Embed Size (px)
Citation preview

Beyond ecological traps: perceptualerrors and undervalued resourcesJames J. Gilroy1 and William J. Sutherland2
1 School of Biological Sciences, University of East Anglia, Norwich, NR4 7TJ, UK2 Department of Zoology, University of Cambridge, Cambridge, CB2 3EJ, UK
Opinion TRENDS in Ecology and Evolution Vol.22 No.7
Glossary
Ecological trap: a sink habitat that is preferred above better available habitats.
Habitat preference: the selection decision that would be made if two different
habitats were available to an organism in equal quantity.
Pseudo-sink: a source habitat that has a population density higher than its
carrying capacity, such that it appears to function as a sink (i.e. has a temporary
negative population growth rate).
Settlement cues: habitat features that trigger evolved habitat preferences,
causing individuals to settle in a given site. These can be direct (i.e. influence
fitness explicitly) or indirect (i.e. correlate with features that influence fitness).
Sink: a habitat or site that supports a negative population growth rate, even at
small population densities.
Ecological traps arise when animals choose to settle inpoor-quality habitats, leading to a reduction in theirsurvival or productivity. Although this phenomenonhas received comprehensive theoretical treatment inthe recent literature, the corollary of the ecological trap(when animals choose to avoid good-quality habitats)is rarely discussed. Failure to recognize high-quality sitescould influence the ability of a population to reach itsthreshold size. An exploration of this phenomenonbrings new insights into existing cases of ecologicaltraps. By considering the full range of pitfalls facedduring settlement, the relationship between habitatquality and population size might become clearer. Weargue here that an improved understanding of factorsdetermining the colonization of high-quality sites couldaid conservationists in mitigating the damaging effectsof maladaptive habitat selection.
Perceptual errors and habitat selectionEcological traps occur when organisms choose poor-qualityhabitats abovebetteralternatives [1]. Thisphenomenonhasreceived increasing attention from researchers interested inthe relationship between population maintenance andhabitat alteration. Both theoretical [1–4] and field studies[5–7] have suggested that errors made in judging habitatquality influence the fate of populations under environmen-tal change. Examples include the wayward orientation ofsea turtle hatchlings in the presence of an artificial light [8],or the high rate of nest predation suffered by birds nestingpreferentially at the edges of forest fragments [6,9,10]. Ourunderstanding of ecological traps has advanced with thepublication of an expanding library of cases, together withthe development of analytical tools to diagnose their pre-sence and function [11–13]. The current focus of interest inecological traps might, however, be fostering a neglect ofalternative implications of maladaptive habitat selection.
Ecological traps are symptomatic of the rapid changesoccurring in natural landscapes worldwide. Many humanactivities are re-shaping the appearance and biologicalfunctioning of habitats, challenging native organisms toadapt to novel surroundings. If habitat alterations cause aformerly suitable area to become inhospitable [6,14,15]or if a poor-quality novel habitat mimics features of abetter one [8,16,17], organisms might be ‘trapped’ by theirevolved habitat preferences, settling in sites that cannot
Corresponding author: Gilroy, J.J. ([email protected]).Available online 9 April 2007.
www.sciencedirect.com 0169-5347/$ – see front matter � 2007 Elsevier Ltd. All rights reserve
support them. In the absence of rapid counteradaptation,these ecological traps might lead to extinction [1–4,18].However, populations might also be affected by a reversalof this phenomenon. If aminor changemakes a habitat lessattractive to settlers, without affecting the underlyingpattern of habitat quality, opportunities for colonizationmight go unexploited, creating what we call an under-valued resource (see Glossary). The principle is illustratedby the age-old tradition of using scarecrows to deter avianpests from settling on arable crop fields. The presence of acrude human effigy acts as an erroneous indicator of risk,discouraging settlement in an otherwise freely availablehabitat. The perceptual error in this example falls within asuite of behavioural phenomena that can influence thecolonization of good-quality sites, working either in tan-dem with ecological traps or in isolation.
Here, we review the conceptual basis for ecological trapsand show that, by considering other outcomes of maladap-tive habitat selection, we could gain new perspectiveson the use of human-altered environments by wild species.We go on to evaluate strategies that could be used to reducethe impacts of maladaptive habitat choice, highlighting thepotential for conservationists to exploit undervaluedresources to offset the negative effects of ecological traps.Finally, we consider the future role of research on mala-daptive habitat selection, focussing on the potential forexperimental manipulations of species distribution, bothas a conservation tool and as a means of studying habitat–productivity relationships.
