Embed Size (px)
Citation preview

289
Size 7.25 x 10 inches
Bacterial Infection in Tasmanian Farmed Abalone: Causes,Pathology, Farm Factors and Control Options
JUDITH HANDLINGER, JEREMY CARSON, LINDA DONACHIE,LES GABOR AND DAVID TAYLOR
Fish Health Unit of the Tasmanian Aquaculture & Fisheries Institute and DPIWEAnimal Health Laboratory, Tasmania, PO Box 46, Kings Meadows,
Tasmania, 7249, Australia
ABSTRACT
Abalone aquaculture based on artificial feeds and intensive generally land-based systemsis increasing in Australia. Intensive systems are vulnerable to diseases of poor environmentcontrol. This paper reports several types of bacterial diseases investigated in grow-outstock of Haliotis rubra, H. laevigata and their hybrids over approximately 10 years, theirpathology, farm factors precipitating disease outbreaks, and the potential for disease controlusing antibiotics. Vibrio species infections have been the most common cause of infection,with a variety of species isolated from moribund animals, generally irrespective of theprimary insult. Specific disease outbreaks, with differing pathology, have been associatedwith two species, Vibrio harveyi and V. splendidus I. Specific pathology has also beenassociated with Flavobacterium-like species. In most cases, stress factors precipitatingdisease has been identified, and control has been largely directed to eliminating this stress.Antibiotics have also been used, with equivocal results, precipitating preliminary studiesof antibiotic absorption and efficacy in these species. The limited potential for antibioticuse, and the requirement for understanding and control of on-farm stress, is discussed.
INTRODUCTION
Despite major wild abalone industries, there are few described diseases of abalone. Withthe emergence of abalone aquaculture industries, more diseases are to be expected, partlydue to closer observation, and partly as the more crowded conditions of animals underculture are more likely to favor disease spread. That the majority of described molluscandiseases are major primary pathogens is partly due to inappropriate animal translocation,but this also reflects the relatively natural food and environment which extensive sea-basedbivalve culture systems provide. In contrast, land-based abalone cultured on artificial feeds,and tank held wild caught stock, are also vulnerable to diseases of poor environment. Theseare often expressed as infections with organisms which are probably ubiquitous.
Understanding the pathogenesis of these diseases and farm factors precipitating diseaseoutbreaks will assist in control through early recognition and correction of the underlyinghusbandry faults. This paper reports characteristics of bacterial disease investigated over
Diseases in Asian Aquaculture V
Handlinger, J., J. Carson, L. Donachie, L. Gabor and D. Taylor. 2005. Bacterial infection in Tasmanian farmed abalone:Causes, pathology, farm factors and control options. In P. Walker, R. Lester and M.G. Bondad-Reantaso (eds). Diseases inAsian Aquaculture V, pp. 289-299. Fish Health Section, Asian Fisheries Society, Manila.
Judith Handlinger et al
290
Size 7.25 x 10 inches
10 years in Tasmanian grow-out stock of Blacklip (Haliotis rubra), Greenlip (H. laevigata)and their hybrids.
MATERIAL AND METHODS
Abalone ranging upward from 1 cm shell size were received either live or fixed whole inseawater formalin as part of routine diagnostic investigations of mortality occurring inaquacultured and occasionally wild harvest stock in temporary holding. Live animals wereexamined using a dissecting microscope and surface lesions were examined by phase contrastmicrocopy of wet smears from areas of pallor. For histopathology, tissues or whole juvenileabalone were fixed in seawater formalin, after benzocaine euthanasia and shell removal.After 24 hours fixation, haematoxylin and eosin stained sections including visible lesions,foot, mantle, mouth, gill, heart, kidneys, stomach, and digestive system, were prepared bystandard methods.
For microbiology, affected tissue or haemolymph were inoculated onto TCBS agar forVibrio species, Johnson’s marine agar (a variant of ZoBell’s 2216), and selective and non-selective marine Shiehs medium for the Flexibacter-Flavobacterium-Cytophaga-like groupof bacteria (FFCLB). Smears for Gram stain were also prepared.
Culture plates were incubated at 25°C for three days. Vibrio isolates were identified usingMicroSys(TM) Vibrio test panels and probabilistic identification software. The test panelcomprises 48 conventional phenotypic tests and the reference database is regularly updatedto include data for 56 species in the genera Vibrio, Photobacterium and Moritella. Yellowpigmented spreading and non-spreading colonies from marine Shieh’s medium werecharacterised by cell morphology, Gram reaction, oxidase and acid production from glucose.Colonies suggestive of Tenacibaculum maritimum were identified by PCR using a 16SrRNA primer set (Carson, 1998; Wilson, et al., 2002).
Table 1. Characteristics of Vibrio splendidus I and Vibrio harveyi isolated from diseased abalone.
