Embed Size (px)
Citation preview

7An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
Bases anatómicas del sueñoAnatomical basis of sleep
J. L. Velayos, F. J. Moleres, A. M. Irujo, D. Yllanes, B. Paternain
RESUMENEl sueño es un estado biológico activo, periódico,
en el que se distinguen las etapas NREM y REM, que sealternan sucesivamente durante la noche. Intervienenlos relojes biológicos en la modulación del sistema, asícomo neurotransmisores específicos. Se trata de unared neuronal compleja, en la que intervienen diversaszonas del sistema nervioso central. Los procesos oníri-cos están controlados además de forma neural.
Se resume la historia de las investigaciones sobreel tema, desde el siglo XIX hasta nuestra época. Hayque destacar los recientes descubrimientos de Lugare-si y su equipo, que, al describir el insomnio familiargrave, dieron importancia al núcleo dorsomedial deltálamo en la instauración de la fase de sueño profundo.Al grupo de Reinoso se debe el hallazgo de que el“director de orquesta” en la instauración del sueñoREM es la zona ventral paramediana del núcleo reticu-lar pontino oral
Palabras clave. Sueño. Vigilia. Centros neurales.REM. NREM.
ABSTRACTSleep is an active and periodic biological state
composed of NREM and REM phases, which alternateduring the night. Both biological clocks and specificneurotransmitters are involved in the modulation ofthis system. It is a complex neuronal network in whichseveral areas of the central nervous system areinvolved. The oneiric processes are also controlledneurally.
This work summarises the history of theinvestigations on this topic from the 19th century todate. It is worth mentioning the recent findings ofLugaresi and colleages who described fatal familialinsomnia, a disease that helped to show theimportance of the mediodorsal thalamic nucleus in thegenesis of slow-wave sleep. Reinoso´s group found outthat the paramedian ventral area of the oral pontinereticular nucleus is the conductor in the establishmentof REM sleep.
Key words. Sleep. Wakefulness. Neural centres.REM. NREM.
Correspondencia:José Luis Velayos JorgeDepartamento de AnatomíaFacultad de MedicinaUniversidad de NavarraC/ Irunlarrea, 131008 PamplonaTfno. 948 425600Fax 948 425649E-mail: [email protected]
Departamento de Anatomía. Facultad deMedicina. Universidad de Navarra. Pamplo-na
An. Sist. Sanit. Navar. 2007; 30 (Supl. 1): 7-17.

J.L. Velayos y otros
8 An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
EL SUEÑO: ASPECTOS GENERALES
Todos los animales, incluidos los uni-celulares, tienen períodos de actividad yde reposo, habiendo en este último unainmovilidad relativa. Por otra parte, segúnse avanza en la escala filogenética, el pro-ceso del sueño es cada vez más complejo.
El sueño, biológicamente, no es unafalta total de actividad, sino que se puedeconsiderar como un estado biológico con-creto, un estado conductual, según seviene diciendo desde los años ochenta delpasado siglo. En el sueño se precisa de unambiente y una postura adecuados, queson variables en distintas especies: hayanimales que pueden dormir de pie, yotros que pueden hacerlo con los ojosabiertos. En contraposición al coma, elestado de sueño es reversible en respues-ta a estímulos adecuados y genera cam-bios electroencefalográficos que lo distin-guen del estado de vigilia. La disminuciónen la motricidad de la musculatura esque-lética y en el umbral de reactividad a estí-mulos son otras dos características deeste estado.
El sueño es periódico y en generalespontáneo, y se acompaña en el hombrede una pérdida de la conciencia vigil. Sinembargo, aun cuando el hombre tengasueño, puede, voluntariamente, no dormir.
El sueño tiene distintos grados de pro-fundidad, y se presentan modificacionesfisiológicas concretas en cada una de lasetapas del mismo. Para el estudio de loscambios funcionales que se dan duranteel sueño se atiende a unas variables quese denominan indicadores del sueño: elelectroencefalograma (EEG), los movi-mientos oculares y el tono muscular (lapolisomnografía es el registro de los tresindicadores)1-3.
ETAPAS DEL SUEÑO: ASPECTOSANATÓMICOS GLOBALES
Según estos indicadores, se distinguenvarias etapas en el sueño:
La etapa I, de somnolencia o adorme-cimiento, en que tiene lugar la desapari-ción del ritmo alfa del EEG (típico delestado de vigilia), hay tono muscular y
no hay movimientos oculares o, si loshay, son muy lentos.
La etapa II - III, de sueño ligero, secaracteriza por una disminución aúnmayor del ritmo electroencefalográfico,con la aparición de los típicos husos desueño y los complejos K, fenómenos de losque es responsable el núcleo reticular deltálamo; sigue existiendo tono muscular, yno hay movimientos oculares.
La etapa IV, de sueño profundo, presen-ta un ritmo electroencefalográfico menor,no hay movimientos oculares y el tonomuscular se mantiene o puede estar muydisminuido. En la instauración de esta fasedel sueño intervienen, entre otras estruc-turas, la corteza prefrontal y el núcleo dor-somedial del tálamo. El Insomnio FamiliarGrave es una enfermedad de tipo priónicoy evolución fatal que fue descrita por pri-mera vez por Lugaresi y su equipo en losaños ochenta4, cuyo estudio permitió des-cubrir la importancia de tal estructuratalámica para la instauración del sueñolento o profundo. Es la fase del sueño másreparadora. Hay movimientos organizadosdel dorso; el individuo da vueltas en lacama, cambia de postura. Esta fase duraaproximadamente un 25% del total deltiempo del sueño.
