Embed Size (px)
Citation preview

BEHAVIORAL ECOLOGY OF WASP-SPIDER INTERACTIONS: THE ROLE OF
WEBS, CHEMICALS, AND DECEPTION
A Dissertation
submitted to the Faculty of the
Graduate School of Arts and Sciences
of Georgetown University
in partial fulfillment of the requirements for the
degree of
Doctor of Philosophy
in Biology
By
Divya Bellur Uma, M.S.
Washington, D.C.
April 14, 2010

ii
Copyright 2010 by Divya Bellur Uma
All Rights Reserved

iv
BEHAVIORAL ECOLOGY OF WASP-SPIDER INTERACTIONS: THE ROLE
OF WEBS, CHEMICALS, AND DECEPTION
Divya Bellur Uma, M.S.
Dissertation Advisor: Martha R. Weiss, Ph.D.
ABSTRACT
Predator-prey interactions are integral to the maintenance of community structure and
function. Predators and prey use multiple cues to detect and assess each other, and
identification of these cues is necessary to understand how selection operates to shape
predator-prey interactions. Mud-dauber wasps (Sphecidae), one of the main predators of
spiders, prefer araneids (two-dimensional web-building spiders) over derived araneoids,
(three-dimensional web-building spiders). Predation pressure by spider-hunting wasps is
considered a key factor in the evolution of 3D web-building spiders. However, the
proximate basis of such preference was not known.
By conducting behavioral and chemical assays I have determined that Sceliphron
caementarium wasps recognize araneids as potential prey due to the presence of
chemicals present on their silk and cuticle. By analyzing cuticular extracts of spiders in
several families, I have shown that all spiders that are taken by wasps have similar
cuticular chemistry; however, all spiders that have that similar chemistry are not taken by

iv
wasps. Indeed, spiders’ antipredator behavior, morphology and web architecture are
important factors that influence prey capture by wasps.
Though not captured by S. caementarium, derived araneoids are readily captured by
another sphecid, Chalybion californicum. Examinations of the hunting behavior of C.
californicum towards Achaearanea tepidariorum (cobweb spider, Theridiidae, derived
araneoid) reveals that the antipredator behavior of the cobweb spider changes over its
ontogeny. Whereas small-sized spiders drop off their webs (i.e., seem to treat wasp as
predators), medium and large-sized spiders attempt to warap wasps in silk (i.e., seem to
treat wasps as prey).
Besides araneids, Sceliphron also capture jumping spiders: however, they do not capture
Peckhamia picata, a jumping spider that mimics ants. Chemical analyses demonstrate
that ant mimics lack the chemical signature of a typical jumping spider but they
chemically resemble their model ant, Camponotus sp. My results suggest that P. picata
uses visual mimicry to avoid predatory jumping spiders, and employs chemical
information to elude wasps and perhaps ants. Thus P. picata uses different sensory
modalities to deceive multiple audiences.
My study highlights the importance of spider webs, chemistry and behavior in shaping
interactions between wasps and spiders, and contributes to our understanding of the use
of multimodal cues in predator-prey interactions.

v
To amma and ajji

vi
ACKNOWLEDGEMENTS
I am grateful to a number of people who have encouraged me and stood by my side
during several years of grad school life. Honestly, I do not have words to thank my
advisor, Martha Weiss. She has been a wonderful mentor, friend, and a source of
inspiration. I am grateful to Martha for allowing me to explore the world of spiders, an
area which was not related to her own research. Working in Martha’s lab was a pleasure,
as I could learn and participate in numerous on-going projects in the lab, which allowed
me to broaden my horizon and to appreciate biology in a more holistic manner. I feel
proud to say that I was of her student. By interacting with her, I have also developed a
deep appreciation towards plants around me. I will miss the summer days spent on the
Observatory hill, where we were always exploring something new, designing
experiments or coming up with a new arts and craft project. This dissertation would not
have been possible without Martha’s constant encouragement, her brilliant thoughts, and
her help with writing proposals and papers. I hope, one day I could write as beautifully as
you do.
Besides my advisor, I am indebted to Todd Blackledge, my committee member for being
my wasp and spider guru. Thank you for providing several insightful inputs through these
six years, for giving critical feedback on papers and proposals and for helping me in
collecting spiders. Only after following your own PhD work on wasps and spiders, I
learnt the tricks of the trade in working with solitary sphecid wasps!

vii
I would not have known that spider-hunting wasps existed but for a conversation I had
with my other committee member, Jonathan Coddington. I am grateful to you for
allowing me to spend time looking through the spider collection at the Natural History
museum. Thanks also to Janet Mann, my other committee member who graciously agreed
to be on my PhD committee, and for her inputs on my dissertation.
This work would not have been possible without the collaboration with Larry Phelan,
chemical ecologist at Ohio State University. I owe my deepest gratitude to Larry Phelan,
Rodrigo Chorba and Jenny for helping with the chromatograms. I am also grateful to
Gudrun Herzner, and Erhard Strohm from University of Regensburg for their help with
the ant mimicry project, and their hospitality while I visited Regensburg. I also sincerely
thank Catalina Estrada and Johannes Kroiss for your guidance with the chemical analysis.
Much appreciation goes to John Lill and Shannon Murphy and other members of the D.C.
Plants Insect Group (DC PIG) for their excellent inputs while I was working on the
manuscripts. I would like to thank Peter Armbruster for being always available to talk
about ideas, and Edd Barrows, for sharing his vast natural history knowledge about
anything we saw out in the Observatory garden! My deepest gratitude to Jeremy Miller
who taught me how to identify spiders, and Fernando Pallida, Lara Lopardo and G. B.
Edwards for identifying spiders. This research would not have been possible but for the
timely statistical help from Gina Wimp and Aaron Howard, who devoted much time with
me to answer my naïve questions. My earnest thanks to a number of others with whom I
corresponded via emails to learn about different aspects of wasps and spiders. These

viii
include Michael Greene, Mark Elgar, Gudrun Herzner, Allan Hook, Stefan Schulz and
James Pitts.
I would not have enjoyed being in grad school but for my wonderful lab mate Heather
Mallory. Thanks Heather for all your help, your insights about anything I wanted to
know, for your company, and for your efforts to expose me to the American culture and
way of life!—I will truly miss you. I owe you a big treat for helping me format the page
numbers on the last day—I could not have done this without your help! I was lucky to
have very good lab mates and fellow graduate students throughout these six years in
Georgetown- Thank you Aaron Howard, Caitlin Durkee, Karin Bothwell, Doug
Blackiston, Dani Rodrigues, Sheena Pooter, Eric Oberdorf, Jean Tsai, Debbie Ladner,
Dan Kjar and Tenley Archer and many more.
I would not have survived graduate school without interactions with a number of dear
friends, most of who do not live in the DC area, but kept in touch through emails! Thank
you, Srini, Smita, Roy, Daniel, Avanti, Tanu, Anju, Swati, Rhitu, Manu, Elu, Suneela and
Vivek for being available when I needed you most. Thanks to Raghvendra who did not
think I am crazy, but cheerfully to part in collecting spiders in India. I am also grateful to
a number of close friends in the area with whom I have explored the multicultural events
that DC has to offer. Thank you Agomoni, Isha, Srinjoy, Shailaja, Krishna, Kirk and Alpa
for your wonderful company and friendship. I am indebted to my housemate Alpa, who
cooked for me, and took care of things during the stressful times before my defense. I
would not have pulled through the last few months before my defense but for the

ix
affection, friendship and solid support of my dear friend Kirk—thanks for just being there
for me.
Most importantly I would like to thank my close friend and husband Sayantan, without
whom I would not have joined grad school in the first place! Thanks for believing in me,
sharing my excitement about new ideas and for showing me that the only way to see
whether something works is to try it! I would also like to thank my in-laws for their love
and support. My love for nature comes from having spent many hours during my
childhood in the backyard of my home in Shimoga, India, either attempting to feed Ashy
Wren Warbler babies, watching ant trails, or shooing monkeys that came to eat guava
fruits! I feel proud and privileged to have grown up in such surroundings, and I owe it to
the love of my mom and grand mother, without whom I would not have come so far.
This research has been supported by funding from Georgetown University, American
Arachnological Society, Washington Biologists’ Field Club, Sigma Xi, Animal Behavior
Society student grant, American Museum of Natural History (Theodore Roosevelt award)
and a NSF dissertation improvement grant, and travel grants from the Center for
Environment, Georgetown University.

x
TABLE OF CONTENTS
Introduction………………………………………………………………………………1
Chapter 1: Chemical mediation of prey recognition by spider-hunting wasps…….23
Methods…………………………………………………………………………..26
Results…………………………………………………………………………....33
Discussion………………………………………………………………………..36
References…………………………………………………………………….….44
Figures…………………………………………………………………………....51
Appendix I. Chemical basis of wasps’ preference of araneids over derived
araneoids……………………………………………..………………………….55
Results…………………………………………………………………….……...57
Chapter 2: Spider chemistry, web architecture and behavior: factors
influencing prey capture success by sphecid wasps…………………………..61
Methods…………………………………………………………………………..63
Results…………………………………………………………………………....69
Discussion………………………………………………………………….…….73
References……………………………………………………………….….……79
Figures…………………………………………………………………….……...83
Chapter 3: Prey turns predator: ontogenetic changes in the antipredator behavior
of theridiid spiders against mud-dauber wasps………………………………90
Methods……………………………………………………………………….….93
Results…………………………………………………………………….……...96

xi
Discussion…………………………………………………………….………….98
References…………………………………………………………….…...……103
Figures………………………………………………………………….……….106
Chapter 4: Double deception: ant-mimicking spiders fool both visually and
chemically-oriented predators………………………………………………..111
Methods………………………………………………………………………..115
Results………………………………………………………………………….119
Discussion……………………………………………………………………...121
References……………………………………………………………………...127
Figures………………………………………………………………………….132

xii
LIST OF FIGURES AND TABLES
Chapter 1: Chemical mediation of prey recognition by spider-hunting wasps
Figure 1. Mud-dauber wasp Sceliphron caementarium and its nest
contents…………………………………………………………………………..51
Figure 2. Spiders captured by S. caementarium when spiders were on or off their
webs……………………………………………………………………………...52
Figure 3. Amount of time wasps spent antennating filter paper discs containing
spider silk and chemical residues………………………………………………...53
Figure 4. Amount of time wasps spent antennating spider dummies……………54
Chapter 1: Appendix Chemical basis of wasps’ preference of araneids over derived
araneoids
Figure 1a.Wasps’ stinging response toward spiders……………………………..59
Figure 1b. Average time wasps spent antennating spider dummies……………..60
Chapter 2: Spider chemistry, web architecture and behavior: factors influencing
prey capture success by sphecid wasps
Figure 1. Schematic diagram of phylogenetic relationship between web and non-
web-building spiders……………………………………………………………..83
Figure 2. Non-metric multidimensional scaling plot visualizing chemical
similarities of spider genera…………………………………………….………..84
Figure 3. Wasps captured most araneids and did not capture derived araneoids or
uloborids…………………………………………………………………………85

xiii
Figure 4. Antipredatory behaviors of spiders differ based on taxonomic affiliation
regardless of web architecture…………………………………………………...86
Figure 5a. Chromatogram of Larinioides sclopetarius…………………………..87
Figure 5b. Cuticular chemical profile of Achaearanea tepidariorum…………...88
Figure 5c. Chemical profiles of Larinioides, Phidippus and Misumena……….89
Chapter 3: Prey turns predator: ontogenetic changes in the anti-predator behavior
of theridiid spiders against mud-dauber wasps
Figure 1. Achaearanea tepidariorum (Theridiidae) on its cobweb…………….106
Figure 2. Spider morphometrics………………………………………………..107
Figure 3. Spider behavior changes with size………………………………...…108
Table 1. Four outcomes of wasp-spider interactions………………………...…109
Table 2. Wasp and spider behaviors……………………………………………110
Chapter 4: Double deception: ant-mimicking spiders fool both visually and
chemically-oriented predators
Figure 1. Ant-mimicking spider, Peckhamia picata and its model ant
Camponotus sp………………………………………………………………….132
Figure 2. Predatory spiders are fooled by ant-mimicking spiders……………..133
Figure 3. Chromatographs of ant, mimic and non-mimetic spider……………..134
Figure 4. Nonmetric multi-dimensional scaling plot of ant, mimic and
non-mimetic spider cuticular chemical profiles……………….………………..135

1
INTRODUCTION
Spiders as predators and prey: the influence of webs, chemicals and behavior
Spiders as prey
Spiders are top predators of arthropods in terrestrial ecosystems, and they play a critical
ecological role by keeping insect populations in check (Schmitz 2008). Spiders, however,
also fall prey to both vertebrate and invertebrate predators (Bristowe 1941; Coville 1987;
Gunnarsson 2007). Vertebrate predators such as birds and lizards differentially affect
spider densities. For example, birds captured large spiders, mobile spiders and those
belonging to the families Linyphiidae, Theridiidae and Agelinidae more often than they
did small spiders, stationary spiders and spiders belonging to the family Lycosidae
(Gunnarsson 2007). Lizards, on the other hand, not only negatively affected spider
densities, but also altered prey abundance and composition (Schoener & Spiller 1987).
Lizard predation also affected web-building spiders that had no shelter/protection
(Manicom et al. 2008).
Invertebrates that prey on spiders include spiders themselves (e.g., spiders in the family
Mimitidae and Salticidae), as well as solitary wasps in the families Pompilidae and
Sphecidae. In addition, spiders are parasitized by nematodes, ichneumonid wasps and

2
parasitic acrocerid flies (Foelix 1996). Pompilids provision their larval nests with a single
large paralyzed spider. The majority of pompilids hunt their prey first and then prepare
their nests, the spider being placed nearby while the nest is under construction (Evans
1953). On the other hand, sphecids construct nests first and then provision them with
multiple prey items(Evans 1953, Coville 1987). Mass provisioning of nests with prey
seems like an efficient strategy, as small prey are often abundant, and easily caught and
carried, to the nest. Sphecids are thus able to build nests in favorable sites, and not
constrained by local prey abundances (Coville 1987).
Sphecids prey upon spiders and as many as 10 different insect orders(Evans 1953).
Unlike other wasps, members of the family Sphecidae exhibit remarkable host
specificity, in that different subfamilies, genera, and even species show different degrees
of host specialization, so that prey type often provides a useful taxonomic character
(Evans 1953).
Six genera belonging to two subfamilies, Sphecinae and Larrinae, contain wasps that hunt
both web-building as well as non-web-building spiders (Bohart & Menke 1976; Coville
1987). Individual female wasps provision a mud nest with a dozen or more paralyzed
spiders, and build multiple nests over the course of their three to six week-long adult
lifespans (Coville 1987). Mud-dauber wasps are selective in their choice of spider:
numerous examinations of mud cell contents, made over a span of a century, demonstrate
that spiders that built two-dimensional webs (2D spiders; Araneidae) are
disproportionately represented relative to spiders that built three-dimensional webs (3D

3
spiders; derived araneoids) (Hartman 1905; Rau 1935; Muma & Jeffers 1945; Bohart &
Menke 1976; references in Blackledge et al. 2003), even though the latter are more
abundant in habitats where the wasps forage (Blackledge et al. 2003). Mud-dauber wasps
hunt araneids in over 45 genera compared to only 16 genera of theridiids (spiders that
build 3D cobwebs) (appendix, Blackledge et al. 2003).
Why do wasps take araneids and not derived araneoids? Chapter One focuses on the
proximate mechanism of prey recognition by the mud-dauber wasp Scelipheron
caementarium (Sphecidae). By conducting a series of behavioral experiments, I
determined that wasps use chemical cues present on spiders’ silk and cuticle to recognize
and differentiate between prey (Larinioides sclopetarius, Araneidae) and non-prey
(Achaearanea tepidariorum, Theridiidae) spiders.
To understand spiders as ‘prey,’ in the following sections I will summarize spiders’
cuticular chemicals; antipredator behavior and deceptive mimicry and within this
framework will highlight the questions addressed in each of the chapters.
1. Use of cuticular hydrocarbons in communication
Insect cuticular hydrocarbons function primarily to prevent desiccation, but have been
used by insects for both intra- and interspecific communication in recognizing species,
gender, and nestmates. They also serve as task-specific cues, dominance and fertility
cues, as territorial pheromones, as alarm, recruitment and chemical defenses, as

4
thermoregulartory pheromones, and as semiochemical cues for predators and parasitoids
(Singer 1998).
Spiders, like many insects, make use of chemical information in a number of activities,
including finding prey location or (Persons & Uetz 1996, Punzo & Kukoyi 1997, Persons
& Rypstra 2000), mate location (Kasumovic & Andrade 2004, reviewed in Gaskett
2007), enemy avoidance (Persons et al. 2001), social recognition (Pollard et al. 1987),
species recognition or reproductive isolation (Barth 2002). Chemical information can be
volatile or substrate-borne, and is obtained either through either spiders’ cuticle and/or
silk.
While chemical information derived from silk or cuticle is mainly used among
conspecific or heterospecific spiders, little is known about whether such information is
also exploited by other insects. Gelis festinans, an eggsac parasitoid wasp, is attracted to
the silk of its linyphiid host spider (Van Baarlen et al. 1996). The pompilid wasp, Pepsis
formosa is attracted to the olfactory cues of its host theraphosid spider (Punzo 2006).
Results of my first chapter reveal that the sphecid wasp Sceliphron caementarium also
uses spiders’ cuticular chemicals to recognize and discriminate among potential spider
prey.
Based on the examination of nest contents, Sceliphron can be considered a generalist
predator: it is known to capture spiders in at least 21 araneid genera with varying web
architecture, as well as spiders in some non-web building families (see appendix,

5
Blackledge et al. 2003). In the second chapter I test the generality of S. caementarium’s
use of spider’s cuticular chemicals to recognize particular spiders as prey. Specifically,
by conducting chemical and behavioral assays, I determined that most spiders captured
by wasps have similar cuticular chemistry, but that most spiders that share that particular
cuticular chemistry are not necessarily captured by wasps.
2. The role of web architecture and antipredatory behavior of spiders against
predators
Defensive behaviors of spiders are intimately associated with the web architectures and
resting position of spiders. Cloudsley-Thompson (1995) classifies defensive behavior of
spiders into primary and secondary defenses. Primary defenses are those that operate
regardless of whether a predator is in the vicinity or not. They include the presence of
retreats, barriers, stabilimenta (conspicuous silk decorations), and communal webs, as
well as spines on spiders’ abdomens. Secondary defenses operate when predators have
actually found the spiders, and include spiders moving away from the site of attack,
spiders dropping to the ground, and vibrating their webs. In general, most studies on
defensive behaviors have focused on two-dimensional web-building spiders (e.g.,
Argiope), reflecting a general bias toward studying them (Cloudsley-Thompson 1995).
Primary defenses
Retreats: Retreats are places where spiders rest or hide; typically a silken tube, a tangled
matrix of silk, a dry leaf or a crevice. Manicom et al. 2008 found that web-building

6
spiders that included retreats were better protected from lizards than those were spiders
that did not have retreats. Retreats do not seem to be effective against pompilid and
sphecid wasps, which often succeeded in pursuing Araneus cornutus (Araneidae) out of
their silken-tubes either by entering the retreats, thereby forcing the spiders to drop down,
or tearing apart the retreat by their mandibles (Eberhard 1970, Divya Uma personal
observation of another retreat-dwelling orb-weaver). The spiral silken retreat of
Achaearanea globispira (Theridiidae) is covered with sand grains, and is thought to be
protective against ants and wasps (Henschel & Jocque 1994), a hypothesis that has not
been explicitly tested.
Barrier webs, silk and debris decorations: Two-dimensional web-building spiders in the
genera Nephila and Metepeira often build three dimensional silk matrices that are
attached behind and/ or above the spiders’ 2D webs. These retreats are thought to prevent
or delay predator attacks (Edmunds & Edmunds 1986). In our own study, Metepeira
labyrinthea that rested within their prey carcass-barrier webs escaped S. caementarium
attacks 56.3% of the time (Chapter 2). Prey-carcass decorations built by Cyclosa sp.
(Araneidae) redirected paper wasp attacks, and thus enhanced the survival of spiders
(Chou et al 2005). Allocyclosa bifurca incorporated their exoskeletal remains in their
detritus stabilimenta (Eberhard 2003). Freshly shed spider skins contain chemical cues on
them, and are often stung by S. caementarium (personal observation). Thus, the presence
of exoskeletal remnants may divert attention of spider-hunting wasps as a result of
chemistry. Debris in the web of a 3D spider Latrodectus revivensis (Theridiidae)
provided both defense and concealment against the grey shrike (Konigswald 1990).

7
Communal webs: While the majority of spiders are solitary, a small minority live in
colonies and even cooperate in web building, prey capture and brood care. In one other
communal spider, Metepeira incrassata (Araneidae), individuals that are located at the
core of large colonies experienced lower risk of attack by predatory wasps, as they
benefited from ‘early warning’ of predator approach through vibrations generated by
predators, as well as evasive responses of other spiders (Rayor & Uetz 2000). Vollarath
(1982) found that survival of single Anelosimus eximius (Theridiidae) females on their
webs was rare and that such mortality was mainly due to Trypoxylon wasps, damsel flies,
and kleptoparasitic spiders. However, rate of survival increased with colony size.
Whether communal webs of A. eximius physically protected spiders, or web-borne
vibrations helped spiders to avoid capture was not clear.
Spines: Diurnal 2D spiders such as Micrathena and Gasteracantha have leathery
abdomens and spines on the ventral side of their abdomens. These morphological
characters are thought to be defensive against wasp,s as these spiders were rarely found
in mud nests of spider-hunting wasps (Muma & Jeffers 1945, Gonzaga &Vasconcellos-
Neto 2005, Polidori et al. 2007, and personal observation). One likely explaination is that
the early instars of wasp larva may find the hard spiny exoskeletons of Micrathena and
Gastercantha difficult to penetrate. Early instars of sphecid wasp larvae may not have
well-developed chewing mandibular mouthparts, and thus may feed on softer parts of the
available prey. Indeed, in the majority of the solitarty wasps, the larva does not consume

8
the hard body parts of the prey (e.g., head capsule, wings, legs, body wall) (Evans &
Eberhard 1970).
Secondary defenses operate when predators have actually found the spiders. Some
widespread responses include spiders moving away from the site of attack, spiders
dropping to the ground and vibrating their webs and throwing silk.
Dropping off the web and web vibration: Dropping to the ground from a web or a retreat
by a drag line is a common and effective escape behavior, especially among araneids
(Eberhard 1970, Rayor 1996, Blackledge & Pickett 2000, Divya Uma personal
observation). Upon disturbance, juveniles may drop more often than adults (e.g., Argiope
sp., Jackson et al.1993). Vibrating, flexing, pumping and shuttling around the web are
other defensive behaviors, and have been recorded mostly in Argiope (Jackson et al.
1993). However, in a recent study, most Argiope just dropped in response to sphecid
wasp attacks, and were not found to vibrate or shuttle around their webs (Blackledge &
Pickett 2000).
Defensive behaviors of spiders change with ontogeny and web architecture (Li et al.
2003). Upon a simulated predatory attack, juvenile Argiope versicolor spiders shuttled on
their stabilimentum-decorated-webs more often than spiders on undecorated webs. Adults
of the same species pumped their stabilimenta-decorated webs more than spiders on webs
without stabilimenta (Li et al. 2003). Predation risk can also cause spiders to alter their
web-building behavior: when exposed to chemical cues of a predatory spider, Portia

9
labiata, juvenile A. versicolor decreased the frequency of incorporating stabilimenta to
their webs and decreased web capture area (Lee & Li 2004).
Anti-predator behaviors of web-building spiders are varied, complex, context-dependent
and closely associated with their webs. Only carefully controlled manipulative
experiments can separate out the contributions of spiders and their webs in escaping their
enemies. In the second chapter, I examine the antipredatory behavior of spiders in
multiple araneid and derived araneoid taxa against Sceliphron caementarium wasps. In
addition, in the fourth chapter, I examine the ontogenetic changes in antipredator
behavior of a cobweb spider, Achaearanea tepidariorum, against spider-hunting
Chalybion wasps.
3. Ant mimicry in spiders
Insects and spiders belonging to over 200 genera in 50 families are known to resemble
ants morphologically, behaviorally or chemically, and thereby to gain protection from
predators and/or access to resources within ant nests (McIver & Stonedhal 1993, Cushing
1997). In the following paragraphs, I briefly review different, non-mutually exclusive
categories of ant mimicy in spiders and highlight the areas where further research is
needed.
A. Morphological and behavioral resemblance to ants (myrmecomorphs)

10
Myrmecomorphy in spiders is reported in 13 families, though it is seen most often in
Salticidae (jumping spiders) and Corinnidae (sac spiders) (Cushing 1997). Ant-
mimicking spiders exhibit a range of spectacular adaptations, including shape and color
modifications; an ant-like gait, and an ‘antennal illusion’, achieved by holding up and
waving the first or the second pair of legs in a manner that suggests an ant’s antennae.
Most myrmecomorphs are thought to be Batesian mimics of ants (Cutler 1991, McIver &
Stonedahl 1993, Cushing 1997, Nelson et al. 2005, 2006, Nelson & Jackson 2006), and
are thought to be protected from visually-oriented predators. Myrmecomorphic spiders
are generally found within the same microhabitat as ants (Cushing 1997) and, in some
cases build their nests next to ant nests, thereby risking aggression or predation by their
potential models (Nelson et al., 2004, 2005, Nelson & Jackson 2008). Whether
myrmecomorphs also chemically resemble their model ants, has not been known untill
now.
B. Myrmecomorphs that feed on ants (aggressive mimicry)
Some myrmecomorphs are myrmecophages; i.e., they feed on ants. The crab spider
Amyciaea forticeps does not normally resemble its host Oecophylla ants morphologically,
but during hunting, it adopts a posture of a struggling or dying ant, there by drawing
worker ants. When an ant is close by, the spider quickly snatches the ant (Mathew 1944).
Another spider, Corinna vertebrata (Clubionidae), is a myrmecomorph that is also a
myrmecophage (Fowler 1983). Ant-mimics in at least 6 spider families are thought to be
myrmecophages (Cushing 1997).

11
C. Chemical resemblance to ants (myrmecophiles). Myrmecophily in spiders is seen in
12 families, and these spiders are found living with ants-- either close to ant nests, inside
ants’ brood chambers or refuse piles (Cushing 1997). They do not resemble ants
morphologically or behaviorally, but rather mimic them chemically (Cushing 1997).
Chemical mimicry is achieved by acquiring the colony odor or host-specific compounds
by coming in contact with ants (Vander Meer et al. 1989) or feeding on ant brood
(references in Cushing 1997, Allan & Elgar 2001, and Allan et al. 2002). Thus,
myrmecophiles can also be aggressive mimics.
D. Morphological, behavioral and chemical resemblance to ants (Multimodal
mimicry)
Myrmecomorphic spiders are often found close to ants, and as a consequence they risk
aggression or predation by their own models (Nelson et al., 2004, 2005, Nelson &
Jackson 2008). Indeed, if the spiders are closely associated with ants to the point that they
evolve visual resemblance, then it is also likely that they will be vulnerable to ant
aggression. Because ants mainly rely on chemical cues to discriminate nestmates from
non-nestmates (Lahav et al. 1998, Ozaki et al. 2005), it is possible that ant-mimicking
spiders will evolve chemical resemblance to avoid ant aggression. A few anecdotal
evidence suggests that myrmecomorphic spiders may resemble their model ants
chemically (Cushing 1997), but whether they actually do so has not been tested. In the
third chapter I examine whether the ant mimic Peckhamia picata chemically resemble its
model ant Camponotus sp.

