Embed Size (px)
Citation preview

Biochemische und funktionelle Charakterisierung
eines anaerob induzierten Ferredoxins aus
Chlamydomonas reinhardtii
Dissertation zur Erlangung des Grades eines
Doktors der Naturwissenschaften der Fakultät für Biologie und Biotechnologie
an der Internationalen Graduiertenschule Biowissenschaften
angefertigt im Lehrstuhl Biochemie der Pflanzen
Arbeitsgruppe Photobiotechnologie
vorgelegt von
Jessica Jacobs
aus
Wuppertal
Bochum
Juli 2009

Biochemical and functional characterisation of an
anaerobically induced ferredoxin from
Chlamydomonas reinhardtii
Dissertation to obtain the degree
Doctor Rerum Naturalium (Dr. rer. nat.) at the Faculty of Biology and Biotechnology
Ruhr-University Bochum
International Graduate School of Bioscience
Ruhr-University Bochum
Department of Plant Biochemistry
Work group Photobiotechnology
submitted by
Jessica Jacobs
from
Wuppertal, Germany
Bochum
Juli 2009

Teile dieser Arbeit wurden bereits veröffentlicht:
Jacobs J., Pudollek S., Hemschemeier A. und Happe T. (2008) A novel, anaerobically
induced ferredoxin in Chlamydomonas reinhardtii. FEBS Lett., 583, 2, 325-329.
Referent: Prof. Dr. Thomas Happe, Lehrstuhl für Biochemie der Pflanzen, AG
Photobiotechnologie
Korreferent: Prof. Dr. Elmar Weiler, Lehrstuhl für Pflanzenphysiolgie
Tag der Abgabe: 1. Juli 2009

Diese Arbeit wurde gefördert durch das Projekt „SOLAR-H“ der Europäischen Union und die
Deutsche Forschungsgemeinschaft im Sonderforschungsbereich 480.

Inhaltsverzeichnis
1 Einleitung .......................................................................................................................... 1
1.1 Photofermentation in Chlamydomonas reinhardtii........................................................ 1
1.2 Ferredoxine sind universelle Elektronenüberträger ....................................................... 5
1.3 Ziele dieser Arbeit .......................................................................................................... 8
2 Material und Methoden ................................................................................................. 10
2.1 Organismen und Anzucht............................................................................................. 10
2.1.1 Chlamydomonas reinhardtii..................................................................................... 10
2.1.2 Escherichia coli........................................................................................................ 11
2.2 Plasmide ....................................................................................................................... 11
2.3 Oligonukleotide............................................................................................................ 12
2.4 Chemikalien ................................................................................................................. 13
2.5 Enzyme......................................................................................................................... 13
2.6 Kits ............................................................................................................................... 14
2.7 Molekulargenetische Methoden ................................................................................... 14
2.7.1 Präparation von genomischer DNA aus C. reinhardtii............................................ 14
2.7.2 Transformation von C. reinhardtii........................................................................... 14
2.7.3 Präparation von RNA und mRNA aus C. reinhardtii.............................................. 14
2.7.4 Elektrophoretische Trennung von Gesamt-RNA und Northern Blotting................. 15
2.7.5 Gen-„Silencing“ in C. reinhardtii durch artifizielle microRNAs ........................... 15
2.7.6 Polymerase-Kettenreaktion (PCR) und Reverse Transkriptase PCR (RT-PCR) ..... 16
2.7.7 Klonierung................................................................................................................ 16
2.7.8 Heterologe Expression von Genen in E.coli mit dem Strep-Tag II System............. 18
2.8 Biochemische Methoden .............................................................................................. 18
2.8.1 Konzentrationsbestimmung von Proteinlösungen....................................................18
2.8.2 Polyacrylamidgelelektrophorese (PAGE) und Western Blotting............................. 18
2.8.3 Eisenbestimmung ..................................................................................................... 20
2.8.4 Strep-Tag II Affinitätsaufreinigung ......................................................................... 20
2.8.5 Aufreinigung von nativem Fdx5 .............................................................................. 20
2.8.6 Herstellung von polyklonalen Fdx5 Antikörpern..................................................... 21
2.8.7 Massenspektrometrische Identifizierung und N-terminale Sequenzierung.............. 21
2.8.8 Bestimmung des Redoxpotentials mittel cyclischer Voltammetrie (CV) ................ 21
2.8.9 Ferredoxin-abhängige Aktivitätstests....................................................................... 22

2.8.10 Isolierung von C. reinhardtii Chloroplasten und Mitochondrien............................. 23
2.9 Spektroskopische Methoden ........................................................................................ 23
2.9.1 Aufnahme von EPR (Electron Paramagnetic Resonance)- Signalen von Fdx5....... 23
2.9.2 Konfokale Laser Scanning Fluoreszenz Mikroskopie.............................................. 23
2.9.3 Eisenbestimmung mittels Röntgenfluoreszenzanalyse ............................................ 24
2.10 Physiologische Messungen .......................................................................................... 24
2.10.1 Chlorophyll- und Carotinoidbestimmung ................................................................ 24
2.10.2 In vitro- Test zur Messung der Hydrogenaseaktivität .............................................. 24
2.10.3 Nachweis von Fermentationsprodukten ................................................................... 25
3 Ergebnisse ....................................................................................................................... 26
3.1 Im Genom von C. reinhardtii kodieren sechs Gene für Ferredoxine........................... 26
3.2 Heterologe Expression von FDX5 aus C. reinhardtii in E.coli.................................... 29
3.3 Aufreinigung von heterolog synthetisiertem Fdx5 durch Strep-Tactin
Affinitätschromatographie............................................................................................ 30
3.4 Gegen heterolog produziertes Fdx5 gerichtete Antiköper zeigen die Akkumulation von
Fdx5 in C. reinhardtii auf Proteinebene....................................................................... 32
3.5 Partielle Aufreinigung von Fdx5 aus C. reinhardtii für die N-terminale Sequenzierung
...................................................................................................................................... 33
3.6 Biochemische Charakterisierung von Fdx5 ................................................................. 34
3.7 Zelluläre Lokalisation von Fdx5 .................................................................................. 37
3.8 Funktionelle Charakterisierung von Fdx5 durch Ferredoxin-abhängige Aktivitätstests..
...................................................................................................................................... 40
3.9 Postranskriptionelles Gen-„Silencing“ von FDX5 mittels artifizieller miRNAs ......... 43
3.9.1 Physiologische Analysen der amiRNA-Transformanden ........................................ 45
4 Diskussion ....................................................................................................................... 49
5 Zusammenfassung.......................................................................................................... 70
6 Summary ......................................................................................................................... 72
7 Literaturverzeichnis....................................................................................................... 73
8 Anhang ............................................................................................................................ 83
8.1 Auflistung der Ferredoxin-kodierenden Gene aus C. reinhardtii................................ 83
8.2 Auflistung der verwendeten Arbeitsgeräte................................................................... 83
9 Lebenslauf ....................................................................................................................... 85

Abkürzungsverzeichnis
Abkürzungsverzeichnis
Abb. Abbildung
Ack Acetat-Kinase
ad auf, bis (hinzufügen)
A. dest destilliertes Wasser
Adh Alkohol-Dehydrogenase
AdhE Acetaldehyd- und Alkohol-Dehydrogenase
Aox alternative Oxidase
Bp Basenpaare
c Konzentration
°C Grad Celsius
Chl Chlorophyll
cDNA komplementäre DNA
Cox Cytochrom-c- Oxidase
Cpx1 Coproporphyrinogen III Oxidase
Crd1 Isoform der oxidativen Cyclase (Mg-Protoporphyrin IX Monomethyl
Ester Cyclase)
Crd2 noch unbekanntes Genprodukt des Gens CRD2
Cyc6 Cytochrom c6
Da Dalton
DIG Digoxygenin
DNA Desoxyribonukleinsäure
E Extinktion
ε Extinktionskoeffizient
EDTA Ethylen-Diamin-Tetra-Acetat
EPR Electron Paramagnetic Resonance
Fd Ferredoxin
Fdx2, 3, 4, 5, 6 Ferredoxin 2, 3, 4, 5, 6 aus C. reinhardtii
Fnr Ferredoxin-NADP-Oxidoreduktase
Ftr Ferredoxin-Thioredoxin-Reduktase
GFP Grün-fluoreszierendes Protein

Abkürzungsverzeichnis
Gogat Glutamin-Oxoglutarat-Aminotransferase
h Stunde
H+ Proton
HydA1 [FeFe]-Hydrogenase 1 aus C. reinhardtii
JGI „Joint Genome Institute“
kDa Kilodalton
L10a ribosomales Protein L10a
mRNA messengerRNA
miRNA microRNA
NAD(P)H Nicotinamidadenindinukleotid (-phosphat)
Nar Nitrat-Reduktase
NCBI “National Center for Biotechnologial Information”
Nir Nitrit-Reduktase
OD optische Dichte (optical density)
p.A. per Analysis
PAGE Polyacrylamid-Gelelektrophorese
Pc Plastocyanin
PCR Polymerase-Ketten-Reaktion
Pdc Pyruvat-Decarboxylase
PEG Polyethylenglycol
PetF in der Photosynthese aktives Gen aus C. reinhardtii
Pfl Pyruvat Formiat-Lyase
PflA Pfl-Aktivase
Pfo Pyruvat-Ferredoxin-Oxidoreduktase
Pq Plastochinon
PS I, PS II Photosystem I und II
Ptox putative chloroplastidäre Plastochinon / Sauerstoff-Oxidoreduktase
RNA Ribonukleinsäure
RNAi RNA-Interferenz
rpm “rotations per minute” (Umdrehungen pro Minute)
RT Raumtemperatur (ca. 22°C)
RT-PCR reverse Transkriptase PCR
Rubisco Ribulose-1,5-bisphosphat Carboxylase/ Oxygenase
SAM S-adenosylmethionine

Abkürzungsverzeichnis
SDS Natrium-Dodecylsulfat
SIR Sulfit-Reduktase
Tab. Tabelle
TAP Tris-Acetat-Phosphate
TcdE 2-Ketobutyrat Formiat-Lyase aus E. coli
Tris Tris(hydroxymethyl)aminomethan
U Unit
UTR untranslatierte Region
Vol. Volumen
(v/v) volume per volume
(w/v) weight per volume
Verzeichnis der wichtigsten Gene Gen kodiertes Protein
BLE Phleomycin-Resistenz
CTH1 Isoform der oxidativen Cyclase (Mg-Protoporphyrin-IX-Monomethyl-
Ester Cyclase)
CPX1 Coproporphyrinogen III Oxidase
CRD1 Isoform der oxidativen Cyclase (Mg-Protoporphyrin-IX-Monomethyl-
Ester Cyclase)
CRD2 Genprodukt bislang unbekannt
CYC6 Cytochrom c6
FDX2, 3, 4, 5, 6 Ferredoxine 2, 3, 4, 5 und 6
HydA1 [FeFe]-Hydrogenase HydA1
PETF PetF
PFL1 Pyruvat-Formiat-Lyase
PFLA Pfl-Aktivase
RBCS kleine Unterheit der Rubisco (RbcS)

Einleitung
1
1 Einleitung
1.1 Photofermentation in Chlamydomonas reinhardtii
Die einzellige Grünalge C. reinhardtii ist ein photosynthetischer Flagellat, welcher der
Ordnung der Volvocales in der Klasse der Chlorophyceae angehört. Die Alge, die mitunter
auch als „photosynthetic yeast“ bezeichnet wurde, hat sich in vergangenen Jahrzehnten zu
einem bedeutenden Modellorganismus in der Zell- und Molekularbiologie entwickelt
(ROCHAIX, 1995). Die Anzucht von C. reinhardtii ist unproblematisch und kann unter sehr
definierten Bedingungen erfolgen, wobei die Grünalge sowohl photoautotroph,
photoheterotroph als auch heterotroph in Abwesenheit von Licht mit Acetat als
Kohlenstoffquelle wachsen kann. Der einfache Lebenszyklus und die leichte genetische
Manipulierbarkeit haben C. reinhardtii zu einem idealen Modellsystem für die Erforschung
der Photosynthese, der Chloroplasten- und der Flagellen-Biogenese (HARRIS, 2001; TAM und
LEFEVBRE, 1993; WEBBER et al., 1995) gemacht. Die in den letzten Jahren abgeschlossene
Sequenzierung aller drei Genome der einzelligen Grünalge hat die Chlamydomonas-
Forschung dabei einen weiteren großen Schritt vorangebracht (MICHAELIS et al., 1990; MAUL
et al., 2002; MERCHANT et al., 2007).
Während C. reinhardtii jahrzehntelang insbesondere als einzelliges Pflanzenmodell
untersucht wurde, haben Studien besonders aus den letzten Jahren gezeigt, dass die Alge auch
viele säugetier- (MERCHANT et al., 2007) und sogar bakterienartige Eigenschaften
(HEMSCHEMEIER und HAPPE, 2005) aufweist. So kann C. reinhardtii im Dunkeln unter
anaeroben Bedingungen innerhalb weniger Minuten einen komplexen
Fermentationsstoffwechsel ausbilden, der für Pflanzen untypisch ist. Dieser ist
gekennzeichnet durch die Produktion von Wasserstoff, Formiat, Ethanol, Acetat und CO2,
sowie Spuren von Glycerol und Lactat (GFELLER und GIBBS, 1981; HEMSCHEMEIER und
HAPPE, 2005; HEMSCHEMEIER et al., 2008 a).
Eine Besonderheit von C. reinhardtii ist, dass dieser Fermentationsstoffwechsel nicht nur im
Dunkeln, sondern auch unter Belichtung ausgebildet werden kann, wenn den Zellen das
Nährelement Schwefel entzogen wird und sie gleichzeitig luftdicht verschlossen sind (MELIS
et al. 2000). Da Schwefel essentieller Bestandteil von Proteinen (Cystein, Methionin), Lipiden
(Sulfoquinosovyl-Diglyceride) und Kofaktoren (Biotin, Thiaminpyrophosphat, [FeS]-Cluster)
ist, induziert der Mangel an Schwefel in der Alge daher eine Reihe von Reaktionen, die unter
anderem zu einer erhöhten Sulfataufnahme und -assimilation führen (LIEN und SCHREINER,
1975; YILDIZ et al., 1996; RAVINA et al., 1999; SATO et al., 2000). Der Grund für die

Einleitung
2
Anaerobisierung einer luftdicht verschlossenen Schwefelmangelkultur liegt jedoch in einer
drastischen Abnahme der photosynthetischen Aktivität der Zellen (WYKOFF et al., 1998), die
generell auf einen Verlust an PSII- Aktivität zurückzuführen ist, da die fortlaufende
Neusynthese des PSII-Kernproteins D1, das besonders bei hohen Lichtintensitäten oxidativen
Schäden und einem dadurch verstärkten Abbau (MELIS et al., 2000; OHAD et al., 1984)
unterliegt, nicht aufrechterhalten werden kann. Weitere Gründe sind die Akkumulation von
„Plastochinon-nicht-reduzierenden“ PSII-Zentren (WYKOFF et al., 1998; GUENTHER et al.,
1990) und das Auftreten von State I/ State II Transition (WYKOFF et al., 1998). Die bei
gleichbleibender Respirationsrate (MELIS et al., 2000) daraus resultierende Netto-
Sauerstoffaufnahme der Zellen führt somit trotz Belichtung zur Anaerobisierung einer
luftdicht verschlossenen Algenkultur und damit zur Induktion des Gärungsstoffwechsels.
Der Fermentationsmetabolismus von C. reinhardtii ist verwandt mit der gemischten
Säuregärung der Enterobakterien, wie z.B. von Escherichia coli. Die Produktion von Formiat,
Ethanol und Acetat im Verhältnis von 2:1:1, wie sie bei dunkel gärenden C. reinhardtii-
Kulturen zu beobachten ist (GFELLER und Gibbs, 1984), wird im Allgemeinen dem
Fermentationsenzym Pyruvat-Formiat-Lyase (Pfl) zugeschrieben, welches ein typisch
bakterielles Gärungsenzym ist (KREUZBERG, 1984). In Anwesenheit von Coenzym A
katalysiert es die thioklastische Spaltung von Pyruvat in Formiat und Acetyl-CoA.
Auch unter S-Mangelbedingungen konnte die Produktion von Formiat und Ethanol in C.
reinhardtii nachgewiesen werden (HEMSCHEMEIER und HAPPE, 2005). Real-Time PCR
Experimente haben gezeigt, dass die Transkription des PFL1-Gens unter S-
Mangelbedingungen induziert wird (HEMSCHEMEIER et al., 2008 a; JACOBS, Diplomarbeit).
Auch auf Proteinebene konnte eine aktive Pfl in C. reinhardtii nachgewiesen werden
(HEMSCHEMEIER et al., 2008 a; JACOBS, Diplomarbeit). Die Pfl1 gehört zur Klasse der
sogenannten Glycylradikal-Enzyme (KNAPPE et al., 1984; WAGNER et al., 1992) und wird
posttranslational durch die Pfl-Aktivase aktiviert, einem Radikal-S-Adenosylmethionin-
(SAM) Protein, dass für die Reaktion SAM und in E. coli, reduziertes Flavodoxin als
Elektronendonor benötigt (BLASCHKOWSKI et al., 1982). Bislang wurden im Genom von C.
reinhardtii keine Flavodoxine gefunden, so dass es sich bei dem benötigten Ein-
Elektronenüberträger auch um ein Ferredoxin handeln könnte. Auch die der Pfl-Reaktion
nachfolgenden typischen Enzyme, wie die Alkoholdehydrogenase konnten in C. reinhardtii
nachgewiesen werden, sowie mögliche Ethanol-produzierende Enzyme wie die Pyruvat-
Decarboxylase und die Pyruvat-Ferredoxin-Oxidoreduktase (ATTEIA et al., 2006;
HEMSCHEMEIER et al., 2008 a).

Einleitung
3
Abb. 1-1: Schematische Darstellung des Fermentationsmetabolismus in C. reinhardtii (nach
Hemschemeier et al., 2008 a). Während der Fermentation wird Glucose über die Glykolyse zu Pyruvat
abgebaut. Pyruvat kann anschließend über drei mögliche Wege umgesetzt werden. Die Pyruvat-Decarboxylase
decarboxyliert Pyruvat zu Acetaldehyd, welches durch die Alkohol-Dehydrogenase zu Ethanol reduziert wird.
Die Pyruvat Formiat-Lyase spaltet Pyruvat thioklastisch in Formiat und AcetylCoA. Das AcetylCoA wird
anschließend durch eine bifunktionale Acetaldehyd/Alkoholdehydrogenase zu Ethanol reduziert oder durch die
Phosphotransacetylase und die Acetat-Kinase zu Acetat umgesetzt. Durch die Pyruvat-Ferredoxin-
Oxidoreduktase kann Pyruvat zu AcetylCoA decarboxyliert werden. Ack = Acetat-Kinase, Adh = Alkohol-
Dehydrogenase, AdhE = Alkoholdehydrogenase, Pdc = Pyruvat-Decarboxyalse, Pfl = Pyruvat Formiat-Lyase,
PflA = Pfl-Aktivase, Pfo = Pyruvat-Ferredoxin-Oxidoreduktase, Pta = Phosphotransacetylase
Neben den typischen Fermentationsprodukten Formiat, Ethanol und Acetat produziert C.
reinhardtii unter anaeroben Bedingungen auch Wasserstoff. Dieser wird durch die monomere
[FeFe]-Hydrogenase HydA1 (HAPPE und NABER, 1993) gebildet, welche die Reduktion von
Protonen zu Wasserstoff katalysiert. Da die Wasserstoffbildung in C. reinhardtii jedoch vor
allem über die photosynthetische Elektronentransportkette mit Elektronen versorgt wird
(HEMSCHEMEIER et al., 2008 b, WINKLER, Dissertation, 2009), bilden die Zellen im Dunkeln
kaum Wasserstoff. Belichtete Schwefelmangelkulturen der Grünalge dagegen produzieren
signifikante Mengen des energiereichen Gases (MELIS et al., 2000), weshalb dieser spezielle
Wasserstoffmetabolismus auch intensiv von Biotechnologen untersucht wird (MELIS, 2007).
Ebenso wie die Pfl1, ist HydA1 ein sauerstoffsensitives Enzym, das durch die Reaktion mit
Sauerstoff irreversibel inaktiviert wird (HAPPE und NABER, 1993). Die Regulation der
HYDA1-Expression erfolgt nach bisherigen Studien auf transkriptioneller Ebene durch

Einleitung
4
Promotorelemente, die durch Anaerobiose reguliert werden (STIRNBERG und HAPPE, 2004)
und erfolgt parallel zur Transkription des PFL1-Gens. Der Elektronendonor von HydA1 ist
vermutlich das in der Photosynthese aktive [2Fe2S]- Ferredoxin PetF (HAPPE und NABER,
1993; WINKLER, Dissertation, 2009), das somit die Wasserstoffproduktion durch die
Hydrogenase an die photosynthetische Elektronentransportkette koppelt.
Abb. 1-2: Schematische Darstellung des Wasserstoffmetabolismus in C. reinhardtii. Elektronen für die
lichtabhängige Produktion von Wasserstoff in Schwefelmangelkulturen von C. reinhardtii stammen aus zwei
Quellen. Zum einen werden Elektronen, die aus der Restaktivität von PSII stammen, über die photosynthetische
Elektronentransportkette auf HydA1 übertragen. Zum anderen werden über die NAD(P)H-Plastochinon-
Oxidoreduktase Nda2 Elektronen, die durch den oxidativen Stärkeabbau freigesetzt werden, auf den
Plastochinonpool übertragen (JANS et al., 2008). Über eine noch nicht näher identifizierte chloroplastidäre
Plastochinon/Sauerstoff Oxidoreduktase können Elektronen aus dem Plastochinonpool wieder auf Sauerstoff
übertragen werden. Cyt b6f = Cytochrom-b6f-Komplex, Fd = Ferredoxin, Nda2 = NAD(P)H-Plastochinon-
Oxidoreduktase, Pc = Plastocyanin, Pq = Plastochinon, PS II, I = Photosystem II, I, Ptox = putative
chloroplastidäre Plastochinon/Sauerstoff Oxidoreduktase.
Unter Schwefelmangel werden die Elektronen, die auf die Hydrogenase übertragen werden,
von unterschiedlichen Quellen bereitgestellt. Es konnte gezeigt werden, dass die Elektronen
fast zu gleichen Teilen aus einer Restaktivität von PSII, sowie aus der nicht-photochemischen
Reduktion des Plastochinonpools (HEMSCHEMEIER et al., 2008 b) herrühren. Bei Letzterem
stammen die Elektronen aus dem oxidativen Abbau organischer Substrate, wie z.B. Stärke
und werden über eine NAD(P)H-Plastochinon-Oxidoreduktase auf den Plastochinonpool

Einleitung
5
übertragen. Dieser Prozess ist möglicherweise Teil der so genannten Chlororespiration
(FOUCHARD et al., 2005).
Die Bedeutung, die der Hydrogenase unter Schwefelmangelbedingungen zukommt, liegt
wahrscheinlich in ihrer Funktion als Elektronensenke (HAPPE et al., 2002). Schwefelmangel
bewirkt in den Algenzellen eine Drosselung der CO2-Fixierung, da das zentrale Protein des
Calvin-Zyklus, die Ribulose-1,5-bisphosphat-carboxylase/-oxygenase (Rubisco), abgebaut
wird (ZHANG et al., 2002). Durch die Einstellung des Calvin-Zyklus und weiterer
anabolischer Prozesse sinkt der Bedarf der Zelle an Reduktionsäquvivalenten und ATP, was
zu einem erhöhten Redoxpotential führt (ANTAL et al., 2003), das offenbar durch die
Hydrogenase abgeleitet wird.
Der außergewöhnlich komplexe, bakterienartige Photofermentationsmetabolismus von C.
reinhardtii, bietet folglich diverse metabolische und biosynthetische Stoffwechselprozesse in
denen Ferredoxine eine Funktion ausüben könnten, angefangen von typisch pflanzlichen bis
hin zu typisch bakteriellen Reaktionen
1.2 Ferredoxine sind universelle Elektronenüberträger
[FeS]-Proteine gehören neben Flavodoxinen, Cytochromen und Kupferproteinen zu den
wichtigsten elektronenübertragenden Proteinen in der Natur. Als prosthetische Gruppe
enthalten sie einen [FeS]-Cluster, in dem Eisen über schwefelhaltige Liganden gebunden
wird. Es wird angenommen, dass [FeS]-Cluster zu den ersten katalytischen Zentren gehörten,
die während der chemischen Evolution gebildet wurden (HUBER und WÄCHTERSHÄUSER,
1998). Unter anaeroben Bedingungen können sich die einfacheren [FeS]-Cluster in einer
Mischung aus Eisen, Sulfid und Thiolen, wie sie auch auf der primitiven Erde vorkamen,
selbst zusammenbauen (BERG und HOLM, 1982). Die Entstehung und die Entwicklung der
ersten [FeS]-Proteine fanden in anaeroben Bakterien statt (HALL et al., 1974). Davon
ausgehend entwickelte sich eine Vielzahl von Proteinen mit verschiedenen Kombinationen
von [FeS]-Clustern. Bis heute sind über 120 verschiedene Gruppen von Proteinen und
Enzymen bekannt, die [FeS]-Cluster besitzen (HALL et al., 1974; JOHNSON, 1994; BEINERT
2000). Die Funktionen, die [FeS]-Proteine ausüben, sind mannigfaltig und umfassen eine
Vielzahl von Stoffwechselwegen. Neben der Elektronenübertragung katalysieren sie unter
anderem Hydratationen / Dehydratationen, dienen als Sauerstoff- und Eisensensoren,
generieren und stabilisieren Radikale und spielen eine Rolle in regulatorischen Prozessen
(JOHNSON et al., 2005).

Einleitung
6
Unter den [FeS]-Proteinen bilden die Ferredoxine eine große, ubiquitär verbreitete Gruppe.
Bei ihnen handelt es sich um kleine, lösliche Proteine mit einer unter physiologischen
Bedingungen negativen Nettoladung, die in allen Organismengruppen gefunden werden
können. Sie sind an der Übertragung von Elektronen beteiligt, weisen ein niedriges
Redoxpotential auf und zeigen EPR-(Electron Paramagnetic Resonance) Signale, die typisch
für den jeweiligen Cluster-Typ sind (BRUSCHI und GUERLESQUIN, 1988). Der Name
Ferredoxin wurde erstmalig von MORTENSEN et al. (1962) für ein eisenhaltiges Protein aus
Clostridium pasteurianum benutzt, das als Elektronenüberträger in der Wasserstoffproduktion
des Bakteriums fungierte. Gleichzeitig wurde von TAGAWA und ARNON (1962) aus Spinacia
oleocera ein eisenhaltiges Protein isoliert, das die Reduktion von NADP+ zu NADPH durch
belichtete Chloroplasten katalysierte. Zudem konnten sie zeigen, dass das Ferredoxin aus C.
pasteurianum das Protein aus S. oleocera funktionell ersetzen konnte. Aufgrund der ähnlichen
Redoxpotentiale und ihrer funktionellen Austauschbarkeit wurde auch das pflanzliche Protein
als Ferredoxin bezeichnet. Auch andere Proteine mit diesen Eigenschaften wurden von
diesem Zeitpunkt in die Gruppe der Ferredoxine eingeordnet.
In Bakterien vorkommende Ferredoxine weisen unterschiedliche Typen von [FeS]-Clustern
auf. Neben den einfachen [2Fe2S]-Clustern können auch Ferredoxine mit [3Fe3S]- oder
[3Fe4S]-, sowie [4Fe4S]- oder auch 2[4Fe4S]- Cluster gefunden werden (BRUSCHI und
GUERLESQUIN, 1988). Sie nehmen an vielen elektronenübertragenden Prozessen teil, wie z.B.
dem Wasserstoff-, CO2- und Formiat-Metabolismus oder auch der phosphoroklastischen
Spaltung von Pyruvat (BRUSCHI und GUERLESQUIN, 1988). Eine Besonderheit stellen die
sogenannten Hochpotential-Eisenschwefel-Proteine (HiPIP) dar. Bei ihnen handelt es sich um
[4Fe4S]-Ferredoxine, die in photosynthetisch aktiven Bakterien vorkommen und
außergewöhnlich hohe Redoxpotentiale (+ 350 mV) aufweisen (HALL et al., 1974). Neben
den HiPIP tritt in einigen Bakterien eine weitere ungewöhnliche Klasse von Ferredoxinen auf,
die den Thioredoxinen ähnlich ist. Diese besitzen [2Fe2S]-Cluster und bilden Homodimere.
Die Funktion dieser Ferredoxine ist noch weitestgehend unbekannt. Es wird jedoch vermutet,
dass sie eine Rolle in der Stickstofffixierung spielen (MEYER, 2001).
Neben den oben aufgeführten Gruppen tritt in der Natur noch eine weitere Klasse von
Ferredoxinen auf, und zwar die überwiegend in Pflanzen, Algen und Cyanobakterien
vorkommenden [2Fe2S]-Ferredoxine. Diese Ferredoxine, die aus historischen Gründen auch
als Ferredoxine des „Pflanzen-Typs“ bezeichnet werden, fungieren auch hier als Ein-
Elektronen-Überträger in verschiedenen Reaktionen. Eine der bedeutensten und am besten
erforschten Reaktionen ist dabei die sogenannte Photoreduktion von

Einleitung
7
Nicotinamidadenindinukleotidphosphat (NADP). Ferredoxin wird durch Photosystem I (PSI)
reduziert und überträgt die Elektronen in einer durch Ferredoxin-NADP-Oxidoreduktase
(Fnr) katalysierten Reaktion auf NADP+ (ARNON, 1965).
In vielen Pflanzen und Algen können Ferredoxine in unterschiedlichen Isoformen gefunden
werden (BERTINI et al., 2002). So konnten beispielsweise in Zea mays vier unterschiedliche
Isoproteine nachgewiesen werden (HASE et al., 1991 a). Diese Isoformen können
gewebespezifisch gebildet werden und dabei funktionelle Unterschiede aufweisen (HASE et
al., 1991 b). Auch in nicht photosynthetisch aktiven Pflanzenzellen wurden bereits
Ferredoxin-Isoformen gefunden (SUZUKI et al., 1985; WADA et al., 1986). In den nicht
Photosynthese betreibenden Zellen von Wurzeln und Früchten werden Reduktionsäqvivalente
durch den oxidativen Pentosephosphatzyklus bereitgestellt und Elektronen über die Fnr auf
Ferredoxin übertragen, was genau dem umgekehrten Elektronenfluss in der Photosynthese
entspricht (FUKUYAMA , 2004). Das reduzierte Ferredoxin stellt wiederum Elektronen für
andere reduktive Prozesse wie die Nitrat- und Schwefelassimilation bereit (WADA et al.,
1986).
Aus der einzelligen Grünalge C. reinhardtii wurde bisher lediglich ein Ferredoxin isoliert und
charakterisiert. Es handelt sich hierbei um das für die NADP+-Photoreduktion von C.
reinhardtii verantwortliche Ferredoxin PetF. PetF ist ein typisches [2Fe2S]-Ferredoxin des
„Pflanzen-Typs“, das eine sehr negative Nettoladung und mehrere saure Reste an
konservierten Stellen der Aminosäuresequenz aufweist (ROGERS et al., 1992). Das native
Protein besteht aus 94 Aminosäuren und hat ein Molekulargewicht von 9908 Dalton
(SCHMITTER et al., 1988). Das Absorptionsspektrum des oxidierten Proteins weist die für
[2Fe2S]-Ferredoxine typischen Maxima bei 330, 420 und 465 nm auf (SCHMITTER et al.,
1988). Das PETF-Gen befindet sich im Kerngenom, das kodierte und im Cytoplasma
synthetisierte Protein wird aber mittels eines 32 Aminosäuren umfassenden Transitpeptids in
den Chloroplasten transportiert, wo es als Elektronenakzeptor für PSI dient (SCHMITTER et al.,
1988; FISCHER et al., 1999). Mittels „Cross-Linking“ wurde nachgewiesen, dass PetF auch
Substrat der Glutamat-Synthase und Nitrit-Reduktase ist (GARCIA-SÁNCHEZ et al., 2000).
Auch die Interaktion zwischen diesem Ferredoxin und Ferredoxin-Thioredoxin-Reduktase
wurde bereits bestätigt (JAQUOT et al., 1997). Daneben sind pflanzliche Ferredoxine auch bei
der Schwefel-Assimilation und der Aminosäure- und Fettsäuresynthese von Bedeutung
(KNAFF, 1996). Im Fall von C. reinhardtii kann PetF auch als Elektronenüberträger zwischen
PSI und der eingangs beschriebenen monomeren [FeFe]-Hydrogenase HydA1 fungieren, die

Einleitung
8
ein zentrales Element des komplexen anaeroben Stoffwechsels der Alge ist (WINKLER,
Dissertation, 2009).
Abb. 1-3: Typische Reaktionspartner von [2Fe2S]-Ferredoxinen des Pflanzen-Typs. In den Chloroplasten
wird Ferredoxin (Fd) von Photosystem I (PSI) reduziert und transferiert Elektronen auf diverse Redoxpartner,
wie die Acyl-Trägerprotein (Acp) -Desaturasen, die Ferredoxin-NADP+-Oxidoreduktase (Fnr ), die Ferredoxin-
Thioredoxin-Reduktase (Ftr ), die Glutamin-Oxoglutarat-Aminotransferase (Gogat), die Nitrit-Reduktase (Nir ),
die Sulfit-Reduktase (Sir) und im Fall von C. reinhardtii auf die Hydrogenase (HydA1) (KNAFF, 1996,
WINKLER, Dissertation, 2009). Cyt b6f = Cytochrom-b6f-Komplex, Fd = Ferredoxin, Pc = Plastocyanin, Pq =
Plastochinon, PS II = Photosystem II
Bei den Reaktionen, die PetF mit seinen Redoxpartnern eingeht, spielen elektrostatische
Wechselwirkungen eine große Rolle. PetF bildet mit seinen Reaktionspartnern spezifische
elektrostatische Komplexe aus, die durch eine Anzahl konservierter, carboxylierter Reste auf
Seite des Ferredoxins und von Arginin- oder Lysinresten auf Seite der Interaktionspartner
stabilisiert werden (GARCIA-SÁNCHEZ et al., 2000; JAQUOT et al., 1997; MAYORAL et al. 2005;
Knaff, 1996).
1.3 Ziele dieser Arbeit
Bei Untersuchungen der Expression verschiedener Gene des Fermentations- und
Wasserstoffmetabolismus von C. reinhardtii zeigte sich in Northern Blot und Real-Time PCR
Analysen, dass die Transkriptmenge des PETF-Gens unter Schwefelmangel abnimmt,
während bei FDX5, einem weiteren Ferredoxin-kodierenden Gen aus C. reinhardtii, eine
außergewöhnlich starke Akkumulation des Transkriptes beobachtet werden konnte (JACOBS,

Einleitung
9
2005, Diplomarbeit; HEMSCHEMEIER, 2002, Diplomarbeit). Die Akkumulation des FDX5-
Transkriptes und die gleichzeitige Abnahme des PETF-Transkriptes deuten daraufhin, dass
der Ferredoxingehalt in C. reinhardtii unter photofermentativen Bedingungen moduliert und
eventuell die Synthese spezialisierter Ferredoxine induziert wird, die im veränderten
Stoffwechsel der Zellen aktiv sind. Ziel dieser Arbeit ist eine umfassende biochemische und
funktionelle Charakterisierung des FDX5-Genproduktes, bei dem es sich um ein potentielles
Ferredoxin handelt. Hierfür soll ein heterologes Expressionsystem in E. coli und ein
effektives Protokoll für die Aufreinigung des rekombinanten Proteins etabliert werden. Das
rekombinante Protein dient als Ausgangspunkt für die biochemische Charakterisierung und
die Herstellung polyklonaler Antikörper. Mittels Lokalisationsstudien und Interaktionstests
sollen Erkenntnisse über die mögliche Funktion von Fdx5 in der Photofermentation von C.
reinhardtii gewonnen werden.