Identifying ecological traps and undervaluedresourcesUnder their strictest definition, ecological traps are habitatsthat support net negative population growth rates (i.e.
Source: a habitat or site that supports a positive population growth rate.
Undervalued resource: a habitat that receives fewer settlers than it could
support under ideal conditions, owing to a lack of settlement cues.
d. doi:10.1016/j.tree.2007.03.014

352 Opinion TRENDS in Ecology and Evolution Vol.22 No.7
immigration exceeds emigration), but are preferred bysettlers above better alternative habitats, even at low popu-lation densities [1–4]. As such, true ecological traps functionas population sinks, having the potential to cause extinctioneven in the presence of abundant suitable habitat [3,4].These phenomena arise when the settlement cues used byorganisms in making preference choices become decoupledfrom underlying patterns of habitat quality, such that poorhabitats appear preferable to better ones.
Most studies of maladaptive habitat selection focus onthe selection of poor habitats [12]. However, populationsize might also be reduced if high-quality habitats lack theappropriate cues to encourage settlement. Such habitatsmight receive fewer settlers than they could support underideal free settlement conditions [19], creating an under-valued resource. Both forms of maladaptive habitat selec-tion can influence source–sink dynamics in complex ways[20]. Importantly, both phenomena could create imbal-ances between population density and the availability orquality of habitat, with predictable consequences for rea-lized population size (Box 1). Until now, these scenarioshave not been considered independently, although theirimplications for conservation might differ widely.
Quantifying the use and misuse of habitats
Ecological traps can be identified in the field by measuringpopulation processes and habitat preference [11–13].Undervalued resources might be more difficult to detect,as their symptoms are effectively non-events, wherebyorganisms fail to exploit available habitat opportunitiesfully. Conclusive evidence that a suitable habitat is beingunder used requires knowledge of the maximum carryingcapacity of the habitat, as well as any density-dependenteffect of population increase on individual fitness [22,23].Achieving this level of knowledge in a natural system isbeyond the scope of most field studies; however, the pre-sence of an undervalued resource could be demonstratedthrough experimentalmanipulation of the cues that encou-rage settlement.
In appraising habitat quality, organisms often rely oncues that can be assessed rapidly to estimate the suit-ability of sites, as time or energy constraints prevent thedirect evaluation of all the resources that they require [24].Often, these cues correlate with habitat quality indirectly,for example the presence or success of conspecifics [25], orthe presence of heterospecifics with similar ecologicalrequirements [26]. Alternatively, settlement cues mighthave direct effects on fitness [24]. Red-eyed vireos Vireoolivaceous, for example, are known to use foliage density asan indicator of habitat quality in woodlands, as this featuredirectly influences the abundance of their caterpillar prey[27]. In many species, cues are ‘imprinted’ during early lifestages in the natal habitat, a phenomenon known as natalhabitat preference induction (NHPI) [28]. Identification ofthe specific settlement cues used by organisms, eitherthrough detailed studies of settlement patterns or throughchoice experiments [13], is an essential stage in the studyof all forms of maladaptive habitat choice.
A first stage in the search for undervalued resourceswould be to consider known ecological trap cases. Manypublished examples of ecological traps have shown
www.sciencedirect.com
that poor habitats are preferred above more productivealternatives. It follows that these better habitats mightlack the appropriate cues to encourage settlement. Some ofthemost widely acknowledged examples of ecological trapsinvolve passerine birds breeding in North American wood-lands. Many species nest at higher densities close to wood-land edges, where they suffer high failure rates as a resultof increased predator or brood parasite activity in theseareas [6,9,10]. Woodland edges are thought to trigger anevolved habitat preference for natural light gaps that wereused by these species in ancient woodland mosaics [6,29].In modern forest fragments, the forest interior mightfunction as an undervalued resource, as it supports highbreeding productivity, but generally has low nest densitiesas it lacks the appropriate cue to encourage settlement[6,9]. However, this is not conclusive evidence that theforest interior is being used below carrying capacity, as themaximum threshold population size of the habitat isunknown. The settlement cue could, however, be exper-imentally introduced into the woodland interior by creat-ing small clear felled patches. If settlement rates were toincrease, without a subsequent reduction in breeding pro-ductivity, such an experiment would provide strong evi-dence for the existence of an undervalued resource. In fact,supporting evidence for this hypothesis has already beenprovided by studies of the effects of selective logging onpopulation regulation in edge-specialist birds [30,31], aswell studies of natural disturbance events, such as icestorms [32].