12
Spiders as Predators
Spiders are one of the most ubiquitous groups of predators in terrestrial ecosystems that
feed almost exclusively on insects. Spiders are also excellent generalist predators, and
assemblages of spiders are used as biological control agents in many natural as well as
agroecosystems (Riechert & Leslie 1990). Hunting modes of spiders (i.e., sit-and-wait
predators or active foragers) are known to alter prey populations, and can have far-
reaching effects on ecosystem functions (Schmitz 2008). In the following paragraphs I
will briefly discribe the factors affecting prey selection by spiders, as well as multimodal
cues used by spiders to perceive prey and to communicate with their surroundings.
Factors affecting prey selection by spiders
Web-building spiders were once thought as passive filterers, eating just about anything
and showing ‘no trace of discrimination’. However, we now know that spiders are quite
selective in their prey capture and consumption (Bristowe 1941). Infact, prey
discrimination can be concidered as an example of optimal foraging, where within the
range of available prey, there exsists an optimal subset of prey type the capture of which
is most profitable for a particular spider. Prey size (not larger than the spider, but not
smaller than 1/6th
of the spider, (Bristowe 1941)), prey type (not chemically noxious or
distasteful, predatory insects, or very large prey, (Foelix 1996)), relative prey availability,
physiological state of a spider (pre or post-molt, hunger level, etc., reviewed by Riechert
& Luczak 1982), spider web’s charecteristic such as mesh size (webs with larger mesh
width capture larger insects and vise versa, (Uetz et al. 1978)), and habituation

13
(probability of attack increases with familiarity with a given prey, Turnbull 1960) all
affect prey selection by spiders (reviewed by Riechert & Luczak 1982).
Multimodal communication in spiders
Spiders use multiple modalities to percieve and respond to different signals in their
surroundings. Spiders detect prey, predators, mates and conspecifics by using a
combination of different senses: a) Mechanical sense- specialized hairs called
trichobothria and slit sensillae detect vibratory, acoustic and tacticle cues. b) Chemical
sense- several chemoreceptor cells present on the tarsi, pedipalps and pit organs percieve
pheromones and prey odor; c) Visual sense- several pairs of eyes detect shape, motion
and color. In the following paragraphs, I will highlight how spiders use these different
sensory modalities to detect prey.
Vibrational cues - Air-borne and substrate-borne vibratory cues, are used by spiders for
detection and localization of prey (Barth 1982, Barth 2002), are best studied in orb
weaving spiders, as the geometry of a two-dimensional orb makes it easy to examine the
effect of vibrations. An orb-weaving spider, sitting at the hub (center) of its web, is
thought to localize a prey item or an object stuck in the web mainly by sensing the
difference in vibrations on the radii (strands radiating from the center of the web) and the
spiral threads of a web (reviewed in Barth 1982). Interestingly, web-building spiders that
remain at the periphery of their webs or within a retreat, place the tarsus of one of their
front legs on a signal thread, which transmitts vibrations from the hub.

14
Chemical cues- Chemical cues, thought to be most primative signals, and have been
mostly studied in the context of mate selection. Pheromonal cues are thought to convey
location of a mate, sexual receptivity, timing and frequency of mating and promote or
avoid sperm competition (Gaskett 2007). Chemical cues are also used by spiders in prey
and predator detection (Persons et al. 2001, Persons & Uetz 1996). Recent studies have
shown that wolf spiders, Hogna helluo, are preferentially attracted to odor cues
associaetd with their most recent prey (i.e. what ever they last fed on) (Persons & Rypstra
2000).
Visual cues -Though most spiders lack acute vision, salticids (jumping spiders) and
lycosids (wolf spiders) possess well-developed eyes and employ visual cues
predominantly in foraging and courtship (Forster 1982). While wolf spiders have good
image resoution, jumping spiders have excellent pattern recognition, motion detection,
and color perception of objects while they themselves are moving (Rovner 1989). In
general, different eyes are used for diffeernt activities--whether to catch aprey, avoid
predators or find a suitable mate. A typical predatory sequence involves detection and
localization of prey (orienting towards an object with lateral eyes); evaluation of the
target (assess size, distance, and rate of movement), stalking, or chasing (through
principle anterior lateral eyes), and finally attacking a prey (reviewed by Forster 1982).
Spiders often use multiple modes of communication, and there is considerable diversity
in signalling behavior in spiders, perhaps because of the variation seen across species,

15
genera and families. There could be several reasons why spiders (and also other animals)
use multiple sensory modalities to communicate: a) multiple signals may be presented
either simultaneously or sequentially to convey different messages to the same or
different recievers, b) multiple signals may convey the same meaning (i.e., redundancy
in signalling), and this may help to reduce mistakes made by receivers, c) multiple signals
may componsate for environmental variability, and finally d) multiple signalling may be
a response to multiple selective pressures (Uetz & Roberts 2002). In Chapter Three I
examine spiders as predators and prey of Chalybion, a mud-dauber wasp, and discuss the
potential cues used by spiders to assess and categorize wasps as predators or prey. In
Chapter Four, I examine how predatory jumping spiders respond to visual signals of an
ant-mimicking spider prey.
REFERENCES
Allan, R. A. & Elgar, M. A. 2001: Exploitation of the green tree ant Oecophylla
smaragdina by the salticid spider Cosmophasis bitaeniata. Aust. J. Zool. 49,129-
137.
Allan, R. A., Capon, R. J., Brown, W. V. & Elgar, M. A. 2002: Mimicry of host cuticular
hydrocarbons by salticid spider Cosmophasis bitaeniata that preys on larvae of
tree ants Oecophylla smaragdina. J. Chem. Ecol. 28, 835-848.
van Baarlen, P., Topping, C. J. & Sunderland, K. D. 1996: Host location by Gelis
festinans, an eggsac parasitoid of the linyphiid spider Erigone atra. Entomol. Exp.
Appl. 81, 155-163.

16
Barth, F. G. 1982: Spiders and vibratory signals: sensory reception and behavioral
significance. In Spider Communication: Mechanisms and Ecological Significance
(ed. by P.W. Witt and J.S. Rovner), Princeton Univ. Press, Princeton, pp. 67–120.
Barth, F. G. 2002: A Spider’s World: Senses and Behavior. Springer, Heidelberg,
Germany.
Blackledge, T. A. & Pickett, K. M. 2000: Predatory interactions between mud-dauber
wasps (Hymenoptera, Sphecidae) and Argiope (Araneae, Araneidae) in captivity.
J. Arachnol. 28, 211-216.
Blackledge, T. A., Coddington, J. A. & Gillespie, R. G. 2003: Are three-dimensional
spider webs defensive adaptations? Ecol. Lett. 6, 13-18.
Bohart, R. M. & Menke, A. S. 1976: Sphecid Wasps of the World. Berkeley: University
of California Press, California.
Bristowe, W. S. 1941: The Comity of Spiders. The Ray Society, London.
Chou, I. C., Wang, P. H., Shen, P. & Tso, I. M. 2005: A test of prey-attracting and
predator defence functions of prey carcass decorations built by Cyclosa spiders.
Anim. Behav. 69, 1055-1061.
Cloudsley-Thompson, J. L. 1995: A review of the anti-predator devices of spiders. Bull.
Brit. Arach. Soc. 10, 81-96.
Coville, R. E. 1987: Spider-hunting sphecid wasps. In: Ecophysiology of Spiders.
(Nentwig, W., ed). Springer-Verlag, Berlin. pp. 309-318.
Cushing, P. E. 1997: Myrmecomorphy and myrmecophily in spiders: a review. Florida
Entomol. 80, 165–193.

17
Cutler, B. 1991: Reduced predation on the antlike jumping spider Synagelis occidentalis
(Araneae: Salticidae). J. Insect Behav. 4, 401–407.
Eberhard, W. G. 1970: The predatory behaviour of two wasps, Agenoideus humils
(Pompilidae) and Sceliphron caementarium (Sphecidae), on the orb weaving
spider Araneus cornatus (Araneidae). Psyche (Cambridge), 77, 243-251.
Eberhard, W. G. 2003: Substitution of silk stabilimenta for egg sacs by Allocyclosa
bifurca (Araneae: Araneidae) suggests that silk stabilimenta function as
camouflage devices. Behav.140, 847-868.
Edmunds, J. & Edmunds, M. 1986: The defensive mechanisms of orb weavers (Araneae,
Araneidae) in Ghana West Africa. In: Proceedings of the Ninth International
Congress of Arachnology, Panama 1983 (Ed. by W. G. Eberhard, Y. D. Lubin &
B. C. Robinson), pp. 73-89. Washington, D.C.: Smithsonian Institution Press.
Evans, H. E. 1953: Comparative Ethology and the Systematics of Spider Wasps. Syst.
Zool. 2 (4), 155-172.
Evans, H. E. & Eberhard, M. J. W. 1970: The wasps. Michigan University Press,
Michigan.
Foelix, R. 1996: Biology of spiders 2nd edn. New York: Oxford University Press
Fowler, H. G. 1984: Note on a clubionid spider associated with attine ants. J. Arachnol.
12, 117-118.
Forster, L. M. 1982: Vision and prey catching strategies in jumping spiders.
Am. Sci. 70, 165-75
Gaskett, A. C. 2007: Spider sex pheromones: emission, reception, structures, and
functions. Biol. Rev. 82, 27-48.

18
Gonzaga, M. O. & Vasconcellos-Neto, J. 2005: Orb-web spiders (Araneae:
Araneomorphae; Orbiculariae) captured by hunting-wasps (Hymenoptera:
Sphecidae) in an area of Atlantic Forest in south-eastern Brazil. J. Nat. Hist. 39,
2913-2933.
Gunnarsson, B. 2007: Predation on spiders: Ecological mechanisms and evolutionary
consequences. J. Arachnol. 35, 509-529.
Hartman, C. 1905: Observations on the habits of some solitary wasps of Texas. Bull.
Univ. Texas, Scientific Series 7, 65, 1-72.
Henschel, J. R. & Jocque, R. 1994: Bauble spiders: a new species of Achaearanea
(Araneae, Theridiidae) with ingenious spiral retreats. J. Nat. Hist. 28, 1287-1295.
Jackson, R. R., Rowe, R. J. & Wilcox, R. S. 1993: Anti-predator defences of Argiope
appensa (Araneae, Araneidae), a tropical orb-weaving spider. J. Zool. Lond. 229,
121-132.
Jackson, R. R. & Pollard, S. D. 1996: Predatory behavior of jumping spiders. Ann. Rev.
Entomol. 41, 287-308.
Kasumovic, M. M. & Andrade, M. C. B. 2004: Discrimination of airborne pheromones
by mate-searching black widow males: species- and population-specific
responses. Can J. Zool. 82, 1027–1034.
Konigswald, A., Lubin, Y, & Ward, D. 1990: The effectiveness of the nest of a desert
widow spider, Latrodectus revivensis, in predator deterrence. Psyche 97, 75-80.
Lahav, S., Soroker, V., Hefetz, A. & Vander Meer, R. K. 1999: Direct behavioral
evidence for hydrocarbons as ant recognition discriminators. Naturwissenschaften
86(5), 246-249.

19
Li, D., Kok, L. M., Seah, W. K. & Lim, M. L. M. 2003: Age-dependent stabilimentum-
associated predator-avoidance behaviours in orb-weaving spiders. Behaviour 140:
1135-1152.
Li, D. & Lee, W. S. 2004: Predator-induced plasticity in web-building behaviour. Anim.
Behav. 67, 309-318.
Mathew, A. P. 1944: Observations on the habits of the two spider mimics of the red ant
Oecophylla smaragdina (Fabr.) Bombay Nat. Hist. Soc. 52:249-63.
Muma, M. H. & Jeffers, W. F. 1945: Studies of the spider prey of several mud-dauber
wasps. Ann. Entomol. Soc. Am. 38, 245-255.
Manicom, C., Schwarzkopf. L., Alford R. A. & Schoener, T. W. 2008: Self-made shelters
protect spiders from predation. Proc. Nat. Acad. Sci. 105, 14903-14907.
McIver, J. D. & Stonedahl, G. 1993: Myrmecomorphy: morphological and behavioral
mimicry of ants. Ann. Rev. Entmol. 38, 351-379.
Nelson, X. J., Jackson, R. R., Pollard, S. D., Edwards, G. B. & Barrion, A. T. 2004:
Predation by ants on jumping spiders (Araneae: Salticidae) in the Philippines.
New Zeal. J. Zool. 31, 45-56.
Nelson, X. J., Jackson, R. R., Edwards, G. B. & Barrion, A. T. 2005: Living with the
enemy: jumping spiders that mimic weaver ants. J. Arachnol. 33, 813-819.
Nelson, X. J., Jackson, R. R., Li, D., Barrion, A. T. & Edwards, G. B. 2006:
Innate aversion to ants (Hymenoptera: Formicidae) and ant mimics: experimental
findings from mantises. Biol. J. Linn. Soc. 88, 23-32.
Nelson, X. J. & Jackson, R. R. 2006: Vision-based innate aversion to ants and ant
mimics. Behav. Eco.17, 676-681.

20
Nelson, X. J. & Jackson, R. R. 2008: Anti-predator crèches and aggregations of ant-
mimicking jumping spiders (Araneae: Salticidae). Biol. J. Linn. Soc. 94, 475-481.
Ozaki, M., Wada-Katsumata, A., Fujikawa, K., Iwasaki, M., Yokohari, F., Satoji, Y.,
Nisimura, T. & Yamaoka, T. 2005: Ant nestmate and non-nestmate discrimination
by a chemosensory sensillum. Science 309, 311-314.
Persons, M. H. & Uetz, G. W. 1996: Wolf spiders vary patch residence time in the
presence of chemical cues from prey (Araneae, Lycosidae). J.Arachnol. 24, 76-79.
Persons, M. H. & Rypstra, A. L. 2000: Preference for chemical cues associated with
recent prey in the wolf spider Hogna helluo (Araneae: Lycosidae).Ethology. 106,
27-35.
Persons, M. H., Walker, S. E., Rypstra, A. L. & Marshall, S. D 2001: Wolf spider
predator avoidance tactics and survival in the presence of diet-associated predator
cues (Araneae: Lycosidae). Anim. Behav. 61:43-51.
Pollard, S. D., Macnab, A. M. & Jackson, R. R. 1987: Communication with Chemicals:
Pheromones and Spiders. In Ecophysiology of Spiders (ed. W. Nentwig), Springer
Verlag, Berlin.pp.133-141.
Polidori, C., Federici, M., Pesarini, C. & Andrietti, F. 2007: Factors affecting spider prey
selection by Sceliphron mud-dauber wasps (Hymenoptera: Sphecidae) in northern
Italy. Anim. Biol. 57, 11-28.
Punzo, F. & Kukoyi, O. 1997: The effects of prey chemical cues on patch residence time
in the wolf spider Trochosa parthenus (Chamberlin)(Lycosidae) and the lynx
spider Oxyopes salticus Hentz (Oxyopidae).Bull. Brit. Arach. Soc.10, 323-326.

21
Punzo, F. 2006: Female spider wasps, Anoplius splendens Driesbach (Hymenoptera:
Pompilidae), learn to associate the odor of host feces with the presence of the
host. J. Entomol. Science 41, 202-210.
Rau, P. 1935: The spider prey of the mud wasp, Sceliphron caementarium (Araneae,
Hymen: Sphecidae). Entomol. News. 46, 267–270.
Rayor, L. S. 1996: Attack strategies of predatory wasps (Hymenoptera: Pompilidae;
Sphecidae) on colonial orb web-building spiders (Araneidae: Metepeira
incrassata). J. Kansas Entomol. Soc. 69, 67-75.
Rayor, L. S. & Uetz, G. W. 2000: Age-related sequential web-building as an adaptive
spacing strategy in the colonial spider Metepeira incrassata (Araneidae). Anim.
Behav. 59, 1251-1259.
Riechert, S. E. & Luczak, J. 1982: Spider foraging: behavioral responses to prey. In
Spider communication: mechanisms and ecological significance. (P.N. Witt, and
J. Rovner. Ed) Princeton University Press.
Rovner, J. S. 1989: Wolf spiders lack mirror-image responsiveness seen in jumping
spiders. Anim. Behav. 38, 526-533.
Schmitz, O. J. 2008: Effects of predator hunting mode on grassland ecosystem function.
Science 319, 952-954.
Singer, T. L. 1998: Roles of hydrocarbons in the recognition systems of insects. Am.
Zool. 38, 394-405.
Schoener, T. S., & Spiller, D. A. 1987: Effect of lizards on spider populations:
manipulative reconstruction of a natural experiment. Science 236, 949-952.

22
Turnbull, A. L. 1960: The prey of the spider Linyphia triangularis (Clerck) (Araneae,
Linyphiidae).Can. J. Zool. 38, 859-873.
Uetz, G. W., Johnson, A. D. & Schemske, D. W. 1978: Web placement, web structure
and prey capture in orb-weaving spiders. Bull. Brit. Arach. Soc. 4, 41-148.
Uetz, G. W. & Roberts, J. A. 2002: Multi-sensory cues and multi-modal communication
in spiders: insights from video/audio playback studies. Brain Behav. & Evol. 59,
222-230.
Uetz, G. W., Boyle, J., Hieber, C. S. & Wilcox, R. S. 2002: Anti-predator benefits of
group living in colonial web-building spiders - the “Early Warning” effect. Anim.
Behav. 63, 445-452.
Vollrath, F. 1982: Colony foundation in a social spider. Z. Tierpsychol. 60, 313-324.
Van Baarlen, P., Toping, C. J. & Sunderland, K. D. 1996: Host location by Gelis
festinans, an eggsac parasitoid of the linyphiid spider Erigone atra. Ent. Exp.
Appl. 81, 155–163.
Vander Meer, R. K., Jouvenaz, D. P. & Wojcik, D. P. 1989: Chemical mimicry in
a parasitoid (Hymenoptera: Eucharitidae) of fire ants (Hymenoptera: Formicidae).
J. Chem. Ecol. 15: 2247-2261.

23
Chapter 1
Chemical mediation of prey recognition by spider-hunting wasps
INTRODUCTION
Predator-prey interactions are integral to the maintenance of community structure and
function; they are important in mediating competition, regulating population size, and
influencing behavior (Sih et al. 1985; Schmitz 2008). Impacts of predator-prey and
predator-predator interactions can cascade across trophic levels, often dramatically
changing community composition and food web structure (Polis et al. 1999; Matsuda et
al. 1996).
In terrestrial as well as aquatic ecosystems, predators and prey use cues from a range of
sensory modalities, including visual (Cronin 2005), auditory (Surlykke & Moss 2000),
chemical (Greenstone & Dickens 2002), vibratory (Barth 1982), and electrical (Nelson &
Maciver 1998) information to detect and assess one another, For a given interaction,
identification of these cues is necessary if we are to understand the behaviors of both
parties, and, more fundamentally, how selection operates to shape the interactions.
Spiders are important predators of arthropods in terrestrial ecosystems, and they play a
critical ecological role by keeping insect populations in check (Schmitz 2008). Spiders,
however, also fall prey to wasps, toads, birds, and lizards, as well as other spiders

24
(Bristowe 1941; Coville 1987; Gunnarsson 2007). Six genera of mud-dauber wasps
(Sphecidae) hunt spiders (both web-building and non-web-building) exclusively, and
provision their mud nests with paralyzed spiders as food for their developing larvae (Fig.
1, Bohart & Menke 1976; Coville 1987). Individual female wasps capture up to 20
spiders daily and build multiple nests over the course of their three to six week-long adult
lifespans (Coville 1987). Mud-dauber wasps are selective in their choice of spider;
numerous examinations of mud cell contents, made over a span of a century, demonstrate
that spiders that built two-dimensional webs (2D spiders; Araneidae) are
disproportionately represented relative to spiders that built three-dimensional webs (3D
spiders; derived araneoids) (Hartman 1905; Rau 1935; Muma & Jeffers 1945; Bohart &
Menke 1976; references in Blackledge et al. 2003; personal observation), even though the
latter are more abundant in habitats where the wasps forage (Blackledge et al. 2003).
Thus, these wasps have a major impact on the spider populations with respect to species
composition as well as numbers of individuals. Investigation of the cues that mud-dauber
wasps use to choose spider prey will help us to understand the dynamics of this common
and important interaction, and will also shed light on selection pressures that are likely to
have contributed to spider diversification (Blackledge et al. 2003).
Why might mud-dauber wasps specialize on 2D web-building spiders? Their preference
could be based on an assessment of web architecture, as spider webs are important not
only in prey capture, but also in protecting spiders against predators (Nentwig & Heimer
1987; Blackledge et al. 2003). Blackledge and colleagues speculated that 2D spiders,
which sit on the exposed surface of their webs, may be more vulnerable to wasp

25
predation than are 3D spiders, which sit within a tangled matrix of web. They further
speculated that wasp predation may have favored the evolution of 3D webs from 2D
webs, the hypothesized ancestral character state. However, whether 3D webs physically
block wasp attacks has never been tested explicitly. Direct observational studies of prey
selection in this system are few and have been limited to mud-dauber wasps hunting 2D
spiders (Eberhard 1970; Rayor 1997; Blackledge & Pickett 2000; Pasquet et al. 2007).
Alternatively, it is possible that mud-dauber wasp prey preferences could be based on
chemical cues present on the spiders and/or their webs, rather than on web architecture.
Pompilid wasps that hunt non-web-building spiders detect and respond to spider chemical
cues (Punzo & Ludwig 2005; Punzo 2006), and many other predatory and parasitoid
hymenopterans use chemical cues to locate and identify their prey or hosts (Vet & Dicke
1992; van Baarlen et al. 1996; Anton & Gnatzy 1998; Steidle & Ruther 2000; Steidle et
al. 2001; Weiss 2003; Herzner et al. 2005; Roux et al. 2007). Thus it is plausible that
chemical cues play a role in prey recognition by S. caementarium.
Indeed, several recent studies reveal a chemical distinction between the silk of 2D spiders
and 3D spiders (Bagneres et al.1997; Papke 2000; Schulz 2004). Furthermore, our
preliminary Gas Chromatography and Mass Spectrometric (GC/MS) analyses indicate
that cuticular extracts of 2D spiders are dominated by saturated branched hydrocarbons,
while those of 3D spiders contain mostly esters of fatty acids, with trace amounts of
hydrocarbons (Chapter 2). Thus, 2D and 3D spiders present markedly different chemical
profiles to potential predators. Since most araneids build 2D webs, the presumed

26
preference for spiders inhabiting two-dimensional webs could in fact be a preference for
chemical cues associated with 2D spiders and their webs.
In this study we used behavioral experiments to investigate the roles of both web
architecture and chemical cues in prey selection by the mud-dauber wasp, Sceliphron
caementarium (Drury 1773). Specifically, we offered foraging S. caementarium wasps
2D and 3D spiders both on and off their webs to examine the importance of web
architecture on wasp prey preference. Furthermore, we conducted a series of behavioral
assays to investigate the importance of spider chemical cues in mediating prey
recognition by mud-dauber wasps.
METHODS
Spider web diversity
The family Araneidae (2D spiders), one of the largest families of web-building spiders, is
comprised of close to 3000 described species in 166 genera (Platnick 2008). Spiders in
this family typically build two-dimensional orb webs, but web modifications and
reductions occur in many genera. Araneids normally build webs overnight, and the
spiders sit in the center or periphery of their webs. Another large group of spiders, termed
the ‘derived araneoids’ (3D spiders), evolved from the Araneidae and is comprised of 11
families, 814 genera and 8286 species (Platnik 2008; Griswold et al.1998). These spiders
typically build and sit within a tangled three-dimensional matrix of silk; 3D webs are
usually built over several days.

27
Experimental set-up
We collected female Sceliphron caementarium (Drury 1773) wasps as they gathered mud
along the George Washington Memorial Parkway near metropolitan Washington, D.C.
during the months of June – September, from 2005 through 2009. Wasps were uniquely
marked using lead-free paint, and five or six wasps were housed per 2m3 outdoor mesh
cage. Each cage was provisioned with wet mud for nest construction and contained three
or four wooden shelters, as wasps prefer to build their nests in sites protected from rain.
Because mud-gathering and nest construction always precede spider-hunting (Evans and
Eberhard 1971), the caged wasps were motivated to hunt once they had built a mud cell.
A mixture of 1:1 honey: water was offered as a carbohydrate source. Petri dishes
containing the honey-water mixture were placed on inverted cups standing within a large
water-filled trough to reduce ant invasion. Grasses and forbs growing within the cage
provided resting sites for wasps and hiding places for fleeing spiders.
Spiders selected for the trials were available in the habitats where mud-dauber wasps
forage. Larinioides sclopetarius (Araneidae, 2D spider) and Achaearanea tepidariorum
(Theridiidae, 3D spider) were used as representative 2D and 3D taxa. We collected
immature spiders by visual search from Plummers Island, MD, and around the
Washington DC area from June to September 2005 to 2009. We did not use adult spiders,
as they were beyond the prey size (0.02-0.08 g) taken by S. caementarium (Blackledge
and Pickett 2000). Individual spiders were allowed to build webs in square wooden
frames (25x25cm²x 8 cm deep) covered with removable plastic wrap. Plastic wrap

28
prevented the spiders from escaping from the wooden frames. Once a week we fed the
spiders Drosophila or crickets, and misted their webs to provide moisture.
Wasp training and testing protocol
Once a wasp had built a mud cell, we lured her down to a ‘testing arena’ (a stool,
29x32x40cm, located in the centre of the cage): we held in front of her a 2D web-
building spider within its wooden frame and then slowly moved the frame down to the
stool. After the wasp touched the silk, she generally followed the frame down to the
testing arena, at which point she began to investigate the spider and its web. We
considered a successful training session one in which the wasp captured the spider, either
on or off its web. After one to three training sessions, the foraging wasp developed an
association between the testing arena and availability of a spider, and would
spontaneously come down to the testing arena when a wooden frame was placed there.
To be conservative, we used a different genus of 2D spider (Neoscona crucifera) rather
than Larinioides sclopetarius as our training spider; we could not use 3D spiders as prey
to train the wasps, because S. caementarium rarely hunt 3D spiders (Blackledge et al.
2003). Furthermore, in Hymenoptera, recognition of host- or prey-derived chemicals
seems to be innate (Vinson 1976; Vet et al. 1995; Raveret-Richter 2000), while prey
location (the testing arena in our experiments) is commonly learned (Vet et al. 1995).
Thus we believe that use of a 2D spider in training did not bias the results of our
subsequent behavioral assays.