Material und Methoden
10
2 Material und Methoden
2.1 Organismen und Anzucht
2.1.1 Chlamydomonas reinhardtii
Folgende C. reinhardtii Stämme wurden im Rahmen dieser Arbeit verwendet:
Stamm Genotyp Referenz
C. reinhardtii CC-125 mt+ 137c HARRIS, 1989
C. reinhardtii SAG 83.81 cw15 mt+ DAVIES und PLASKITT, 1971
C. reinhardtii CW 388 Arg- cw15 arg7 nitI mt+ PURTON und ROCHAIX, 1994
SAG: „Sammlung von Algenkulturen der Universität Göttingen“, Göttingen, Deutschland; CC: „Culture
Collection at Duke University“, Durham, NC, USA.
2.1.1.1 Kultivierung von C. reinhardtii
Die Anzucht von C. reinhardtii erfolgte photoheterotroph in 200 mL Tris-Acetat-Phosphat
(TAP)- Medium (HARRIS, 1989) in Erlenmeyerkolben unter konstanter Belichtung mit 100
µmol Photonen x m-2 x s-1 und konstantem Schütteln bei 20°C.
2.1.1.2 Schwefelmangelinduktion
Für die Schwefelmangelinduktion wurden die Zellen durch Zentrifugation (2,5 min, 3000
rpm) in der exponentiellen Wachstumsphase geerntet und in TAP-S Medium (TAP Medium,
in dem alle sulfathaltigen Salze durch die entsprechenden Chloride ersetzt wurden)
gewaschen und resuspendiert. Anschließend wurden die Zellen in Vierkantflaschen gefüllt,
mit luftdichten Subastopfen („red rubber Suba seals 37“, Sigma-Aldrich, St. Louis, Mo, USA)
verschlossen und unter kontinuierlicher, einseitiger Belichtung mit 100 µmol Photonen x m-2
x s-1 inkubiert.
2.1.1.3 Kupfermangelinduktion
Die Herstellung von TAP-Cu, sowie die Anzucht unter Cu-freien Bedingungen wurden wie
von QUINN und MERCHANT (1998) bechrieben, durchgeführt.
2.1.1.4 Anaerobe Induktion
Für die anaerobe Induktion wurden Zellen durch Zentrifugation (2,5 min, 3000 rpm) in der
exponentiellen Wachstumsphase geerntet und die Zelldichte auf einen Wert von 100 µg/ml

Material und Methoden
11
Chlorophyll in einem Volumen von 10 ml eingestellt. Anschließend erfolgte eine Begasung
der Kulturen mit Argon.
2.1.2 Escherichia coli
Für molekularbiologische sowie biochemische Techniken wurden folgende E. coli Stämme
verwendet:
Stamm Genotyp Referenz/Firma
E.coli DH5α MCR F’end A1 hsd 17 (rk-mk+) sup E44 thi-1 rec A1� ∆
(laclzyA-argF) u 169 deo R
RALEIGH et al., 1989;
WOODCOCK et al., 1989 /
(Novagen/ Merck,
Darmstadt, Deutschland)
E.coli BL21(DE3)pLysS F-, ompT, hsdSB (rB-, mB-), dcm, gal, λ (DE3),
pLysS, Cmr
STUDIER und MOFFAT, 1986
(Novagen/ Merck,
Darmstadt, Deutschland)
2.1.2.1 Kultivierung von E. coli
Gewöhnlich wurden E. coli Zellen in LB-Medium (25 g x l-1 Luria Broth Base, Invitrogen
Gibco, Carlsbad, CA, USA) oder auf LB-Platten (30 g x l-1 Luria Agar, Invitrogen Gibco) bei
37 °C über Nacht angezogen. Dafür wurden alle Medien bei 120°C für 20 min. autoklaviert.
Hitzelabile Komponenten wurden sterilfiltriert und dem Medium nach dem Autoklavieren
hinzugefügt. Zur Selektion plasmidtragender E. coli Zellen wurden Antibiotika in folgenden
Konzentrationen hinzugefügt: Ampicillin: 100 µg x ml-1; Chloramphenicol: 20 µg x ml-1;
Kanamycin: 10 µg/ml.
2.2 Plasmide
Folgende Plasmide wurden während dieser Arbeit verwendet oder hergestellt:
Plasmid Charakteristika Referenz/Firma
pGEM TEasy linearer Klonierungsvektor mit 3’- Thyminüberhängen, welche die
Klonierung von PCR-Fragmenten der Taq-Polymerase erleichtern;
ermöglicht Blau-Weiß-Selektion; ApR 3,02 kb
Promega
(Mannheim,
Deutschland)
pASK-IBA7 Expressionsvektor, der das Anhängen eines N-terminalen Strep-Tag
II an ein rekombinantes Protein ermöglicht; Regulation über
Anhydrotetracyclin (Vektor enthält tet-Promotor und Operator
sowie das Gen für den tet-Repressor); ApR; 3,25 kb
IBA
(Göttingen,
Deutschland)
pJJ6 151 bp Fragment von RPL10A in pGEM (Sondenherstellung) JACOBS,
Diplomarbeit, 2005
Material und Methoden
12
pJJ7 260 bp Fragment von HYDA1 in pGEM (Sondenherstellung) JACOBS,
Diplomarbeit, 2005
pJJ10 271 bp Fragment von FDX5 in pGEM (Sondenherstellung) JACOBS,
Diplomarbeit, 2005
pJJ20 orf des PETF Gens, exklusive der Transitsequenz, in pASK IBA7 diese Arbeit
pJJ23 orf des FDX5 Gens, exklusive der Transitsequenz, in pASK IBA7 diese Arbeit
pMF59 Konstrukt zur Expression eines Ble/ GFP-Fusionsproteins mit zwei
rbcS2-Intron-Elementen vor bzw. innerhalb der BLE Sequenz,
nachgeschalteter cgfp-Sequenz, sowie 5’- und 3’-UTR des rbcS2-
Gens von C. reinhardtii. Die Expression der Gensequenzen
unterliegt der Kontrolle des artifiziellen HSP70A/ RBCS2-
Tandempromotors. ApR; 5,07 kb
FUHRMANN et al.,
1999
pJJ24 Konstrukt zur Expression eines Fdx5-Transitpeptid/ GFP-
Fusionsproteins. Ausgangsplasmid für dieses Konstrukt war
pMF59.
diese Arbeit
pChlamiRNA2 Konstrukt zur Expression einer artifiziellen miRNA. Der
Transkriptionsterminator beinhaltet die 3’-UTR des RPL12 Gens.
Die Expression der amiRNA-Sequenz unterliegt der Kontrolle des
artifiziellen HSP70A/ RBCS2-Tandempromotors. Der Vektor trägt
die Sequenz des ARG7-Gens. ApR; 12,65 kb
MOLNÁR et al., 2009
pJJ27 Fdx5 amiRNA Variante 1 in pChlamiRNA2 miRNA diese Arbeit
pJJ28 Fdx5 amiRNA Variante 2 in pChlamiRNA2 miRNA diese Arbeit
2.3 Oligonukleotide
Folgende Oligonukleotide wurden in dieser Arbeit abgeleitet und verwendet:
Primer Sequenz Verwendung
Ferredoxin_5_157 CCGCTGACACCTACATCCTG
Ferredoxin_3_504 AACCGCTCATAGTGCTAGGC
RT-PCR
Fdx2_5_29 GCCACGCTGTAGCTCATTCT
Fdx2_3_344 AGCATGTTCTGGTCGGACTG
RT-PCR
Fdx3_5_55 CCGGACAACCAATACATCCT
Fdx3_3_215 TCCTCCTCGTCTAGCGTGAA
RT-PCR
Fdx4_5_19 GAAGGCAAGCAGGTGGATCT
Fdx4_3_296 CTGGTCATCAGCTGCATGTC
RT-PCR
Fdx5_5_777 CGGCTTGAAGAGTCAAGAGT
Fdx5_3_1047 CCACACCAGAGCCATACATA
RT-PCR
Fdx6_5_494 CGACCTGTGGATGGAAGATG
Fdx6_3_658 ATCTCCTGGTTGCCGTAGTG
RT-PCR
Fdx5_5_Strep ATGGTAGGTCTCAGCGCTTTCAGGTGACGCTGCGCATGC Klonierung in

Material und Methoden
13
Fdx5_3_Strep ATGGTAGGTCTCATATCACTGGTGCTTGCCGTACTCGCA pASK-IBA7
PetF_5_Strep ATGGTAGGTCTCAGCGCTACAAGGTCACCCTGAAGACCC
PetF_3_Strep ATGGTAGGTCTCATATCAGTACAGGGCCTCCTCCTGGTG
Klonierung in
pASK-IBA7
Fdx5_5_pMF CGACCCATTTGCAGGCTGGCACAGCAGTTTCAGGCATTG
Fdx5_3_pMF GTCGGGCCGGCCCTGGTGCTTGCCGTACTCGC
Klonierung in
pMF59
amiFor_Fdx5_1 CTAGTGTGGCGACTCTTCCGGGAGAATCTCGCTGATCGGC
ACCATGGGGGTGGTGGTGATCAGCGCTATTCTTTCGGAAG
AGTCGCCACG
amiRev_Fdx5_1 CTAGCGTGGCGACTCTTCCGAAAGAATAGCGCTGATCAC
CACCACCCCCATGGTGCCGATCAGCGAGATTCTCCCGGA
AGAGTCGCCACA
Klonierung in
pChlamiRNA2
amiFor_Fdx5_2 CTAGTAGACGGGTATGTATGGGTCTATCTCGCTGATCGGC
ACCATGGGGGTGGTGGTGATCAGCGCTATAGAGCCATAC
ATACCCGTCTG
amiRev_Fdx5_2 CTAGCAGACGGGTATGTATGGCTCTATAGCGCTGATCACC
ACCACCCCCATGGTGCCGATCAGCGAGATAGACCCATAC
ATACCCGTCTA
Klonierung in
pChlamiRNA2
2.4 Chemikalien
Die eingesetzten Chemikalien wurden in größtmöglicher Reinheit (p.A. Qualität) von den
Firmen Acros, LKB-Pharmacia, Merck, Roche, Roth, Serva und Sigma bezogen.
2.5 Enzyme
Es wurden folgende Enzyme verwendet, die nicht Bestandteil eines Kit-Systems waren:
Enzym Hersteller
Cytochrom c (Pferdeherz) Sigma, St. Louis, Mo, USA
Ferredoxin-NADP+-Oxidoreduktase (S. oleocera) Sigma, St. Louis, Mo, USA
Hydrogenase HydA1 (C. reinhardtii) S. Stripp (Ruhr-Universität Bochum, Lehrstuhl
Biochemie der Pflanzen, AG
Photobiotechnologie)
Pfu Ultra Hotstart Polymerase Stratagene, La Jolla, CA, USA
Photosystem I (C. reinhardtii) M. Hippler (Westfälische Wilhelms-Universität
Münster, Institut für Biochemie und
Biotechnologie der Pflanzen, Münster,
Deutschland)
Plastocyanin (C. reinhardtii) M. Hippler
Restriktionsendonukleasen Fermentas, Burlington, Canada
Superscript II Reverse Transkriptase Invitrogen Gibco, Carlsbad, CA, USA

Material und Methoden
14
Taq-DNA-Polymerase Fermentas, Burlington, Canada
T4-DNA Ligase Fermentas, Burlington, Canada
2.6 Kits
Folgende Kits wurden verwendet:
Bezeichnung Hersteller
DIG RNA Labeling Kit (Sp6/T7) Roche Applied Science,
Mannheim, Deutschland
Gene JetTM Plasmid MiniPrepKit Fermentas, Burlington, Canada
GFXTM
PCR DNA and Gel Band Purification Kit GE Healthcare, Chalfont, UK
Mini EluteTM
Reaction Cleanup Kit Macherey-Nagel, Düren, Deutschland
Nucleo Trap mRNA-Isolierungskit Macherey-Nagel, Düren, Deutschland
OneStep RT-PCR Kit Qiagen, Hilden, Deutschland
pGEM-TEasy Vektor System Promega, Promega, Madison, WI, USA),
Testsystem zum Nachweis von Formiat und Ethanol R-Biopharm AG, Darmstadt, Deutschland
2.7 Molekulargenetische Methoden
2.7.1 Präparation von genomischer DNA aus C. reinhardtii
Die Isolierung von genomischer DNA aus C. reinhardtti erfolgte nach NEWMAN et al. (1990).
2.7.2 Transformation von C. reinhardtii
Die Transformation von zellwandlosen C. reinhardtii Stämmen erfolgte nach der „Glasperlen-
Methode“ (KINDLE, 1990)
2.7.3 Präparation von RNA und mRNA aus C. reinhardtii
Die Präparation von Gesamt-RNA aus C. reinhardtii erfolgte nach Johanningmeierund
Howell, 1984. Die RNA-Konzentration wurde photometrisch bei einer Wellenlänge von 260
nm gegen Wasser als Referenz bestimmt. Eine Absorption von 1,0 entspricht einer
Konzentration von 40 µg RNA pro ml. Zur Bestimmung des Reinheitsgrades der RNA wurde
der Quotient der Absorption bei 260 und 280 nm ermittelt.
Die Isolierung der mRNA erfolgte mit Hilfe des Oligotex mRNA Mini Kits von Macherey-
Nagel nach Angaben des Herstellers. Vor der mRNA-Isolierung wurde die einzusetzende
RNA-Menge bestimmt. Die Lagerung der mRNA erfolgte bei -80°C.

Material und Methoden
15
2.7.4 Elektrophoretische Trennung von Gesamt-RNA und Northern Blotting
Die Auftrennung von Gesamt-RNA in 1,5 % Formaldehyd-MOPS Agarose-Gelen und
Northern Blotting erfolgte nach bereits beschriebenen Standardmethoden unter Nutzung von
Digoxigenin-markierten RNA-Sonden (Sambrook et al., 1989).
2.7.5 Gen-„Silencing“ in C. reinhardtii durch artifizielle microRNAs
Gemäß dem von MOLNÁR et al. (2009) entwickelten Protokoll wurden basierend auf der
Zielsequenz des FDX5-Gens mittels der Web MicroRNA Designer Plattform (WMD2,
http://wmd2.weigelworld.org/cgi-bin/mirnatools.pl) geeignete miRNA Kandidaten gesucht.
Um die Wahrscheinlichkeit des Gen-„Silencing“ zu erhöhen, wurden zwei miRNAs
ausgewählt, die unterschiedliche Bereiche des Gens zum Ziel haben. miRNA Variante 1
(TTCTTTCGGAAGAGTCGCCAC) ist komplementär zu einer in der 5’-UTR des FDX5-
Gens gelegenen Sequenz, während die Zielsequenz von amiRNA Variante 2
(TAGAGCCATACATACCCGTCT) in der 3’-UTR liegt. Ausgehend von diesen miRNA-
Sequenzen wurden mit dem WMD2 „Oligo“-Programm 90 nt umfassende Oligonukleotide
entworfen, die nach Annealing zu doppelsträngigen DNA-Molekülen bereits einen SpeI
kompatiblen Überhang aufwiesen.
CTAGTGTGGCGACTCTTCCGGGAGAA TCTCGCTGATCGGCACCA
TGGGGGTGGTGGTGATCAGCGCTATTCTTTCGGAAGAGTCGCCA
CG
miRNA Variante 1
(amiFor_Fdx5_1,
amiRev_Fdx5_1)
CTAGCGTGGCGACTCTTCCGAAAGAA TAGCGCTGATCACCACCA
CCCCCATGGTGCCGATCAGCGAGATTCTCCCGGAAGAGTCGCCA
CA
CTAGTAGACGGGTATGTATGGGTCTA TCTCGCTGATCGGCACCA
TGGGGGTGGTGGTGATCAGCGCTATAGAGCCATACATACCCGTC
TG
miRNA Variante 2
(amiFor_Fdx5_2,
amiRev_Fdx5_2)
CTAGCAGACGGGTATGTATGGCTCTA TAGCGCTGATCACCACCA
CCCCCATGGTGCCGATCAGCGAGATAGACCCATACATACCCGTC
TA
Für das Annealing wurden die betreffenden Oligonukleotide in einer Konzentration von 100
µM für fünf min in Annealingpuffer (20 mM Tris ph 8, 2 mM EDTA, 100 mM NaCl) gekocht
und langsam über Nacht (ü.N.) abgekühlt. Um die hohen Salzkonzentrationen zu entfernen,
wurde die doppelsträngige DNA mit dem „GFX™ PCR DNA and Gel Band Purification Kit“

Material und Methoden
16
(GE Healthcare) aufgereinigt. Die Klonierung in das Expressionsplasmid pChlamiRNA2
(MOLNÁR et al., 2009) erfolgte über die Restriktionsschnittstelle SpeI. Die SpeI-
Restriktionsschnittstelle in pChlamiRNA2 werden flankiert von den modifizierten Armen des
miRNA-Vorläufermoleküls cre-MIR1157, die zusammen mit dem eingefügten
doppelsträngigen Oligonukleotid den artifiziellen miRNA-Vorläufer bilden, der zur miRNA
prozessiert wird. Im Anschluss an die Klonierung erfolgte die Transformation von C.
reinhardtii CW 388 Arg- mittels „Glasperlen“-Methode.
2.7.6 Polymerase-Kettenreaktion (PCR) und Reverse Transkriptase PCR (RT-PCR)
Für Kontrollexperimente wurde die PCR mit Taq DNA Polymerase durchgeführt. Für die
sequenzexakte Amplifizierung von DNA-Fragmenten wurde Pfu Ultra Hotstart Polymerase
verwendet. Die Primer wurden von Sigma-Genosys Ltd (Sigma-Aldrich) bezogen und in einer
Konzentration von 0,4 µM eingesetzt.
„One Step“ RT-PCRs wurden mit dem „OneStep RT-PCR- Kit“ von Qiagen nach
Herstellerangaben durchgeführt. Für die „Two-Step“ RT-PCR wurde die Superscript II
Reverse Transkriptase von Invitrogen verwendet. Das Protokoll für die reverse Transkription
orientierte sich an den Herstellerangaben. Im Anschluss an die cDNA-Synthese wurden dem
Ansatz NaOH und EDTA zugegeben und 15 min bei 65 °C inkubiert, um noch vorliegende
RNA zu hydrolysieren. Die Aufreinigung der cDNA erfolgte mit dem „Mini Elute Reaction
Cleanup Kit“ von Quiagen nach Herstellerangaben. Eluiert wurde in 10 µl Wasser. Die
aufgereinigte cDNA wurde als Matrize für eine PCR mit einer ausgewählten
Primerkombination eingesetzt.
2.7.7 Klonierung
2.7.7.1 Transformation von E .coli
Kompetente E. coli Dh5α- bzw BL21-Zellen wurden nach SAMBROOK et al. (1989) unter
Aufnahme in 200µl eines TB-Puffers (250 mM KCl, 10 mM PIPES, pH 6,7) mit 15 mM
CaCl2 hergestellt und bei -80°C gelagert. Die Transformation erfolgte nach der CaCl2-
Standardmethode (COHEN et al., 1972), modifiziert nach SAMBROOK (1989).
2.7.7.2 Präparation von Plasmid-DNA aus E. coli
Die Gewinnung von Plasmid-DNA aus E. coli erfolgte mittels des „GFX™ Micro Plasmid
Prep Kit“ von Amersham nach den Herstellerangaben.

Material und Methoden
17
2.7.7.3 Elektrophoretische Trennung von DNA-Fragmenten
Die Auftrennung von DNA-Fragmenten erfolgte durch Flachbett-Elektrophorese in 1 % TAE
(Tris-Acetat-EDTA)-Agarose-Gelen. Die Analyse und Photographie erfolgte auf einem UV-
Lichttransilluminator.
2.7.7.4 Bestimmung der DNA-Konzentration
Die DNA-Konzentration wurde photometrisch bei einer Wellenlänge von 260 nm gegen
Wasser als Referenz bestimmt. Eine Absorption von 1,0 entspricht einer Konzentration von
50 µg DNA pro ml. Als Maß für die Reinheit der der Nukleinsäure wurde der Quotient OD 260
/OD 280 ermittelt, der bei einer reinen Lösung zwischen 1,8 und 2,0 liegen sollte.
2.7.7.5 Ligation von DNA-Fragmenten
Für die Ligation von DNA-Fragmenten mit Vektoren wurden das DNA-Fragment und das
Plasmid im Verhältnis von 3:1 in einem Reaktionsansatz mit T4-Ligase und dem
entsprechenden Puffer zusammengegeben. Der Ligationsansatz wurde ü.N. bei 4°C inkubiert.
PCR-Produkte wurden, abhängig von der Sauberkeit der PCR-Probe, direkt kloniert oder
vorher mit dem „GFXTM PCR DNA and Gel Band Purification Kit“ von GE Healthcare (
Chalfont, UK) aufgereinigt.
2.7.7.6 Restriktion von Plasmid-DNA
Die Restriktion von DNA mit Restriktionsendonukleasen erfolgte in dem jeweiligen vom
Hersteller empfohlenen Puffersystem. Die Restriktionstemperatur und -dauer sowie die
Inaktivierung des Enzyms richtete sich je nach verwendeter Endonuklease nach den
Herstellerangaben. Plasmid-DNA wurde mit 1 bis 2 U Enzym pro µg DNA verdaut.
2.7.7.7 Isolierung von DNA-Fragmenten aus Agarosegelen
DNA-Fragmente wurden mittels des „GFXTM PCR DNA and Gel Band Purification Kit“ von
GE Healthcare ( Chalfont, UK) isoliert. Die gewünschte Bande wurde bei schwacher UV-
Bestrahlung mit dem Skalpell aus dem Agarosegel herausgeschnitten. Das Gelstück wurde in
ein Eppendorf-Gefäß überführt und in 300 µl Capture-Puffer bei 60°C gelöst. Die weiteren
Schritte der Isolierung erfolgten nach den Herstellerangaben.
2.7.7.8 Sequenzierung
Die DNA-Sequenzanalyse erfolgte im Rahmen einer Sequenzierservice-Einrichtung des
Lehrstuhls für Biochemie I an der Ruhr-Universität Bochum nach dem
Didesoxyribonukleotid-Kettenabbruch-Prinzip (SANGER et al., 1977).

Material und Methoden
18
2.7.8 Heterologe Expression von Genen in E.coli mit dem Strep-Tag II System
Für die heterologe Expression von Genen aus C. reinhardtii in E. coli wurde der Vektor
pASK-IBA7 des Strep-Tag Systems der Firma IBA (Göttingen, Deutschland) eingesetzt, bei
dem der Strep-Tag II N-terminal an das Zielprotein angefügt wird. Das Prinzip des Strep-Tag
Systems beruht auf der Bindung von Biotin an Streptavidin. Der Strep-Tag II besteht aus acht
Aminosäuren (Trp-Ser-His-Pro-Gln-Phe-Glu-Lys) und bindet an die Biotin-Bindedomäne des
Streptavidins. Damit wird eine affinitätschromatographische Aufreinigung des Strep-Tag II
Fusionsproteins über eine Strep-Tactin Säule, einer molekularbiologisch erzeugten Form des
Streptavidin, ermöglicht. Die Expression eines Genes in Vektor pASK-IBA7 unterliegt der
Kontrolle des TET-Promotors und ist durch die Zugabe von Anhydrotetracyclin (AHT)
induzierbar. Der TET-Repressor ist auf dem pASK-IBA Vektor kodiert und wird konstitutiv
exprimiert. Die Expression des klonierten Gens wird bis zur Induktion durch AHT strikt
unterdrückt. Im Gegensatz zum LAC-Promotor ist die Kontrolle durch den TET-Promotor
streng kontrolliert, so dass vor der Induktion keine Grundexpression des rekombinanten
Proteins zu beobachten ist (SKERRA, 1994)
Für die heterologe Expression wurde E. coli Stamm BL21 DE3 pLys mit dem entsprechenden
Expressionsplasmid transformiert. Die Protein-Produktion erfolgte in 3 l Vogel-Bonner
Medium (VOGEL und BONNER, 1955) Die Zellkultur wurde unter Schütteln bei 37°C
inkubiert, bis sie eine OD550 von 0,4 erreichte. Daraufhin erfolgte die Induktion der
Genexpression durch die Zugabe von 0,1 µg/ml Anhydrotetracyclin. Anschließend wurden
die Zellen für 4 h bei 37°C geschüttelt, danach durch Zentrifugation geerntet und das Pellet
bei -20 °C eingefroren.
2.8 Biochemische Methoden
2.8.1 Konzentrationsbestimmung von Proteinlösungen
Zur Bestimmung der Proteinkonzentration wurde ein Bio-Rad-Mikroassay nach BRADFORD
(1976) durchgeführt. Die Durchführung erfolgte nach Herstellerangaben unter Verwendung
von BSA als Standard.
2.8.2 Polyacrylamidgelelektrophorese (PAGE) und Western Blotting
Der Nachweis und die semiquantitative Analyse von spezifischen Proteinen in C. reinhardtii
Proteinrohextrakten und in Proteinproben erfolgte durch SDS-PAGE und Western Blotting.

Material und Methoden
19
2.8.2.1 Herstellung von Protein-Rohextrakten
1 ml Zellkultur wurde abzentrifugiert und das Pellet in 200 µl Proteinprobenpuffer (0.25 M
Tris-HCl, pH 8, 25 % Glycerin, 7.5 % SDS, 0, 25 mg/ml Bromophenolblau, 12.5% v/v β-
Mercaptoethanol) resuspendiert. Der Ansatz wurde für 5 min bei 95 °C inkubiert. Die Proben
wurden bei 4°C aufbewahrt und vor dem Auftragen auf das SDS-Gel ein weiteres Mal
aufgekocht, dann 1 min bei 14.000 rpm abzentrifugiert.
2.8.2.2 SDS-PAGE
Die elektrophoretische Auftrennung von Proteinen unter denaturierenden Bedingungen
erfolgte unter Verwendung diskontinuierlicher Glycin-SDS-Polyacrylamidgele nach
LAEMMLI (1970). Für die bessere Auflösung von Proteinen < 30 kDa wurden Tricin-SDS-
Polyacrylamidgele nach SCHÄGGER (2006) verwendet.
2.8.2.3 Native PAGE
Die native PAGE wurde analog zur SDS-PAGE durchgeführt. Jedoch enthielten die
Polyacrylamidgele und der Laufpuffer kein SDS.
2.8.2.4 Coomassie- Färbung von PAA-Gelen
Für die Coomassie-Färbung wurden SDS-Gele ca. 30 min in Färbelösung (0,1 % Coomassie
R250 in 10x Entfärber) inkubiert und anschließend mehrmals in 1x Entfärber (45 % Ethanol,
45 % Essigsäure, 10 % Wasser) gewaschen.
2.8.2.5 Silberfärbung von PAA-Gelen
Die Silberfärbung von Gelen erfolgte nach Standardmethoden (BLUM et al., 1987).
2.8.2.6 Western Blotting
Western Blotting wurde nach Standardmethoden, wie bereits beschrieben (HEMSCHEMEIER et
al., 2008 a), durchgeführt.
Folgende Antikörper wurden im Rahmen dieser Arbeit benutzt:
Antikörper Verdünnung Referenz
AdhE 1:2000 KESSLER , persönl. Mitteilung
AOX1 1:10000 Agrisera (Vännäs, Schweden)
Cyc6 1:1000 MERCHANT, persönl. Mitteilung
Fdx5 1:1000 diese Arbeit
HydA1 1:5000 HAPPE et al, 1994
TdcE (Kreuzreaktion gegen PFL) 1:5000 SAWERS, persönl. Mitteilung

Material und Methoden
20
PetF 1:1000 BÖHME, 1977
Rubisco (große Untereinheit) 1:5000 NEALE, 1993
Strep Tag II (Strep-Tactin AP Konjugat) 1:4000 IBA (Göttingen, Deutschland)
“ImmunoPure Goat Anti Rabbit IgG”, Peroxidase
gekoppelt
1:5000 Pierce (Rockford, IL, USA)
2.8.3 Eisenbestimmung
Die Bestimmung des Eisengehaltes in Proteinen erfolgte gemäß der Methode nach FISH
(1988). Dafür wurden 400 µl Proteinlösung mit 200 µl Lösung A (0.14 M
Kaliumpermanganat, 0.6 M HCl) versetzt und 2 h bei 60°C inkubiert. Nach kurzem Abkühlen
wurde 200 µl Lösung B (1.3 mM Ferrospektral, 2.6 mM Neocuproin, 0.4 M Natriumascorbat,
1 M Ammoniumacetat) dazugegeben und für 30 min bei RT inkubiert. Die Aufnahme der
Absorption bei 562 nm folgte nach einem kurzen Zentrifugationsschritt. Die Berechnung des
Fe-Gehaltes orientierte sich an einer mit definierten Eisenmengen erstellten Eichgerade.
2.8.4 Strep-Tag II Affinitätsaufreinigung
Alle Schritte der Aufreinigung wurden unter anaeroben Bedingungen in einem Anaerob-Zelt
durchgeführt. Das Zellpellet (siehe Kapitel 2.7.8) wurde eins zu eins in 100 mM Tris-HCl, pH
8, resuspendiert und die Zellen durch Ultraschall aufgeschlossen (6 Pulse á 30 sec, 1 min
Pause, 30 % - 40 % Power). Das Zelllysat wurde für 1 h bei 45000 rpm und 4°C
abzentrifugiert, um Zelltrümmer zu beseitigen. Der Überstand wurde mit 5 mg Avidin
versetzt und 30 min auf Eis inkubiert, um biotinylierte Proteine zu komplexieren.
Anschließend wurde der Proteinextrakt durch einen 0,2 µM Sterilfilter filtriert. Dieser Schritt
verhindert die Bindung biotinylierter Proteine aus der Zelle an die Strep-Tactin Säule. Der
klare Überstand wurde für die Affinitätsreinigung auf eine mit 100 mM Tris-HCl, pH 8 mit
150 mM NaCl äquilibrierte Strep-Tactin Säule gegeben. Nach mehrmaligem Waschen mit 20
ml 100 mM Tris-HCl, pH 8 erfolgte die Elution des Strep-TagII Fusionsproteins mit 2,5 mM
Desthiobiotin in 10 Fraktion von jeweils 1 ml.
2.8.5 Aufreinigung von nativem Fdx5
Für die Isolation von nativem Fdx5 aus C. reinhardtii wurden 10 l einer
Schwefelmangelkultur in der H2-produzierenden Phase durch Zentrifugation geerntet. Alle
Schritte der Aufreinigung wurden unter anaeroben Bedingungen durchgeführt. Der
Zellaufschluss erfolgte durch Ultraschall (4 Pulse á 30 sec, 1 min Pause, 30 % - 40 % Power).
Die Zelltrümmer wurden durch Zentrifugation (1 h, 45000 rpm, 4°C) entfernt und der

Material und Methoden
21
Proteinüberstand mit Ammoniumsulfat gefällt. Nach dem ersten Schritt (40 % Sättigung)
wurde die Proteinlösung zentrifugiert (1 h, 12000 rpm, 4°C) und der Überstand zu 80 %
Ammoniumsulfat gesättigt. Im Anschluss erfolgte eine weitere Zentrifugation. Das Pellet
wurde in 15 ml 50 mM Tris–HCl, pH 8 resuspendiert und gegen das 100-fache Volumen des
gleichen Puffers ü.N. dialysiert. Die Proteinlösung wurde auf eine mit 50 mM Tris-HCl, pH 8
äquilibrierte Q-Sepharose „fast-flow“ Säule (30 x 60 mm) (Amersham Bioscience, München,
Germany) aufgetragen. Nach dem Waschen mit dem gleichen Puffer wurde Fdx5 mit einem
linearen Salzgradienten (0–750 mM NaCl) eluiert. Die Fdx5 enthaltenden Fraktionen wurden
vereinigt, auf eine zweite Q-Sepharose „fast-flow“ Säule (20 x 35 mm) aufgetragen und mit
einem weiteren linearen Salzgradienten (0–500 mM NaCl) eluiert.
2.8.6 Herstellung von polyklonalen Fdx5 Antikörpern
Polyklonale Antikörper wurden durch die Immunisierung eines Kaninchens mit heterolog
produziertem Fdx5 hergestellt, von welchem der Strep-Tag durch Faktor Xa Protease nach
den Anleitungen des Herstellers (Novagen/ Merck, Darmstadt, Deutschland) entfernt wurde.
Die Antikörperproduktion wurde von BioGenes- Gesellschaft für Biopolymere (Berlin,
Deutschland) nach Standard-Immunisierungsprotokollen durchgeführt.
2.8.7 Massenspektrometrische Identifizierung und N-terminale Sequenzierung
Die massenspektrometrische Identifizierung wurde in Zusammenarbeit mit M. Nowaczyk
(Ruhr-Universität Bochum, Lehrstuhl Biochemie der Pflanzen, Bochum, Deutschland)
durchgeführt und erfolgte wie zuvor beschrieben (LAX et al., 2007). Die NH2-terminale
Sequenzierung erfolgte durch Edman-Abbau und wurde von der Proteome Factory AG
(Berlin, Deutschland) durchgeführt.
2.8.8 Bestimmung des Redoxpotentials mittel cyclischer Voltammetrie (CV)
Die Bestimmung des Redoxpotentials mittels cyclischer Voltammetrie erfolgte in
Kooperation mit H. Krassen (Universität Bielefeld, Fakultät für Chemie, Physikalische
Chemie III, Bielefeld, Deutschland). Die CV-Messung wurde in einem elektrochemischen
Drei-Elektroden-Aufbau mit der Goldelektrode als Arbeits-, dem Platinnetz als Gegen-, und
Ag/AgCl als Referenzelektrode durchgeführt. Die Konzentration der Proteinlösung betrug
100 µM.

