Embed Size (px)
Citation preview

Biochimica et Biophysica Acta 850 (1986) 359-368 359 Elsevier
BBA 42042
B i n d i n g a n d h y d r o l y s i s o f 2 - a z i d o - A T P a n d 8 - a z i d o - A T P by i so l a t ed
m i t o c h o n d r i a l FI: c h a r a c t e r i s a t i o n o f h i g h - a f f i n i t y b i n d i n g s i tes
M . B . M . v a n D o n g e n , J .P. de G e u s , T. K o r v e r , A . F . H a r t o g a n d J .A. B e r d e n
Laboratory of Biochemistry, Unioersity of Amsterdam, P.O. Box 20151, 1000 HD A msterdam (The Netherlands)
(Received January 8th, 1986) (Revised manuscript received March 28th, 1986)
Key words: Photoaffinity labelling; Nucleotide binding; ATP-analogue; ATP synthase; Azido-ATP
The kinetic parameters for the hydrolysis by F 1 of the photoreactive nucleotide analogue 2-azido-ATP were determined (Vma x, 105 U / r a g Fl; K m, 250 pM, in the presence of 1.0 mM SO~-). In the absence of an activating anion, a non-linear relationship in a Lineweaver-Burk plot was found for the hydrolysis of 2-azido-ATP. The 2-azido-analogues of ATP and ADP proved to be good photoaffinity labels causing notable inactivation of the Ft-ATPase activity upon irradiation at 360 nm. This inhibition was also used to demonstrate high-affinity binding of these analogues to a catalytic binding site on the F t. High-affinity binding proved to be an Mg2+-requiring process, occurring with both 2-azido-ATP and 2-azido-ADP but hardly or not occurring with 8-azido-AT(D)P. Covalent binding of 2-nitreno-ATP upon irradiation of F I containing tightly bound [fl-S2P]2-azido-ATP results in a proportional inhibition of ATPase activity, extrapolating to 0.92 mol of covalently bound label per mol of F t needed for the complete inactivation of the enzyme. When the F 1 was irradiated in the presence of excess [fl-S2P]2-azido-AT(D)P, 3-4 tool of label were bound when the enzyme was fully inactivated. In all cases, all or most of the radioactivity was found on the fl subunits.
Introduction
The F 1 part of the ATP synthase complex is composed of five different subunits in the follow- ing stoicheiometry ot 3, f13, •, 3, c [1]. It has been demonstrated that the six nucleotide binding sites present on the F 1 are located on the a and fl subunits [2-4]. The nucleotide binding sites can be divided in catalytic and non-catalytic binding sites, the actual number of catalytic sites having been a matter of debate for a long time [2,5,6]. Evidence has accumulated that the catalytic sites are located on the fl subunits [4,7], in agreement with the
Abbreviations: 8(2)-N-AT(D)P, 8(2)-nitrenoadenosine-5'- tri(di)-phosphate; Mes, 4-morpliolineethanesulphonic acid; Mops, 4 morpholinepropanesulphonic acid.
finding that most ATPase inhibitors interact with the fl subunits.
Besides, isolated F 1 is known to contain 2 or 3 nucleotides tightly bound to the enzyme [2,3,8,9]. The function and the localisation of these tightly bound nucleotides is currently being investigated in our laboratory, the question being if and how they are involved in the reaction mechanism of ATP-hydrolysis and -synthesis. To investigate this, the photoreactive nucleotides 2-azido-AT(D)P and 8-azido-AT(D)P are used. With the 8-azido ana- logues, catalytic and non-catalytic binding sites could be demonstrated on the F 1, located on the fl subunits and the interfaces between ct and fl sub- units, respectively [7,10]. It was also shown that the tightly bound nucleotides cannot be ex- changed with the 8-azido nucleotides [2,7,10]. As
0005-2728/86/$03.50 © 1986 Elsevier Science Publishers B.V. (Biomedical Division)

360
the tightly bound nucleotides were shown to ex- change very slowly with medium nucleotides [11], and as the function of the tight nucleotide binding sites appeared to be at most regulatory [12], they were not considered to be directly involved in catalysis. This is supported by experiments with nucleotide-depleted F 1 which could bind five molecules of [14C]ATP per F1, only two of which are exchangeable [3]. On the other hand, evidence is accumulating that tight binding may be an intermediary phase in the reaction mechanism of catalysis [5,7,13-17].
As at least two catalytic sites could be demon- strated on two fl subunits with 8-azido-AT(D)P [2,7,10,18], it is theoretically possible that the third fl subunit contains a tightly bound nucleotide involved in catalysis of conversion of substrates that can be bound tightly to the enzyme. It has been proposed [19] that for tight binding an 'anti ' configuration of the bound nucleotide is required. This made it necessary to expand our studies with azido analogues in the 'anti ' configuration. In the present paper, it is shown that the very hydrolys- able 2-azido-ATP is also a valuable photoaffinity label which can be bound with high affinity, due to its 'anti ' configuration. It is demonstrated that this tight binding to a catalytic site is an inter- mediate phase in the reaction mechanism.
Materials and Methods
Bovine heart mitochondrial F 1 was isolated according to Knowles and Penefsky [20] and stored in liquid nitrogen in a medium comprising 10 mM Tris-HC1 (pH 7.5), 4 mM ATP, 250 mM sucrose and 4 mM EDTA. Prior to use, F 1 was subjected to ammonium sulphate precipitation and was sub- sequently filtered three times by centrifugation through a column (Penefsky column [21]) which contained Sephadex G-50 coarse pre-equilibrated in the same medium, except that ATP was omitted (medium A). In some of the experiments, the Sephadex used was pre-equilibrated in medium in which EDTA was replaced by 6 mM MgC12 (medium B). When optimal turnover conditions (ATP hydrolysis) were needed for the experiment, 10 mM HCO~- was added to medium B and the pH was raised to 8.