Las etapas I a IV se denominan en suconjunto sueño no REM (NREM).
La siguiente etapa es la de sueño para-dójico, que se caracteriza por una activi-dad EEG que recuerda al estado de vigilia(por eso se habla de sueño paradójico),debida a una activación cortical por partede estructuras encefálicas profundas,como es la formación reticular activadora.Fue descubierto por Kleitman y Aserinsky,junto con Dement 5, en los años cincuentadel siglo pasado. Hay una desincronizacióndel EEG, que se asemeja a una situación devigilia, de alerta. Se observan movimientosoculares rápidos (también se habla desueño MOR, de movimientos oculares rápi-dos o sueño REM, de rapid eye move-ments), dependientes de la actividad deestructuras profundas tales como la for-mación reticular pontina. Se produce unaatonía (desaparición del tono muscular),de lo que son responsables estructurascomo la formación reticular bulbar, el

9An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
BASES ANATÓMICAS DEL SUEÑO
locus coeruleus, etc. El músculo diafragmasigue manteniendo el tono, y contrayéndo-se, permitiendo la respiración. La fase desueño REM constituye un 25 % del sueñototal. En el recién nacido, el sueño REMconstituye el 50% del tiempo total desueño. El tiempo de vigilia va aumentandocon la edad, cada vez se duerme menos, ycada vez hay menos sueño REM. Sólo exis-te sueño REM en los mamíferos, excepto elconejo macho, el oso hormiguero y el del-fín de nariz en botella. Parece ser, en líneasgenerales, que el sueño paradójico se pro-duce, filogenéticamente, cuando la cortezacerebral está más desarrollada. De la ins-tauración del sueño REM es responsable eltronco del encéfalo, concretamente ungrupo de neuronas que también descarganen la vigilia (el centro nodal es el núcleoreticular pontino oral, cuyas porcionesventral y paramediana reciben conexionesde múltiples estructuras relacionadas conel control del ciclo vigilia-sueño), produ-ciendo una activación de los sistemas coli-nérgicos.
Las fases de sueño NREM y REM sealternan sucesivamente, cuatro a cincoveces por la noche. En total, la fase desueño NREM dura unas 6 horas; y la fasede sueño REM, dos horas, por términomedio. Es más fácil despertar al sujeto enla fase de sueño REM que en la fase NREM.Los medicamentos antidepresivos reducenel sueño REM y las benzodiacepinas acor-tan o suprimen las fases III y IV.
El metabolismo cerebral y en conse-cuencia la temperatura cerebral disminuyecon la profundidad del sueño NREM. Sinembargo, en el sueño REM pueden incre-mentarse estas cifras con respecto al esta-do de vigilia, ya que hay una activación dela corteza cerebral.
En el sueño NREM se da una progresivadesactivación de la formación reticularactivadora junto a una inhibición de lasneuronas relé talámicas. Todo esto quieredecir que las estimulaciones sensorialeshan de tener un umbral determinado paraprovocar el despertar, ya que en el sueño,y sobre todo en la fase de sueño profundo,no hay una concienciación de lo sensorial.Sin embargo, es curioso que se pueda darun despertar ante situaciones determina-
das: la madre se despierta cuando su bebénecesita algo, aun cuando no le despiertenotros estímulos más potentes; algunas per-sonas pueden despertarse a una hora pre-determinada, con gran precisión. Deben dejugar un papel en este sentido sistemas deinhibiciones que desconocemos aún.
RELOJES BIOLÓGICOSEl sueño es por lo tanto un estado
dinámico, en el que se activan e inhibendistintas zonas del encéfalo, con las reper-cusiones funcionales que se han indicado.Obedece a un ritmo biológico, circadiano(cada 24 horas), relacionado con el ritmodía-noche, nictemeral, al que se ajusta elproceso. De por sí, el ritmo sueño-vigilia escada 25-29 horas, según se ha estudiadoexperimentalmente en voluntarios ence-rrados en una habitación a la que no lleganlas influencias exteriores. Pero la presióndel sueño aumenta en torno a las dos de latarde, lo que explica que sea fisiológicosentir sueño después de comer. La volun-tad puede evitar el dormir después decomer.
Existen unos relojes biológicos en elsistema nervioso central. Uno de ellos,situado en el hipotálamo (núcleo supra-quiasmático), establece el ritmo sobre losotros relojes biológicos, situados caudal-mente, y hace que el sueño NREM y elsueño REM duren un tiempo fijado. Inter-vienen en su regulación no sólo, y de modofundamental, los impulsos retinianos, sinotambién otras influencias, como es el pHde la sangre o la glucemia. El núcleo supra-quiasmático no es responsable en símismo del ritmo vigilia - sueño, pero síforma parte de las redes neurales implica-das en el proceso, redes neurales sobre lasque deben de actuar diversos sistemaspara hacer que el proceso quede anuladopor un tiempo, y al fin y al cabo retrasado.Las conexiones eferentes del núcleo supra-quiasmático son muy profusas, e inclusobilaterales: hacia el hipotálamo posterior,región preóptica, núcleos del septo, núcleoparatenial, núcleo paraventircular del tála-mo, núcleo ventral lateral del tálamo yglándula pineal 6-8.