In principle, experimental manipulations of cuedistribution could be applied to any ecological trap whereresearchers have determined the cues that promote ordiscourage settlement. In practice, manipulation of thosecues might be feasible only in some cases.
Undervalued resources in isolation
If a change in habitat appearance causes a decline infavourability for settlement, without influencing under-lying habitat quality, the resulting habitat might functionas an undervalued resource. This scenario is most likely tooccur following the introduction of a novel habitat feature,or an entirely new habitat, to a natural landscape. Theexample of the farmer’s scarecrow illustrates this possib-ility: the novel feature (scarecrow) causes a decline infavourability of a field without influencing underlyinghabitat quality. Many human disturbances could createsimilar gaps in resource exploitation if the risks perceivedby potential settlers are not realized. For example, on UKheathlands, nesting woodlarks Lullula arborea showavoidance of areas surrounding busy footpaths [33]. Inreality, the risk posed by pedestrians to woodlarks isminimal, and the few individuals that do choose to settlein disturbed areas experience heightened breeding pro-ductivity in comparison to birds in other areas of the heath,owing to a release from density-dependent competition.Habitats surrounding these footpaths therefore representan undervalued resource, albeit on a small scale. Otherstudies have reported cases of recreational disturbanceinfluencing species distribution, often without presentinga genuine risk to survival or fitness of the species inquestion [34–36].
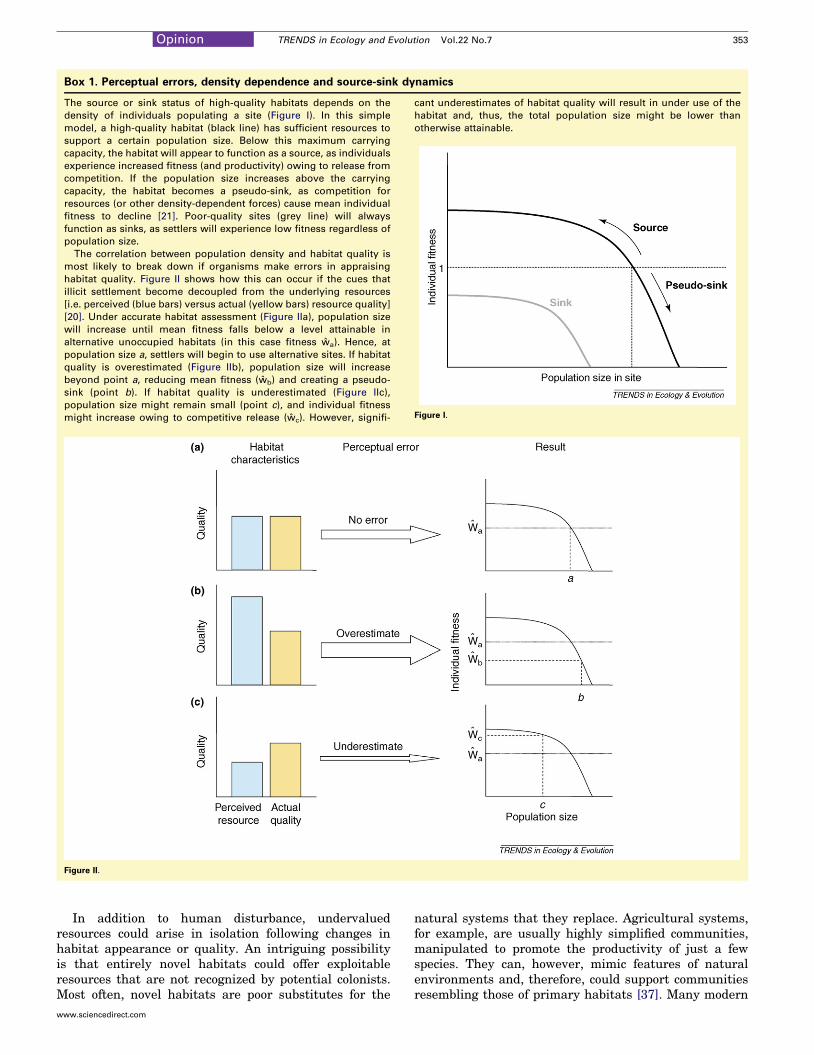
Box 1. Perceptual errors, density dependence and source-sink dynamics
The source or sink status of high-quality habitats depends on the
density of individuals populating a site (Figure I). In this simple
model, a high-quality habitat (black line) has sufficient resources to
support a certain population size. Below this maximum carrying
capacity, the habitat will appear to function as a source, as individuals
experience increased fitness (and productivity) owing to release from
competition. If the population size increases above the carrying
capacity, the habitat becomes a pseudo-sink, as competition for
resources (or other density-dependent forces) cause mean individual
fitness to decline [21]. Poor-quality sites (grey line) will always
function as sinks, as settlers will experience low fitness regardless of
population size.