The photoaffinity labels 2-azido-ATP and 2-
azido-ADP were synthesized in our laboratory; 2-chloroadenosine was mixed with anhydrous hy- drazine under a nitrogen atmosphere at room tem- perature for 16 h according to the method of Schaefer and Thomas [22]. The 2-hydrazine- adenosine formed was isolated by evaporating the hydrazine and, after coevaporating residual hy- drazine with added isopropanol, was subsequently solubilised in 5% acetic acid. After the addition of the sodium nitrite solution at 0°C, the 2-azido- adenosine started to crystallise after 10 min. The 2-azidoadenosine (75%) was isolated on a Biachner filter, washed with cold water and dried over phosphorpentoxide. Phosphorylation of 2-azido- adenosine was carried out as described by Sowa and Ouchi [23]. 2-Azidoadenosine 5'-monophos- phate (60%) was isolated and purified on a DEAE-Sephadex A-25 column (50 x 2 cm) by eluting with a 0-300 mM triethylamine-HCO 3 gradient (pH 7.8). Synthesis of 2-azido-ADP and 2-azido-ATP was carried out as described by Hoard and Ott [24], starting with a dry solution of 2-azido-AMP as a tributylamine salt in dimethyl- formamide. After purification on a DEAE-Se- phadex A-25 column (50 X 2 cm) with 100-800 mM triethylamine-HCO 3 gradient (pH 7.8), 50-60% of the purified product was solubilised in acetone ethanol. After addition of LiC1, the Li salt was centrifuged and solubilised in a 10 mM Tris- HC1 solution (pH 7.0). The azido analogues were stored at - 2 0 ° C . Spectral and analytical data were in agreement with those published by Czar- necki et al. [25]. [fl-32p]2-azido-ADP was synthe- sized in the same way, using 10 mCi [32p]p i (45 /~mol). About 30% of the 2-azido-AMP (20/~mol) was transformed into the labelled compound which was phosphorylated in [fl-32p]2-azido-ATP using the phosphoenolpyruvate/pyruvate kinase sys- tem. 8-Azido-AT(D)P and [2-3H]8-azido-AT(D)P were synthesized as described previously [26].
ATP hydrolysis activity was measured spectro- photometrically by following the oxidation of NADH in an ATP-regenerating system as de- scribed by Sloothaak et al. [26]. For K m de- terminations, the ATPase medium contained vary- ing concentrations of MgATP, Mg-8-azido-ATP or Mg-2-azido-ATP. Furthermore, the pyruvate kinase concentration was raised 5-times and 10 mM HCO 3 or 1 mM SO32- was added as an
activating anion. ATP hydrolysis, as measured by phosphate production, was performed by taking samples from the Fl-containing reaction medium at several time intervals. The amount of phosphate produced was measured as described by Fiske and Subbarow [27].
Photoaffinity labelling was performed by irradiation at room temperature at 360 nm using a CAMAG Universal ultraviolet lamp. Irradiation of tightly incorporated 2-azido-nucleotides was performed on hollow sheets of aluminium. The total amount of covalently bound [fl-32p]2-N- AT(D)P was measured after heat denaturation of the F 1 (5 min, 95 ° C) and centrifugation by count- ing the radioactivity in the pellet. The distribution of radioactivity on a and /3 subunits was determined via SDS-urea polyacrylamide gel elec- trophoresis as described previously [7].
Protein concentrations were determined as described by Lowry et al. [28], using bovine serum albumin as a standard. ATP, phosphoenolpyru- vate, lactate dehydrogenase and NADH were purchased from Boehringer Mannheim. All other chemicals used were of analytical grade.
Results
Hydrolysis The nucleotide analogue 2-azido-ATP can be
considered as a better analogue of ATP than 8-azido-ATP. This is illustrated in Table I where K m and Vma x values are reported for the steady- state hydrolysis of ATP and the two photoreactive nucleotides 2-azido-ATP and 8-azido-ATP. It can be seen that the Vm~ x value for the hydrolysis of 2-azido-ATP approaches a value much closer to that of ATP than the Vma x value 8-azido-nucleo- tide. Furthermore, it has to be taken into account that the 2-azido-ATP is in equilibrium with tetra- zolic iso-mers: under the conditions used (aqueous solution, pH 8.0) about 55% can be expected to be in one of these two tetrazolic forms [25]. The real K m value for 2-azido-ATP, as reported in Table I, is, therefore, about 2.2-times lower than the mea- sured apparent value.
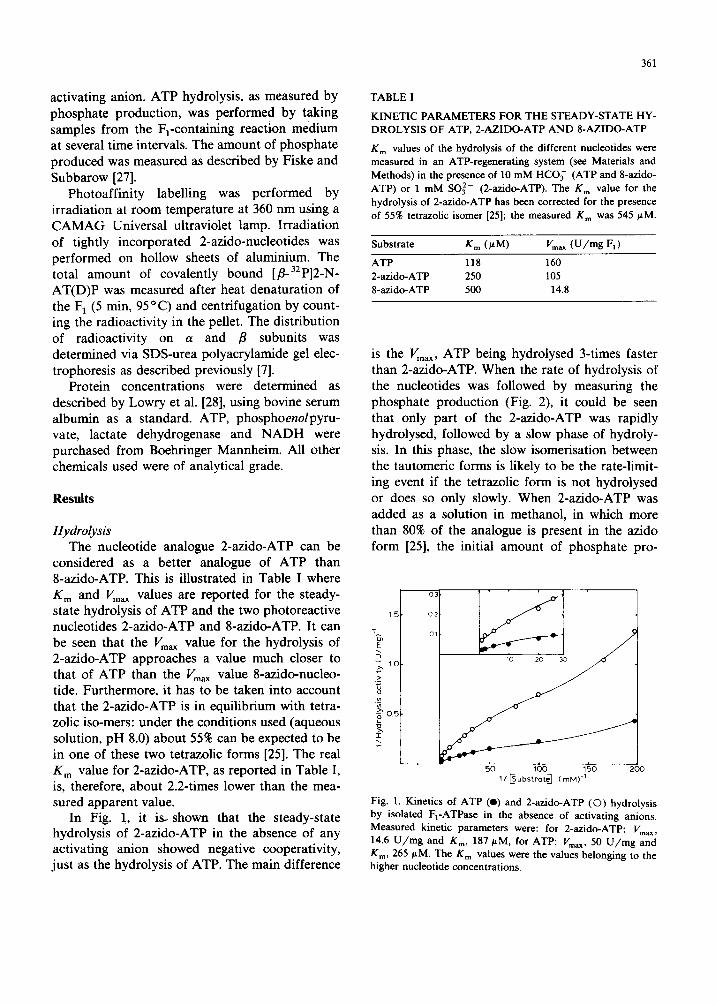
In Fig. 1, it is .shown that the steady-state hydrolysis of 2-azido-ATP in the absence of any activating anion showed negative cooperativity, just as the hydrolysis of ATP. The main difference
361
TABLE I
KINETIC PARAMETERS FOR THE STEADY-STATE HY- DROLYSIS OF ATP, 2-AZIDO-ATP A N D 8-AZIDO-ATP
K m values of the hydrolysis of the different nucleotides were measured in an ATP-regenerating system (see Materials and Methods) in the presence of 10 mM HCO 3 (ATP and 8-azido- ATP) or 1 m M SO 2 - (2-azido-ATP). The K m value for the hydrolysis of 2-azido-ATP has been corrected for the presence of 55% tetrazolic isomer [25]; the measured K m was 545 btM.