El proceso del ciclo vigila-sueño estáregulado por una red neuronal compleja en

J.L. Velayos y otros
10 An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
la que intervienen diversas zonas del siste-ma nervioso central, a base de activacionesy de inhibiciones, cuyo resultado es la vigilao el sueño. Dentro del sueño, la fase desueño REM es regulada por una complicadared neural en la que intervienen diversosneurotransmisores. Una explicación simpli-ficada no es posible (Tabla 1, Fig. 1).
Se trata de un sistema homeostático enel que el sueño llama a la vigilia y la vigilaal sueño, sistema que voluntariamentepuede alterarse: un joven californiano de23 años estuvo 288 horas sin dormir. Laprivación del sueño lleva a la muerte,sobre todo debido a la falta de la fase pro-funda del sueño. La privación del sueño
Tabla 1. Resumen de los mecanismos celulares implicados en el sueño y la vigilia.
Núcleos del tronco del encéfalo Neurotransmisor involucrado Estado de actividad de las neuronas responsables correspondientes del tronco
del encéfalo
VigiliaNúcleos colinérgicos de la unión Acetilcolina Activasponto-mesencefálicaLocus coeruleus Noradrenalina ActivasNúcleos del rafe Serotonina Activas
Sueño no-REMNúcleos colinérgicos de la unión Acetilcolina Inactivasponto-mesencefálicaLocus coeruleus Noradrenalina InactivasNúcleos del rafe Serotonina Inactivas
Sueño REM CONECTADONúcleos colinérgicos de la unión Acetilcolina Activas (ondas ponto-encefálica pontogeniculooccipitales)Núcleos del rafe Serotonina Inactivas
Sueño REM desconectadoLocus coeruleus Noradrenalina Activas
Corteza cerebral(fuente de las ondasEEG oscilatorias)
El circuitotalamocorticalgenera oscilaciones
Tálamo
Modulaciónde la actividadtalámica
Núcleos del TE:ColinérgicosSerotoninérgicosNoradrenérgicos
Modulaciónde la actividadcortical
El control del sueño y la vigiliadepende de la modulación deltálamo y de la corteza porel tronco del encéfalo (TE).
Los principales componentes delsistema modulador son losnúcleos colinérgicos,noradrenérgicos ysetoninérgicos.
Estas estructuras tienenproyecciones tanto a la cortezacomo al tálamo, donde tienenefectos directos e indirectos sobrela función cortical.
Corteza cerebral(fuente de las ondasEEG oscilatorias)
Figura 1. Circuitos implicados en la regulación del ritmo vigilia-sueño

11An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
BASES ANATÓMICAS DEL SUEÑO
REM da lugar a alteraciones hipotalámicas,con agresividad, aumento del peso y final-mente muerte, según se ha visto experi-mentalmente.
LOS ENSUEÑOS: BASESANATÓMICAS
Durante el sueño hay actividad mental:se trata de las experiencias oníricas, losensueños, que pueden relatarse en unaproporción del 80% al despertar al sujetodespués de pasada una fase de sueño para-dójico. Por lo tanto, el sueño no significafalta de vida ni falta de actividad mental.Los sueños de la fase REM son de tipo cine-matográfico y en color. Algunos piensanque el movimiento de los ojos en esta faseREM va en seguimiento de las imágenesvisuales del ensueño. También se sueñadurante la fase de sueño profundo, peroestos sueños son más bien de tipo abs-tracto. El procesamiento de las imágenesde los ensueños, según los hallazgos clíni-cos, se hace de forma preferente en la cor-teza occipitotemporal.
Para Hobson9-11, en el sueño REM habríauna activación del sistema reticular activa-dor del tronco del encéfalo y del prosencé-falo basal, estructuras que también estánactivas en la vigilia. Para este autor, estruc-turas como los núcleos talámicos relé, queson los que reciben la estimulación senso-rial, quedarían activados, lo que contribui-ría a la aparición de las diversas modalida-des psicológicas que aparecen en lossueños. Las estructuras límbicas, talescomo la amígdala y la corteza cingular,también estarían activadas, lo que explica-ría los fenómenos emotivos durante la fasede sueño REM. Los ganglios basales y elcerebelo, que regulan la motricidad, tam-bién activados, explicarían los movimien-tos ficticios de los sueños en la fase REM.Están activas cortezas asociativas talescomo las del lobulillo parietal inferior y dela corteza occipitotemporal, todo lo cualexplica las imágenes visuales de esta fasedel sueño. La corteza prefrontal, importan-te en los procesos mentales, está inhibida,lo cual puede explicar la falta de lógica enlos razonamientos que se experimentan enel sueño. Precisamente, como dice Reino-so12, las mismas estructuras que son nece-
sarias para la consolidación de la memo-ria, tales como el tálamo medial, la amíg-dala, el hipocampo, las estructuras parahi-pocampales, la corteza orbitofrontal ycortezas asociativas monomodales estánactivas en el sueño REM.
