Embed Size (px)
Citation preview

Basal body replication in green algae ± when andwhere does it start?
Karl-Ferdinand Lechtreck1)a, Michel Bornensb
a Botanisches Institut, Universität zu Köln, Cologne/Germanyb Institut Curie, Section Recherche, UMR 1444 du CNRS, Paris/France
Received May 30, 2001Received in revised version July 20, 2001Accepted July 25, 2001
Centrioles ± centrin ± polyglutamylation ± Spermatozopsis ±Xenopus egg extract
Basal body duplication in the green alga Spermatozopsissimilis was reinvestigated using GT335, an antibody binding topolyglutamylated tubulins, and antibodies directed to p210, acomponent of the flagellar transition region which representsthe distal border of the basal body. p210 was also detected insmall spots at the base of each basal body which increased insize prior to mitosis. The presence of p210 on one of themicrotubular flagellar roots suggested a transport of basal bodymaterial along these tracks. Immunogold electron microscopyconfirmed the presence of p210 in the probasal bodies. Further,small probasal bodies are apparently connected to the maturebasal bodies by centrin fibers as observed after artificiallyinduced basal body separation in Xenopus egg extract. Whilebasal bodies grew, most of the p210 remained at the tip ofelongating basal bodies, but two or four additional spots wereobserved in distinct patterns near the base of the basal bodies.In cytokinesis, basal body pairs separated and p210 wasobserved in a strong signal at the tip and a weaker one in thevicinity of the proximal end of each basal body. We interpret thedata as indicating that a new p210-containing structure formsnear the proximal end of the basal bodies during basal bodyelongation, representing the precursor of the next generation ofbasal bodies. Thus, basal bodies appear to seed the succeedinggeneration already during their own development, a mecha-nism which could ensure the correct number and position ofbasal bodies.
Introduction
Basal bodies and centrioles share the same highly conservedultrastructure of nine triplet microtubules forming a cylinder.
Two mechanisms of basal body assembly have been described:templated assembly in the vicinity of preexisting basal bodiesand de novo assembly, for example at fibrous granules inmulticiliated metazoan cells (Dirksen, 1991; Marshall et al.,2001). The latter can occur either in cells with or withoutcentrioles. Ultrastructural studies, mostly on multiciliated cells,established the following step-by-step scheme for basal bodydevelopment: it starts from fibrous structures which, in the caseof multiple basal body formation, are termed e.g. deutero-somes, blepharoplasts, or fibrous granules. The first micro-tubular structure to arise is a ring of nine microtubular singlets,held together by a cartwheel consisting of a central hub and ninespokes. The cartwheel is thought to appear first during basalbody development (Mignot, 1996) and is more pronounced ingrowing basal bodies/centrioles while it may disappear uponcentriole maturation (Lange and Gull, 1996; Lechtreck andGrunow, 1999). Then, as the basal body elongates, doublets andtriplets appear, which are interconnected at least at certainlevels of the barrel by A-C- and sometimes by A-A-tubulelinkers (Lechtreck and Grunow, 1999). Alternative scenarios ofcentriole development by breaking of a part of the mother(Matioli, 1999) or by unfolding of a condensed nine-foldsymmetrical precursor (Albrecht-Bühler, 1990) are not wellsupported.
Basal bodies or centrioles usually replicate once per cellcycle. In metazoan cells new centrioles are formed during Sphase laterally on the two preexisting ones. During mitosis,pairs of centrioles each consisting of an old and a newly formedone are present at the spindle poles and subsequently inheritedto the daughter cells (Kochanski and Borisy, 1990). A stringentcontrol of basal body/centriole number seems a prerequisite forcorrect duplication of the centrosome and the formation of abipolar spindle: cells with aberrant centriole numbers tend toform uni- or multipolar spindles which subsequently can resultin genomic instability (Winey, 1996). Recent work by severalgroups has started to reveal the link between the cell cyclemachinery and centriole/basal body replication. Using spermnuclei resuspended in Xenopus egg extract or culture cellswhich overproduce centrioles due to a block in DNA synthesis
0171-9335/01/80/10-631 $15.00/0
1) Dr. K.-F. Lechtreck, Botanisches Institut, Universität zu Köln,Gyrhofstr. 15, D-50931 Cologne/Germany, e-mail: karl.lechtreck@ uni-koeln.de, Fax: � 492214705181.
EJCB 631European Journal of Cell Biology 80, 631 ± 641 (2001, October) ´ � Urban & Fischer Verlag ´ Jenahttp://www.urbanfischer.de/journals/ejcb

it was shown that cdk2/cyclin E or cdk2/cyclin A are necessaryto trigger centriole replication (Hinchcliffe et al., 1999; Laceyet al., 1999; Meraldi, 1999). p34cdc2 and cycB1 have been shownto concentrate in fact at the centrosome in G2-M (Bailly et al.1989, 1992). Mutant Drosophila embryos with defects in mitoticregulators are blocked in certain steps of the centriolar cycle,like initiation, elongation or disengagement of centrioles(Vidwans et al. 1999). Further, centriole separation can beinhibited by proteasome inhibitors indicating that proteolysis isinvolved in the centrosomal cycle (Freed et al., 1999).
Despite its complex ultrastructure, only a limited number ofbasal body proteins have been identified up to now. Amongthose are several members of the tubulin multiprotein family. g-tubulin is tightly associated with the wall of the centriolarcylinder (Moudjou et al., 1996), and g-tubulin is indispensablefor basal body assembly as demonstrated by blocking itsexpression in Paramecium (Ruiz et al., 1999). Recent work hascharacterized the role of a ring complex containing g-tubulinduring microtubule assembly (for a review see (Schiebel,2000)), but whether a similar complex is involved in theassembly of triplet microtubules is unknown. Mutant analysis ofChlamydomonas has identified d-tubulin, which is necessaryfor the assembly or maintenance of the C-tubule (Dutcher andTrabuco, 1998; Garreau De Loubresse et al., 2001). Recently, itwas shown that h-tubulin, another new member of the tubulinfamily, is required for basal body duplication in Paramecium(for review see (Dutcher, 2001)). Further, basal bodies of bald-2mutation, which has not yet been characterized, consist of onlynine singlet microtubules (Goodenough and St. Clair, 1975).Centrin is present in the distal lumen of centrioles inmammalian cells (Paoletti et al., 1996), and it is the predomi-nant protein of various contractile, basal body-associated fibersin protists (Melkonian et al., 1991). The major component ofbasal bodies and centrioles is the ab-tubulin heterodimer. Basalbody a-tubulin is polyglutamylated with long side chains(Geimer et al., 1997b) and antibodies directed against poly-glutamylated tubulin (GT335) destabilize centrioles whenintroduced into the cells (Bobinnec, 1998). A comparativeanalysis in green flagellates using GT335 indicated that newbasal bodies become polyglutamylated while they elongate(Lechtreck and Geimer, 2000).
Recently, we identified p210, a protein present in the flagellartransition region of the green flagellate Spermatozopsis similis(the part of the flagellum located between the distal end of thebasal body and the axoneme (Lechtreck et al., 1999)). Thepartial cDNA sequence of p210 encodes a highly repetitiveprotein which has no homologue in the data base exceptunconvincing similarities to proteins with a similar amino acidcomposition (rich in A, P, and Q). Using monoclonal andpolyclonal antibodies, we located p210 in the fibrous materialcovering the outer surface of the doublet microtubules andlinking them to the plasma membrane. Further, p210 wasalready observed at the distal end of probasal bodies wellbefore the transition region is formed or membrane contact ofbasal bodies is established. Thus, p210 could also have a roleduring basal body development. Reexamination of the distri-bution of p210 in the green alga Spermatozopsis similis nowrevealed that p210 is present at both the distal and proximal endof elongating basal bodies. Based on this observation and on thebehavior of the connecting centrin fibers in Xenopus eggextract, we discuss general models of basal body developmentwhich imply the formation of basal body precursors alreadyduring the assembly of the preceding generation of basal bodies.
Material and methods
Indirect immunofluorescenceCells of S. similis were cultured as described previously (McFadden andMelkonian, 1986). Cells were harvested by centrifugation, washed oncein MT (30 mM HEPES, 5 mM Na-EGTA, 15 mM KCl, 5 mM MgSO4,pH 7.0), and permeabilized by the addition of Nonidet P-40 (0.25% finalconcentration in MT). Immediately after permeabilization, cytoskele-tons were fixed for approximately 25 minutes in 3% formaldehyde/0.25% glutaraldehyd and allowed to adhere to poly-L-lysine coatedmultiwell slides. After washing with PBS and blocking (20 minutes withPBS/0.05% Tween/0.2% gelatine/1% BSA), multiwell slides werestored at ÿ20 8C in PBS/glycerol (1 :2) or used directly. Cytoskeletonswere incubated with a mixture of the primary antibodies (anti-p210,1 : 100, and GT335, 1 :1500 ± 2500, or anti-centrin BAS6.8, 1 : 2 inblocking buffer). For origin of antibodies, refer to (Wolff et al., 1992;Geimer et al., 1997b; Lechtreck et al., 1999). After washing with PBSand brief blocking, secondary antibodies (cyanine 3 (Jackson Immu-noResearch) and FITC (Sigma)) were applied for 90 minutes in thedilutions suggested by the manufacturers. Specimen were mounted inPBS/glycerol containing DABCO as an anti-fading reagent.