Material und Methoden
22
2.8.9 Ferredoxin-abhängige Aktivitätstests
2.8.9.1 NDAP+-Photoreduktion
Die NDAP+-Photoreduktion entspricht in ihrem Ablauf einem Teil der photosynthetischen
Elektronentransportkette. Dabei wird Ascorbat als künstlicher Elektronendonor eingesetzt, der
mittels Plastocyanin als Elektronenüberträger Elektronen auf Photosystem I überträgt. Die
Elektronen werden durch Ferredoxin auf die Fnr transportiert, welche NADP+ zu NADPH
reduziert. Die Bildung von NADPH kann photometrisch bei 340 nm bestimmt werden. C.
reinhardtii Plastocyanin und PSI wurden im Rahmen einer Kooperation mit der
Arbeitsgruppe von Prof. M. Hippler am Institut für Biochemie und Biotechnologie der
Pflanzen der Universität Münster (Deutschland) bereitgestellt.
Der Testansatz für die NDAP+-Photoreduktion (FINAZZI et al., 2005) enthielt 10 mM MgCL2,
0,03 % β−Dodecylmaltosid, 5 mM Na-Ascorbat, 60 µM DCPIP, 10 µM NADP+, 5 µg Chl/ ml
PSI, 4 µM Plastocyanin 0,5 µM Fnr (S. oleocera) und variierende Ferredoxin-
Konzentrationen in 20 mM Tricin, pH 7,6. Die Reaktion wurde durch die Zugabe von
Ferredoxin gestartet und der Ansatz mit Weißlicht in einer Intensität von ca. 1000 µE/ m2 x s
bestrahlt. Alle 30 s wurde dem Ansatz ein Aliquot entnommen und die Absorption bei 340 nm
bestimmt. Der Extinktionskoeffizient von NADPH bei 340 nm beträgt ε340 = oder 6,22 x mM-
1 x cm-1.
2.8.9.2 Cytc-Reduktion
In dieser rekonstituierten Redoxkaskade wird die Fnr durch NADPH reduziert und überträgt
Elektronen auf Ferredoxin, welches wiederum Cytc reduziert. Die Reduktion von Cytc kann
photometrisch bei einer Wellenlänge von 550 nm bestimmt werden. Der Testansatz für die
Cytc-Reduktion enthielt 2 mM NADPH, 5 mM Cytc (Pferdeherz), 0,1 U/ml Fnr (S. oleocera)
und variierende Ferredoxin-Konzentrationen in 50 mM Tris-HCl, pH 8. Die Reaktion wurde
durch die Zugabe von Ferredoxin gestartet. Die Änderung der Absorption wurde
kontinuierlich mit dem Photometer gemessen. Der Extinktionskoeffizient für reduziertes Cytc
beträgt ε550 = 19,1 x mM-1 x cm-1.
2.8.9.3 HydA1-Reduktion
Um die Reduktion von HydA1 durch Ferredoxin zu untersuchen, wurde durch Natrium-
Dithionit reduziertes Ferredoxin mit 0,5 µg HydA1 (aufgereinigt und zur Verfügung gestellt
von S. Stripp aus dem Lehrstuhl für Biochemie der Pflanzen, AG Photobiotechnologie, Ruhr-
Universität Bochum, Deutschland) in 100 mM Tris HCl pH 8, 150 mM NaCl für 20 min bei

Material und Methoden
23
37°C inkubiert. Die Inkubation erfolgte unter anaeroben Bedingungen in einem
Gesamtvolumen von 200 µl in luftdicht verschlossenen Reaktionsgefäßen. Der freigesetzte
Wasserstoff wurde mittels Gaschromatographie nachgewiesen und quantifiziert.
2.8.10 Isolierung von C. reinhardtii Chloroplasten und Mitochondrien
S-Mangel-induzierte C. reinhardtii SAG 83.81 Zellen wurden in einem GSA Rotor (Sorvall/
Thermo Fisher Scientific, Waltham, USA) für 5 min zentrifugiert und in isotonischer Lösung
(0.3 M Sorbitol, 10 mM Tricin-HCl, pH 7.8, 5 mM MgCl2) resuspendiert. Die Zellsuspension
wurde mit dem „BioNeb Cell Disruption“ System (Glas-Col, Terre Haute, USA) bei einem
N2-Gasdruck von 20 p.s.i. aufgeschlossen und kurz bei 5000 rpm in einem SS34 Rotor
(Sorvall/ Thermo Fisher Scientific, Waltham, USA) zentrifugiert. Das Pellet wurde für die
Isolierung von intakten Chloroplasten (ZERGES und ROCHAIX, 1998), der Überstand für die
Isolierung von Mitochondrien genutzt (ERIKKSON et al., 1995)
2.9 Spektroskopische Methoden
2.9.1 Aufnahme von EPR (Electron Paramagnetic Resonance)- Signalen von Fdx5
Die EPR-spektroskopische Untersuchung von Fdx5 erfolgte in Zusammenarbeit mit S.
Pudollek an der Freien Universität Berlin, Fachbereich Physik, AG EPR-Spektroskopie von
biologischen Systemen. Für die Untersuchung wurden die Proteinproben auf eine
Konzentration von 200 µM eingestellt und mit 20 mM Natrium-Dithionit reduziert. Sie
wurden in EPR-Röhrchen überführt und in Isopropanpol eingefroren. Alle Experimente
wurden an einem Bruker Biospin Elexsys E580 Spektrometer (Bruker Corporation, Billerica,
USA), ausgestattet mit einem Super HighQ Resonator, durchgeführt. Die Aufnahme der
Spektren erfolgte bei einer Temperatur von T = 10 K und einer Mikrowellen-Frequenz von m
= 9.38 GHz. Es wurde eine Mikrowellen-Leistung von P = 4 mW und eine
Amplitudenmodulation von 0.5 mT bei 100 kHz Modulationsfrequenz genutzt.
2.9.2 Konfokale Laser Scanning Fluoreszenz Mikroskopie
Die Fluoreszenz-Emissionen von C. reinhardtii Transformanden wurden an einem Zeiss LSM
510 META Mikroskop (Carl Zeiss, Jena, Deutschland) untersucht. cGFP und Chloroplasten
wurden mit der 488 nm Linie eines Argon-Ionen-Lasers angeregt. Die Fluoreszenz-
Emissionen wurden unter Nutzung der Strahlenteiler HFT UV/488/543/633 und NFT545
durch den Bandpassfilter BP505-530 und den Langpassfilter LP560 selektiert.

Material und Methoden
24
2.9.3 Eisenbestimmung mittels Röntgenfluoreszenzanalyse
Die Röntgenfluoreszenzanalyse erfolgte in Kooperation mit M. Haumann an der freien
Universität Berlin, Fachbereich Physik, AG Biophysik und Photosynthese. Für die
Untersuchung wurden Proteinproben mit einer Konzentration von 100 µM eingesetzt. Die
Messung wurde an einem Bruker S2 PICOFOX (Bruker Corporation, Billerica, USA)
durchgeführt.
2.10 Physiologische Messungen
2.10.1 Chlorophyll- und Carotinoidbestimmung
1 ml einer C. reinhardtii Zellkultur wurde durch Zentrifugation sedimentiert und das Pellet
anschließend in 1 ml Aceton resuspendiert. Daraufhin wurde der Ansatz für 2 min bei 80°C
inkubiert. Nach erneuter Zentrifugation erfolgte die photometrische Bestimmung des
Chlorophyllgehalts des Überstandes bei 652 nm. Der Chlorophyllgehalt lässt sich mit
folgender Gleichung berechnen: Chlorophyllgehalt [µg x ml-1] = E652 x 27,8.
Die Extinktion der Carotinoide wurde unter Beachtung der Absorption von Chlorophyll a und
Chloropyll b berechnet: Carotinoidgehalt [µg x ml-1] = E480 + 0,114 x E663 - 0,638 x E645.
Der Carotinoridgehalt [µg x ml-1] kann dann folgendermaßen bestimmt werden: c = E480 x
0,2-1. (SCHOPFER, 1989)
2.10.2 In vitro- Test zur Messung der Hydrogenaseaktivität
Für die Bestimmung der in vitro Hydrogenaseaktivität wurden 0,5 ml Zellkultur in einem
anaeroben Reaktionsansatz (60 mM Kaliumphosphatpuffer pH 6,8, 1 % Triton X-100, 10 mM
Methylviologen und 100 mM Natrium-Dithionit) durch Schütteln aufgeschlossen. Im
Anschluss wurde der Ansatz eine Minute mit Argon gespült und 20 min bei 37°C und
konstantem Schütteln inkubiert. Der freigesetzte Wasserstoff wurde mittels
Gaschromatographie gemessen. Die Hydrogenaseaktiviät wurde auf den Chlorophyllgehalt
der Zellen und die Zeit (h) bezogen und kann folgendermaßen berechnet werden:
Hydrogenaseaktivität [nmol H2 x h-1 x µg Chl-1] = (area x 3 x 40 x nmol) / (0,5 x µg Chl x
Eichfaktor x h).

Material und Methoden
25
2.10.3 Nachweis von Fermentationsprodukten
Für den Nachweis von Fermentationsprodukten wurde 1 ml Zellkultur abzentrifugiert und der
Überstand mittels Enzymtests zum Nachweis von Formiat und Ethanol (r-biopharm,
Darmstadt, Deutschland) analysiert. Die Enzymtests wurden gemäß den Herstellerangaben
durchgeführt.

Ergebnisse
26
3 Ergebnisse
3.1 Im Genom von C. reinhardtii kodieren sechs Gene für Ferredoxine
Unter anaeroben S-Mangelbedingungen findet im Metabolismus von C. reinhardtii eine
grundlegende physiologische Umstrukturierung statt, durch welche die Transkription
verschiedenster Gene ein- oder abgeschaltet wird (JACOBS, Diplomarbeit, 2005). Auch die
Transkriptmengen der Ferredoxin-kodierenden Gene PETF und FDX5 sind unter diesen
Bedingungen einer Veränderung unterlegen (JACOBS Diplomarbeit, 2005; HEMSCHEMEIER
Diplomarbeit, 2002). Da Ferredoxine wichtige Elektronenüberträger sind, die auch in
verschiedenen anaeroben Stoffwechselwegen aktiv sind, war ein Ziel dieser Arbeit die
Klärung der Frage, ob noch weitere Ferredoxine-kodierende Gene in C. reinhardtii durch die
veränderten physiologischen Bedingungen unter S-Mangel und Anaerobiose reguliert werden.
Abb. 3-1: Sequenzvergleich der im Genom von C. reinhardtii kodierten Ferredoxine. Dargestellt ist ein
Sequenzvergleich zwischen PetF und den putativen Ferredoxinen Fdx2, Fdx3, Fdx4, Fdx5 und Fx6. Die den
Cluster koordinierenden Cysteinreste sind rot markiert. In PetF und Fdx5 sind die chloroplastidären
Proteaseschnittstellen, an denen das Transitpeptid abgetrennt wird, durch rote Pfeile gekennzeichnet (SCHMITTER
et al, 1988; diese Arbeit). Der Sequenzvergleich wurde mit dem Programm SECentral unter Verwendung der
Scoring-Matrix BLOSUM 62 erstellt.
Protein-BLAST Analysen gegen die JGI Datenbank (Version 3.0) des C. reinhardtii Genoms,
in denen die Proteinsequenz von PetF als Vergleich diente, deckten das Vorhandensein von
insgesamt sechs Genen in C. reinhardtii auf, die für Ferredoxine bzw. Ferredoxin-ähnliche
Proteine kodieren. Ein Sequenzvergleich der putativen Ferredoxine mit PetF zeigt, dass sie
signifikante Ähnlichkeiten zueinander aufweisen und dass in allen Polypetiden die
konservierten Cysteinreste vorkommen, die für die Koordination eines [2Fe2S]-Clusters

Ergebnisse
27
FDX5
PETF
FDX2
FDX3
FDX4
FDX60 h 24 h 48 h 72 h 0 h 0,5 h 4 h
A B
notwendig sind. Im Vergleich zu PetF zeigen insbesondere Fdx3 und Fdx6 auffälige C-
terminale Extensionen, wobei Fdx6 insgesamt große Sequenzabweichungen und Insertionen
gegenüber den anderen Ferredoxinen aufweist (Abb. 3-1).
Um die Transkription der Ferredoxin-kodierenden Gene unter S-Mangelbedingungen, sowie
unter artifiziell durch Argonbegasung induzierten anaeroben Bedingungen zu untersuchen,
wurden semiquantitative RT-PCRs durchgeführt, in denen mit spezifischen Primern für
PETF, FDX2, FDX3, FDX4, FDX5 und FDX6 kurze DNA-Fragmente der jeweiligen
Ferredoxin-kodierenden Gene amplifiziert wurden.
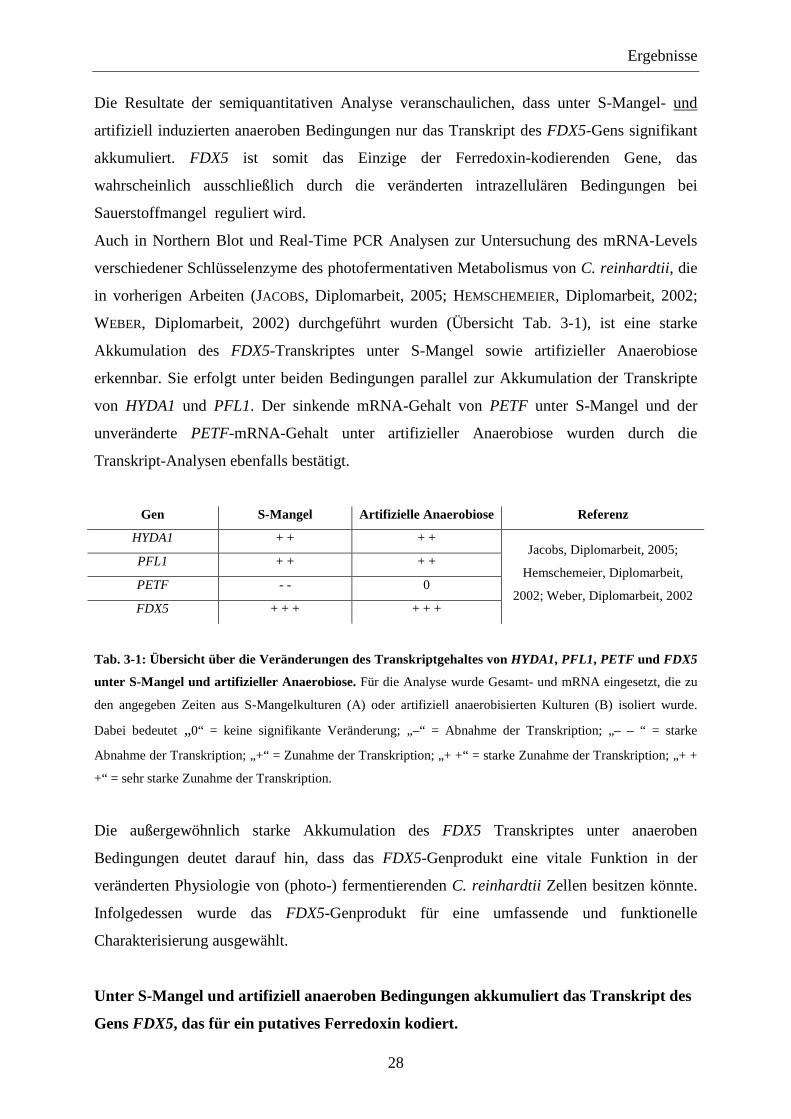
Abb. 3-2: Semiquantitative RT-
PCR Analyse zur Untersuchung
der Transkription der
Ferredoxin-kodierenden Gene
PETF, FDX2, FDX3, FDX4,
FDX5 und FDX6 aus C.
reinhardtii. Es wurde mRNA
eingesetzt, die zu den angegebenen
Zeitpunkten aus S-Mangelkulturen
(A) oder aus artifiziell durch
Begasung mit Argon anaerob
induzierten C. reinhardtii Kulturen
(B) isoliert wurde. Alle RT-PCRs
wurden mit 23 Zyklen
durchgeführt.
Die Ergebnisse der RT-PCRs zeigten (Abb. 3-2), dass sowohl unter S-Mangel als auch unter
artifiziell erzeugten anaeroben Bedingungen die Transkriptmengen aller Ferredoxin-
kodierenden Gene beeinflusst wird.
So ist unter S-Mangel eine Abnahme der Transkriptlevel von PETF, FDX3, FDX4 und FDX6
zu beobachten, während bei FDX2 und FDX5 eine Zunahme zu erkennen ist. Auch unter
artifiziell anaeroben Bedingungen wird eine Abnahme des FDX3, FDX4 und FDX6
Transkriptgehaltes deutlich, der Transkriptlevel von PETF bleibt unter diesen Bedingungen
jedoch konstant. Bei FDX2 kann im Gegensatz zu S-Mangelbedingungen kein Transkript
nachgewiesen werden, während bei FDX5 eine starke Akkumulation des Transkriptes auch
unter artifiziell erzeugten anaeroben Bedingungen deutlich wird.
Ergebnisse
28
Die Resultate der semiquantitativen Analyse veranschaulichen, dass unter S-Mangel- und
artifiziell induzierten anaeroben Bedingungen nur das Transkript des FDX5-Gens signifikant
akkumuliert. FDX5 ist somit das Einzige der Ferredoxin-kodierenden Gene, das
wahrscheinlich ausschließlich durch die veränderten intrazellulären Bedingungen bei
Sauerstoffmangel reguliert wird.
Auch in Northern Blot und Real-Time PCR Analysen zur Untersuchung des mRNA-Levels
verschiedener Schlüsselenzyme des photofermentativen Metabolismus von C. reinhardtii, die
in vorherigen Arbeiten (JACOBS, Diplomarbeit, 2005; HEMSCHEMEIER, Diplomarbeit, 2002;
WEBER, Diplomarbeit, 2002) durchgeführt wurden (Übersicht Tab. 3-1), ist eine starke
Akkumulation des FDX5-Transkriptes unter S-Mangel sowie artifizieller Anaerobiose
erkennbar. Sie erfolgt unter beiden Bedingungen parallel zur Akkumulation der Transkripte
von HYDA1 und PFL1. Der sinkende mRNA-Gehalt von PETF unter S-Mangel und der
unveränderte PETF-mRNA-Gehalt unter artifizieller Anaerobiose wurden durch die
Transkript-Analysen ebenfalls bestätigt.
Gen S-Mangel Artifizielle Anaerobiose Referenz
HYDA1 + + + +
PFL1 + + + +
PETF - - 0
FDX5 + + + + + +
Jacobs, Diplomarbeit, 2005;
Hemschemeier, Diplomarbeit,
2002; Weber, Diplomarbeit, 2002
Tab. 3-1: Übersicht über die Veränderungen des Transkriptgehaltes von HYDA1, PFL1, PETF und FDX5
unter S-Mangel und artifizieller Anaerobiose. Für die Analyse wurde Gesamt- und mRNA eingesetzt, die zu
den angegeben Zeiten aus S-Mangelkulturen (A) oder artifiziell anaerobisierten Kulturen (B) isoliert wurde.
Dabei bedeutet „0“ = keine signifikante Veränderung; „–“ = Abnahme der Transkription; „– – “ = starke
Abnahme der Transkription; „+“ = Zunahme der Transkription; „+ +“ = starke Zunahme der Transkription; „+ +
+“ = sehr starke Zunahme der Transkription.
Die außergewöhnlich starke Akkumulation des FDX5 Transkriptes unter anaeroben
Bedingungen deutet darauf hin, dass das FDX5-Genprodukt eine vitale Funktion in der
veränderten Physiologie von (photo-) fermentierenden C. reinhardtii Zellen besitzen könnte.
Infolgedessen wurde das FDX5-Genprodukt für eine umfassende und funktionelle
Charakterisierung ausgewählt.
Unter S-Mangel und artifiziell anaeroben Bedingungen akkumuliert das Transkript des
Gens FDX5, das für ein putatives Ferredoxin kodiert.

Ergebnisse
29
3.2 Heterologe Expression von FDX5 aus C. reinhardtii in E.coli
Für die heterologe Expression von FDX5 und PETF wurde das Strep-Tag System der Firma
IBA BioTAGnology (IBA GmbH, Göttingen, Deutschland) mit dem dazugehörigen
Expressionsplasmid pASK-IBA7 verwendet.
In diesem Plasmid ist der einklonierten Gensequenz eine 5’-terminal gelegene DNA-Sequenz
vorgeschaltet, die für einen Strep-Tag II kodiert. Der Strep-Tag ist durch einen kurzen Linker und
eine Faktor Xa- Schnittstelle mit dem Ziel-Protein verbunden. Die Expression eines Genes in
diesem System unterliegt der Kontrolle des TET-Promotors und ist durch die Zugabe von
Anhydrotetracyclin (AHT) induzierbar. Der TET-Repressor ist auf dem pASK-IBA Vektor kodiert
und wird konstitutiv exprimiert. Die Expression des klonierten Gens wird bis zur Induktion durch
AHT strikt unterdrückt. Im Gegensatz zum lac-Promotor ist der tet-Promotor streng kontrolliert,
so dass vor der Induktion keine Grundexpression des rekombinanten Proteins zu erwarten ist.
Das FDX5-Gen liegt auf Chromosom 17:702791-704769 in JGI4.0 (GenBank ABC88604)
und kodiert laut Annotation ein 1078 Bp langes Transkript. Das Transkript umfasst einen aus
393 Bp bestehenden offenen Leserahmen, der für ein 130 Aminosäuren (AS) großes Protein
kodiert. Basierend auf der Konsensusequenz Val-X-Ala für chloroplastidäre Signalpeptide
(FRANZEN et al., 1990) konnte eine putative Proteaseschnittstelle zwischen Position 27 und 28
des Proteins (Sequenz V-Q-A27↓F28) identifiziert werden (Abb. 3-3). Für die Klonierung in
ein Expressionsplasmid wurde daher nur die Gensequenz berücksichtigt, die für das native
Protein, exklusive des Transitpeptides, kodiert.
Abb. 3-3: Sequenz des offenen Leserahmens des FDX5-Gens
Dargestellt ist die cDNA-Sequenz des offenen Leserahmens des FDX5-Gens. Unterhalb der cDNA-Sequenz ist in
grüner Farbe die kodierte AS-Sequenz abgebildet. Die putative Proteaseschnittstelle für ein chloroplastidäres
Signalpeptid ist durch einen Pfeil gekennzeichnet. Das resultierende Transitpeptid ist grau unterlegt.
Für die Klonierung wurde die FDX5-cDNA mit den durch den IBA Primer D'Signer1.1
generierten Primern Fdx5_5_Strep und Fdx5_3_Strep, welche die für die gerichtete
Klonierung notwendige BsaI-Restriktionsschnittstelle bereits enthielten, amplifiziert. Das
durch die Klonierung erhaltene Expressionplasmid wurde als pJJ23 bezeichnet.
Um das Expressionsvermögen des erhaltenen Konstruktes zu überprüfen, wurde der
Expressionsstamm E. coli BL21 De3 plys mit pJJ23 transformiert (im Folgenden wird dieser
Ergebnisse
30
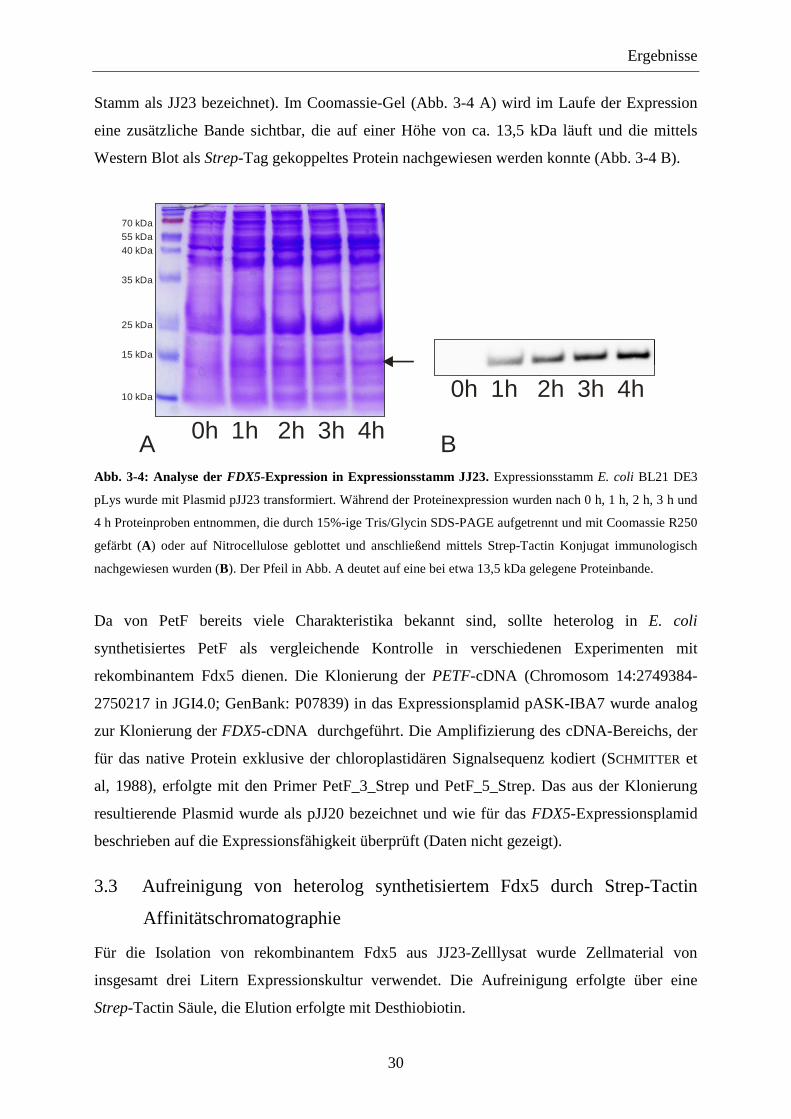
Stamm als JJ23 bezeichnet). Im Coomassie-Gel (Abb. 3-4 A) wird im Laufe der Expression
eine zusätzliche Bande sichtbar, die auf einer Höhe von ca. 13,5 kDa läuft und die mittels
Western Blot als Strep-Tag gekoppeltes Protein nachgewiesen werden konnte (Abb. 3-4 B).
0h 1h 2h 3h 4h
0h 1h 2h 3h 4h A B
10 kDa
15 kDa
25 kDa
35 kDa
40 kDa55 kDa70 kDa
Abb. 3-4: Analyse der FDX5-Expression in Expressionsstamm JJ23. Expressionsstamm E. coli BL21 DE3
pLys wurde mit Plasmid pJJ23 transformiert. Während der Proteinexpression wurden nach 0 h, 1 h, 2 h, 3 h und
4 h Proteinproben entnommen, die durch 15%-ige Tris/Glycin SDS-PAGE aufgetrennt und mit Coomassie R250
gefärbt (A) oder auf Nitrocellulose geblottet und anschließend mittels Strep-Tactin Konjugat immunologisch
nachgewiesen wurden (B). Der Pfeil in Abb. A deutet auf eine bei etwa 13,5 kDa gelegene Proteinbande.
Da von PetF bereits viele Charakteristika bekannt sind, sollte heterolog in E. coli
synthetisiertes PetF als vergleichende Kontrolle in verschiedenen Experimenten mit
rekombinantem Fdx5 dienen. Die Klonierung der PETF-cDNA (Chromosom 14:2749384-
2750217 in JGI4.0; GenBank: P07839) in das Expressionsplamid pASK-IBA7 wurde analog
zur Klonierung der FDX5-cDNA durchgeführt. Die Amplifizierung des cDNA-Bereichs, der
für das native Protein exklusive der chloroplastidären Signalsequenz kodiert (SCHMITTER et
al, 1988), erfolgte mit den Primer PetF_3_Strep und PetF_5_Strep. Das aus der Klonierung
resultierende Plasmid wurde als pJJ20 bezeichnet und wie für das FDX5-Expressionsplamid
beschrieben auf die Expressionsfähigkeit überprüft (Daten nicht gezeigt).
3.3 Aufreinigung von heterolog synthetisiertem Fdx5 durch Strep-Tactin
Affinitätschromatographie
Für die Isolation von rekombinantem Fdx5 aus JJ23-Zelllysat wurde Zellmaterial von
insgesamt drei Litern Expressionskultur verwendet. Die Aufreinigung erfolgte über eine
Strep-Tactin Säule, die Elution erfolgte mit Desthiobiotin.
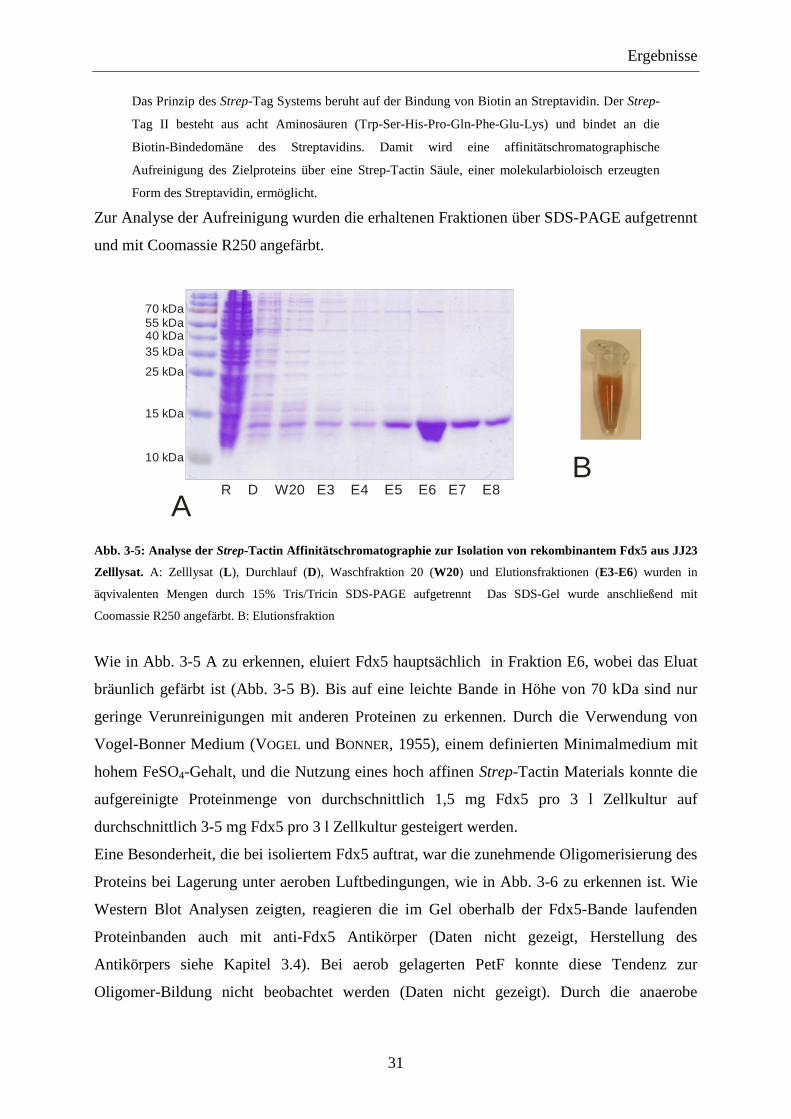
Ergebnisse
31
Das Prinzip des Strep-Tag Systems beruht auf der Bindung von Biotin an Streptavidin. Der Strep-
Tag II besteht aus acht Aminosäuren (Trp-Ser-His-Pro-Gln-Phe-Glu-Lys) und bindet an die
Biotin-Bindedomäne des Streptavidins. Damit wird eine affinitätschromatographische
Aufreinigung des Zielproteins über eine Strep-Tactin Säule, einer molekularbioloisch erzeugten
Form des Streptavidin, ermöglicht.
Zur Analyse der Aufreinigung wurden die erhaltenen Fraktionen über SDS-PAGE aufgetrennt
und mit Coomassie R250 angefärbt.
AB10 kDa
15 kDa
25 kDa
35 kDa40 kDa55 kDa70 kDa
R D W20 E3 E4 E5 E6 E7 E8
Abb. 3-5: Analyse der Strep-Tactin Affinitätschromatographie zur Isolation von rekombinantem Fdx5 aus JJ23
Zelllysat. A: Zelllysat (L ), Durchlauf (D), Waschfraktion 20 (W20) und Elutionsfraktionen (E3-E6) wurden in
äqvivalenten Mengen durch 15% Tris/Tricin SDS-PAGE aufgetrennt Das SDS-Gel wurde anschließend mit
Coomassie R250 angefärbt. B: Elutionsfraktion
Wie in Abb. 3-5 A zu erkennen, eluiert Fdx5 hauptsächlich in Fraktion E6, wobei das Eluat
bräunlich gefärbt ist (Abb. 3-5 B). Bis auf eine leichte Bande in Höhe von 70 kDa sind nur
geringe Verunreinigungen mit anderen Proteinen zu erkennen. Durch die Verwendung von
Vogel-Bonner Medium (VOGEL und BONNER, 1955), einem definierten Minimalmedium mit
hohem FeSO4-Gehalt, und die Nutzung eines hoch affinen Strep-Tactin Materials konnte die
aufgereinigte Proteinmenge von durchschnittlich 1,5 mg Fdx5 pro 3 l Zellkultur auf
durchschnittlich 3-5 mg Fdx5 pro 3 l Zellkultur gesteigert werden.
Eine Besonderheit, die bei isoliertem Fdx5 auftrat, war die zunehmende Oligomerisierung des
Proteins bei Lagerung unter aeroben Luftbedingungen, wie in Abb. 3-6 zu erkennen ist. Wie
Western Blot Analysen zeigten, reagieren die im Gel oberhalb der Fdx5-Bande laufenden
Proteinbanden auch mit anti-Fdx5 Antikörper (Daten nicht gezeigt, Herstellung des
Antikörpers siehe Kapitel 3.4). Bei aerob gelagerten PetF konnte diese Tendenz zur
Oligomer-Bildung nicht beobachtet werden (Daten nicht gezeigt). Durch die anaerobe

Ergebnisse
32
10 kDa
15 kDa
25 kDa
35 kDa
40 kDa55 kDa70 kDa
Aufreinigung und auch Lagerung des isolierten Proteins konnte die Oligomerisiung des
ansonsten monomer vorliegenden Proteins vermieden werden.
Abb. 3-6: Oligomer-Bildung in einer aerob
gelagerten Probe von rekombinantem Fdx5.
Unter aeroben Bedingungen aufgereinigtes und
gelagertes Protein wurde durch Elektropherese
eines 15%-igen Tris/Glycin SDS-Geles
aufgetrennt. Die Färbung des Geles erfolgte mit
Coomassie R250.
Rekombinantes Fdx5 kann in ausreichenden Mengen aus E. coli aufgereinigt werden.
Unter Lufteinfluss neigt das Protein zur Bildung von Oligomeren.
3.4 Gegen heterolog produziertes Fdx5 gerichtete Antiköper zeigen die
Akkumulation von Fdx5 in C. reinhardtii auf Proteinebene
Für die Herstellung von polyklonalen Antikörpern wurde ein Kaninchen nach einem
Standardprotokoll mit heterolog produziertem Fdx5 immunisiert. Um die Spezifität des
Antikörpers zu erhöhen, wurde der Strep-Tag vor der Immunisierung durch den Verdau mit
Faktor Xa vom Protein entfernt (Abb. 3-7 A)
Abb. 3-7: SDS-PAGE Analyse des mit Faktor Xa
verdauten Fdx5 und Western Blot mit anti-Fdx5
Antikörper zur Kontrolle von Kreuzreaktionen
mit PetF. A: 300 µg Fdx5 wurden mit 10 U Faktor
Xa ü.N. bei RT verdaut. Zur Kontrolle wurde ein
Aliquot des verdauten Proteins (-Tag) und des
unverdauten Original-Proteins (+Tag) in einem 15%-
igen SDS-Gel aufgetrennt und anschließend mit
Coomassie-R250 angefärbt. B: Zur Kontrolle auf
Kreuzreaktionen des anti-C. reinhardtii Fdx5
Antikörpers mit PetF wurden gleiche Mengen an
rekombinanten Fdx5 und PetF durch SDS-PAGE
aufgetrennt und geblottet. Der immunologische
Nachweis erfolgte durch Inkubation mit anti-C.
reinhardtii Fdx5 Antikörper.
- Tag + Tag
Fdx5 PetFB
A

Ergebnisse
33
In Immunoblots reagierte der hergestellte Antikörper mit heterolog produziertem Fdx5 und
zeigte keine Kreuzreaktionen mit PetF, wie in Abb. 3-7 B zu erkennen ist.
Abb. 3-8: Western Blot Analysen mit
anti-Fdx5 Antikörper weisen die
Akkumulation von Fdx5 in
schwefelmangelinduzierten C. reinhardtii
Zellen nach. Äqvivalente Mengen
Rohextrakt wurden mittels SDS-PAGE
aufgetrennt und nach Transfer auf
Nitrocellulose mittels anti-C. reinhardtii
HydA1 Antikörper, anti-E. coli TcdE
Antikörper, anti-S.oleocera PetF Antikörper
und anti-C. reinhardtii Fdx5 Antikörper
nachgewiesen.
Mit dem anti-Fdx5 Antikörper war es erstmals möglich die parallel zur Akkumulation von
FDX5-Transkript (Abb. 3-2, Tab. 3-1) verlaufende Akkumulation von Fdx5-Protein in
Rohextrakten von schwefelmangelinduzierten C. reinhardtii Zellen nachzuweisen (Abb. 3-8).
Unter aeroben Bedingungen kann kein Fdx5 detektiert werden. Die Zunahme von Fdx5
erfolgt gleichzeitig mit der Synthese von HydA1 und Pfl1. Interessanterweise bleibt die
Menge an PetF-Protein unter diesen Bedingungen konstant, obwohl auf Transkriptebene eine
Abnahme beobacht werden kann. Der für die Western Blots eingesetzte anti-S.oleocera PetF
Antikörper wurde auf Kreuzreaktionen mit Fdx5 untersucht und zeigt keinerlei Interaktion mit
dem rekombinanten Protein (Daten nicht gezeigt).
Unter S-Mangel kann eine Akkumulation von Fdx5-Protein beobachtet werden, die
parallel zur Akkumulation von HydA1 und Pfl1 verläu ft.
3.5 Partielle Aufreinigung von Fdx5 aus C. reinhardtii für die N-terminale
Sequenzierung
Um die N-terminale Sequenz des maturierten Fdx5-Proteins zu verifizieren, wurde Fdx5 aus
einer durch S-Mangel induzierten C. reinhardtii Kultur isoliert.
Die erhaltenen Elutionsfraktionen wurden durch Western Blot Analyse unter Anwendung des
anti-Fdx5 Antikörpers analysiert. Fdx5 eluiert dabei hauptsächlich in Fraktion drei bis fünf
PetF
Fdx5
HydA1
Pfl1
0 h 24 h 48 h 72 h