29
Once the wasps were trained to visit the testing arena, we assessed their responses to
spider stimuli in four sets of experiments. We offered them 1) wooden frames containing
live 2D or 3D spiders on their webs; 2) freshly killed 2D and 3D spiders; 3) filter paper
discs containing spider silk and chemical cues; and 4) chemically manipulated spiders or
spider dummies. Within an experiment, each wasp was tested only once, but the same
wasps participated in different experiments. Spiders that survived wasp attacks were not
reused, to avoid any possible bias in the results if the spiders were to improve at avoiding
wasps over successive encounters. During a trial, all wasps in the field cage except the
one being tested were enclosed in a smaller mesh cage (30cm³) to prevent interference.
Once the trial was over, we released the wasps back into the field cage.
Influence of web architecture
Wasps’ responses to spiders in the presence of webs
To examine whether 3D webs act as barriers to wasp attacks relative to 2D webs, we
presented frames (without plastic wrap) containing either Larinioides sclopetarius (2D)or
Achaearanea tepidariorum (3D) spiders of similar weight to individual wasps, one at a
time, in a randomized sequence. Each trial, which consisted of a wasp being presented
with either a 2D or a 3D spider, commenced when a wasp first antennated the web or
spider and lasted until she successfully captured the spider or until 15 min had elapsed,
whichever happened first. Thus, each wasp participated in two trials, encountering one
2D and one 3D spider, with a 15 min interval between trials. The number of 2D and 3D
spiders captured by wasps was analyzed by binomial probability test. We recorded both
wasp and spider behaviors. Wasp behaviors included antennating the web, searching for

30
the spider on the web or on the ground, and capturing and stinging the spider. We also
noted whether the wasps’ initial contact was with the web or the spider. In 2005 and 2006
we recorded the amount of time wasps spent pursuing 2D and 3D spiders. Spider
behaviors included remaining on the web or moving to the periphery of the web,
dropping from and abandoning the web, and attacking the wasps.
Wasps’ responses to spiders in the absence of webs
In order to examine the response of wasps to spiders in the absence of their webs, we
simultaneously presented individual wasps with freshly killed 2D and 3D spiders,
positioned six cm apart on a filter paper disc (11 cm diameter). The paper disc itself was
placed on the horizontal surface of the testing arena. We obtained freshly killed spiders
by placing vials containing individual 2D or 3D spiders in the freezer for five minutes;
the spiders were used in trials within 10 minutes of removal from the freezer. Each trial,
which consisted of an individual wasp encountering a pair of freshly killed 2D and 3D
spiders, commenced when a wasp walked or flew to the paper disc and ended when the
wasp antennated both of the spiders, either stinging or rejecting them, or until 30 minutes
had elapsed, whichever happened first. We excluded three trials in which wasps failed to
encounter both spiders. The position of 2D and 3D spiders was switched between each
trial and a fresh paper disc was used each time. We used a binomial probability test to
analyze the number of freshly killed 2D and 3D spiders captured by wasps.
Influence of chemical cues
Spider cues on filter paper discs

31
To examine whether wasps base their prey choices on spiders’ chemical cues, we
presented wasps with filter papers containing silk and chemical residues from both 2D
and 3D spiders. We housed 2D and 3D spiders in individual cylindrical plastic vials (4x
8cm; 85 ml) lined with filter-paper discs (9 cm diameter) for two days prior to the
experiment. Spiders deposited drag-line silk as they walked on the papers; some also
defecated in the vials. Spiders were not fed while they were in the vials. Filter-paper discs
placed in vials without spiders served as controls. On the day of a trial, we placed an
empty wooden frame (no spider) on the testing arena, as the wasps had learned to come
down to the arena and search for spiders when the frame was present. We then placed
three paper discs, one each from 2D, 3D and control vials, on a horizontal plastic mat
(0.5x7x24 cm) in front of the frame. We positioned the three discs randomly for each
trial and did not reuse them. The plastic mat was wiped with ethanol after each trial to
remove any residual odors. We recorded the amount of time the focal wasp spent walking
and antennating on the paper discs within a five minute trial period. Antennation time
was considered as a proxy for interest in prey (Anton & Gnatzy 1998). We used
Friedman nonparametric analysis of variance (ANOVA) followed by Dunn’s post-hoc
tests, which correct for multiple comparisons, to compare antennation time between
treatments (Zar 1999).
Manipulations of cuticular chemistry
To investigate whether spider cuticular chemicals were necessary for prey recognition by
wasps, we obtained spiders without cuticular substances by washing freshly-frozen 2D
and 3D spiders twice in 1.5 ml of ACG-grade hexane for 15 minutes and subsequently

32
air-drying them on filter paper for 20 minutes. Hexane and other non-polar solvents are
routinely used to extract the hydrocarbons from insect and spider cuticles (Singer 1998;
Allan et al. 2002; Greene & Gordon 2003). The hexane solution from the first wash
contained one spider-equivalent of cuticular substances. To examine whether cuticular
cues were sufficient to elicit prey recognition (antennation followed by sting), we tested
wasps’ responses to spider-sized paper balls (approximately 5 mm in diameter) made
from black-colored construction paper and impregnated with spider cuticular extract. One
day prior to the bioassay, paper balls were soaked in 1.5 ml of 2D or 3D spider extract or
of hexane (control) and air dried for 24 hrs.
A single spider or paper ball was placed on an 11 cm diameter filter paper disc in the
testing arena, and wasps’ responses to the following seven treatments were examined in a
completely randomized sequence: a) 2D spiders lacking cuticular chemicals, b) 3D
spiders lacking cuticular chemicals, c) paper balls soaked with 2D spider chemicals, d)
paper balls soaked with 3D spider chemicals, e) paper balls soaked with hexane, f)
freshly killed 2D spiders and g) freshly killed 3D spiders. Presentations to the wasps of
any two treatments were separated by a 10-15 minute interval, and each wasp participated
in all seven treatments within one to two days. Wasp behaviors, including antennation,
stinging, and rejection of spiders and paper balls, were recorded for two minutes from the
time wasps landed on filter paper discs. As in the experiments involving responses of
wasps to chemical cues on filter paper discs, antennation time was considered as a proxy
for interest in prey (Anton & Gnatzy 1998).

33
We performed a series of pre-planned comparisons in order to evaluate the wasps’
responses to the treatments described above, To examine the importance of cuticular
chemicals for recognition of intact spiders as prey, we compared wasp responses to 1) 2D
spiders lacking cuticular chemicals vs freshly killed 2D spiders, and 2) 3D spiders
lacking cuticular chemicals vs freshly killed 3D spiders. We used a binomial probability
test to compare the number of freshly killed and hexane-washed spiders stung by the
wasps for comparisons 1 and 2. To assess the relative importance of visual and chemical
cues, and to test whether chemical cues are sufficient to elicit prey recognition by wasps,
we compared wasp responses to 3) 2D spiders lacking cuticular chemicals vs paper balls
soaked with 2D spider chemicals; 4) 3D spiders lacking cuticular chemicals vs paper
balls soaked with 3D spider chemicals; To examine whether 2D spider chemicals are
more attractive than 3D spider chemicals, we compared wasp responses to 5) paper balls
soaked with 2D spider chemicals vs paper balls soaked with 3D spider chemicals.
Finally, as controls, we compared wasp responses to 6) paper balls soaked with 2D spider
chemicals vs paper balls soaked with hexane and 7) paper balls soaked with 3D spider
chemicals vs paper balls soaked with hexane. We used Friedman nonparametric ANOVA
followed by Dunn’s post-hoc correction (Zar 1999) to determine whether antennation
time differed significantly between treatments for comparisons 3 to 7.
RESULTS
Influence of web architecture
Response of wasps to spiders in the presence and absence of webs

34
When offered L. sclopetarius (2D) and P. tepidariorum (3D) spiders on their respective
webs in a randomized sequence, wasps (N = 15) did not capture any 3D spiders, but took
11 of the 15 2D spiders (Fig. 2, binomial probability p = 0.0004, N = 11). The behavior
of wasps also differed when they encountered 2D or 3D spiders on their webs. In 2005
and 2006, the wasps (N = 10) pursued 2D spiders significantly longer than they did 3D
spiders (Mann Whitney U = 27.50, p = 0.04). They pursued 2D spiders for a median time
of 354 sec (±192.8 Median Absolute Deviation); while one wasp captured a 2D spider
that rested in the middle of its web in only two seconds, others spent as long as 11
minutes searching the corners of the wooden frame, vegetation and the ground below the
testing arena for spiders that hid at the periphery or dropped off their webs. In contrast,
the wasps searched the webs of 3D spiders for a median time of 36±65.4 sec (MAD)
before abandoning the hunt, even though these spiders never left their tangled 3D webs.
In almost all cases, the wasps touched spider silk before they encountered the spider; only
once, in 30 encounters, did a wasp catch a 2D spider at the centre of its web without first
touching the silk.
When offered a simultaneous choice between freshly killed 2D and 3D spiders without
their webs, wasps (N = 19) stung 18 of the 19 2D spiders and carried them back to their
nests. However, wasps stung and carried away only one out of 19 3D spiders; the rest
were antennated, but ignored (Fig. 2, binomial probability p<0.00001, N = 18)
Spider responses to wasps

35
Just as wasp behavior varied when hunting 2D and 3D spiders, spider behavior in the
presence of a hunting wasp also varied both between and within groups of 2D and 3D
spiders. While 2D spiders tended to hide at the periphery or drop off their webs, 3D
spiders stayed on their webs. Of the 15 2D spiders tested, five spiders that remained on
their webs (one at the centre, and four at the periphery), as well as the six that dropped off
their webs when wasps approached, were caught. The remaining four 2D spiders that
dropped down and abandoned their webs escaped. In contrast, 13 of the 15 3D spiders
tested remained on their webs when the wasps approached, and none were caught. The
remaining two 3D spiders acted as predators rather than prey; they came down from their
resting places and captured their erstwhile hunters!
Influence of chemical cues
Spider cues on filter paper discs
When offered a simultaneous choice of filter paper discs containing silk and chemical
residues from 2D and 3D spiders and a control disc, wasps (N = 10) spent significantly
more time antennating 2D than 3D or control discs (Fig. 3, Friedman ANOVA: F =
12.25, p = 0.0008; Dunn’s multiple comparison tests: 2D vs 3D, p<0.05; 2D vs control,
p<0.001; 3D vs control, p>0.05).
Manipulations of cuticular chemistry
Wasps stung all 13 of the freshly killed 2D spiders, but stung only one out of 13 hexane-
washed 2D spiders (comparison 1) (binomial probability test, p = 0.002, N = 13). In
contrast to the freshly killed 2D spiders, wasps stung only two of the 13 freshly killed 3D

36
spiders; again, they did not sting any of the hexane-washed spiders (comparison 2)
(binomial probability test, p = 0.25, N = 2). Furthermore, wasps (N = 13) antennated
paper balls soaked with 2D chemicals significantly longer than they did paper balls
soaked with 3D chemicals, balls soaked with hexane, or hexane-washed spiders
(Friedman ANOVA, F = 26.29, p = 0.0001). Specifically, wasps antennated paper balls
soaked with 2D spider chemicals more than hexane-washed 2D spiders (comparison 3,
Dunn’s post-hoc test, p <0.01), but did not differentiate between paper balls soaked with
3D chemicals and hexane-washed 3D spiders (comparison 4, Dunn’s post-hoc test,
p>0.5). Additionally, wasps spent significantly more time antennating paper balls soaked
with 2D spider chemicals than they did balls soaked with 3D spider chemicals
(comparison 5, Dunn’s post-hoc test, p <0.05), or balls soaked with hexane (comparison
6, Dunn’s post-hoc test, p < 0.01). Wasps did not differentiate paper balls soaked with 3D
spider chemicals and balls soaked in hexane (comparison 7, Dunn’s post- hoc test, p>0.5)
(Fig. 4).
DISCUSSION
Chemical cues, rather than web architecture, mediate wasps’ prey preferences
The results of our behavioral experiments clearly demonstrate that chemical cues, rather
than web architecture, mediate prey recognition by Sceliphron caementarium. We found
that wasps captured Larinioides sclopetarius (2D spider) and did not take Achaearanea
tepidariorum (3D spider) with or without their webs (Fig. 2). Responses of the wasps to

37
filter paper discs covered with spider silk and cuticular chemicals, as well as to
manipulations of spider cuticular chemistry, further underscore the importance of
chemical cues in prey recognition. The wasps spent significantly more time antennating
2D than 3D filter paper discs (Fig. 3), and similarly, antennated paper balls soaked with
2D chemicals significantly longer than they did paper balls soaked with 3D chemicals
(Fig. 4). Furthermore, wasps antennated, stung, and captured freshly killed 2D spiders,
but antennated and did not sting hexane-washed 2D spiders, indicating that presence of a
cuticular chemical is necessary for prey recognition.
Clearly S. caementarium recognize 2D spiders, but not 3D spiders, as prey. The lack of
wasps’ interest in 3D spiders is supported not only by our experimental results, but also
by over a century of natural history records, which document the absence of 3D spiders in
S. caementarium mud cells (Hartman 1905; Rau 1935; Elgar & Jebb 1999; Volkova et al.
1999). The basis for recognition of 2D spiders as prey could be due to a) presence of an
attractant or absence of a repellant on 2D spiders, and/or b) presence of a repellant or
absence of an attractant on 3D spiders. Our chemical manipulation studies demonstrated
that wasps did not differentiate between paper balls coated with 3D spider extracts and
hexane-washed 3D spiders; in addition, they antennated paper balls coated with 2D
spider extract significantly longer than they did hexane-washed 2D spiders, suggesting
that they are responding to the presence of an attractant on 2D spiders, rather than the
presence of a repellant on 3D spiders.
Wasps’ use of visual and chemical cues

38
Mud-dauber wasps seem to rely on visual and chemotactile cues at different stages of
prey location and recognition. Visual cues appear to be used by wasps to locate a
foraging habitat or nesting site (Evans & Eberhard 1971, personal observation). Once a
wasp arrives at a foraging area, however, both visual and chemical cues come into play.
We have observed two scenarios, one beginning with chemical, the other with visual
information, both of which can lead to prey recognition and capture. In the first scenario,
the wasps initially encounter chemical information that influences their subsequent search
behavior. In 29 of the 30 encounters between S. caementarium and spiders (both 2D and
3D) on their webs, the wasps touched the webs before encountering the spiders. The
chemotactile cue then seemed to release different patterns of search behavior: wasps
pursued a prolonged search if they encountered araneid spiders’ silk, but abbreviated the
search upon antennating silk spun by derived araneoid spiders.
In a second scenario, which we call “pounce first, ask questions later”, wasps attack
small, dark-colored objects that are visually contrasting or moving (e.g., a rusty nail,
falling debris, a dark spot on a leaf, a spider silhouetted against a web) (Eberhard 1970;
Blackledge & Pickett 2000; Pasquet et al. 2007; personal observation) and then use
chemotactile information to confirm their identity as suitable prey. In our chemical
manipulation studies, we found that the wasps approached objects (freshly killed spiders,
hexane-washed spiders, and dark spider-sized paper balls), based on visual contrast
against the filter paper on which they were presented, but that their subsequent behaviors
depended on the chemical information they received at the initial encounter. That the

39
wasps antennated paper balls soaked with 2D spider chemicals significantly longer than
they did hexane-washed 2D spiders suggests that chemical cues are more important than
visual or textural cues in prey recognition. However, only two out of 13 wasps stung balls
coated with 2D chemicals, implying that 2D chemicals are necessary but not sufficient
for prey recognition. Lack of contrast or movement sometimes facilitated a spider’s
escape from a hunting wasp, as spiders that dropped from their webs and remained
motionless on the ground were able to ‘disappear’ into the complex background. Some
2D web-building spiders that incorporate prey debris into their webs are able to
camouflage themselves among the prey carcasses and successfully escape wasp
predators, as wasps attack the debris more than the spiders (Chou et al. 2005).
Once a wasp locates a potential prey item, additional factors, including spider age, sex,
morphology, and mass, can influence the forager’s subsequent behaviors (Rau 1928;
Muma & Jeffers 1945; Elgar & Jebb 1999; see also references in O’Neill 2001, Gonzaga
& Vasconcellos-Neto 2005; Polidori et al. 2007).
Anti-predator behaviors of spiders
When attacked by wasps, the majority of 2D spiders (66.6%) dropped off the webs.
These behaviors, as well as vibration of the web by the resident spider, are common
antipredatory responses of araneids (2D spiders) to attack by wasps and other invertebrate
predators (Eberhard 1970; Cloudsley-Thompson 1995; Rayor 1999; Blackledge & Pickett
2000). However, such behaviors did not seem to be particularly effective against S.

40
caementarium, as 60% of those that moved or dropped off were nevertheless caught by
wasps. Because S. caementarium do not recognize 3D spiders as prey, and so do not
attack and pursue these spiders, we cannot describe the responses of 3D spiders to wasp
‘attack’. However, when wasps approached, the majority of 3D spiders (86.6%) remained
on their webs. Although to our knowledge no studies have documented defensive
behaviors of derived araneoids against wasp predators, Latrodectus hesperus
(Theridiidae) actively defend themselves against mice by throwing sticky, viscous silk on
the predator (Vetter 1980).
Why do Sceliphron spp. avoid 3D spiders?
Our behavioral results, as well as those of numerous studies of mud nest contents
(references in Bohart & Menke 1976; Blackledge et al. 2003), demonstrate that
Sceliphron wasps specialize on araneid (2D) spiders, and discriminate against derived
araneoids (3D). We have shown that, on a proximate level, S. caementarium rely on
chemical cues to recognize araneids as suitable prey. On an ultimate level, what might
account for Sceliprhon’s lack of interest in derived araneoid (3D) spiders? We can think
of four possible explanations: 1) toxicity of 3D spiders; 2) physical protection of the
spider by the web; 3) risk of attack by 3D spiders, and 4) historical artifact.
First, it is possible that S. caementarium might avoid derived araneoids (3D) if the spiders
were non-palatable or even poisonous for the developing wasp larvae. This explanation is
unlikely, as another closely related mud-dauber wasp, Chalybion sp., provisions its larval

41
nests with both araneids and derived araneoid spiders. In addition, we have successfully
raised a single S. caemenatrium larva on freshly killed A. tepidariorum spiders, and are
currently investigating S. caementarium’s larval performance on paralyzed 3D spiders.
Second, wasps may find tangled 3D webs difficult to negotiate, and/or 3D webs may
transmit wasp vibrations through silk strands, thus facilitating an early warning of the
attack (Blackledge et al. 2003). Our results outlined above demonstrate that wasps avoid
A. tepidariorum, (as well as three other genera of 3D spiders; Uma, Phelan & Weiss in
prep.), not because of their 3D web architecture, but because of the chemicals present on
the silk and the spiders. It is possible that at one time 3D webs served as defensive
adaptations against wasp predation, but that over time the wasps may have come to
associate chemical cues with web architecture, such that chemical signals now provide
the first available proximate cues by which wasps can assess prey and non-prey.
However, we can take advantage of variation in web architecture seen in an araneid (2D)
species to assess the utility of 3D architecture, independent of chemistry, as a defense
against wasp attack. Mecynogea lemniscata sit in the middle of a three-dimensional
dome-shaped web, but are readily captured by S. caementarium (Chapter 2), suggesting
that three-dimensional web architecture per se is not necessarily a hinderance for wasps.
Third, derived araneoid (3D) spiders may be dangerous to wasps: a small number of our
wasps became entangled in the sticky silk at the base of A. tepidariorum (3D) webs and
were subsequently caught by the spiders. We did not see any wasps caught by 2D
spiders, over an equivalent number of encounters. As mentioned earlier, it is possible that

42
Sceliphron now use chemicals present on the 3D spider web as a proximate cue that
allows them to avoid dangerous 3D spiders.
Fourth, Sceliphron’s avoidance of derived araneoids may be a historical artifact: while
araneids (2D) first evolved in the Jurassic (Selden 1989), sphecid wasps first evolved and
diversified in the early to mid-Cretaceous (Bohart & Menke 1976). However, derived
araneoids (3D) diversified in the late Cretaceous (Penney & Selden 2002). It is possible
that Sceliphron wasps developed the capacity to recognize araneids (2D) as potential prey
because of the spiders’ abundance in the early Cretaceous. Mechanistically, wasps’
antennal receptors (and associated carrier binding proteins) may have evolved to
recognize a certain set of cuticular chemicals that were present on araneid spiders (2D).
If the later-evolving derived araneoids had different cuticular chemicals, the spiders may
have gone unnoticed by Sceliphron spp., though they were perfectly edible prey. On the
other hand, three other sphecid wasp genera prefer derived araneoids over araneid
spiders. It is possible that these wasp genera diversified after the radiation of 3D spiders,
and thus were able evolve the capacity to recognize both araneids and derived araneoid
spiders as potential prey. A better understanding of the timing of sphecid evolution in
relation to spider evolution, as well as a generic level wasp phylogeny will help us to
clarify the evolution of prey preferences in mud-dauber wasps. Both risk of attack by 3D
spiders and the historical artifact scenarios could explain Sceliphron’s lack of interest in
derived araneoids and need further investigation.
Generality of our results

43
We have demonstrated that wasps recognize L. sclopetarius (2D) but not A. tepidariorum
(3D) as prey, and that this recognition is based on spiders’ chemical cues rather than web
architecture. Can we generalize these results across other araneid and derived araneoid
taxa? As mentioned above, the fact that 2D spiders are disproportionately represented in
wasp nests (Blackledge et al. 2003) suggests a general recognition signal. In addition, we
have conducted behavioral observations (Uma, Phelan & Weiss in prep.) that S.
caemantarium readily capture three additional taxa of araneids, and avoid three additional
derived araneoid genera, regardless of web architecture, support the generality of our
behavioral results. Furthermore, as mentioned above, GC/MS analyses of a number of
araneid taxa, including L. sclopetarius, reveal that their cuticular extracts are dominated
by saturated branched hydrocarbons, while those of derived araneoids, including A.
tepidariorum, contain mostly esters of fatty acids (Uma, Phelan & Weiss in preparation).
Thus it seems likely that S. caementarium rely on a common set of chemical cues to
recognize araneids as prey.
Surprisingly, until now the factors involved in prey recognition by spider-hunting sphecid
wasps have rarely been investigated -- perhaps because the web is so prominent and
seemingly important a feature of spider life history that other possibilities had not been
considered. Our results provide the first evidence that spiders’ cuticular cues mediate
prey recognition by sphecid wasps and add to the growing list of studies that document
use of chemical information in prey recognition by predatory and parasitoid wasps (van
Baarlen 1996; Anton & Gnatzy 1998; Weiss 2003; Herzner et al. 2005, Punzo 2006).

44
REFERENCES
Allan, R. A., Capon, R. J., Brown, W.V. & Elgar, M.A. 2002: Mimicry of host cuticular
hydrocarbons by salticid spider Cosmophasis bitaeniata that preys on larvae of
tree ants Oecophylla smaragdina. J. Chem. Ecol. 28, 835-848.
Anton, S. & Gnatzy, W. 1998: Prey specificity and the importance of close-range
chemical cues in prey recognition in the digger wasp, Liris niger. J. Insect.Behav.
11, 671-690.
Bagneres, A.G., Trabalon, M., Blomquist, G. J. & Schulz, S. 1997: Waxes of the social
spider Anelosimus eximius (Araneae, Theridiidae): Abundance of novel n-propyl
esters of long-chain methyl-branched fatty acids. Arch. Insect Biochem. Physiol.
36, 295-314.
van Baarlen, P., Topping, C. J. & Sunderland, K.D. 1996: Host location by Gelis
festinans, an eggsac parasitoid of the linyphiid spider Erigone atra. Entomol. Exp.
Appl. 81, 155-163.
Barth, F.G. 1982: Spiders and vibratory signals: sensory reception and behavioral
significance. In: Spider communication: Mechanisms and ecological significance.
(Witt, P.N. & Rovner, J.S., eds). Princeton University Press, New Jersey. pp. 67–
122.
Blackledge, T. A. & Pickett, K. M. 2000: Predatory interactions between mud-dauber
wasps (Hymenoptera, Sphecidae) and Argiope (Araneae, Araneidae) in captivity.
J. Arachnol. 28, 211-216.

45
Blackledge, T. A., Coddington, J. A. & Gillespie, R. G. 2003: Are three-dimensional
spider webs defensive adaptations? Ecol. Lett. 6,13-18.
Bohart, R. M. & Menke, A. S. 1976: Sphecid Wasps of the World. Berkeley: University
of California Press, California.
Bristowe, W. S. 1941: The Comity of Spiders. The Ray Society, London.
Chou, I. C., Wang, P. H., Shen, P. & Tso, I. M. 2005: A test of prey-attracting and
predator defence functions of prey carcass decorations built by Cyclosa spiders.
Anim. Behav. 69, 1055-1061.
Cloudsley-Thompson, J. L. 1995: A review of the anti-predator devices of spiders. Bull.
Brit. Arach. Soc. 10, 81-96.
Coville, R. E. 1987: Spider-hunting sphecid wasps. In: Ecophysiology of Spiders.
(Nentwig, W., ed). Springer-Verlag, Berli,. pp. 309-318.
Cronin, T. W. 2005: Visual ecology of predator-prey interactions. In: Ecology of
predator-prey interactions (Barbosa, P. & Castellanos, I. eds.) Oxford University
Press, New York, pp. 105-138.
Eberhard, W. G. 1970: The predatory behaviour of two wasps, Agenoideus humils
(Pompilidae) and Sceliphron caementarium (Sphecidae), on the orb weaving
spider Araneus cornatus (Araneidae). Psyche (Cambridge), 77, 243-251.
Elgar, M. A. & Jebb, M. 1999: Nest provisioning in the mud-dauber wasp Sceliphron
laetum (F. Smith): Body mass and taxa specific prey selection. Behaviour 136,
147-159.
Evans, H. E. & Eberhard, M. J. W. 1971: The wasps. Michigan University Press,
Michigan.

46
Griswold, C. E., Coddington, J. A., Hormiga, G., Scharff, N. 1998: Phylogeny of the orb-
web building spiders (Araneae, Orbiculariae: Deinopoidea, Araneoidea). Zool. J.
Linn. Soc. 123, 1-99.
Greene, M. J. & Gordon, D. M. 2003: Social insects -cuticular hydrocarbons inform task
decisions. Nature 423, 32-32.
Greenstone, M. H. & Dickens 2005: The production and appropriation of chemical
signals among plants, herbivores and predators. In: Ecology of predator-prey
interactions. (Barbosa, P. & Castellanos, I. eds.) Oxford University Press, New
York, pp. 139-165.
Gonzaga, M. O. & Vasconcellos-Neto, J. 2005: Orb-web spiders (Araneae:
Araneomorphae; Orbiculariae) captured by hunting-wasps (Hymenoptera:
Sphecidae) in an area of Atlantic Forest in south-eastern Brazil. J. Nat. Hist. 39,
2913-2933.
Gunnarsson, B. 2007: Predation on spiders: Ecological mechanisms and evolutionary
consequences. J. Arachnol. 35: 509-529.
Hartman, C. 1905: Observations on the habits of some solitary wasps of Texas. Bull.
Univ. Texas, Scientific Series 7, 65, 1–72.
Herzner, G., Schmitt, T., Linsenmair, K., Strohm, E. 2005: Prey recognition by females
of the European beewolf and its potential for a sensory trap. Anim. Behav. 70,
1411-1418.
Matsuda, H., Hori, M. & Abrams, P. A. 1996: Effects of predator-specific defense on
biodiversity and community complexity in two-trophic-level communities. Evol.
Ecol. 10, 13–28.

47
Muma, M. H. & Jeffers, W. F. 1945: Studies of the spider prey of several mud-dauber
wasps. Ann. Entomol. Soc. Am. 38, 245-255.
Nelson M. E. & MacIver, M. A. 1999: Prey capture in the weakly electric fish
Apteronotus albifrons: sensory acquisition strategies and electrosensory
consequences. J. Exp. Biol. 202, 1195-1203.
Nentwig, W. & Heimer, S. 1987: Ecological aspects of spider webs. In: Ecophysiology of
Spiders. (Nentwig, W., ed). Springer-Verlag, Berlin, pp. 211-228.
O'Neill, K. M. 2001: Solitary Wasps: Natural History and Behaviour. Cornell University
Press, New York.
Papke, M. D. 2000: Isolierung, identifizierung und synthese von signalstoffen aus
spinnen. Ph.D. thesis, University of Hamburg, Germany.
Pasquet, A., Cardot, J. & Lenorgne, R. 2007: Wasp attacks and spider defense in the orb
weaving species Zygiella x-notata. J. Insect Behav. 20, 553-564.
Platnick, N. I. 2008. The World Spider Catalog, Version 8.5. American Museum of
Natural History, online at
http://research.amnh.org/entomology/spiders/catalog/index.html.
Polis, G. A., Myers, C. A. & Holt, R. D. 1989: The ecology and evolution of intraguild
predation: Potential competitors that eat each other. Ann. Rev. Ecol. Syst. 20,
297–330.
Polidori, C., Federici, M., Pesarini, C. & Andrietti, F. 2007. Factors affecting spider prey
selection by Sceliphron mud-dauber wasps (Hymenoptera: Sphecidae) in northern
Italy. Anim. Biol. 57, 11-28.

48
Punzo, F. & Ludwig, L. 2005: Behavioral responses to Pepsis thisbe (Hymenoptera:
Pompilidae) to chemosensory cues associated with host spiders. J.Insec.Behav.
18, 757-766.
Punzo, F. 2006: Female spider wasps, Anoplius splendens Driesbach (Hymenoptera:
Pompilidae), learn to associate the odor of host feces with the presence of the
host. J. Entomol. Science. 41, 202-210.
Rau, P. 1935: The spider prey of the mud wasp, Sceliphron caementarium (Araneae,
Hymen: Sphegidae). Entomol. News. 46, 267–270.
Raveret Richter, M. 2000: Social wasp (Hymenoptera: Vespidae) foraging behavior. Ann.
Rev. Entomol. 45, 121–150.
Rayor, L. S. 1996. Attack strategies of predatory wasps (Hymenoptera: Pompilidae;
Sphecidae) on colonial orb web-building spiders (Araneidae: Metepeira
incrassata). J. Kansas Entomol. Soc. 69, 67-75.
Roux, O., Gers, C., Tene-Ghomsi, J. N., Arvanitakis, L., Bordat, D. & Legal, L. 2007:
Chemical characterization of contact semiochemicals for host-recognition and
host-acceptance by the specialist parasitoid Cotesia plutellae (Kurdjumov).
Chemoecology.17, 13-18.
Singer, T. L. 1998: Roles of hydrocarbons in the recognition systems of insects. Am.
Zool. 38, 394-405.
Schulz, S. 2004: Semiochemistry of spiders. In: Advances in Insect Chemical Ecology
(Carde, R.T. & Miller, J.G. eds). Cambridge University Press, Cambridge, pp.
110-150.