The correlation between population density and habitat quality is
most likely to break down if organisms make errors in appraising
habitat quality. Figure II shows how this can occur if the cues that
illicit settlement become decoupled from the underlying resources
[i.e. perceived (blue bars) versus actual (yellow bars) resource quality]
[20]. Under accurate habitat assessment (Figure IIa), population size
will increase until mean fitness falls below a level attainable in
alternative unoccupied habitats (in this case fitness wa). Hence, at
population size a, settlers will begin to use alternative sites. If habitat
quality is overestimated (Figure IIb), population size will increase
beyond point a, reducing mean fitness (wb) and creating a pseudo-
sink (point b). If habitat quality is underestimated (Figure IIc),
population size might remain small (point c), and individual fitness
might increase owing to competitive release (wc). However, signifi-
cant underestimates of habitat quality will result in under use of the
habitat and, thus, the total population size might be lower than
otherwise attainable.
Figure I.
Figure II.
Opinion TRENDS in Ecology and Evolution Vol.22 No.7 353
In addition to human disturbance, undervaluedresources could arise in isolation following changes inhabitat appearance or quality. An intriguing possibilityis that entirely novel habitats could offer exploitableresources that are not recognized by potential colonists.Most often, novel habitats are poor substitutes for the
www.sciencedirect.com
natural systems that they replace. Agricultural systems,for example, are usually highly simplified communities,manipulated to promote the productivity of just a fewspecies. They can, however, mimic features of naturalenvironments and, therefore, could support communitiesresembling those of primary habitats [37]. Many modern
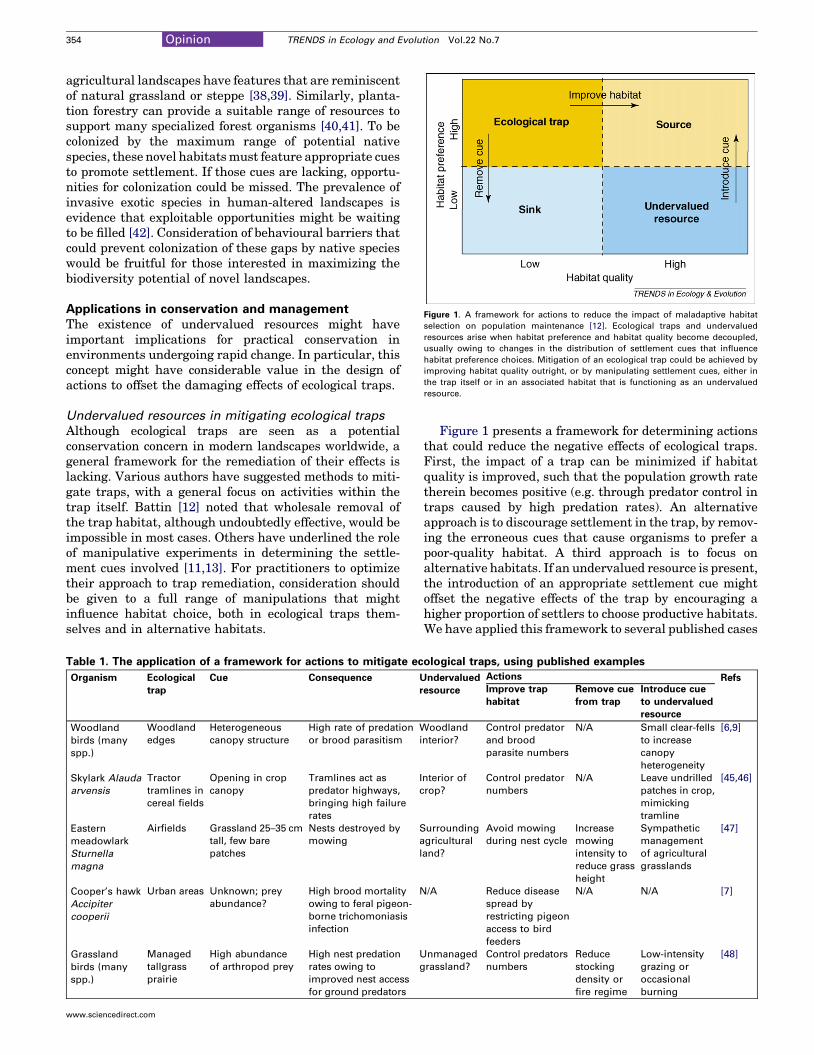
Figure 1. A framework for actions to reduce the impact of maladaptive habitat
selection on population maintenance [12]. Ecological traps and undervalued
resources arise when habitat preference and habitat quality become decoupled,
usually owing to changes in the distribution of settlement cues that influence
habitat preference choices. Mitigation of an ecological trap could be achieved by
improving habitat quality outright, or by manipulating settlement cues, either in
the trap itself or in an associated habitat that is functioning as an undervalued
resource.