Vibrio splendidus I Vibrio harveyiTest 01/2567 Expected 03/0411 ExpectedSwarming - - - 13Growth in 0% NaCl - - - -Thornley’s Arginine + + - -Moller’s Lysine - - + +Moller’s Ornithine - - - 96Nitrate reduction + + + +Oxidase + + + +Indole + + + +ONPG + 751 - 67Voges Proskauer - - - -Resistance to O129 10 µg . - - + 71Resistance to O129 150 µg - - - 17Resistance to Ampicillin 10 µg - - - 92Resistance to Polymyxin B 50 iu - - - 13Resistance to Tellurite (0.0005%) - - - -Aesculin hydrolysis + 50 + 53Alginase - 50 + 38
Bacterial Infection in Tasmanian Farmed Abalone:Causes, Pathology, Farm Factors and Control Options
291
Size 7.25 x 10 inches
RESULTS
There was no obvious species difference in susceptibility to the bacterial diseases observed,and the results combine findings from both species and their hybrids.
Septicaemic vibriosis
Vibrio species were commonly associated with moribund abalone, irrespective of the primaryinsult. Spread within contact populations has so far been associated with outbreaks of onlytwo Vibrio species, V. harveyi and V. splendidus I (see Table 1 for typical characteristics),the pathology of which is described below. Outbreaks typically occurred as peaks of mortalityin summer, with few gross or clinical signs of infection, following a sharp increase in watertemperature, or other stresses related to handling or water quality.
Urease - - - 50Fermentation: L-arabinose - - - -
Arbutin acid - 17 - 71Mannose acid + 75 + +Salicin acid - - - 88Sorbitol acid - 17 - 4Sucrose acid - 67 - 29Mannitol acid + + + +
Utilisation: L-arabinose - - - -Cellobiose + + + +D-galactose + 75 - 58D-glucose + + + +D-mannose + 75 + +Melibiose - - - 8Lactose - - - -Melizitose - 50 - -Sucrose - 50 - 25Trehalose + + + +Xylose - - - -Ethanol - - - 4Glycerol + + - 71Propanol - - - 13D-sorbitol - 50 - 4Gluconate + 25 + 97D-glucuronate - 75 + 89Amygdalin - - - 8Arbutin - - - 33L-citrulline - 75 - 8L-hydroxy proline - - - 17L-leucine - - - -D-glucosamine - - + 4DL-3-hydroxybutyrate - - - -(-ketoglutarate - + - 75Succinate + + + 88