Substrate K m (p.M) F~. ( U / m g F1)
ATP 118 160 2-azido-ATP 250 105 8-azido-ATP 500 14.8
is the Vmax, ATP being hydrolysed 3-times faster than 2-azido-ATP. When the rate of hydrolysis of the nucleotides was followed by measuring the phosphate production (Fig. 2), it could be seen that only part of the 2-azido-ATP was rapidly hydrolysed, followed by a slow phase of hydroly- sis. In this phase, the slow isomerisation between the tautomeric forms is likely to be the rate-limit- ing event if the tetrazolic form is not hydrolysed or does so only slowly. When 2-azido-ATP was added as a solution in methanol, in which more than 80% of the analogue is present in the azido form [25], the initial amount of phosphate pro-
15
1.0
_>
0 5
T
o3!
02
0 1
5 0 100 150 2 0 0 1 / ~ u b s t r ' o t ~ ( r n M ) -1
Fig. 1. Kinetics of ATP (e) and 2-azido-ATP (©) hydrolysis by isolated F1-ATPase in the absence of activating anions. Measured kinetic parameters were: for 2-azido-ATP: Vmax, 14.6 U / m g and Km, 187/~M, for ATP: Vm~ , 50 U / m g and K m, 265 #M. The Km values were the values belonging to the higher nucleotide concentrations.
362
~ 80 I~~ Z
8 ~_ 6Ol- ~
I z 2 0
0 2 4 6 B 10 ! | I i Incubation - t ime (minutes) 0 5 10 15 20
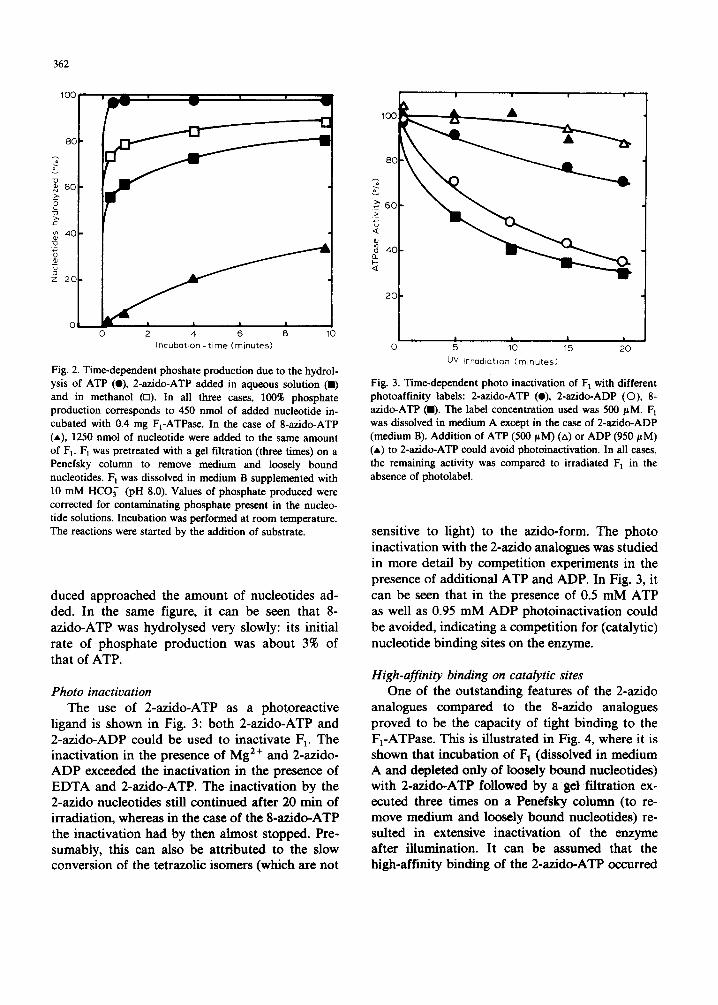
Fig. 2. Time-dependent phoshate production due to the hydrol- ysis of ATP (O), 2-azido-ATP added in aqueous solution (11) and in methanol (m). In all three cases, 100% phosphate production corresponds to 450 nmol of added nucleotide in- cubated with 0.4 mg F1-ATPase. In the case of 8-azido-ATP (*), 1250 nmol of nucleotide were added to the same amount of F~. 1=1 was pretreated with a gel filtration (three times) on a Penefsky column to remove medium and loosely bound nucleotides. F~ was dissolved in medium B supplemented with 10 mM HCO~- (pH 8.0). Values of phosphate produced were corrected for contaminating phosphate present in the nucleo- tide solutions. Incubation was performed at room temperature. The reactions were started by the addition of substrate.
duced approached the amount of nucleotides ad- ded. In the same figure, it can be seen that 8- az ido-ATP was hydrolysed very slowly: its initial rate of phosphate product ion was about 3% of that of ATP.
Photo inactivation The use of 2-azido-ATP as a photoreact ive
ligand is shown in Fig. 3: bo th 2-azido-ATP and 2-az ido-ADP could be used to inactivate 1::1. The inactivation in the presence of Mg 2+ and 2-azido- A D P exceeded the inactivation in the presence of E D T A and 2-azido-ATP. The inactivation by the 2-azido nucleotides still cont inued after 20 rain of irradiation, whereas in the case of the 8-azido-ATP the inactivation had by then almost stopped. Pre- sumably, this can also be at tr ibuted to the slow conversion of the tetrazolic isomers (which are not
UV Irradiat ion (minutes)
Fig. 3. Time-dependent photo inactivation of F 1 with different photoaffinity labels: 2-azido-ATP (e), 2-azido-ADP (O), 8- azido-ATP (11). The label concentration used was 500 pM. F 1 was dissolved in medium A except in the case of 2-azido-ADP (medium B). Addition of ATP (500/~M) (zx) or ADP (950 pM) (A) to 2-azido-ATP could avoid photoinactivation. In all cases, the remaining activity was compared to irradiated F 1 in the absence of photolabel.
sensitive to light) to the azido-form. The photo inactivation with the 2-azido analogues was studied in more detail by competi t ion experiments in the presence of addit ional A T P and ADP. In Fig. 3, it can be seen that in the presence of 0.5 m M ATP as well as 0.95 m M A D P photoinact ivat ion could be avoided, indicating a competi t ion for (catalytic) nucleotide b inding sites on the enzyme.
