaders. Can. J. Fish. Aquat. Sci. 59, 1175–1188.
Łotocka, M., 2009. The first recorded bloom of Pseudochattonella farcimen(Dictyochophyceae, Heterokonta), (Riisberg I.2008) in the Gulf of Gdansk.Oceanologia 51 (1), 139–143.
Lodge, D., Williams, S., MacIsaac, H., Hayes, K.R., Leung, B., Reichard, S., Mack, R.N.,Moyle, P.B., Smith, M., Andow, D.A., Carlton, J.T., McMichael1, A., 2006.Biological invasions: recommendations for US Policy and management. Ecol.Appl. 16 (6), 2035–2054.
Mackiewicz, T., 1995. Nanoplankton Zatoki Gdanskiej, Studia i Materiały MIR, seriaA, Nr 32.
Manoharan, K., Lee, T.K., Cha, J.M., Kim, J.H., Lee, W.S., Chang, M., Park, C.W., Cho,J.H., 1999. Acclimation of Prorocentrum minimum (Dinophyceae) to prolongeddarkness by use of an alternative carbon source from triacylglycerides andgalatolipids. J. Phycol. 35, 287–292.
Minchin, D., Gollasch, S., Cohen, A.N., Hewitt, C.L., Olenin, S., 2009. In: G. Rilov, J.A.Crooks, Jeffrey, A. (Eds.), Characterizing Vectors of Marine Invasion. BiologicalInvasions in Marine Ecosystems Ecological, Management, and GeographicPerspectives. Series: Ecological Studies, 204 (XXVI), 109–116.
Nakazima, M., 1965. Studies on the source of shellfish poison in Lake Hamana. I.Relation of the abundance of a species of dinoflagellate Prorocentrum sp. toshellfish toxicity. Bull. Japan Soc. Sci. Fish. 31, 198–203.
Nehring, S., 1994. First living Alexandrium minutum resting cysts in WesternBaltic.–Harmful Algae News. Unesco IOC Newslett. 9, 1–2.

1700 I. Olenina et al. / Marine Pollution Bulletin 60 (2010) 1691–1700
Nikolaev, I.I., 1951. On new introductions in fauna and flora of the North and theBaltic Seas from distant areas [O novyh vselencach v faune i flore Severnogomorja i Baltici iz otdalennyh rajonov]. Zoologicheskij Zhurnal., 30, 556–561 (InRussian).
NOBANIS, 2009. European Network on Invasive Alien Species. Gateway toInformation on Invasive Alien Species in North and Central Europe.(download 2009-09-07).
Occhipinti-Ambrogi, A., Galil, B., 2004. A uniform terminology on bioinvasions: achimera or an operative tool? Mar. Pollut. Bull. 49, 688–694.
Okaichi, T., Imatomi, Y., 1979. Toxicity of Prorocentrum minimum var. mariae-lebouriae assumed to be a causative agent of short-necked clam poisoning. In:Taylor, D.L., Seliger, H.H. (Eds.), Toxic Dinoflagellate Blooms. Elsevier/North-Holland, New York, pp. 385–388.
Olenin, S., and Didziulis,V., 2009. Introduction to the list of alien taxa. In: DAISIE.Handbook of Alien Species in Europe. Invading Nature – Springer Series InInvasion Ecology. Vol. 3, Springer, pp. 129–132.
Olenin, S., Minchin, D., Daunys, D., 2007. Assessment of biopollution in aquaticecosystems. Mar. Pollut. Bull. 55, 379–394.
Olenin, S., Gollasch, S., Jonusas, S., Rimkute, I., 2000. En-route investigations ofplankton in ballast water on a ships voyage from the Baltic Sea to the openAtlantic coast of Europe. Int. Rev. Hydrobiol. 85 (5–6), 577–596.
Orendt, C., Schmitt, C., van Liefferinge, C., Wolfram, G., de Deckere, E., 2009.Include or exclude? A review on the role and suitability of aquaticinvertebrate neozoa as indicators in biological assessment with specialrespect to fresh and brackish European waters. Biological invasions, doi:10.1007/s10530-009-9448-x.
Ostenfeld, C.H., 1908. On the immigration of Biddulphia sinensis Grev. and itsoccurrence in the North Sea during 1903–1907 and on its use for the study ofthe direction and rate of flow of the currents.–Medd. Komm. Havundersøgelser.Plankton 1, 1–44.
Pankow, H., 1990. Ostsee – Algenflora. – Gustav Fischer Verlag, Jena, 648pp.Pertola, S., Faust, M.A., Kuosa, H., 2006. Survey on germination and species
composition of dinoflagellates from ballast tanks and recent sediments in portson the South Coast of Finland, North-Eastern Baltic Sea. Mar. Pollut. Bull. 52,900–911.