Identification V. splendidus I V. harveyiWillcox probability score 0.997 0.9991Percent strains positive

Judith Handlinger et al
292
Size 7.25 x 10 inches
Frequently, low levels of mixed Vibrio species were isolated. Other Vibrios isolated and ofpathogenic potential included V. tubiashii, V. anguillarum, and recently V. tapetis from amixed infection in a wild caught abalone. Many Vibrio isolates could not be speciated.
V. splendidus I
One sustained outbreak of V. splendidus I was studied, though this was also a commonsporadic isolate from moribund animals. On two other occasions this was consistently isolatedfrom all animals in an affected group, sometimes as part of a mixed infection. Howeverthese outbreaks were not sustained.
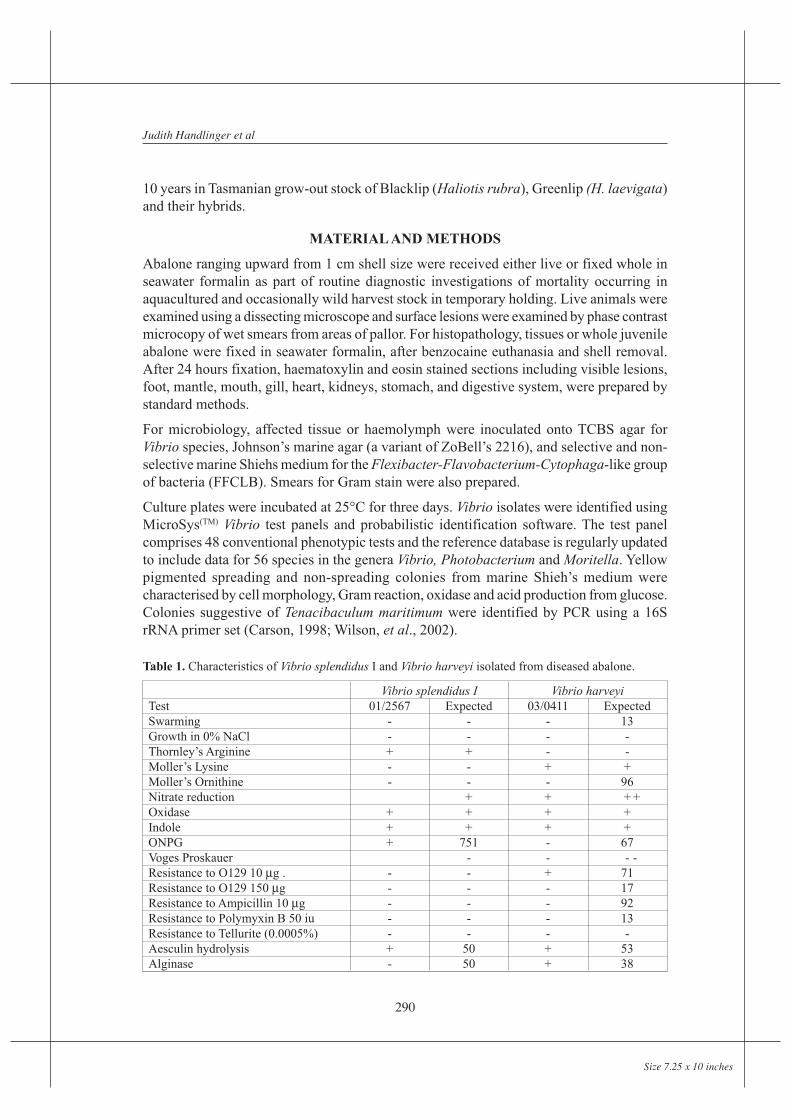
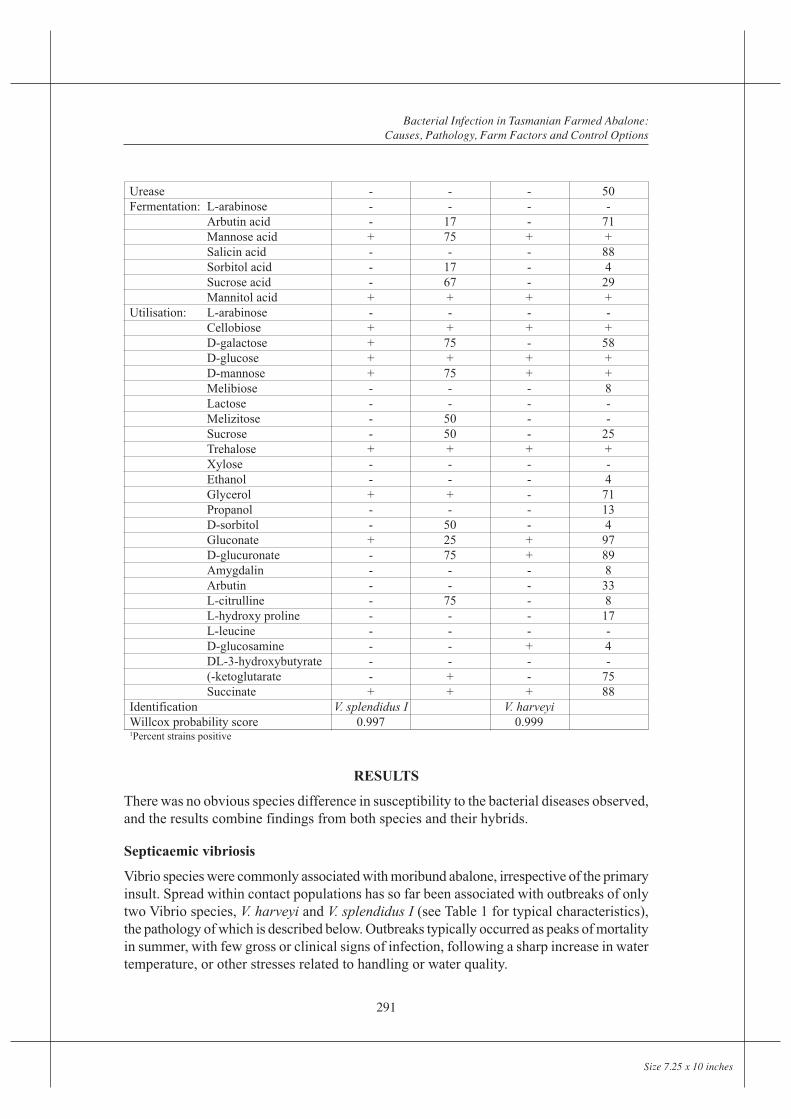
The studied outbreak showed similar pathology to most septicaemias, and had the followingcharacteristics: As well as being detectable in haemolymph, bacteria were typicallydetectable, often in large numbers, in the peri-gut haemocyte region. Selective degenerationof the peri-gut haemocytes despite an intact gut epithelium, distinguished the infectionfrom peri-mortem invasion, but not necessarily from other Vibrio infections (Figure 1).Localization occurred less frequently within other vascular beds such as heart muscle whichappeared selectively targeted by this species (Figure 2), gills, left kidney, or connectivetissue round major ganglia (Figure 3). Localization was often intense in the left kidney,which appeared as a site of selective filtration. Bacteria were often trapped within proteindeposits, which were common in the gills as well as left kidney, and were occasionally seenin other locations. Light focal haemocyte responses were seen in tissues such as the heartmuscle, but were generally minimal. Affected peri-gut interstitial tissue frequently appearedoedematous, occasionally hypercellular, suggesting acute and chronic stages of the infection.Very large numbers of bacteria were occasionally present here, despite low numbers inhaemolymph, suggesting this is a site of selective entrapment.