354 Opinion TRENDS in Ecology and Evolution Vol.22 No.7
agricultural landscapes have features that are reminiscentof natural grassland or steppe [38,39]. Similarly, planta-tion forestry can provide a suitable range of resources tosupport many specialized forest organisms [40,41]. To becolonized by the maximum range of potential nativespecies, these novel habitatsmust feature appropriate cuesto promote settlement. If those cues are lacking, opportu-nities for colonization could be missed. The prevalence ofinvasive exotic species in human-altered landscapes isevidence that exploitable opportunities might be waitingto be filled [42]. Consideration of behavioural barriers thatcould prevent colonization of these gaps by native specieswould be fruitful for those interested in maximizing thebiodiversity potential of novel landscapes.
Applications in conservation and managementThe existence of undervalued resources might haveimportant implications for practical conservation inenvironments undergoing rapid change. In particular, thisconcept might have considerable value in the design ofactions to offset the damaging effects of ecological traps.
Undervalued resources in mitigating ecological traps
Although ecological traps are seen as a potentialconservation concern in modern landscapes worldwide, ageneral framework for the remediation of their effects islacking. Various authors have suggested methods to miti-gate traps, with a general focus on activities within thetrap itself. Battin [12] noted that wholesale removal ofthe trap habitat, although undoubtedly effective, would beimpossible in most cases. Others have underlined the roleof manipulative experiments in determining the settle-ment cues involved [11,13]. For practitioners to optimizetheir approach to trap remediation, consideration shouldbe given to a full range of manipulations that mightinfluence habitat choice, both in ecological traps them-selves and in alternative habitats.
Table 1. The application of a framework for actions to mitigate ec
Organism Ecological
trap
Cue Consequence U
r
Woodland
birds (many
spp.)
Woodland
edges
Heterogeneous
canopy structure
High rate of predation
or brood parasitism
W
i
Skylark Alauda
arvensis
Tractor
tramlines in
cereal fields
Opening in crop
canopy
Tramlines act as
predator highways,
bringing high failure
rates
I
c
Eastern
meadowlark
Sturnella
magna
Airfields Grassland 25–35 cm
tall, few bare
patches
Nests destroyed by
mowing
S
a
l
Cooper’s hawk
Accipiter
cooperii
Urban areas Unknown; prey
abundance?
High brood mortality
owing to feral pigeon-
borne trichomoniasis
infection
N
Grassland
birds (many
spp.)
Managed
tallgrass
prairie
High abundance
of arthropod prey
High nest predation
rates owing to
improved nest access
for ground predators
U
g
www.sciencedirect.com
Figure 1 presents a framework for determining actionsthat could reduce the negative effects of ecological traps.First, the impact of a trap can be minimized if habitatquality is improved, such that the population growth ratetherein becomes positive (e.g. through predator control intraps caused by high predation rates). An alternativeapproach is to discourage settlement in the trap, by remov-ing the erroneous cues that cause organisms to prefer apoor-quality habitat. A third approach is to focus onalternative habitats. If an undervalued resource is present,the introduction of an appropriate settlement cue mightoffset the negative effects of the trap by encouraging ahigher proportion of settlers to choose productive habitats.We have applied this framework to several published cases
ological traps, using published examples
ndervalued
esource
Actions RefsImprove trap
habitat
Remove cue
from trap
Introduce cue
to undervaluedresource
oodland
nterior?