After immunostaining, cytoskeletons were imaged on LeicaDMRXA or Leica DMRD microscopes. The latter was equipped witha 8-bit CCD camera (Hamamatsu). The Leica DMRXA allowed therecording of image stacks (200-nm steps) using a piezoelectric objectivepositioning device and a MicroMAX CCD camera (Princeton Instru-ments). With a 100� 1.4 NA objective the final magnification on thechip was 67 nm/pixel. Metamorph was used for deconvolution andstacking of images. Image processing was done in Photoshop andIllustrator.
Use of Xenopus egg extractLow speed Xenopus oocyte extracts (15000g) blocked in metaphase IIof meiosis were prepared according to the protocol of Murray (1991).For shifting extract from mitosis to interphase 0.3 mM CaCl2 wereadded. Histone H1 kinase activity was measured essentially as describedby Murray (1991). Cells of S. similis were harvested, resuspended in MT,and glued for 2 minutes to poly-L-lysine-coated multiwell slides. Cellswere lysed with NP-40 (0.5% in MT), washed several times in extractbuffer and covered with Xenopus egg extract. Preincubation withmonoclonal anti-centrin (BAS6.8 diluted 1 : 1 in extract buffer) wasperformed for 15 ± 20 min. After 60 ± 90 minutes at 20 8C, slides werecovered with BRB80 (80 mM K-Pipes, pH 6.8, 1 mM MgSO4, 1 mMEGTA) containing 10 mM Nocodazol and cooled on ice for 5 ± 15minutes. Alternatively, cytoskeletons were prepared as described above,washed once in MT, and resuspended in 50 ml extract. After 40 min,10 mM Nocodazol was added, the samples were incubated on ice for10 min, and diluted with BAB80 containing 10 % glycerol. Cytoskele-tons were centrifuged (10000g, 10 min, 4 8C) on poly-L-lysine-treatedcoverslips. AMP-PNP was used at 2.5 mM in extracts prepared withoutenergy mix (1 mM ATP, 7.5 mM creatine phosphate, 1 mM MgSO4, finalconcentrations). Finally, slides were washed in BAB80, fixed andimmunostained as described above.
Immunogold electron microscopyPreembedding immunogold electron microscopy of isolated cytoskele-tons of S. similis has been described previously (Lechtreck et al., 1999).To visualize primary antibodies, protein A conjugated to 20-nm goldparticles diluted in blocking buffer was used. Ultrathin sections(<100 nm) were cut with a diamond knife (DDK, Wilmington, DE)on an RMC MT-6000 microtome (RMC, Tucson, AZ) and mounted onPioloform-coated slot grids. The sections were stained with lead citrateand uranyl acetate (Reynolds, 1963) and observed with a Philips CM 10or CM 120 transmission electron microscope.
632 K.-F. Lechtreck, M. Bornens EJCB

Results
Basal body precursors in interphase cellsIn this article the term probasal body is used for immature,flagella-less basal bodies and distinguished from fibrousprobasal bodies (fibrous disks without nine-fold symmetricmircotubular scaffold) and from basal body precursors (singledots observed with anti-p210 but otherwise structurally ill-defined).
Interphase cells of Spermatozopsis similis contain twoflagella-bearing basal bodies, which, in indirect immunofluo-rescence, can be visualized as two dots with anti-p210 (Fig. 1).The p210 signal is located in the transition region, which indouble labeling experiments with GT335, an antibody specificfor polyglutamylated tubulin, was visible as a gap between theaxonemes and the strongly polyglutamylated basal bodies(Fig. 1 a ± c, Fig. 4). Interphase cells often contained twoadditional tiny dots stained with anti-p210 in the space betweenthe two basal bodies (Fig. 1a, h). Deconvoluted images showedthat the two small dots are located near the proximal ends of themature basal bodies (Fig. 1a, b). The merged and enlargedversion of a and b is shown in Fig. 1c and is cartooned in Fig. 1d.These dotty p210 signals occurred consistently in logarithmi-cally growing cultures, e.g. when most of the cells were going todivide, whereas in lag-phase cultures signals tended to be veryweak or absent (see Fig. 4B l). In logarithmic cultures of S.similis cell division occurs between 9 and 12 hours after theonset of light in a L/D cycle of 14/10. In cytoskeletons preparedfrom cells in the second half of light phase, we observed fourdots as before but the tiny anti-p210 dots near the base of thebasal bodies often had increased in size and intensity andtherefore are addressed as probasal bodies (Fig. 1e). We alsoobserved elongated structures stained with anti-p210 whichterminate in one of the two smaller dots (Fig. 2). Preembeddingelectron microscopy suggested that these signals represented
the five-stranded microtubular roots, which originate in thespace between the two basal bodies below the centrin-contain-ing connecting fiber (Fig. 3b, c; see also Fig. 1d). Serial thinsection analysis of prelabeled material identified the decoratedroot as the one which links the younger basal body to the eyespot apparatus, a specialized part of the chloroplast and theoverlaying plasma membrane utilized by various protists forlight perception (n� 2, not shown). The gold particles repre-senting p210 were observed only on that side of these flatmicrotubular bundles which faces the cell nucleus. Thedecoration could be traced for up to five consecutive sections,approximately to the region where the microtubules bypass the
Fig. 1. Early basal body development in S. similis as visualized withanti-p210. a, b) Deconvoluted images from an interphase cells. a) Anti-p210 staining revealing two large and two smaller dots. b) Thecorresponding GT335 staining shows the two axonemes and thebrightly stained basal bodies. c) Enlarged and merged image of a andb showing that the most of p210 (displayed in white) is present in thetransition region (arrows) between the axoneme (ax) and the basalbody (bb, shown in black), whereas the smaller p210 dots (arrowheads)are located near the base of each basal body. d) Schematic drawing of
the green algal basal apparatus with the immunoreactive parts shown inwhite (anti-p210) and black (GT335). The gray lines represent themicrotubular root system with the number of microtubules indicated. e)Cytoskeleton stained with anti-p210, which marked two large and twosmaller dots, and the corresponding GT335 staining (f). g, h) Twodifferent optical sections through a cytoskeleton containing small p210signals in addition to the decoration of the transition regions. Images ofthis figure were recorded with the Leica DMRXA microscope. Bar�1 mm.
Fig. 2. Cytoskeletons stained with anti-p210 showing elongatedstructures in addition to the decoration of the basal bodies andprobasal bodies. Note that the elongated signals terminate close to theprobasal bodies. a, b) Cytoskeletons with basal bodies in a 1808configuration (as in Fig. 1d), d) in the parallel (08) configuration, and c)basal bodies with an intermediate configuration. Bar� 2 mm.