Figure 1. Degeneration of haemocytes (arrow) under intactgut epithelium associated with septicaemia, in this case withVibrio harveyi infection. (Scale bar = 20 mm)
Bacterial Infection in Tasmanian Farmed Abalone:Causes, Pathology, Farm Factors and Control Options
293
Size 7.25 x 10 inches
No specific predisposing factors, and no specific relationship to temperature, have so farbeen identified for initiating V. splendidus infection. However the prolonged outbreakoccurred during winter, while infections at other times were not sustained.
V. harveyi pustule disease
Outbreaks typically occurred after exposure to summer temperatures, and prolongedoutbreaks were often precipitated by short periods of stress, such as several hours ofinterruption to water supply and resulting transient increase in temperature.
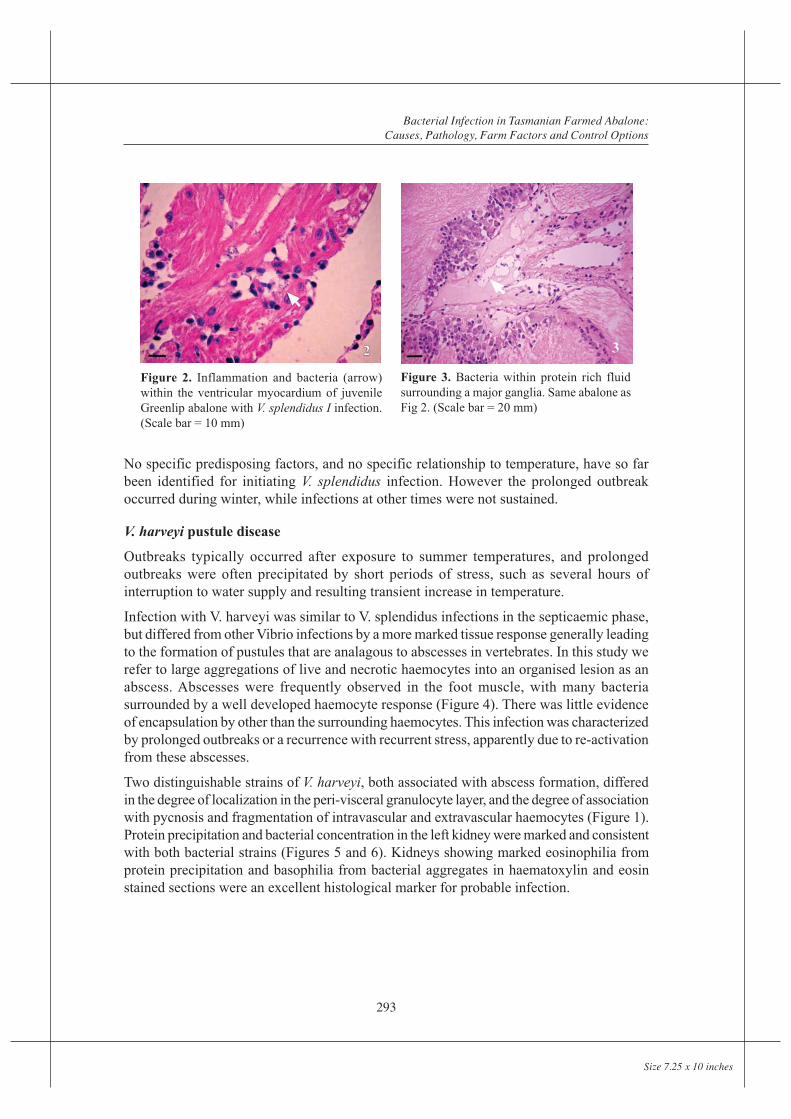
Infection with V. harveyi was similar to V. splendidus infections in the septicaemic phase,but differed from other Vibrio infections by a more marked tissue response generally leadingto the formation of pustules that are analagous to abscesses in vertebrates. In this study werefer to large aggregations of live and necrotic haemocytes into an organised lesion as anabscess. Abscesses were frequently observed in the foot muscle, with many bacteriasurrounded by a well developed haemocyte response (Figure 4). There was little evidenceof encapsulation by other than the surrounding haemocytes. This infection was characterizedby prolonged outbreaks or a recurrence with recurrent stress, apparently due to re-activationfrom these abscesses.