Control predator
and brood
parasite numbers
N/A Small clear-fells
to increase
canopy
heterogeneity
[6,9]
nterior of
rop?
Control predator
numbers
N/A Leave undrilled
patches in crop,
mimicking
tramline
[45,46]
urrounding
gricultural
and?
Avoid mowing
during nest cycle
Increase
mowing
intensity to
reduce grass
height
Sympathetic
management
of agricultural
grasslands
[47]
/A Reduce disease
spread by
restricting pigeon
access to bird
feeders
N/A N/A [7]
nmanaged
rassland?
Control predators
numbers
Reduce
stocking
density or
fire regime
Low-intensity
grazing or
occasional
burning
[48]

Opinion TRENDS in Ecology and Evolution Vol.22 No.7 355
to illustrate the appraisal process (Table 1). Of the fiveexamples considered, removal of settlement cues in thetrap habitat was likely to be effective in just one case,whereas the introduction of cues to undervalued resourcesmight have been effective in four. Improving habitat qual-ity in a trap was deemed to be feasible in all five examples,generally involving the control of predator or brood para-site numbers to improve the success rate of nesting birds.Application of this framework reveals several key require-ments for effective ecological trap studies, including thespecific identification of settlement cues, consideration ofavailable alternative habitats, and quantification of thepopulation response to manipulations of both habitat qual-ity and cue distribution.
Future directionsExpansion of the ecological trap concept to includeundervalued resources provides a more complete concep-tual model of habitat use in human-altered environments.Although there is no conclusive evidence of the existence ofundervalued resources, we hope that the current interestin ecological traps, combined with a broadened view oftheir underlying principles, will confirm their inevitableexistence. Here, we have underlined the potential for usingexperimental manipulation to explore the relationshipbetween settlement cues and habitat quality. Futureadvances could be made in the measurement of cues andthe processes that trigger them, particularly through amultidisciplinary approach considering both behaviouralecology and cognitive processes [43,44]. Manipulation ofcues both in the traps themselves and in their alternativescould give rise to insights into the relationship betweenhabitat quality, cue recognition and the role of densitydependence in determining productivity. These lines ofenquiry will also contribute to conservation strategies inenvironments where populations are threatened by mala-daptive behaviour.
ConclusionsEcological traps could become increasingly prevalent asnatural communities acclimatize to continuing humanperturbation. Landscape conversion will inevitably leadto the loss of biodiversity, but the potential remains forthese altered landscapes to support important and diversewild communities. For this potential to be achieved, organ-ismsmust be capable of recognizing and colonizing suitablenovel habitats, without being drawn to false opportunitiesthat could lead to their decline. Organisms can adapt tonew environments, but the rate at which habitat is lostcould outstrip the potential for evolutionary response.Activities that mitigate the effects of ecological traps, orencourage settlement in suitable but unoccupied areas,could help to minimize the impact of habitat change onnatural environments. Further field and experimentalstudies of both ecological traps and undervalued resourceswill be essential for the success of future attempts toreconcile habitat change with the natural world.
AcknowledgementsJ.J.G. was supported by PhD funding from Natural England, RoyalSociety for the Protection of Birds and British Trust for Ornithology. We
www.sciencedirect.com
thank G.Q.A. Anderson, J.A. Vickery, P. Grice and A.C. Lees and fouranonymous referees for helpful comments.