633Basal body replication in green algaeEJCB
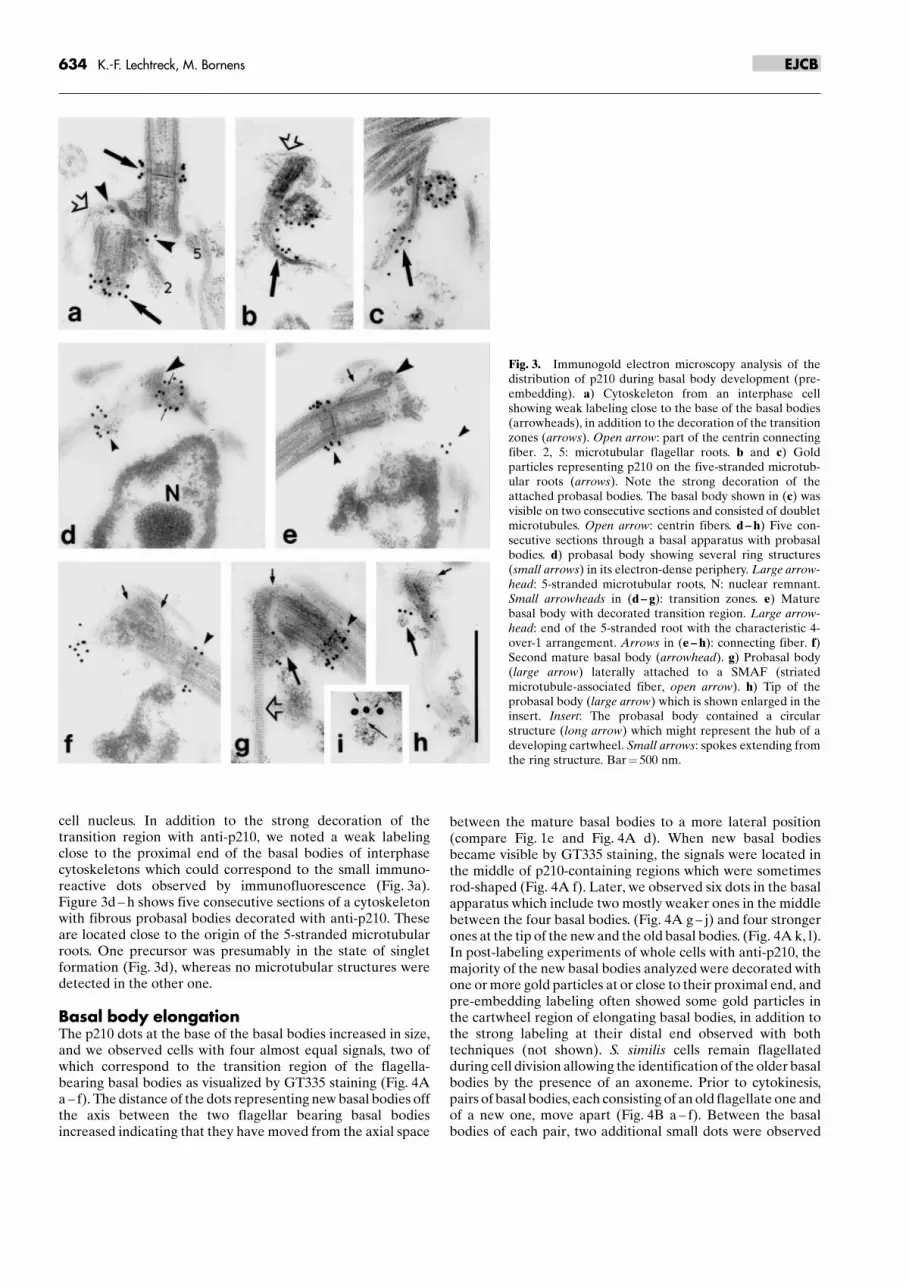
cell nucleus. In addition to the strong decoration of thetransition region with anti-p210, we noted a weak labelingclose to the proximal end of the basal bodies of interphasecytoskeletons which could correspond to the small immuno-reactive dots observed by immunofluorescence (Fig. 3a).Figure 3d ± h shows five consecutive sections of a cytoskeletonwith fibrous probasal bodies decorated with anti-p210. Theseare located close to the origin of the 5-stranded microtubularroots. One precursor was presumably in the state of singletformation (Fig. 3d), whereas no microtubular structures weredetected in the other one.
Basal body elongationThe p210 dots at the base of the basal bodies increased in size,and we observed cells with four almost equal signals, two ofwhich correspond to the transition region of the flagella-bearing basal bodies as visualized by GT335 staining (Fig. 4Aa ± f). The distance of the dots representing new basal bodies offthe axis between the two flagellar bearing basal bodiesincreased indicating that they have moved from the axial space
between the mature basal bodies to a more lateral position(compare Fig. 1e and Fig. 4A d). When new basal bodiesbecame visible by GT335 staining, the signals were located inthe middle of p210-containing regions which were sometimesrod-shaped (Fig. 4A f). Later, we observed six dots in the basalapparatus which include two mostly weaker ones in the middlebetween the four basal bodies. (Fig. 4A g ± j) and four strongerones at the tip of the new and the old basal bodies. (Fig. 4A k, l).In post-labeling experiments of whole cells with anti-p210, themajority of the new basal bodies analyzed were decorated withone or more gold particles at or close to their proximal end, andpre-embedding labeling often showed some gold particles inthe cartwheel region of elongating basal bodies, in addition tothe strong labeling at their distal end observed with bothtechniques (not shown). S. similis cells remain flagellatedduring cell division allowing the identification of the older basalbodies by the presence of an axoneme. Prior to cytokinesis,pairs of basal bodies, each consisting of an old flagellate one andof a new one, move apart (Fig. 4B a ± f). Between the basalbodies of each pair, two additional small dots were observed
Fig. 3. Immunogold electron microscopy analysis of thedistribution of p210 during basal body development (pre-embedding). a) Cytoskeleton from an interphase cellshowing weak labeling close to the base of the basal bodies(arrowheads), in addition to the decoration of the transitionzones (arrows). Open arrow: part of the centrin connectingfiber. 2, 5: microtubular flagellar roots. b and c) Goldparticles representing p210 on the five-stranded microtub-ular roots (arrows). Note the strong decoration of theattached probasal bodies. The basal body shown in (c) wasvisible on two consecutive sections and consisted of doubletmicrotubules. Open arrow: centrin fibers. d ± h) Five con-secutive sections through a basal apparatus with probasalbodies. d) probasal body showing several ring structures(small arrows) in its electron-dense periphery. Large arrow-head: 5-stranded microtubular roots, N: nuclear remnant.Small arrowheads in (d ± g): transition zones. e) Maturebasal body with decorated transition region. Large arrow-head: end of the 5-stranded root with the characteristic 4-over-1 arrangement. Arrows in (e ± h): connecting fiber. f)Second mature basal body (arrowhead). g) Probasal body(large arrow) laterally attached to a SMAF (striatedmicrotubule-associated fiber, open arrow). h) Tip of theprobasal body (large arrow) which is shown enlarged in theinsert. Insert: The probasal body contained a circularstructure (long arrow) which might represent the hub of adeveloping cartwheel. Small arrows: spokes extending fromthe ring structure. Bar� 500 nm.
634 K.-F. Lechtreck, M. Bornens EJCB
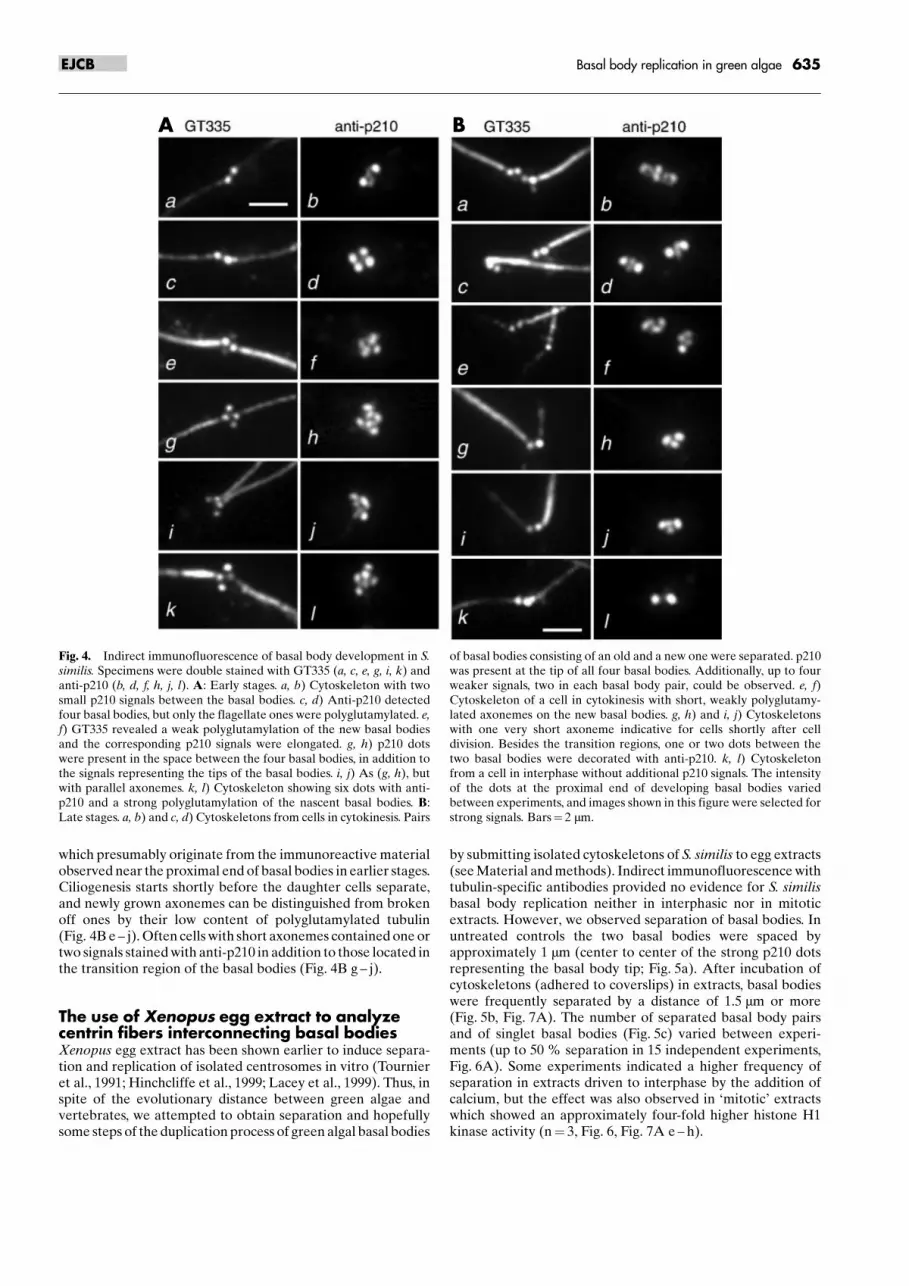
which presumably originate from the immunoreactive materialobserved near the proximal end of basal bodies in earlier stages.Ciliogenesis starts shortly before the daughter cells separate,and newly grown axonemes can be distinguished from brokenoff ones by their low content of polyglutamylated tubulin(Fig. 4B e ± j). Often cells with short axonemes contained one ortwo signals stained with anti-p210 in addition to those located inthe transition region of the basal bodies (Fig. 4B g ± j).