49
Schmitz, O. J. 2008: Effects of predator hunting mode on grassland ecosystem function.
Science. 319, 952-954.
Sih, A.1985: Predation, competition, and prey communities: a review of field
experiments. Ann. Rev. Ecol. Syst. 16, 269–311.
Steidle, J. L. M. & Ruther, J. 2000: Chemicals used for host recognition by the granary
weevil parasitoid Lariophagus distinguendus. J. Chem. Ecol. 26, 2665-2675.
Steidle, J. L. M., Lanka, J., Muller, C. & Ruther, J. 2001: The use of general foraging
kairomones in a generalist parasitoid. Oikos. 95, 78-86.
Surlykke, A. & Moss, C. F. 2000: Echolocation behavior of big brown bats, Eptesicus
fuscus, in the field and the laboratory. J.Acoust.Soc.Am. 108, 2419-2429.
Vetter, R. S. 1980: Defensive behavior of black widow spider Latrodectus hesperus
(Araneae: Theridiidae). Behav. Ecol. Sociobiol. 7, 187-193.
Vet, L. E. M. & Dicke, M. 1992: Ecology of infochemical use by natural enemies in a
tritrophic context. Ann. Rev. Entomol. 37, 141-172.
Vet, L.E. M., Lewis, W. J. & Carde, R. T. 1995: Parasitoid foraging and learning. In:
Chemical Ecology of Insects, 2nd
edn. (Bell, W. & Cardé, R. T., eds).Chapman &
Hall, New York, pp. 65-101.
Vinson, S. B. 1976: Host selection by insect parasitoids. Annu. Rev. Entomol. 21, 109–
133.
Volkova, T., Matthews, R. W. & Barber, M. C. 1999: Spider prey of two mud dauber
wasps (Hymenoptera: Sphecidae) nesting in Georgia's Okefenokee Swamp. J.
Entomol. Sci. 34, 322–327.

50
Weiss, M. R. 2003: Good housekeeping: why do shelter-dwelling caterpillars fling their
frass? Ecol. Lett. 6, 361-370.
Zar, J. H. 1999: Biostatistical analysis, 4th
edn. Prentice Hall, New Jersey.

51
Fig.1. Mud-dauber wasp Sceliphron caementarium and its nest contents. 1a. S.
caementarium capturing a two-dimensional web-building spider on the web. 1b. Sphecid
wasp larva feeding on a paralyzed two-dimensional web-building spider. 1c. Paralyzed
spiders found within a mud nest.
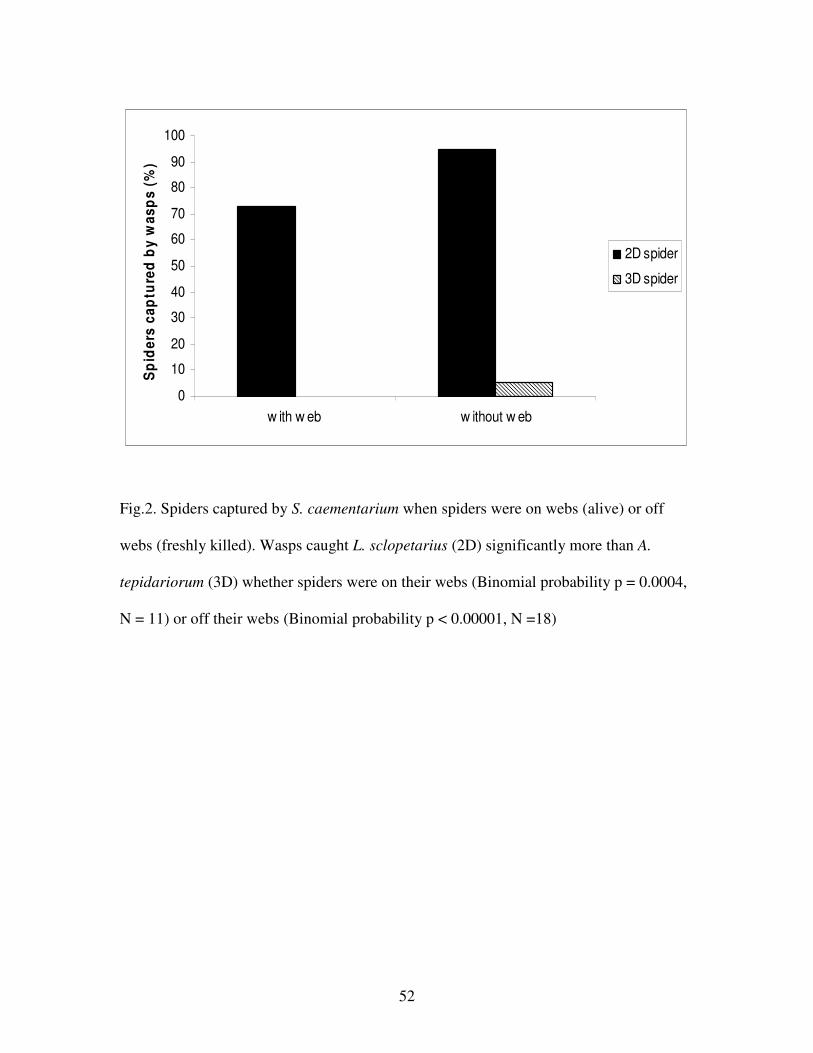
52
Fig.2. Spiders captured by S. caementarium when spiders were on webs (alive) or off
webs (freshly killed). Wasps caught L. sclopetarius (2D) significantly more than A.
tepidariorum (3D) whether spiders were on their webs (Binomial probability p = 0.0004,
N = 11) or off their webs (Binomial probability p < 0.00001, N =18)
0
10
20
30
40
50
60
70
80
90
100
w ith w eb w ithout w eb
Sp
ide
rs c
ap
ture
d b
y w
as
ps
(%
)
2D spider
3D spider
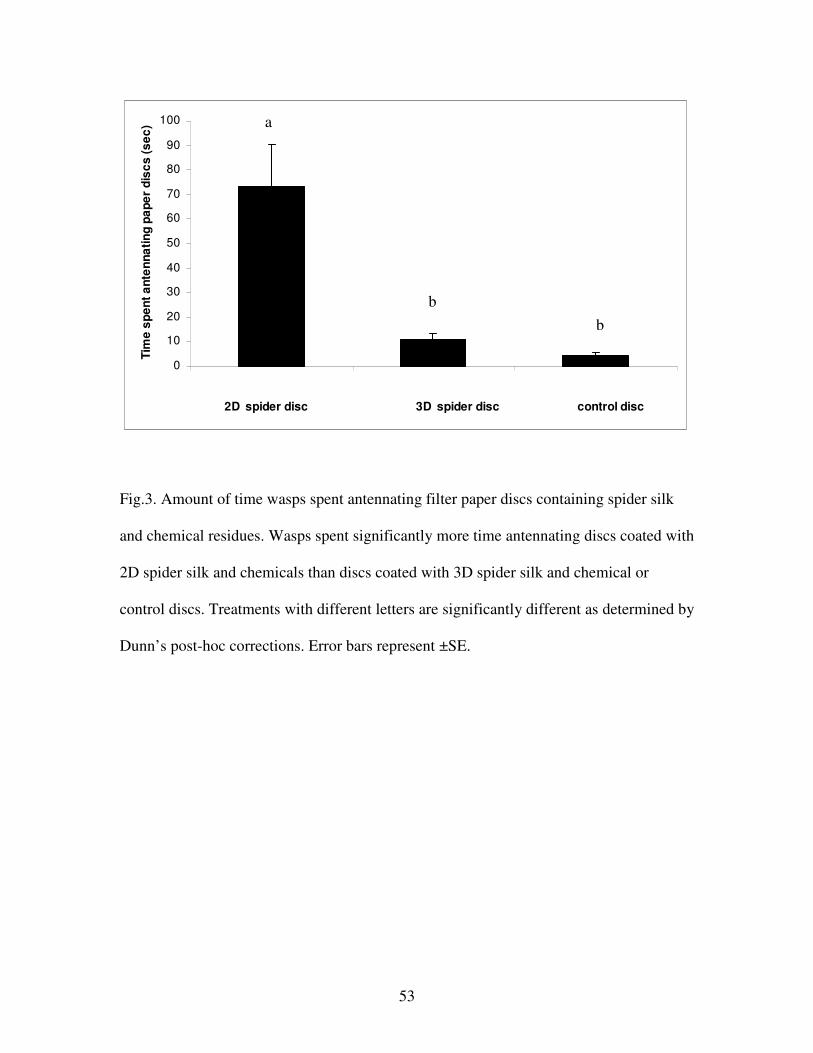
53
0
10
20
30
40
50
60
70
80
90
100
1 2 3
2D spider disc 3D spider disc control disc
Tim
e s
pe
nt
an
ten
na
tin
g p
ap
er
dis
cs
(s
ec
)
Fig.3. Amount of time wasps spent antennating filter paper discs containing spider silk
and chemical residues. Wasps spent significantly more time antennating discs coated with
2D spider silk and chemicals than discs coated with 3D spider silk and chemical or
control discs. Treatments with different letters are significantly different as determined by
Dunn’s post-hoc corrections. Error bars represent ±SE.
a
b
b
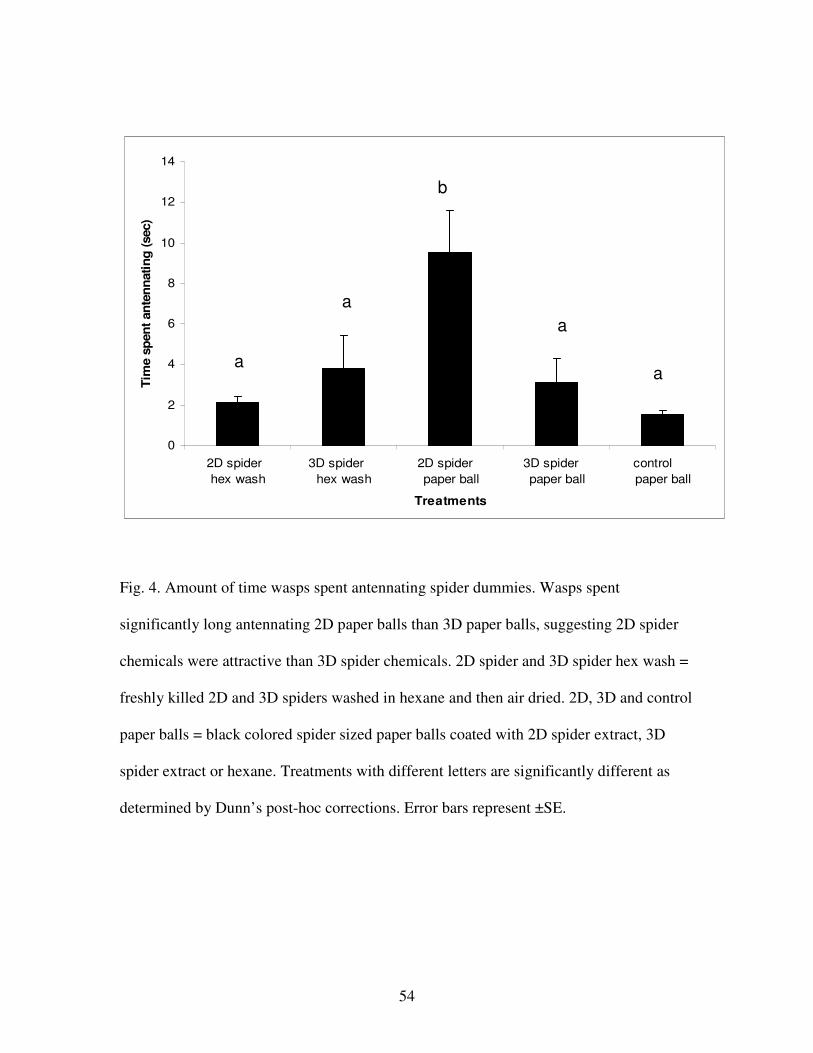
54
0
2
4
6
8
10
12
14
2D spider
hex wash
3D spider
hex wash
2D spider
paper ball
3D spider
paper ball
control
paper ball
Treatments
Tim
e s
pen
t an
ten
nati
ng
(sec)
Fig. 4. Amount of time wasps spent antennating spider dummies. Wasps spent
significantly long antennating 2D paper balls than 3D paper balls, suggesting 2D spider
chemicals were attractive than 3D spider chemicals. 2D spider and 3D spider hex wash =
freshly killed 2D and 3D spiders washed in hexane and then air dried. 2D, 3D and control
paper balls = black colored spider sized paper balls coated with 2D spider extract, 3D
spider extract or hexane. Treatments with different letters are significantly different as
determined by Dunn’s post-hoc corrections. Error bars represent ±SE.
b
a
a
a
a

55
Chapter 1 Appendix
Chemical basis of wasps’ preference of araneids over derived araneoids
Wasps’ preference for araneids over derived araneoids may be due to the presence of
stimulants or absence of deterrent chemicals on araneids; and/or the presence of
deterrents or absence of stimulant chemicals on derived araneoids. We used Larinioides
sclopetarius and Achaearanea tepidariorum as representative araneid (2D) and derived
araneoid (3D) taxa. We presented individual wasps with chemically treated Larinioides
(henceforth 2D spider) and Achaearanea (henceforth 3D spider) spiders in a fully
randomized sequence: a) hexane-washed 2D spider coated with 3D spider extract (looks
like 2D, but smells like 3D), b) hexane-washed 3D spider coated with 2D spider extract
(looks like 3D, but smells like 2D), c) hexane-washed 2D spider (looks like 2D, but has
no spider ordor), d) hexane-washed 3D spider (looks like 3D, but has no spider odor), e)
freshily-killed 2D spider (looks and smells like 2D spider) and f) freshly killed 3d spider
(looks and smells like 3D spider).
To remove chemicals from spiders’ cuticle, we washed each freshly-frozen spider twice
in 1.5 ml of ACG-grade hexane for 15 minutes. The spider thus treated lacked cuticular
chemicals, but remained otherwise intact. To obtain 2D spiders with 3D extracts, we
soaked hexane-washed 2D spiders in 1.5 ml of 3D extract and allowed hexane to
evaporate overnight. Similarly, to obtain 3D spiders with 2D extract, we soaked hexane-

56
washed 3D spiders in 1.5 ml of 2D extract and allowed hexane to evaporate overnight. To
obtain freshly killed 2D and 3D spiders, we simply killed the spiders by placing them in
the freezer for 5 mins before an experiment.
Spiders were placed one at a time on 9cm diameter paper discs. The paper disc itself was
placed at the base of an empty wooden frame, where wasps came down to search for
spiders. (The wasps used here were from an earlier experiment, in which they had been
trained to come down to the wooden frame). Wasp responses to spiders including time
spent antennating, and stinging/not stinging the spiders were recorded for two minutes.
Antennation time was considered as a proxy for interest in prey. Typically, once a wasp
recognizes a spider as a potential prey (whether alive or freshily killed), it immediately
captures and stings the spider usually within a couple of seconds. Presentations to the
wasps of any two treatments were separated by a 10 minute interval, and each wasp
participated in all six treatments within one to two days.
To examine whether wasps treated hexane-washed 3D spiders with 2D extracts similarly
as they treated freshly killed 2D spiders, we recorded wasps’ response (sting/no sting)
towards these treatments. Hexane-washed 2D spiders that lacked cuticular chemicals
were used as a control.
We carried out three pre-planned comparisons in order to determine the chemical basis of
prey specificity by S. caementarium. To examine whether 2D spiders were captured by
wasps due to the presence of stimulant chemicals (or absence of repellant), we compared

57
the antennation time of wasps on 1) hexane-washed 3D spiders coated with 2D spider
extract vs. hexane-washed 3D spiders. Similarly, to examine whether 3D spiders lack
stimulant or possess a repellant, we compared antennation time of wasps on 2) hexane-
washed 2D spider coated with 3D spider extract vs. hexane-washed 2D spiders. To
examine wasp response towards spiders lacking cuticular chemicals, we compared
antennation time of wasps on 3) hexane-washed 2D spiders vs. hexane-washed 3D
spiders. We used Friedman nonparametric ANOVA followed by Dunn’s Bonferroni post-
hoc correction to determine whether antennation time differed significantly between
treatments.
RESULTS
2D spiders have stimulant chemicals
While wasps readily stung all nine freshly killed 2D spiders, they did not sting any
hexane-washed 2D spiders, suggesting that cuticular chemical cue is essential for prey
recognition (Fig. 1a). Wasps stung four of the nine 3D spiders coated with 2D extract,
suggesting that these spiders had ‘prey recognition’ chemicals on their cuticle (Fig 1b).
Though these four spiders were stung, wasps antennated them from 1 to 11 seconds. This
was in contrast to wasps’ antennating freshly killed spiders for just 1 sec before stinging
them. The difference in antennation time may reflect differences in chemical profile or
morphology of the treated spiders: the chemical compostion of the extract is unlikely to

58
mimic exactly the chemical profile of an intact spider, and the morphology of the spiders
changes somewhat after they have soaked in hexane.
Wasps (n=9) antennated hexane-washed 3D spider coated with 2D extract longer than
they antennated hexane-washed 3D spider, hexane-washed 2D spider with 3D extract and
hexane-washed 2D spider (Fig 1b, Friedman ANOVA F= 12.60, p= 0.0056). They did
not differentiate between hexane-washed 2D spiders and hexane-washed 2D spiders with
3D extract (Dunn’s Bonferroni test p>0.05), suggesting that 3D spiders do not have a
repellant compound that dissuades wasps. However, wasps antennated hexane-washed
3D spider with 2D extract significantly more than hexane-washed 3D spiders (Fig 1b,
Dunn’s Bonferroni test p<0.05), suggesting that the 2D spiders have a blend of
compounds that may serve as stimulants. Wasps did not differentiate between hexane-
washed 2D and 3D spiders (Dunn’s Bonferroni test p>0.05), indicating the importance of
chemical cues in prey recognition.
Thus we believe that wasps recognize 2D spiders as potential prey as these spiders have
stimulant chemicals on their cuticles, and ignore 3D spiders because they lack the
stimulant rather than have repellant chemicals on their cuticles.
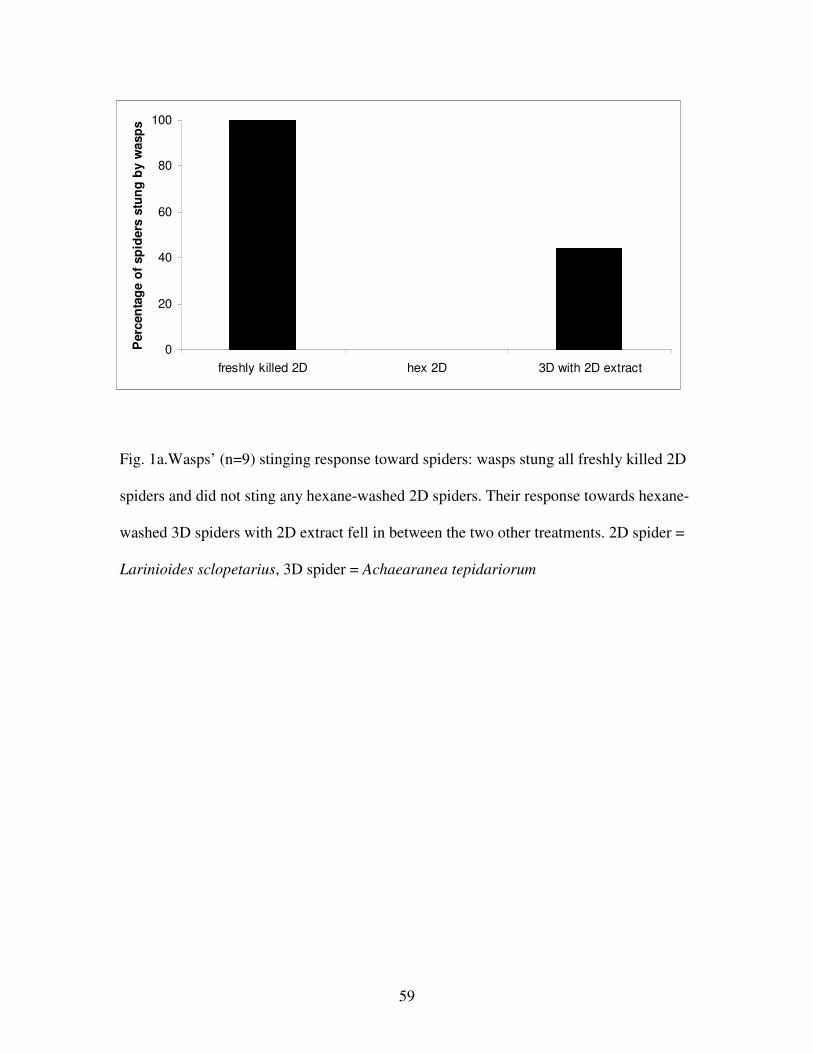
59
0
20
40
60
80
100
freshly killed 2D hex 2D 3D with 2D extract
Perc
en
tag
e o
f sp
iders
stu
ng
by w
asp
s
Fig. 1a.Wasps’ (n=9) stinging response toward spiders: wasps stung all freshly killed 2D
spiders and did not sting any hexane-washed 2D spiders. Their response towards hexane-
washed 3D spiders with 2D extract fell in between the two other treatments. 2D spider =
Larinioides sclopetarius, 3D spider = Achaearanea tepidariorum
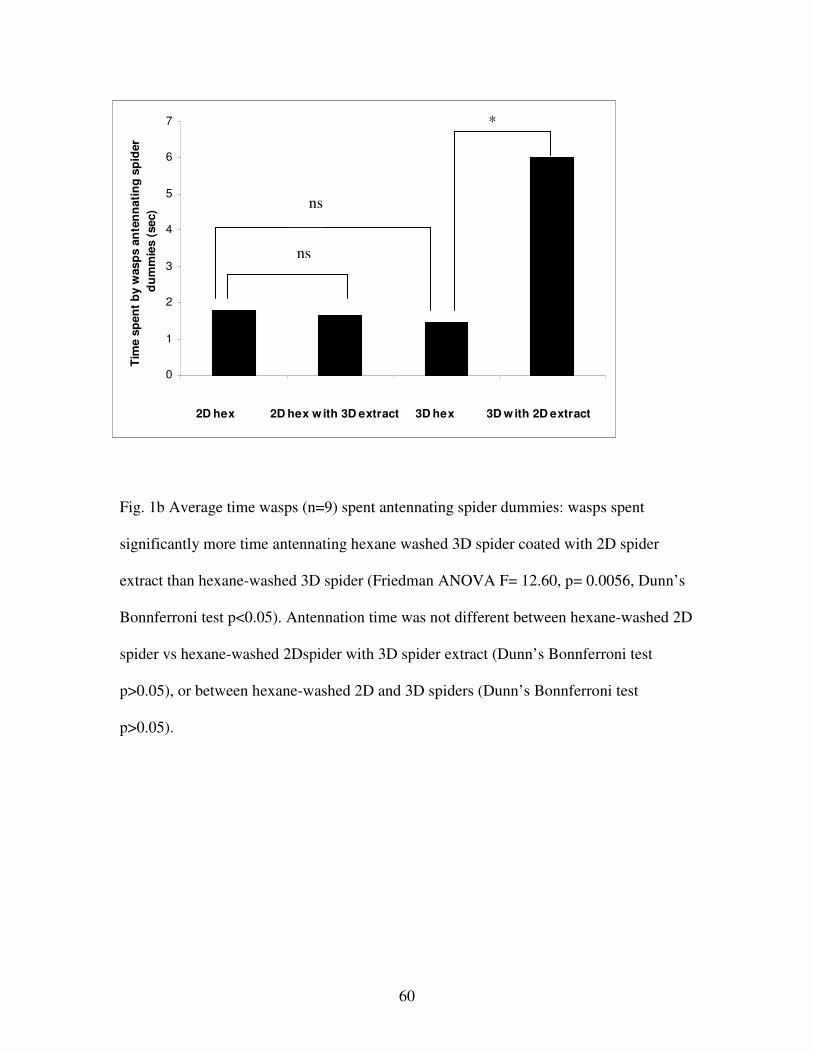
60
0
1
2
3
4
5
6
7
1 2 3 4
2D hex 2D hex w ith 3D extract 3D hex 3D w ith 2D extract
Tim
e s
pen
t b
y w
asp
s a
nte
nn
ati
ng
sp
ider
du
mm
ies (
sec)
Fig. 1b Average time wasps (n=9) spent antennating spider dummies: wasps spent
significantly more time antennating hexane washed 3D spider coated with 2D spider
extract than hexane-washed 3D spider (Friedman ANOVA F= 12.60, p= 0.0056, Dunn’s
Bonnferroni test p<0.05). Antennation time was not different between hexane-washed 2D
spider vs hexane-washed 2Dspider with 3D spider extract (Dunn’s Bonnferroni test
p>0.05), or between hexane-washed 2D and 3D spiders (Dunn’s Bonnferroni test
p>0.05).
*
ns
ns

61
Chapter 2
Spider chemistry, web architecture and behavior: factors influencing prey capture
success by sphecid wasps
INTRODUCTION
For both vertebrate and invertebrate animals, chemical cues play an important role in
mediation of many activities, including foraging (Nevitt 2000), mate location (Gaskett
2007, Milinski et al. 2005), and localization and recognition of both prey and enemies
(Conover 2007, Persons & Rypstra 2000, Persons & Uetz 1996, Punzo 2006, Dicke &
Grostal 2001).
Potential prey subjects can use a number of means to localize (and try to avoid) their
predators. In addition to making use of visual, vibrational and auditory information, prey
can also use chemical cues recognizing not only those that are directly produced by
predators (Kats & Dill 1998), but also alarm chemicals produced by both conspecifics
and heterospecific prey (Chivers et al. 1996, Kats & Dill 1998, Chivers & Mirza 2001,
Kortet & Hedrick 2004, Schoeppner & Relyea 2005, Conover 2007).
Just as prey can use chemical information to avoid predators, predators can also make use
of volatile or contact chemical cues to locate prey (Vinson 1976, Weiss 2003, Herzner et
al. 2005, Punzo 2006). Less is known about the response of generalist predators towards

62
chemically-related prey groups. The present study examines whether the mud-dauber
wasp, Sceliphron caementarium, a spider-hunting invertebrate predator, chooses its prey
based on chemical similarity of the spider taxa.
Six genera of mud-dauber wasps (Sphecidae) hunt spiders (both web-building and non-
web-building) exclusively, and provision their mud nests with paralyzed spiders as food
for their developing larvae (Coville 1987). Examining the contents of wasp nests suggests
that these wasps preferentially capture spiders in the family Araneidae, which typically
build two-dimensional webs (2D spiders), over derived araneoid spiders belonging to
several families, most of which typically build three-dimensional webs (3D spiders),
though the latter are more abundant in areas where wasps forage (Blackledge et al.2003).
Why might mud-dauber wasps specialize on araneids? Blackledge et al. (2003)
hypothesized that araneids, which typically sit exposed on their 2D webs, may be more
vulnerable to wasp predation than are derived araneoids, which sit within a tangled
matrix of web. They further speculated that wasp predation may have favored the
evolution of 3D web-building spiders from their ancestral 2D web-building spiders.
A recent study demonstrates that Sceliphron caementarium (Sphecidae) wasps use
chemicals present on spiders’ cuticle and silk to distinguish araneid from derived
araneoid spiders, regardless of their web architecture (Uma & Weiss 2010). While wasps
pursued and readily captured Larinioides sclopetarius (araneid), they rarely followed and
captured Achaearanea tepidariorum (derived ananeoid) with or without their webs. Thus

63
chemicals present on spider silk and cuticles may serve as proximate cues that allow S.
caemantarium to differentiate between prey and non-prey spiders. Not only did the
wasps’ behavior towards L. sclopeterius and A. tepidariorum differ, but the two spiders’
defensive responses towards wasps were also different (Uma & Weiss 2010): when
approached by a foraging wasp, L. sclopetarius often moved to the periphery or dropped
off their 2D webs, whereas A. tepidariorum often remained on their 3D webs.
Based on the examination of nest contents, S. caementarium is known to capture spiders
in a number of other araneid genera with varying web architecture, as well as spiders in
some non-web building families (reviewed by Blackledge et al. 2003). In this study, we
sampled spiders with varied web architectures belonging to multiple genera within
Araneidae and four derived araneoid families to examine the extent to which spiders’
cuticular chemistry, web architecture and antipredator behavior influence prey capture by
wasps. Specifically, by conducting chemical and behavioral assays, we asked 1a) do all
spiders captured by wasps have similar cuticular chemistry? and 1b) are all spiders that
share that cuticular chemistry acceptable to wasps? In addition, we took advantage of the
diversity of web architecture within spider families in an attempt to uncouple the relative
importance of taxonomic affiliation and web architecture. We asked, 2) do wasps capture
spiders with certain taxonomic affiliation (i.e., capture araneids and do not capture
derived araneoids etc) regardless of their web architecture? Finally, we also examined
whether 3) anti-predatory behaviors of spiders differ based on taxonomic affiliation.
METHODS