References1 Delibes, M. et al. (2001) Attractive sinks, or how individual behavioural
decisions determine source-sink dynamics. Ecol. Lett. 4, 401–4032 Donovan, T.M. and Thompson, F.R., III (2001) Modelling the ecological
trap hypothesis: a habitat and demographic analysis for migrantsongbirds. Ecol. Appl. 11, 871–882
3 Kokko, H. and Sutherland, W.J. (2001) Ecological traps in changingenvironments: ecological and evolutionary consequences of abehaviourally mediated Allee effect. Evol. Ecol. Res. 3, 537–551
4 Kristan, W.B., III (2003) The role of habitat selection behavior inpopulation dynamics: source–sink systems and ecological traps.Oikos 103, 457–468
5 Dwernychuk, L.W. and Boag, D.A. (1972) Ducks nesting in associationwith gulls – ecological trap. Can. J. Zool. 50, 559–563
6 Gates, J.E. and Gysel, L.W. (1978) Avian nest dispersion and fledgingsuccess in field–forest ecotones. Ecology 59, 871–883
7 Boal, C.W. and Mannan, R.W. (1999) Comparative breeding ecology ofCooper’s hawks in urban and exurban areas of southeastern Arizona.J. Wildl. Manage. 63, 77–84
8 Witherington, B.E. (1997) The problem of photopollution for sea turtlesandothernocturnalanimals. InBehaviouralApproaches toConservationin the Wild (Clemmons, J.R. and Bucholz, R., eds), pp. 303–329,Cambridge University Press
9 Flaspohler, D.J. et al. (2001) Species-specific edge effects on nestsuccess and breeding bird density in a forested landscape. Ecol.Appl. 11, 32–46
10 Flaspohler, D.J. et al. (2001) Effects of forest edges on ovenbirddemography in amanaged forest landscape. Conserv. Biol. 15, 173–183
11 Schlaepfer, M.A. et al. (2002) Ecological and evolutionary traps. TrendsEcol. Evol. 17, 474–480
12 Battin, J. (2004) When good animals love bad habitats: ecological trapsand the conservation of animal populations. Conserv. Biol. 18, 1482–1491
13 Robertson, G.A. andHutto, R.L. (2006) A framework for understandingecological traps and an evaluation of existing evidence. Ecology 87,1075–1085
14 Chasko, G.G. and Gates, J.E. (1982) Avian habitat suitability along atransmission-line corridor in an oak-hickory forest region. Wildl.Monogr. 82, 5–41
15 Kolbe, J.J. and Janzen, F.J. (2002) Impact of nest-site selection on nestsuccess and nest temperature in natural and disturbed habitats.Ecology 83, 269–281
16 Kriska, G. et al. (1998) Why do mayflies lay their eggs en masse on dryasphalt roads? Water-imitating polarized light reflected from asphaltattracts Ephemeroptera. J. Exp. Biol. 201, 2273–2286
17 Remes, V. (2003) Effects of exotic habitat on nesting success, territorydensity, and settlement patterns in the blackcap (Sylvia atricapilla).Conserv. Biol. 17, 1127–1133
18 Donovan, T.M. and Lamberson, R.H. (2001) Area-sensitivedistributions counteract negative effects of habitat fragmentation onbreeding birds. Ecology 82, 1170–1179
19 Fretwell, S.D. and Lucas, H.L. (1970) On territorial behavior and otherfactors influencing habitat distribution in birds. I. Theoreticaldevelopment. Acta Biotheor. 19, 16–36
20 Remes, V. (2000) How can maladaptive habitat choice generate source-sink population dynamics? Oikos 91, 579–582
21 Watkinson, A.R. and Sutherland, W.J. (1995) Sources, sinks andpseudo-sinks. J. Anim. Ecol. 64, 126–130
22 Pulliam, H.R. (1988) Sources, sinks, and population regulation. Am.Nat. 132, 652–661
23 Pulliam, H.R. and Danielson, B.J. (1991) Sources, sinks, and habitatselection - a landscape perspective on population-dynamics. Am. Nat.137, S50–S66
24 Stamps, J. and Krishnan, V.V. (2005) Nonintuitive cue use in habitatselection. Ecology 86, 2860–2867
25 Doligez, B. et al. (2002) Public information and breeding habitatselection in a wild bird population. Science 297, 1168–1170
26 Thomson, R.L. et al. (2003) Positive interactions between migrant andresident birds: testing the heterospecific attraction hypothesis.Oecologia 134, 431–438

356 Opinion TRENDS in Ecology and Evolution Vol.22 No.7
27 Marshall, M.R. and Cooper, R.J. (2004) Territory size of a migratorysongbird in response to caterpillar density and foliage structure.Ecology 85, 432–445