High-affinity binding on catalytic sites One of the outs tanding features of the 2-azido
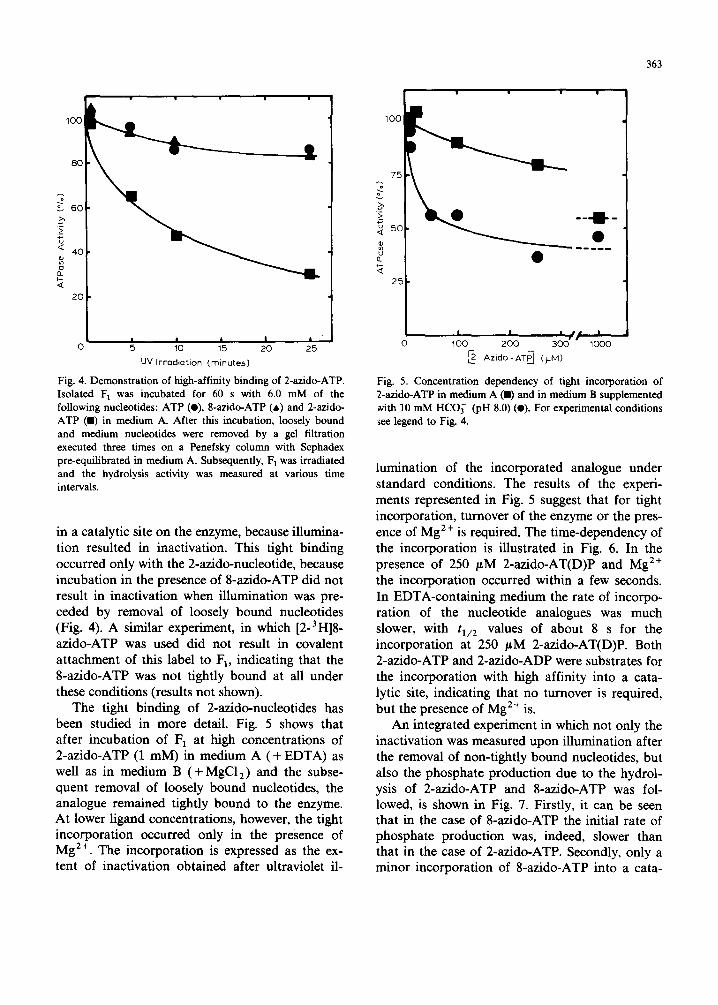
analogues compared to the 8-azido analogues proved to be the capacity of tight binding to the F1-ATPase. This is illustrated in Fig. 4, where it is shown that incubat ion of F 1 (dissolved in medium A and depleted only of loosely bound nucleotides) with 2-azido-ATP followed by a get filtration ex- ecuted three times on a Penefsky column (to re- move medium and loosely bound nucleotides) re- suited in extensive inactivation of the enzyme after illumination. It can be assumed that the high-affinity binding of the 2-azido-ATP occurred
363
1 1 0 0 ~
i i I e,.e • ---m--
o loo 3od; 17oo o 5 10 15 20 25 uv Irradiation (minutes)
Fig. 4. Demonstration of high-affinity binding of 2-azido-ATP. Isolated F 1 was incubated for 60 s with 6.0 mM of the following nucleotides: ATP (O), 8-azido-ATP (a) and 2-azido- ATP (11) in medium A. After this incubation, loosely bound and medium nucleotides were removed by a gel filtration executed three times on a Penefsky column with Sephadex pre-equilibrated in medium A. Subsequently, F 1 was irradiated and the hydrolysis activity was measured at various time intervals,
in a catalytic site on the enzyme, because illumina- tion resulted in inactivation. This tight binding occurred only with the 2-azido-nucleotide, because incubation in the presence of 8-azido-ATP did not result in inactivation when illumination was pre- ceded by removal of loosely bound nucleotides (Fig. 4). A similar experiment, in which [2-3H]8 - azido-ATP was used did not result in covalent attachment of this label to F 1, indicating that the 8-azido-ATP was not tightly bound at all under these conditions (results not shown).
The tight binding of 2-azido-nucleotides has been studied in more detail. Fig. 5 shows that after incubation of 1=1 at high concentrations of 2-azido-ATP (1 mM) in medium A (+ EDTA) as well as in medium B (+ MgC12) and the subse- quent removal of loosely bound nucleotides, the analogue remained tightly bound to the enzyme. At lower ligand concentrations, however, the tight incorporation occurred only in the presence of Mg 2÷. The incorporation is expressed as the ex- tent of inactivation obtained after ultraviolet il-
I~ - Azido - ATP--J (}JM)
Fig. 5. Concentration dependency of tight incorporation of 2-azido-ATP in medium A (11) and in medium B supplemented ~¢ith 10 mM HCO~- (pH 8.0) (O). For experimental conditions ~ee legend to Fig. 4.
lumination of the incorporated analogue under standard conditions. The results of the experi- ments represented in Fig. 5 suggest that for tight incorporation, turnover of the enzyme or the pres- ence of Mg 2+ is required. The time-dependency of the incorporation is illustrated in Fig. 6. In the presence of 250 /~M 2-azido-AT(D)P and Mg 2+ the incorporation occurred within a few seconds. In EDTA-containing medium the rate of incorpo- ration of the nucleotide analogues was much slower, with 11/2 v a l u e s of about 8 s for the incorporation at 250 /~M 2-azido-AT(D)P. Both 2-azido-ATP and 2-azido-ADP were substrates for the incorporation with high affinity into a cata- lytic site, indicating that no turnover is required, but the presence of Mg 2+ is.