64
1. a, b. Do all spiders captured by wasps have similar chemical profiles? Are all
spiders that share that cuticular chemistry acceptable to wasps?
To examine whether all spiders that are captured by wasps share similar chemical
profiles, we examined the cuticular extracts of spiders, both web-building (araneids,
derived araneoids, uloborids), and non-web-building (salticids and thomisids), that are
found in the habitats where wasps forage. (See Fig.1 for spider genera and their
phylogenetic relationship). Among the web-building spiders we analyzed four araneids
(total number of spiders = 13; Larinioides sclopetarius n=3, Mecynogea lemniscata n=3,
Micrathena gracilis n=5, Metepeira labyrinthea n=2); five derived araneoids in three
families (total number of spiders = 21) (Achaearanea tepidariorum (Theridiidae)n=4,
Steatoda triangulosa (Theridiidae) n=5, Frontinella sp. (Linyphidae) n=4, Tetragnatha
sp. (Tetragnathidae) n=4, Leucauge sp. (Tetragnathidae) n=4); and two uloborids (total
number of spiders n = 5)(Uloborus sp.(Uloboridae) n=2, Hyptiotes sp.(Uloboridae) n=3).
Among the non-web-building spiders, we analyzed the cuticular extracts of Phiippus sp.
(Salticidae n=3), and Misumena sp. (Thomisidae n=2). Adult and subadult spiders were
collected by visual searches, and by using ‘beat sheet’ technique around the Washington
D.C. area from June to September 2006-2009, and from Bath Nature Preserve, Akron,
Ohio in July 2008.
Spiders were killed by freezing for 15 mins @ -4°C. Spiders’ cuticular chemicals were
extracted by soaking individuals in 1.5 ml of HPLC-grade hexane for15 min at room

65
temperature. After removal of the spiders, the extracts were stored at –20°C in glass vials
with Teflon-lined lids until ready for analysis. For chemical characterization, 2µl of
extract was injected into an Agilent Technologies 6890 gas chromatograph interfaced to a
5973 mass selective detector (GC/MS), using a splitless injection at 300°C onto a ZB1ms
column (30mx0.25mm) and a temperature program of 100°C-320°C @ 30°C/min.
Chemical constituents were identified by computer matching EI mass spectra against the
2005 NIST database of ca. 200,000 spectra.
Data analysis
Chemicals present on spiders’ cuticle were analyzed by multivariate methods. To
consider a chromatographic peak characteristic of a genus (due to our limited sampling,
each genus was represented by a single species), at least half of the individuals within a
species had to posses the corresponding peak. (e.g., if there were only two individuals in
a species, then both of them had to possess the corresponding peak). In addition, only
hydrocarbons with carbon chain length C21 or more were included as these are
considered to be typical insect and spider cuticular compounds (Lockey 1988). Peaks that
did not match either of the above criteria were excluded from the analysis. Since relative
peak areas of a sample are not statistically independent, we transformed the data
according to Aitchison’s method (Aitchison 1959). Analysis of similarity (ANOSIM) was
used to test significance between groups based on chemical similarities by using
DECODA software (Minchin 2005). We grouped araneids, salticids, thomisids and
uloborids together in one group based on the chromatographs obtained from GCMS and

66
with prior knowledge of wasp behavior. We grouped derived araneoids (tetragnathids,
theridiids, linyphids) as a separate group based on prior knowledge of wasp behavior. We
used non-metric-multidimensional scaling (NMDS) to visualize the chemical distances of
different spider taxa.
2. Do wasps capture spiders based on taxonomic affiliation regardless of their web
architecture?
Our previous study on prey choice by S. caementarium demonstrated that the wasps
readily capture Larinioides sclopetarius (araneid) but do not capture Achaearanea
tepidariorum (derived araneoid) spiders irrespective of their web-architecture (Uma &
Weiss 2010). To assess wasps’ response to other spider taxa we offered S. caementarium
spiders belonging to four additional araneid and three derived araneoid taxa, as well as
Uloborus sp., spiders basal to both araneid and derived araneoid groups. We took
advantage of the natural diversity of web architecture within spider families in an attempt
to uncouple the relative importance of taxonomic affiliation and web architecture.
Spiders tested in the family Araneidae included two genera that spin characteristic 2D
webs (Neoscona sp., Micrathena gracilis), as well as two genera that spin 3D webs
(Mecynogea lemniscata, Metepeira labyrinthea). M. lemniscata, build and rest under a
3D dome-shaped web, and M. labyrinthea attach a debris-covered 3D retreat to their 2D
web. Derived araneoids included spiders that typically construct 3D webs (Frontinella
sp., (Linyphiidae) and Steatoda sp., (Theridiidae)), and those that retain the ancestral 2D

67
webs (Tetragnatha sp., (Tetragnathidae)). We also tested Uloborus glomosus
(Uloboridae), sister to both araneids and derived araneoids, which spins non-sticky 2D
webs composed of highly coiled cribellate silk that differs from the ecribellate, sticky
silk, characteristic of araneid webs.
Spiders selected for the trials were found in the habitats where Sceliphron caementarium
commonly forage. We collected adult and subadult spiders by visual search around the
Washington DC area from June to September 2005 to 2009. Neoscona, Frontinella, and
Steatoda were allowed to build webs in square wooden frames (25x25cm²x 8 cm deep).
Frontinella were provided with a forked twig inside their wooden cages, for structural
support on which to build their webs. Mecynogea, Metepeira, Tetragnatha and Uloborus
characteristically build large webs and so were provided with larger wooden frames
(30x30cmx12cm deep). Plastic wrap prevented the spiders from escaping from the
wooden frames. Once a week we fed the spiders 5-10 Drosophila or one subadult cricket,
and misted their webs to provide moisture. Micrathena did not spin webs within wooden
frames, but readily built webs in the outdoor mesh field cages (2m3). We placed only one
Micrathena in a field cage at any given point so that we did not test the same spider more
than once. Field cages contained a variety of plants and grasses that served as places
where spiders could build webs, and hide from wasps.
Female wasps were collected as they gathered mud along the George Washington
Memorial Parkway near metropolitan Washington, D.C., and from the Georgetown
University campus during the months of June – September, from 2005 through 2009.

68
About five to six wasps were released per field cages (2m³), each of which were
provisioned with wet mud for nest construction and contained three or four wooden
shelters, as wasps prefer to build their nests in sites protected from rain. Because mud-
gathering and nest construction always precede spider-hunting (Evans & Eberhard 1971),
the caged wasps were motivated to hunt once they had built a mud cell.
Once a wasp had built a mud cell, we lured her down to a ‘testing arena’ (a stool,
29x32x40cm, located in the centre of the cage): we held in front of her a 2D web-
building spider (Larinioides sp.) within its wooden frame and then slowly moved the
frame down to the stool. After the wasp touched the silk, she generally followed the
frame down to the testing arena, at which point she began to investigate the spider and its
web. We considered a successful training session one in which the wasp captured the
spider, either on or off its web. After one to three successful training sessions, the
foraging wasp developed an association between the testing arena and availability of a
spider, and would spontaneously come down to the testing arena when a wooden frame
was placed there.
We opportunistically presented foraging wasps with spiders that were available at the
time of testing. Not all spider taxa were available all the time: for example, Mecynogea
were abundant early in the season (June-July), but, Micrathena were available during late
summer (July-Aug). Individual wasps trained to come down to the testing arena were
offered one to eight different spider species belonging to araneid, derived araneoid and
uloborid taxa over the course of 4-5 days. However, a single wasp was presented with
only one individual of a particular spider species. Each trial, which consisted of an

69
individual wasp encountering a particular spider species, commenced when a wasp first
antennated the web and lasted until the wasp successfully captured the spider or until 15
min had elapsed, whichever happened first. We examined whether identity of spiders
and/or wasps affected the outcome of wasp response by using logistic regression (SPSS
v. 18).
3. Do antipredator behaviors of spiders differ based on taxonomic affiliation?
Besides examining wasps’ responses towards spiders, we also recorded spiders’
responses towards wasp attacks. Spiders’ responses included moving to the periphery of
their web or dropping off the web or resting on the web. We used logistic regression to
determine whether taxonomic affiliation (i.e. araneid, derived araneoid or uloborid), web
architecture (2D/3D) or acceptability to wasps (captured or not captured) correlated with
antipredator behavior.
RESULTS
1. All spiders captured by wasps have similar chemical profiles; but not all spiders
with that similar chemical profile are captured by wasps.
We identified a total of 60 hydrocarbons (HC) among web-building as well as non-web-
building spiders. GC/MS revealed unbranched and branched HC, as well as HC with
different functional groups (ester, ether) on the cuticles of all the spiders. Cuticular

70
extracts of the four araneid genera contained 12 to 21 compounds, the majority being
branched HCs (average amount of branched HCs present on four araneid genera: mean ±
SE 71.5%±5.8). Uloborus and Hyptiotis, the cribellate silk spiders, had 4-10 compounds
on their cuticles, and, similar to the araneids, predominantly contained branched
hydrocarbons (average amount of branched HC present on two uloborid genera 60.9 ±
30.3%). The cuticles of two non-web-building spiders Phidippus and Misumena had 14
and 11 compounds respectively (average amount of HCs present on the non-web builders
44.7 ± 6.9%). Cuticular extracts of derived araneoids, on the other hand, contained 5 to
20 compounds, which, in contrast with the rest of the spider taxa, were largely HC with
functional groups such as esters and ethers (average amount of compound present on
three genera 78.1 ± 8.5%). A typical araneid and a derived araneoid profile are
represented in Fig 5a and 5b. Chemical profiles of spiders that are captured by wasps in
three different families (araneid, salticid and thomisid) are represented in Fig 5c.
Across all types of cuticular compounds, the maximum carbon chain length increased
from basal to derived spider groups (Fig 1). For example, Uloborus and Hyptiotes
contained cuticular compounds with carbon chain length of C21-C29; their sister group
Araneidae had compounds as long as C33 (C21-C33) and derived araneoids had
compounds up to C40 (C21-C40). The cuticular carbon chain length of non-web-building
spiders (salticids and thomisids) ranged between C21-C29.
The two groups (group1: araneids, uloborids, salticids and thomisids; group 2: derived
araneoids), separated based on their chemical profiles, were significantly different as

71
visualized by NMDS (Fig 2, ANOSIM: R =0.51 P<0.00001). Spiders whose cuticular
profiles were dominated by branched hydrocarbons were represented with pink symbols;
those that were dominated by hydrocarbons with esters and ethers as their functional
groups were given blue symbols. We further coded spiders with respect to whether or not
they were taken by wasps; those that were captured by wasps were represented with
squares and those that were not captured were represented as triangles (Fig 2). Thus pink
squares represent spiders whose cuticular profiles were dominated with branched
hydrocarbons and were captured by wasps; pink triangles represent spiders whose
cuticular profiles were dominated with branched hydrocarbons, but were not captured by
wasps; blue triangles represent spiders whose cuticular profiles were dominated with
esters of fatty acids and were not captured by wasps. This suggests that cuticular
chemistry is necessary for recognizing a particular spider as a potential prey, but not
sufficient for its capture.
2. Wasps capture spiders based on taxonomic affiliation regardless of their web
architecture
Wasps (n=31) captured spiders in three of four araneid genera regardless of their web
architecture. Of the taxa that spin 2D webs, wasps readily captured individuals of
Neoscona (55.5%) (Fig.1), but did not capture any of the six Micrathena (0%), though
these spiders sat right in the middle of a 2D web and had cuticular chemistry similar to
the rest of the araneids. With respect to araneids that incorporated a 3D component into
their webs, nine of 11 Mecynogea (81.81%) and seven of 16 Metepeira (43.75%) were

72
nonetheless caught. In contrast, wasps did not capture any spiders belonging to the three
derived araneoid genera, including Tetragnatha, which built 2D webs. Wasps did not
capture any uloborids, though their chemical profiles were similar to those of araneids
(Fig. 3). And they make an araneid-like 2D web. Spider genus was a significant predictor
of the outcome of wasp responses (logistic regression: spider genus X² = 12.45, df =1,
P<0.0001), but the identity of individual wasps was not (wasp ID X²= 1.52, df =1, P <
0.218).
3. Antipredatory behavior of spiders differed based on taxonomic affiliation
Antipredatory behavior of different groups of spiders were markedly different: upon
wasp attack araneids mostly moved to the periphery and/or dropped off their webs, but
derived araneiods and uloborids rarely moved away from their resting positions, and
never dropped off their webs (Fig 4). Taxonomic affiliation (araneid or derived araneoid
or uloborid) was a significant predictor of spiders’ antipredatory behavior (Logistic
regression: X² =4.65, df = 1, P= 0.03) but web architecture (Logistic regression: X² =
1.62, df =1, P=.202) and acceptability to wasps (captured/not captured) were not
(Logistic regression: X² = 2.74, df =1, P=0.09).
Though most araneids (27 of 36, 75%) moved to the periphery and/or dropped off their
webs when attacked by a wasp, this behavior did not ensure escape; indeed 21 of the 27
araneids that moved were nonetheless caught by wasps. Of the nine araneids that did not
move or drop off the webs, six were Micrathena, the spiny orb web spider that was not

73
taken by the wasps. The remaining three were individuals of Neoscona sp. (n=1), and
Mecynogea (n=2) that were caught right away, before the spiders had a chance to move
or drop down. On the other hand, though wasps landed on the webs of derived araneoids
and uloborids, the spiders were not pursued or caught by wasps, and these spiders rarely
moved or dropped down from their webs: Whereas spiders in two derived araneoid taxa
Tetragnatha and Frontinella, remained on their webs and did not move upon wasps’
approach, three of eight Steatoda sp. came down from their webs by dragline silk to
inspect the apparent disturbance caused by wasps, but then returned to their webs. Upon
disturbance, Uloborus sometimes stayed at the center (n=3) or moved to the periphery of
their webs (n=2), but never dropped off their webs.
DISCUSSION
Through chemical and behavioral analyses we have determined that all spiders taken by
wasps, regardless of web type have similar chemistry, but not all spiders with that
chemistry are taken by wasps (Fig. 2, Fig. 3). Araneids, uloborids, salticids and thomisids
have similar cuticular chemical profiles, but wasps took no spiders in one araneid genus
(Micrathena), nor did they take any uloborids. Derived araneoids have cuticular chemical
profiles different from those of araneids (Fig. 2), and although wasps antennated these
spiders and/or their webs, they did not pursue or attack the spiders (Fig. 3).
Similar to the differential response of wasps towards spiders, the antipredatory behavior
of spiders also varied according to taxonomic affiliation: all araneids, excepting

74
Micrathena, either hid or dropped down from their webs in response to wasp attacks (Fig.
4). These behaviors seem to be a common strategy used by araneids in response to a
range of disturbances including wasp attacks (Eberhard 1970; Cloudsley-Thompson
1995; Rayor 1999; Blackledge & Pickett 2000, Uma & Weiss 2010). Unlike araneids,
derived araneoid and uloborid spiders rarely moved to the webs’ periphery, and never
dropped off the webs.
Spider chemistry, web architecture, morphology and behavior: important at
different stages of prey capture
Many factors influence the prey capture success of mud-dauber wasps, but their relative
importance varies over time. For example, seasonal patterns of spider availability
influence opportunities for prey choice at a broader level. On the other hand, spider
chemistry, web architecture, and defensive behavior, as well as spider shape and size play
significant roles once the wasp has encountered a spider. Such a ‘hierarchy’ of sequence
is also seen influencing prey capture success in a number of parasitoid wasps (Vinson
1976). Once a mud-dauber wasp approaches a prey habitat, it may accidentally bump into
a web, or encounter a spider resting on its web or on vegetation. In either case,
Sceliphron uses chemical cues present on silk and spider cuticle to recognize a potential
prey (Uma & Weiss 2010). Thus, chemistry is the first available cue that allows the wasp
to determine whether a particular spider or web is worth pursuing. If the ‘right’ chemistry
is established, the wasp may continue to search for the spider for an extended period; if
the spider has the ‘wrong’ chemistry, on the otherhand, the wasp is likely to stop

75
searching. For example, after touching the web of Larinioides sclopetarius, an araneid
spider, a Sceliphron forager searched the web and the surrounding area for 11 min;
however, gave up searching in less than 34 secs after it encountered a web of
Achaearanea tepidariorum, a derived araneoid (Uma & Weiss 2010).
Once a wasp bumps into a web with the ‘right blend’ of chemical cues, the web
architecture, encompassing its two or three dimensionality and presence of 3D retreats,
debris, stabilimenta, egg sacs, prey carcasses, or a leaf for hiding, can influence prey
capture. Variation in web architecture within araneid genera contributed to almost a two-
fold variation in prey capture success by wasps (82%-44%). In case of Mecynogea
lemniscata, sitting within a 3D dome-shaped web was actually detrimental to the spiders,
as they appeared as contrasting dark objects against their ‘invisible’ webs, and wasps
flew directly towards them and captured most of them (capture success 81.81%). The
spiders used in our study were sub-adults, but we speculate that adult spiders that adorn
their webs with egg sacs may be better protected from wasp predators, as egg sacs of
some araneids (Cyclosa sp.) serve as decoys to distract predators (Tseng & Tso 2009).
Wasps were less successful in capturing Metepeira hidden behind their three-dimensional
carcass-covered retreats: while wasps continued to search the debris, the spiders moved
to the periphery or dropped off their webs. Thus the prey-carcass decorations helped
spiders to buy time from the imminent danger. A similar example of the use of prey-
carcass decorations to avoid wasps is seen in Cyclosa (Araneidae) (Chou et al. 2005,
Tseng & Tso 2009).

76
Highly coiled non-sticky silk (cribellate silk) of the Uloborus web could deter wasp
attacks. Uloborus built and sat in the middle of their 2D webs, but were never captured
by wasps. In four of the six instances we observed, wasps bumped into the webs of
Uloborus only to get temporarily stuck in spiders’ silk threads. After freeing themselves,
wasps were found to heavily groom their antenna and legs. Cribellate orb webs are
energetically expensive and time-consuming to spin (Opell 1997, 1998) compared to the
orb webs of araneid spiders; however the cribellate webs may be better at keeping spider-
hunting wasps at bay. A couple of times, we saw wasps fly directly towards and
antennate a spider sitting at the center of its web, but never pursued or captured them.
This behavior was similar to how wasps treated Micrathena or Tetragnatha. In another
experiment, a wasp antennated and stung a freshly killed Uloborus sp., not on its web,
(Divya Uma personal observation) suggesting that it recognized Uloborus as a potential
prey.
Spider morphology may also determine the ability of wasps to capture spiders: wasps
never captured Micrathena though the spiders sat right in the middle of their webs, and
had cuticular profiles similar to those of other araneids. Wasps often hovered close to the
spiders, antennated them and then flew away. Our observation corroborates the finding
that spiny orb weavers are rarely found in wasps’ nests (Elgar & Jebb 1999). We can
think of two equally plausible explanations for this pattern: first, early instars of wasp
larva may find the hard spiny exoskeleton of Micrathena difficult to penetrate and
consequently adult wasps have come to avoid Micrathena over an evolutionary time; next

77
Micrathena may be difficult to pack into a mud cell due to the presence of projecting
spines, such that capturing the spiders may not be profitable (Blackledge, personal
communication). Further experiments are necessary to explore these possibilities.
Spider behavior becomes important at the very end of the prey capture sequence:
defensive strategies of araneids such as hiding at the periphery or dropping off the web
are some times effective, but not always, as 77.7% of the araneids (n=27) that
moved/dropped down were nevertheless caught by S.caementarium. Other araneids such
as Argiope are also known to drop off their webs in response to wasps’ attacks, but most
were nevertheless captured by wasps (Blackledge & Pickett 2000). Derived araneoids
such as Steatoda (Theridiidae) mostly stayed on their webs (n=5), but some came down
to assess the apparent disturbance caused by wasps. Sceliphron does not recognize
derived araneoids as prey, and hence do not stay in the web area long enough to interact
with the resident spider. However, theridiid spiders can be dangerous to wasps (Uma &
Weiss 2010).
Are spiders’ cuticular chemicals defensive adaptations to avoid wasp attacks?
Blackledge et al. (2003) hypothesized that the three-dimensional spider webs built by
derived araneoid spiders serve in part as defensive adaptations, and that escape from
wasp predation may have facilitated the evolution of three-dimensional webs. Indeed, 3D
web architecture of some araneids (e.g., Metepiera) may aid in escape from wasp attacks.
But we now know that Sceliphron do not capture derived araneoids, not because of their

78
3D webs, but due to the presence of a different chemical signature on their cuticles. Our
GCMS data indicates that the hydrocarbons present on the derived araneoids are much
longer (up to C40) than those present on araneids (upto C33). Moreover, the majority of
the hydrocarbons present on derived araneoids have different functional groups (esters
and ethers) than those present on araneids. In fact, several recent studies reveal a
chemical distinction between the silk of araneids and derived araneoids (Bagneres
et.al.1997; Papke 2000; Schulz 2004).
It is possible that a shift in identity of chemical compounds present on derived araneoids
before the evolution of 3D webs may have led to the escape, and subsequently
diversification of derived araneoids. Though our sampling of spiders for chemical
analysis is limited, five spider genera in three different families (Theridiidae, Linyphiidae
and Tetragnathidae) all have cuticular chemistry that are not recognized by wasps.
Alternatively, some derived araneoids (e.g. theridiids) are dangerous to wasps (Chapter
3), and wasps may have evolved to sense the characteristic hydrocarbons that are present
on the derived araneoids as a means to avoid contact with their webs. Further sampling of
spiders is required to determine the generality of this pattern across multiple wasp genera.

79
REFERENCES
Aitchison, J. 1986: The statistical analysis of compositional data. Chapman & Hall,
London.
Bagneres, A.G., Trabalon, M., Blomquist, G. J. & Schulz, S. 1997: Waxes of the social
spider Anelosimus eximius (Araneae, Theridiidae): Abundance of novel n-propyl
esters of long-chain methyl-branched fatty acids. Arch. Insect Biochem. Physiol.
36, 295-314.
Blackledge, T. A. & Pickett, K. M. 2000: Predatory interactions between mud-dauber
wasps (Hymenoptera, Sphecidae) and Argiope (Araneae, Araneidae) in captivity.
J. Arachnol. 28, 211-216.
Blackledge, T. A., Coddington, J. A. & Gillespie, R. G. 2003: Are three-dimensional
spider webs defensive adaptations? Ecol. Lett. 6, 13-18.
Chivers, D. P., Brown, G. E. & Smith, R. J. 1996: The evolution of chemical alarm
signals: attracting predators benefits alarm signal senders. Am. Nat.148, 649–659.
Chivers, D. P. & Mirza, R. S. 2001: Predator diet cues and the assessment of predation
risk by aquatic vertebrates: a review and prospectus. In: Chemical signals in
vertebrates, Volume 9 (Marchlewska-Koj A., Lepri, J. J & Müller-Schwarze, D.
eds). Plenum Press, New York, pp. 277-284.
Chou, I. C., Wang, P. H., Shen, P. & Tso, I. M. 2005: A test of prey-attracting and
predator defense functions of prey carcass decorations built by Cyclosa spiders.
Anim. Behav. 69, 1055-1061.

80
Cloudsley-Thompson, J. L. 1995: A review of the anti-predator devices of spiders. Bull.
British Arachnol. Soc.10, 81-96.
Conover, M. R. 2007: Predator-Prey Dynamics: The Use of Olfaction. Taylor and
Francis, NewYork, pp.250
Dicke, M. & Grostal, P. 2001: Chemical detection of natural enemies by arthropods: an
ecological perspective. Ann. Rev. Ecol. Syst. 32, 1-23.
Eberhard, W. 1970: The predatory behavior of two wasps, Agenoideus humilis
(Pompilidae) and Sceliphron caementarium (Sphecidae), on the orb weaving
spider Araneus cornutus (Araneidae). Psyche. 77, 243–251.
Elgar, M. A. & Jebb, M. 1999: Nest provisioning in the mud-dauber wasp Sceliphron
laetum (F. Smith): Body mass and taxa specific prey selection. Behaviour 136,
147-159.
Evans, H. E. & Eberhard, M. J. W. 1971: The wasps. Michigan University Press,
Michigan.
Gaskett, A. C. 2007: Spider sex pheromones: emission, reception, structures, and
functions. Biol. Rev. 82, 27-48.
Herzner, G., Schmitt, T., Linsenmair, K., Strohm, E. 2005: Prey recognition by females
of the European beewolf and its potential for a sensory trap. Anim. Behav. 70,
1411-1418.
Kortet, R. & Hedrick, A. 2004: Detection of the spider predator, Hololena nedra by naive
juvenile field crickets (Gryllus integer) using indirect cues. Behaviour. 141, 1189-
1196.

81
Kats, L. B. & Dill, L. M. 1998: The scent of death: chemosensory assessment of
predation risk by prey animals. Ecoscience. 5, 361-394.
Minchin, P. R. 2005: DECODA statistical software 3.0. Southern Illinois University
Edwardsville. IL.
Milinski, M., Griffiths, S., Wegner, K. M., Reusch, T. B. H., Haas-Assenbaum, A. &
Boehm, T. 2005: Mate choice decisions of stickleback females predictably
modified by MHC peptide ligands. Pro. Nat. Aacd. Sci. 102(12), 4414-4418.
Nevitt, G. A. 2000: Olfactory foraging by Antarctic Procellariiform seabirds: life at high
Reynolds numbers. Biol. Bull.198, 245-253.
Opell, B. D. 1997: The material cost and stickiness of capture threads and the evolution
of orb-weaving spiders. Biol. J. Linn. Soc. 62, 443-458.
Opell, B. D. 1998: Economics of spider orb-webs: the benefits of producing adhesive
capture thread and of recycling silk. Fun. Ecol. 12, 613-624.
Punzo, F. 2006: Female spider wasps, Anoplius splendens Driesbach (Hymenoptera:
Pompilidae), learn to associate the odor of host feces with the presence of the
host. J. Entomol. Science. 41, 202-210.
Papke, M. D. 2000: Isolierung, identifizierung und synthese von signalstoffen aus
spinnen. Ph.D. thesis, University of Hamburg, Germany.
Persons, M. H. & Uetz, G. W. 1996: Wolf spiders vary patch residence time in the
presence of chemical cues from prey (Araneae, Lycosidae). J. Arachnol. 24, 76-
79.

82
Persons, M. H. & Rypstra, A. L. 2000: Preference for chemical cues associated with
recent prey in the wolf spider Hogna helluo (Araneae: Lycosidae).Ethology. 106,
27–35.
Rayor, L.S. 1997: Attack strategies of predatory wasps (Hymenoptera: Pompilidae;
Sphecidae) on colonial orb web-building spiders (Araneidae: Metepeira
incrassata). J. Kansas Entomol. Soc. 69(4), 67–75.
Schoeppner, N. M. & Relyea, R.A. 2005: Damage, digestion, and defense: the roles of
alarm cues and kairomones for inducing prey defenses. Ecol. Lett. 8, 505-512.
Schulz, S. 2004: Semiochemistry of spiders. In: Advances in Insect Chemical Ecology
(Carde, R.T. & Miller, J.G. eds). Cambridge University Press, Cambridge, pp.
110-150.
Tseng, L. & I-Min Tso. 2009: A risky defense by a spider using conspicuous decoys
resembling itself in appearance. Anim. Behav. 78(2), 425-431.
Uma, D. B. & Weiss, M. R. 2010: Chemical mediation of prey recognition by spider-
hunting wasps. Ethology. 116, 85-95.
Vinson, S. B. 1976: Host selection by insect parasitoids. Ann. Rev. Entomol. 21, 109-
133.
Weiss, M. R. 2003: Good housekeeping: why do shelter-dwelling caterpillars fling their
frass? Ecol. Lett. 6, 361-370.