The use of Xenopus egg extract to analyzecentrin fibers interconnecting basal bodiesXenopus egg extract has been shown earlier to induce separa-tion and replication of isolated centrosomes in vitro (Tournieret al., 1991; Hinchcliffe et al., 1999; Lacey et al., 1999). Thus, inspite of the evolutionary distance between green algae andvertebrates, we attempted to obtain separation and hopefullysome steps of the duplication process of green algal basal bodies
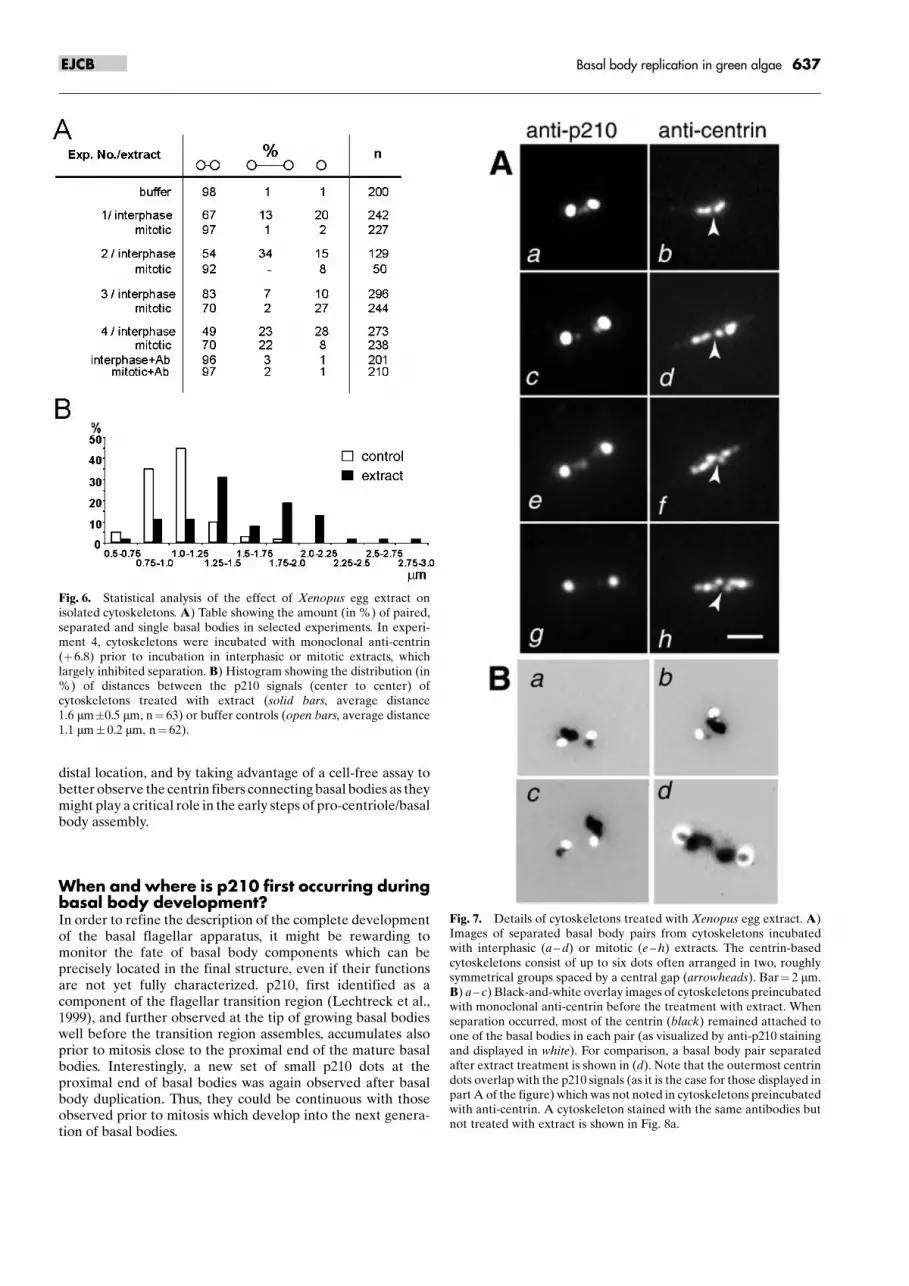
by submitting isolated cytoskeletons of S. similis to egg extracts(see Material and methods). Indirect immunofluorescence withtubulin-specific antibodies provided no evidence for S. similisbasal body replication neither in interphasic nor in mitoticextracts. However, we observed separation of basal bodies. Inuntreated controls the two basal bodies were spaced byapproximately 1 mm (center to center of the strong p210 dotsrepresenting the basal body tip; Fig. 5a). After incubation ofcytoskeletons (adhered to coverslips) in extracts, basal bodieswere frequently separated by a distance of 1.5 mm or more(Fig. 5b, Fig. 7A). The number of separated basal body pairsand of singlet basal bodies (Fig. 5c) varied between experi-ments (up to 50 % separation in 15 independent experiments,Fig. 6A). Some experiments indicated a higher frequency ofseparation in extracts driven to interphase by the addition ofcalcium, but the effect was also observed in �mitotic� extractswhich showed an approximately four-fold higher histone H1kinase activity (n� 3, Fig. 6, Fig. 7A e ± h).
Fig. 4. Indirect immunofluorescence of basal body development in S.similis. Specimens were double stained with GT335 (a, c, e, g, i, k) andanti-p210 (b, d, f, h, j, l). A: Early stages. a, b) Cytoskeleton with twosmall p210 signals between the basal bodies. c, d) Anti-p210 detectedfour basal bodies, but only the flagellate ones were polyglutamylated. e,f) GT335 revealed a weak polyglutamylation of the new basal bodiesand the corresponding p210 signals were elongated. g, h) p210 dotswere present in the space between the four basal bodies, in addition tothe signals representing the tips of the basal bodies. i, j) As (g, h), butwith parallel axonemes. k, l) Cytoskeleton showing six dots with anti-p210 and a strong polyglutamylation of the nascent basal bodies. B:Late stages. a, b) and c, d) Cytoskeletons from cells in cytokinesis. Pairs
of basal bodies consisting of an old and a new one were separated. p210was present at the tip of all four basal bodies. Additionally, up to fourweaker signals, two in each basal body pair, could be observed. e, f)Cytoskeleton of a cell in cytokinesis with short, weakly polyglutamy-lated axonemes on the new basal bodies. g, h) and i, j) Cytoskeletonswith one very short axoneme indicative for cells shortly after celldivision. Besides the transition regions, one or two dots between thetwo basal bodies were decorated with anti-p210. k, l) Cytoskeletonfrom a cell in interphase without additional p210 signals. The intensityof the dots at the proximal end of developing basal bodies variedbetween experiments, and images shown in this figure were selected forstrong signals. Bars� 2 mm.