An integrated experiment in which not only the inactivation was measured upon illumination after the removal of non-tightly bound nucleotides, but also the phosphate production due to the hydrol- ysis of 2-azido-ATP and 8-azido-ATP was fol- lowed, is shown in Fig. 7. Firstly, it can be seen that in the case of 8-azido-ATP the initial rate of phosphate production was, indeed, slower than that in the case of 2-azido-ATP. Secondly, only a minor incorporation of 8-azido-ATP into a cata-
364
100
8 0
60
< 40
o_
2 0
0
• i
- / - ' 8
I I / I •
2 0 4 0 6 0 " " 120
Incuba t ion t i m e ( seconds )
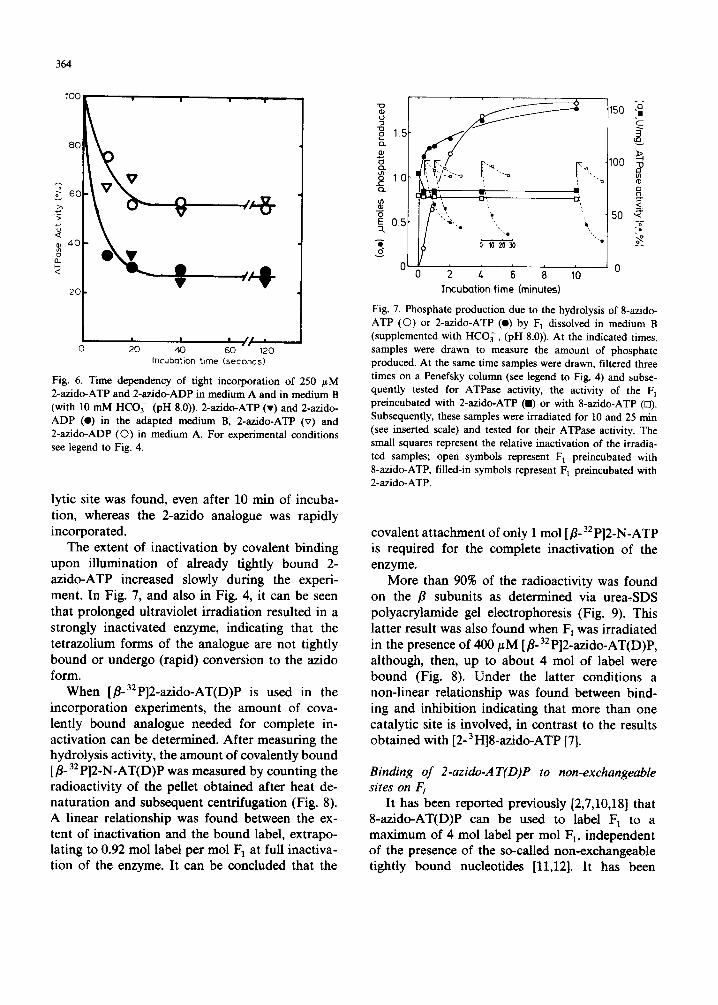
Fig. 6. Time dependency of tight incorporation of 250 /~M 2-azido-ATP and 2-azido-ADP in medium A and in medium B (with 10 mM HCO~- (pH 8.0)). 2-azido-ATP (v) and 2-azido- ADP (I) in the adapted medium B, 2-azido-ATP (v) and 2-azido-ADP (O) in medium A. For experimental conditions see legend to Fig. 4.
lytic site was found, even after 10 min of incuba- tion, whereas the 2-azido analogue was rapidly incorporated.
The extent of inactivation by covalent binding upon i l lumination of already tightly bound 2- az ido-ATP increased slowly during the experi- ment. In Fig. 7, and also in Fig. 4, it can be seen that prolonged ultraviolet irradiation resulted in a strongly inactivated enzyme, indicating that the tetrazolium forms of the analogue are not tightly bound or undergo (rapid) conversion to the azido form.
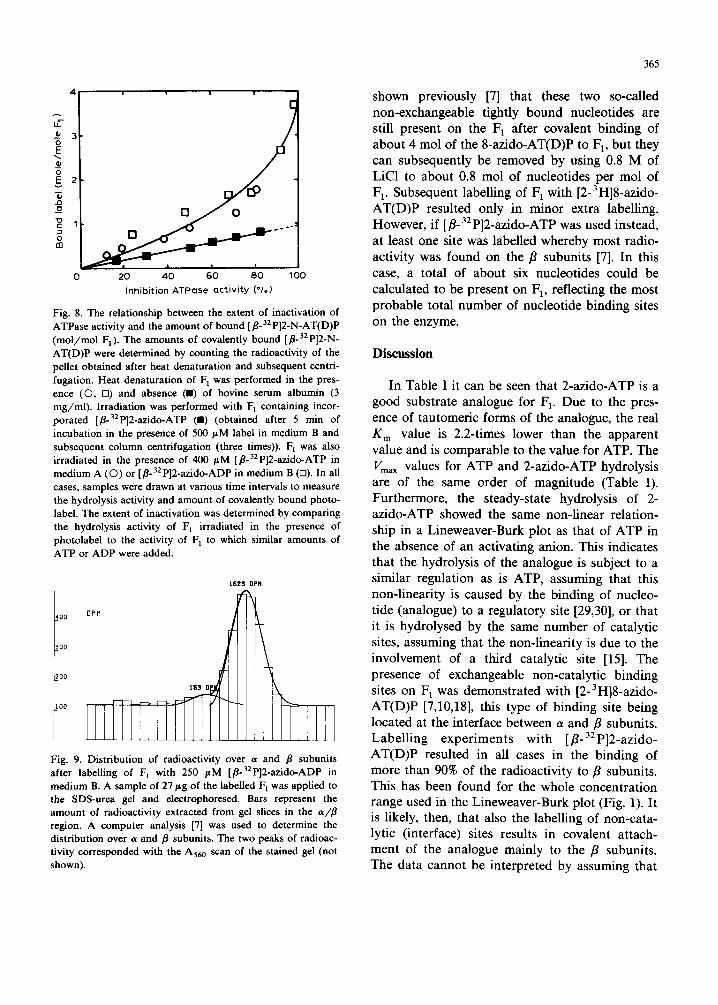
When [fl-32p]2-azido-AT(D)P is used in the incorporat ion experiments, the amount of cova- lently bound analogue needed for complete in- activation can be determined. After measuring the hydrolysis activity, the amount of covalently bound [•o 32 P]2-N-AT(D)P was measured by count ing the radioactivity of the pellet obtained after heat de- naturat ion and subsequent centrifugation (Fig. 8). A linear relationship was found between the ex- tent of inactivation and the bound label, extrapo- lating to 0.92 mol label per mol F] at full inactiva- tion of the enzyme. It can be concluded that the
o o
N
E
N
15
10
0.5
0
Incubation lime (minutesl
• '"';i'-"-- r"-.. t
'a ,
8 ~2b~
C
.g loo ~>
Q
50 ~
Fig. 7. Phosphate production due to the hydrolysis of 8-azido- ATP (©) or 2-azido-ATP (e) by F 1 dissolved in medium B (supplemented with HCO~-, (pH 8.0)). At the indicated times, samples were drawn to measure the amount of phosphate produced. At the same time samples were drawn, filtered three times on a Penefsky column (see legend to Fig. 4) and subse- quently tested for ATPase activity, the activity of the F 1 preincubated with 2-azido-ATP (I) or with 8-azido-ATP (n). Subsequently, these samples were irradiated for 10 and 25 min (see inserted scale) and tested for their ATPase activity. The small squares represent the relative inactivation of the irradia- ted samples; open symbols represent F 1 preincubated with 8-azido-ATP, filled-in symbols represent F 1 preincubated with 2-azido-ATP.