En el sueño NREM, en cambio, hay unainhibición de las zonas que están activa-das en el sueño REM y activación de lasinhibidas, lo que puede explicar que en talsueño NREM haya ensoñaciones más detipo abstracto que en el sueño REM. El des-pertar en esta fase da lugar a estados con-fusionales y un rápido retorno al sueño.
OTROS FENÓMENOSOcurren otros fenómenos durante el
proceso del sueño: por ejemplo, secreciónde hormonas sexuales y de la hormona decrecimiento, sobre todo en el sueño NREM,de ahí la importancia del sueño en losniños y adolescentes. La fase de sueñoREM es importante además para los proce-sos de memoria; parece ser que la consoli-dación de la memoria y la eliminación delos datos "inútiles" se hace durante la faseREM. Precisamente en los ancianos laduración del sueño REM disminuye bas-tante, en cambio en el feto, la fase REMocupa la mayor parte del tiempo de sueño.Conforme avanza la edad, el estado de vigi-lia va aumentando en duración; cada vezse duerme menos, y cada vez hay menossueño REM. Independientemente delsueño, la temperatura corporal cae en lamadrugada. En el sueño REM hay tumefac-ción del pene, con independencia del tipode ensueños.
Se puede resumir que el sueño es unestado fisiológico activo, en que participandiversas estructuras encefálicas, forman-do una red neuronal, en que se dan activa-ciones e inhibiciones complejas, con unaregulación cíclica, y sobre la que puedeactuar la voluntad, modulando y estable-ciendo determinadas actitudes, comporta-mientos. Por ejemplo, cada persona tienesu "liturgia", sus hábitos para dormir: unosnecesitan leer un poco antes de conciliarel sueño; otros han de tomar un vaso deagua; otros, precisan de una oscuridadtotal; otros, de cierto grado de luminosi-dad, etc. Los sueños son distintos de unas

J.L. Velayos y otros
12 An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
personas a otras; así por ejemplo, las per-sonas creativas tienen sueños particular-mente ricos, mientras que las más simples,los tienen más elementales. Por tanto, laimpronta personal también deja su huellaen estos procesos.
HACIENDO HISTORIA (ANATOMÍADEL SUEÑO)
Podemos decir que ya a finales delsiglo XIX Gayet, en Francia, observó lesio-nes del mesencéfalo en una autopsia de uncaso de letargia crónica. Más conocidosson los hallazgos de Von Economo13, quedescribe en la I Guerra Mundial, lesionesdel hipotálamo posterior y del mesencéfa-lo rostral en casos de encefalitis letárgica.Son clásicos los trabajos de Bremer14 en losaños treinta del siglo XX, en que seccionael sistema nervioso central del gato, entrebulbo y médula ("encephale isolé"), yobserva alternancias del sueño y la vigilia.Pero si la sección era más craneal ("cerve-au isolé"), el animal entraba en un estadode sueño total. En base a los experimentos,Bremer afirmaba que lo que provocaría elsueño sería una desaferentización del sis-tema nervioso central, una interrupción delas vías ascendentes.
A principios del siglo XX, Claparède15
sugirió que el sueño era provocado poruna intoxicación del organismo. Partida-rios de esta teoría fueron Hess y Moruzzi,así como Piéron, anteriormente a Claparè-de. Pavlov16, en 1923 aplica su teoría delreflejo condicionado a la explicación de losfenómenos del sueño.
Realmente es von Ecónomo17 quiendefiende la existencia de centros neuralesresponsables de la mecánica del sueño:habla de un centro situado en hipotálamoposterior y porción rostral del mesencéfa-lo, cuya lesión produce hipersomnolenciay estupor; y de un centro situado en hipo-tálamo anterior, para los estados de sueño.Hess18 confirmó las hipótesis de von Ecó-nomo, pero con una salvedad, realmenteera el tálamo el que contenía el verdaderocentro del sueño, ya que la estimulación desu zona anteromedial (a baja latencia) pro-ducía estado de sueño.
La invención del electroencefalogramapor parte de Berger19, inicia una nueva era
en el estudio de los mecanismos de la vigi-lia y el sueño. Gracias a estos progresospueden explicarse los trabajos de Bremer,y en este sentido, también son clásicos lostrabajos de Magoun y Rhines20 y de Moruz-zi y Magoun21, que llevan a concluir que laformación reticular caudal tendría que vercon el estado de sueño, y la craneal, con elde vigilia, correspondiéndose asimismocon datos obtenidos por Rhines yMagoun22, referidos a la influencia de laformación reticular sobre las neuronasmotoras de la médula. En la década de loscincuenta, Aserinsky y Kleitman5 y sugrupo23,24 describen la etapa del sueño REM,en la que hay una inhibición del tono mus-cular, según demostrara por esos añosJouvet, así como una “paradójica” desin-cronización electroencefalográfica. Jou-vet25 pensaba que el centro responsabledel mecanismo ejecutor del sueño REM erala porción rostral de la formación reticularpontina oral. Jouvet fue el que bautizó alsueño REM como sueño paradójico. Sinembargo, Reinoso26 ha demostrado congran precisión que el centro que regula,como “director de orquesta”, la fase desueño REM, se halla en la región ventralparamediana de la formación reticularpontina oral, zona por otra parte impor-tante en la regulación de movimientos ocu-lares complejos.