Pertola, S., 2006. Diffusive and ship-mediated spread of dinoflagellates in the BalticSea with Prorocentrum minimum as a special case. Doctoral dissertation.University of Helsinki, Faculty of Biosciences, Department of Biological andEnvironmental Sciences and Finnish Institute of Marine Research.
Pertola, S., Kuosa, H., Olsonen, R., 2005. Is the invasion of Prorocentrum minimum(Dinophyceae) related to the nitrogen enrichment of the Baltic Sea? HarmfulAlgae 4, 481–492.
Pyšek, P., Hulme, P.E., Nentwig, W., 2009. Glossary of the main technical terms usedin the handbook. In: DAISIE, The Handbook of Alien Species in Europe, SpringerScience + Business Media B.V., pp. 375–381.
Reise, K., Olenin, S., Thieltges, D.W., 2006. Are aliens threatening European aquaticcoastal ecosystems? Helgoland Mar. Res. 60 (2), 106–112.
Riisberg, I., 2008. Genetic characterization of the marine ichthyotoxic flagellatePseudochattonella farcimen (Heterokonta) and phylogenetic relationshipsamong heterokonts, Ph.D. thesis, Univ. Oslo, Oslo, pp. 1–33.
Riisberg, I., Edvardsen, B., 2008. Genetic variation in bloom-forming ichthyotoxicPseudochattonella species (Dictyochophyceae, Heterokonta) using nuclear,mitochondrial and plastid DNA sequence data, Eur. J. Phycol. 43 (4), 413–422.
Sagert, S., Jensen, D.K., Henriksen, P., Rieling, T., Schubert, H., 2004. Integratedecological assessment of Danish Baltic Sea coastal areas by means ofphytoplankton and macrophytobenthos Estuarine. Coastal Shelf Sci. 63 (1–2),109–118.
Smayda, T.J., 1990. Novel and nuisance phytoplankton blooms in the sea: evidencefor a global epidemic. In: Granéli, E., Sundström, B., Edler, L. & Anderson, D.M.(Eds.), Toxic Marine Phytoplankton. Elsevier, New York, pp. 29–40.
Smith, G.B., 1975. Phytoplankton blooms and reef kills in the mid-eastern Gulf ofMexico. Fla. Mar. Res. Publ. 8, 8.
Stoecker, D.K., Li, A., Coats, D.W., Gustafson, D.E., Nannen, M.K., 1997. Mixotrophy inthe dinoflagellate Prorocentrum minimum. Mar. Ecol. Prog. Ser. 152, 1–12.
Subba Rao, D.V., Sprules, W.G., Locke, A., Carlton, J.T., 1994. Exotic phytoplanktonfrom ships’ ballast waters: risk of potential spread to mariculture sites onCanada’s east coast. Can. Data Rep. Fish. Aquatic Sci. 937, 1–51.
Tango, P.J., Magnien, R., Butler, W., Lacouture, R., Sellner, K., Glibert, P., Luckenbach,M., Poukish, C., Luckett, C., 2005. Characterization of impacts and potentialeffects due to Prorocentrum minimum blooms in Chesapeake Bay. Harmful Algae4, 525–531.
Tyler, M.A., Seliger, H.H., 1981. Selection for a red tide organism: physiologicalresponses to the physical environment. Limnol. Oceanogr. 26, 310–324.
Wasmund, N., Andrushaitis, A., Lysiak-Pastuszak, E., Müller-Karulis, B., Nausch, G.,Neumann, T., Ojaveer, H., Olenina, I., Postel, L., Witek, Z., 2001. Trophic status ofthe South-Eastern Baltic Sea: a comparison of costal and open areas. Estuarine,Coastal and Shelf Science 53, 849–864.
Wasmund, N., and Siegel, H., 2008. Phytoplankton. In: Feistel, R., Nausch, G.,Wasmund, N. (Eds.), State and Evolution of the Baltic Sea, 1952 – 2005. ADetailed 50-Year Survey of Meteorology and Climate, Physics, Chemistry,Biology, and Marine Environment. Wiley, Hoboken, 2008, pp. 441–481.
Witek, B., Plinski, M., 2000. The first recorded bloom of Prorocentrum minimum(PAVILLARD) SCHILLER in the coastal zone of the Gulf of Gdansk. Oceanologia42, 29–36.














![Classifying the Baltic Sea Shallow Water Habitats …...organic matter [9] and suspended particles [9,10], as well as frequent phytoplankton blooms [11]. High concentrations of optically](https://img.pdfslide.net/doc/110x75/5f806076914e67117e6e1831/classifying-the-baltic-sea-shallow-water-habitats-organic-matter-9-and-suspended.jpg)