A B
635Basal body replication in green algaeEJCB
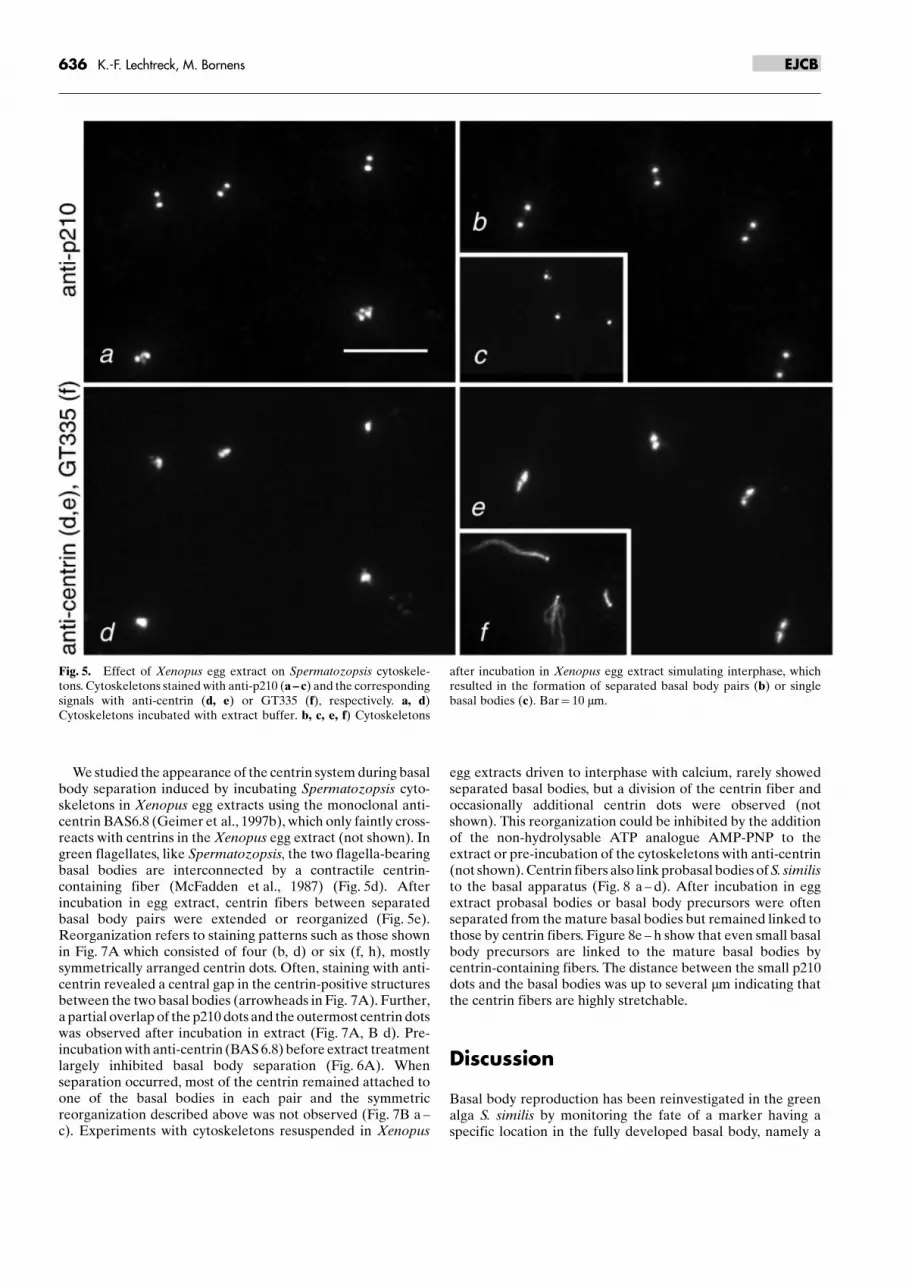
We studied the appearance of the centrin system during basalbody separation induced by incubating Spermatozopsis cyto-skeletons in Xenopus egg extracts using the monoclonal anti-centrin BAS6.8 (Geimer et al., 1997b), which only faintly cross-reacts with centrins in the Xenopus egg extract (not shown). Ingreen flagellates, like Spermatozopsis, the two flagella-bearingbasal bodies are interconnected by a contractile centrin-containing fiber (McFadden et al., 1987) (Fig. 5d). Afterincubation in egg extract, centrin fibers between separatedbasal body pairs were extended or reorganized (Fig. 5e).Reorganization refers to staining patterns such as those shownin Fig. 7A which consisted of four (b, d) or six (f, h), mostlysymmetrically arranged centrin dots. Often, staining with anti-centrin revealed a central gap in the centrin-positive structuresbetween the two basal bodies (arrowheads in Fig. 7A). Further,a partial overlap of the p210 dots and the outermost centrin dotswas observed after incubation in extract (Fig. 7A, B d). Pre-incubation with anti-centrin (BAS 6.8) before extract treatmentlargely inhibited basal body separation (Fig. 6A). Whenseparation occurred, most of the centrin remained attached toone of the basal bodies in each pair and the symmetricreorganization described above was not observed (Fig. 7B a ±c). Experiments with cytoskeletons resuspended in Xenopus
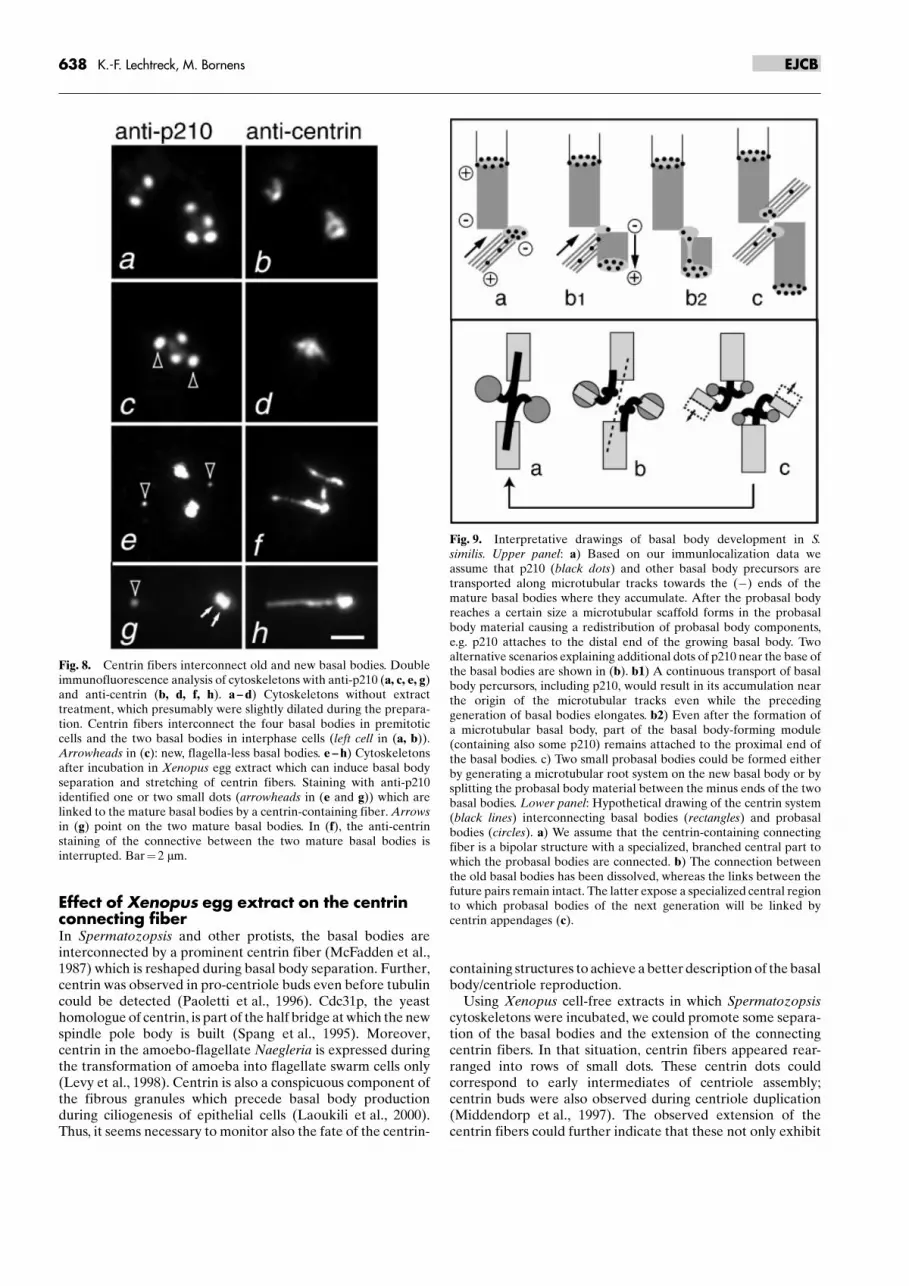
egg extracts driven to interphase with calcium, rarely showedseparated basal bodies, but a division of the centrin fiber andoccasionally additional centrin dots were observed (notshown). This reorganization could be inhibited by the additionof the non-hydrolysable ATP analogue AMP-PNP to theextract or pre-incubation of the cytoskeletons with anti-centrin(not shown). Centrin fibers also link probasal bodies of S. similisto the basal apparatus (Fig. 8 a ± d). After incubation in eggextract probasal bodies or basal body precursors were oftenseparated from the mature basal bodies but remained linked tothose by centrin fibers. Figure 8e ± h show that even small basalbody precursors are linked to the mature basal bodies bycentrin-containing fibers. The distance between the small p210dots and the basal bodies was up to several mm indicating thatthe centrin fibers are highly stretchable.