Es en los años 80 del pasado siglo cuan-do se empieza sospechar de la importan-cia del tálamo en los fenómenos del sueño.En este sentido, son cruciales los trabajosde Villablanca y col27 que observan cómoel gato diencefálico (en que la cortezacerebral era ablacionada) presentaba lamisma patología que el gato atalámico. Porello, Villablanca pensó en la importanciade los circuitos corticotalámicos en laregulación del sueño. Por esa época, Ste-riade28 demuestra la importancia delnúcleo reticular del tálamo en la genera-ción de espigas en el sueño.
El insomnio familiar grave es una enfer-medad de tipo priónico descrita por Luga-resi en los años ochenta del pasadosiglo4,29,30. En esta entidad, se da una impo-sibilidad para iniciarse el período desueño lento, junto con alteraciones en elsueño REM. En el inicio se afectan losnúcleos dorsomedial y anteriores del tála-

13An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
BASES ANATÓMICAS DEL SUEÑO
mo y en sus periodos finales, se afecta lacorteza cerebral, y especialmente las cor-tezas cingular y prefrontal. En cambio, enla enfermedad de Creutzfeldt-Jakob, tam-bién de tipo priónico y descrita a princi-pios del siglo XX, las lesiones comienzanen corteza, y termina por afectarse el tála-mo. Gracias a Lugaresi y su equipo, pode-mos hablar de la importancia del núcleodorsomedial del tálamo en la instauracióndel sueño profundo.
En lo que a nosotros respecta (Figs. 2, 3y 4), hemos visto que los citados núcleostalámicos dorsomedial y anteriores tienenconexiones específicas, que pueden expli-car en parte la fisiopatología del insomniofamiliar grave31-33.
Gracias a los estudios referidos y aotros, cuya larga lista sería prolijo relatar,hoy día se sabe que en la regulación delciclo vigilia-sueño intervienen diversasestructuras del sistema nervioso central,actuando como una compleja red neural,cuyo fisiologismo íntimo está sin embargotodavía por aclarar. Decía Cajal34: "Pasaránsiglos y acaso millares de años antes que elhombre pueda entrever algo del insonda-ble arcano del mecanismo no sólo de nues-
tra psicología, sino de la más sencilla, deun insecto". Cajal tiene un trabajo relacio-nado con el tema que nos ocupa, titulado"Las teorías sobre el ensueño", de 190835.En el trabajo, cita a Duval, que en 1895 diceque en el sueño, y en base a los hallazgosde Cajal, habría una retracción de las rami-ficaciones nerviosas, interrumpiendo así lacorriente, de lo cual no es partidario Cajal,pues, según él, las arborizaciones termina-les tienen siempre la misma extensión,forma y grado de aproximación a los cuer-pos celulares, cualquiera que sea el modode muerte del animal. En cambio es parti-dario de que las células neuróglicas peri-vasculares tendrían la misión de ampliarlos espacios vasculares, para provocaralteraciones en la congestión vascular, enrelación a los procesos mentales. "Estascontracciones (de la neuroglia) puedenproducirse automáticamente, pero más amenudo son provocadas por el estímulode la voluntad, que logra de esta suerte, yobrando sobre un grupo particular decélulas neuróglicas, llevar el proceso de laasociación en direcciones determinadas."También dice que no ignora que esta hipó-tesis puede ser susceptible de ser modifi-cada36. Aunque no exactamente, sí se
Cx
RT
MD
A
BP
MB
Hpt
BS
Figura 2. Proyecciones aferentes de los núcleos dorsomedial (MD) y anteriores (A) del tálamo delgato. BP: prosencéfalo basal; BS: tronco del encéfalo; Cx: corteza cerebral; Hpt: hipotálamo;MB: cuerpo mamilar; RT: núcleo reticular del tálamo.