83
Fig. 1 Schematic diagram of phylogenetic relationship between web and non-web-
building spiders: spider genera that are used only in the chemical analysis, and not in
behavioral assays are indicated with an ‘*’next to web/spider drawings. Neoscona (~)
was used only for the behavioral assays, but not for chemical analysis. Carbon chain
length of cuticular compounds is written below each family. Tree modified and redrawn
from www.tolweb.org
Uloboridae Uloborus
Hyptiotes
Araneoidea
Araneidae
Neoscona
Metepeira
C21-29
Larinioides
Mecynogea
Micrathena
Tetragnathidae Tetragnatha
Leucauge Linyphiidae
Steatoda
Achaearanea
Orbiculariae
Salticidae
Phidippus
Thomisidae Misumena
Frontinella
Theridiidae
C21-33
C23-31
C29-40
C21-40
C21-29
C23-28
Derived araneoids
*
*
*
*
*
*
~
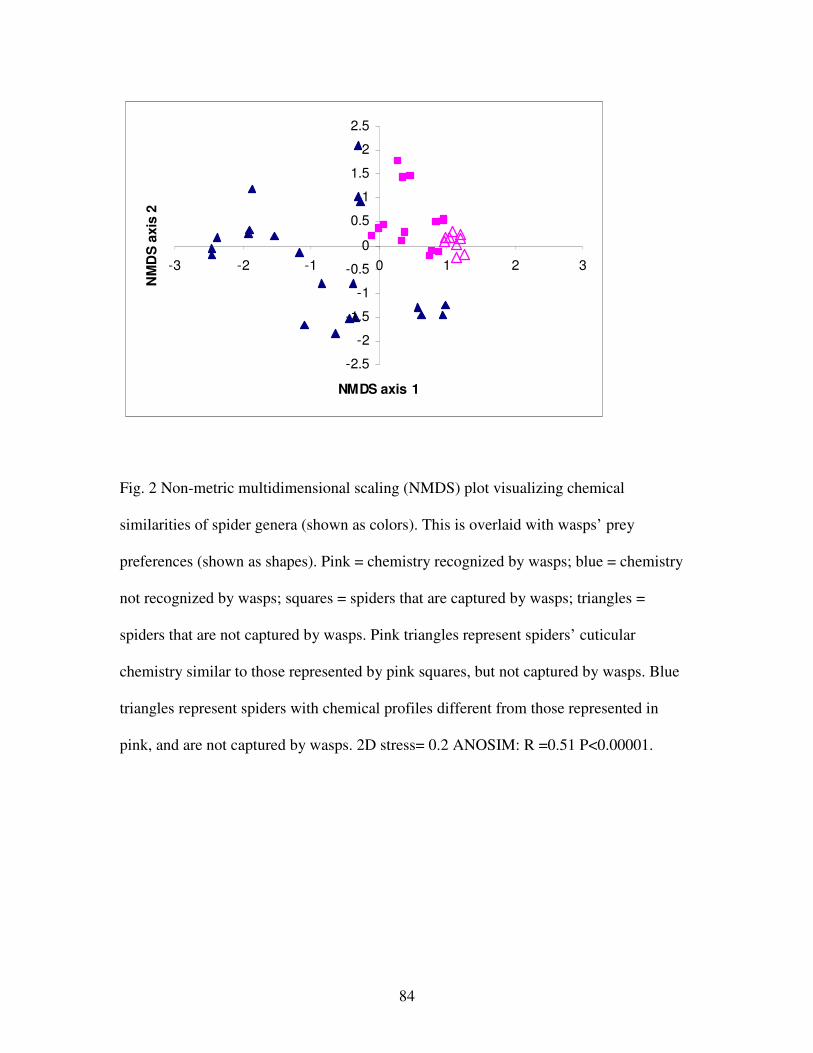
84
-2.5
-2
-1.5
-1
-0.5
0
0.5
1
1.5
2
2.5
-3 -2 -1 0 1 2 3
NMDS axis 1
NM
DS
axis
2
Fig. 2 Non-metric multidimensional scaling (NMDS) plot visualizing chemical
similarities of spider genera (shown as colors). This is overlaid with wasps’ prey
preferences (shown as shapes). Pink = chemistry recognized by wasps; blue = chemistry
not recognized by wasps; squares = spiders that are captured by wasps; triangles =
spiders that are not captured by wasps. Pink triangles represent spiders’ cuticular
chemistry similar to those represented by pink squares, but not captured by wasps. Blue
triangles represent spiders with chemical profiles different from those represented in
pink, and are not captured by wasps. 2D stress= 0.2 ANOSIM: R =0.51 P<0.00001.

85
Fig. 3 Wasps captured most araneids and did not capture derived araneoids or uloborids
regardless of web architecture. Micrathena and Uloborus were not captured by wasps
though their cuticular chemistry was similar to other araneids, and they sat on 2D webs.
Derived araneoids, which had different cuticular chemistry, were never captured by
wasps. Spider genus was a significant predictor of the outcome of wasp responses
(logistic regression: spider genus X² = 12.45 d f =1 P<0.0001). Total number of spiders
tested is written in parenthesis.
0 10 20 30 40 50 60 70 80 90
Steatoda (n=8)
Frontinella (n=10)
Tetragnatha (n=6)
Micrathena (n=6)
Mecynogea (n=11)
Metepeira (n=16)
Neoscona (n=9)
Uloborus (n=6)
percentage of spiders captured by wasps
Derived araneoids
Araneids
Uloborid

86
Fig. 4 Antipredatory behaviors of spiders differ based on taxonomic affiliation: while
most araneids moved or dropped off their webs, most derived araneoids stayed on their
webs. Steatoda (grey bar) came down from their webs, perhaps due to the disturbance
caused due to breaking off gumfooted strands. Uloborus moved or stayed on their webs.
Taxonomic affiliation (whether spider belonged to uloborid, araneid or derived araneoid
groups) was a significant predictor of spiders’ antipredatory behavior (Logistic
regression: X² = 4.65, df = 1, P= 0.03) but web architecture (Logistic regression: X² =
1.62, df =1, P =.202) or wasp responses were not (Logistic regression: X² = 2.74, df =1, P
= 0.09). Number of spiders tested is indicated in parentheses.
Derived araneoids
Araneids
Uloborid
0 30 60 90
Steatoda (n=8)
Frontinella (n=10)
Tetragnatha (n=5)
Micrathena (n=6)
Mecynogea (n=9)
Metepeira (n=13)
Neoscona (n=8)
Uloborus (n=6)
Percent spiders that move or drop off their webs on wasp attack
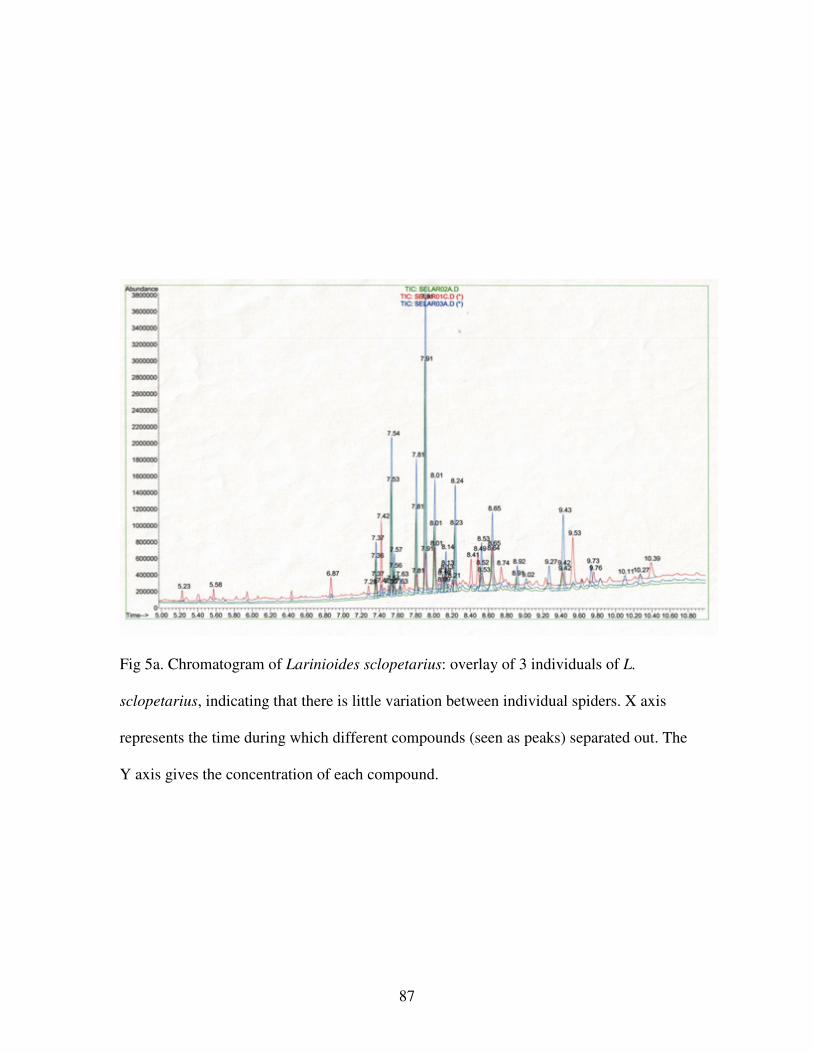
87
Fig 5a. Chromatogram of Larinioides sclopetarius: overlay of 3 individuals of L.
sclopetarius, indicating that there is little variation between individual spiders. X axis
represents the time during which different compounds (seen as peaks) separated out. The
Y axis gives the concentration of each compound.
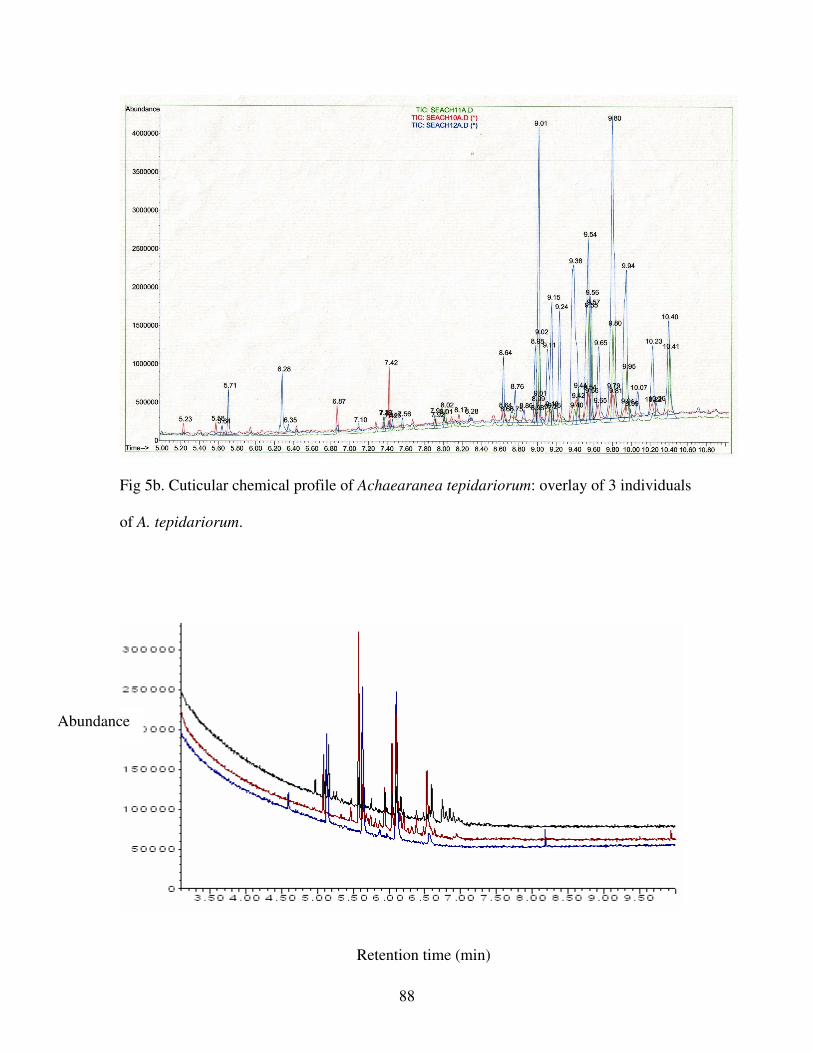
88
Fig 5b. Cuticular chemical profile of Achaearanea tepidariorum: overlay of 3 individuals
of A. tepidariorum.
Retention time (min)
Abundance

89
Fig. 5c Chemical profiles of Larinioides, (Araneidae, black), Phidippus, (Salticidae, red)
and Misumena, (Thomisidae, blue). These three spiders belong to 3 different families, yet
their chemical profiles are similar; and are taken by wasps.

90
Chapter 3
Prey turns predator: ontogenetic changes in the anti-predator behavior of theridiid
spiders against mud-dauber wasps
INTRODUCTION
Predation, one of the main factors affecting ecosystem function, can affect prey traits,
abundances and community interactions (Barbosa and Castellanos 2005). The ability of
animals to assess the risk of being preyed upon often shapes prey behavior, but predation
risk can also generate indirect community interactions, including trophic cascades and
interactions amongst multiple predators (Lima and Dill 1990, Sih et al. 2000). To avoid
predation, prey species may undergo temporal, spatial, morphological, life history or
behavioral changes at different developmental stages (Sih 1987,Lima and Bednekoff
1999, Stankowich and Blumstein 2005). For example, the bluegill sunfish undergoes
size-specific ontogenetic shifts in habitat use to avoid predation from largemouth bass
(Werner and Hall 1988); panic moth caterpillars color change from cryptic to aposematic
at later instars (Grant 2007). Understanding how the risk of predation influences
decision-making in animals is important in understanding the nature of predator-prey
interactions, and ultimately their ecological consequences.
Spiders are ideal organisms with which to study ontogenetic changes in predation risk, as
they are not only top predators of insects, but also fall prey to them. Web-building spiders
in particular are excellent models as they are stationary and hence easily tractable, and

91
their webs are physical representations of their foraging decisions (Sherman 1994,
Blackledge 1998, Blackledge and Zevenbergen 2007).
Spiders in the family Theridiidae spin and rest within three-dimensional cobwebs. Unlike
many araneid webs, which are constructed anew every night, cobwebs remain for many
days, and are expanded and repaired, but rarely abandoned (Benjamin and Zschokke
2003). Achaearanea tepidariorum (Theridiidae) is a cosmopolitan spider, commonly
found near human habitats. The web of A. tepidariorum is comprised of sticky ‘gum-
footed’ threads which are held under high tension, connected to a matrix of supporting
threads at one end, and the substrate at the other end (Fig. 1) (Benjamin and Zschokke
2003). Gumfooted threads, as their name suggests, have sticky glue droplets at their ends;
these can adhere to cursorial prey, either pulling smaller prey up from the ground or
restraining larger prey (Szlep 1965). When a resident spider is alerted by vibrational cues,
it comes down from its retreat, wraps the prey with silk, and takes it back to its retreat to
consume it.
Sphecid wasps are one of the main invertebrate predators of spiders, as individual wasps
provision cells of their mud nests with a dozen or more paralyzed spiders as larval food
(Coville 1987). The spider taxa subject to wasp predation in a given area can be easily
detected by examining the nest contents of wasps. In fact, research over the past century
has revealed that four of the six genera of spider-hunting mud-dauber wasps
preferentially capture orb-web weavers, which build two-dimensional flat webs, and
rarely take cobweb spiders, which build three-dimensional webs, though the latter are

92
more abundant in nature (Blackledge et al. 2003 and references therein). However, wasps
in the genus Chalybion, commonly known as the blue mud-dauber wasp, hunt both orb
and cobweb spiders equally (Muma & Jefferson 1945, Landes et al. 1987, Coville 1976,
Blackledge & Pickett 2000, also see appendix of Blackledge et al. 2003). Chalybion does
not build its own mud nests; it cohabits with another mud-dauber wasp, Sceliphron, and
occupies those wasps’ old and abandoned nests (Coville 1976).
Based on detailed accounts of the hunting behaviors of Sceliphron caementarium
(Eberhard 1970, Blackledge and Pickett 2000, Uma & Weiss 2010) and Chalybion spp.,
(Coville 1976, Blackledge and Pickett 2000) it is clear that the two wasp taxa utilize
different strategies when hunting orb web spiders. Foraging S. caementarium randomly
bump into webs, and then actively search the web area and ground below the web site to
locate the spider, using a combination of chemotactile and visual cues. Chalybion, in
contrast, is known to employ aggressive mimicry once it encounters an orb web: after
landing on a web, the wasp plucks the silk strands with its second pair of legs and flickers
its wings to imitate a stuck insect. When the spider comes down from its resting position
to investigate, the wasp captures and stings the spider (Coville 1976, Blackledge and
Pickett 2000). Though cobweb spiders are commonly found in its nests, it is not known
how Chalybion hunts them.
In this study we examine the hunting behavior of Chalybion californicum towards the
cobweb spider Achaearanea tepidariorum. In particular, from the wasps’ perspective, we
examined whether a) wasps’ hunting behaviors change when they encounter different

93
size classes of spiders, and b) wasps use aggressive mimicry to capture cobweb spiders.
From the spiders’ perspective, we examined whether the defensive behaviors of A.
tepidariorum change over spider ontogeny.
METHODS
Experimental set-up
We collected female Chalybion californicum from Fort Washington and the Theodore
Roosevelt National Parks from June to September in 2008 and 2009 while they were
resting on their mud-nests. Wasps were uniquely marked using lead-free paint, and three
or four wasps were housed per outdoor mesh cage (2m3). We glued old mud nests of S.
caementarium to wooden shelters in the cage to encourage C. californicum to use them,
as these wasps do not build their own nests, but modify old nests of Sceliphron for
provisioning spiders (Coville 1976). Petri dishes containing a 1:1 honey-water mixture
as a carbohydrate source were placed on inverted cups standing within a large water-
filled trough to reduce ant invasion. Grasses and forbs growing within the cage provided
resting sites for wasps and hiding places for fleeing spiders.
We collected different sizes of female Achaearanea tepidariorum (Theridiidae) by visual
searches around the Washington D.C. area from June to September 2008 and 2009.
Spiders were measured to the nearest mm (tibia-patella length of the first right leg) and
weighed to the nearest mg, and grouped into small (0.003-0.008g), medium (0.008-0.03g)

94
and large (0.03-0.07g) weight classes. Though these weight classifications are somewhat
arbitrary, we attempted to group spiders based on their age classes (i.e., small spiders as
juveniles, medium spiders as subadults and large spiders as adults). Typically different
instars of spiders are determined based on the length of the tibia or other body parts that
does not vary much with weight. We determined that tibia-patella length is highly
correlated with spider weight, and used it as a proxy for estimating spider age (Fig 2).
Individual spiders were allowed to build webs in square wooden frames (25cm²x 8 cm
deep) covered with removable plastic film, which prevented the spiders from escaping.
We used spiders with a 2-4 day old web for the experiments. Spiders were not fed in the
lab, as most were used within one week of capture.
Wasp training and testing
Chalybion wasps, after 2-3 days of acclimatization in the field cage, usually rested on one
of the mud nests. We used only these wasps for our behavioral experiments. We lured
each wasp down to a ‘testing arena’ (a stool, 29x32x40cm, located in the center of the
cage) by holding in front of her a cobweb spider (Steatoda grossa) within its wooden
frame and then slowly moving the frame down to the stool. After the wasp touched the
silk, she generally followed the frame down to the testing arena, at which point she began
to investigate the web. We considered a successful training session one in which the wasp
attacked the spider. After a couple of training sessions, the foraging wasp developed an
association between the testing arena and availability of a spider, and would
spontaneously come down to the arena when a wooden frame was placed there.

95
We presented each Chalybion wasp with one small, medium and large A. tepidariorum
spider, within its frame, one at a time and in a randomized sequence. Each trial
commenced when a wasp first touched the web, and lasted until either the wasp or the
spider had captured the other or until 30 min had elapsed, whichever happened first. We
recorded both wasp and spider behaviors. Wasp hunting behaviors were classified as
follows: a) wasps see and approach spiders: a wasp flies or walks towards the spider that
is sitting on its web, or follows a spider that has dropped to the ground; b) wasps lure
spiders: a wasp plucks on the silk strands of the web and lures the spider towards it; c)
wasps encounter a gum-footed strand of silk: a wasp walks on the base of the web,
breaking the high tension gum-footed threads, causing spiders to come down from their
webs to the site of disturbance. In both ‘b’ and ‘c’ categories, the spider approached the
wasp, as a result of the wasp’s behavior.
Spider behaviors were classified into two categories: a) spiders moving to the periphery
or dropping off their webs, and b) spiders throwing silk on the wasp. Moving to the
periphery or dropping off is a typical antipredatory behavior of spiders (Cloudsley-
Thompson 1995, Uma & Weiss 2010), and we considered that spiders behaved ‘as prey’
if they moved or dropped off their webs. On the other hand, throwing silk and wrapping
prey is a typical predatory behavior (Robinson 1969), and we considered spiders to be
behaving ‘as predators’ of wasps when they did so. For each encounter, we recorded both
wasp and spider behaviors, including approaches, attacks, escapes, and captures. We
analyzed whether wasp behaviors (approach, pluck the web, walk on gum-foot strands)

96
and prey capture success varied with spider size by Fisher’s exact and Chi square tests.
We analysed whether spider size and/or wasp identity influenced spiders’ behavior
(behaving like a prey or a predator) by using logistic regression (SPSS version 18).
RESULTS
Chalybion capture success and hunting behavior
Neither the wasps’ capture success nor hunting behaviors (approaching, plucking the web
or walking on gum-foot strands) varied with size of the spider prey. Overall, Chalybion
wasps successfully captured 15 of the 31 spiders presented to them. Wasps did not
preferentially capture any one size class (Chi square X2
= 2.23, df =2), (Table 1).
a. Wasp approaches to the spider: Wasp approaches to the spider did not vary by size
class (Fisher’s exact tests: small vs. medium, P = 0.3; medium vs. large, P = 1; small vs.
large, P = 0.47). Though wasps approached the small spiders in 90% of the trials, either
by locating them on their webs or by following them on the ground, only 50% of those
spiders were eventually caught (Table 2). Wasps approached the medium spiders in
33.3% of the trials, but successfully caught all of them. Wasps approached large spiders
44.4% of the time, but did not capture any of them (Table 2). In three instances, wasps
did not pursue large spiders after an initial encounter, as spiders started throwing silk. As
a result, these three large spiders successfully evaded wasp attacks.

97
b. Wasp lures the spider: Wasp web plucking behavior did not vary by spiders size
class (Fisher’s exact tests: small vs. medium, P = 0.49; medium vs.large, and small vs.
large, P = 1). In three instances (twice with a medium, and once with a large spider) we
observed wasps plucking the silk strands of the resident spider’s web (Table 2).
Typically, wasps landed on a web, remained motionless for a few seconds and then
plucked the web and / or vibrated their entire body for a few seconds. This whole
sequence was repeated until the spider came down from its resting place. In two of the
three instances were the wasps were successful in capturing the spiders.
c. Wasp walks on gum-footed threads: Wasps behaved similarly when they
encountered the gum-footed strands of webs spun by each of the three spider classes
(Fisher’s exact: small vs. medium, P = 0.2; medium vs. large, P = 1; small vs. large, P =
0.32). While the wasp behavior in the above two categories were ‘intentional’ (wasps
approached spiders or lured the spiders towards them), wasps walking on gum-footed
threads seem to be ‘by accident’. While searching for the spiders, wasps often walked on
the base of the wooden frame, which was covered with sticky gum-footed threads held
under tension. Wasps broke a number of such sticky strands, providing vibrational cues
via the web to the resident spider. Following gumfoot damage, only one small spider
approached a wasp, and it escaped attack by walking away from the wasp. In contrast, six
medium spiders approached the wasps, and 50% of them were stung by wasps. Four large
spiders approached the wasps, and 75% (3/4) of them were stung by wasps.
Spider behavior

98
Considering all categories of wasp behaviors, spider behavior towards wasps was
influenced by spider size. Whereas 85.7% (6/7) of the small spiders moved to the
periphery or dropped off their webs, thus seeming to treat the wasps as predators, 87.5%
(7/8) of the medium and 88.8% (8/9) of the large spiders threw gluey silk on the wasps,
thus seeming to treat them as prey (Fig. 2, logistic regression, small: X²= 7.64, df =2,
P<0.022; medium and large: X²= 5.7 df =1 P< 0.016). Identity of individual wasp did not
influence spider behavior (Logistic regression: X²=0.585 df =1 P=0.44).
DISCUSSION
Hunting behavior of Chalybion on Achaearanea tepidariorum
Our study provides the first documentation of hunting behavior by Chalybion in pursuit
of the cobweb spider, Achaearanea tepidariorum. We observed several instances of
aggressive mimicry, in which wasps repeatedly plucked the web with their legs, flickered
their wings or shook their entire body, sending vibrational signals which were picked up
by the spiders, through the silk strands. In our study, wasps plucked the web and lured
spiders towards them in three out of 31 (9.6%) trials. In a comparable study using orb
weaving spiders, wasps plucked the web to lure spiders in 23% of observed attacks
(Blackledge & Pickett 2000). Such aggressive mimicry may indeed be more common
against orb-web than cobweb spiders, though more experimental trials are needed to
confirm this trend. We can think of two possible reasons that aggressive mimicry might
be more common against orb weavers than cobweb spiders.

99
First, most orb weavers are nocturnal and hide in retreats or at the periphery of their webs
during the day; thus they are not always visible during the day. Plucking could be an
effective way for diurnally active predators to bring forth retreat-dwelling spiders
(Coville 1976). In a similar strategy, a vespid wasp taps the silken strands of an orbweb
with its antenna to lure the spiders from their hiding places (MacNulty 1961).
In contrast, the cobweb spider, A. tepidariorum is diurnal, resting within its three
dimensional matrix, and has gum-footed silk at the base of its web. In majority of the
cases, wasps either approached the spiders visually or brought the spiders towards them
as a result of walking on the gum-footed strands. Thus, gum-footed threads may serve as
an additional way to lure cobweb spiders near wasps.
Cobweb spiders as prey and predators
Cobweb spiders behaved both as prey and predators of wasps, and the role that they
played seems to depend on the spiders’ size. Close to 90% (6/7) of the small spiders
moved to the periphery or dropped down from their webs when they encountered wasps;
thus, small spiders seem to treat wasps as predators. In contrast, close to 90% (15/17) of
the medium and large spiders attempted to wrap wasps with sticky silk, seemingly
considering wasps as potential prey (Table 2). On several occasions in the field, we have
observed Chalybion entangled in the webs of large adult A. tepidariorum spiders,
confirming that large spiders are indeed dangerous to wasps.

100
Attempting to capture wasps can be risky, even for a large spider. Several spiders lost
their hind legs, as wasps bit spiders while they were throwing silk. A couple of the large
spiders managed to wrap the wasps, but in the process got stung, leaving spiders
paralyzed and wasps dead. Only one large spider actually captured a wasp without
injuring herself.
How do spiders assess prey vs. predators?
Our results suggest that small, medium and large spiders react differently to the same
sized wasp. How do they assess whether it represents a potential predator or a prey item?
Most web-building spiders have poor vision, and rely on air-borne or web-borne
vibratoional cues for prey detection (Barth 1982). Both orb and cobweb spiders are
known to detect vibrations transmitted through silk strands (Uetz and Hieber 1994, Uetz
et al. 2002, personal observations). It is plausible that web-building spiders could use
vibrational information to discriminate prey from predators.
Spider silk diameter is known to increase with spider mass, presumably because spinneret
diameter increases with each molt (Blackledge, unpublished results). Thus small spiders
will produce silk strands of thinner diameter than those produced by medium or large
spiders. The vibrational quality produced by the proximity of a given sized wasp is
transmitted differently across silk of different thickness. Size of the predator in
conjunction with diameter of the silk strand may aid spiders in assessing whether to treat

101
wasps as predators or as prey. Alternatively, mechanical shock of a large insect such as a
wasp could break several weak gumfooted strands in the web of a small spider, resulting
in the collapse of the supporting silk matrix, causing the spider to drop down. Thicker
gumfooted strands and supporting threads, comprising the webs of medium and large
spiders, would be better able to absorb the mechanical shock.
Basis of prey preference by Chalybion
Chalybion do not construct mud nests, but reuse old and abandoned nests of Sceliphron.
Hence, they are restricted to areas that are occupied by Sceliphron. Though both of these
wasps hunt in similar habitats, they provision their nests with different kinds of spiders,
perhaps reflecting their different hunting techniques. Sceliphron specialize on orb
weavers (in the family Araneidae), and also capture non-web weavers such as jumping
spiders and crab spiders. Chalybion, on the other hand, provisions its nests with orb and
cobweb building spiders. Thus, these sympatrically occurring sphecid wasps use different
hunting techniques, perhaps due to niche partitioning (Blackledge and Pickett 2000).
Why does Chalybion capture cobweb spiders, while Sceliphron does not? Prey
recognition by Sceliphron caementarium is mediated by chemical cues present on spider
cuticle and silk (Uma and Weiss 2010). Our preliminary results suggest that spiders (3
genera of orb weavers, 1 jumping and 1 crab species) taken by wasps have similar
cuticular chemical profiles. Mechanistically, Sceliphron’s antennal receptors may have
evolved to recognize a certain set of cuticular chemicals that were present on orb weaving

102
araneid spiders, but are not tuned to chemicals present on the derived cobweb spiders. On
the other hand, Chalybion hunt both orb and cobweb spiders, and most likely use
cuticular chemicals to recognize prey, as they readily stung freshly killed A.
tepidariorum, but never stung freshly killed hexane-washed A. tepidariorum
(unpublished results). It is likely that certain wasp genera, including Chalybion,
diversified after the radiation of cobweb spiders and thus were able to evolve the capacity
to recognize both araneids and derived araneoid spiders as potential prey. A better
understanding of the timing of sphecid evolution in relation to spider evolution, as well as
a generic level wasp phylogeny will help us to clarify the evolution of prey preferences in
mud-dauber wasps.