Two distinguishable strains of V. harveyi, both associated with abscess formation, differedin the degree of localization in the peri-visceral granulocyte layer, and the degree of associationwith pycnosis and fragmentation of intravascular and extravascular haemocytes (Figure 1).Protein precipitation and bacterial concentration in the left kidney were marked and consistentwith both bacterial strains (Figures 5 and 6). Kidneys showing marked eosinophilia fromprotein precipitation and basophilia from bacterial aggregates in haematoxylin and eosinstained sections were an excellent histological marker for probable infection.
Figure 2. Inflammation and bacteria (arrow)within the ventricular myocardium of juvenileGreenlip abalone with V. splendidus I infection.(Scale bar = 10 mm)
Figure 3. Bacteria within protein rich fluidsurrounding a major ganglia. Same abalone asFig 2. (Scale bar = 20 mm)
Judith Handlinger et al
294
Size 7.25 x 10 inches
Figures 4-6. The same juvenile abalone from a sustained outbreak of V. harveyi infection. Fig 4 shows anabscess (A) within the foot muscle (M), scale bar = 100 mm. Fig 5 shows protein precipitation (whitearrow) in several of the folia of the left kidney (scale bar = 200 mm). Fig 6 is a higher magnification of thesame kidney, showing diffuse protein deposition as intense eosinophilia (white arrow) in one of the folia,and masses of bacteria in the central vessel of another (black arrow). (Scale bar = 20 mm)
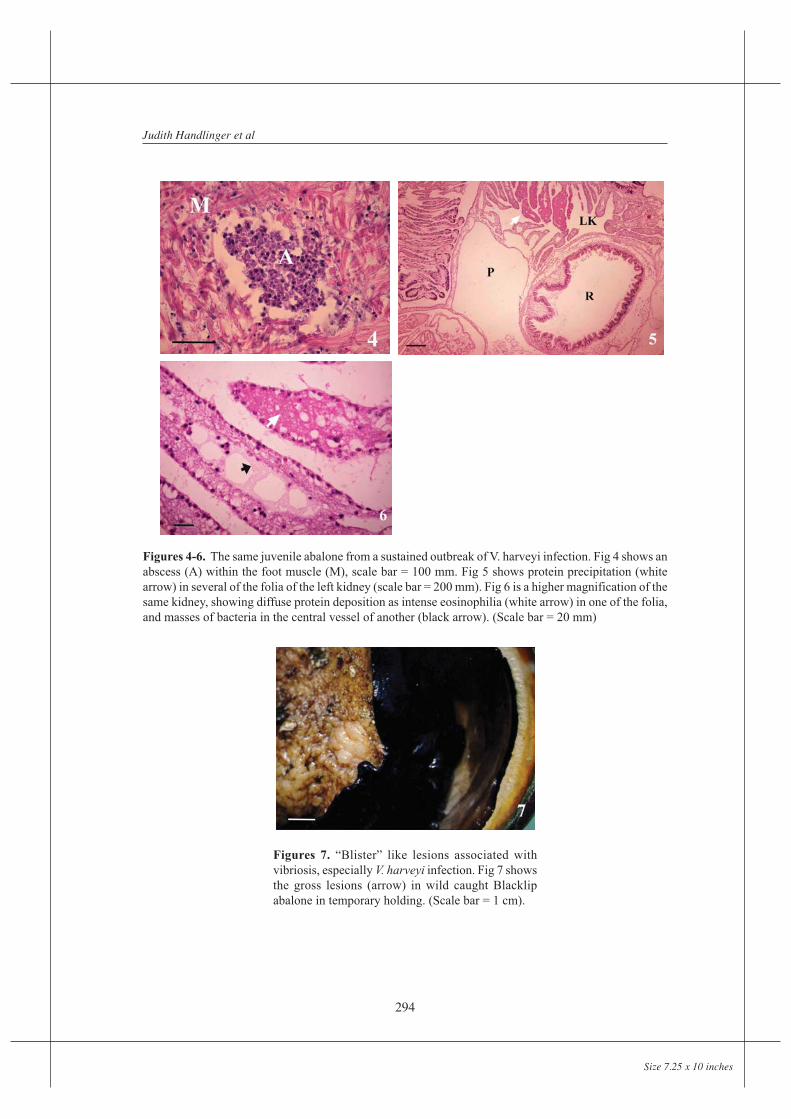
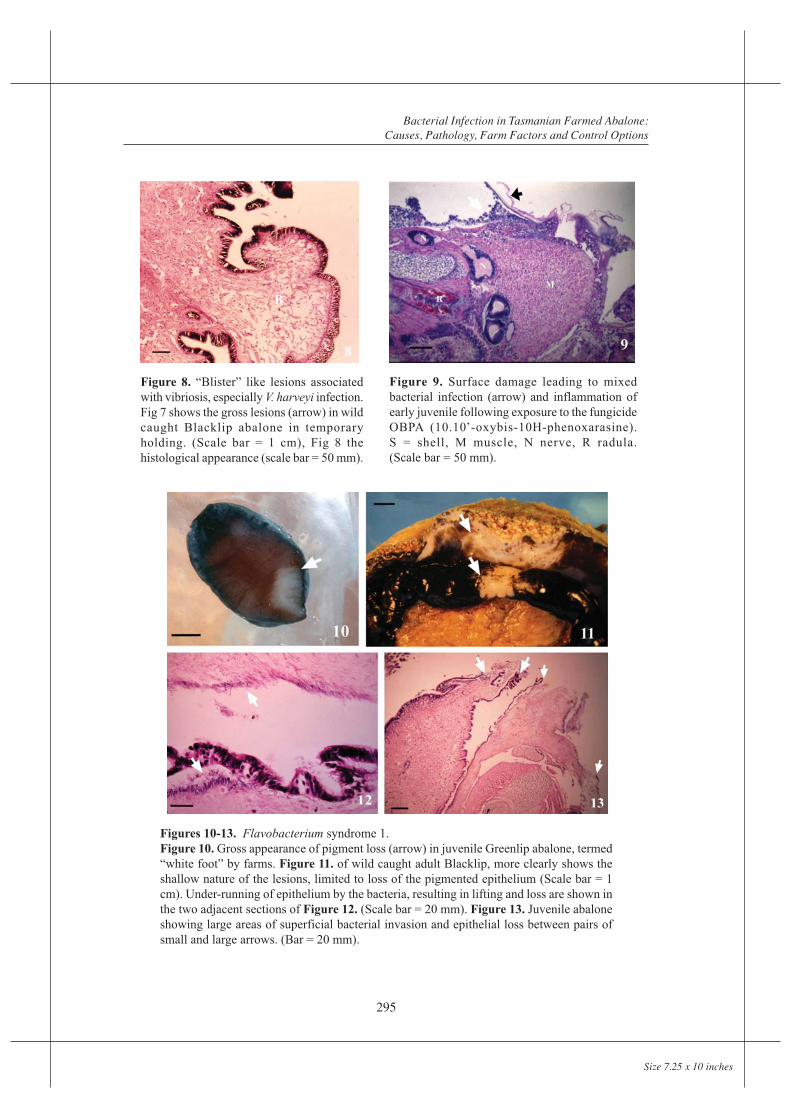
Figures 7. “Blister” like lesions associated withvibriosis, especially V. harveyi infection. Fig 7 showsthe gross lesions (arrow) in wild caught Blacklipabalone in temporary holding. (Scale bar = 1 cm).
Bacterial Infection in Tasmanian Farmed Abalone:Causes, Pathology, Farm Factors and Control Options
295
Size 7.25 x 10 inches
Figure 8. “Blister” like lesions associatedwith vibriosis, especially V. harveyi infection.Fig 7 shows the gross lesions (arrow) in wildcaught Blacklip abalone in temporaryholding. (Scale bar = 1 cm), Fig 8 thehistological appearance (scale bar = 50 mm).