covalent a t tachment of only 1 mol [fl-32 P]2-N-ATP is required for the complete inactivation of the enzyme.
More than 90% of the radioactivity was found on the fl subunits as determined via urea-SDS polyacrylamide gel electrophoresis (Fig. 9). This latter result was also found when F 1 was irradiated in the presence of 400 # M [fl-32p]2-azido-AT(D)P, although, then, up to about 4 mol of label were bound (Fig. 8). Under the latter condit ions a non-l inear relationship was found between bind- ing and inhibition indicating that more than one catalytic site is involved, in contrast to the results obtained with [2-3 H]8-azido-ATP [7].
Binding of 2-azido-AT(D)P to non-exchangeable sites on
It has been reported previously [2,7,10,18] that 8-azido-AT(D)P can be used to label F 1 to a max imum of 4 mol label per mol F1, independent of the presence of the so-called non-exchangeable tightly bound nucleotides [11,12]. It has been
E 3
to
0
c9 O o
. I
20 40 GO 80 100
Inhibition ATPose a c t i v i t y ( * / . )
Fig. 8. The relationship between the extent of inactivation of ATPase activity and the amount of bound [fl.32 P]2-N-AT(D)P (mo l /mo l F1). The amounts of covalently bound [fl-32p]2-N- AT(D)P were determined by counting the radioactivity of the pellet obtained after heat denaturation and subsequent centri- fugation. Heat denaturat ion of F 1 was performed in the pres- ence (©, El) and absence ( I ) of bovine serum albumin (3 mg /ml ) . Irradiation was performed with F 1 containing incor- porated [fl-32p]2-azido-ATP (ll) (obtained after 5 min of incubation in the presence of 500 #M label in medium B and subsequent column centrifugation (three times)). F 1 was also irradiated in the presence of 400 /.¢M [fl-32p]2-azido-ATP in medium A (O) or [fl-32p]2-azido-ADP in medium B (El). In all cases, samples were drawn at various time intervals to measure the hydrolysis activity and amount of covalently bound photo- label. The extent of inactivation was determined by comparing the hydrolysis activity of F 1 irradiated in the presence of photolabel to the activity of F 1 to which similar amounts of ATP or ADP were added.
1623 0PM
_400 OF~M
300
I
O0
Fig. 9. Distribution of radioactivity over a and fl subunits after labelling of F 1 with 250 #M [fl-32p]2-azido-ADP in medium B. A sample of 27 Fg of the labelled F 1 was applied to the SDS-urea gel and electrophoresed. Bars represent the amount of radioactivity extracted from gel slices in the a/fl region. A computer analysis [7] was used to determine the distribution over a and fl subunits. The two peaks of radioac- tivity corresponded with the A56 o scan of the stained gel (not shown).
365
shown previously [7] that these two so-called non-exchangeable tightly bound nucleotides are still present on the F] after covalent binding of about 4 mol of the 8-azido-AT(D)P to F t, but they can subsequently be removed by using 0.8 M of LiC1 to about 0.8 mol of nucleotides per mol of F 1. Subsequent labelling of F 1 with [2-3H]8-azido - AT(D)P resulted only in minor extra labelling. However, if [fl-aEp]2-azido-ATP was used instead, at least one site was labelled whereby most radio- activity was found on the fl subunits [7]. In this case, a total of about six nucleotides could be calculated to be present on F1, reflecting the most probable total number of nucleotide binding sites on the enzyme.
Discuss ion
In Table I it can be seen that 2-azido-ATP is a good substrate analogue for F]. Due to the pres- ence of tautomeric forms of the analogue, the real K m value is 2.2-times lower than the apparent value and is comparable to the value for ATP. The Vma x values for ATP and 2-azido-ATP hydrolysis are of the same order of magnitude (Table I). Furthermore, the steady-state hydrolysis of 2- azido-ATP showed the same non-linear relation- ship in a Lineweaver-Burk plot as that of ATP in the absence of an activating anion. This indicates that the hydrolysis of the analogue is subject to a similar regulation as is ATP, assuming that this non-linearity is caused by the binding of nucleo- tide (analogue) to a regulatory site [29,30], or that it is hydrolysed by the same number of catalytic sites, assuming that the non-linearity is due to the involvement of a third catalytic site [15]. The presence of exchangeable non-catalytic binding sites on F] was demonstrated with [2-3H]8-azido - AT(D)P [7,10,18], this type of binding site being located at the interface between a and fl subunits. Labelling experiments with [fl-3:p]2-azido- AT(D)P resulted in all cases in the binding of more than 90% of the radioactivity to fl subunits. This has been found for the whole concentration range used in the Lineweaver-Burk plot (Fig. 1). It is likely, then, that also the labelling of non-cata- lytic (interface) sites results in covalent attach- ment of the analogue mainly to the fl subunits. The data cannot be interpreted by assuming that

366
the 2-azido analogue does not bind to the (regu- latory?) interface binding sites, as does the 8- azido-analogue. The difference between the two types of analogue in labelling the a and /3 sub- units must be attributed to the different location of the photoreactive azido group in the adenine ring.