J.L. Velayos y otros
14 An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
puede decir que hoy día se da una impor-tancia más relevante que antes a los astro-citos, por su participación en la modula-ción de la transmisión sináptica37.Analizando concretamente los fenómenosoníricos, dice Cajal que "el ensueño perte-
nece en la inmensa mayoría de los casos ala esfera visual". "Los ensueños auditivos ytactiles son menos frecuentes, y excepcio-nales los olfativos"35. Se presentan prefe-rentemente durante la madrugada, y setrata de imágenes no sujetas a mecanis-
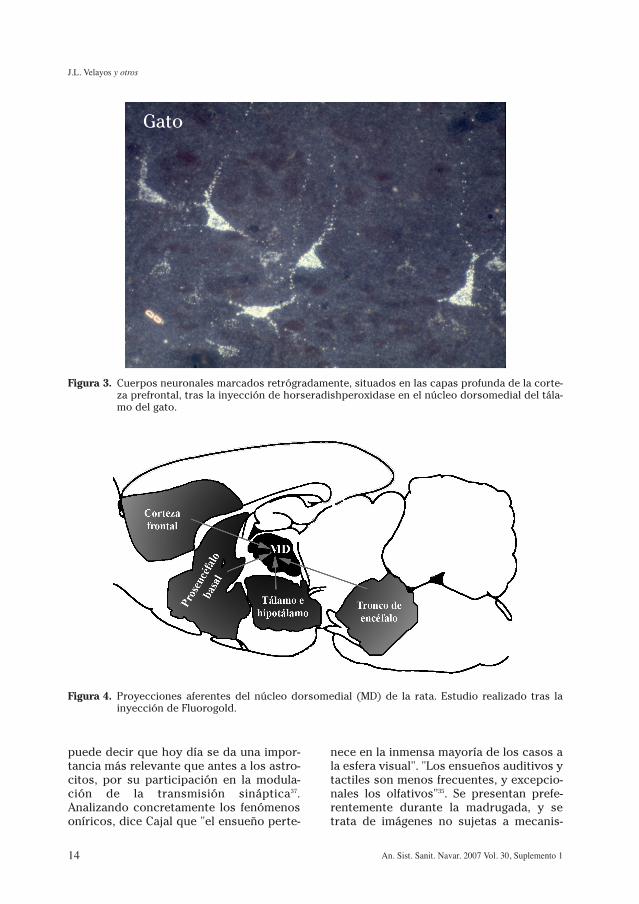
Figura 3. Cuerpos neuronales marcados retrógradamente, situados en las capas profunda de la corte-za prefrontal, tras la inyección de horseradishperoxidase en el núcleo dorsomedial del tála-mo del gato.
Figura 4. Proyecciones aferentes del núcleo dorsomedial (MD) de la rata. Estudio realizado tras lainyección de Fluorogold.
Gato

15An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
BASES ANATÓMICAS DEL SUEÑO
mos asociativos del estado vigil. "El albo-rear de la razón y del sentido crítico mar-can de ordinario el ocaso del sueño y anun-cia el próximo despertar"35. Tambiénafirma que los ensueños son ajenos a laspreocupaciones del día o a una actividadintelectual enérgica. Por introspecciónpaciente, autosugestionándose, con el finde analizar algunas facetas del sueño,observó que hay ensueños con relieve,con sensación de ejercer la convergenciaocular; que el color puede alcanzar lamisma vivacidad que la realidad exterior;también afirma que el campo de la imagendel ensueño no varía, aunque se muevanlos ojos o la cabeza. (Hoy se sabe bien quela participación de las redes neurales quecontrolan el mecanismo es esencial). Y citaCajal observaciones de otros autores y deél mismo, que relatan cómo los ciegos tar-díos sueñan con imágenes ópticas, auncareciendo de retina y nervio óptico, loque confirma una vez más que se engen-dran en el propio sistema nervioso central.
Se puede decir, de acuerdo a los hallaz-gos realizados hasta ahora, que de la vigi-lia son responsables los grupos serotonér-gicos del tronco del encéfalo (núcleos delrafe y otras neuronas serotonérgicas dis-persas)38, grupos aminérgicos tales comoel locus coeruleus, la sustancia negra y elárea tegmental ventral de Tsai, neuronashistaminérgicas y orexinérgicas del hipotá-lamo posterior (sustancias estas últimas,descubiertas en 1998)39,40, el núcleo teg-mentopedunculopontino, neuronas gluta-matérgicas del tegmento pontomesencefá-lico y del hipotálamo lateral y posterior,neuronas gabaérgicas (como moduladorasde todo el sistema)41. Todas ellas, neuronascuya actuación se realiza de forma impor-tante sobre la corteza cerebral, en sus dife-rentes capas, y de un modo específico paracada neurotransmisor.
En cuanto al sueño NREM, y en lo querespecta a la fase de somnolencia, yahemos dicho que es importante el núcleoreticular del tálamo, para la instauraciónde los husos de sueño. Según demostraranSterman y Clemente42, el prosencéfalobasal es una estructura hipnogénicaimportante en el establecimiento delsueño profundo, pero también lo es el tála-mo, y en concreto el núcleo dorsomedial,
como demostraron Lugaresi y su grupo;también lo son las conexiones corticotalá-micas, según demostrara Villablanca27. Elnúcleo reticular del tálamo, como pacema-ker diencefálico, es esencial para el esta-blecimiento de los husos de sueño28,43 .Tam-bién es importante el tronco del encéfalo44.
En cuanto al sueño REM, ya hemosdicho que es el núcleo reticular pontinooral en su zona ventral y paramediana el“director de orquesta” que regula la situa-ción26, estando implicadas otras estructu-ras, interconectadas en una compleja red.Parece ser que se atribuyó a la acetilcolinala responsabilidad de la desincronizacióndel EEG que se da en el sueño REM, ya quela inyección de carbacol en el tronco delencéfalo desencadena el sueño REM. Sinembargo, el grupo de Reinoso observó quecuando se inyectaba el carbacol en la por-ción ventral paramediana del núcleo reti-cular pontino oral, se producía el REM,pero cuando se inyectaba en el tegmentodorsal, se producía atonía, que es una delas características del REM45. También elcitado grupo ha observado cómo la supre-sión de las neuronas serotonérgicas delnúcleo dorsal del rafe y la activación de lasneuronas colinérgicas del tegmento meso-pontino, da lugar no sólo a la inhibición detodas estas estructuras, sino de otrasestructuras del tallo cerebral y del prosen-céfalo, que a su vez pueden activar o inhi-bir la mencionada zona del núcleo reticu-lar pontino oral.