28 Davis, J.M. and Stamps, J.A. (2004) The effect of natal experience onhabitat preferences. Trends Ecol. Evol. 19, 411–416
29 Morgan, K.A. and Gates, J.E. (1982) Bird population-patterns in forestedge and strip vegetation at Remington Farms, Maryland. J. Wildl.Manage. 46, 933–944
30 Webb, W.L. et al. (1977) Effect of logging on songbird populations in anorthern hardwood forest. Wildl. Monogr. 55, 1–35
31 Robinson, W.D. and Robinson, S.K. (1999) Effects of selective loggingon forest bird populations in a fragmented landscape.Conserv. Biol. 13,58–66
32 Faccio, S.D. (2003) Effects of ice storm-created gaps on forest breedingbird communities in central Vermont. For. Ecol. Manage. 186, 133–145
33 Mallord, J.W. et al. (2007) Predicting the consequences of recreationaldisturbance for a woodlark (Lullula arborea) population on heathlandsin southern England. J. Appl. Ecol. 44, 185–195
34 Yalden, D.W. (1992) The influence of recreational disturbance oncommon sandpipers Actitis hypoleucos breeding by an uplandreservoir, in England. Biol. Conserv. 61, 41–49
35 Brown, B.T. and Stevens, L.E. (1997) Winter bald eagle distribution isinversely correlated with human activity along the Colorado River,Arizona. J. Raptor Res. 31, 7–10
36 Rodriguez-Prieto, I. and Fernandez-Juricic, E. (2005) Effects of directhuman disturbance on the endemic Iberian frog Rana iberica atindividual and population levels. Biol. Conserv. 123, 1–9
Endeav
The quarterly magazine
philosophy o
You can access Ende
ScienceDirect, where you
editorial comment and a c
illustrated articles on th
Featuri
Information revolution: William ChambeDoes history count?
Waking up to shell shock: psychiatry in the UDeserts on the sea floor: Edward Forbes and his azoic hypoth‘Higher, always higher’: technology, the military and aviation m
Bully for Apatosauru
Coming
Environmentalism out of the IndusPandemic in print: the spread of influen
Earthquake theories in the early mScience in fiction - attempts to make a scie
The birth of botanical Dro
And much, mu
Endeavour is available on Science
www.sciencedirect.com
37 Green, R.E. et al. (2005) Farming and the fate of wild nature. Science307, 550–555
38 Bignal, E.M. and McCracken, D.I. (1996) Low-intensity farmingsystems in the conservation of the countryside. J. Appl. Ecol. 33,413–424
39 Kleijn, D. and Sutherland, W.J. (2003) How effective are Europeanagri-environment schemes in conserving and promoting biodiversity?J. Appl. Ecol. 40, 947–969
40 Hartley, M.J. (2002) Rationale andmethods for conserving biodiversityin plantation forests. For. Ecol. Manage. 155, 81–95
41 Carnus, J.M. et al. (2006) Planted forests and biodiversity. J. For. 104,65–77
42 Mack, R.N. et al. (2000) Biotic invasions: causes, epidemiology, globalconsequences, and control. Ecol. Appl. 10, 689–710
43 Healy, S. and Braithwaite, V. (2000) Cognitive ecology: a field ofsubstance? Trends Ecol. Evol. 15, 22–26
44 Farina, A. and Belgrano, A. (2006) The eco-field hypothesis: toward acognitive landscape. Landscape Ecol. 21, 5–17
45 Donald, P.F. et al. (2002) Survival rates, causes of failure andproductivity of skylark Alauda arvensis nests on lowland farmland.Ibis 144, 652–664
46 Odderskær, P. et al. (1997) Skylark (Alauda arvensis) utilisation ofmicro-habitats in spring barley fields.Agric. Ecosyst. Environ. 62, 21–29
47 Kershner, E.L. and Bollinger, E.K. (1996) Reproductive success ofgrassland birds at east-central Illinois airports. Am. Midl. Nat. 136,358–366
48 Shochat, E. et al. (2005) Ecological traps in isodars: effects of tallgrassprairie management on bird nest success. Oikos 111, 159–169
our
for the history and
f science.
avour online on
’ll find book reviews,
ollection of beautifully
e history of science.
ng:
rs, the publishing pioneer by A. Fyfe
by K. Anderson
S military during World War II by H. Pols
esis for a lifeless deep ocean by T.R. Anderson and T. Rice
edicine during the age of the two world wars by C. Kehrt
s by P. Brinkman
soon:
trial Revolution by C. Macleod
za in the Fin de Siecle by J. Mussell
odern period by F. Willmoth
nce out of literary criticism by J. Adams
sophila by S. Leonelli
ch more. . .
Direct, www.sciencedirect.com