In Fig. 2, it can be seen that the phosphate production by 2-azido-ATP follows a biphasic pattern. It is very likely that the slow (second) phase is due to the rate-limiting conversion of tetrazolic forms to the hydrolysable azido form. This notion is supported by the finding that when 2-azido-ATP was dissolved in methanol (in this medium 80% of the analogue is present in the azido form [25]), the initial amount of phosphate produced was indeed 80% of the total 2-azido-ATP added. The kinetic parameters for the hydrolysis of 8-azido-ATP (Table I) show that this analogue is a less easily hydrolysable substrate. A Line- weaver-Burk plot for the steady-state hydrolysis of 8-azido-ATP by isolated F~ has recently been re- ported by Sloothaak et al. [26]. The presence of the (non-photoreactive) tetrazolic isomers of 2- azido-AT(D)P was at least partly responsible for the less impressive inactivation of F1 upon il- lumination compared to 8-azido-ATP (Fig. 3). The results of the competition experiments between the 2-azido-analogues and AT(D)P are in agree- ment with those of competition experiments be- tween 8-azido-AT(D)P and AT(D)P [31,32].
Besides the demonstration of the very good kinetic parameters for the hydrolysis of 2-azido- ATP by F~, it is shown in this paper that the 2-azido analogues have the unique feature of bind- ing with high affinity to catalytic sites on the Fa (Fig. 4), as has also been demonstrated in CF 1 [35]. In Fig. 4, it can be seen that such tight binding to a catalytic site does not occur with 8-azido-ATP in the presence of EDTA. Possible tight binding to non-catalytic sites (in which case illumination would not result in inactivation) could be excluded on the basis of an experiment with [2-3H]8-azido-ATP in which no label was found on Fa after repeated Sephadex filtration and il- lumination. In the presence of Mg 2÷, small amounts of the 8-azido analogue were not re- moved by repeated Sephadex filtration (Fig. 7). The capacity of tight binding of the 2-azido ana-
logue has been described to be dependent on its intramolecular 'anti' configuration [19]. This also explains why 'syn' 8-azido-ATP is not tightly bound to F 1 under the conditions in which 2- azido-ATP is. As illumination of F 1 containing incorporated 2-azido-AT(D)P resulted in inactiva- tion, it can be concluded that a high-affinity state during the turnover exists on (one of the) catalytic sites. The experimental finding of a low Vma x for the hydrolysis of 8-azido-ATP can then be ex- plained by the fact that the 8-azido-ATP is hardly capable (due to its 'syn' configuration) of passing through this high-affinity state. This is supported by the finding that 8-azido-ATP is hardly able to bind tightly to the F 1.
Although inhibition of the F1-ATPase activity is observed upon illumination of F 1 with incorpo- rated 2-azido-ATP, it cannot be distinguished whether the analogue was only bound in its azido form or (also) in its tetrazolic forms and whether it was bound in such a way that the interconver- sion still continued. Fig. 2 suggests that it is only the azido form which is hydrolysed.
Especially at lower concentrations of 2-azido- ATP, the tight incorporation seems to be a Mg 2+- requiring process (Fig. 5). This is also illustrated by following the time course of the incorporation (Fig. 6). As both 2-azido-ATP and 2-azido-ADP could be tightly bound, it can be concluded that this tight binding does not require real turnover, i.e., dissociation of ADP due to the addition of ATP. In the presence of EDTA, competition for the Mg 2+ present between the MgZ+-chelating quality of EDTA and the Mg 2+ binding of the 2-azido-ATP could be responsible for the lowered incorporation. An integrated experiment, repre- sented in Fig. 7, shows both phosphate production due to the hydrolysis of 2-azido-ATP and 8-azido- ATP, and the tight incorporation of the analogues. The minor incorporation of the 8-azido-ATP pos- sibly reflects the lower rate of hydrolysis of this analogue compared to 2-azido-ATP.
When [fl-32p]2-azido-ATP was incorporated into a high-affinity site on the F1, 0.92 mol of label per mol of F1 were needed for complete inactivation (Fig. 8). It can be concluded that the covalent attachment of only 1 mol of 2-N-ATP per mol of F~ is required for the complete in- activation of the enzyme. The data of Melese and

367
Boyer [35] interpreted by the authors as suggestive for the retention of catalytic cooperativity by the two remaining subunits after one subunit has al- ready been labelled with 2-azido-ATP, can easily be explained by the assumption that under their conditions not one specific catalytic site was labelled but several, both catalytic and non-cata- lytic. Just one centrifugation through a Penefsky column removes only part of the loosely bound 2-azido-ATP. The high-affinity binding of the 2- azido analogues is very likely similar to the tight binding of ADP to catalytic sites, as proposed by Drobinskaya et al. [17].
As this high-affinity binding is to a catalytic site, the catalytic sites involved cooperate in an alternating site or sequential mechanism. These results are in agreement with results found with [2-3H]8-azido-ATP when this label was used at a low concentration (20/~M): also, in that case, only 1 mol of label per mol of t:1 was required for the complete inactivation of the enzyme, and the ra- dioactivity was found on the /3 subunits only [7,10,18]. At higher concentrations of [2-3H]8 - azido-ATP, an a//3 interface binding site was colabeUed, resulting in a typical 1 : 3 ratio for label bound to a//3 [7,10,18]. These results indicate that with [2-3H]8-azido-ATP a catalytic as well as a non-catalytic binding site can be labelled, located on the/3 subunit and the a//3 interface, respec- tively. This differentiation could not be made with [fl-32p]2-azido-AT(D)P, as the radioactivity was mainly (more than 90%) found on the/3 subunits, also when more than two sites were labelled.
At higher concentrations of [/3-32p]2-azido- AT(D)P, without the removal of medium and loosely bound nucleotides, 3-4 mol of label were bound. From this result, it can be concluded that 2-azido-ATP binds to both types of site. This will be investigated further by determining the amino acid residues labelled by the 2-azido analogue, as has been performed for 8-azido-ATP [34]. With this latter photolabel, indeed two different sites were demonstrated on the /3 subunits. The non- linearity between binding and inactivation can be explained by the assumption that more than one catalytic site was labelled, all causing the inhibi- tion of the enzyme. The shape of the curve corre- sponds to the theoretical curve for the case in which, among other (non-catalytic) sites, two cata-
lytic sites, both essential for hydrolysis, are being labelled. When using low concentrations of [a- 32p]2-azido-ADP (1-20 #M), Boulay et al. [36] found that 2 mol of bound label were needed for inactivation of F 1, the relationship between bind- ing and inhibition being linear. Since at least three sites bind 2-azido-ADP under the conditions used [36], it is likely that the linear extrapolation to 2 is not correct.