103
REFERENCES
Barbosa, P. & Castellanos, I. 2005: Ecology of predator-prey interactions. Oxford
University Press, New York.
Barth, F. G. 1982: Spiders and vibratory signals: sensory reception and behavioral
significance. In Spider Communication: Mechanisms and Ecological Significance
(ed. by P.W. Witt and J.S. Rovner), Princeton Univ. Press, Princeton, pp. 67–120.
Benjamin S. P. & Zschokke S. 2003: Webs of theridiid spiders: construction, structure
and evolution. Biol. J. Linn. Soc. 78, 293-305.
Blackledge, T. A. 1998: Stabilimentum variation and foraging success in Argiope
aurantia and Argiope trifasciata (Araneae, Araneidae). J. Zool. London. 246, 21-
27.
Blackledge, T. A. & Pickett, K. M. 2000: Predatory interactions between mud-dauber
wasps (Hymenoptera: Sphecidae) and Argiope (Araneae: Araneidae) in captivity.
J. Arachnol. 28, 211-216.
Blackledge, T. A., Coddington, J. A. & Gillespie, R. G. 2003: The evolution of three-
dimensional spider webs as predator defenses. Ecol Lett. 6, 13-18.
Blackledge, T. A. & Zevenbergen, Z. M. 2007: Condition-dependent web architecture in
the western black widow spider, Latrodectus hesperus. Anim. Behav. 73, 855-864.
Cloudsley-Thompson, J. L. 1995: A review of the anti-predator devices of spiders. Bull.
Brit. Arach. Soc. 10, 81-96.
Coville, R. E.1976: Predatory behavior of the spider wasp, Chalybion californicum. Pan-
Pac Entomol. 52, 229–233.

104
Coville, R. E. 1987: Spider-hunting sphecid wasps. In: Ecophysiology of Spiders.
Nentwig, W., ed. Springer-Verlag, Berlin. pp. 309-318.
Eberhard, W. G. 1970: The predatory behavior of two wasps, Agenoideus humilis
(Pompilidae) and Sceliphron caementarium (Sphecidae), on the orb weaving
spider Araneus cornutus (Araneidae). Psyche 77, 243-251.
Grant, J. B. 2007: Ontogenetic color change and the evolution of aposematism: a case
study in panic moth caterpillars. J. Anim. Ecol. 76(3), 439-447.
Landes, D. A., Obin, M. S., Cady, A. B. & Hunt, J. H. 1987: Seasonal and latitudinal
variation in spider prey of the mud dauber Chalybion californicum (hymenoptera:
Sphecidae). J. Arachnol. 15: 249-256.
Lima, S. L. & Dill, L. M. 1990: Behavioral decisions made under the risk of predation: a
review and prospectus. Can. J. Zool. 68, 619–640.
Lima, S. L. & Bednekoff, P. A. 1999: Temporal variation in danger drives antipredator
behavior: the predation risk allocation hypothesis. Am. Nat. 153, 649-659.
MacNulty, B. J. 1961: Field observations on someWest African Hymenoptera. Proc. Soc.
London Entomol.Nat. Hist. Soc.118–133.
Muma, M. H. & Jeffers, W. F. 1945: Studies of the spider prey of several mud-dauber
wasps. Ann. Entomol. Soc. Am. 38, 245-255.
Robinson, M. H. 1969: Predatory behavior of Argiope argentata (Fabricius). Am. Zool.
9, 161-173.
Sherman, P. M. 1994: The orb web: an energetic and behavioral estimator of a spider’s
dynamic foraging and reproductive strategies. Anim. Behav. 48, 19-34.
Sih, A.1987. Prey refuges and predator–prey stability.Theor. Popul. Biol. 31, 1–12.

105
Sih, A., Ziemba, R. & Harding, K. C. 2000: New insights on how temporal variation in
predation risk shapes prey behavior. Trends Eco Evol. 15, 3-4.
Stankowich, T, & Blumstein, D. T. 2005: Fear in animals: a meta-analysis and review of
risk assessment. Proc. R. Soc. 272, 2627-2634.
Szlep, R. 1965: The web-spinning process and web-structure of Latrodectus
tredecimguttatus, L. pallidus and L. revivensis. Proc. Zool. Soc. Lond 14, 75–89.
Uetz, G. W. & Hieber, C. S. 1994: Group size and predation risk in colonial web-building
spiders: analysis of attack abatement mechanisms. Behav. Eco. 5, 326–333.
Uetz, G. W. & Roberts, J. A. 2002: Multisensory cues and multimodal communication in
spiders: insights from video/audio playback studies. Brain Behv. Evol. 59 (4),
222-230.
Uma, D. B. & Weiss, M. R. 2010: Chemical mediation of prey recognition by spider-
hunting wasps. Ethology. 116, 85-95.
Werner, E. E. & Hall, D. J.1988: Ontogenetic habitat shifts in bluegill: the foraging rate-
predation risk trade-off. Ecology. 69(5), 1352-1366.
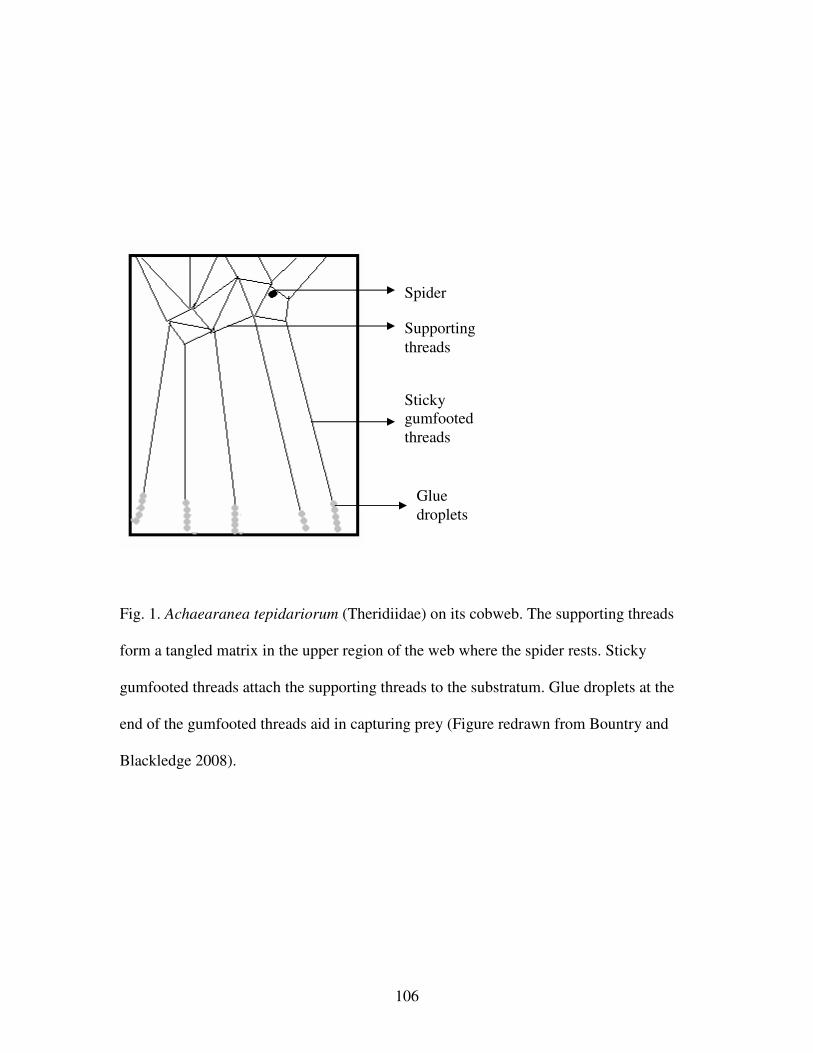
106
Fig. 1. Achaearanea tepidariorum (Theridiidae) on its cobweb. The supporting threads
form a tangled matrix in the upper region of the web where the spider rests. Sticky
gumfooted threads attach the supporting threads to the substratum. Glue droplets at the
end of the gumfooted threads aid in capturing prey (Figure redrawn from Bountry and
Blackledge 2008).
Supporting
threads
Sticky
gumfooted
threads
Glue
droplets
Spider
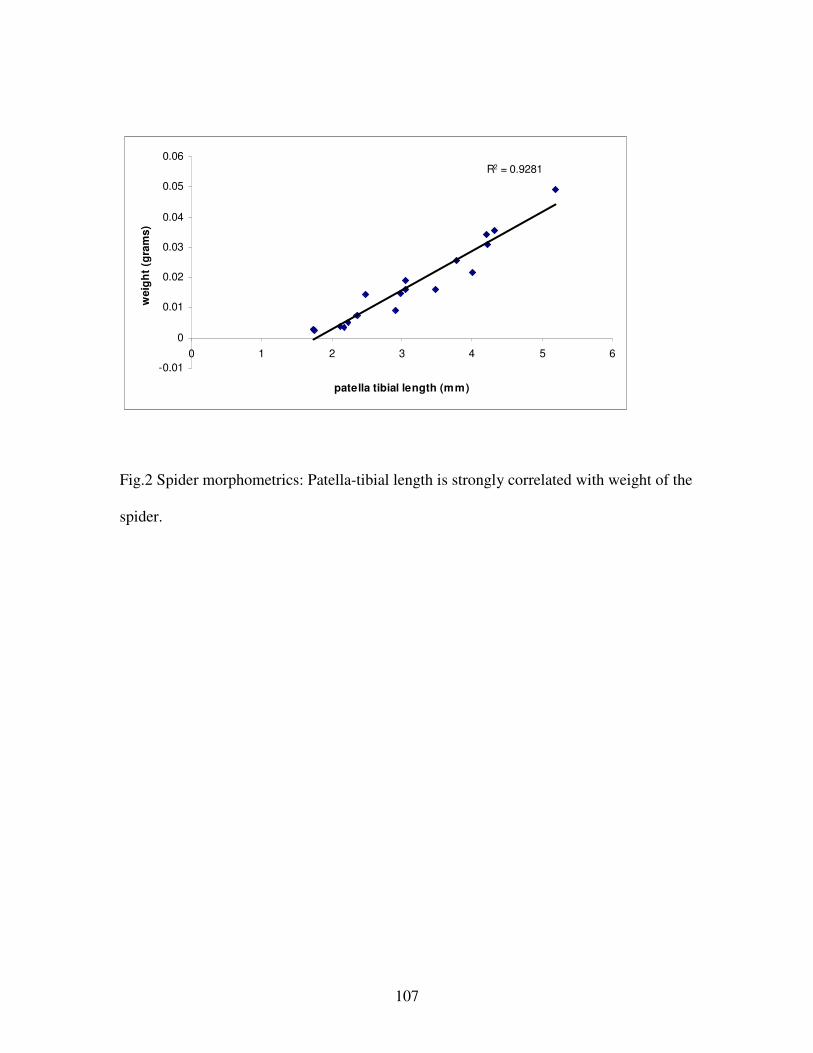
107
R2 = 0.9281
-0.01
0
0.01
0.02
0.03
0.04
0.05
0.06
0 1 2 3 4 5 6
patella tibial length (mm)
weig
ht
(gra
ms)
Fig.2 Spider morphometrics: Patella-tibial length is strongly correlated with weight of the
spider.
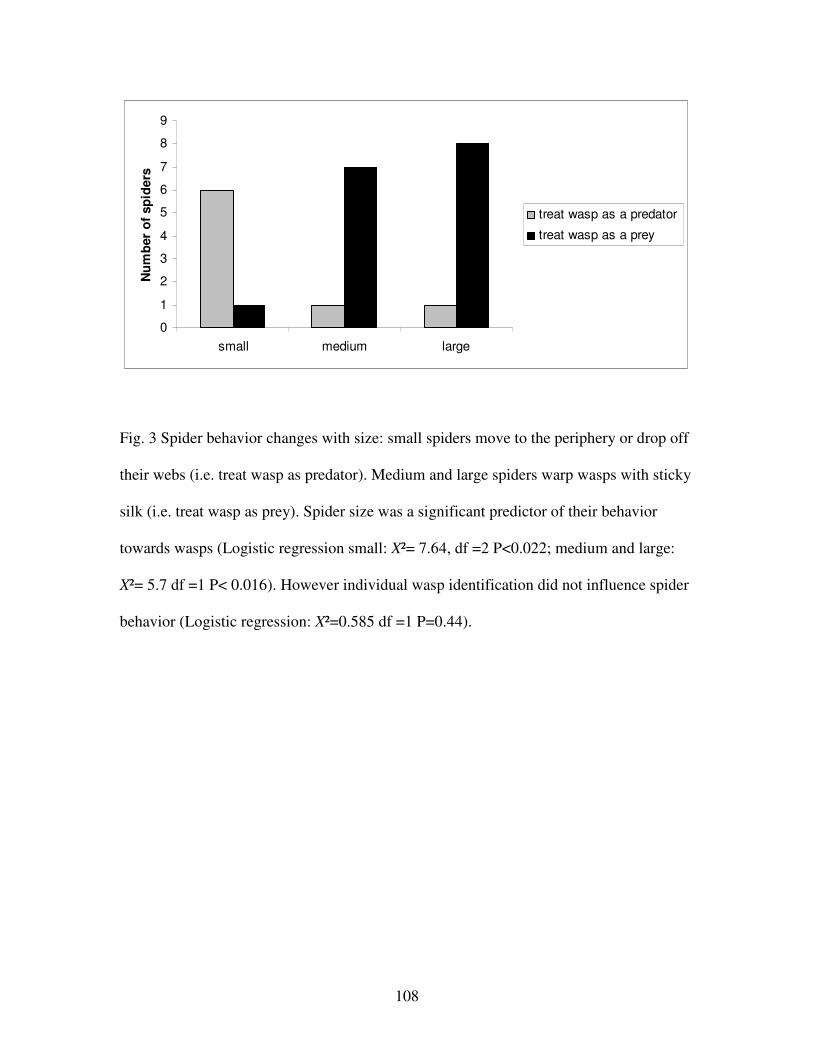
108
0
1
2
3
4
5
6
7
8
9
small medium large
Nu
mb
er
of
sp
iders
treat wasp as a predator
treat wasp as a prey
Fig. 3 Spider behavior changes with size: small spiders move to the periphery or drop off
their webs (i.e. treat wasp as predator). Medium and large spiders warp wasps with sticky
silk (i.e. treat wasp as prey). Spider size was a significant predictor of their behavior
towards wasps (Logistic regression small: X²= 7.64, df =2 P<0.022; medium and large:
X²= 5.7 df =1 P< 0.016). However individual wasp identification did not influence spider
behavior (Logistic regression: X²=0.585 df =1 P=0.44).
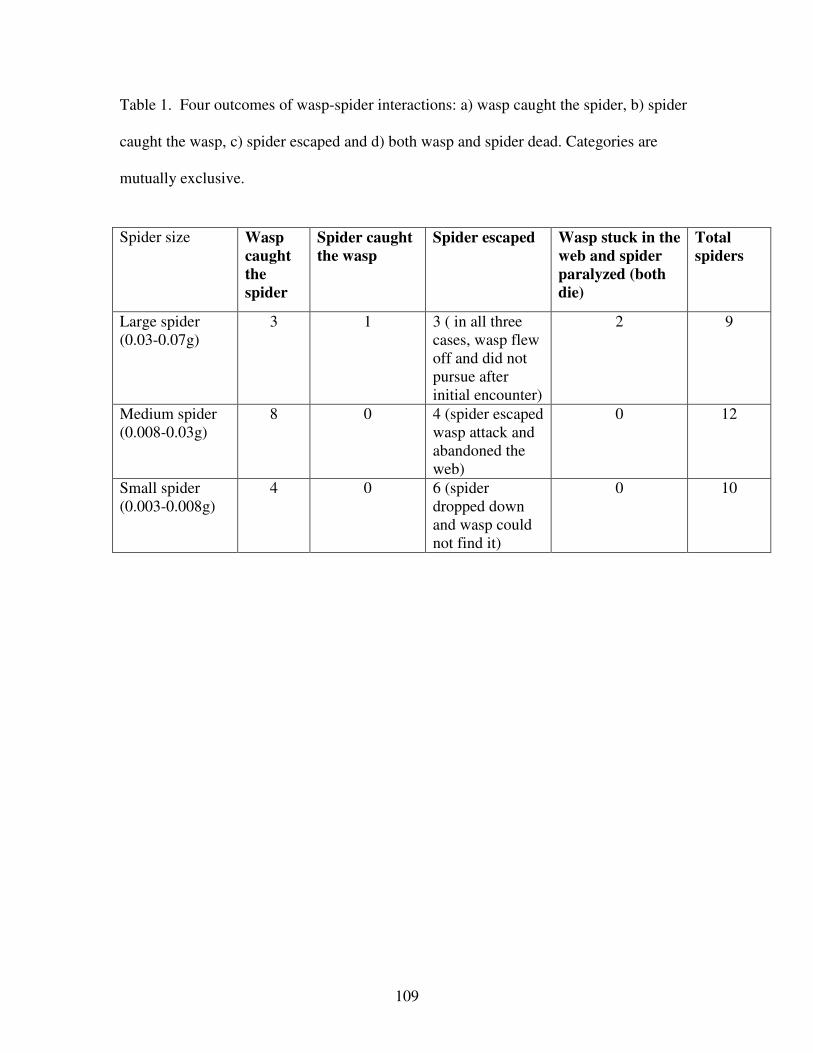
109
Table 1. Four outcomes of wasp-spider interactions: a) wasp caught the spider, b) spider
caught the wasp, c) spider escaped and d) both wasp and spider dead. Categories are
mutually exclusive.
Spider size Wasp
caught
the
spider
Spider caught
the wasp
Spider escaped Wasp stuck in the
web and spider
paralyzed (both
die)
Total
spiders
Large spider
(0.03-0.07g)
3 1 3 ( in all three
cases, wasp flew
off and did not
pursue after
initial encounter)
2 9
Medium spider
(0.008-0.03g)
8 0 4 (spider escaped
wasp attack and
abandoned the
web)
0 12
Small spider
(0.003-0.008g)
4 0 6 (spider
dropped down
and wasp could
not find it)
0 10
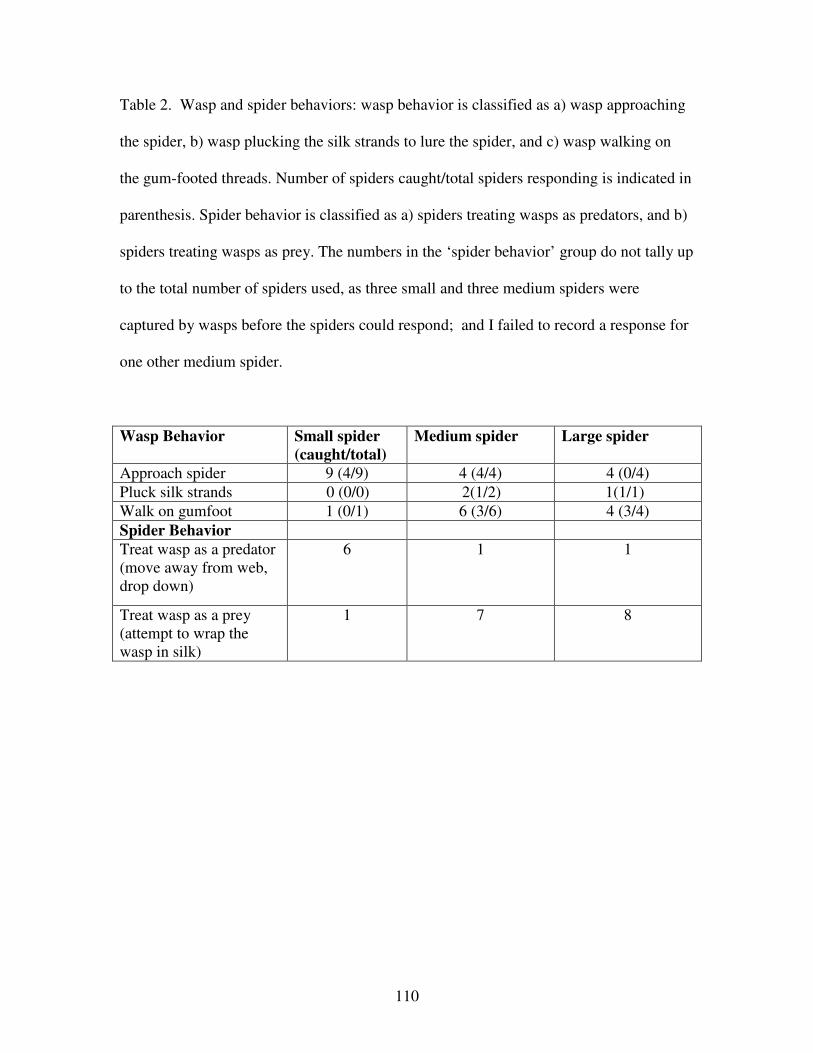
110
Table 2. Wasp and spider behaviors: wasp behavior is classified as a) wasp approaching
the spider, b) wasp plucking the silk strands to lure the spider, and c) wasp walking on
the gum-footed threads. Number of spiders caught/total spiders responding is indicated in
parenthesis. Spider behavior is classified as a) spiders treating wasps as predators, and b)
spiders treating wasps as prey. The numbers in the ‘spider behavior’ group do not tally up
to the total number of spiders used, as three small and three medium spiders were
captured by wasps before the spiders could respond; and I failed to record a response for
one other medium spider.
Wasp Behavior Small spider
(caught/total)
Medium spider Large spider
Approach spider 9 (4/9) 4 (4/4) 4 (0/4)
Pluck silk strands 0 (0/0) 2(1/2) 1(1/1)
Walk on gumfoot 1 (0/1) 6 (3/6) 4 (3/4)
Spider Behavior
Treat wasp as a predator
(move away from web,
drop down)
6 1 1
Treat wasp as a prey
(attempt to wrap the
wasp in silk)
1 7 8

111
Chapter 4
Double deception: ant-mimicking spiders fool both visually and chemically-oriented
predators
INTRODUCTION
Biological mimics provide some of the most astonishing examples of adaptation in the
natural world. Some spectacular examples include the inflorescence of the dead horse
arum, the brownish-maroon color and carrion odor of which attract flies that mistake it
for a rotting carcass (Stensmyr et al. 2002), and hoverflies that resemble wasps in varied
degrees of perfection and yet serve to fool visually-oriented predators (Chittka & Osorio
2007). Typically there are three participants in a mimicry system: two signal emitters
(model and mimic) and one signal receiver (e.g., predator, pollinator, or seed disperser).
Evolutionary and ecological dynamics of mimicry are often influenced by the sensory
capabilities, as well as the limitations of both signalers and receivers (Chittka & Osorio
2007).
Mimics can deceive a receiver by deploying cues in a range of sensory modalities (visual,
chemical, acoustical, textural, behavioral, and movement). Many cases of mimicry
illustrate the use of a single sensory modality to deceive a specific audience: warning
wing-color patterns in heliconid butterflies deceive visually-oriented avian predators
(Kapan 2001) and the cuticular chemicals of cuckoo wasps allow them access to the nests

112
of their beewolf hosts (Kroiss et al. 2008). However, signaling may also involve the
production of multiple cues targeted towards a particular audience: examples of such
multi-modal mimicry include the chemical and acoustical signals that allow lyceanid
caterpillars to parasitize the nests of Myrmica ants (Pierce 2002); the coordinated
movement of patterned wings used by metalmark moths to deceive predation by jumping
spiders (Rota & Wagner 2006); and the chemical, visual and textural resemblance to a
female wasp that leads male wasps to attempt to mate with Ophrys orchid flowers (Borg-
Karlson et al. 1985, Schiestl et al. 2000). However, to our knowledge, mimicry involving
visual, and chemical modalities in a single organism, in which different sensory
modalities serve to deceive different predators, has not been reported. Here we describe
an ant-mimicking jumping spider, Peckhamia picata (Salticidae), which deceives at least
two different arthropod predators: a visually-oriented jumping spider (Phidippus
johnsoni) and a chemically-oriented mud-dauber wasp (Sceliphron caementarium), and
may also deceive chemically-oriented ants (e.g., Camponotus sp.)
Ants dominate terrestrial ecosystems, and are generally avoided by many predators
because of their nasty bite, acid taste, and aggressive behavior. Insects and spiders
belonging to over 200 genera in 50 families are known to resemble ants morphologically,
behaviorally or chemically to gain protection from predators and/or to access resources
within ant nests (McIver & Stonedhal 1993, Cushing 1997). Insect and spider mimics that
live within ant nests employ a combination of strategies to protect themselves ant attacks:
while some employ morphological, chemical or behavioral strategies, others use

113
defensive and appeasement secretions (Dettner & Liepert 1994, Lenoir et al. 2001, Allan
et al .2002).
Myrmecomorphy (morphological and behavioral resemblance to ants) in spiders is
reported in 13 families, though it is seen most often in Salticidae (jumping spiders) and
Clubionidae (sac spiders) (Cushing 1997). Ant-mimicking spiders exhibit a range of
spectacular adaptations, including shape and color modifications that make the two-
segemented arachnid body form resemble that of a three-segmented insect; an ant-like
gait, and ‘antennal illusion’, achieved by holding up and waving the first or the second
pair of legs in a manner that suggests an ant’s antennae. Visual ant mimicry in spiders is
thought to be Batesian, where a palatable spider, by mimicking signals of an unpalatable,
noxious ant model, gains protection from visually-oriented vertebrate (birds and lizards)
as well as invertebrate (jumping spiders, mantids and sphecid and pompilid wasps)
predators (Cutler 1991, McIver & Stonedahl 1993, Cushing 1997, Nelson et al. 2005,
2006a, 2006b). Myrmecomorphic spiders are generally found within the same
microhabitat as ants (Cushing 1993) and, in some cases build their nests next to ant nests,
thereby risking aggression or predation by their potential models (Nelson et al., 2004,
2005, Nelson & Jackson 2008). Indeed, if the spiders are closely associated with ants to
the point that they evolve visual resemblance, then it is also likely that they will be
vulnerable to ant aggression. Because ants mainly rely on chemical cues to discriminate
nestmates from non-nestmates (Lahav et al. 1998, Ozaki et al. 2005), it is possible that
ant-mimicking spiders will evolve chemical resemblance to avoid ant aggression.