Figures 10-13. Flavobacterium syndrome 1.Figure 10. Gross appearance of pigment loss (arrow) in juvenile Greenlip abalone, termed“white foot” by farms. Figure 11. of wild caught adult Blacklip, more clearly shows theshallow nature of the lesions, limited to loss of the pigmented epithelium (Scale bar = 1cm). Under-running of epithelium by the bacteria, resulting in lifting and loss are shown inthe two adjacent sections of Figure 12. (Scale bar = 20 mm). Figure 13. Juvenile abaloneshowing large areas of superficial bacterial invasion and epithelial loss between pairs ofsmall and large arrows. (Bar = 20 mm).
Figure 9. Surface damage leading to mixedbacterial infection (arrow) and inflammation ofearly juvenile following exposure to the fungicideOBPA (10.10’-oxybis-10H-phenoxarasine).S = shell, M muscle, N nerve, R radula.(Scale bar = 50 mm).

Judith Handlinger et al
296
Size 7.25 x 10 inches
Abscess formation within foot muscle was the most characteristic lesion, though focal fluidpooling within the foot was also common. This presented grossly as surface “blisters”(Figure 7). These are recognized by farms as an indicator of infection, usually but notalways with V. harveyi, leading to their term of “blister disease” for this infection. Bacteriawere only occasionally present in blisters in numbers sufficient to be detectable by normallight microscopy, and cell responses were minimal, the appearance being that of localizedoedema, with apparent rupture of fibres (Figure 8). Vacuolation of epithelium over theseareas was common, often followed by loss of epithelium and rupture of the vesicle.
Secondary surface infection
Surface infection with mixed Vibrio-like bacteria was seen following shell damage by spionidpolychaetes, and exposure to plastic incorporating the fungicide OBPA (10.10’-oxybis-10H-phenoxarasine), which is commonly used in plastics such as swimming pool liners (Figure9). Two syndromes of surface infection with Flavobacterium-like bacteria, were also seen.