Ergebnisse
34
Fdx5
AB
des zweiten NaCl Gradienten Die Identifizierung des Fdx5-Bande im Coomassie R250
gefärbten SDS-Gel erfolgte im Anschluss durch Massenspektrometrie (Abb11).
Abb. 3-9: Fdx5 wurde mittels
Massenspektrometrie in Elutions-
fraktion vier identifiziert. A: Die
Auftrennung von Elutionsfraktion 4
erfolgte im 15 %-igen Tris-Tricin
Gel. Der Pfeil kennzeichnet die
Fdx5-Bande. B: Fdx5 wurde durch
Massenspektrometrie identifiziert
(B).
Die auf PVDF-Membran transferierte Fdx5-Bande wurde für die N-terminale Sequenzierung
durch Edman-Abbau genutzt und ermöglichte die Identifizierung der ersten fünf Aminosäuren
Phe-Gln-Val-Thr-Leu des nativen Proteins. Damit bestätigt die N-terminale Sequenzierung,
dass heterolog produziertes Fdx5 in seiner Aminosäuresequenz mit nativem, aus der Zelle
aufgereinigtem Fdx5, übereinstimmt. Folglich hat Fdx5 eine Länge von 103 AS und eine
Größe von 11,56 kDa. Darüber hinaus verifiziert das Ergebnis der Sequenzierung das
Vorhandensein eines N-terminalen Transitpeptides mit einer Größe von 27 AS.
3.6 Biochemische Charakterisierung von Fdx5
Das heterolog produzierte und aufgereinigte Ferredoxin, das in seiner Aminosäuresequenz
dem nativen in C. reinhardtii synthetisierten Protein entspricht, diente als Grundlage für eine
umfassende biochemische Charakterisierung, welche u.a. die Frage nach dem [FeS]-Cluster-
Typ von Fdx5 und dem Redoxpotential klären sollte.
Die Bestimmung des Eisengehaltes von rekombinantem Fdx5 mittels der Methode nach FISH
(1988) ergab, basierend auf einem Molekulargewicht von 11,56 kDa, ein Verhältnis von 2,05
± 0,23 mol Eisen/ Mol Protein. Die zur Kontrolle der Eisenbestimmung durchgeführte
Röntgenfluoreszenzspektroskopie bestätigte dieses Ergebnis mit einem Verhältnis von 2,27 ±
0,1 mol Eisen/ Mol Protein. Dies deutet daraufhin, dass das redoxaktive Zentrum von Fdx5
aus einem [2Fe2S]-Cluster besteht.
Die Eisenbestimmung nach FISH beruht auf einer sauren Lyse der Proteine, durch die der [FeS]-
Cluster freigesetzt und das Eisen in Lösung gebracht wird. Das freie Eisen wird durch
Ascorbinsäure zu Fe2+reduziert und von Ferrospektral komplexiert. Die Komplexierung des
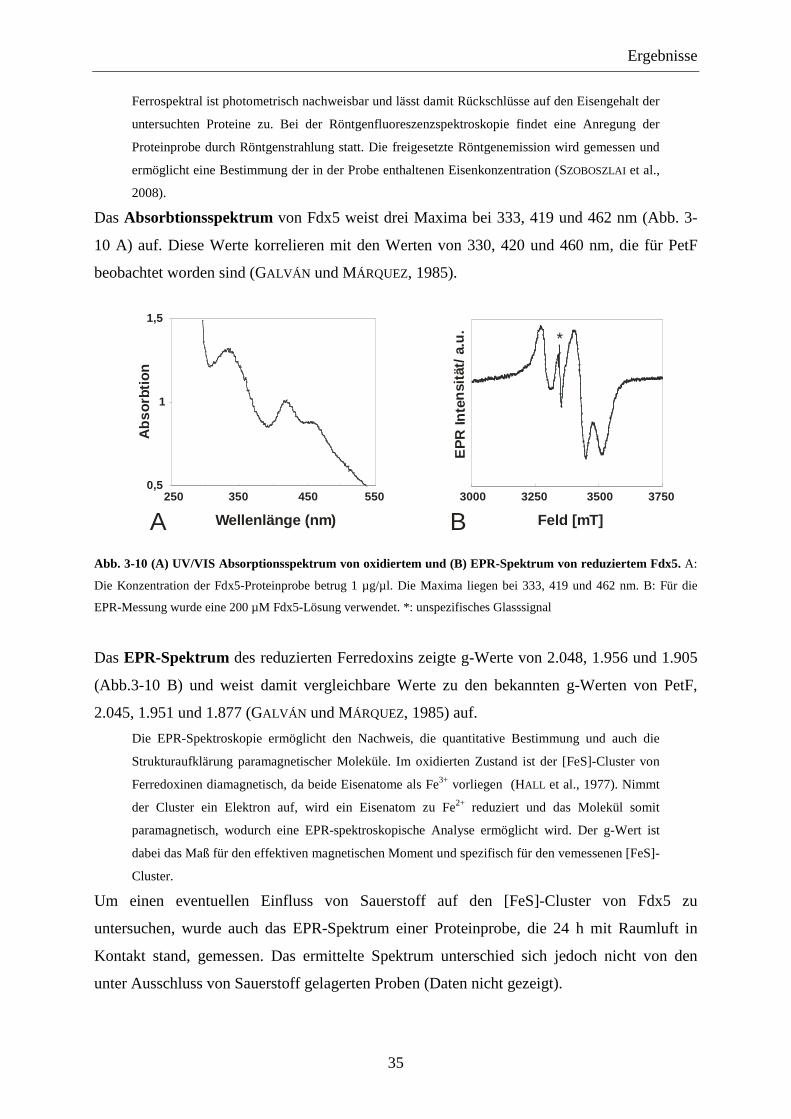
Ergebnisse
35
Ferrospektral ist photometrisch nachweisbar und lässt damit Rückschlüsse auf den Eisengehalt der
untersuchten Proteine zu. Bei der Röntgenfluoreszenzspektroskopie findet eine Anregung der
Proteinprobe durch Röntgenstrahlung statt. Die freigesetzte Röntgenemission wird gemessen und
ermöglicht eine Bestimmung der in der Probe enthaltenen Eisenkonzentration (SZOBOSZLAI et al.,
2008).
Das Absorbtionsspektrum von Fdx5 weist drei Maxima bei 333, 419 und 462 nm (Abb. 3-
10 A) auf. Diese Werte korrelieren mit den Werten von 330, 420 und 460 nm, die für PetF
beobachtet worden sind (GALVÁN und MÁRQUEZ, 1985).
Wavelength [nm] 3000 3250 3500 3750
0,5
1,0
1,5
250 300 350 400 450 500 550 3000 3250 3500 3750250 350 450 550
1,5
1
0,5
Ab
sorb
tio
n
EP
R In
ten
sitä
t/ a
.u.
Feld [mT]
*
Wellenlänge (nm)A B
Abb. 3-10 (A) UV/VIS Absorptionsspektrum von oxidiertem und (B) EPR-Spektrum von reduziertem Fdx5. A:
Die Konzentration der Fdx5-Proteinprobe betrug 1 µg/µl. Die Maxima liegen bei 333, 419 und 462 nm. B: Für die
EPR-Messung wurde eine 200 µM Fdx5-Lösung verwendet. *: unspezifisches Glasssignal
Das EPR-Spektrum des reduzierten Ferredoxins zeigte g-Werte von 2.048, 1.956 und 1.905
(Abb.3-10 B) und weist damit vergleichbare Werte zu den bekannten g-Werten von PetF,
2.045, 1.951 und 1.877 (GALVÁN und MÁRQUEZ, 1985) auf.
Die EPR-Spektroskopie ermöglicht den Nachweis, die quantitative Bestimmung und auch die
Strukturaufklärung paramagnetischer Moleküle. Im oxidierten Zustand ist der [FeS]-Cluster von
Ferredoxinen diamagnetisch, da beide Eisenatome als Fe3+ vorliegen (HALL et al., 1977). Nimmt
der Cluster ein Elektron auf, wird ein Eisenatom zu Fe2+ reduziert und das Molekül somit
paramagnetisch, wodurch eine EPR-spektroskopische Analyse ermöglicht wird. Der g-Wert ist
dabei das Maß für den effektiven magnetischen Moment und spezifisch für den vemessenen [FeS]-
Cluster.
Um einen eventuellen Einfluss von Sauerstoff auf den [FeS]-Cluster von Fdx5 zu
untersuchen, wurde auch das EPR-Spektrum einer Proteinprobe, die 24 h mit Raumluft in
Kontakt stand, gemessen. Das ermittelte Spektrum unterschied sich jedoch nicht von den
unter Ausschluss von Sauerstoff gelagerten Proben (Daten nicht gezeigt).
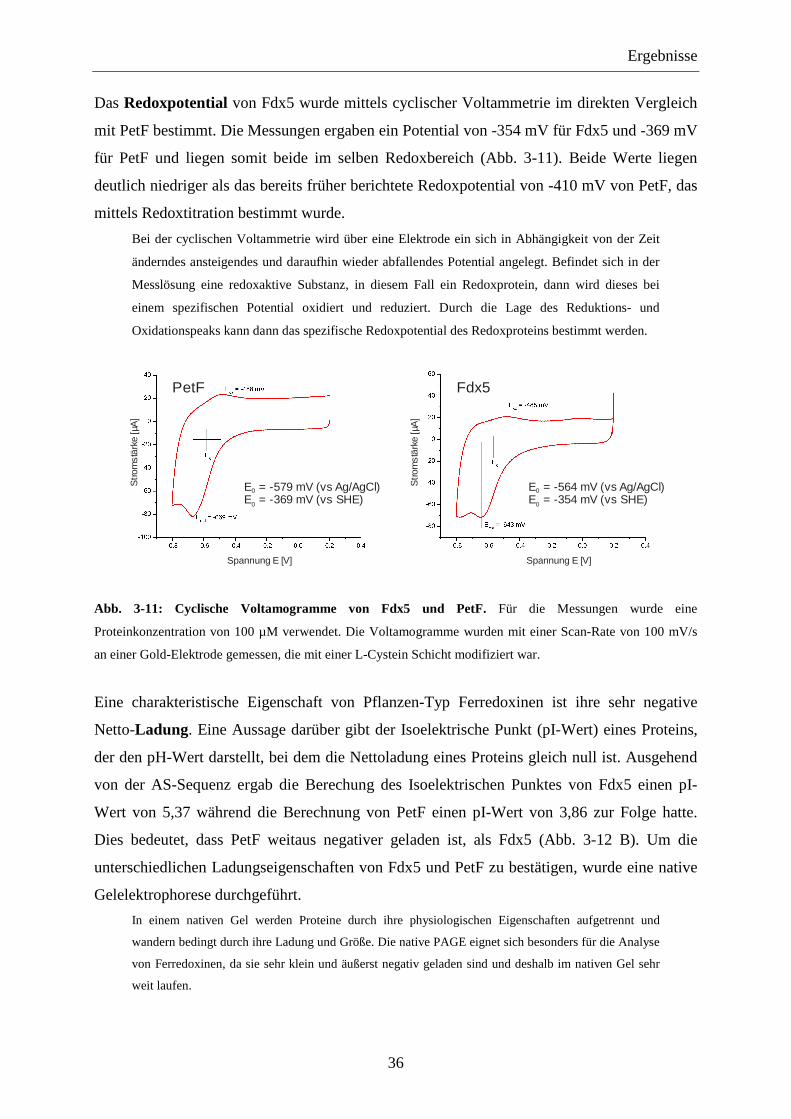
Ergebnisse
36
Das Redoxpotential von Fdx5 wurde mittels cyclischer Voltammetrie im direkten Vergleich
mit PetF bestimmt. Die Messungen ergaben ein Potential von -354 mV für Fdx5 und -369 mV
für PetF und liegen somit beide im selben Redoxbereich (Abb. 3-11). Beide Werte liegen
deutlich niedriger als das bereits früher berichtete Redoxpotential von -410 mV von PetF, das
mittels Redoxtitration bestimmt wurde.
Bei der cyclischen Voltammetrie wird über eine Elektrode ein sich in Abhängigkeit von der Zeit
änderndes ansteigendes und daraufhin wieder abfallendes Potential angelegt. Befindet sich in der
Messlösung eine redoxaktive Substanz, in diesem Fall ein Redoxprotein, dann wird dieses bei
einem spezifischen Potential oxidiert und reduziert. Durch die Lage des Reduktions- und
Oxidationspeaks kann dann das spezifische Redoxpotential des Redoxproteins bestimmt werden.
E = -579 mV (vs Ag/AgCl)E = -369 mV (vs SHE)
0
0
PetF Fdx5
E = -564 mV (vs Ag/AgCl)E = -354 mV (vs SHE)
0
0
Stro
mst
ärke
[µA
]
Stro
mst
ärke
[µA
]
Spannung E [V] Spannung E [V]
Abb. 3-11: Cyclische Voltamogramme von Fdx5 und PetF. Für die Messungen wurde eine
Proteinkonzentration von 100 µM verwendet. Die Voltamogramme wurden mit einer Scan-Rate von 100 mV/s
an einer Gold-Elektrode gemessen, die mit einer L-Cystein Schicht modifiziert war.
Eine charakteristische Eigenschaft von Pflanzen-Typ Ferredoxinen ist ihre sehr negative
Netto-Ladung. Eine Aussage darüber gibt der Isoelektrische Punkt (pI-Wert) eines Proteins,
der den pH-Wert darstellt, bei dem die Nettoladung eines Proteins gleich null ist. Ausgehend
von der AS-Sequenz ergab die Berechung des Isoelektrischen Punktes von Fdx5 einen pI-
Wert von 5,37 während die Berechnung von PetF einen pI-Wert von 3,86 zur Folge hatte.
Dies bedeutet, dass PetF weitaus negativer geladen ist, als Fdx5 (Abb. 3-12 B). Um die
unterschiedlichen Ladungseigenschaften von Fdx5 und PetF zu bestätigen, wurde eine native
Gelelektrophorese durchgeführt.
In einem nativen Gel werden Proteine durch ihre physiologischen Eigenschaften aufgetrennt und
wandern bedingt durch ihre Ladung und Größe. Die native PAGE eignet sich besonders für die Analyse
von Ferredoxinen, da sie sehr klein und äußerst negativ geladen sind und deshalb im nativen Gel sehr
weit laufen.

Ergebnisse
37
PetF Fdx5
Ferredoxin pI-Wert
PetF 3,86
Fdx5 5,87
A
B
Im nativen Gel (Abb. 3-12 A) ist zu erkennen, dass PetF, wie erwartet, als deutlich definierte
Bande sehr weit unten migriert. Fdx5 dagegen ist nicht als distinkte Bande erkennbar, sondern
zeigt sich als gering gewanderter, undefinierter Streifen sehr weit oberhalb der PetF-Bande.
Dies bestätigt damit die durch die pI-Werte angedeuteten Ladungseigenschaften der beiden
Ferredoxine PetF und Fdx5.
Abb. 3-12: Native PAGE
zur Analyse der Ladungs-
eigenschaften von Fdx5 im
Vergleich zu PetF native
Page und Darstellung der
jeweiligen PI-Werte. A: PetF
und Fdx5 wurden in
äqvivalenten Mengen durch
eine native PAGE
aufgetrennt. B: Darstellung
der pI-Werte von PetF und
Fdx5, ausgehend von der
Aminosäuresequenz.
Fdx5 ist ein [2Fe2S]-Ferredoxin mit ähnlichem Redoxpotential wie PetF. Fdx5 ist jedoch
sehr viel positiver geladen.
3.7 Zelluläre Lokalisation von Fdx5
Die N-terminale Sequenzierung von nativem Fdx5 bestätigte das Vorhandensein eines 27 AS
umfassenden N-terminalen Transitpetides, das möglicherweise den Transport des Proteins in
den Chloroplasten vermittelt. Um eine chloroplastidäre Lokalisation des Ferredoxins Fdx5 zu
überprüfen, wurden zwei unterschiedliche Strategien angewendet. Zum einen wurde ein
Fusionsprotein aus dem Fdx5-Transitpeptid und dem Grün Fluoreszierenden Protein (GFP)
hergestellt, um im Anschluss die Lokalisation des Fusionsproteins in vivo mittels Fluoreszenz
Mikroskopie zu beobachten. Zum anderen wurde die Technik der Zellfraktionierung
angewandt, um den Gehalt von Fdx5 in chloroplastidären und mitochondrialen Fraktionen
mittels Western Blot zu bestimmen.
Als Grundlage für die Herstellung von FDX5-Transitsequenz/ GFP-Konstrukten wurde das
Plasmid pMF59 (FUHRMANN et al., 1999) verwendet, dass für ein Ble/ GFP-Fusionsprotein
kodiert, das unter der Kontrolle des konstitutiven HSP70A/RBCS2-Tandempromotors steht

Ergebnisse
38
Das BLE-Gen kodiert für die Phleomycin-Resistenz aus Streptoalloteichus hindustanus. Das
kodierte bakterielle Protein wird in den Zellkern von C. reinhardtii transportiert (FUHRMANN et
al., 1999).
Die in pMF59 verwendete GFP-Sequenz wurde an die „Codon-Usage“ von C. reinhardtii
angepasst und besitzt einige zusätzliche Modifikationen, die die spektralen Eigenschaften des
Proteins verbessern (FUHRMANN et al., 1999). Durch Restriktion mit XcmI und FseI wurde ein
Großteil des BLE-Gens entfernt und an diese Position die das Transitpeptid kodierende
cDNA-Sequenz des FDX5-Gens eingefügt. Der verbleibende Rest des BLE-Gens verblieb als
eine Art Linker zum GFP-Gen. Abb. 3-13 A zeigt eine schematische Darstellung des
hergestellten Konstruktes. Das aus der Klonierung resultierende Plasmid pJJ24 wurde
zusammen mit einer Paromomycin-Resistenzkasette für die Co-Transformation von C.
reinhardtii CW 388- eingesetzt. Aus der Transformation hervorgegangene Klone wurden
durch Kolonie-PCR auf die Integration des FDX5-Transitsequenz/ GFP-Konstruktes
überprüft (Daten nicht gezeigt). Aus dieser Analyse gingen vier positive Transformanden
hervor, die im Anschluss durch konfokale Fluoreszenzmikroskopie analysiert wurden. In
diesen Messungen diente der mit pMF59 transformierte C. reinhardtii Stamm CC-125 als
Positivkontrolle, als Negativkontrolle wurde der untransformierte C. reinhardtii
Wildtypstamm CW 388- genutzt. Wie in Abb. 3-13 B zu erkennen ist, zeigte der
untransformierte Wildtyp während der Anregung von GFP keine beachtenswerte Fluoreszenz,
während die Positivkontrolle eine deutliche Akkumulation von GFP-Fluoreszenz im Zellkern
aufwies. Bei den das FDX5-Transitsequenz/ GFP-Konstrukt tragenden Transformanden
konnte ebenfalls eine eindeutige GFP-Fluoreszenz, die mit der Autofluoreszenz des
Chloroplasten co-lokalisierte, beobachtet werden. Zudem konnten in diesen Zellen
punktartige Akkumulationen von GFP-Fluoreszenz beobachtet werden, die in den
Randbereichen der Chlorophyll-Autofluoreszenz auftraten.

Ergebnisse
39
Durchlicht GFP Chlorophyll Überlagerung
Wildtyp
Ble/GFP
Fdx5-Transit-peptid/GFP
HSP70/RBCS2 BLEFDX5-Transits. GFP
A
B
Abb. 3-13: Konfokale Fluoreszenzmikroskopie für die Aufklärung der subzellulären Lokalisation von Fdx5. A:
Schematische Darstellung des hergestellten FDX5 Transitsequenz/ GFP-Konstruktes. B: Floureszenzmikroskopie von
C. reinhardtii CW 388- Transformanden wurden durch Durchlicht- oder konfokale Fluoreszenzmikroskopie
untersucht. Als Kontrolle dienten der Wildtyp C. reinhardtii CW 388- (= Wildtyp) und der mit pMF59 transformierte
C. reinhardtii Stamm CC-125 (= Ble/GFP).
Um die Lokalisation von Fdx5 durch eine weitere Methode zu bestätigen, wurden, wie bereits
oben erwähnt, chloroplastdidäre und mitochondriale Fraktionen von C. reinhardtii mittels
Immunoblotanalyse untersucht.
Für die Zellfraktionierung wurde der C. reinhardtii Stamm SAG 83.81 in der Wasserstoff-
produzierenden Phase einer S-Mangel Induktion verwendet. Nach einem schonenden
Zellaufschluss und einer kurzen Zentrifugation wurde das Pellet für die Isolation von intakten
Chloroplasten über einen diskontinuierlichen Percollgradienten aufgetrennt. Die Isolation der
Mitochondrien aus dem Überstand erfolgte über ein Percollkissen.
Die Coomassie R250 Färbung eines SDS-Gels, in dem Aliquots der erhaltenen Fraktionen
aufgetrennt wurden, zeigte, dass sich chloroplastidäre und mitochondriale Fraktionen in ihrem
Bandenmuster eindeutig unterschieden (Daten nicht gezeigt). Um die Reinheit der erhaltenen
Fraktionen zu überprüfen, wurde das Vorkommen spezifischer Markerproteine untersucht.
Als Marker für die chloroplastidäre Fraktionen dienten die große Untereinheit der Ribulose-

Ergebnisse
40
RbcL
HydA1
Aox1
Fdx5L C M
1,5-Bisphosphat-Carboxylase (Rubisco) und HydA1 (HAPPE et al., 1994), als Marker für die
mitochondriale Fraktion diente die Alternative Oxidase 1 (Aox1).
Abb. 3-14: Analyse der subzellulären Verteilung von
Fdx5 im Vergleich zur großen Untereinheit der
Rubisco (RbcL), zur Alternativen Oxidase (Aox1)
und zur Hydrogenase (HydA1). Für die Analyse
wurde ein Äqvivalent von 80 µg Protein des Zelllysats
(L), der Chloroplasten (C) und der Mitochondrien (M)
über eine 15%-ige Tris/Tricin SDS-PAGE aufgetrennt.
Aufgrund einer geringen Ausbeute wurde von den
Mitochondrien nur ein Äquivalent von 40 µg Protein
aufgetragen. Nach Transfer auf Nitrocellulose erfolgte
der immunologische Nachweis mit den entsprechenden
polyklonalen Antikörpern.
Eine Western Blot Analyse (Abb. 3-14) wies nach, dass die mitochondriale Fraktion frei von
chloroplastidären Proteinen war, die chloroplastidäre Fraktion hingegen mit mitochondrialen
Proteinen verunreinigt war. Das Fdx5 Signal ist in diesen Fraktionen ausschließlich auf die
chloroplastidäre Fraktion beschränkt und zeigt damit das gleiche Bandenmuster wie die
chloroplastidären Proteine Rubisco und HydA1.
Konfokale Fluoreszensmikroskopie und Western Blot Analysen mit isolierten
Chloroplasten und Mitochondrien von C. reinhardtii belegen eine chloroplastidäre
Lokalisation von Fdx5.
3.8 Funktionelle Charakterisierung von Fdx5 durch Ferredoxin-abhängige
Aktivitätstests
Um einen ersten Eindruck über die mögliche Funktion von Fdx5 im Metabolismus von
C. reinhardtii zu gewinnen, wurden verschiedene Ferredoxin-abhängige Aktivitätstests
durchgeführt. Als Positivkontrolle in diesen Analysen wurde heterolog produziertes PetF
genutzt.
Da die Transkription von Fdx5 ausschließlich unter anaeroben Bedingungen stattfindet, lag es
nahe, zunächst eine mögliche Interaktion mit der Hydrogenase HydA1 zu untersuchen, die

Ergebnisse
41
ein Schlüsselenzym des anaeroben Metabolismus von C. reinhardtii ist und Ferredoxin als
Elektronendonor benötigt.
Für die in vitro Tests mit der Hydrogenase wurde Ferredoxin mit NaDT reduziert und 20 min bei
37°C zusammen mit einem Aliquot HydA1 in luftdichten Reaktionsgefäßen inkubiert. Eine Probe
der Gasphase wurde anschließend durch Gaschromatographie analysiert.
In diesen Versuchen (Abb.3-15 A) konnte mit Fdx5 als Elektronendonor keine signifkante
Wasserstoffproduktion gemessen werden. Mit PetF als Elektronendonor hingegen konnte die
Reduktion von HydA1 durch die Bildung deutlicher Mengen an Wasserstoff beobachtet
werden. Die kinetischen Parameter dieser Reaktion mit PetF lagen bei einer maximalen
Hydrogenaseaktivität von 250 µmol H2 pro h und mg Protein und einem Km(PetF)-Wert von 5
µM und sind somit im Einklang mit früheren Beobachtungen (HAPPE und NABER, 1993). In
Folge dieses Ergebnisses scheint die Funktion von Fdx5 als nativer Redoxpartner von HydA1
ausgeschlossen zu sein.
Da die biochemischen Parameter von Fdx5 eine hohe Ähnlichkeit zu den Eigenschaften von
PetF aufweisen, wurde im nächsten Schritt geprüft, ob Fdx5 in typischen Elektronentransfer-
Reaktionen wie der NADP+-Photoreduktion mit PSI und Fnr aktiv ist.
In der photosynthetischen Elektronentransportkette werden Elektronen durch Plastocyanin auf PSI
übertragen, das wiederum Ferredoxin reduziert. Reduziertes Ferredoxin dient neben NADP+ und
H+ als Substrat für die Fnr, die NADPH als Reduktionsmittel für die Zelle bereitstellt. Für die
Messung einer nachweisbaren Interaktion zwischen PSI-Ferredoxin-Fnr wurde daher ausgehend
von Plastocyanin eine in vitro rekonstituierte Redoxkette genutzt (FINAZZI et al., 2005). Als
Reduktionsmittel für Plastocyanin diente in diesem in vitro System Natrium-Ascorbat, das mittels
DCPIP Elektronen auf Plastocyanin überträgt. Als weitere Komponenten wurden PSI, Ferredoxin,
Fnr und NADP+ eingesetzt. Durch Belichtung des in vitro Ansatzes wurde die Reaktion gestartet
und die Bildung von NADPH photometrisch gemessen.
Dabei konnte in Reaktionsansätzen, die PetF enthielten, eine eindeutige NADPH Produktion
beobachtet werden, während Reaktionsansätze mit Fdx5 keine NADPH Bildung aufwiesen
(Abb. 3-15 B). Somit scheint eine mögliche Funktion von Fdx5 in der NADP+-
Photoreduktion ebenfalls ausgeschlossen. Jedoch gibt dieser Versuch keinen Aufschluss
darüber, ob Fdx5 nicht durch PSI reduziert werden kann, ob Fdx5 nicht mit der Fnr
interagieren kann oder ob eine Wechselwirkung mit beiden Reaktionspartnern nicht möglich
ist.
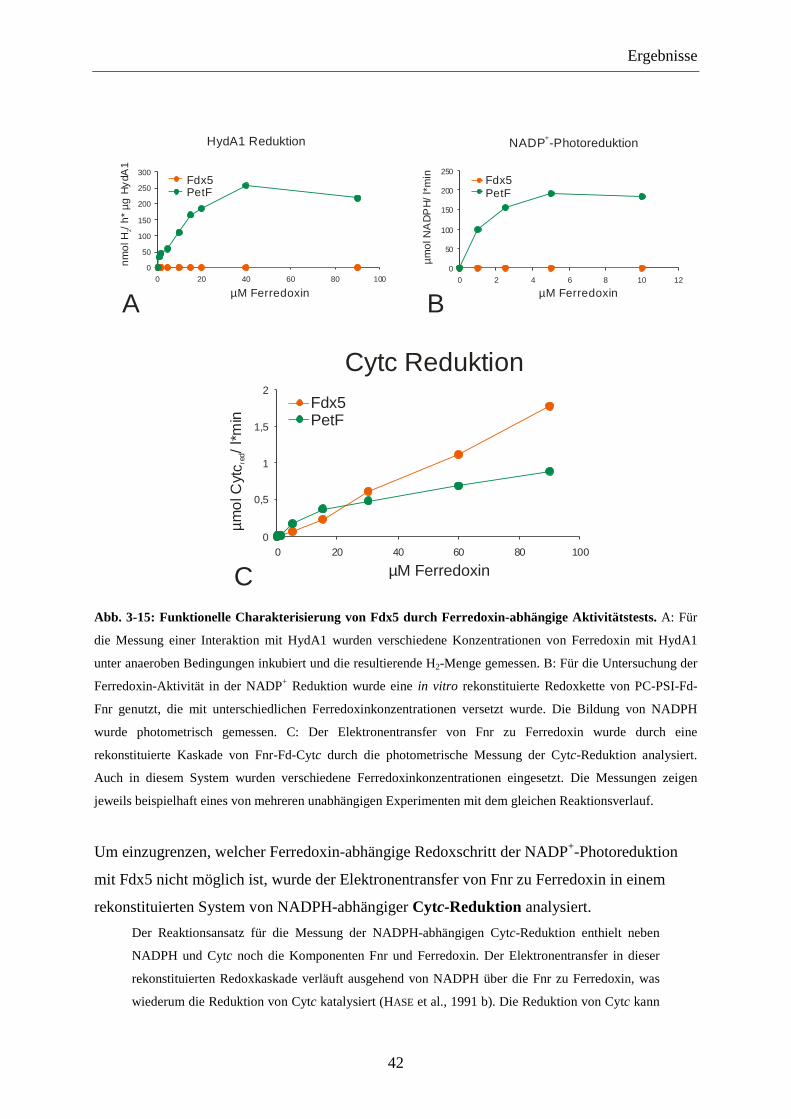
Ergebnisse
42
0
50
100
150
200
250
0 2 4 6 8 10 12
Fdx5
PetF
NADP -Photoreduktion+
0
50
100
150
200
250
300
0 20 40 60 80 100
Fdx5
PetF
HydA1 Reduktion
0
0,5
1
1,5
2
0 20 40 60 80 100
Fdx5
PetF
Cytc Reduktion
nm
ol H
/ h*
µg
Hy
dA1
2
µm
ol C
yt/
l*m
inc r
ed
µm
ol N
AD
PH
/ l*
min
µM Ferredoxin
µM Ferredoxin
µM FerredoxinA B
C
Fdx5 Fdx5PetF PetF
Fdx5PetF
Abb. 3-15: Funktionelle Charakterisierung von Fdx5 durch Ferredoxin-abhängige Aktivitätstests. A: Für
die Messung einer Interaktion mit HydA1 wurden verschiedene Konzentrationen von Ferredoxin mit HydA1
unter anaeroben Bedingungen inkubiert und die resultierende H2-Menge gemessen. B: Für die Untersuchung der
Ferredoxin-Aktivität in der NADP+ Reduktion wurde eine in vitro rekonstituierte Redoxkette von PC-PSI-Fd-
Fnr genutzt, die mit unterschiedlichen Ferredoxinkonzentrationen versetzt wurde. Die Bildung von NADPH
wurde photometrisch gemessen. C: Der Elektronentransfer von Fnr zu Ferredoxin wurde durch eine
rekonstituierte Kaskade von Fnr-Fd-Cytc durch die photometrische Messung der Cytc-Reduktion analysiert.
Auch in diesem System wurden verschiedene Ferredoxinkonzentrationen eingesetzt. Die Messungen zeigen
jeweils beispielhaft eines von mehreren unabhängigen Experimenten mit dem gleichen Reaktionsverlauf.
Um einzugrenzen, welcher Ferredoxin-abhängige Redoxschritt der NADP+-Photoreduktion
mit Fdx5 nicht möglich ist, wurde der Elektronentransfer von Fnr zu Ferredoxin in einem
rekonstituierten System von NADPH-abhängiger Cytc-Reduktion analysiert.
Der Reaktionsansatz für die Messung der NADPH-abhängigen Cytc-Reduktion enthielt neben
NADPH und Cytc noch die Komponenten Fnr und Ferredoxin. Der Elektronentransfer in dieser
rekonstituierten Redoxkaskade verläuft ausgehend von NADPH über die Fnr zu Ferredoxin, was
wiederum die Reduktion von Cytc katalysiert (HASE et al., 1991 b). Die Reduktion von Cytc kann

Ergebnisse
43
dabei photometrisch bestimmt werden (HASE et al., 1991 b) und ermöglicht eine Aussage über den
stattgefundenen Elektronentransfer.
In diesem System konnte sowohl mit PetF als auch mit Fdx5 eine signifikante Cytc-
Reduktion gemessen werden (Abb. 3-15 C). Zudem scheint mit Fdx5 eine deutlich höhere
Maximalgeschwindigkeit der Reaktion möglich zu sein. Dieses Ergebnis bestätigt somit eine
mögliche Interaktion zwischen Fnr und Ferredoxin und lässt indirekt die Aussage zu, dass
Fdx5 in der NADP+-Photoreduktion nicht von PSI reduziert werden kann.
Fdx5 fungiert nicht als Elektronendonor für HydA1, noch kann es als
Elektronenakzeptor von PSI dienen. Eine Interaktion zwischen Fnr und Fdx5 hingegen
ist möglich.
3.9 Postranskriptionelles Gen-„Silencing“ von FDX5 mittels artifizieller
miRNAs
Da die Ferredoxin-abhängigen Aktivitätstests keine genaue Aussage über die mögliche
Funktion von Fdx5 ermöglichten, wurde nach einer weiteren Strategie gesucht, um den
Funktionsbereich des Ferredoxins zu identifizieren. Ein häufig verwendetes Verfahren, um
die Funktion von Proteinen zu untersuchen, ist die Unterdrückung der Synthese der fraglichen
Proteine.
Auch in C. reinhardtii wurden bereits Protokolle etabliert, die, vermittelt durch die Methode der
RNA Interferenz, zum gezielten Abschalten eines Gens führen. Dabei werden in der Zelle lange
doppelsträngige RNA-Moleküle exprimiert, welche die Expression auf dem transkriptionellen oder
posttranskriptionellen Level beeinflussen (SCHRODA, 2006). Oftmals werden diese langen
Transgene jedoch selber durch das zelleigene „Silencing“-System ausgeschaltet, wodurch die
Phänotypen der Mutanten nach einiger Zeit verschwinden. (SCHRODA, 2006). Ein weiteres,
ähnliches System ist das „Silencing“ von Genen durch artifizielle miRNAs. microRNAs sind
kleine, 21-22 nt umfassende RNAs, die in Eukaryonten die post-transkriptionale Regulation durch
gerichteten mRNA Abbau und translationale Arretierung steuern (NAQVI et al., 2009). Kürzlich
wurde nachgewiesen, dass auch in C. reinhardtii die Genexpression durch microRNAs kontrolliert
wird (MOLNÁR et al, 2007). Von dieser Erkenntnis ausgehend wurde von Molar et al. ein potentes
Protokoll entwickelt, in dem artifizielle miRNAs genutzt werden, um die gezielte Degradation von
Transkripten in C. reinhardtii herbeizuführen (MOLNÁR et al.., 2009). Da miRNAs normalerweise
nicht Ziel des zelleigenen „Silencing“-Systems sind, werden die oben angeführten Nachteile der
RNAi-Methode mit dieser neuen „Silencing“-Methode beseitigt.