114
Whether myrmecomorphs also chemically resemble their model ants has not been
investigated explicitly, although some anecdotal evidence suggest that they may (see
references in Cushing 1997). Chemical resemblance of myrmecomorphs to ants may have
evolved primarily to decrease ant aggression towards mimics, but may also serve to
provide protection from other chemically-oriented predators, such as sphecid wasps.
Mud-dauber wasps (Sphecidae) are one of the main predators of spiders, as individual
wasps paralyze dozens of spiders to deposit in their larval nests (Bohart & Menke 1976;
Coville 1987). Although mud-dauber wasps specialize on spiders in the family Araneidae
(spiders that build two-dimensional webs, Blackledge et al. 2003), they also capture
jumping spiders (Salticidae) and crab spiders (Thomisidae); however, they rarely take ant
mimic salticids or clubionids. It has been suggested that ant-mimicking spiders escape
wasp predation as a result of their visual resemblance to ants (Edmunds 1993). However,
although wasps, including mud-daubers, may use visual cues to locate prey habitat, they
rely on chemosensory cues to recognize spiders as potential prey (Herzner et al. 2005,
Uma & Weiss 2010). We have determined that the cuticular chemical profiles of all
spiders taken by wasps of the species Sceliphron caementarium are similar (Chapter 2).
Thus it is possible that ant-mimicking salticid spiders do not have the same cuticular
chemicals as other salticids that are taken by wasps, and therefore are not captured by
wasps.
Using behavioral and chemical assays, we examined whether ant mimics uses different
modalities to fool different predators. Specifically we tested the hypotheses that the ant-
mimicking spider, Peckhamia picata a) deceives visually-oriented jumping spiders, b)

115
deceives chemically-oriented mud-dauber wasps, and c) chemically resembles its model
ants and thus avoids aggressive encounters.
METHODS
In each of the three experiments described below, Peckhamia picata serves as the mimic;
however, the identities of the predator and the non-mimic (used as a control) differ in
each experiment and are described accordingly.
1. Are visually-oriented jumping spiders fooled by ant mimics?
Subjects: We collected model ant (Camponotus sp.), mimic (Peckhamia picata) and non-
mimics (Metaphidippus sp.) by visual search from the same habitat near the Potomac
River in Washington DC near the Georgetown university campus between May and
October, 2009. Predatory spiders (Phidippus johnsoni, Salticidae) that were field
collected were obtained from the Spider Pharm, a commercial establishment in Arizona.
(We could not use local predatory jumping spiders, as we did not find sufficiently
numbers of a particular species to do behavioral assays). Spiders were housed
individually in plastic cups provided with wet cotton and four to five Drosophila
melanogaster every other day. Four to five ants were housed together in plastic cups and
given wet cotton sprinkled with sugar. Spiders and ants were kept in the laboratory
approximately for 10 days until all of them were tested, and then they were released back
in the field or used for other ongoing experiments in the lab.

116
Experimental design: To examine whether predatory spiders are deceived by Peckhamia
picata’s visual resemblance to Camponotus sp. (Fig 1), we performed the following
experiment, in which each trial included the following three treatments: 1) predator vs.
mimic, to determine whether predatory spiders avoid palatable mimics; 2) predator vs.
ant model, to determine whether predatory spiders avoid unpalatable ants; and 3) predator
vs. non-mimic spider, used as a control to assess the predatory spiderr’s motivation to
hunt.Within a Petri dish (9cm diameter), we presented each predatory jumping spider
(n=19), in random sequence, with an ant-mimic spider, a non-mimic spider, and a model
ant and videotaped each paired interaction for 5 mins, with a 5 min interval between
treatments. Regardless of the treatment, the predatory spider was always introduced first
inside the Petri dish, and was allowed 5 min to acclimate. On the day before a trial,
predatory spiders were fed with three or four Drosophila so that they were not hungry. If
the predatory spiders ate a mimic, non-mimic or an ant during a trial, we performed the
remaining treatments the following day, so that the hunger status of the predator did not
confound the results. Total number of mimics, non-mimics and ants captured by
predatory spiders were recorded and analyzed by Fisher’s exact test.
2. Do chemically-oriented wasps recognize ant-mimicking spiders as prey?
Subjects: Female Sceliphron caementarium (Sphecidae) wasps were collected from the
George Washington Memorial Parkway near metropolitan Washington, D.C. during the
months of June – September, from 2008 through 2009. Wasps were individually marked,

117
and housed in outdoor mesh cages (2m3) that were provisioned with wooden shelters and
wet mud for nest construction. Once wasps had built a mud nest, we trained them to come
down to a ‘testing-arena’ (a 0.5 m tall camp stool) by placing a spider (Larinioides sp or
Metpeira labyrinthea, Araneidae) on its web on the stool, and allowing a wasp to locate
and capture the spider. Wasps were trained until they came down on their own (see Uma
& Weiss 2010 for details on wasp set-up and training). Web-building spiders used for
training wasps were collected from the same area as the wasps, and individual spiders
were allowed to build webs in square wooden frames (25x25cm²x 8 cm deep) covered
with removable plastic wrap, which prevented the spiders’ escaping from the wooden
frames prior to the experiment. Mimetic and non-mimetic spiders were collected and
housed as in experiment 1, above.
Experimental Design: To examine whether wasps discriminated between mimetic
(Peckhamia) and non-mimetic spiders (Phidippus sp.), we presented individual wasps
(n=8) with paired freshly frozen spiders of comparable weight, positioned six cm apart on
a filter paper disc (11 cm diameter). (Wasps do not distinguish between freshly-killed
spiders and live spiders, Uma & Weiss 2010). The paper disc itself was placed on a
plastic mat (0.5cm*7cm*24cm) located on the testing arena. Each trial, which consisted
of an individual wasp encountering a pair of freshly killed (by freezing for 15 min @-4C)
mimic and non-mimic spiders, commenced when a wasp walked or flew to the paper disc
and ended when the wasp antennated both of the spiders, and subsequently either sting or
rejected them, or until 30 minutes had elapsed, whichever happened first. New spiders
were used in each trial and their positions were switched between trials, and a fresh paper

118
disc was used each time. Number of mimic and non-mimic spiders captured by wasps
was analyzed using a binomial probability test.
3. Do ant-mimicking spiders chemically resemble their model ant?
Chemical Preparation: To examine whether mimics chemically resemble ants, we
analyzed cuticular compounds of both model ants (Camponotus sp), and mimic spiders
(Peckhamia) by gas chromatography and mass spectrometry (GC/MS). Non-mimetic
spiders (Phidippus johnsoni) were used as a control. Model ants and adult spider mimics
were collected from the same microhabitat near the Potomac River, Washington DC, and
adult non-mimetic spiders were obtained from Spider Pharm, a commercial establishment
in Arizona. Though we would have preferred to collect the non-mimicking spiders from
the same habitat as ants and mimics, we did not find enough number non-mimics of the
same species at the time of the experiment. P. johnsoni were field caught in Arizona.
While the cuticular chemicals of ant (n = 3) and ant mimic (n = 3) were extracted by
soaking freshly killed individuals in 150 ul of HPLC grade hexane for15 min at room
temperature, those of non-mimetic spiders (n = 3) were extracted by soaking individual
spiders in 1.5 ml HPLC grade hexane for 15 min. After removal of the spiders and ants,
the extracts were stored at –20°C in glass vials with Teflon-lined lids until ready for
analysis. For chemical characterization, 2µl of extract was injected into an Agilent
Technologies 6890 gas chromatograph interfaced to a 5973 mass selective detector
(GC/MS), using a splitless injection at 300°C onto a ZB1ms column (30mx0.25mm) and
a temperature program of 100°C-320°C @ 30°C/min. Chemical constituents were

119
identified by computer matching EI mass spectra against the 2005 NIST database of ca.
200,000 spectra.
Data analysis: Chemicals present on the cuticle of spiders were analyzed by multivariate
methods. To consider a chromatographic peak characteristic of a genus, at least half of
the individuals (two out of three in our case) within that genus had to posses that
corresponding peak. We excluded from the analysis peaks that did not match these
criteria. Only hydrocarbons with carbon chain length C21 or more were included in the
analysis, as these are considered to be typical insect and spider cuticular compounds (ref).
Since for a given sample, relative peak areas are not statistically independent, we
transformed the data according to Aitchison’s method (1986). Non-parametric statistical
procedures were applied that suited the distribution of our data. We used non-metric-
multidimensional scaling (NMDS) to visualize the chemical distances of different spider
taxa. Analysis of similarity (ANOSIM) was used to test significance between a priory
groups by using PAST software.
RESULTS
1. Visually-oriented jumping spiders are fooled by ant mimics
We found that Phidippus johnsoni, the predatory jumping spider captured non-mimics
significantly more than it captured mimics (Fisher’s exact test P = 0.034) or model ants
(Fisher’s exact test P = 0.009) (Fig.2). Furthermore, numbers of ants and ant mimics

120
captured by the predatory spider were not different, (Fisher’s exact test, P = 0.5),
suggesting that the predators were fooled by ant mimics’ resemblance to ants.
2. Chemically-oriented wasps do not recognize ant-mimicking spiders as prey
When offered a simultaneous choice of freshly-killed mimic (Peckhamia picata) and
non-mimic (Phidippus sp.) spiders, matched for weight (T test: P = 0.11, t = 1.68 df =
12), S. caementarium wasps (n=8) antennated both members in eight pairs; they stung
seven of the eight non-mimics, but did not sting any of the mimics (binomial probability
P = 0.035, N=7).
3. The ant-mimicking spider Peckhamia picata chemically resembles its model ant
Chemical profiles of ant and ant mimics were similar, although the mimics had reduced
amounts of hydrocarbons compared to ants (Fig. 3). In contrast, the profile of non-mimic
spiders (Phidippus johnsoni) was quite different from both ant and mimic. Saturated and
branched hydrocarbons were the predominant chemicals found on the cuticles of all three
taxa (ants, mimics and non-mimics). However, they differed in both number of
compounds, and length of HC chains. While we found 21 compounds (C23-C35) on the
cuticle of Camponotus ants, ant-mimic spiders had only six compounds (C25 to C33),
and the non-mimic had 10 compounds (C21 to C29). Surprisingly, whereas all six of the
mimics’ compounds were shared with those present on ants, none were shared with those
of the non-mimic, though both mimic and non-mimic belong to the same spider family.

121
Based on the chemical data, ants and mimics were significantly different from non-
mimetic spiders (ANOSIM R=1 P<0.012); this difference was visualized by non-metric
multidimensional scaling (NMDS Stress =0.11).
DISCUSSION
Mimics fool visually-oriented predators
Results of our experiments clearly demonstrate that the ant-mimicking jumping spider
Peckhamia picata deceives visually-oriented predatory jumping spiders. Predatory
jumping spiders readily captured non-mimics, but treated mimics and ants similarly (Fig.
2). This result corroborates existing evidence that ant-mimicking spiders are effective
Batesian mimics (Cutler 1991, Nelson & Jackson 2006, Nelson et al. 2006). Jumping
spiders are well known for their elaborate vision-mediated hunting behavior (Forster
1982a, Jackson & Pollard 1996). But they are also known to use olfactory, vibrational
and tactile information along with or in the absence of visual cues (Forster 1982b, Taylor
et al. 1998, Clark et al. 2000). Myrmecophages, spiders that feed on ants use olfactory
cues to locate ants (Clark et al. 2000). However, use of olfaction to detect prey has not
been reported in non-ant-eating jumping spiders. Hoefler et al. (2002) found no evidence
of chemosensory response to prey by Phidippus audax. Furthermore, our preliminary
result, that Phidippus johnsoni stalk and capture hexane-washed house flies (with intact
visual cues, but lacking chemical cues), suggests that it is unlikely that P. johnsoni use

122
olfaction to hunt prey. (In our behavioral assays we have not ruled out the possibility of
predatory jumping spiders using vibrational cues to distinguish mimic and non-mimics)
Avoidance of ant mimics could be a learned or an innate behavior: predatory spiders
Phidippus sp. used in this experiment did not come from the same area from where the
mimic, model and non mimics were collected. However, they rarely captured P. picata,
implying that avoidance of mimic may be innate. If the model is nasty and widespread (as
many ants are), the predators need not have a priori experience with a particular mimic,
but can generalize across time and space (Ruxtom et al. 2004). Indeed, Batesian visual
mimicry can be innate (Nelson & Jackson 2006, Nelson et al. 2006).
Mimics escape detection by chemically-oriented mud-dauber wasps
When offered a simultaneous choice between non-mimics (Phidippus sp.) and mimics,
(Peckhamia), Sceliphron caemantarium wasps antennated and stung the non-mimics, but
antennated and rejected the mimics. This behavioral result would explain the absence of
ant-mimicking spiders from wasp nests -- examinations of the nests of mud-dauber wasps
over the past century have revealed that wasps mostly take araneids, but also capture
jumping spiders and crab spiders (Blackledge et al. 2003 and references there in).
However, Edmunds (1993) rarely found ant-mimicking jumping spiders in mud-daubers’
nests. Edmunds (1993) speculated that mud-dauber wasps are deceived by the mimics’
visual resemblance to ants. However, we now know that wasps rely on visual and
chemical cues at different stages of prey location and recognition (Uma & Weiss 2010).

123
Whereas visual cues seem to be important in locating a foraging habitat or spotting dark
colored or moving objects, chemical cues present on silk and spiders cuticle are important
for prey recognition by mud-dauber wasps (Uma & Weiss 2010). Our preliminary results
suggest that cuticular chemical profiles of araneids and salticids, though they belong to
unrelated families, are similar (Chapter 2). We had expected wasps to recognize
Peckhamia as prey, as they are also salticids, and spiders within one family often share
similar chemical signatures. However, our results indicated otherwise. We can identify
two non-mutually exclusive reasons for wasps failing to recognize ant-mimicking spiders
as prey. First, the mimics do not have a typical araneid/ salticid chemical profile--indeed,
the chemical signatures of Peckhamia and Phidippus are different (Fig. 3). Second, the
total amount of hydrocarbons present on mimic’s cuticle may be below wasps’ threshold
detection level. The total amount of hydrocarbons present on mimic’s cuticular surface
was a fifth of the non-mimic spider (preliminary results). Whether this amount of HC is
below a wasp’s detection level is not known. Further experiments are needed to separate
out these reasons.
Mimic resemble ant’s chemical profile
While the chemical signature of Peckhamia closely resembles that of its model ant
Camponotus sp, its profile doesn not match with Phidippus jonhsoni, the non-mimic,
though both are salticids (Fig 3, 4). Moreover, we found the total amount of
hydrocarbons present on the mimic’s cuticular surface was a tenth of the model ant. Thus,

124
ant mimic may rely on two different strategies, chemical mimicry and chemical
insignificance (being ‘odorless’) to stay close to ants.
Chemical insignificance, the deficiency of cuticular hydrocarbons, is often thought to
precede chemical mimicry (Lenoir et al. 2001). For example, obligate parasites of social
insects are chemically insignificant when they enter host colonies, and later acquire
colony-specific odor (Lorenzi & Bagneres 2002). On the other hand, chemical
insignificance and mimicry can coexist as a dual strategy in some scenarios (Kroiss et al.
2009). Myrmecomorphs such as Peckhamia are typically found outside ant colonies,
where they reside amidst their own models, as well as other ants in the same
microhabitat. Being chemically insignificant can ‘liberate’ the mimics to freely move
among different ants.
Though we have observed chemical mimicry/insignificance in only one ant-mimicking
spider, we believe it could be much more widespread across other spider and insect
myrmecomorphs. Further research is needed to examine the generality of such a
phenomenon.
Our results suggest that Peckhamia resemble the ant Camponotus’s chemical profile.
However, whether the chemical signals are perceived by ants is not known. Ants are not
only models for ant-mimicking spiders, but also potential predators (Nelson et al. 2004).
Not many studies that have looked into ants’ response towards ant-mimicking spiders;
however studies by Nelson et al. (2004, 05) suggest that ant-mimicking

125
(myrmecomorphs) and ant-eating spiders (myrmecophages) survive better than non-
mimicking salticids in the presence of ants. Theses spiders were lab-raised, and thus had
no prior contact with ants. Nelson et al. (2004, 05) attribute the survival of the mimic to
its behavioral and textural adaptation. However, ants mainly rely on chemical cues to
differentiate between nestmates and non-nestmates (Ozaki et al. 2005). We believe that
the higher rate of survival of ant-mimicking spiders in Nelson et al.’s study is due to
chemical resemblance (or insignificance) with the ants. It is likely that Peckhamia’s
chemical resemblance (and/or insignificance) may offer additional protection from
chemically-oriented ants.
Multimodal cues for multiple audiences
Much of the previous research on animal communication has examined signals in
isolation (i.e., only visual, chemical, or auditory, etc.) (reviewed in Hebets & Papaj
2005). However, recent research has revealed multimodal signaling to be extremely
common across different taxa in various contexts (reviewed in Uetz & Roberts 2002,
Hebets & Papaj 2005). Multimodal signaling targeted towards different audiences is
likely to evolve when different receivers require different kinds of signal information
(called the receiver variability hypothesis, Hebets & Papaj 2005). For example, female
satin bowerbirds of different ages use different male traits to choose different males
(Coelman et al. 2004). In this case, a complex signal can evolve to simultaniously
address age or stage specific preferences females have.

126
We have shown that ant mimic Peckhamia uses both visual and chemical modalities to
deceive different predators. To our knowledge, use of two signal modalities, each used to
fool different audience in two different classes of arthropods (spiders and insects) has not
been previously reported. We believe that the utilization of multimodal cues to deceive
multiple audiences will be extremely common, but has yet to be explored in mimicry
systems. Understanding multimodal signaling will reveal different selective pressures
under which such cues have evolved, and will help us understand the evolution of form
and function of complex signals.

127
REFERENCES
Aitchison, J. 1986: The statistical analysis of compositional data. Chapman & Hall,
London.
Allan, R. A., Capon, R. J., Brown, W. V. & Elgar, M. A. 2002: Mimicry of host cuticular
hydrocarbons by salticid spider Cosmophasis bitaeniata that preys on larvae of
tree ants Oecophylla smaragdina. 28(4), 835-848.
Blackledge, T. A., Coddington, J. A. & Gillespie, R. G. 2003: Are three-dimensional
spider webs defensive adaptations? Ecol. Lett. 6, 13-18.
Borg-Karlson, A. K., Berstrom, G., and Groth, I. 1985: Chemical basis for the
relationship between Ophrys orchids and their pollinators II. Volatile compounds
of Orphrys lutea and O. fusca as insect mimetic attractants/excitants. Chem. Scr.
25: 283-94.
Bohart, R. M. & Menke, A. S. 1976: Sphecid Wasps of the World. Berkeley: University
of California Press, California.
Chittka, L. & Osorio, D. 2007: Cognitive dimensions of predator responses to imperfect
mimicry? PloS Biol. 5(12), 2754-2758.
Clark, R. J., Jackson, R. R. & Cutler, B. 2000: Chemical cues from ants influence
predatory behavior in Habrocestum pulex, an ant-eating jumping spider (Araneae,
Salticidae). J. Arachnol. 28, 309-318.
Coleman S. W, Patricelli, G. L, Borgia, G 2004: Variable female preferences
drive complex male displays. Nature 428, 742-745.

128
Coville, R. E. 1987: Spider-hunting sphecid wasps. In: Ecophysiology of Spiders.
(Nentwig, W., ed). Springer-Verlag, Berlin. pp. 309-318.
Cushing, P. E. 1997: Myrmecomorphy and myrmecophily in spiders: a review. Florida
Entomol 80, 165–193.
Cutler, B. 1991: Reduced predation on the antlike jumping spider Synagelis occidentalis
(Araneae: Salticidae). J. Insect Behav. 4, 401–407.
Dettner, K. & Liepert, C. 1994: Chemical mimicry and camouflage. Ann. Rev. Entomol.
39, 129-154.
Edmunds, M. E. 1993: Does mimicry of ants reduce predation by wasps on salticid
spiders? Mem. Queens. Mus. 33, 507-12.
Forster, L. M. 1982a: Vision and prey catching strategies in jumping spiders.
Am. Sci. 70, 165-75.
Forester, L. M. 1982b: Non-visual prey-capture in Trite planiceps, a jumping spider
(Araneae, Salticidae). J. Arachnol.10, 179-183.
Hebets, E. A. & D. R. Papaj. 2005: Complex signal function: Developing a framework of
testable hypotheses. Beh. Eco. Sociobiol. 57, 197-214.
Herzner, G., Schmitt, T., Linsenmair, K. & Strohm, E. 2005: Prey recognition by females
of the European beewolf and its potential for a sensory trap. Anim. Behav. 70,
1411-1418.
Hoefler, C., Taylor, M. & Jakob, E. M. 2002: Chemosensory response to prey in
Phidippus audax (Araneae, Salticidae) and Pardosa milvina (Araneae,
Lycosidae). J. Arachnol. 30, 155-158.

129
Jackson, R. R. & Pollard, S. D. 1996: Predatory behavior of jumping spiders. Ann. Rev.
Entomol. 41, 287-308.
Kapan, D. D. 2001: Three-butterfly system provides a field test of Müllerian mimicry.
Nature. 409, 338-340.
Kroiss, J., Bordon, S. & Strohm, E. 2008: Hydrocarbons in the nest material of a solitary
digger wasp represent a kairomone for a specialized cuckoo wasp. Anim. Behav.
76, 1555-1563.
Lahav, S., Soroker,V., Hefetz, A. & Vander Meer, R. K. 1999: Direct behavioral
evidence for hydrocarbons as ant recognition discriminators. Naturwissenschaften
86(5), 246-249.
Lenoir, A., D’Ettorre, P. & Errard, C. 2001: Chemical ecology and social parasitisim in
ants. Ann. Rev. Entomol. 46, 573-591.
Lorenzi, M. C. & Bagneres, A. G. 2003: Concealing identity and mimicking hosts: a dual
chemical strategy for a single social parasite? (Polistes atrimandibularis,
Hymenoptera: Vespidae). Parasitology 125, 507-512.
McIver, J. D. & Stonedahl, G. 1993: Myrmecomorphy: morphological and behavioral
mimicry of ants. Ann. Rev. Entmol. 38, 351-379.
Nelson, X. J., Jackson, R. R., Pollard, S. D., Edwards, G. B. & Barrion, A. T. 2004:
Predation by ants on jumping spiders (Araneae: Salticidae) in the Philippines. N Z
J. Zool. 31, 45-56.
Nelson, X. J., Jackson, R. R., Edwards, G. B. & Barrion, A.T. 2005: Living with the
enemy: jumping spiders that mimic weaver ants. J. Arachnol. 33, 813–819.
Nelson, X. J., Jackson, R. R., Li, D., Barrion, A. T. & Edwards, G. B. 2006a:

130
Innate aversion to ants (Hymenoptera: Formicidae) and ant mimics: experimental
findings from mantises. Biol. J. Linn. Soc. 88, 23-32.
Nelson, X. J. & Jackson, R. R. 2006b: Vision-based innate aversion to ants and ant
mimics. Behav. Eco.17, 676-681.
Nelson, X. J. & Jackson, R. R. 2008: Anti-predator crèches and aggregations of ant-
mimicking jumping spiders (Araneae: Salticidae). Biol. J. Linn. Soc. 94, 475-481.
Ozaki, M., Wada-Katsumata, A., Fujikawa, K., Iwasaki, M., Yokohari, F., Satoji, Y.,
Nisimura, T. & Yamaoka, T. 2005: Ant nestmate and non-nestmate discrimination
by a chemosensory sensillum. Science. 309, 311-314.
Pierce, N. E., Braby, M. F., Heath, A., Lohman, D. J., Mathew, J., Rand, D. B. &
Travassos, M. A. 2002: The ecology and evolution of ant association in the
Lycaenidae (Lepidoptera). Ann. Rev. Entomol. 47, 733-471.
Rota, J. & Wagner, D. L. 2006: Predator mimicry: Metalmark moths mimic their jumping
spider predators. PloS ONE 1(1), e45 doi:10.1371/ journal.pone.0000045.
Ruxton, G. D. Sherratt, T. N. and Speed, M. P. 2004: Avoiding attack: the evolutionary
ecology of crypsis, warning signals, and mimicry. Oxford Biology
Schiestl, F. P., Ayasse, M., Paulus, H. F., Löfstedt, C., Hansson, B. C., Ibarra, F. and
Francke, W. 2000: Sex pheromone mimicry in the early spider orchid ( Ophrys
sphegodes ): patterns of hydrocarbons as the key mechanism for pollination by
sexual deception. J. comp. Phisiol A. 186 (6), 567-574.
Taylor, P. W., Jackson, R. R. & Robertson, M. B. 1998. A case of blind spider's buff?:
Prey-capture by jumping spiders (Araneae, Salticidae) in the absence of visual
cues. J. Arachnol. 26, 177-189.

131
Uetz, G. W. & Roberts, J. A. 2002: Multi-sensory cues and multi-modal communication
in spiders: insights from video/audio playback studies. Brain Behav. & Evol. 59,
222-230.
Uma, D. B. & Weiss, M. R. 2010: Chemical mediation of prey recognition by spider-
hunting wasps. Ethology. 116, 85-95.

132
Fig.1 Ant-mimicking spider, Peckhamia picata (Salticidae, left) and its model ant
Camponotus sp (Formicine, right). Peckhamia morphologically (size, color, shape) and
behaviorally (uses its second pair of legs as ‘antenna’) resembles its model ant.
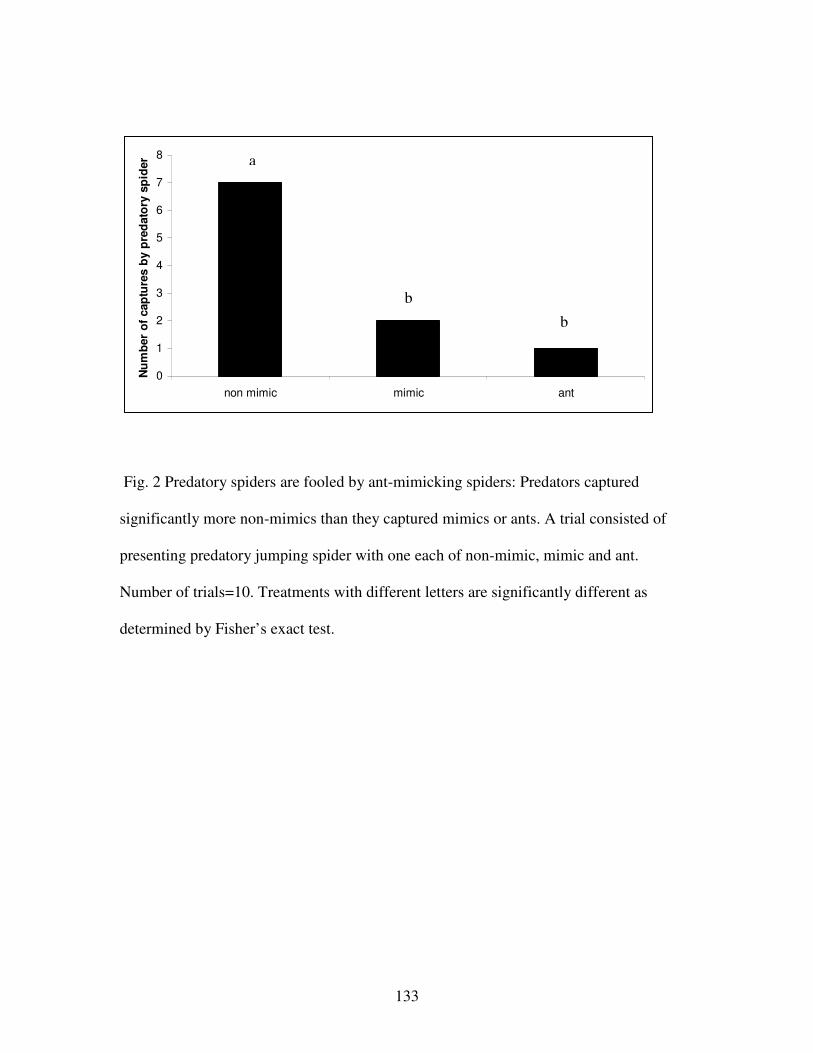
133
0
1
2
3
4
5
6
7
8
non mimic mimic ant
Nu
mb
er
of
cap
ture
s b
y p
red
ato
ry s
pid
er
Fig. 2 Predatory spiders are fooled by ant-mimicking spiders: Predators captured
significantly more non-mimics than they captured mimics or ants. A trial consisted of
presenting predatory jumping spider with one each of non-mimic, mimic and ant.
Number of trials=10. Treatments with different letters are significantly different as
determined by Fisher’s exact test.
a
b
b

134
Fig. 3. Chromatographs of ant (Camponotus, red), mimic (Peckhamia, blue) and non-
mimetic spider (Phidippus black). Mimics share all of their peaks with ants, but have
very reduced amounts. The chromatograph of the non-mimic is not clean, but it doesn’t
share its peaks with the mimic, though both are jumping spiders. (I will not be publishing
it like this… but this is just for the time being)

135
-0.15
-0.1
-0.05
0
0.05
0.1
-0.6 -0.4 -0.2 0 0.2 0.4 0.6
NMDS axis1
NM
DS
axis
2
Mimic
NON-MIMIC
Ant
Fig. 4. Nonmetric multi-dimensional scaling (NMDS) plot of ant, mimic and non-
mimetic spider cuticular chemical profiles. Stress is a ‘quality’ measure of NMDS.
(Usually stress is less than 0.2).





