Flavobacterium syndrome 1
Presenting signs of loss of pigmentation and contact avoidance were seen on at least nineoccasions, including six tank outbreaks (Figures 10 and 11). These lesions progressed topale flaking epithelium and shallow epithelial erosions of the foot, epipodium, occasionallythe mantle and superficial mouth parts, with masses of filamentous bacteria on the surfaceand under-running and replacing the epithelium (Figure 12). There was limited superficialpenetration by bacteria and minimal host response. Coalescence of lesions often resulted inlarge areas being denuded of epithelium (Figure 13).
Smears from eroded surfaces showed a mat of debris containing slender filamentousFlavobacterium-like bacteria, often in large numbers, plus debris and other bacteria. Arange of yellow pigmented colonies were isolated from sites of erosion in which filamentousbacteria were visible in smears or from haemolymph. On marine Shieh’s medium, typicalcolony types were 1-3 mm in diameter, smooth, butyrous and non-adherent. The bacteriaappeared as filamentous Gram negative rods, oxidase positive some of which fermented
Figures 14-15. Flavobacterium syndrome 2.Figure 14. Deep surface necrosis, adult wild caught Greenlip from the summer outbreak.Figure 15. Superficial (black arrows) and deep bacterial invasion (white arrows), by largesometimes beaded bacteria, with minimal visible host response. (Scale bar = 20 µm)

Bacterial Infection in Tasmanian Farmed Abalone:Causes, Pathology, Farm Factors and Control Options
297
Size 7.25 x 10 inches
glucose. There was a marked disparity in cell morphology in isolates between relativelyshort slender rods, 3-5mm long to robust cell types 1.0-1.5 mm wide and 8-10 mm inlength. The variation in cell type was also reflected in colony pigmentation which variedbetween pale to deep golden yellow. Recovery rates were variable and ranged from light toheavy, sometimes represented by a single colony type or mixed colonies discernible byvariable size and pigmentation. No further characterisation was undertaken but on the basisof culture conditions, colony and cell morphology, bacteria of this type are consistent withthe Flexibacter-Flavobacterium-Cytophaga-like group of bacteria. Most isolates could notbe speciated. In some instances colonies typical of Tenacibaculum maritimum (formerlyFlexibacter maritimus) were detected and their identity was confirmed by 16S rRNA PCR.Such outbreaks occurred during the cooler months of May to October, when temperatureswere 11 to 16°C, well within the range of these species. Initiating stress factors were generallytraumatic, or in one case caused by high salinity, suggesting compromised healing andsecondary infection at a time when host defenses may be reduced. Subsequent spread withintanks suggests bacterial load is also important.
Flavobacterium syndrome 2
More aggressive bacterial invasion was seen in one February outbreak in wild Greenlipabalone held at high stocking densities in tanks for 4-8 days at 19-21°C, conditions whichare outside the species optimum (Edwards, 1996). Erosions progressed rapidly within oneto several days to deep necrotic lesions (Figure 14). Larger and occasionally beadedFlavobacterium-like bacteria were present throughout affected sections, sometimes inassociation with other bacteria, and penetrating deeply into the expanding zone of necrosis(Figure 15). Host response was generally minimal. No consistent lesions were detected inother organs.
DISCUSSION
Vibriosis due to Vibrio splendidus has not previously been described from abalone of theseage groups, but the non-specific pathology may make many cases indistinguishable fromterminal opportunist infections. Nor has toxicity to molluscs been described for exposureto plastics containing the fungicide OBPA, though similar toxicity to salmon has beendescribed once previously (Zitko et al., 1985). In both situations the presence of the fungicidemade use of swimming pool liners inappropriate for aquaculture use, at least until leachinghad reduced the fungicide levels.
Mortality associated with white focal foot lesions due to V. harveyi (reported asV. ‘carchariae’, a junior synonym of Vibrio harveyi) has been described from Japan(Nishimori et al., 1998) and France (Nicolas et al., 2002). The pathology of V. harveyiinfection in Tasmania resembles published reports of abalone pustule disease in China dueto V. fluvialis II (Li et al., 1998), although there is a general lack of publication of detailedaspects of abalone pathology such the protein precipitates seen in gills, left kidney andother organs. Though the origin and significance of such precipitates is uncertain, they areapparently an indication of tissue damage, as this is also observed alone with non-infectiousinsults.

Judith Handlinger et al
298
Size 7.25 x 10 inches
Although other Vibrio species were isolated, their occurrence was isolated and probablyopportunistic, or may represent normal bacterial carriage as low levels of mixed Vibriospecies were also isolated from clinically normal animals held for experimental trials(Handlinger et al., 2002).