Ergebnisse
44
Gemäß dem von MOLNÁR et al. entwickelten System wurden in dieser Arbeit artifizielle
miRNA (amiRNA)-Konstrukte hergestellt, um den gezielten Abbau des FDX5-Transkriptes in
C. reinhardtii zu induzieren. Dafür wurden basierend auf der Zielsequenz mittels der Web
MicroRNA Designer Plattform (WMD2, http://wmd2.weigelworld.org/cgi-bin/mirnatools.pl)
zwei geeignete, unterschiedliche Bereiche des Zielgens abdeckende, miRNA Kandidaten für
FDX5 gesucht und mit diesen Sequenzen 90 nt umfassende Oligonukleotide erstellt, die nach
Annealing zu doppelsträngiger DNA in das Plasmid pChlamiRNA2 (MOLNÁR et al., 2009)
ligiert wurden. Die Expression der amiRNAs wird in diesem System durch den konstitutiven
HSP70A/RBCS2-Tandempromotor angetrieben. Zudem trägt pChlamiRNA2 den
Selektionsmarker ARG7. Mit den aus der Klonierung resultierenden Plasmiden pJJ27
(amiRNA 1) und pJJ28 (amiRNA 2) wurde eine Transformation mit C. reinhardtii
Wildtypstamm CW 388- durchgeführt. Jeweils sieben der erhaltenen amiRNA-
Transformanden wurden durch Begasung mit Argon anaerobisiert, um die Induktion der
FDX5-Genexpression auszulösen
Abb. 3-16: Analyse der FDX5- und HYDA1 Expression in artifiziell anaerobisierten amiRNA-
Transformanden. RNA wurde aus jeweils sieben amiRNA1- und amiRNA2-Transformanden nach
vierstündiger Argonbegasung isoliert. Für die Northern Blots wurden äquivalente Mengen an RNA eingesetzt.
Eine L10A (kodierend für das ribosomale Protein L10a) spezifische Sonde wurde als Ladekontrolle benutzt.
Northern Blot Analysen der RNA aus anaerobisierten amiRNA-Transformanden (Abb. 3-16)
zeigen einen im Vergleich zum Wildtypstamm CW 388- niedrigeren FDX5-Transkriptgehalt
in einem großen Teil der Transformanden. Dabei variieren die FDX5-Transkriptmengen in
unterschiedlichem Maße. Um auszuschließen, dass diese Variationen auf einen
unterschiedlichen Grad der Anaerobisierung zurückzuführen sind, wurde der Transkriptgehalt
des anaerob induzierten HYDA1-Gens analysiert. Dieser ist in allen untersuchten
Transformanden nahezu gleich und bestätigt somit einen ähnlichen Anaerobisierungsgrad der

Ergebnisse
45
mit Argon begasten Kulturen. Folglich sind die unterschiedlichen FDX5-Transkriptlevel der
amiRNA-Transformanden auf eine Expression der artifiziellen miRNA zurückzuführen.
Für eine weitergehende Charakterisierung wurden amiRNA-Transformanden 1.10 und 2.2
ausgewählt, da sie einen äußerst geringen FDX5-Transkriptgehalt aufwiesen. Im Vergleich
zum Wildtyp beträgt der FDX5-Transkriptlevel der Transformanden ca. 10 -20 %.
Durch die Expression artifizieller miRNAs konnten mehrere C. reinhardtii
Transformanden generiert werden, die einen verringerten FDX5-Transkriptgehalt
aufweisen.
3.9.1 Physiologische Analysen der amiRNA-Transformanden
Für eine datailierte Charakterisierung wurden amiRNA-Transformanden 1.10 und 2.2 unter S-
Mangelbedingungen sowie anaerober Induktion untersucht. Bei diesen Analysen wurde ein
Schwerpunkt auf die Schlüsselreaktionen des Fermentationsmetabolismus und der
Wasserstoff-, Formiat- und Ethanolproduktion gelegt, um eine eventuelle Beteiligung von
Fdx5 in diesen Stoffwechselwegen zu erkennen.
Es wurde ein Hauptaugenmerk auf die oben angegebenen Fermentionsprodukte gerichtet, da
Ferredoxin als Elektronendonor im Rahmen der Aktivierung der Pyruvat-Formiat-Lyase Pfl1
durch die Pfl-Aktivase und als Substrat der Pyruvat-Ferredoxin-Oxidoreduktase eine Rolle spielen
könnte (HEMSCHEMEIER et al., 2008 a).
In ersten Experimenten konnten unter S-Mangelbedingungen keine gravierenden
Unterschiede zwischen dem Wildtypstamm CW 388- und den amiRNA-Transformanden
beobachtet werden. Der Wasserstoffgehalt in der Gasphase über den Zellen sowie die Mengen
an Formiat und Ethanol in den Medien der jeweiligen Stämme waren ähnlich (Daten nicht
gezeigt).
Auch bei der artifiziellen anaeroben Induktion (Abb. 3-17) zeigten alle drei Stämme ein
übereinstimmendes Verhalten in der gemessenen in vitro Hydrogenaseaktivität und der
Formiat-und Ethanolproduktion.

Ergebnisse
46
0
5
10
15
20
25
1 2 3
3885.2_25.1_10
0
5
10
15
20
1 2 3
3885.2_25.1_10
0
50
100
150
200
250
300
0 1 2 3 4
3885.2_25.1_10
nmol
H/ h
*µg
Chl
2 mg/
l
mg/
l
Zeit [h]Zeit [h]Zeit [h]
in vitroHydrogenaseaktivität Formiatproduktion Ethanolproduktion
Abb. 3-17: Analyse von Schlüsselreaktionen des Fermentationsmetabolismus der artifiziell
anaerobisierten amiRNA Transformanden im Vergleich mit dem Wildtypstamm CW 388-. Für die
Untersuchung wurden die betreffenden C. reinhardtii -Kulturen 3 h mit Argon begast. Nach 1 h, 2 h und 3 h
wurde die in vitro Hydrogenaseaktivität in einem reduziertes Methylviologen enthaltenden Reaktionsansatz
ermittelt und es wurden Proben entnommen, die auf Formiat und Ethanol untersucht wurden. Die Abbildungen
zeigen jeweils beispielhaft eines von zwei unabhängigen Experimenten, welche die gleichen Tendenzen
aufwiesen.
Es ist jedoch noch ein weiterer Metabolismus, an dem Fdx5 beteiligt sein könnte, möglich,
der bislang noch nicht betrachtet wurde. Bei kürzlich durchgeführten Analysen zur
differentiellen Expression von Genen unter Kupfermangel konnte ebenfalls eine starke
Akkumulation des FDX5-Transkriptes beobachtet werden (S. Merchant, persönl.
Kommunikation). Dabei zeigte FDX5 die zweitstärkste Transkriptakkumulation der
untersuchten Gene. Um eine mögliche Beteiligung von Fdx5 am veränderten Metabolismus
unter Cu-Mangel zu untersuchen, wurden deshalb Wachstumskurven der Cu-Mangel-
induzierten amiRNA-Transformanden im Vergleich zum Wildtyp durchgeführt.
Kupfer ist für die meisten Organismen ein bedeutender Nährstoff, da Kupfer ein wichtiger
Kofaktor von Proteinen ist, die an Elektronentransferreaktionen beteiligt sind, wie z.B.
Plastocyanin oder Superoxiddismutasen. Cu-Mangel führt daher, wie S-Mangel, zu weitreichenden
Stoffwechselveränderungen der Zellen (MERCHANT et al., 2006). In C. reinhardtii ist der
veränderte Metabolismus unter Cu-Mangel bereits intensiv untersucht worden (QUINN und
MERCHANT, 1998, ERIKSSON et al., 2004)). Zu den bedeutendsten Adaptionen, die in C.
reinhardtii unter Cu-Mangel beobachtet werden können, gehört beispielsweise die starke
Expression des CYC6-Gens, das für das Elektronentransferprotein Cytochrom c6 kodiert (WOOD,
1978; MERCHANT und BOGORAD, 1987). Cyc6 enthält als Kofaktor ein Häm-Molekül und ersetzt
unter Cu-Mangel vollständig das Cu-enthaltende Plastocyanin der Elektronentransportkette.
Bei den Untersuchungen der Cu-defizienten Stämme konnten interessante Unterschiede
beobachtet werden: Beide amiRNA-Transformanden zeigten unter Cu-Mangel einen im
Vergleich zum Wildtyp Stamm CW 388- eindeutig reduzierten Chlorophyll- und
Carotinoidgehalt (Abb. 3-18 A). Diese Unterschiede sind nicht auf eine eine verringerte
Zellmenge zurückzuführen, da die Zellzahl bei allen drei Stämmen annähernd
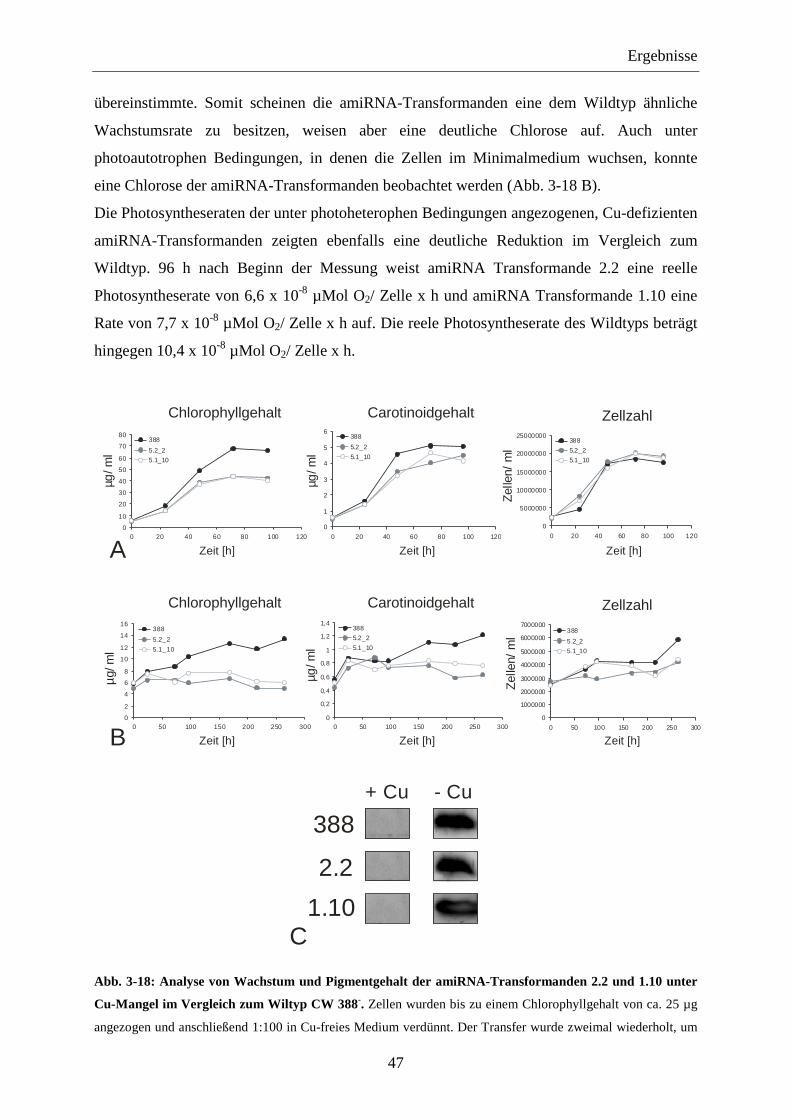
Ergebnisse
47
übereinstimmte. Somit scheinen die amiRNA-Transformanden eine dem Wildtyp ähnliche
Wachstumsrate zu besitzen, weisen aber eine deutliche Chlorose auf. Auch unter
photoautotrophen Bedingungen, in denen die Zellen im Minimalmedium wuchsen, konnte
eine Chlorose der amiRNA-Transformanden beobachtet werden (Abb. 3-18 B).
Die Photosyntheseraten der unter photoheterophen Bedingungen angezogenen, Cu-defizienten
amiRNA-Transformanden zeigten ebenfalls eine deutliche Reduktion im Vergleich zum
Wildtyp. 96 h nach Beginn der Messung weist amiRNA Transformande 2.2 eine reelle
Photosyntheserate von 6,6 x 10-8 µMol O2/ Zelle x h und amiRNA Transformande 1.10 eine
Rate von 7,7 x 10-8 µMol O2/ Zelle x h auf. Die reele Photosyntheserate des Wildtyps beträgt
hingegen 10,4 x 10-8 µMol O2/ Zelle x h.
0
0,2
0,4
0,6
0,8
1
1,2
1,4
0 50 100 150 200 250 300
388
5.2_2
5.1_10
0
10
20
30
40
50
60
70
80
0 20 40 60 80 100 120
388
5.2_2
5.1_10
0
1
2
3
4
5
6
0 20 40 60 80 100 120
388
5.2_2
5.1_10
0
5000000
10000000
15000000
20000000
25000000
0 20 40 60 80 100 120
388
5.2_2
5.1_10
µg/ m
l
Zeit [h] Zeit [h]Zeit [h]
Zel
len/
ml
Zel
len/
ml
Chlorophyllgehalt
Chlorophyllgehalt
Carotinoidgehalt
Carotinoidgehalt
Zellzahl
Zellzahl
0
2
4
6
8
10
12
14
16
0 50 100 150 200 250 300
388
5.2_2
5.1_10
0
1000000
2000000
3000000
4000000
5000000
6000000
7000000
0 50 100 150 200 250 300
388
5.2_2
5.1_10
Zeit [h]Zeit [h]Zeit [h]
µg/
ml
µg/ m
l
µg/ m
l
A
B
C
388
2.2
1.10
- Cu+ Cu
Abb. 3-18: Analyse von Wachstum und Pigmentgehalt der amiRNA-Transformanden 2.2 und 1.10 unter
Cu-Mangel im Vergleich zum Wiltyp CW 388-. Zellen wurden bis zu einem Chlorophyllgehalt von ca. 25 µg
angezogen und anschließend 1:100 in Cu-freies Medium verdünnt. Der Transfer wurde zweimal wiederholt, um

Ergebnisse
48
- Cu+ Cu + S - S
388
2.2
1.10
die Verschleppung von Cu zu vermeiden und sicherzustellen, dass verbleibendes Cu von den Zellen
aufgebraucht wurde. Anschließend erfolgte ein weiterer Transfer in kupferfreies TAP-Medium (A), bzw.
kupferfreies TMP-Medium (B). Die Kulturen wurden dabei auf einen anfänglichen Chlorophyllgehalt von 5 µg/
ml eingestellt. Zu den angegebenen Zeiten wurden Proben entnommen und auf Chlorophyll- und
Carotinoidgehalt untersucht. Zeitgleich erfolgte die Bestimmung der Zellzahl. Die Messungen zeigen jeweils
beispielhaft eines von zwei unabhängigen Experimenten mit dem gleichen Reaktionsverlauf. Um die Cu-Mangel
Induktion zu bestätigen, wurden nach dreimaligem Transfer der Kulturen in kupferfreies Medium Proben
entnommen, mittels SDS-PAGE aufgetrennt und nach Blotting auf ihren Cyc6-Gehalt untersucht (C).
Um zu untersuchen, ob die beobachteten Phänotypen wirklich auf einen veränderten Fdx5-
Gehalt der amiRNA-Transformanden im Vergleich zum Wildtyp CW 388- zurückzuführen
sind, wurden Immuno Blots durchgeführt (Abb. 3-19). Diese zeigen, dass unter Cu-
supplementierten, aeroben Bedingungen kein Fdx5-Protein detektiert werden kann. Unter Cu-
Mangel wie auch unter S-Mangel kann Fdx5 hingegen nachgewiesen werden. Jedoch zeigen
die amiRNA-Stämme dabei unter Cu-Mangel eindeutig eine Reduktion des Fdx5-Gehaltes um
mehr als 50 % im Vergleich zum Wildtypstamm CW 388-. Unter S-Mangelbedingungen
hingegen ist der Fdx5-Level der amiRNA-Transformanden im Vergleich zum Wildtyp
konstant.
Abb. 3-19: Analyse des Fdx5-Gehaltes der
amiRNA-Stämme 2.2 und 1.10 unter Cu-
und S-Mangel. Äquivalente Proteinmengen
wurden über eine 15 %-ige Tris/Tricin-PAGE
aufgetrennt und nach Transfer auf PVDF mit
anti-Fdx5 Antikörper inkubiert. Cu-Mangel
Proben wurden nach dreimaligem Transfer
der Kultur in kupferfreies Medium
genommen. S-Mangelproben 48 h nach
Überführung der Kultur in S-Mangelmedium .
Unter Cu-Mangelbedingungen sind der Pigmentgehalt und die Photsyntheserate der
amiRNA-Transformanden 2.2 und 1.10 reduziert

Diskussion
49
4 Diskussion
C. reinhardtii besitzt sechs Ferredoxin-kodierende Gene, die unter S-Mangel differentiell
exprimiert werden.
Ferredoxine sind in der Natur weit verbreitetete Elektronenüberträger. In Pflanzen und Algen
können Ferredoxine gefunden werden, deren redoxaktives Zentrum aus einem [2Fe2S]-
Cluster besteht und die als Pflanzen-Typ Ferredoxine bezeichnet werden. Sie sind wichtig für
eine Vielzahl von metabolischen Reaktionen und haben ebenfalls eine Funktion in Signal-
Prozessen. Es ist bereits bekannt, dass in vielen Pflanzen mehrere Ferredoxin-Isoformen
gefunden werden können. Beispiele dafür sind die Ferredoxine aus Zea mays oder
Arabidopsis thaliana. In Z. mays wurden bislang vier Ferredoxin-Isoformen (FdI-FdIV)
gefunden, von denen FdI und FdIII die am häufigsten auftetenden Isoformen in Blättern bzw.
im Wurzelgewebe darstellen (KIMATA und HASE, 1989; HASE et al., 1991 b). In Wurzeln kann
Ferredoxin durch die Oxidation von NADPH im Rahmen der Umkehrreaktion der Fnr
reduziert werden (SUZUKI et al., 1985), was die Aufrechterhaltung von Ferredoxin-
abhängigen Stoffwechselwegen wie z.B. Nitrit- und Sulfit-Reduktion in nicht-
photosynthetisch aktiven Geweben ermöglicht. NADPH wird dabei durch den oxidativen
Pentosephosphatweg bereitgestellt (BOWSHER et al., 1989). Untersuchungen an
Wurzelferredoxinen im Vergleich zu Blattferredoxinen haben funktionelle Unterschiede
zwischen beiden Ferredoxin-Gruppen aufgedeckt, da Wurzelferredoxine besser durch
NADPH reduziert werden können, während Blattferredoxine höhere Raten in der NADP+-
Photoreduktion erreichen (HASE et al., 1991 b, ONDA et al., 2000, HANKE et al., 2004). In A.
thaliana konnten sechs Ferredoxin-kodierende Gene im Genom gefunden werden. Vier der
kodierten Proteine entsprachen konventionellen [2Fe2S]-Ferredoxinen und wurden
weitergehend untersucht. AtFd1 und AtFd2 konnten dabei als typische Blatt-Ferredoxine
identifiziert werden, während AtFd3 Ähnlichkeiten zu Wurzel-Ferredoxinen zeigt. AtFd4
weist zwar Ähnlichkeiten zu Wurzel-Ferredoxinen auf, besitzt aber ein ungewöhnlich
positives Redoxpotential und kann deshalb keine Elektronen auf die Fnr transferieren (HANKE
et al., 2004).
Auch in C. reinhardtii konnten im Rahmen dieser Arbeit insgesamt sechs Ferredoxin-
kodierende Gene (PETF, FDX2, FDX3, FDX4, FDX5 und FDX6) identifiziert werden, die
unter S-Mangelbedingungen bzw. artifiziell anaeroben Bedingungen differentiell exprimiert
werden. Als Ausgangsbasis für die Analyse der Genexpression mittels semiquantitativer RT-
PCR wurden luftdichtverschlossene S-Mangel-induzierte C. reinhardtii Kulturen verwendet,

Diskussion
50
die bei Belichtung nach ca. 24 h aufgrund einer verringerten Photosyntheserate auf
physiologischem Wege Anaerobiose etablierten (MELIS et al. 2000; WYKOFF et al. 1998).
Dabei konnten zwei Ferredoxin-Gene, FDX5 und FDX2, identifiziert werden, deren
Transkripte unter diesen Bedingungen akkumulierten. Es muss jedoch beachtet werden, dass
sich unter den oben beschriebenen Bedingungen zwei potentiell einflussnehmende Faktoren
ändern: der Schwefelgehalt der Zellen und dadurch bedingt auch der Sauerstoffgehalt der
Kultur. Allein durch den Mangel an Schwefel werden in der Zelle eine Reihe von
Veränderungen induziert, die sich z.B. in einer verstärkten Expression von Genen äußern,
deren Proteinprodukte an der Assimilation von Sulfat beteiligt sind (LIEN und SCHREINER,
1975; YILDIZ et al., 1996; RAVINA et al., 1999; SATO et al., 2000). Dabei ist die
Sulfatassimilation ebenfalls ein Prozess, der das Vorhandensein reduzierter
Elektronenüberträger voraussetzt. Für die Reduktion von Sulfit zu Sulfat werden
beispielsweise 6 Elektronen benötigt, die durch reduziertes Ferredoxin bereitgestellt werden.
In C. reinhardtii konnte bereits PetF als ein Interaktionspartner der Sulfitreduktase
identifiziert werden (GARCIA-SÁNCHEZ et al., 2000). Es wäre jedoch möglich, dass unter S-
Mangel die Synthese eines spezialisierten Ferredoxins stattfindet, das höhere Affinitäten zu
seinen Redoxpartnern aufweist und damit zu einem höheren Sulfatumsatz beiträgt.
In dieser Arbeit waren jedoch die Ferredoxin-Gene von besonderem Interesse, deren
Expression eindeutig durch den Faktor Anaerobiose induziert werden konnte. Für eine weitere
Analyse der Genexpression wurden deshalb Kulturen verwendet, die mit dem inerten Gas
Argon begast wurden. Bei dieser Methode ist der einzige Parameter, der verändert wird, die
Sauerstoffkonzentration der Kultur. Unter diesen artifiziell anaeroben Bedingungen konnte
nur die Akkumulation des FDX5-Transkriptes induziert werden, was den Umkehrschluss
erlaubt, das FDX2 scheinbar nur unter S-Mangelbedingungen verstärkt exprimiert wird.
Interessanterweise scheint auch die Expression des PETF-Gens durch den Faktor Schwefel
beeinflusst zu werden, da unter S-Mangelbedingungen eine eindeutige Abnahme des
Transkriptes beobachtet werden konnte, während unter artifiziell aneroben Bedingungen
keine Veränderung eintrat.
Zusammenfassend erlauben die Ergebnisse der semiquantitativen RT-PCR die Aussage, dass
FDX5 das einzige unter den im Genom zu findenden Ferredoxin-Genen ist, dessen Expression
durch die Abwesenheit von Sauerstoff induziert wird.

Diskussion
51
Die Expression von FDX5 wird durch Hypoxie aktiviert.
Unter den Bedingungen der endogen etablierten Anaerobiose von belichteten S-
Mangelkulturen findet bei C. reinhardtii eine umfassende physiologische Umstrukturierung
statt, die zur Ausbildung des photofermentativen Metabolismus führt, der den Zellen ein
Überleben unter diesen eingeschränkten Bedingungen ermöglicht (MELIS et al. 2000,
HEMSCHEMEIER und HAPPE, 2005, HEMSCHEMEIER et al. 2008 a). Im Rahmen dieser
Umstrukturierung wird die Expression verschiedener Gene induziert oder auch eingestellt,
wie u.a. Real-Time PCR Experimente zeigten (JACOBS, Diplomarbeit, 2005). Zu den Genen,
die selektiv unter anaeroben Bedingungen eingeschaltet werden, gehören u.a. die Gene, die
für Schlüsselenzyme des Fermentations- und Wasserstoffmetabolismus kodieren, wie z.B. das
HYDA1- und das PFL1-Gen (HAPPE und NABER, 1993, HEMSCHEMEIER et al., 2008 a). Die
simultane Aktivierung der FDX5-, HYDA1- und PFL1-Expression lässt darauf schließen, dass
Sauerstoff der entscheidende Faktor für diese Aktivierung ist.
Promotorstudien des FDX5-Gens (persönl. Kommunikation C. Lambertz) haben gezeigt, dass
innerhalb der Promotorsequenz von FDX5 konservierte DNA-Motive der Sequenz G-T-A-C
auftreten, die auch als „copper-response elements“ (CuREs) bezeichnet werden (QUINN et al.
2000, KROPAT et al. 2005). CuRE Motive wurden bereits bei Untersuchungen der durch
Kupfermangel induzierbaren Expression des CYC6-Gens im Promotorbereich des CYC6-Gens
beschrieben. Interessanterweise vermitteln die CuRE-Motive jedoch nicht nur eine Antwort
auf Kupfermangel, sondern auch auf Sauerstoffmangel, wie Reportergenanalysen zeigten
(QUINN et al. 2000; Quinn et al., 2002). Als Regulator, der an die CuRE-Motive bindet,
konnte der spezifische Transkriptionsfaktor Crr1 identifiziert werden (KROPAT et al. 2005).
Obwohl bewiesen ist, dass Crr1 und CuRE-Motiv notwendig sind für eine hypoxische
Induktion der analysierten Gene, ist bislang noch ungeklärt, welcher Mechanismus dafür
verantwortlich ist. Durch Reportergenanalysen mit verkürzten Varianten des FDX5-Promotors
(persönl. Kommunikation C. Lambertz) konnte demonstriert werden, dass auch die CuRE-
Motive von FDX5 notwendig sind für die Aktivierung des Gens unter Anaerobiose.
Da somit FDX5 das Einzige der insgesamt sechs Ferredoxin-kodierenden Gene ist, dessen
Expression nachweislich durch Hypoxie induziert ist, wurde es für weitergehende Analysen
ausgewählt.
In photofermentierenden C. reinhardtii Zellen wird Fdx5-Protein synthetisiert.
Um anaerobe Zellen auf das Vorhandensein von Fdx5 zu untersuchen, wurde ein polyklonaler
anti-Fdx5 Antikörper hergestellt, der in Immunoblots erstmalig den Nachweis einer

Diskussion
52
Akkumulation von Fdx5 auf Proteinebene ermöglichte. Da der Antikörper nicht mit
rekombinanten PetF kreuzreagierte und auch Kreuzreaktionen mit den weiteren C. reinhardtii
Ferredoxinen ausgeschlossen werden können (persönl. Kommunikation S. Schmidt) kann
davon ausgegangen werden, dass die in den Western Blots detektierten Signale spezifisch für
Fdx5 sind. Die Aktivierung der FDX5-Expression durch Hypoxie wird durch die
durchgeführten Western Blots bestätigt, da unter aeroben Bedingungen kein Protein detektiert
werden konnte. Die parallel zur FDX5-Transkriptakkumulation eintretende Fdx5-Synthese ist
ein Hinweis darauf, dass FDX5 auf transkriptioneller Ebene reguliert wird und dass Fdx5 an
Elektronenübertragungen in den durch Hypoxie verursachten Stoffwechselwegen beteiligt ist.
Interessanterweise bleibt unter den untersuchten Bedingungen der PetF-Gehalt der Zellen
gleich, obwohl auf Transkriptebene eine eindeutige Abnahme der PETF-Expression
erkennbar ist. Vorausgesetzt, dass der PetF Antikörper nicht mit den anderen Ferredoxinen
aus C. reinhardtii kreuzreagiert, und es sich bei dem detektierten Protein daher um das PetF-
Protein handelt, kann davon ausgegangen werden, dass PetF ein sehr stabiles Protein ist. Eine
ähnliche Diskrepanz zwischen Transkriptgehalt und Proteinlevel konnte schon bei anderen
Proteinen beobachtet werden. So folgt unter Cu-Mangel die Produktion von Cyc6 und Cpx1
dem CYC6- und CPX1-Transkriptgehalt. Wird Cu jedoch wieder supplementiert, dann können
die Proteine immer noch nachgewiesen werden, wenn die respektiven mRNAs bereits wieder
vollständig abgebaut sind (HILL et al. 1991). Das gleiche Phänomen kann auch bei der durch
Hypoxie induzierten Bildung der beiden Proteine beobachtet werden (QUINN et al., 2002).
Isoliertes Fdx5-Protein reagiert sensitiv auf Sauerstoff.
Für die Aufreinigung von heterolog in E. coli produziertem Fdx5 wurde ein auf dem Strep-
Tag basierendes System gewählt, da die Isolation von Strep-Tag gekoppelten
Metalloproteinen schon beschrieben wurde und ihre Aktivität von der Aufreinigung nicht
beeinflusst wird, wie es etwa bei der Nickel-Nitrilotriessigsäure-Affinitätschromatographie
von Proteinen mit Histidin-Tag beschrieben wurde (TERPE 2003, GIRBAL et al., 2005). Der
Einsatz von E. coli als Expressionswirt begründet sich in der schon dokumentierten Nutzung
dieses Bakteriums für die heterologe Expression von [4Fe4S]-, als auch von [2Fe2S]-
Ferredoxinen (JACQUOT et al., 1997, GUERRINI et al., 2008).
Mit dem etablierten Expressionssystem konnte Fdx5 für nachfolgende Analysen in
ausreichenden Mengen aus E. coli isoliert werden. Dabei zeigte das Protein eine bräunliche
Färbung, wie sie für oxidierte Ferredoxine typisch ist (KNAFF et al., 1996). Die bräunliche
Färbung des Proteins gab einen ersten Hinweis darauf, dass das FDX5-Genprodukt einen

Diskussion
53
[FeS]-Cluster besitzt und das dieser in aktiver Form durch das Maturationssystem von E. coli
in das Apoprotein eingebaut wurde. Interessanterweise konnte bei aerob gelagerten
Proteinproben die Bildung von Oligomeren beobachtet werden. Sauerstoffsensitivität ist für
viele [FeS]-Cluster haltige Proteine bekannt (PETERING et al., 1971). Angriffspunkte des
Sauerstoffs sind dabei u. a. exponierte Fe-Atome des Clusters, die oxidiert werden und
dadurch den Redoxstatus der Clusters verändern, der aufgrund dessen instabil wird und
Eisenatome verliert. Dieser Mechanismus wurde beispielweise ausführlich am Beispiel des
Sauerstoff-abhängigen Transkriptionsfaktors FNR (Fumarate-Nitrate Reduction) aus E. coli
untersucht, der die Expression von über 120 Genen beeinflusst (SAWERS et al., 1988). Unter
anaeroben Bedingungen besitzt die FNR einen Ferredoxin-ähnlichen [4Fe4S]-Cluster. Unter
diesen Bedingungen bilden zwei FNR-Moleküle ein Dimer, welches an DNA binden kann
und so die Transkription beeinflusst. Steigt der Sauerstoffgehalt an, wird der Sauerstoff-labile
[4Fe4S]-Cluster oxidiert und wandelt sich über einen [3Fe4S] in einen [2Fe2S]-Cluster um.
Die mit dem Cluster-Wechsel einhergehende Konformationsänderung verhindert sowohl die
Dimerisierung der FNR-Monomere als auch die Bindung an DNA (KILEY und BEINERT,
1998). Auch für Ferredoxine wurde der starke Einfluss, den Sauerstoff auf den [FeS]-Cluster
haben kann, bereits beschrieben. Das Heterocysten-Ferredoxin FdxH2 aus Anabaena
variabilis besitzt unter Luftbedingungen nur eine Halbwertszeit von ca. 1 h, wie
Absorbtionsmessungen bei 422 nm gezeigt haben (SCHRAUTEMEIER et al., 1995). Analysen
von Proteinmodellen ließen dabei den Schluss zu, dass im Falle von FdxH2 eine Aminosäure,
Valin an Position 77 der Aminosäurekette, die Sauerstoffsensitivität verursacht (SINGH et al.,
2001). Obwohl bei Fdx5 die Oligomerisierung des Proteins unter Luftbedingungen beobachtet
werden kann, scheint der [2Fe2S]-Cluster durch den Sauerstoffeinfluss nicht direkt beeinflusst
zu werden, wie das unveränderte EPR-Spektrum des 24 h unter Raumluft inkubierten
Ferredoxins zeigt. Neben den für die Koordination des Clusters notwendigen Cysteine besitzt
Fdx5 jedoch noch zwei weitere Cysteine, von denen eines nahe des C-Terminus lokalisiert ist.
Es wäre möglich, dass unter Sauerstoffeinfluss die spontane Oxidation dieser Cysteine eintritt
und somit die Ausbildung von Disulfidbrücken gefördert wird. Die spontane Ausbildung von
Disulfidbrücken unter oxidativen Bedingungen ist ein bereits dokumentiertes Phänomen, dass
z.B. beim bakteriellen Transkriptionsfaktor OxyR beobachet werden konnte, der unter
Einfluss von H2O2 eine intramolekulare Disulfidbrücke ausbildet, die Einfluss auf die
Konformation des Proteins hat (BAUER et al., 1999). Es scheint jedoch auch Ferredoxine zu
geben, die funktionell als Dimer vorliegen, wie beispielsweise das [4Fe4S]-Ferredoxin aus