Discussion
Basal body reproduction has been reinvestigated in the greenalga S. similis by monitoring the fate of a marker having aspecific location in the fully developed basal body, namely a
Fig. 5. Effect of Xenopus egg extract on Spermatozopsis cytoskele-tons. Cytoskeletons stained with anti-p210 (a ± c) and the correspondingsignals with anti-centrin (d, e) or GT335 (f), respectively. a, d)Cytoskeletons incubated with extract buffer. b, c, e, f) Cytoskeletons
after incubation in Xenopus egg extract simulating interphase, whichresulted in the formation of separated basal body pairs (b) or singlebasal bodies (c). Bar� 10 mm.
636 K.-F. Lechtreck, M. Bornens EJCB
distal location, and by taking advantage of a cell-free assay tobetter observe the centrin fibers connecting basal bodies as theymight play a critical role in the early steps of pro-centriole/basalbody assembly.
When and where is p210 first occurring duringbasal body development?In order to refine the description of the complete developmentof the basal flagellar apparatus, it might be rewarding tomonitor the fate of basal body components which can beprecisely located in the final structure, even if their functionsare not yet fully characterized. p210, first identified as acomponent of the flagellar transition region (Lechtreck et al.,1999), and further observed at the tip of growing basal bodieswell before the transition region assembles, accumulates alsoprior to mitosis close to the proximal end of the mature basalbodies. Interestingly, a new set of small p210 dots at theproximal end of basal bodies was again observed after basalbody duplication. Thus, they could be continuous with thoseobserved prior to mitosis which develop into the next genera-tion of basal bodies.
Fig. 6. Statistical analysis of the effect of Xenopus egg extract onisolated cytoskeletons. A) Table showing the amount (in %) of paired,separated and single basal bodies in selected experiments. In experi-ment 4, cytoskeletons were incubated with monoclonal anti-centrin(�6.8) prior to incubation in interphasic or mitotic extracts, whichlargely inhibited separation. B) Histogram showing the distribution (in%) of distances between the p210 signals (center to center) ofcytoskeletons treated with extract (solid bars, average distance1.6 mm�0.5 mm, n� 63) or buffer controls (open bars, average distance1.1 mm� 0.2 mm, n� 62).
Fig. 7. Details of cytoskeletons treated with Xenopus egg extract. A)Images of separated basal body pairs from cytoskeletons incubatedwith interphasic (a ± d) or mitotic (e ± h) extracts. The centrin-basedcytoskeletons consist of up to six dots often arranged in two, roughlysymmetrical groups spaced by a central gap (arrowheads). Bar� 2 mm.B) a ± c) Black-and-white overlay images of cytoskeletons preincubatedwith monoclonal anti-centrin before the treatment with extract. Whenseparation occurred, most of the centrin (black) remained attached toone of the basal bodies in each pair (as visualized by anti-p210 stainingand displayed in white). For comparison, a basal body pair separatedafter extract treatment is shown in (d). Note that the outermost centrindots overlap with the p210 signals (as it is the case for those displayed inpart A of the figure) which was not noted in cytoskeletons preincubatedwith anti-centrin. A cytoskeleton stained with the same antibodies butnot treated with extract is shown in Fig. 8a.
637Basal body replication in green algaeEJCB
Effect of Xenopus egg extract on the centrinconnecting fiberIn Spermatozopsis and other protists, the basal bodies areinterconnected by a prominent centrin fiber (McFadden et al.,1987) which is reshaped during basal body separation. Further,centrin was observed in pro-centriole buds even before tubulincould be detected (Paoletti et al., 1996). Cdc31p, the yeasthomologue of centrin, is part of the half bridge at which the newspindle pole body is built (Spang et al., 1995). Moreover,centrin in the amoebo-flagellate Naegleria is expressed duringthe transformation of amoeba into flagellate swarm cells only(Levy et al., 1998). Centrin is also a conspicuous component ofthe fibrous granules which precede basal body productionduring ciliogenesis of epithelial cells (Laoukili et al., 2000).Thus, it seems necessary to monitor also the fate of the centrin-
containing structures to achieve a better description of the basalbody/centriole reproduction.
Using Xenopus cell-free extracts in which Spermatozopsiscytoskeletons were incubated, we could promote some separa-tion of the basal bodies and the extension of the connectingcentrin fibers. In that situation, centrin fibers appeared rear-ranged into rows of small dots. These centrin dots couldcorrespond to early intermediates of centriole assembly;centrin buds were also observed during centriole duplication(Middendorp et al., 1997). The observed extension of thecentrin fibers could further indicate that these not only exhibit
Fig. 8. Centrin fibers interconnect old and new basal bodies. Doubleimmunofluorescence analysis of cytoskeletons with anti-p210 (a, c, e, g)and anti-centrin (b, d, f, h). a ± d) Cytoskeletons without extracttreatment, which presumably were slightly dilated during the prepara-tion. Centrin fibers interconnect the four basal bodies in premitoticcells and the two basal bodies in interphase cells (left cell in (a, b)).Arrowheads in (c): new, flagella-less basal bodies. e ± h) Cytoskeletonsafter incubation in Xenopus egg extract which can induce basal bodyseparation and stretching of centrin fibers. Staining with anti-p210identified one or two small dots (arrowheads in (e and g)) which arelinked to the mature basal bodies by a centrin-containing fiber. Arrowsin (g) point on the two mature basal bodies. In (f), the anti-centrinstaining of the connective between the two mature basal bodies isinterrupted. Bar� 2 mm.
Fig. 9. Interpretative drawings of basal body development in S.similis. Upper panel: a) Based on our immunlocalization data weassume that p210 (black dots) and other basal body precursors aretransported along microtubular tracks towards the (ÿ) ends of themature basal bodies where they accumulate. After the probasal bodyreaches a certain size a microtubular scaffold forms in the probasalbody material causing a redistribution of probasal body components,e.g. p210 attaches to the distal end of the growing basal body. Twoalternative scenarios explaining additional dots of p210 near the base ofthe basal bodies are shown in (b). b1) A continuous transport of basalbody percursors, including p210, would result in its accumulation nearthe origin of the microtubular tracks even while the precedinggeneration of basal bodies elongates. b2) Even after the formation ofa microtubular basal body, part of the basal body-forming module(containing also some p210) remains attached to the proximal end ofthe basal bodies. c) Two small probasal bodies could be formed eitherby generating a microtubular root system on the new basal body or bysplitting the probasal body material between the minus ends of the twobasal bodies. Lower panel: Hypothetical drawing of the centrin system(black lines) interconnecting basal bodies (rectangles) and probasalbodies (circles). a) We assume that the centrin-containing connectingfiber is a bipolar structure with a specialized, branched central part towhich the probasal bodies are connected. b) The connection betweenthe old basal bodies has been dissolved, whereas the links between thefuture pairs remain intact. The latter expose a specialized central regionto which probasal bodies of the next generation will be linked bycentrin appendages (c).
638 K.-F. Lechtreck, M. Bornens EJCB

calcium-dependent contractility but also have elastic proper-ties. The latter was already noted for the stalk of Vorticellawhich consists predominately of the centrin-related proteinspasmin (Weis-Fogh and Amos, 1972; Maciejewski et al., 1999;Adams and Kilmartin, 2000). After incubation in extract, weoften noted a partial overlap of the p210 dots with the centrinstructures in indirect immunofluorescence (see Fig. 7), whichmight indicate that centrin was recruited into the basal body/centrioles where it is also located in animal cells (Paoletti et al.,1996). Alternatively, the incubation in extract could partiallydisintegrate the basal bodies allowing a better access of anti-centrin to the stellate structure. Further, we noted that basalbodies often remained associated even when a gap in thecentrin structure connecting them was observed. Thus, weassume that non-centrin fibers contribute to the basal bodylinker. The phenotype of vfl2 mutation in the Chlamydomonascentrin gene is characterized by errors in basal body segrega-tion (Taillon et al. 1992).
Strikingly, during the incubation in the extract, small p210dots were separated from the mature basal bodies but remainedlinked to them by extended centrin fibers. Our observationswould fit the idea that centrin filaments could provide anchor-ing sites for the basal body precursors and somehow determinethe number and place of basal bodies in the next generation (see(Adams and Kilmartin, 2000)).
How are the number and position of new basalbodies controlled?The complete reproduction of the flagellar apparatus can becompared to an assembly line in which several requirementsconverge to set the correct sequence of events which eventuallylead to the final product. We do not know, however, whether allsteps in the assembly line are carried out at the same pace: arethe building blocks for basal body assembly synthesized andbrought into the line in a rate-limiting manner, or are some ofthem stockpiled ahead of their use, and others are rate limiting?Depending on the answer to this question for any given antigenunder study, one would interpret its staining pattern in one wayor another. Are some mechanisms continuous during all phasesof the cell cycle progression, and are others discrete, triggeredby cell cycle regulators? Are some components participating inthe assembly process as transient scaffolds which otherwise donot participate in the final product? These questions will haveto be answered before one arrives at a comprehensive descrip-tion of basal body reproduction.