Flavobacterium infections of abalone resemble those of fin-fish and have been brieflydescribed previously (Handlinger et al., 2001b), and seen recently in New Zealand (B.Diggles, pers com). Although there were apparent differences in bacterial morphology,clinical expression, pathology and initiating stress factors between the summer and winteroutbreaks, it is uncertain if these differences are due to bacterial type, or to a combinationof elevated bacterial activity at high temperatures and reduced host responses with stressand with low temperatures. Nor do bacterial characteristics determine at this stage if asingle type of FFCLB is causing the problem. On the evidence so far it is probably a syndromethat involves a range of similar opportunist bacteria of the FFCLB group. Further work isrequired to characterize fully this poorly described group of bacteria.
The potential devastating losses from these infections makes antibiotic use potentiallyvaluable, especially where pustules result in on-going disease. However a preliminary trialof antibiotic absorption and efficacy in H. rubra suggests the value may be limited(Handlinger et al., 2001a; 2002). There was moderate absorption of oxytetracycline (OTC)from in-feed treatment, but poor absorption of sulphadiazine, amoxicillin and the quinoloneoxolinic acid. Only oxolinic acid was well absorbed by bath treatment. Prolonged residuetimes with OTC and an uncertain future for quinolones in aquaculture reduce their usefulness.OTC was only partially effective against V. harveyi infection, and infection persisted withinthe population; trimethoprim was ineffective. Innate soluble antibacterial factors variedsignificantly between groups of abalone, suggesting variable susceptibility may be significantand may offer better avenues for control and monitoring. Thus antibiotic treatment appearsat best short-term, with significant environmental and food safety regulation concerns.
CONCLUSIONS
It was concluded that abalone react differently to different bacterial pathogens, which hasimplications for disease control, especially for V. harveyi and similar bacteria which resultin abscess formation leading to persistence of infection in the population. Disease controlby the use of antibiotics is problematic, and future emphasis should be on other means ofcontrol, including management factors.
As environmental and management are significant factors in establishing infections, diseaseinvestigation is of more immediate importance to abalone farmers than to most bivalvemollusc culture, as is more detailed information of mollusc immune responses and of factorswhich may affect these.

Bacterial Infection in Tasmanian Farmed Abalone:Causes, Pathology, Farm Factors and Control Options
299
Size 7.25 x 10 inches
REFERENCES
Carson, J. 1998. Development of molecular probes for use in bacterial disease diagnosis and healthmonitoring of farmed and wild finfish in Australia. Final Report on Project 93/128, FisheriesResearch and Development Corporation, Canberra.
Edwards, S. 1996. Thermal preferences and tolerances of greenlip and blacklip abalone. Proceedingsof the 3rd Annual Abalone Aquaculture Workshop, Port Lincoln, SARDI, Adelaide, pp.65-67.
Handlinger, J., Harris, J., Carson, J. and Taylor, D. 2001. Investigation into the potential use ofantibiotics for disease control in abalone: A preliminary report. In Proceedings of the 8thAnnual FRDC Abalone Aquaculture Workshop, July 2001, Fremantle, Western Australia. pp.83-96.
Handlinger, J., Taylor, D. and Carson, J. 2001a. Flavobacterium-like infection of abalone. (Abstract).In Proceedings of the European Association of Fish Pathologists Tenth InternationalConference, September 2001. P P-209.
Handlinger, J., Carson, J. Harris, J and Taylor D. 2002. Antibiotic treatment of Vibrio harveyi infectionin Haliotis rubra. In Proceedings of the 9th Annual FRDC Abalone Aquaculture Workshop,July 2002, Queenscliff, Victoria. pp 1-12.
Li, T.W., Ding, M. Zhang, J. Xiang, J. and Liu, R. 1998. Studies on the pustule disease of abalone(Haliotis discus hannai Ino) on the Dalian coast. Journal of Shellfish Research 17, 707-711.
Nicolas, J.L., Basuyaux, O., Mazurié, J. and Th_bault, A. 2002. Vibrio carchariae, a pathogen of theabalone Haliotis tuberculata. Diseases of Aquatic Organisms 50, 35-45.
Nishimori, E., Hasegawa, O., Numata, T. and Wakabayashi, H. 1998. Vibrio carchariae causes massmortalities in Japanese abalone, Sulculus diversicolor supratexta. Fish Pathology 33, 495-502.
Wilson, T., Carson, J. and Bowman, J. 2002. Optimisation of one-tube PCR-ELISA to detectfemtogram amounts of genomic DNA. Journal of Microbiological Methods 51, 163-170.

300
Size 7.25 x 10 inches



























![The Novel ABALONE Photosensor Technology - sensl.comsensl.com/.../irp/2017_Ferenc_The_Novel_ABALONE_Photosensor_Technology.pdf · The novel ABALONE Photosensor Technology [1] has](https://img.pdfslide.net/doc/110x75/5d5a3f8888c993d7478bb96c/the-novel-abalone-photosensor-technology-sensl-the-novel-abalone-photosensor.jpg)