Diskussion
54
Pyrococcus furiosus (HASAN et al., 2002). Die molekularen Hintergründe für diese
Dimerisierung sind jedoch noch unbekannt.
Fdx5 ist ein typisches [2Fe2S]-Ferredoxin des Pflanzen-Typs mit einer außergewöhnlich
positiven Ladung.
Die Bestimmung des Eisengehaltes bewies zum einen, dass Fdx5 Eisenatome und damit einen
[FeS]-Cluster enthält, zum anderen legte das Verhältnis der Eisenatome pro Molekül die
Vermutung nahe, dass es sich bei dem enthaltenen Cluster um einen [2Fe2S]-Cluster handeln
könnte. Die Vermutung, dass Fdx5 ein klassisches [2Fe2S]-Ferredoxin ist, bestätigte sich
durch die UV/VIS- und EPR-Spektren des rekombinanten Proteins. Das Absorptionsspektrum
des oxidierten Ferredoxins zeigt die charakteristschen drei Maxima, die typisch für die Cluster
des [2Fe2S]-Typus sind und die bereits für viele [2Fe2S]-Ferredoxine dokumentiert wurden
(SCHMITTER et al., 1988; HANKE et al., 2004; KNAFF, 1996). [4Fe4S]-Cluster weisen diese
distinkten Maxima nicht auf, sie besitzen ausschließlich eine Schulter bei ca. 390 nm
(MULLINGER et al., 1975; REUBELT et al., 1991). Auch die EPR-Spektren von Fdx5 zeigen
Übereinstimmungen mit den Spektren anderer [2Fe2S]-Ferredoxine (GALVÁN und MÁRQUEZ,
1985, HALL et al. 1973). Ein weiterer Parameter, der das Vorhandensein eines [2Fe2S]-
Clusters bestätigt, ist der Abstand der Cysteine, die diesen Cluster koordinieren, da die
Positionen der Cysteine hoch konserviert sind und kaum variieren (INOUE et al., 1984). Die
meisten Pflanzen-Typ Ferredoxine weisen das Motiv Cys-X4-Cys-X2-Cys-X29-Cys auf, d.h.
sie besitzen vier AS zwischen den Cluster bindenden Cysteinen eins und zwei, zwei AS
zwischen Cystein zwei und drei und exakt 29 AS zwischen Cystein drei und vier (BERTINI et
al., 2002). Dies trifft auch auf die Cluster-koordinierenden Cysteine von Fdx5 zu. Die
Bestimmung des Redoxpotentials mittels cyclischer Voltammetrie zeigte, dass das
Redoxpotential von Fdx5 ca. 15 mV positiver ist als von PetF. Insgesamt sind die gemessenen
Werte jedoch etwas positiver als das früher berichtete Redoxpotential von PetF von -410 mV
gegen SHE (Standard Hydrogen Electrode) das durch Redoxtitration bestimmt wurde
(GALVÁN und MÁRQUEZ, 1985). Diese Variation scheint auf die unterschiedliche Methodik
zurückzuführen zu sein, die für die Bestimmung des Redoxpotentials angewendet wurde, da
solche Abweichungen bereits beschrieben wurden (VERHAGEN et al., 1995; SMITH und
FEINBERG 1990). Abgesehen von der Abweichung zum früher berichteten Potential, scheinen
die Redoxpotentiale von Fdx5 und PetF jedoch sehr ähnlich zu sein und liegen in dem
Wertebereich von -305 und -455 mV, der schon für andere Pflanzen-Typ Ferredoxine
dargestellt wurde. (CAMMACK et al., 1977). Die bislang aufgeführten Pararameter weisen also
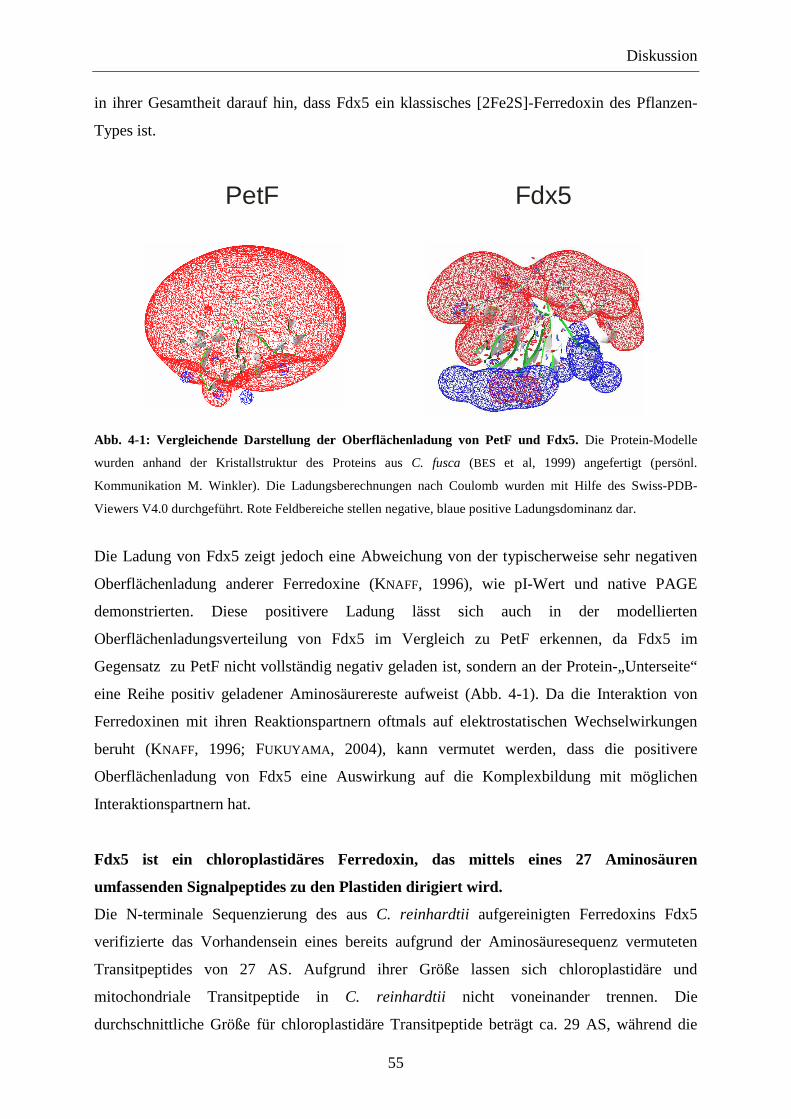
Diskussion
55
in ihrer Gesamtheit darauf hin, dass Fdx5 ein klassisches [2Fe2S]-Ferredoxin des Pflanzen-
Types ist.
PetF Fdx5
Abb. 4-1: Vergleichende Darstellung der Oberflächenladung von PetF und Fdx5. Die Protein-Modelle
wurden anhand der Kristallstruktur des Proteins aus C. fusca (BES et al, 1999) angefertigt (persönl.
Kommunikation M. Winkler). Die Ladungsberechnungen nach Coulomb wurden mit Hilfe des Swiss-PDB-
Viewers V4.0 durchgeführt. Rote Feldbereiche stellen negative, blaue positive Ladungsdominanz dar.
Die Ladung von Fdx5 zeigt jedoch eine Abweichung von der typischerweise sehr negativen
Oberflächenladung anderer Ferredoxine (KNAFF, 1996), wie pI-Wert und native PAGE
demonstrierten. Diese positivere Ladung lässt sich auch in der modellierten
Oberflächenladungsverteilung von Fdx5 im Vergleich zu PetF erkennen, da Fdx5 im
Gegensatz zu PetF nicht vollständig negativ geladen ist, sondern an der Protein-„Unterseite“
eine Reihe positiv geladener Aminosäurereste aufweist (Abb. 4-1). Da die Interaktion von
Ferredoxinen mit ihren Reaktionspartnern oftmals auf elektrostatischen Wechselwirkungen
beruht (KNAFF, 1996; FUKUYAMA , 2004), kann vermutet werden, dass die positivere
Oberflächenladung von Fdx5 eine Auswirkung auf die Komplexbildung mit möglichen
Interaktionspartnern hat.
Fdx5 ist ein chloroplastidäres Ferredoxin, das mittels eines 27 Aminosäuren
umfassenden Signalpeptides zu den Plastiden dirigiert wird.
Die N-terminale Sequenzierung des aus C. reinhardtii aufgereinigten Ferredoxins Fdx5
verifizierte das Vorhandensein eines bereits aufgrund der Aminosäuresequenz vermuteten
Transitpeptides von 27 AS. Aufgrund ihrer Größe lassen sich chloroplastidäre und
mitochondriale Transitpeptide in C. reinhardtii nicht voneinander trennen. Die
durchschnittliche Größe für chloroplastidäre Transitpeptide beträgt ca. 29 AS, während die

Diskussion
56
Länge von mitochondrialen Transitpeptiden mit ca. 30 AS angegeben wird (Fránzen et al.,
1998). Im Fall von Fdx5 wird jedoch das Transitpeptid nach der Aminosäuresequenz Val-
Gln-Ala vom Rest des Proteins abgetrennt. Die Sequenz Val-X-Ala wird häufig in der
Position -3 bis -1 der Proteaseschnittstelle von Stroma-lokalisierten Proteinen gefunden
(Fránzen et al., 1998). Dies ist z.B. der Fall bei den PSI Untereinheiten P28, P30 und P37,
welche die Sequenzen Val-Ser-Ala, Val-Arg-Ala und Val-Arg-Ala in Position -3 bis -1 zur
ihren Proteaseschnittstellen aufweisen (FRÁNZEN et al., 1998). Obwohl die Ähnlichkeit der
chloroplastidären Transitpeptide von C. reinhardtii zu den chloroplastidären Transitpeptiden
höherer Pflanzen im Allgemeinen nicht sehr groß ist – die Transitpeptide höherer Pflanzen
sind viel länger und besitzen eine andere Aminosäurekomposition (FRÁNZEN et al., 1998;
GAVEL und HEIJNE, 1990) – kann auch in höheren Pflanzen das weniger stark konservierte
Motiv (Val/Ile)-X-(Ala/Cys)↓ Ala in chloroplastidären Transitpeptiden gefunden werden
(GAVEL und HEIJNE, 1990). Das Vorkommen dieser eher gering konservierten
Konsensussequenz im N-terminalen Bereich der Proteaseschnittstelle von Fdx5 gibt einen
ersten Hinweis auf eine mögliche chloroplastidäre Lokalisation des reifen Proteins.
Die konfokale Fluoreszenzmikroskopie zur Detektion von GFP-gekoppelten Proteinen ist eine
probate Methode, um die Lokalisation von Proteinen in vivo nachzuweisen. GFP wurde
erstmalig 1962 von SHIMOMURA et al. beschrieben und hat sich seitdem zu einem der am
häufigsten genutzten Reportergene entwickelt (TSIEN, 1998). In C. reinhardtii wurde die
Nutzung von verschiedenen Reportergenen bereits beschrieben (MINKO et al., 1999,
BATEMAN und PURTON, 2000). Ein Problem dabei ist jedoch das in C. reinhardtii oftmals
beobachtete „Silencing“ von Fremdgenen (STEVENS et al., 1996; CERUTTI et al., 1997),
welches die Synthese dieser Reportergene hemmt. Deshalb wurde ein an die Codon-„Usage“
von C. reinhardtii angepasstes GFP-Gen entwickelt, dass die Nutzung dieses optimierten
GFPs als Reportergen ermöglicht (FUHRMAN et al., 1999) und welches bereits für
Lokalisationsstudien benutzt wurde (FUHRMAN et al., 1999, GLANZ et al. 2006; WINKLER,
Dissertation, 2009). Die Fdx5-Lokalisatonsstudien dieser Arbeit, die ebenfalls die optimierte
GFP-Sequenz als Reportergen nutzen, weisen auf eine chloroplastidäre Lokalisation des
Fdx5-Polypeptides hin. Die GFP-Fluoreszenz des Fdx5-Transitpeptid/ GFP-Fusionsproteins
co-lokalisiert mit der Chloroplasten-Fluoreszenz und weist damit Übereinstimmungen zu den
Lokalisationsstudien des chloroplastidär lokalisierten Gruppe II Intron RNA-Bindeproteins
NAPL (GLANZ et al. 2006) und der chloroplastidär lokalisten Proteine HydA1 und HydA2
(WINKLER, Dissertation, 2009) auf. Die beobachteten Akkumulationen von GFP-Fluoreszenz,
die am Rande des Chloroplasten lokalisieren, konnten in dieser Art auch bei den

Diskussion
57
Lokalisaktionsstudien von HydA1 und HydA2 beobachtet werden (WINKLER, Dissertation,
2009).
Der dritte Hinweis auf eine chloroplastidäre Lokalisation von Fdx5 konnte durch Immuno
Blot Analysen mit isolierten Mitochondrien und Chloroplasten ermittelt werden, in denen
Fdx5 das gleiche Bandenmuster wie die Proteine HydA1 und RbcL aufwies, von denen die
plastidäre Lokalisation bereits beschrieben wurde (HAPPE et al, 1994; HARTMANN und
HARPEL, 1994). Dabei war die Reinheit der erhaltenen Chloroplasten nicht absolut, es konnte
eine Verunreinigung mit mitochondrialen Proteinen detektiert werden. Diese
Verunreinigungen sind jedoch oftmals nicht zu vermeiden und wurden auch in anderen
Veröffentlichungen beschrieben (ATTEIA et al, 2006). Zusammenfassend kann aufgrund
dieser Ergebnisse der Schluss gezogen werden, dass Fdx5 mittels eines 27 AS großen
Transitpeptides in den Chloroplasten von C. reinhardtii dirigiert wird.
Fdx5 weist funktionelle Unterschiede zu PetF auf, die sich in den Primärsequenzen
beider Ferredoxine wiederspiegeln. Fdx5 besitzt signifikante Abweichungen in den
konservierten Domänen von Pflanzen-Typ Ferredoxinen, die relevant sind für die
elektrostatische Wechselwirkung des Ferredoxins mit seinen Interaktionpartnern.
Da Fdx5 als ein klassisches [2Fe2S]-Ferredoxin des Pflanzen-Types identifiziert werden
konnte, das in Chloroplasten von C. reinhardtii lokalisiert ist, sollte seine Funktionalität in
typischen Elektronentransferreaktionen getestet werden. Aufgrund der durch Anaerobiose
induzierten Expression von FDX5 wurde untersucht, ob eine Interaktion mit der ebenfalls
unter anaeroben Bedingungen gebildeten Hydrogenase HydA1, die nachweislich von PetF
reduziert werden kann, möglich ist (HAPPE und NABER, 1993, WINKLER, Dissertation, 2009).
In in vitro Hydrogenasetests, in denen reduziertes Fdx5 eingesetzt wurde, konnte im
Gegensatz zu PetF jedoch keine Wasserstoffbildung nachgewiesen werden. Die
Komplexbildung von HydA1 und PetF aus C. reinhardtii beruht auf der Beteiligung von
elektrostatischen Kräften, wie die Analyse der Elektronentransferwechselwirkung zwischen
beiden Proteinen in Abhängigkeit von der Elektrolytkonzentration des Reaktionspuffers zeigte
(WINKLER, Dissertation, 2009). Die insgesamt positivere Ladung von Fdx5 gegenüber PetF
könnte deshalb einen entscheidenden Einfluss auf die Bildung eines elektrostatischen
Komplexes mit HydA1 haben und eine Interaktion zwischen beiden Proteinen verhindern.
Durch Analysen mit PetF Punktmutanten konnten spezifische AS von PetF, die an der
Interaktion mit HydA1 beteiligt sind, identifiziert werden (WINKLER, Dissertation, 2009).
Beim Vergleich der Aminosäuresequenzen von PetF und Fdx5 ist ersichtlich, dass die für eine
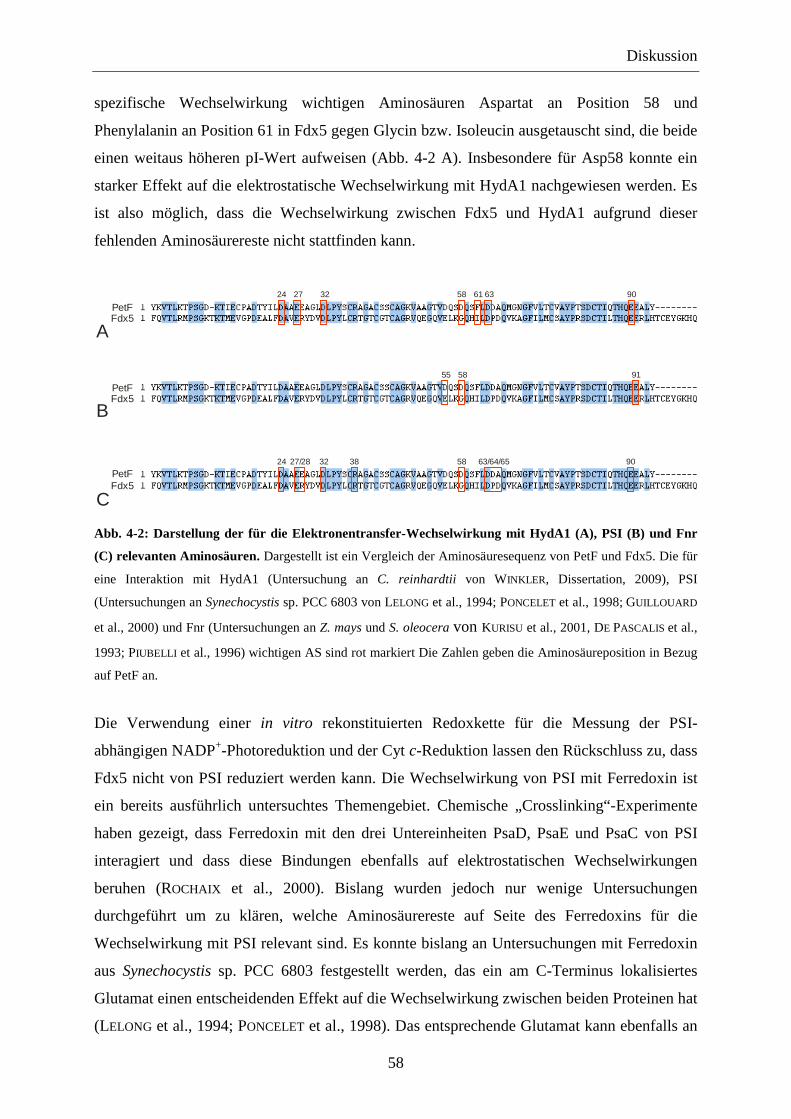
Diskussion
58
spezifische Wechselwirkung wichtigen Aminosäuren Aspartat an Position 58 und
Phenylalanin an Position 61 in Fdx5 gegen Glycin bzw. Isoleucin ausgetauscht sind, die beide
einen weitaus höheren pI-Wert aufweisen (Abb. 4-2 A). Insbesondere für Asp58 konnte ein
starker Effekt auf die elektrostatische Wechselwirkung mit HydA1 nachgewiesen werden. Es
ist also möglich, dass die Wechselwirkung zwischen Fdx5 und HydA1 aufgrund dieser
fehlenden Aminosäurereste nicht stattfinden kann.
24 27 32 58 61 63 90
9155 58
58 9063/64/653827/2824 32
PetF
PetF
PetF
Fdx5
Fdx5
Fdx5
A
B
C
Abb. 4-2: Darstellung der für die Elektronentransfer-Wechselwirkung mit HydA1 (A), PSI (B) und Fnr
(C) relevanten Aminosäuren. Dargestellt ist ein Vergleich der Aminosäuresequenz von PetF und Fdx5. Die für
eine Interaktion mit HydA1 (Untersuchung an C. reinhardtii von WINKLER, Dissertation, 2009), PSI
(Untersuchungen an Synechocystis sp. PCC 6803 von LELONG et al., 1994; PONCELET et al., 1998; GUILLOUARD
et al., 2000) und Fnr (Untersuchungen an Z. mays und S. oleocera von KURISU et al., 2001, DE PASCALIS et al.,
1993; PIUBELLI et al., 1996) wichtigen AS sind rot markiert Die Zahlen geben die Aminosäureposition in Bezug
auf PetF an.
Die Verwendung einer in vitro rekonstituierten Redoxkette für die Messung der PSI-
abhängigen NADP+-Photoreduktion und der Cyt c-Reduktion lassen den Rückschluss zu, dass
Fdx5 nicht von PSI reduziert werden kann. Die Wechselwirkung von PSI mit Ferredoxin ist
ein bereits ausführlich untersuchtes Themengebiet. Chemische „Crosslinking“-Experimente
haben gezeigt, dass Ferredoxin mit den drei Untereinheiten PsaD, PsaE und PsaC von PSI
interagiert und dass diese Bindungen ebenfalls auf elektrostatischen Wechselwirkungen
beruhen (ROCHAIX et al., 2000). Bislang wurden jedoch nur wenige Untersuchungen
durchgeführt um zu klären, welche Aminosäurereste auf Seite des Ferredoxins für die
Wechselwirkung mit PSI relevant sind. Es konnte bislang an Untersuchungen mit Ferredoxin
aus Synechocystis sp. PCC 6803 festgestellt werden, das ein am C-Terminus lokalisiertes
Glutamat einen entscheidenden Effekt auf die Wechselwirkung zwischen beiden Proteinen hat
(LELONG et al., 1994; PONCELET et al., 1998). Das entsprechende Glutamat kann ebenfalls an

Diskussion
59
der äquvivalenten Position in der Aminosäuresequenz von Fdx5 gefunden werden (Abb. 4-2
B). Es wurden jedoch noch zwei weitere Aminosäuren in Synechocystis sp. PCC 6803 PetF
identifiziert, die die Komplexbildung mit PSI stabilisieren. Der Austausch der beiden
Aspartatreste, die in C. reinhardtii PetF Asp 55, bzw. Asp 58 entsprechen, gegen basische
Aminosäuren, führt zu einer 14- bis 19-fachen Abnahme der Affinität (GUILLOUARD et al.,
2000). In Fdx5 sind diese Aspartatereste nicht vorhanden, an ihre Stelle treten Glutamat, bzw.
Glycin, wobei vor allem Glycin als ungeladene Aminosäure einen viel höheren pI-Wert als
das negativ geladene Aspart aufweist.
Einen weiteren Effekt auf die Wechselwirkung von Fdx5 und PSI könnte auch die C-
terminale Extension von Fdx5 hervorufen. Fdx5 weist eine, im Vergleich zu den meisten
Pflanzen-Typ Ferredoxinen, die durchschnittlich 97 bis 99 AS lang sind (BERTINI et al.,
2002), außergewöhnliche Größe von 103 AS auf (siehe auch Abb. 4-3 A). Gegenüber PetF ist
der C-Terminus von Fdx5 um die Aminosäuren Thr-Cys-Glu-Tyr-Gly-Lys-His-Gln
verlängert. An Synechocystis sp. PCC 6803 PetF konnte gezeigt werden, dass eine
Verlängerung des C-Terminus um sieben AS zu einer 10-fachen Abnahme der Affinität zu
PSI führt (PONCELET et al., 1998).
Im Gegensatz zu der Wechselwirkung mit PSI ist die Interaktion zwischen Fdx5 und Fnr
möglich, wie die Reduktion von Cyt c in einer in vitro rekonstituierten
Elektronentransportkette zeigte. Röntgenstrukturanalysen des Komplexes von Z. mays Fnr
und Ferredoxin haben veranschaulicht, dass Aminosäuren Glu 29, Arg 40, Asp 61, Asp 65
und Asp 66 auf Seiten des Ferredoxins für die Ausbildung intermolekularer Salzbrücken mit
der Fnr relevant sind (KURISU et al., 2001). Diese Aminosäuren können in äquivalenten
Positionen auch in C. reinhardtii PetF gefunden werden, in Fdx5 sind jedoch an den zu Asp
61 und Asp 66 entsprechenden Positionen Glycin (Position 58) und Prolin (Position 64) zu
finden (Abb. 4-2 C). Daneben scheinen aber auch Asp 26, Glu 30, Asp 34, Asp 60 und Asp 67
in S. oleocera Ferredoxin für die Interaktion mit Fnr von Bedeutung zu sein (DE PASCALIS et
al., 1993). Bis auf Asp 67, an dessen Stelle ein Glycin tritt, sind diese AS an
korrespondierenden Positionen in PetF zu finden. In Fdx5 ist hingegen Glu 30 gegen ein
Arginin ausgetaucht. Ausserdem scheint auch Glu 92 von S. oleocera Ferredoxin einen Effekt
auf die Wechselwirkung mit der Fnr zu haben (PIUBELLI et al., 1996). Das zu dieser Position
äquvivalente Glu (Position 90) kann sowohl in C. reinhardtii PetF als auch in Fdx5 gefunden
werden.

Diskussion
60
Obwohl somit einige der vorwiegend negativ geladenen Aminosäuren, die für eine Interaktion
mit der Fnr relevant zu sein scheinen, in Fdx5 durch neutrale oder positiv geladene
Aminosäuren ausgetauscht sind, scheinen diese die Wechselwirkung mit der Fnr im Vergleich
zur Reaktion der Fnr mit PetF nicht zu hemmen. Genauere Aussagen zur Interaktion zwischen
Fdx5 und Fnr können jedoch nicht gemacht werden, da in der für die Analyse verwendeten in
vitro Redoxkette auch der Elektronentransfer zwischen Ferredoxin und Cytc einen
massgeblichen Einfluss auf die Messwerte hat und die Wechselwirkung zwischen Fdx5 und
Cytc bzw. PetF und Cytc unterschiedliche kinetische Eigenschaften aufweisen könnte.
Eine Schlussfolgerung, die aus den Analysen der Wechselwirkung von Fdx5 mit typischen
Reaktionspartnern gezogen werden kann, ist, dass die signifikanten Unterschiede in den
Reaktionen insbesondere mit HydA1 und PSI, die bei Fdx5 im Vergleich zu PetF zu
beobachten waren, auf das Auftreten abweichender Aminosäurereste zurückzuführen sind.
Die meisten Ferredoxine besitzen konservierte Domänen (XU et al., 2006), die vor allem
durch das Auftreten negativ geladener Aminosäurereste charakterisiert sind, und die in der
Sequenz von Fdx5 zum Teil deutlich vom Konsensus abweichen. Die entprechenden Bereiche
sind in Abb. 4-3 A anhand eines Sequenzvergleichs verschiedener Pflanzen-Typ Ferredoxine
in Bezug auf C. reinhardtii PetF dargestellt. Es stellt sich die Frage, ob diese
Sequenzabweichungen auch einen Einfluss auf die Wechselwirkung von Fdx5 mit anderen
typischen Ferredoxin-Redoxpartnern, wie z.B der Ferredoxin/ Thioredoxin-Reduktase oder
der Nitrit-Reduktase haben könnten, die auch mit diesen Domänen interagieren (Xu et al.,
2006, JAQUOT et al., 1997, GARCIA-SÁNCHEZ et al., 2000) oder ob Fdx5 eventuell ein
spezialisiertes Ferredoxin ist, dass eine ganz spezifische Funktion im Metabolismus von. C
reinhardtii erfüllt.
Die ungewöhnliche Aminosäuresequenz von Fdx5 stellt sich auch im Homologievergleich zu
verschiedenen anderen Pflanzen-Typ Ferredoxinen dar (Abb. 4-3 A). “BLAST”-Analysen in
der Datenbank von NCBI zeigen, dass die Übereinstimmung von Fdx5 zu PetF nur 50 %
beträgt. Generell weist Fdx5 die größten Homologien zu cyanobakteriellen Ferredoxinen auf.
Das Phylogramm in Abb. 4.3 B, in dem sich der Verwandschaftsgrad von Pflanzen-Typ
Ferredoxinen aus unterschiedlichen Organismengruppen darstellt, verdeutlicht die
Sonderstellung, die Fdx5 im Vergleich zu den anderen Ferredoxinen einnimmt. Während sich
die Ferredoxine der Grünalgen, der Rotalgen, der Cyanobakterien und der höheren Pflanzen
jeweils in Gruppen zusammenfinden, steht Fdx5 weit ausserhalb dieser Gruppen, was den
geringen Verwandschaftsgrad zu den anderen Ferredoxinen verdeutlicht.
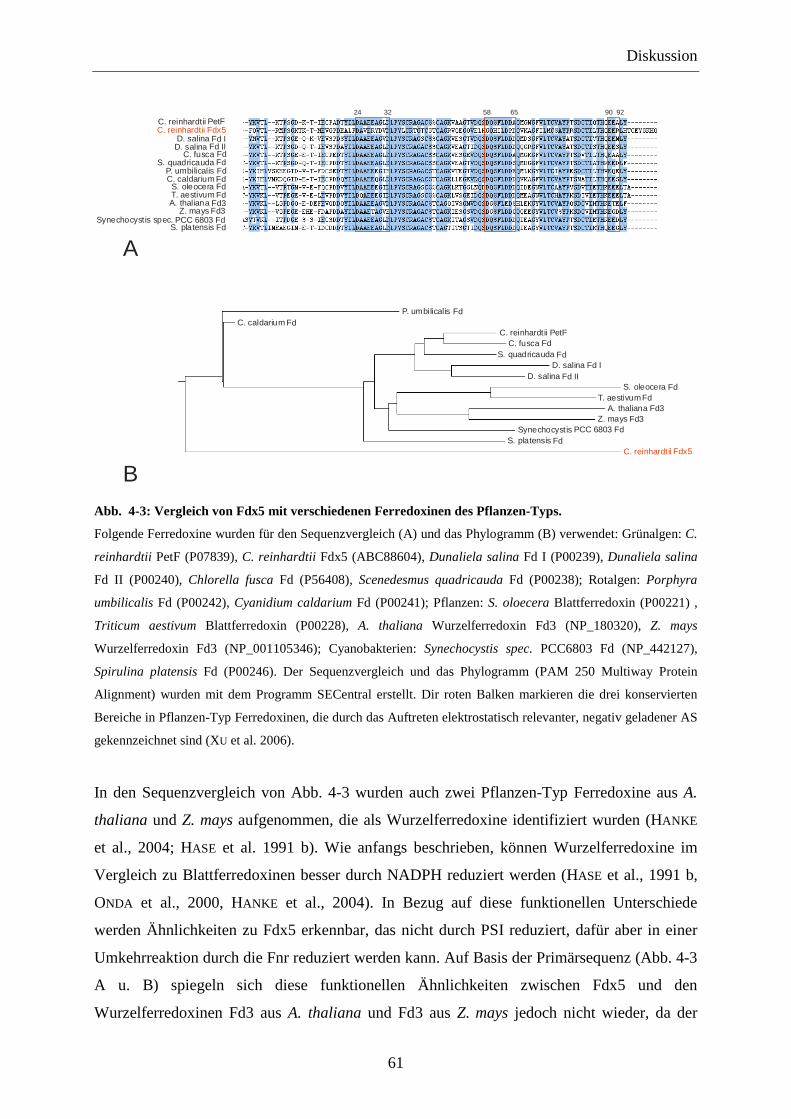
Diskussion
61
C. reinhardtii PetFC. reinhardtii Fdx5
D. salina Fd ID. salina Fd II
C. fusca FdS. quadricauda Fd
P. umbilicalis FdC. caldarium FdS. oleocera FdT. aestivum Fd
Synechocystis spec. PCC 6803 FdS. platensis Fd
A. thaliana Fd3Z. mays Fd3
C. reinhardtii PetF
C. reinhardtii Fdx5
D. salina Fd ID. salina Fd II
C. fusca FdS. quadricauda Fd
P. umbilicalis FdC. caldarium Fd
S. oleocera FdT. aestivum Fd
Synechocystis PCC 6803 FdS. platensis Fd
A. thaliana Fd3Z. mays Fd3
A
B
24 5832 65 90 92
Abb. 4-3: Vergleich von Fdx5 mit verschiedenen Ferredoxinen des Pflanzen-Typs.
Folgende Ferredoxine wurden für den Sequenzvergleich (A) und das Phylogramm (B) verwendet: Grünalgen: C.
reinhardtii PetF (P07839), C. reinhardtii Fdx5 (ABC88604), Dunaliela salina Fd I (P00239), Dunaliela salina
Fd II (P00240), Chlorella fusca Fd (P56408), Scenedesmus quadricauda Fd (P00238); Rotalgen: Porphyra
umbilicalis Fd (P00242), Cyanidium caldarium Fd (P00241); Pflanzen: S. oloecera Blattferredoxin (P00221) ,
Triticum aestivum Blattferredoxin (P00228), A. thaliana Wurzelferredoxin Fd3 (NP_180320), Z. mays
Wurzelferredoxin Fd3 (NP_001105346); Cyanobakterien: Synechocystis spec. PCC6803 Fd (NP_442127),
Spirulina platensis Fd (P00246). Der Sequenzvergleich und das Phylogramm (PAM 250 Multiway Protein
Alignment) wurden mit dem Programm SECentral erstellt. Dir roten Balken markieren die drei konservierten
Bereiche in Pflanzen-Typ Ferredoxinen, die durch das Auftreten elektrostatisch relevanter, negativ geladener AS
gekennzeichnet sind (XU et al. 2006).
In den Sequenzvergleich von Abb. 4-3 wurden auch zwei Pflanzen-Typ Ferredoxine aus A.
thaliana und Z. mays aufgenommen, die als Wurzelferredoxine identifiziert wurden (HANKE
et al., 2004; HASE et al. 1991 b). Wie anfangs beschrieben, können Wurzelferredoxine im
Vergleich zu Blattferredoxinen besser durch NADPH reduziert werden (HASE et al., 1991 b,
ONDA et al., 2000, HANKE et al., 2004). In Bezug auf diese funktionellen Unterschiede
werden Ähnlichkeiten zu Fdx5 erkennbar, das nicht durch PSI reduziert, dafür aber in einer
Umkehrreaktion durch die Fnr reduziert werden kann. Auf Basis der Primärsequenz (Abb. 4-3
A u. B) spiegeln sich diese funktionellen Ähnlichkeiten zwischen Fdx5 und den
Wurzelferredoxinen Fd3 aus A. thaliana und Fd3 aus Z. mays jedoch nicht wieder, da der

Diskussion
62
Verwandschaftsgrad von Fdx5 zu diesen Wurzelferredoxinen ebenso gering erscheint wie zu
typischen Blattferredoxinen.
C. reinhardtii Zellen mit einem reduzierten FDX5-Transkriptgehalt weisen unter Cu-
Mangel einen verringerten Chlorophyllgehalt und eine verringerte photosynthetische
Aktivität auf. Dies weist auf eine Funktionalität von Fdx5 in den unter Cu-Mangel
eintretenden metabolischen Adaptionen von C. reinhardtii hin.
Da die Ferredoxin-abhängigen Aktivitätstests nur einen ersten Eindruck von der
Funktionalität des Fdx5-Proteins vermitteln konnten, wurde als weitere Strategie für die
Aufklärung des Fdx5 abhängigen Stoffwechsels die Ausschaltung des Fdx5-Gens angestrebt.
Dafür wurde eine erst kürzliche vorgestellte Methode des Gen-„Silencing“ durch artifizielle
miRNAs verwendet (MOLNÁR et al., 2009). miRNAs gehören neben siRNAs zur Klasse der
kleinen RNAs, die eine Funktion im „Silencing“-Mechanismus von Zellen haben (BARTEL,
2004). Auch in C. reinhardtii wurde vor kurzem die Existenz von miRNAs entdeckt
(MOLNÁR et al., 2007). Sie werden aus längeren Vorläufer-miRNAs gebildet, die nicht perfekt
gepaarte, doppelsträngige Bereiche ausbilden. Die Prozessierung dieser Vorläufer-miRNAs
verläuft ähnlich, wie für siRNAs beschrieben. Durch das Polymerase III-ähnliche Enzym
Dicer wird aus der Vorläufer-miRNA ein 21-24 nt langes doppelsträngiges RNA-Molekül
freigesetzt (AMBROS, 2001), das daraufhin in den Multienzymkomplex RISC („RNA induced
silencing complex“) aufgenommen wird und an ein Argonaut-Protein bindet. Der Strang mit
der höheren thermodynamischen Stabilität am 5’-Ende, d.h. der Strang mit der stabileren 5’-
Basenpaarung, verbleibt im RISC, während der Passagierstrang, auch als miRNA*
bezeichnet, abgebaut wird (BARTEL, 2004). Der Argonaut-miRNA-Komplex bindet mittels
Watson-Crick-Basenpaarung an komplementäre Sequenzen der Ziel-mRNA und initiiert
dadurch den eigentlichen „Silencing“-Mechanimus. In Abhängigkeit von der
Komplementarität der miRNA zur mRNA wird dann entweder die Degradation der mRNA
oder eine Arretierung der Translation induziert (BARTEL, 2004). Wie in Kapitel 3.9
beschrieben wurde, werden durch die Nutzung von artifiziellen miRNAs als Werkzeug zum
gezielten Ausschalten von Genen die Probleme der RNAi-Methodik umgangen, da miRNAs
in der Regel nicht Ziel des zelleigenen Silencingmechanismus sind (MOLNÁR et al., 2009). In
Pflanzen wurde die Methodik der amiRNAs bereits für das effektive Ausschalten von Genen
beschrieben. So konnten in A. thaliana, Solanum lycopersicum und Nicotiana tabacum schon
verschiedene Gene durch den Einsatz entsprechender amiRNAs ausgeschaltet werden
(SCHWAB et al., 2006, ALVAREZ et al., 2006). Mittels der auch in dieser Arbeit verwendeten

Diskussion
63
Methodik konnten auch in C. reinhardtii bereits verschiedene Gene, z.B. COX90
(Untereinheit der Cytochrom c Oxidase), PSY (Phytoene Synthase), DCL1 (Dicer-ähnliche
Nuklease), sowie MAA7 (Tryptophan Synthase β-UE) und RBCS1/2 (kleine UE der Rubisco)
herunterreguliert werden (MOLNÁR et al, 2009, ZHAO et al., 2008). Dabei zeigten bis zu 72 %
der transgenen C. reinhardtii Linien einen Phänotyp, der auf die Expression der amiRNAs
zurückzuführen war (MOLNÁR et al., 2009). Bei den in dieser Arbeit hergestellten transgenen
C. reinhardtii Transformanden konnte ebenfalls ein hoher Anteil an Zelllinien identifiziert
werden, die unter artifiziell anaeroben Bedingungen einen verringerten FDX5-
Transkriptgehalt aufwiesen. Aus Zeitgründen war eine Untersuchung des Fdx5-
Proteingehaltes unter anaerob artifiziellen Bedingungen nicht mehr möglich. Da ein niedriger
FDX5-Transkriptgehalt jedoch auf einen niedrigen Fdx5-Proteingehalt schließen lässt, wurden
die zwei Stämme mit dem niedrigsten FDX5-Transkriptgehalt für weitere Untersuchungen
ausgewählt.
Dabei wurde zunächst ein Schwerpunkt auf die Analyse des Photofermentationsmetabolismus
gelegt. Die Beteiligung von Fdx5 an der Reduktion der Hydrogenase HydA1 mittels
Elektronen, die aus der photosynthetischen Elektronentransportkette stammen, kann bereits
ausgeschlossen werden, da NADP+-Photoreduktion und in vitro Analysen mit HydA1 gezeigt
haben, dass Fdx5 nicht von PSI reduziert wird und nicht mit HydA1 interagieren kann. Die
gemessene Wasserstoffakkumulation in der Gasphase der S-Mangel induzierten C. reinhardtii
amiRNA-Transformanden im Vergleich zum Wildtyp bestätigt die Vermutung, dass Fdx5
keine Funktion im Elektronentransfer zwischen PSI und HydA1 besitzt.
Auch die Formiat- und Ethanolproduktion der S-Mangel induzierten sowie artifiziell
anaerobisierten C. reinhardtii Stämme zeigte keine signifikanten Unterschiede zum Wildtyp.
Die Produktion von Formiat und Ethanol ist ein Hinweis auf die Aktivität der Pfl1-Reaktion
(HEMSCHEMEIER et al., 2008 a) Wie schon in der Einleitung beschrieben, gehört die Pfl1 zur
Klasse der sogenannten Glycylradikal-Enzyme und wird durch die Pfl-Aktivase, einem
Radikal-SAM-Protein, posttranslational aktiviert (FREY et al., 1994; HEMSCHEMEIER et al,
2008 a). Die Aktivität der Pfl-Aktivase ist dabei abhängig von SAM, Pyruvat und einem
externen Ein-Elektronendonator wie Flavodoxin (BLASCHKOWSKI et al., 1982). Da in C.
reinhardtii bislang noch keine Flavodoxine identifiziert werden konnten, besteht die
Möglichkeit, dass der benötigte Ein-Elektronenüberträger ein Ferredoxin ist. Wäre Fdx5 der
Ein-Elektronenübertrager, der für die Aktivierung der Pfl1 notwendig ist, müsste ein