Besides the presence of four exchangeable bind- ing sites, F 1 contains at least two so-called non-ex- changeable tight nucleotide binding sites [2,3,8,9] which could be partially labelled with [fl-32p]2- azido-AT(D)P but not with the 8-azido analogues when the bound nucleotides were first removed by treating F 1 with LiC1 [7]. This difference can also be explained by the requirement for an 'anti ' configuration of the nucleotide, which is sup- ported by the relatively high substrate specificity of these sites [2]. Although it is theoretically ex- pected that the two so-called nonexchangeable nucleotides are located on a fl subunit and an a/fl interface, it is not possible to verify this assumption by measuring the distribution of [/3- 32p]2-azido-AT(D)P over a and fl subunits. De- termination of the labelled amino acid residues of the/3 subunit is required. The differentiation be- tween high-affinity binding to catalytic sites on the one hand and the so-called non-exchangeable tight nucleotide binding sites on the other is under current investigation in our laboratory, as both types of binding seem to require the intramolecu- lar 'anti ' configuration of the binding nucleotide.
Acknowledgements
We wish to thank M. Leijdekkers and C. van Santen for performing part of the experiments and Ms. G.E.E. van Noppen for her help in preparing the manuscript. This work was supported by the Netherlands Organization for the Advancement of Pure Research (Z.W.O.) under the auspices of the Netherlands Foundation for Chemical Research (S.O.N.).
References
1 Vignais, P.V. and Satre, M. (1984) Mol. Cell Biochem. 60, 33-70

368
2 Wagenvoord, R.J., Kemp, A. and Slater, E.C. (1980) Bio- chim. Biophys. Acta 593, 204-211
3 Cross, R.C. and Nalin, C.M. (1982) J. Biol. Chem. 257, 2874-2881
4 Weber, J., Li~cken, U. and Sch~ifer, G. (1985) Eur. J. Biochem. 148, 41-47
5 Cross, R.L., Grubmeyer, C. and Penefsky, H.S. (1982) J. Biol. Chem. 257, 12101-12105
60 'Nea l , C.C. and Boyer, P.D. (1984) J. Biol. Chem. 259, 5761-5767
7 Van Dongen, M.B.M. and Berden, J.A. (1986) Biochim. BIophys. Acta 850, BBA 42021
8 Harris, D.A., Rosing, J., Van de Stadt, R.J. and Slater, E.C. (1973) Biochim. Biophys. Acta 314, 149-153
9 Slater, E.C., Kemp, A., Van der Kraan, I., Muller, LL.M., Roveri, O.A., Verschoor, G.J., Wagenvoord, R.J. and Wielders, J.P.M. (1979) FEBS Lett. 103, 7-11
10 Van Dongen, M.B.M., Berden, J.A., Hartog, A.F. and Slater, E.C. (1985) 13th International Congress of Biochem- istry, Amsterdam, BBA Abstracts, p. 568, Elsevier Science Publishers, Amsterdam
11 Harris, D.A. and Baltscheffsky, M. (1979) Biochem. Bio- phys. Res. Commun. 86, 1248-1255
12 Rosing, J., Harris, D.A., Slater, E.C. and Kemp, A. (1975) J. Supramol. Struct. 3, 284-296
13 Grubmeyer, C., Cross, R.L. and Penefsky, H.S. (1982) J. Biol. Chem. 257, 12092-12100
14 Grubmeyer, C. and Penefsky, H.S. (1981) J. Biol. Chem. 256, 3728-3734
15 Gresser, M.J., Myers, J.A. and Boyer, P.D. (1982) J. Biol. Chem. 257, 12030-12038
16 Czarnecki, J.J., Abbott, M.S. and Selman, B.R. (1983) Eur. J. Biochem. 136, 19-24
17 Drobinskaya, I.Ye., Kozlov, I.A., Murataliev, M.B. and Vulfson, E.N. (1985) FEBS Lett. 182, 419-424
18 Berden, J.A., Van Dongen, M.B.M., Sloothaak, J.B. and Hartog, A.F. (1985) in Achievements and Perspectives of
Mitochondrial Research (Quagliariello, E., Slater, E.C., Palmieri, F., Saccone, C. and Kroon, A.M., eds.), Vol. I, pp. 257-266, Elsevier Science Publishers, Amsterdam
19 Czarnecki, J.J., Abbott, M.S. and Selman, B.R. (1982) Proc. Natl. Acad. Sci. USA 79, 7744-7748
20 Knowles, A.F. and Penefsky, H.S. (1972) J. Biol. Chem. 247, 6624-6630
21 Penefsky, H.S. (1977) J. Biol. Chem. 252, 2891-2899 22 Schaefer, H.J. and Thomas, H.J. (1958) J. Am. Chem. Soc.
80, 3738-3742 23 Sowa, T. and Ouchi, S. (1975) Bull. Chem. Soc. Jap. 48,
2084-2090 24 Hoard, D.E. and Ott, D.G. (1965) J. Am. Chem. Soc. 87,
1785-1788 25 Czarnecki, J.J. (1984) Biochim. Biophys. Acta 800, 41-51 26 Sloothaak, J.B., Berden, J.A., Herweijer, M.A. and Kemp,
A. (1985) Biochim. Biophys. Acta 809, 27-38 27 Fiske, C.H. and Subbarow, Y. (1925) J. Biol. Chem. 66,
375-400 28 Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall,
R.J. (1951) J. Biol. Chem. 193, 265-275 29 Recktenwald, D. and Hess, B. (1977) FEBS Lett. 76, 25-28 30 Stutterheim, E., Henneke, M.A.C. and Berden, J.A. (1980)
Biochim. Biophys. Acta 592, 415-430 31 Wagenvoord, R.J., Van der Kraan, I. and Kemp, A. (1977)
Biochim. Biophys. Acta 460, 17-24 32 Wagenvoord, R.J., Van der Kraan, I. and Kemp, A. (1979)
Biochim. Biophys. Acta 548, 85-95 33 Abbott, M.S., Czarnecki, J.J. and Selman, B.R. (1984) J,
Biol. Chem. 259, 12271-12278 34 Hollemans, M., Runswick, M.J., Fearnly, I.M. and Walker,
J.E. (1983) J. Biol. Chem. 258, 9307-9313 35 Melese, T. and Boyer, P.D. (1985) J. Biol. Chem. 260,
15398-15401 36 Boulay, F., Dalbon, P. and Vignais, P.V. (1985) Biochem.
24, 7372-7379





