Así pues, dentro de la compleja red neu-ral que regula los mecanismos del ciclo vigi-lia-sueño, son importantes la corteza cere-bral, el prosencéfalo basal, el tálamo, eltronco del encéfalo, entre otras. Se puedendistinguir dos procesos, en cierta medidaindependientes dentro del sueño: la faseNREM, con sus implicaciones hodológicaspropias, y la del REM, con otras implicacio-nes diversas; estando NREM y REM interre-lacionados de forma rítmica, independien-temente del ritmo sueño-vigilia-sueño en sí.El proceso de la vigilia presenta sus meca-nismos propios, relacionados con el proce-so del sueño, en forma homeostática y cro-nológicamente establecida.
Hoy día se da una gran importancia alas cortezas asociativas en el hombre,

J.L. Velayos y otros
16 An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
muy desarrolladas. Precisamente, unalesión de la corteza occipitotemporal dalugar a anoniria46.
BIBLIOGRAFÍA1. MCCARLEY RW. Sleep, dreams and states of
consciousness. En: P. M. Conn. Neuro-science in medicine. Philadelphia, 1995: 535-554.
2. STERIADE M, MCCORMICK, DA,SEJNOWSKI TJ.Thalamocortical oscillations in the sleepingand aroused brain. Science 1993; 262: 679-685.
3. PURVES D, AUGUSTINE GJ, FITZPATRICK, D, KATZ,LC, LAMANTIA, AS AND MCNAMARA, J. Invitacióna la neurociencia. Editorial Médica Panamer-icana. Buenos Aires, 2001.
4. LUGARESI E, MEDORI R, MONTAGNA P, BARUZZI A,CORTELLI P, LUGARESI A, et al. Fatal familialinsomnia and dysautonomia with selectivedegeneration of thalamic nuclei. N Engl JMed 1986; 315: 997-1003.
5. ASERINSKY E, KLEITMAN N. Regularly occurringperiods of eye motility, and concomitantphenomena, during sleep. Science 1953; 118:273-274.
6. MOORE RY. Neural control of the pinealgland. Behav Brain Res 1996; 73: 125-130.
7. TECLEMARIAM-MESBAH R, TER HORST GJ, POSTEMA
F, WORTEL J, BUIJS RM. Anatomical demon-stration of the suprachiasmatic nucleus-pineal pathway. J Comp Neurol 1999; 406:171-182.
8. KALSBEEK A, TECLEMARIAM-MESBAH R, PEVET P.Efferent projections of the suprachiasmaticnucleus in the golden hamster (Mesocrice-tus auratus). J Comp Neurol 1993; 332: 293-314.
9. HOBSON JA, MCCARLEY RW. The brain as adream state generator: an activation-synthe-sis hypothesis of the dream process. Am JPsychiatry 1977; 134: 1335-1348.
10. HOBSON JA. The chemistry of consciousstate. Editor. Boston, 1994.
11. HOBSON JA, STICKGOLD R. The conscious stateparadigm: a neurocognitive approach towaking, sleeping and dreaming. En: M. Gaz-zaniga. The cognitive neurosciences. Cam-bridge, Mass., 1995: 1373-1389.
12. REINOSO-SUAREZ F. Investidura como DoctorHonoris Causa. EUNSA. Madrid, 2002.
13. VON ECONOMO C, EN BRODAL, A. The reticularformation and some related nuclei. En:Oxford University Press. Neurological anato-
my in relation to clinical medicine. NewYork, 1981.
14. BREMER F. Cerveau "isolé" et physiologie dusommeil. C.R. Soc. Biol. 1935; 118: 1235-1241.
15. CLAPAREDE E. Esquisse d´une théoriebiologique du sommeil. Arch Psychol 1905;4: 246-349.
16. PAVLOV IP. "Innere Hemmung" der bedingten.Reflexe und der Schlaf - ein und derselbeProze. Skand Arch Physiol 1923; 44: 42-58.
17. VON ECONOMO C. Encephalitis lethargica.Wien Med Wischr 1923; 73: 777-782, 835-838,1113-1117, 1243-1249, 1334-1338.
18. HESS WR. Das Schlafsyndrom als Folge den-cephaler Reizung. Helv Physiol PharmacolActa 1944; 2: 305-344.
19. BERGER H. Über das Elektroenkephalogrammdes Menschen. J Psychol Neurol 1930; 40:160-179.
20. MAGOUN HW, RHINES R. An inhibitory mecha-nism in the bulbar reticular formation. Jour-nal of Neurophysiology 1946; 9: 165-171.
21. MORUZZI G, MAGOUN HW. Brain stem reticularformation and activation of the EEG. 1949. JNeuropsychiatry Clin Neurosci 1995; 7: 251-267.
22. RHINES R, MAGOUN HW. Brainstem facilitationof cortical motor responses. Journal of Neu-rophysiology 1946; 9: 219-229.