Diskussion
64
verringerter Fdx5-Gehalt einen Einfluss auf den Gehalt an aktiver Pfl1 haben und damit auch
einen Einfluss auf die Produktion von Formiat und Ethanol.
Ein weiterer Ferredoxin-abhängiger Stoffwechselweg, in den Fdx5 involviert sein könnte, ist
die oxidative Decarboxylierung von Pyruvat zu Acetyl-CoA durch die Pfo. Die Expression
des PFO-Gens wird parallel zur Expression der Gene PFL und HYDA1 induziert, wie durch
semiquantitative RT-PCR gezeigt werden konnte. In Wasserstoff-produzierenden Bakterien
ist die Reduktion von Ferredoxin durch die Pfo die Elektronenquelle für verschiedene
Hydrogenasen (PIEULLE et al., 1997; STAL und MOEZELAAR, 1997). Bislang ist noch nicht
geklärt, ob durch die Pfo reduziertes Ferredoxin auch in C. reinhardtii ein Elektronendonor
für HydA1 sein könnte (HEMSCHEMEIER et al., 2008 a). Ebenfalls ungeklärt ist die Frage, ob
durch die Pfo produziertes Acetyl-CoA durch die AdhE zu Ethanol umgesetzt wird
(HEMSCHEMEIER et al, 2008 a). Eine weitere nennenswerte Funktion besitzt die Pfo von E.
coli, die vermutlich reduziertes Flavodoxin für die Aktivierung der Pfl durch die PflA
bereitstellt (BLASCHKOWSKI et al., 1982). Wäre Fdx5 ein Redoxpartner der Pfo, müsste sich
eine verringerte Fdx5-Konzentration, demnach eventuell in einer verringerten Ethanol- oder
auch Wasserstoffproduktion der S-Mangel induzierten oder artifiziell anaerobisierten
amiRNA-Transformanden zeigen. Eine solche Reaktion konnte jedoch nicht beobachtet
werden.
Zusammengefasst lassen die Untersuchungen an S-Mangel induzierten und artifiziell
anaerobisierten C. reinhardtii-Stämmen mit einem verringerten FDX5-Transkriptgehalt keine
Unterschiede in Wasserstoff-, Formiat- und Ethanolproduktion erkennen, so dass eine
Funktionalität von Fdx5 in diesen Stoffwechselwegen zunächst auszuschließen ist. Es könnte
jedoch auch sein, dass C. reinhardtii in der Lage ist, an einen geringen Fdx5-Gehalt zu
adaptieren und dass andere Elektronenüberträger die Funktion von Fdx5 übernehmen und
aufgrund dessen keine Unterschiede beobachtet werden können. Gleichwohl könnte auch der
Fdx5-Gehalt der Zellen trotz eines verringerten Transkriptgehaltes unter artifiziell anaeroben
Bedingungen unverändert sein. Western Blot Analysen geben einen ersten Hinweis darauf,
dass zumindest unter S-Mangel der Fdx5-Gehalt der amiRNA-Transformanden
gleichbleibend ist. Ein Grund hiefür könnte der artifizielle HSP70A/RBCS2-Promotor sein,
der die Expression der amiRNAs antreibt. Es ist bekannt, dass unter S-Mangel eine
Degradation der Rubisco eintritt (Zhang et al., 2002). Auf Transkriptebene kann eine
Abnahme des mRNA Levels des RBCS-Gens beobachtet werden (ZHANG et al., 2004,
HEMSCHEMEIER, Diplomarbeit, 2002). Es ist also nicht auszuschließen, dass gerade unter S-

Diskussion
65
Mangel die Aktivität des RBCS-Promotors inhibiert ist und deshalb die Expression der
amiRNAs nicht stattfindet. Aus zeitlichen Gründen war eine Untersuchung des FDX5-
Transkriptgehaltes der amiRNA-Transformanden unter S-Mangel nicht mehr möglich. Unter
artifiziellen anaeroben Bedingungen hingegen scheint die Aktivität des Promotors nicht
beeinflusst, wie das „Screening“ nach amiRNA-Transformanden mit verringertem FDX5-
Transkriptgehalt gezeigt hat.
Parallel zu dieser Arbeit durchgeführte Analysen des Transkriptoms von Cu-defizienten C.
reinhardtii Kulturen haben gezeigt, dass auch unter Cu-Mangel die Expression des FDX5-
Gens aktiviert wird und das FDX5 überdies die zweitstärkste Transkriptakkumulation unter
diesen Bedingungen zeigt (S. Merchant, persönl. Mitteilung). Wie schon eingangs bei der
Analyse der FDX5-Expression beschrieben, können in der Promotorsequenz des FDX5-Gens
die sogenannten CuRE-Motive gefunden werden, die eine Antwort auf Cu- als auch auf
Sauerstoffmangel vermitteln (QUINN et al. 2000; Quinn et al., 2002). So wird z.B. auch die
Expression des CYC6-Gens und des CPX1-Gens nicht nur unter Cu-Mangel, sondern auch
unter anaeroben Bedingungen aktiviert (QUINN et al. 2000; QUINN et al., 2002). Die
Gemeinsamkeiten, die hinter den Cu- und Sauerstoffmangel Antworten stehen, sind bislang
noch nicht geklärt, ein evolutionärer Zusammenhang zwischen Cu- und Sauerstoffmangel
Antwort kann jedoch nicht ausgeschlossen werden (QUINN et al., 2002), da Kupfer-Ionen
unter anaeroben Bdingungen extrem unlösliche und somit nicht bioverfügbare Cu(I)-Sulfide
bilden (ÖSTERBERG, 1974). Somit ist zumindest nicht auszuschließen, dass Fdx5 auch eine
Funktion im Cu-Mangel Metabolismus von C. reinhardtii besitzten könnte.
Die in C. reinhardtii am häufigsten vorkommenden Cu-haltigen Enzyme sind die Cytochrom
Oxidase der Mitochondrien (FERGUSON-M ILLER und BABCOCK, 1996) das Plastocyanin der
Chloroplasten (MERCHANT et al., 1991) und eine Multi-Kupfer Oxidase (HERBIK et al., 2002;
CHEN et al., 2008), die an der Assimilation von Eisen beteiligt ist. Neben diesen Enzymen
gibt es noch weitere kupferhaltige Proteine, wie z.B. Metalloproteinasen, Aminoxidasen oder
Ascorbat-abhängige Monooxygenasen (HEITZER und HALLMANN 2002; BUFFONI und IGNESTI
2000; MERCHANT et al., 2006) deren Anteil am intrazellulären Cu- Bedarf jedoch
wahrscheinlich nicht so hoch ist (MERCHANT et al., 2006). Kupfer ist folglich ein essentieller
Nährstoff für die Zelle, dessen Mangel zu einer weitreichenden Umstrukturierung des
Metabolismus führt. Diese Umstrukturierung führt zum einen zu einer erhöhten Cu-
Assimilation zum anderen zur Synthese von sogenannten „Back up“-Enzymen, welche die
Cu-haltigen Proteine unter Cu-Mangel ersetzen (MERCHANT et al., 2006).

Diskussion
66
Ein Beispiel für eines dieser Cu-freien „Back up“ Enzyme ist das Häm-enthaltende Cyt c6, das
in Cu-supplementierten Zellen nicht nachgewiesen werden kann und dessen Synthese durch
Cu-Mangel induziert wird (MERCHANT und BORGORAD, 1987). Cyt c6 ersetzt unter Cu-
Mangel das Plastocyanin der photosynthetischen Elektronentransportkette und übernimmt
seine Elektronentransferfunktion in der Übertragung von Elektronen vom Cytochtom-b6f-
Komplex zu PSI. Ein weiteres „Back up“ Enzym wird wahrscheinlich durch das Gen CRD2
kodiert. Wie bereits erwähnt, ist eine an der Eisen-Assimilation beteiligte Multi-Kupfer
Oxidase ein in C. reinhardtii häufig vorkommendes Protein. Unter Cu-Mangel können jedoch
keine Symptome beobachtet werden, die auf Eisenmangel zurückzuführen wären, was im
Umkehrschluss vermuten lässt, dass die Eisenaufnahme in Cu-freien Medium durch ein Cu-
freies „Ersatz“-Enzym aufrechterhalten wird (LA FONTAINE et al., 2002). Dies wurde an
Untersuchungen einer CRD2-Mutante bestätigt, die unter Cu-Mangel einen verringerten
Chlorophyllgehalt und ein verringertes Wachstum besitzt (ERIKSSON et al., 2004). Bislang ist
das Genprodukt von CRD2 jedoch noch unbekannt (MERCHANT et al., 2006). Zu den weiteren
Genen, die unter Cu-Mangel hochreguliert werden, gehören das Gen CPX1, das für die
Coprophoryrinogen III Oxidase und das Gen CRD1, das für eine Isoformen der aeroben
oxidativen Cyclase (Mg-Protoporphyrin-IX-Monomethyl-Ester Cyclase) kodiert.
Coprophoryrinogen III Oxidase und oxidative Cyclase sind Enzyme, die eine Funktion in der
Tetrapyrrol-Biosynthese der Zellen besitzen. Coprophoryrinogen III Oxidase katalysiert dabei
den fünften Schritt der Tetrapyrrol-Biosynthese, die Umsetzung von Coproporphyrinogen III
zu Protoporphyrinogen III, während die oxidative Cyclase an der Sauerstoff-abhängigen
Bildung von Protochlorophyllid beteiligt ist (TIMKO , 1998). Die metabolischen Hintergründe,
die eine Hochregulation von CPX1 und CRD1 notwendig machen, sind jedoch noch ungeklärt
(MERCHANT et al., 2006).
Gen Protein Metabolismus
CYC6 Cytochrom c6 (ersetzt Plastocyanin) Photosynthese
CPXI Coproporphyrinogen III Oxidase Tetrapyrrol-Biosynthese
CRD1 Oxidative Cyclase Tetrapyrrol-Biosynthese
CRD2 ? (ersetzt vermutlich eine Multi-Kupfer Oxidase) Eisenassimilation
FDX5 Ferredoxin Fdx5 ?
Tab. 4-1: Übersicht über Proteine, die unter Cu-Mangel akkumulieren. Unter Cu-Mangel wird die Synthese
diverser Proteine aktiviert, die an unterschiedlichen Stoffwechselwegen beteiligt sind. Einige dieser Proteine
fungieren als „Backup“-Enzyme für kupferhaltige Proteine, die nicht mehr gebildet werden können (MERCHANT
und BORGORAD, 1987; QUINN und MERCHANT, 1998; ERIKSSON et al., 2004, MERCHANT et al., 2006).

Diskussion
67
Interessanterweise konnte bei Untersuchungen der FDX5 amiRNA Transformanden unter Cu-
Mangel ein verringerter Chlorophyll- und Carotinoidgehalt der Transformanden im Vergleich
zum Wildtyp beobachtet werden. Dieser Unterschied zeigte sich sowohl unter
photoheterotrophen als auch unter photoautotrophen Bedingungen. Die Photosyntheseraten
der amiRNA-Transformanden sind gleichermaßen verringert. Western Blot Analysen gegen
Cyc6, von dem die auschließliche Bildung unter Cu-defizienten Bedingungen bekannt ist
(MERCHANT und BORGORAD, 1987) bestätigten die Cu-Mangel Induktion der Zellen, so dass
davon ausgegangen werden kann, dass der beobachtete Phänotyp wirklich auf eine Cu-
Defizienz zurückzuführen ist. Im Gegensatz zu S-Mangel scheint sich die Reduktion des
FDX5-Transkriptgehaltes der amiRNA-Transformanden unter Cu-Mangel auch in einer
Reduktion des Fdx5-Proteingehaltes wiederzuspiegeln, was ein Hinweis darauf ist, dass der
Phänotyp der amiRNA Transformanden unter Cu-Mangel durch den „Knock down“ des
FDX5-Gens verursacht wird.
Ein stark verringerter Chlorophyllgehalt von pflanzlichen Zellen, sowie eine verringerte
Photosyntheserate sind oft Symptom für bestehende Eisendefizienz (SPILLER und TERRY,
1980). Dabei bedingt der Mangel an Eisen eine verringerte Akkumulation an
Photosynthesekomplexen, die sich auch in einer Abnahme der photosynthetischen Aktivität
äußert. Dieser Effekt konnte auch an Eisen-defizienten C. reinhardtii Zellen beobachtet
werden, die unter Eisenmangel eine Chlorose ausbilden und eine verringerte
photosynthetische Aktivität besitzen (MOSELEY et al., 2000). Der Grund für die verringerte
photosynthetische Aktivität liegt dabei in einer veränderten Zusammensetzung der
Photosynthesekomplexe und ihrer Lichtsammelantennen (MOSELEY et al., 2000; NAUMANN et
al., 2005). In Hefe und Säugetierzellen sind Cu-Metabolismus und Fe-Metabolismus
miteinander vernetzt, da Cu-Mangel auch zu einer sekundären Eisendefizienz führt (ASKWITH
und KAPLAN, 1998). Wie schon oben beschrieben, ist in C. reinhardtii vermutlich eine Multi-
Kupfer Oxidase an der Assimilation von Eisen beteiligt (LA FONTAINE, 2002), die unter Cu-
Mangel durch das noch unbekannte Genprodukt des Gens CRD2 ersetzt wird. Möglicherweise
ist Fdx5 ebenfalls in einen an der Eisenassimilation beteiligten Stoffwechselschritt involviert,
was die Ausbildung einer sekundären Eisendefizienz unter Cu-Mangel erklären würde.
Einen ähnlicher Phänotyp, wie von den amiRNA-Transformanden unter Cu-Mangel gezeigt,
weisen auch C. reinhardtii Mutanten mit einem Defekt im CRD1-Gen auf (MOSELEY et al.,
2002). Es konnte jedoch gezeigt werden, dass der bei den CRD1-Mutanten aufgetretene
Phänotyp, ein verringerter Chlorophyllgehalt und eine verminderte Photosyntheseaktivität,
bedingt durch die geringe Akkumulation von PSI und einem veränderten Gehalt der

Diskussion
68
Lichtsammelkomplexe, nicht auf einer sekundären Eisendefizienz beruht. Gleichermaßen wie
die Expression von CYC6 und CPX1, ist die CRD1-Expression auch durch Sauerstoffmangel
induzierbar (MOSELEY et al., 2000). Wie schon oben beschrieben, kodiert CRD1 für das
Enzym oxidative Cyclase des Chlorophyll-Stoffwechsels. In C. reinhardtii kann noch eine
weitere Isoform dieser oxidativen Cyclase gefunden werden, die durch das Gen CTH1 kodiert
wird (MOSELEY et al., 2000). CRD1 und CTH1 werden unter Cu- und Sauerstoffmangel
differentiell exprimiert. So konnte nachgewiesen werden, dass CRD1 ausschließlich unter Cu-
und Sauerstoffmangel exprimiert wird, während CTH1 in Cu- und Sauerstoff
supplementierten Zellen aktiv ist (MOSELEY et al., 2002). Aufgrund der phänotypischen
Ähnlichkeiten zwischen CRD1-Mutante und den amiRNA-Transformanden wäre somit auch
eine Funktionalität von Fdx5 in einem Schritt des Chlorophyllbiosyntheseweges möglich, der
in vielen Schritten abhängig von Reduktionsäquivalenten ist, wie z.B. im Falle der NADPH
Protochlorophyllid-Oxidoreduktase (Timko et al., 1998). Da bei den amiRNA-
Transformanden auch eine Reduktion des Carotinoidgehaltes zu beobachten ist, könnte die
Möglichkeit bestehen, dass bei den amiRNA-Transformanden ebenso wie bei den CRD1-
Mutanten eine Störung in der Akkumulation von PSI und Lichtsammelkomplexen vorliegt.
Abb.4-4: Schematische Darstellung von metabolischen Prozessen, die unter Cu-Mangel modifiziert
werden. Unter Cu-Mangel wird die Synthese diverser Proteine (rot markiert) aktiviert, die an verschiedenen
Stoffwechselwegen beteiligt sind. Cpx1 und Crd1 katalysieren Schritte der Tetrapyrrol-Biosynthese, während
Cyc6 das kupferhaltige Plastocyanin der photosynthetischen Elektronentransportkette ersetzt. Crd2 ist an der
Eisenassimilation beteiligt und ersetzt vermutlich eine Multi-Kupfer Oxidase (MERCHANT und BORGORAD,

Diskussion
69
1987; QUINN und MERCHANT, 1998; ERIKSSON et al., 2004, MERCHANT et al., 2006). Die genaue Funktion von
Fdx5 ist bislang ungeklärt. Diese Abbildung stellt eine Übersicht über die bisher untersuchten metabolischen
Adaptionen unter Cu-Mangel dar. Es ist jedoch wahrscheinlich, dass es noch weitere, bislang nicht untersuchte
Stoffwechselprozesse gibt, die ebenfalls von Cu-Mangel beeinflusst werden.
Cyc6 = Cytochrom c6; Crd1 = oxidative Cyclase; Crd2 = bislang noch unbekannt Cyt b6f = Cytochrom-b6f-
Komplex, Cpx1 = Coproporphyrinogen Oxidase; Fd = Ferredoxin; Fdx5 = Ferredoxin5; Pc = Plastocyanin, Pq
= Plastochinon, PS II, I = Photosystem II, I,.
Um jedoch genaue Aussagen zur Funktionalität von Fdx5 tätigen zu können, muss noch eine
detailliertere Analyse des unter Cu-Mangel ausgebildeten Phänotyps der amiRNA-
Transformanden erfolgen. Wichtig wäre hierbei die Untersuchung der photosynthetischen
Aktivität, z.B. durch Chlorophyll-Fluoreszenz Messungen oder die Supplementation der Cu-
Mangelkulturen mit Eisen, um eine sekundäre Eisendefizienz der Zellen zu analysieren.
Interessant wäre zudem eine Untersuchung des Wachstumsverhaltens der amiRNA-
Transformanden unter anaeroben Bedingungen, um zu ermitteln, ob der Phänotyp auch durch
Hypoxie induzierbar ist, wie z.B. im Falle der CRD1-Mutanten.
Das „Silencing“ des FDX5-Gens mittels artifizieller miRNAs liefert somit die ersten
Hinweise auf die Funktion des Fdx5-Proteins und zeigt, dass eine Reduktion des Fdx5-
Gehaltes auch eine phänotypische Reaktion hervorruft. Genaue Analysen sind jedoch
notwendig, um den genauen Wirkungsort von Fdx5 zu bestimmen. Es scheint allerdings
naheliegend, dass Fdx5 ein spezialisiertes Ferredoxin ist, das an spezifischen
Stoffwechselprozessen beteiligt ist, die unter bestimmten Stressbedingungen wie Cu- und
Sauerstoffmangel in C. reinhardtii induziert werden.

Zusammenfassung
70
5 Zusammenfassung
Durch Homologievergleiche konnten im Genom von C. reinhardtii sechs Gene (PET, FDX2,
FDX3, FDX4, FDX5 und FDX6) identifiziert werden, die für (putative) Ferredoxine kodieren.
Semiquantitative RT-PCR-Analysen zeigten, dass diese Ferredoxin-kodierenden Gene in
belichteten, anaeroben S-Mangelkulturen sowie in artifiziell anaerobisierten C. reinhardtii
Kulturen differentiell exprimiert werden. Dabei war FDX5 das einzige der identifizierten
Gene, dessen Transkript unter beiden Bedingungen akkumulierte. Da dies eine vitale Funktion
des FDX5-Genproduktes in der veränderten Physiologie anaerober C. reinhardtii Zellen
andeutet, wurde FDX5 heterolog in E. coli exprimiert und das Fdx5-Protein via Strep-Tag
Affinitätschromatographie aufgereinigt. Das rekombinante Protein wurde für die Produktion
von polyklonalen Antikörpern verwendet, die mittels Immuno Blot Analysen erstmalig den
Nachweis einer Akkumulation von Fdx5-Protein in anaeroben C. reinhardtii Zellen
ermöglichten. Durch N-terminale Sequenzierung konnten die ersten fünf Aminosäuren des
aus C. reinhardtii in nativer Form aufgereinigten Proteins identifiziert und das Vorhandensein
eines N-terminalen Signalpeptides verifiziert werden. Mittels konfokaler
Fluoreszensmikroskopie und Zellfraktionierung konnten anschließend Hinweise auf eine
chloroplastidäre Lokalisation des Fdx5-Proteins ermittelt werden. Die biochemische
Charakterisierung des rekombinanten Proteins zeigte, dass Fdx5 ein [2Fe2S]-Ferredoxin des
Pflanzen-Typs ist. Seine Aminosäuresequenz unterscheidet sich jedoch signifikant von der
typischen Primärsequenz von Pflanzen-Typ Ferredoxinen, welche konservierte negativ
geladene Bereiche aufweist, die bedeutend für die Wechselwirkung des Ferredoxins mit
seinen Reaktionspartnern sind. Diese Abweichungen bedingen eine insgesamt positivere
Nettoladung des Fdx5-Proteins im Vergleich zu anderen Pflanzen-Typ Ferredoxinen. Um
einen ersten Eindruck von der Funktionalität von Fdx5 zu erhalten, wurden Ferredoxin-
abhängige Aktivitätstests durchgeführt. Diese zeigten, dass Fdx5 in vitro keine Elektronen auf
die Hydrogenase HydA1 übertragen und nicht von Photosystem I reduziert werden kann,
während eine Interaktion mit der Ferredoxin-NADP+-Oxidoreduktase möglich ist. Mittels
Gen-„Silencing“ durch artifizielle miRNAs konnte die FDX5-Transkriptakkumulation in zwei
C. reinhardtii Stämmen gehemmt werden. Die Charakterisierung dieser Stämme unter
anaeroben Bedingungen zeigte, dass eine Funktionalität von Fdx5 in typischen
Stoffwechselwegen des photofermentativen Metabolismus eher auszuschließen ist. Unter Cu-
Mangel konnten dabei eindeutige Unterschiede in Pigmentgehalt und photosynthetischer

Zusammenfassung
71
Aktivität der amiRNA-Transformanden im Vergleich zum Wildtyp beobachtet werden. Diese
Unterschiede deuten auf eine Beteiligung des Fdx5-Proteins an den unter Cu-Mangel
stattfindenden Stoffwechselprozessen hin.

Summary
72
6 Summary
BLAST-analyses revealed that the genome of the unicellular green alga C. reinhardtii
contains six genes encoding for (putative) ferredoxins (PET, FDX2, FDX3, FDX4, FDX5 and
FDX6). Semi-quantitative RT-PCR analyses showed that these genes were differentially
expressed in C. reinhardtii cultures that were either anaerobic in the light due to S-deprivation
or that had been artificially deprived of oxygen. Only the FDX5 transcript did accumulate
under both conditions, indicating a vital role of the encoded protein in the anaerobic
metabolism of the cells. For further analyses, FDX5 was heterologously expressed in E. coli
and the Fdx5-protein was purified via Strep-Tag affinity chromatography. Recombinant Fdx5
was used for the generation of polyclonal antibodies, whose application for the first time
demonstrated the accumulation of Fdx5 protein in anaerobic C. reinhardtii cells. N-terminal
sequencing allowed the identification of the first five amino acids of the mature protein,
which was purified from S-deprived C. reinhardtii cells, and verified the occurrence of an N-
terminal signal peptide. Results of confocal fluorescence microscopy and cell fractionation
analyses indicated a chloroplastic localisation of the Fdx5-protein. According to the
biochemical characterisation, Fdx5 is a typical plant-type [2Fe2S]-ferredoxin. However, its
amino acid sequence differs significantly from the typical sequence of plant-type ferredoxins.
These have conserved regions including several negatively charged amino acids which are
essential for the electrostatic interaction of the ferredoxin and its reaction partner. The
variations of the Fdx5-sequence result in a more positive net charge of Fdx5 compared to
other plant-type ferredoxins. Ferredoxin dependent activity assays showed that Fdx5 is not
able to transfer electrons to the hydrogenase HydA1 in vitro nor to accept electrons from PSI,
while it can interact with Fnr. Via gene silencing using artificial miRNAs two transgenic C.
reinhardtii strains were generated that showed significantly decreased FDX5 transcript levels
under anaerobic conditions. Under Cu-deficiency, these strains exhibited considerable
variations in pigment content and photosynthetic activity compared to the wild type. These
variations indicate a participation of the Fdx5-protein in the metabolic processes under Cu-
deficiency

Literaturverzeichnis
73
7 Literaturverzeichnis
Alvarez, J.P., Pekker, I., Goldshmidt, A., Blum, E., Amsellem, Z. and Eshed, Y. (2006) Endogenous and
synthetic microRNAs stimulate simultaneous, efficient, and localized regulation of multiple targets in diverse
species. Plant cell, 18, 1134–1151.
Ambros, V. (2001) microRNAs: tiny regulators with great potential. Cell, 107, 823–826
Antal T.K., Krendeleva T.E., Laurinavichene T.V., Makarova V.V., Ghirardi M.L., Rubin A.B.,Tsygankov
A.A. and Seibert M. (2003) The dependence of algal H2-production on photosystem II and O2consumption
activities in sulphur-deprived Chlamydomonas reinhardtii cells. Biochim Biophys Acta 1607, 153-160
Arnon, D. I. (1965) Ferredoxin and photosynthesis. Science 149, 1460-1470
Askwith C. and Kaplan J. (1998) Iron and copper transport in yeast and its relevance to human disease. Trends in
Biochem. Sci. 23, 135-138
Atteia, A., van Lis R., Gelius-Dietrich G., Adrait A., Garin J., Joyard J., Rolland N., and Martin W.. (2006).
Pyruvate formate-lyase and a novel route of eukaryotic ATP synthesis in Chlamydomonas mitochondria. J. Biol.
Chem.281, 9909-9918.
Bartel, D. P. (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281–297.
Bateman, J.M. and Purton, S. (2000) Tools for chloroplast transformation in Chlamydomonas: expression vectors
and a new dominant selectable marker. Mol. Gen. Genet. 263, 404-410.
Bauer C. E., Elsen S. and Bird T. H. (1999) Mechanisms for redox control of gene expression. Annu. Rev.
Microbiol. 53, 495-523
Beinert H. (2000) Iron-sulfur proteins: ancient structures, still full of surprises. J. Biol. Inorg. Chem. 5, 2-15
Berg, J. M. and Holm, R. H. (1982) Structures and reactions of iron sulfur proteins clusters and their synthetic
analogs, in Iron Sulphur Proteins (Spiro T.G., Ed.), Wiley-Interscience Publications, 1-66
Bertini I., Luchinat C., Provenzani A., Rosato A., Vasos P. R. (2002) Browsing gene banks for Fe2S2
ferredoxins and structural modelling of 88 planttype sequences: an analysis of fold and function. Proteins 46, 110-
127
Bes M. T., Parisini E., Inda L. A., Saraiva L. M.,Peleato M. L.and Sheldrick G. M. (1999) Crystal structure
determination at 1.4 Å resolution of ferredoxin from the green alga Chlorella fusca. Structure 7, 1201-1213.
Blaschkowski, H. P., Neuer G., Ludwig-Festl M., and Knappe J. (1982) Routes of flavodoxin and ferredoxin
reduction in Escherichia coli. CoA acetylating pyruvate:flavodoxin and NADP:flavodoxin oxidoreductase
participatingin the activation of pyruvate formate-lyase. Eur. J. Biochem. 123, 563-569.
Blum, H., Beier, H. & Gross, H.J. (1987) Improved silver staining of plant proteins, RNA and DNA in
polyacrylamid gels. Electrophoresis 8, 93-99.
Böhme, H. (1977) On the role of ferredoxin and ferredoxin-NADP+-reductase in cyclic electron transport of
spinach chloroplasts. Eur. J. Biochem. 72, 283-289.
Bowsher C. G., Hucklesby D. .P, Emes M. J. (1989) Nitrite reduction and carbohydrate metabolism in plastids
purified from roots of Pisum sativum L. Planta 177, 359-366.
Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein
utilizing the principle of protein-dye binding. Anal Biochem 72, 248-54.

Literaturverzeichnis
74
Bruschi M. and Guerlesquin F. (1988) Structure, function and evolution of bacterial ferredoxins. FEMS
Microbiology Reviews 54, 155-176.
Buffoni F. and Ignesti G. (2000) The copper-containing amine oxidases: biochemical aspects and functional role.
Mol. Genet. Metab. 71, 559–564.
Cammack, R., Roa, K. K. , Bargeron, C. P., Hutson, K. G., Andrew, P. W., & Rogers, L. J. (1977) Midpoint
Redox Potentials of Plant and Algal Ferredoxins. Biochem. J. 168,205-209.
Cerutti, H., Johnson, A. M., Gillham, N. W. and Boynton, J. E. (1997) A eubacterial gene conferring
spectinomycin resistance on Chlamydomonas reinhardtii: Integration into the nuclear
genome and gene expression. Genetics 145, 97-110.
Chen J. C., Hsieh S. I., Kropat J., and Merchant S. S. (2008) A Ferroxidase Encoded by FOX1 Contributes to
Iron Assimilation under Conditions of Poor Iron Nutrition in Chlamydomonas. Eukaryotic cell, 7, 3, 541–545
Cohen, S. N., Chang, A. C. and Hsu, L. (1972) Nonchromosomal antibiotic resistance in bacteria: genetic
transformation of Escherichia coli by R-factor DNA. Proc Natl Acad Sci U A 69, 2110-2114.
Davies, D. R. and Plaskitt, A. (1971) Genetical and structural analyses of cell-wall formation in Chlamydomonas
reinhardtii. Genet. Res. (Camb.) 17, 33–43
De Pascalis A. R., Jelesarov I., Ackermann F.,Koppenol W. H., Hirasawa M., Knaff D. B. and Bosshard H.
R. (1993) Binding of ferredoxin to ferredoxin : NADP+ oxidoreductase: The role of carboxyl groups, electrostatic
surface potential, and molecular dipole moment. Protein Science 2, 1126-1135.
Erikkson, M., Gardeström, P. and Samuelsson, G. (1995) Isolation, purification and characterization of
mitochondria from Chlamydomonas reinhardtii. PlantPhysiol. 107, 479-483.
Eriksson M., Moseley J. L., Tottey S., del Campo J. A., Quinn J. M., Kim Y., Merchant S. (2004) Genetic
dissection of nutritional copper signaling in Chlamydomonas distinguishes regulatory and target genes. Genetics
168, 795–807.
Ferguson-Miller S. and Babcock G. T. (1996) Heme/Copper Terminal Oxidases. Chem. Rev. 96, 7, 2889–2908
Finazzi, G., Sommer, F. and Hippler, M. (2005) Release of oxidized plastocyanin from photosystem I limits
electron transfer between photosystem I and cytochrome b6f complex in vivo. Proc. Natl. Acad. Sci. USA 102,
7031–7036.
Fischer N., Setif P., und Rochaix J.-D. (1999) Site-directed Mutagenesis of the PsaC Subunit of Photosystem I. J
Biol Chem. 274, 33, 23333-23340.
Fish, W. W. (1988) Rapid colorimetric micromethod for the quantitation of complexed iron in biological samples.
Meth. Enzymol. 158, 357-364.
Fouchard S., Hemschemeier A., Caruana A., Pruvost J., Legrand J., Happe T., Peltier G., Cournac L. (2005)
Autotrophic and mixotrophic hydrogen photoproduction in sulfur-deprived Chlamydomonas cells. Appl Environ
Microbiol 71, 6199-6205.
Franzen L.-G., Rochaix J.-D. and von Heijne G. (1990) Chloroplast transit peptides from the green alga
Chlamydomonas reinhardtii share features with both mitochondrial and higher plantchloroplast presequences.
FEBS Letters 260, 2, 165-168.
Frey, M., Rothe M., Wagner A. F. V. and Knappe J. (1994). Adenosylmethionine dependent synthesis of the
glycyl radical in pyruvate formate-lyase by abstraction of the glycine C-2 pro-S hydrogen atom. J. Biol. Chem. 269,
12432–12437.

Literaturverzeichnis
75
Fuhrmann M., Oertel W. and Hegemann P. (1999) A synthetic gene coding for the green fluorescent protein
(GFP) is a versatile reporter in Chlamydomonas reinhardtii. Plant J 19, 353-61.
Fukuyama K. (2004) Structure and function of plant-type ferredoxins. Photosynthesis Research 81, 289–301.
Galvan F., Marquez A. J. und Fernandez E. (1985) Physicochemical Properties of Ferredoxin from
Chlamydomonas reinhardtii. Z.Naturforsch. 40c, 373-378.
García-Sánchez M. I., Díaz-Quintana A., Gotor C., Jacquot J.-P., De la Rosa M. A., Vega J. M. (2000)
Homology predicted structure and functional interaction of ferredoxin from the eukaryotic alga Chlamydomonas
reinhardtii with nitrite reductase and glutamate synthase. J Biol Inorg Chem. 5, 6, 713-719.
Gavel, Y. and von Heijne, G. (1990) A conserved cleavage-site motif in chloroplast transit peptides. FEBS Lett.
261, 455-458.
Gfeller R. P. and Gibbs M. (1984) Fermentative metabolism of Chlamydomonas reinhardtii. I. Analysis of
fermentative products from starch in dark and light. Plant Physiol. 75, 212-218.
Girbal L., von Abendroth G., Winkler M., Benton P. M., Meynial-Salles I., Croux C., Peters J. W., Happe T.,
Soucaille P. (2005) Homologous and heterologous overexpression in Clostridium acetobutylicum and
characterization of purified clostridial and algal Fe-only hydrogenases with high specific activities. Appl Environ
Microbiol. 71, 5, 2777-2781.
Glanz, S., Bunse, A., Wimbert, A., Balczun, C., and Kuck, U. (2006) A nucleosome assembly protein-like
polypeptide binds to chloroplast group II intron RNA in Chlamydomonas reinhardtii. Nucleic Acids Res 34, 5337-
51.
Guenther J. E. and Melis A. (1990) The physiological significance of photosystem II heterogeneity in
chloroplasts. Photosynth. Res. 23, 105-109.
Guerrini O., Burlat B., Leger C., Guigliarelli B. , Soucaille P. and Girbal L. (2008) Characterization of Two
2[4Fe4S] Ferredoxins from Clostridium acetobutylicum. Curr Microbiol 56, 261–267.
Guillouard I., Lagoutte B., Moal G. and Bottin H. (2000) Importance of the Region Including Aspartates 57 and
60 of Ferredoxin on the Electron Transfer Complex with Photosystem I in the Cyanobacterium Synechocystis sp.
PCC 6803. Biochem Biophys Res Commun 271, 647–653.
Hall D. O., Cammack R., Rao K. K. (1973) The plant ferredoxins and their relationship to the evolution of
ferredoxins from primitive life. Pure and Applied Chem. 34, 553-557.
Hall D. O., Cammack R., Rao K. K. ( 1974) The iron sulphur proteins: Evolution of a ubiquitous protein from
model systems to higher organisms. Orig Life, 5, 3, 363-86.
Hall D. O., Cammack R., Rao K. K. (1977) Chemie und Biologie der Eisen-Schwefel-Proteine. Chemie in
unserer Zeit 71, 165-173.
Hanke G. T., Kimata-Ariga Y., Taniguchi I., and Hase T (2004) A Post Genomic Characterization of
Arabidopsis Ferredoxins. Plant Physiol. 134, 255-264.
Hanukoglu I. (1996) Electron transfer proteins of cytochrome P450 systems. Adv Mol Cell Biol 14, 29-55.
Happe, T. and Naber, J. D. (1993) Isolation, characterization and N-terminal amino acid sequence of hydrogenase
from the green alga Chlamydomonas reinhardtii. Eur. J. Biochem. 214, 475-481.
Happe, T., Mosler, B. and Naber, J. D. (1994) Induction, localization and metal content of hydrogenase in the
green alga Chlamydomonas reinhardtii. Eur. J.Biochem. 222, 769-774.
Happe T. and Kaminski A. (2002) Differential regulation of the Fe-hydrogenase during anaerobic adaptation in
the green alga Chlamydomonas reinhardtii. Eur J Biochem 269, 1022-1032.