We have interpreted our data on p210 into two alternativemodels, depending on the assumptions one could make on thebehavior of p210. But in both cases, the basic rules would beprovided by two structures that we consider functionallyessential elements for the whole assembly process, namely thecentrin fibers (see above), and the microtubules themselves.
One can safely state that the basic property of microtubulesshould play a critical role in the whole assembly process. Aspolar structures they are able to govern motor-driven trans-portation of molecules, protein complexes or vesicles towardsone end or the other, and thus are able to concentrate locallyplus- or minus end-binding proteins. This would hold for themicrotubule subsets of the centriole/basal body ((Bobinnecet al., 1998) and for further discussion (Tassin and Bornens,1999)) and for other subsets of stable microtubules associatedwith the basal bodies. In that respect, the green algae, becauseof the stereotyped organization of microtubular roots associ-ated with the flagellar apparatus, may allow us to observe
important features that might be more difficult to observeduring centrosome duplication in vertebrate cells. We assumethe existence of a basal body-producing module, containing asubset of proteins which, in the case of S. similis, also containsp210. The number of different proteins in such a module is noteasy to predict: if one considers that the nucleation of a singlemicrotubule is catalyzed by the gTuRC, a complex of sixdifferent polypeptides (Zheng et al., 1995), one can assume thatmany more proteins are involved in the assembly of basalbodies and centrioles. Among those are presumably MAPs,minus- and plus end microtubule-binding proteins, chaperones.
We observed p210 on the five-stranded microtubular roots ofS. similis, indicating a transport of basal body precursors alongthese tracks. Recently, it was shown that pericentrin and g-tubulin are transported to the centrosome in a microtubule-dependent manner (Young et al., 2000). A minus end-directedtransport of basal body precursor material along certainmicrotubular bundles would cause its accumulation, togetherwith other components of the precursor module, near the (ÿ)ends of the microtubule bundles, in the vicinity of the proximalend of basal bodies, i.e. the (ÿ) ends of the microtubule triplets(Fig. 9, upper panel, a). During this phase, p210, which in thefirst model (Fig. 9 b1) is assumed to be a strict distal componentof basal bodies, would be stuck there as long as the formationand elongation of the microtubular wall of the new basal bodiesis not triggered. When this happens, upon an independent signal(for example a cell cycle regulator), this would induce a sortingof the proteins accumulated in the basal body precursor (plusend microtubule-binding proteins to the plus end, p210 to thetip, etc.). This could be motor driven for some proteins, or couldinvolve a mechanism similar to what has been recentlysubstantiated for proteins like EB1, or CLIP170 which associ-ate only to the tips of growing MTs and dissociate when theyshrink (Perez et al., 1999; Mimori-Kiyosue et al., 2000). In thismodel, the formation of new p210-containing structures nearthe base of the old and the young basal bodies would simplyreflect the continuous supply of p210 along the 5-strandedmicrotubular roots. A prediction would be that the firstoccurrence of p210 is observed only after the new roots areset in place (Fig. 9c).
In the alternative model (Fig. 9, upper panel, b2), the newp210 dots would originate by a split of the p210-containingmaterial during basal body elongation resulting in a largeportion at the distal end and a smaller portion close to the baseof the new basal body or between the new and the parental basalbody. The sites of future basal body assembly would remainimprinted by some permanent scaffolding proteins near thebase of existing basal bodies. In the amoebo-flagellate Para-urostyla, a transient fibrous system was observed whichprecedes basal body assembly and forms tracks along whichbasal bodies will be later formed (Fleury et al., 1993). Aminiature version of such a scaffolding system, which wouldallow the formation of only one new basal body, could beattached to each parental basal body. An additional mechanismwould be necessary to ensure a distribution of the scaffoldingmaterial between the mother and daughter basal body whichcould be driven e.g. by an affinity of the precursor material forthe (ÿ) end of the triplets (see also legend of Fig. 9). In that caseone must predict that the additional dots of p210 would beobserved as soon as new basal bodies are formed.
639Basal body replication in green algaeEJCB

How are the centrin fibers re-organized duringbasal body development?The interplay between the two types of microtubules (basalbodies and rootlets) and the centrin fibers is likely to be a keyissue, in order to keep spatial landmarks with one system whenthe other is remodeled or assembled. Our experiments showeda stable connection between the centrin fibers and theadditional small p210 dots, even when the latter were separatedfrom the basal bodies. The molecular structure and polarity ofcentrin-containing fibers is not known, but in the case of thenucleus-basal body connector an ultrastructural polarity isapparent (the basal body at one end, the cell nucleus at theother). In contrast, the centrin fiber connecting the two flagella-bearing basal bodies of green flagellates is a symmetricstructure as indicated by its mirror striation pattern or by thepresence of a specialized mid region (Ringo, 1967, Geimeret al., 1997a). Thus, the connecting fiber could actually consistof two half-fibers. New basal bodies seem to be linked to theparental basal bodies via branches of the centrin-connectingfiber (see Fig. 8e) (Lechtreck and Grunow, 1999) which laterdevelop into half-fibers of the new basal body connective(Fig. 9, lower panel). Such a scenario where the attachment siteof the new basal body is located on a centrin-containingstructure originating from the parental basal body resemblesthe formation of a new spindle pole body (SPB) at the end of thehalf-bridge. The major difference in this analogy is that newbasal bodies form already before their parents are separatedduring mitosis and cell division (three generations of centrioles/basal bodies in one cell), whereas SPB development is delayedby one cell cycle (only two generations of SPB in one cell).
Basal body assembly ± a self-sustainedprocess ?In summary, we obviously need to have a refined description ofthe behavior of many more components besides p210 during thebasal body apparatus development. We also need to know theprecise and comparative timing of the assembly of each type ofmicrotubular roots and of centrin fibers with respect to that ofthe basal bodies. Our present data suggest that basal bodydevelopment could be a self-sustained process, in which eachgeneration of basal bodies induces the formation of a structurewhich subsequently develops into the succeeding generation.By forming new microtubular flagellar roots, the new basalbodies may reset the supply in building blocks for the next basalbody generation. Alternatively, the growing basal body mayseed a docking site. The asynchrony between different assemblycycles (basal bodies assembly, microtubular roots and centrinfibers assembly) might be the key of the whole process: if thesecycles were set to overlap, this would ensure that both, spatialand temporal clues are preserved.
Acknowledgement. We gratefully acknowledge Thomas Kunzinger forprofound advice on the preparation of Xenopus egg extracts, MathieuPiel for his help with the recording of fluorescence images, and BernardEdde for kindly providing the GT335 antibody. This work was supportedby the DFG (grant number Le 806/3 ± 2 and 4 ± 1).
References
Adams, I. R., Kilmartin, J. V. (2000): Spindle pole duplication: a modelfor centrosome duplication? Trends Cell Biol. 10, 329 ± 335.
Albrecht-Buehler, G. (1990): The iris diaphragm model of centriole andbasal body formation. Cell Motil. Cytoskeleton 17, 197 ± 213.
Bailly, E., Doree, M., Nurse, P., Bornens, M. (1989): p34cdc2 is located inboth nucleus and cytoplasm; part is centrosomally associated at G2/Mand enters vesicles at anaphase. EMBO J. 8, 3985 ± 3995.
Bailly, E., Pines, J., Hunter, T., Bornens, M. (1992): Cytoplasmicaccumulation of cyclin B1 in human cells: association with adetergent-resistant compartment and with the centrosome. J. CellSci. 101, 529 ± 545.
Bobinnec, Y., Khodjakov, A., Mir, L. M., Rieder, C. L., Edde, B.,Bornens, M. (1998): Centriole disassembly in vivo and its effect oncentrosome structure and function in vertebrate cells. J. Cell Biol. 143,1575 ± 1589.
Dirksen, E. R. (1991): Centriole and basal body formation duringciliogenesis revisited. Biol. Cell 72, 31 ± 38.
Dutcher, S. K. (2001): The tubulin fraternity: alpha to eta. Curr. Opin.Cell Biol. 13, 49 ± 54.
Dutcher, S. K., Trabuco, E. C. (1998): The UNI3 gene is required forassembly of basal bodies of Chlamydomonas and encodes delta-tubulin, a new member of the tubulin superfamily. Mol. Biol. Cell 9,1293 ± 1308.
Fleury, A., Le Guyader, H., Iftode, F., Laurent, M., Bornens, M. (1993):A scaffold for basal body patterning revealed by a monoclonalantibody in the hypotrich ciliate Paraurostyla weissei. Dev. Biol. 157,285 ± 302.
Freed, E., Lacey, K. R., Huie, P., Lyapina, S. A., Deshaies, R. J., Stearns,T., Jackson, P. K. (1999): Components of an SCF ubiquitin ligaselocalize to the centrosome and regulate the centrosome duplicationcycle. Genes Dev. 13, 2242 ± 2257.
Garreau De Loubresse, N., Ruiz, F., Beisson, J., Klotz, C. (2001): Role ofdelta-tubulin and the C-tubule in assembly of Paramecium basalbodies. BMC Cell Biol. 2, 4.