23. DEMENT W, KLEITMAN N. Cyclic variations inEEG during sleep and their relation to eyemovements, body motility, and dreaming.Electroencephalogr Clin Neurophysiol Suppl1957; 9: 673-690.
24. DEMENT W, WOLPERT EA. The relation of eyemovements, body motility, and externalstimuli to dream content. J Exp Psychol1958; 55: 543-553.
25. JOUVET M. Recherches sur les structuresnerveuses et les mécanismes responsalesdes différentes phases du sommeil physi-ologique. Arch Ital Biol 1962; 100: 125-206.
26. DE LA ROZA C, REINOSO-SUAREZ F. Ultrastruc-tural synaptic organization of axon termi-nals in the ventral part of the cat oral pon-tine reticular nucleus. J Comp Neurol 2000;427: 31-53.
27. VILLABLANCA JR, DE ANDRES I, OLMSTEAD CE.Sleep-waking states develop independentlyin the isolated forebrain and brain stem fol-lowing early postnatal midbrain transectionin cats. Neuroscience 2001; 106: 717-731.
28. STERIADE M, DOSSI RC, NUNEZ A. Network mod-ulation of a slow intrinsic oscillation of catthalamocortical neurons implicated in sleepdelta waves: cortically induced synchroniza-

17An. Sist. Sanit. Navar. 2007 Vol. 30, Suplemento 1
BASES ANATÓMICAS DEL SUEÑO
tion and brainstem cholinergic suppression.J Neurosci 1991; 11: 3200-3217.
29. LUGARESI E, MONTAGNA P. Thalamus, sleep andcircadian functions. En: M. Mancia and G.Marini. The diencephalon and sleep. NewYork, 1994: 215-220.
30. LUGARESI E, OMICINI L. El sueño, los sueños, unmundo misterioso. EUNSA. Pamplona, 1999.
31. VELAYOS JL, REINOSO-SUAREZ F. Prosencephalicafferents to the mediodorsal thalamic nucle-us. J Comp Neurol 1985; 242: 161-181.
32. VELAYOS JL, OLIVA M, ALFAGEME F. Afferent pro-jections to the mediodorsal and anteriorthalamic nuclei in the cat. Anatomical-clini-cal correlations. Brain Pathol 1998; 8: 549-552.
33. VELAYOS JL, ULLÁN J, AMAT A, RAMOS P, SIEIRA
JA. Expression of the cellular prion protein(PrPC) in the cat central nervous system.Some findings. Eur J Anat 2002; 6: 23-29.
34. RAMÓN Y CAJAL S. Recuerdos de mi vida: his-toria de mi labor científica. Alianza Editorial.Madrid, 1981.
35. RAMÓN Y CAJAL S. Las teorías sobre elensueño. Revista de Medicina y Cirugía de laFacultad de Medicina de Madrid. 1908; II: 87-98.
36. RAMÓN Y CAJAL S. Sobre las relaciones de lascélulas nerviosas con las neuróglicas.Revista trimestral micrográfica 1896; 1:123-126.
37. PEREA G, ARAQUE A. Nuevas vías de informa-ción en el sistema nervioso: comunicaciónentre astrocitos y neuronas. Rev Neurol2003; 36: 137-144.
38. RODRIGO-ANGULO ML, RODRIGUEZ-VEIGA E,REINOSO-SUAREZ F. Serotonergic connectionsto the ventral oral pontine reticular nucleus:implication in paradoxical sleep modula-tion. J Comp Neurol 2000; 418: 93-105.
39. SAKURAI T, AMEMIYA A, ISHII M, MATSUZAKI I,CHEMELLI RM, TANAKA H et al. Orexins andorexin receptors: a family of hypothalamicneuropeptides and G protein-coupled recep-tors that regulate feeding behavior. Cell1998; 92: 573-585.
40. SUTCLIFFE JG, DE LECEA L. The hypocretins:setting the arousal threshold. Nat Neurosci2002; 3: 339-349.
41. KING DP, TALAHASHI JS. Molecular mechanismsof circadian rhythms in mammals. Annu RevB Neurosci 2000; 23: 713-742.
42. STERMAN MB, CLEMENTE CD. Forebraininhibitory mechanisms: sleep patternsinduced by basal forebrain stimulation inthe behaving cat. Exp Neurol 1962; 6: 103-117.
43. SILLITO AM, JONES HE, GERSTEIN GL, WEST DC.Feature-linked synchronization of thalamicrelay cell firing induced by feedback fromthe visual cortex. Nature 1994; 369: 479-482.
44. FORCADAS MI, ZARRANZ JJ. Insomnio y alucina-ciones tras lesiones vasculares del tegmen-to protuberancial en el hombre. Neurología1994; 9: 211-223.
45. REINOSO-SUÁREZ F, DE ANDRÉS I, RODRIGO-ANGULO
M, GARZÓN M. Brain structures and mecha-nisms involved in the generation of REMslepp. Sleep Med Rev 2001; 5: 63-78.
46. POZA JJ, MARTI MASSO JF. Pérdida completa deensoñaciones tras una lesión cerebral tem-porooccipital. Neurologia 2006; 21:152-154.