Geimer, S., Lechtreck, K.-F., Melkonian, M. (1997a): The cytoskeletonof the naked green flagellate Spermatozopsis similis (Chlorophyta):analysis of the isolated basal apparatuses in the parallel configura-tion. J. Phycol. 33, 241 ± 253.
Geimer, S., Teltenkötter, A., Plessmann, U., Weber, K., Lechtreck, K.-F.(1997b): Purification and characterization of basal apparatuses froma flagellate green alga. Cell Motil. Cytoskeleton 37, 72 ± 85.
Goodenough, U. W., St Clair, H. S. (1975): BALD-2: a mutationaffecting the formation of doublet and triplet sets of microtubulesin Chlamydomonas reinhardtii. J. Cell Biol. 66, 480 ± 491.
Hinchcliffe, E. H., Li, C., Thompson, E. A., Maller, J. L., Sluder, G.(1999): Requirement of Cdk2-cyclin E activity for repeated centro-some reproduction in Xenopus egg extracts. Science 283, 851 ± 854.
Kochanski, R. S., Borisy, G. G. (1990): Mode of centriole duplicationand distribution. J. Cell Biol. 110, 1599 ± 1605.
Lacey, K. R., Jackson, P. K., Stearns, T. (1999): Cyclin-dependent kinasecontrol of centrosome duplication. Proc. Natl. Acad. Sci. USA 96,2817 ± 2822.
Lange, B. M. H., Gull, K. (1996): Structure and function of the centriolein animal cells: progress and questions. Trends Cell Biol. 6, 348 ± 352.
Laoukili, J., Perret, E., Middendorp, S., Houcine, O., Guennou, C.,Marano, F., Bornens, M, Tournier, F. (2000): Differential expressionand cellular distribution of centrin isoforms during human ciliatedcell differentiation in vitro. J. Cell Sci. 113, 1355 ± 1364.
Lechtreck, K.-F., Geimer, S. (2000): Distribution of polyglutamylatedtubulin in the flagellar apparatus of green flagellates. Cell Motil.Cytoskeleton 47, 219 ± 235.
Lechtreck, K.-F., Grunow, A. (1999): Evidence for a direct role ofnascent basal bodies during spindle pole initiation in the green algaSpermatozopsis similis. Protist 150, 163 ± 181.
Lechtreck, K.-F., Teltenkötter, A., Grunow, A. (1999): A 210 kDaprotein is located in a membrane-microtubule linker at the distal endof mature and nascent basal bodies. J. Cell Sci. 112, 1633 ± 1644.
Levy, Y. Y., Lai, E. Y., Remillard, S. P., Fulton, C. (1998): Centrin issynthesized and assembled into basal bodies during Naegleriadifferentiation. Cell Motil. Cytoskeleton 40, 249 ± 260.
Maciejewski, J. J., Vacchiano, E. J., McCutcheon, S. M., Buhse, H. E. Jr.(1999): Cloning and expression of a cDNA encoding a Vorticella
640 K.-F. Lechtreck, M. Bornens EJCB

convallaria spasmin: an EF-hand calcium-binding protein. J. Eukar-yot. Microbiol. 46, 165 ± 173.
Marshall, W. F., Vucica, Y., Rosenbaum, J. L. (2001): Kinetics andregulation of the de novo centriole assembly: implications for themechanism of centriole duplication. Curr. Biol. 11, 308 ± 317.
Matioli, G. T. (1999): Centriole duplication and orientation in mamma-lian cells. Med. Hypotheses 52, 13 ± 14.
McFadden, G. I., Melkonian, M. (1986): Use of Hepes buffer formicroalgal culture media and fixation for electron microscopy.Phycologia 25, 551 ± 557.
McFadden, G. I., Schulze, D., Surek, B., Salisbury, J. L., Melkonian, M.(1987): Basal body reorientation mediated by a Ca2�-modulatedcontractile protein. J. Cell Biol. 105, 903 ± 912.
Melkonian, M., Beech, P. L., Katsaros, C., Schulze, D. (1991): Centrin-mediated cell motility in algae. In: Melkonian M (ed.): Algal cellmotility. Chapman and Hall, New York, London, pp. 179 ± 221.
Meraldi, P., Lukas, J., Fry, A.M., Bartek, J., Nigg, E. A. (1999):Centrosome duplication in mammalian somatic cells requires E2Fand Cdk2-cyclin A. Nature Cell Biol. 1, 88 ± 93.
Middendorp, S., Paoletti, A., Schiebel, E., Bornens, M. (1997):Identification of a new mammalian centrin gene, more closelyrelated to Saccharomyces cerevisiae CDC31 gene. Proc. Natl. Acad.Sci. USA 94, 9141 ± 9146.
Mignot, J.-P. (1996): Nouvelles hypotheÁses sur la reÂplication descentrioles et des corps basaux. CR Acad. Sci. 319, 1093 ± 1099.
Mimori-Kiyosue, Y., Shiina, N., Tsukita, S. (2000): The dynamicbehavior of the APC-binding protein EB1 on the distal ends ofmicrotubules. Curr. Biol. 10, 865 ± 868.
Moudjou, M., Bordes, N., Paintrand, M., Bornens, M. (1996): Gamma-tubulin in mammalian cells: the centrosomal and the cytosolic forms.J. Cell Sci. 109, 875 ± 887.
Murray, A. W. (1991): Cell cycle extracts. Methods Cell Biol. 36, 581 ±605.
Paoletti, A., Moudjou, M., Paintrand, M., Salisbury, J. L., Bornens, M.(1996): Most of centrin in animal cells is not centrosome-associatedand centrosomal centrin is confined to the distal lumen of centrioles.J, Cell Sci. 109, 3089 ± 3102
Perez, F., Diamantopoulos, G. S., Stalder, R., Kreis, T. E. (1999): CLIP-170 highlights growing microtubule ends in vivo. Cell 96, 517 ± 527.
Ringo D. L. (1967): Flagellar motion and fine structure of the flagellarapparatus in Chlamydomonas. J. Cell Biol. 33, 543 ± 571.
Ruiz, F., Beisson, J., Rossier, J., Dupuis-Williams, P. (1999): Basal bodyduplication in Paramecium requires gamma-tubulin. Curr. Biol. 14,43 ± 46.
Schiebel, E. (2000): Gamma-tubulin complexes: binding to the centro-some, regulation and microtubule nucleation. Curr. Opin. Cell Biol.12, 113 ± 118.
Spang, A., Courtney, I., Grein, K., Matzner, M., Schiebel, E. (1995): TheCdc31p ± binding protein Kar1p is a component of the half bridge ofthe yeast spindle pole body. J. Cell Biol. 128, 863 ± 877.
Taillon, B. E., Adler, S. A., Suhan, J. P., Jarvik, J. W. (1992): Mutationalanalysis of centrin: an EF-hand protein associated with three distinctcontractile fibers in the basal body apparatus of Chlamydomonas. J.Cell Biol. 119, 1613 ± 1624.
Tassin, A. M., Bornens, M. (1999): Centrosome structure and micro-tubule nucleation in animal cells. Biol. Cell 91, 343 ± 354.
Tournier, F., Cyrklaff, M., Karsenti, E., Bornens, M. (1991): Centro-somes competent for parthenogenesis in Xenopus eggs supportprocentriole budding in cell-free extracts. Proc. Natl. Acad. Sci. USA88, 9929 ± 9933.
Vidwans, S. J., Wong, M. L., O�Farrell, P. H. (1999): Mitotic regulatorsgovern progress through steps in the centrosome duplication cycle. J.Cell Biol. 147, 1371 ± 1378.
Weis-Fogh, T., Amos, W. B. (1972): Evidence for a new mechanism ofcell motility. Nature 236, 301 ± 304.
Winey, M. (1996): Genome stability. Keeping the centrosome cycle ontrack. Curr. Biol. 6, 962 ± 964.
Wolff, A., de NeÂchaud, B., Chillet, D., Mazurguil, H., DesbruyeÁres, EÂ .,Auderbert, S., EddeÂ, B., Gros, F., Denoulet, P. (1992): Distribution ofglutamylated alpha- and beta-tubulin in mouse tissue using a specificmonoclonal antibody, GT335. Eur. J. Cell Biol. 59, 425 ± 432.
Young, A., Dictenberg, J. B., Purohit, A., Tuft, R., Doxsey, S. J. (2000):Cytoplasmic dynein-mediated assembly of pericentrin and gamma-tubulin onto centrosomes. Mol. Biol. Cell 11, 2047 ± 2056.
Zheng, Y., Wong, M. L., Alberts, B., Mitchison, T. (1995): Nucleation ofmicrotubule assembly by a gamma-tubulin-containing ring complex.Nature 378, 578 ± 583.
641Basal body replication in green algaeEJCB