Literaturverzeichnis
76
Happe T., Hemschemeier A., Winkler M. and Kaminski A. (2002) Hydrogenases in green algae: Do they save
the algae´s life and solve our energy problems? Trends Plant Sci 7, 6, 246-250.
Harris E. H. (1989) The Chlamydomonas sourcebook. Academic Press, Inc, San Diego
Harris E. H. (2001) Chlamydomonas as a model organism. Annu Rev Plant Physiol Plant Mol. Biol. 52, 363-406.
Hartman F. C. and Harpel M. R. (1994) Sturcture, function, regulation and assembly of D-Ribulose-1,5-
Bisohosphate Carboxylase/ Oxygenasae. Annu. Rev. Biochem. 63, 197-234.
Hasan M. N., Hagedoorn P. L. and Hagen W. R. (2002) Pyrococcus furiosus ferredoxin is a functional dimer.
FEBS Letters 53, 335-338.
Hase T., Kimata Y., Yonekura K., Matsumura T., Sakakibara H. (1991 a) Molecular cloning and differential
expression of the maize ferredoxin gene family. Plant Physiol 96, 77-83.
Hase T., Mizutani S. and Mukohata Y. (1991 b) Expression of Maize Ferredoxin cDNA in Escherichia coil.
Comparison of Photosynthetic and Nonphotosynthetic Ferredoxin Isoproteins and their Chimeric Molecule Plant
Physiol. 97, 1395-1401.
Heitzer M. and Hallmann A. (2002) An extracellular matrix-localized metalloproteinase with an exceptional
QEXXH metal binding site prefers copper for catalytic activity. J. Biol. Chem. 277, 28280–28286.
Hemschemeier A. (2002) Der H2-Metabolismus von Chlamydomonas reinhardtii unter Schwefelmangel: eine
physiologische und molekularbiologische Studie. Diplomarbeit an der Friedrich-Wilhelms Universität, Bonn,
Deutschland.
Hemschemeier, A. and T. Happe. (2005) The exceptional photofermentative hydrogen metabolism of the green
alga Chlamydomonas reinhardtii. Biochem.Soc. Trans. 33, 39-41.
Hemschemeier, J. Jacobs and T. Happe (2008 a), Biochemical and Physiological Characterization of the
Pyruvate Formate-Lyase Pfl1 of Chlamydomonas reinhardtii, a Typically Bacterial Enzyme in a Eukaryotic Alga.
Eukaryotic Cell 7, 3, 518-526.
Hemschemeier A., Fouchard S., Cournac L., Peltier G., Happe T. (2008 b) Hydrogen production by
Chlamydomonas reinhardtii: an elaborate interplay of electron sources and sinks. Planta 227, 397-407.
Herbik A., Bolling C., Buckhout T.J. (2002) The involvement of a multicopper oxidase in iron uptake by the
green algae Chlamydomonas reinhardtii. Plant Physiol. 130, 2039–2048.
Hill K. L., Li H. H., Singer J., Merchant S. (1991) Isolation and structural characterization of the
Chlamydomonas reinhardtii gene for cytochrome c6 analysis of the kinetics and metal specificity of its copper-
responsive expression. J Biol Chem 266, 15060–15067.
Huber C. and Wächtershäuser G. (1998) Peptides by Activation of Amino Acids with CO on (Ni,Fe)S Surfaces:
Implications for the Origin of Life Science 281, 670-672.
Inoue K., Hase T., Matsubara H., Fitzgerald M. P. and Rogers L. J. (1984) Amino acid sequence of a
ferredoxin from Rhodymenia palmata, a red alga in the Florideophyceae. Phytochemistry 23, 773-776.
Jacobs J. (2005) Der anaerobe Stoffwechsel von Chlamydomonas reinhardtii: Neue Erkenntnisse mit Real-Time
PCR und heterologer Expression. Diplomarbeit an der Ruhr-Universität Bochum, Bochum, Deutschland.
Jacquot J. P., Stein M., Suzuki A., Liottet S., Sandoz G., Miginiac-Maslow M. (1997) Residue Glu-91 of
Chlamydomonas reinhardtii ferredoxin is essential for electron transfer to ferredoxin-thioredoxin reductase. FEBS
Lett. 400, 3,293-296.

Literaturverzeichnis
77
Jans F., Mignoleta E., Houyouxa P.-A., Cardola P., Ghysels B., Cuine S., Cournac L. , Peltierc G., Remacle
C., Francka, F. (2008) A type II NAD(P)H dehydrogenase mediates light-independent plastoquinone reductionin
the chloroplast of Chlamydomonas. Proc Natl Acad Sci USA. 105, 51, 20546-20551.
Johanningmeier U. and Howell S. H. (1984). Regulation of light-harvesting chlorophyll-binding protein mRNA
accumulation in Chlamydomonas reinhardtii. Possible involvement of chlorophyll synthesis precursors. J. Biol.
Chem. 259, 13541–13549.
Johnson M. K. (1994) Iron-sulfur proteins. In Encyclopedia of Inorganic Chemistry, ed. RB King,.1896-1915.
Chichester, UK.
Johnson D. C., Dean D. R., Smith A. D. and Johnson M. K. (2005) Stucture, function and formation of
biological iron-sulfur clusters. Annu. Rev. Biochem. 74, 247-281.
Kiley P. J. and Beinert H. (1998) Oxygen sensing by the global regulator, FNR: the role of the iron-sulfur cluster.
FEMS Microbiology Reviews 22, 5, 341-352.
Kimata Y. and Hase T. (1989) Localization of Ferredoxin Isoproteins in Mesophyll and Bundle Sheath Cells in
Maize Leaf. Plant Physiol. 89, 1193-1197.
Kindle K. L. (1990) High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proc. natl. Acad. Sci
USA 87, 1228-1232.
Knaff D. B. (1996) Ferredoxin and ferredoxin dependent enzymes. In DR Ort, CF Yocum, eds, Oxygenic
Photosynthesis: The Light Reactions. Kluwer Academic Publishers, Dordrecht, The Netherlands, 333-361.
Knappe J., Neugebauer F. A., Blaschkowski H. P. and Gänzler M. (1984) Post-translational activation
introduces a free radical into pyruvate formate-lyase. Proc. Natl. Acad. Sci. USA 81, 1332-1335.
Kreuzberg K. (1984) Starch fermentation via a formate producing pathway in Chlamydomonas reinhardtii,
Chlorogonium elongatum and Chlorella fusca. Physiol. Plant. 61, 87-94.
Kropat J., Tottey S., Birkenbihl R. P., Depège N., Huijser P. and Merchant S. (2005) A regulator of nutritional
copper signaling in Chlamydomonas is an SBP domain protein that recognizes the GTAC core of copper response
element. Proc Natl Acad Sci USA 102, 51,18730-18735.
Kurisu G., Kusunoki M., Katoh E., Yamazaki T., Teshima K., Onda Y., Kimata-Ariga Y. and Hase T. (2001)
Structure of the electrontransfer complex between ferredoxin and ferredoxin-NADP+ reductase. Nature Structural
Biology 8, 117 – 121.
Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature
227, 680-685
La Fontaine S., Quinn J.M., Nakamoto S.S., Page M.D., Göhre V., Moseley J.L., Kropat J., Merchant S.,
(2002) Copper-dependent iron assimilation pathway in the model photosynthetic eukaryote
Chlamydomonasreinhardtii. Eukaryotic Cell 1,5, 736–757.
Lax, J., Arteni, A., Boekema, E., Pistorius, E., Michel, K.-P. and Rögner, M. (2007) Structural response of
Photosystem 2 to iron deficiency: Characterization of a new Photosystem 2-IdiA complex from the cyanobacterium
Thermosynechococcus elongatus BP-1. Biochim. Biophys. Acta 1767, 528-534.
Lelong C., Setif P., Lagoutte B. and Bottin H. (1994) Identication of the amino acids involved in the functional
interaction between photosystem I and ferredoxin from Synechocystis sp. PCC 6803 by chemical crosslinking. J.
Biol. Chem. 269, 10034-10039.
Lien T. and Schreiner O. (1975) Purification of a derepressible arylsulfatase from Chlamydomonas reinhardtii.
Biochim. Biophys. Acta 384, 168-179.

Literaturverzeichnis
78
Maul J. E., Lilly J. W., Cui L., dePamphilis C. W., Miller W., Harris E. H. and Stern D. B. (2002) The
Chlamydomonas reinhardtii plastid chromosome: islands of genes in a sea of repeats. Plant Cell. 11, 2659-2679.
Mayoral, T., Martinez-Julvez M., Pérez-Dorado I., Sanz-Aparicio J., Gómez-Moreno C., Medina M.and
Hermoso J. A. (2005) Structural analysis of interactions for complex formation between Ferredoxin-NADP+
reductase and its protein partners. Proteins 59, 3, 592-602.
Melis A., Zhang L., Forestier M., Ghirardi M. L. and Seibert M. (2000) Sustained photobiological hydrogen
gas production upon reversible inactivation of oxygen evolution in the green alga Chlamydomonas reinhardtii.
Plant Physiol. 122, 127-135.
Melis A. (2007) Photosynthetic H2 metabolism in Chlamydomonas reinhardtii (unicellular green algae) Planta.
226, 5, 1075-86.
Merchant S. and Bogorad L. (1987) Metal ion regulated gene expression: use of a plastocyanin-less mutant of
Chlamydomonas reinhardtii to study the Cu (II)-dependent expression of cytochrome c-552, EMBO J. 6, 2531–
2535.
Merchant S. S., Hill K. and Howe G. (1991) Dynamic interplay between two copper-titrating components in the
transcriptional regulation of cyt c6. EMBO J. 10, 6,1383–1389.
Merchant S., Allen M. D., Kropat J., Moseley J. L., Long J. C., Tottey S. and Terauchi A. M. (2006) Between
a rock and a hard place: trace element nutrition in Chlamydomonas. Biochim. Biophys. Acta 1763, 578-594.
Merchant S. S., Prochnik S. E., Vallon O., Harris E. H., Karpowicz S. J., Witman G. B., Terry A., Salamov
A., Fritz-Laylin L. K., Maréchal-Drouard L., Marsha ll W. F., Qu L. H., Nelson D. R., Sanderfoot A. A.,
Spalding M. H., Kapitonov V. V., Ren Q., Ferris P., Lindquist E., Shapiro H., Lucas S. M., Grimwood J.,
Schmutz J., Cardol P., Cerutti H., Chanfreau G., Chen C. L., Cognat V., Croft M. T., Dent R., Dutcher S.,
Fernández E., Fukuzawa H., González-Ballester D., González-Halphen D., Hallmann A., Hanikenne M.,
Hippler M., Inwood W., Jabbari K., Kalanon M., Kura s R., Lefebvre P. A., Lemaire S. D., Lobanov A. V.,
Lohr M., Manuell A., Meier I., Mets L., Mittag M., Mittelmeier T., Moroney J. V., Moseley J., Napoli C.,
Nedelcu A. M., Niyogi K., Novoselov S. V., Paulsen I. T., Pazour G., Purton S., Ral J. P., Riaño-Pachón D.
M., Riekhof W., Rymarquis L., Schroda M., Stern D., Umen J., Willows R., Wilson N., Zimmer S. L., Allmer
J., Balk J., Bisova K., Chen C. J., Elias M., Gendler K., Hauser C., Lamb M. R., Ledford H., Long J. C.,
Minagawa J., Page M. D., Pan J., Pootakham W., Roje S., Rose A., Stahlberg E., Terauchi A. M., Yang P.,
Ball S., Bowler C., Dieckmann C. L., Gladyshev V. N., Green P., Jorgensen R., Mayfield S., Mueller-Roeber
B., Rajamani S., Sayre R. T., Brokstein P., Dubchak I., Goodstein D., Hornick L., Huang Y. W., Jhaveri J.,
Luo Y., Martínez D., Ngau W. C., Otillar B., Poliakov A. , Porter A., Szajkowski L., Werner G., Zhou K.,
Grigoriev I. V., Rokhsar D. S., Grossman A. R. (2007) The Chlamydomonas genome reveals the evolution of
key animal and plant functions. Science. 318, 5848, 245-250.
Meyer J. (2001) Ferredoxins of the third kind. FEBS Letters 509, 1-5.
Michaelis G., Vahrenholz C. and Pratje E. (1990) Mitochondrial DNA of Chlamydomonas reinhardtii: The gene
for apocytochrome b and the complete functional map of the 15.8 kb DNA. Mol Gen Genet 223, 211-216.
Minko I., Holloway S. P., Nikaido S., Carter M., Odom O. W., Johnson C. H. and Herrin D.L. (1999) Renilla
luciferase as a vital reporter for chloroplast gene expression in Chlamydomonas. Mol. Gen. Genet. 262, 421-425.
Molnár A.,Schwach F., Studholme D. J., Thuenemann E. C., Baulcombe D. C. (2007) miRNAs control gene
expression in the single-cell alga Chlamydomonas reinhardtii. Nature.447, 7148, :1126-1129.

Literaturverzeichnis
79
Molnár A., Bassett A., Thuenemann E. C., Schwach F., Karkare S., Ossowski S., Weigel D. and Baulcombe
D. C. (2009) Highly specific gene silencing by artificial microRNAs in the unicellular alga Chlamydomonas
reinhardtii. Plant J. 58, 1, 165-174.
Mortenson L. E., Valentine R. C. and Carnahan J. E. (1962) An electron transport factor from Clostridium
pasteurianum. Biochem. Biophys. Res. Comm. 7, 488-452.
Moseley J.L., Quinn J., Eriksson M. and Merchant S. (2000) The Crd1 gene encodes a putative di-iron enzyme
required for photosystem I accumulation in copper deficiency and hypoxia in Chlamydomonas reinhardtii. EMBO
J. 19, 2139–2151.
Moseley J. L., Page M. D., Alder N. P., Eriksson M., Quinn J., Soto F., Theg S. M., Hippler M. and S.
Merchant (2002) Reciprocal expression of two candidate di-iron enzymes affecting photosystem I and light-
harvesting complex accumulation. Plant Cell 14, 673–688.
Mullinger R. N., Cammack R., Rao K. K. and Hall D. O. (1975) Physicochemical Characterization of the Four-
Iron-Four-Sulphide Ferredoxin from Bacillus stearothermophilus. Biochem. J. 151, 75-83.
Naqvi A. R., Islam M. D. 1, ChoudhuryN. R. and Haq Q. M. (2009) The Fascinating World of RNA
Interference. Int. J. Biol. Sci. 5, 2,97-117.
Naumann B., Stauber E. J., Busch A., Sommer F. and Hippler M. (2005) N-terminal processing of Lhca3 is a
key step in remodeling of the photosystem I- light-harvesting complex under iron-deficiency in Chlamydomonas
reinhardtii. J. Biol. Chem. 280, 21, 20431-20441.
Neale P. J., Cullen J. J., Lesser M. P. and Melis A. (1993) Physiological bases for detecting and predicting
photoinhibition of aquatic photosynthesis by PAR and UV radiation. In: Current Topics in Plant Physiology
(Yamamoto, H.J. and Smith, C.M., Eds.), Photosynthetic responses to the environment, Vol. 8, pp.61-77, American
Society of Plant Physiologists, Rockville, MD, USA
Newman S. M., Boynton J. E., Gillham N. W., Randolph-Anderson B. L., Johnson A. M. and Harris E. H.
(1990) Transformation of Chloroplast Ribosomal RNA Genes in Chlamydomonas: Molecular and Genetic
Characterization of Integration Events Genetics 126; 875–888
Ohad I., Kyle D. J. and Arntzen C. J. (1984) Membrane protein damage and repair: Removal and replacement of
inactivated 32-kilodalton polypeptide in chloroplast membranes. J Cell Biol 89, 481-485.
Onda Y, Matsumura T, Kimata-Ariga Y, Sakakibara H, Sugiyama T, Hase T (2000) Differential interaction of
maize root ferredoxin:NADP(�) oxidoreductase with photosynthetic and non-photosynthetic ferredoxin
isoproteins. Plant Physiol 123, 1037–1045.
Österberg, R. (1974) Origins of metal ions in biology. Nature 249: 382-383
Petering D., Fee J. A., Palmer G. (1971)The oxygen sensitivity of spinach ferredoxin and other iron-sulfur
proteins. The formation of protein-bound sulfur-zero. J Biol Chem 246, 643–653.
Pieulle L., Magro V. and Hatchikian E. C. (1997) Isolation and analysis of the gene encoding the
pyruvateferredoxin oxidoreductase of Desulfovibrio africanus, production of the recombinant enzyme in
Escherichia coli and effect of carboxy-terminal deletions on its stability. J Bacteriol 179, 5684-5692.
Piubelli L, Aliverti A, Bellintani F, Zanetti G. (1996) Mutations of Glu92 in ferredoxin I from spinach leaves
produce proteins fully functional in electron transfer but less efficient in supporting NADP+ photoreduction. Eur J
Biochem. 236, 2,465-469.

Literaturverzeichnis
80
Poncelet M., Cassier-Chauvat C., Leschelle X., Bottin H., Chauvat F. (1998)Targeted deletion and mutational
analysis of the essential (2Fe2S) plant-like ferredoxin in Synechocystis PCC 6803 by plasmid shuffeling, Mol.
Microbiol. 28, 813-821.
Purton, S., and Rochaix, J. D. (1994) Complementation of a Chlamydomonas reinhardtii mutant using a genomic
cosmid library. Plant Mol Biol 24, 533-7.
Quinn J. M. and Merchant S. (1998) Copper-responsieve gene expression during adaption to Copper deficiency.
Methods Enzymol. 297, 263-279.
Quinn J. M., Barraco P., Eriksson M. and Merchant S. (2000) Coordinate Copper- and Oxygen-responsive
Cyc6 and Cpx1 Expression in Chlamydomonas is Mediated by the Same Element. J Biol Chem 275, 9, 6080–6089.
Quinn J. M., Eriksson M., Moseley J. L. and Merchant S. (2002) Oxygen Deficiency Responsive Gene
Expression in Chlamydomonas reinhardtii through a Copper-Sensing Signal Transduction Pathway. Plant Physiol.
128, 463–471.
Raleigh, E. A., Lech, K and Brent, R. (1989) Selected topics in classical bacterial genetics. In: Ausubel, F.M.,
Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., and Struhl, K., Eds. Current Protocols in
Molecular Biology. John Wiley and Sons, New York. pp 1.4.1-1.4.7.
Ravina C. G., Barroso C., Vega J. M. and Gotor C. (1999) Cysteine biosynthesis in Chlamydomonas reinhardtii.
Molecular cloning and regulation of O-acetylserine(thiol)lyase. Eur. J. Biochem. 264, 848 – 853.
Reubelt U., Wohlfarth G., Schmid R. and Diekert G. (1991) Purification and characterization of ferredoxin from
Peptostreptococcus productus (strain Marburg). Arch Microbiol 156, 422-426.
Rochaix J.- D., Fischer N., Hippler M. (2000) Chloroplast site-directed mutagenesis of photosystem I in
Chlamydomonas: Electron transfer reactions and light sensitivity. Biochimie 82, 635−645.
Rochaix J.-D. (1995) Chlamydomonas reinhardtii as the photosynthetic yeast. Annu Rev Genet 29, 209-230.
Rogers W. J., Hodges M., Decottignies P., Schmitt J.-M., Gadal P. and Jacquot J.-P. (1992) Isolation of a
cDNA fragment coding for Chlamydomonas reinhardtii ferredoxin and expression of the recombinant protein in
Escherichia coli. FEBS Lett. 310, 3, 240-245.
Sambrook, J., Tritsch, E. F. and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2nd edn, Cold
Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Sanger, F., Nicklen, S., and Coulson, A. R. (1977) DNA sequencing with chain-terminating inhibitors. Proc Natl
Acad Sci USA 74, 5463-5467.
Sato N., Hagio M., Wada H. and Tsuzuki M. (2000) Environmental effects on acidic lipids of thylakoid
membranes. Biochem Soc Trans 28, 912-914.
Sawers R. G., Zalelein E. and Böck A. (1988) Two-dimensional gel electrophoretis analysis of Escherichia coli
proteins: influence of various anaerobic growth conditions and the fnr gene product on cellular protein composition.
Arch. Microbiol. 149, 240-244.
Schägger H. (2006) Tricine–SDS-PAGE. Nature protocols 1, 16-22
Schmidt H. and Heinz E. (1990) Desaturation of oleoyl groups in envelope membranes from spinach chloroplasts.
Proc. Natl. Acad. Sci. USA 87, 9477-9480.
Schmitter, J. M., Jacquot, J. P., de Lamotte-Guery, F., Beauvallet, C., Dutka, S., Gadal, P. and Decottignies
P. (1988) Purification, properties and complete amino acid sequence of the ferredoxin from a green alga,
Chlamydomonas reinhardtii. Eur. J. Biochem. 172, 405-412.
Schopfer P. (1989) Experimentelle Pflanzenphysiologie II. Einführung in die Anwendungen, Springer, Berlin

Literaturverzeichnis
81
Schrautemeier B, Neveling U, Schmitz S (1995) Distinct and differently regulated Mo-dependent nitrogen-fixing
systems evolved for heterocysts and vegetative cells of Anabaena variabilis ATCC 29413: Characterization of the
FdxH1/2 gene regions as part of the nif1/2 gene clusters. Mol Microbiol 18, 357–369.
Schroda M. (2006) RNA silencing in Chlamydomonas: mechanisms and tools. Curr Genet. 49, 2, 69-84.
Schütz K., Happe T., Troshina O., Lindblad P., Leitao E., Oliveira P., Tamagnini P. (2004) Cyanobacterial H2
production- a comparative analysis. Planta 218, 350-359.
Schwab, R., Ossowski, S., Riester, M., Warthmann, N. and Weigel, D. (2006) Highly specific gene silencing by
artificial microRNAs in Arabidopsis. Plant cell, 18, 1121–1133.
Shimomura O., Johnson F. H. and Saiga Y (1962) Extraction, purification and properties of aequorin, a
bioluminescent protein from the luminous hydromedusan, Aequorea. J Cell Comp Physiol. 59, 223-239.
Singh B. B., Curdt I., Shomburg D., Bisen P. S. and Böhme H. (2001) Valine 77 of heterocystous ferredoxin
FdxH2 in Anabaena variabilis strain ATCC 29413 is critical for its oxygen sensitivity. Mol. Cell Biochem. 217,
137–142.
Skerra, A. (1994). Use of the tetracycline promoter for the tightly regulated production of a murine antibody
fragment in Escherichia coli. Gene 151, 131-135.
Smith T. E. and Feinberg, B. A. (1990) Redox Properties of Several Bacterial Ferredoxins Using Square Wave
Voltammetry*, J. Biol. Che. 265, 24, 14371-14376.
Spiller, S. and Terry, N. (1980) Limiting factors in photosynthesis. II. Iron stress diminishes photochemical
capacity by reducing the number of photosynthetic units. Plant Physiol. 65, 121-125.
Stal L. J. and Moezelaar R. (1997) Fermentation in cyanobacteria. FEMS Microbiol Rev 21, 179-211.
Stevens, D. R., Rochaix, J.-D. and Purton, S. (1996) The bacterial phleomycin resistance gene ble as a dominant
selectable marker in Chlamydomonas. Mol. Gen. Genet. 251, 25-30.
Stirnberg, M. and Happe T. (2004) Identification of a cis-acting element controlling anaerobic expression of the
hydA-gene from Chlamydomonas reinhardtii. In "Biohydrogen III" (Miyake J., Igarashi Y., Roegner M..,eds),
Elsevier Science, Oxford, 117-127.
Studier F. W. and Moffatt B. A. (1986) Use of bacteriophage T7 RNA polymerase to direct selective high-level
expression of cloned genes. J Mol Biol 189, 113-130
Suzuki A.,Oaks A., Jacquot J.-P., Vidal J. and Gadal P. (1985) An electron tranport system in maize roots for
reactions of glutamate synthase and nitrite reductase: physiological and Immunochemical properties of the electron
carrier and pyridine nucleotide reductase. Plant Physiol 78, 374-378.
Szoboszlai N., Polgári Z., Mihucz V. G. and Záray G. (2008) Recent trends in total reflection X-ray fluorescence
spectrometry for biological applications, Anal Chim Acta. 633, 1, 1-18.
Tagawa K. and Arnon D. 1. (1962) Ferredoxins as Electron Carriers in Photosynthesis and in the Biological
Production and Consumption of Hydrogen Gas. Nature 195, 537-543.
Tam L.-W. and Lefebvre P.A. (1993) Cloning of flagellar genes in Chlamydomonas reinhardtii by insertional
mutagenesis. Genetics 135, 375-384.
Terpe, K. (2003) Overview of tag protein fusions: from molecular and biochemical fundamentals to commercial
systems. Appl. Microbiol. Biotechnol. 60, 523-533.
Timko, M. P. (1998) Pigment biosynthesis: Chlorophylls, Heme, and Carotenoids. In The Molecular Biology of
Chloroplasts and Mitochondria in Chlamydomonas. (Rochaix, J.-D., Goldschmidt-Clermont, M., and Merchant, S.,
eds), Kluwer Academic Publishers, Dordrecht, The Netherlands, 377-414

Literaturverzeichnis
82
Tsien R. Y. (1998) The green Fluorescent protein. Annu. Rev. Biochem. 67, 509–544.
Verhagen, M., F., J., M., Link, T., A. and Hagen, W. R. (1995) Electrochemical study of the redox properties of
[2Fe-2S] ferredoxins. Evidence for superreduction of the Rieske [2Fe-2S] cluster, FEBS Letters 361, 75-78.
Vogel, H. J. and D. M. Bonner. (1955) Acetylornithinase of Escherichia coli: partial purification and some
properties. J. Biol. Chem. 218, 97-106.
Wada K., Onda M. and Matsubara H. (1986) Ferredoxin isolated from plant non-photosynthetic tissues:
purification and characterization. Plant Cell Physiol 27, 407-415.
Wagner A.F.V., Frey M., Neugebauer F.A., Schäfer W. and Knappe J. (1992) The free radical in pyruvate
formate-lyase is located on glycine-734. Proc. Natl. Acad. Sci. USA 89, 996-1000
Webber A. N., Scott E. B. and Hyeonmoo L. (1995) Genetic engineering of thylakoid protein complexes by
chloroplast transformation in Chlamydomonas reinhardtii. Photosynthesis Res. 44, 191-205.
Weber A. (2002) Molekularbiologische Untersuchungen zum Wasserstoffmetabolismus der Grünalge
Chlamydomonas reinhardtii. Diplomarbeit an der Friedrich-Wilhelms Universität, Bonn, Deutschland.
Winkler, M. (2009) Charakterisierung der [FeFe]-Hydrogenasen des Chlorophyta-Typs - Struktur-
Funktionsbeziehungen, Verbreitung und Phylogenie, Doktorarbeit an der Ruhr-Universität Bochum, Bochum,
Deutschland
Wood P. M. (1978) Interchangeable copper and iron proteins in algal photosynthesis. Studies on plastocyanin and
cytochrome c-552 in Chlamydomonas. Eur.J.Biochem. 87, 9–19.
Woodcock, D.M., Crowther, P.J., Doherty, J., Jefferson, S., Decruz, E., Noyer-Weidner, M., Smith, S.S.,
Micheal, M.Z. and Graham, M.W. (1989) Quantitative evaluation of Escherichia coli host strains for tolerance to
cytosine methylation in plasmid and phage recombinants. Nuc Acids Res 17, 3469-3478
Wykoff D. D., Davies J. P., Melis A. und Grossman A. R. (1998) The regulation of photosynthetic electron
transport during nutrient deprivation in Chlamydomonas reinhardtii. Plant Physiol. 117, 129-139.
Xu X., Kim S. K., Schürmann P., Hirasawa M., Tripathy J. N., Smith J., Knaff D. B., Ubbink M. (2006)
Ferredoxin/ferredoxin-thioredoxin reductase complex: Complete NMR mapping of the interaction site on
ferredoxin by gallium substitution. FEBS Lett. 580, 28, 6714-6720.
Yildiz F. H., Davies J. P. und Grossman A. R. (1996) Sulfur availability and the SAC1 gene control adenosine
triphosphate sulfurylase gene expression in Chlamydomonas reinhardtii. Plant Physiol. 112, 669-675.
Zerges, W. and Rochaix, J.-D. (1998) Low density membranes are associated with RNA-binding proteins and
thylakoids in the chloroplast of Chlamydomonas reinhardtii. J. Cell Biol. 140, 101-110.
Zhang L., Happe T. und Melis A. (2002) Biochemical and morphological characterization of sulfur-deprived and
H2-producing Chlamydomonas reinhardtii (green alga). Planta 214, 552-561.
Zhang Z., Shrager J., Jain M., Chang C.-W. , Vallon O. and Grossman A. R. (2004) Insights into the Survival
of Chlamydomonas reinhardtii during Sulfur Starvation Based on Microarray Analysis of Gene Expression.
Eukaryotic cell 3, 5, 1331–1348.
Zhao T., Wang W., Bai X., Qi Y. (2008) Gene silencing by artificial microRNAs in Chlamydomonas. Plant J. 58,
1, 157-164.
Anhang
83
8 Anhang
8.1 Auflistung der Ferredoxin-kodierenden Gene aus C. reinhardtii
Gene Protein ID (JGI4.0) Chromosom und Position (JGI4.0) GenBank Nr. (NCBI)
PETF 147787 14:2749384-2750217 P07839
FDX2 159161 16:1387337-1388874 ABC88601
FDX3 196707 6:7017104-7018511 ABC88602
FDX4 196705 7:3072834-3074762 ABC88603
FDX5 156833 17:702791-704769 ABC88604
FDX6 196703 3:4305938-4308048 ABC88605
Angegeben sind Protein ID der C. reinhardtii Datenbank JGI 4.0, die Lokalisation des Gens im Genom und die
Genbank Nr. der NCBI Datenbank.
8.2 Auflistung der verwendeten Arbeitsgeräte
Gerät Modell Hersteller
Anaerobzelt CLc Coy Laboratory, Grass
Lake, MI, USA
Autoklaven
VST 40/60
Varioklav 300/400/500 EP-Z
Zirbus, Bad Grund,
Deutschland
H+P Labortechnik AG,
Oberschleissheim,
Deutschland
Brutschränke
B 28
MBG
WTB
Binder, Tuttlingen,
Deutschland
Binder, Tuttlingen,
Deutschland
Binder, Tuttlingen,
Deutschland
Chemilumineszenz-Detektor FluorChem 8800 Alpha Innotech, San
Leandro, Ca, USA
Distille Cyclon Fistreem, Loughborough,
UK
Elektrophoreseapparaturen Mini-Protean Biorad, Hercules, Ca, USA
Feinwaage AW 320
P1200N
Shimadzu, Duisburg,
Deutschland
Mettler, Columbus, OH,
USA
Anhang
84
Gaschromatograph GC 2010 Shimadzu, Duisburg,
Deutschland
Geldokumentationsgerät Gel Max Intas, Göttingen,
Deutschland
Heizschüttler TM compact Eppendorf, Hamburg,
Deutschland
Impfbank HF 48 Gelaire, Sydney, Australien
Mikroskop 4379553 Carl Zeiss, Jena,
Deutschland
PCR-Geräte Mastercycler personal Eppendorf, Hamburg,
Deutschland
pH-Meter GC 822 Schott, Mainz, Deutschland
Protein-Blotkammer Fastblot B44 Biometra, Göttingen,
Deutschland
Sauerstoff-Elektrode: Elektrode und Kontrollbox
Schreiber
BD 40 Autofocus 3000 Hansatech, Norfolk,
England
Kipp & Zonen, Delft,
Niederlande,
Spektralphotometer Smart spec 3000
NanoDrop ND-1000
Biorad, Hercules, Ca, USA
Peqlab, Erlangen,
Deutschland
Ultraschallgerät Sonifier 250 Branson, Danbury, CT,
USA
Wasserbäder SW-20C
(Schüttelwasserbad)
Julabo, Seelbach,
Deutschland
Zentrifugen
Mini Spin
5417 R
5810 R
L8-M
Sorvall RC-5B
Eppendorf, Hamburg,
Deutschland
Eppendorf, Hamburg,
Deutschland
Eppendorf, Hamburg,
Deutschland
Beckmann Coulter, Krefeld,
Deutschland
DuPont Instruments,
Wilmington, DE, USA

Lebenslauf
85
9 Lebenslauf
Persönliche Daten
Name Jessica Jacobs Geburtsdatum 22.06.1981 Geburtsort Wuppertal Staatsangehörigkeit deutsch Familienstand ledig
Hochschulstudium
12/2005 bis 10/2009 Promotionsstudium an der Ruhr-Universität Bochum am LS Biochemie der Pflanzen, AG Photobiotechnologie, Thema der Dissertation: „Biochemische und funktionelle Charakterisierung eines anaerob induzierten Ferredoxins aus der Grünalge Chlamydomonas reinhardtii“
10/2000 bis 09/2005 Biologiestudium an der Ruhr-Universität Bochum, Thema der Diplomarbeit: „Der anaerobe Stoffwechsel von Chlamydomonas reinhardtii: Neue Erkenntnisse mit Real-Time PCR und heterologer Expression“ (Gesamtnote: 1,1)
Schulausbildung
09/1991-06/2000 Gymnasium Sedanstraße, Wuppertal, Abschluss mit Abitur (Gesamtnote: 2,3), Schwerpunkte: Biologie, Deutsch, Mathematik und Geographie
09/1987-07/1991 Grundschule Alarichstraße, Wuppertal Publikationen
Jacobs J., Pudollek S., Hemschemeier A. and Happe T. (2009) A novel, anaerobically induced ferredoxin in Chlamydomonas reinhardtii. FEBS Lett. 583(2):325-9. Epub 2008 Dec 26
Hemschemeier A., Jacobs J. and Happe T. (2008), Biochemical and Physiological Characterization of the Pyruvate Formate-Lyase Pfl1 of Chlamydomonas reinhardtii, a Typically Bacterial Enzyme in a Eukaryotic Alga. Eukaryotic Cell, March 2008, p. 518-526,
Vol. 7, No. 3

Lebenslauf
86
Konferenzbeiträge
Jacobs J., Lambertz C., Hemschemeier A. and Happe T. (2009) The novel anaerobically induced ferredoxin of Chlamydomonas reinhardtii. Jahrestagung der VAAM, Bochum,
Deutschland. Poster
Jacobs J., Hemschemeier A. and Happe T. (2008) The novel anaerobically induced ferredoxin of Chlamydomonas reinhardtii. 17. Photosynthese-Workshop Nord-West,
Bochum. Poster
Jacobs J., Hemschemeier A. and Happe T. (2008) The novel anaerobically induced ferredoxin of Chlamydomonas reinhardtii. 13th International Conference on the Cell and Molecular Biology of Chlamydomonas, Hyeres, Frankreich. Poster
Jacobs J., Hemschemeier A. and Happe T. (2008) The novel anaerobically induced ferredoxin of Chlamydomonas reinhardtii. Molecular Functions of Botanical Systems,
Bochum, Deutschland. Vortrag Jacobs J., Hemschemeier A., Winkler M., Lambertz C., Kamp C., Stripp S. and Happe T. (2007) The novel anaerobically induced ferredoxin of Chlamydomonas reinhardtii. SFB 480
Workshop, Velen, Deutschland. Vortrag
Jacobs J., Hemschemeier A. and Happe T. (2007) A novel anaerobically induced ferredoxin is involved in photosynthetic hydrogen production. The 8th International Hydrogenase
Conference, Breckenridge, USA, Poster
Hemschemeier A., Jacobs J., Rühle T., v. Abendroth G., Kamp C., Winkler M. and Happe T. (2007) The anaerobic life of Chlamydomonas reinhardtii – uncovering the molecular tools of algal hydrogen production. Solar H Meeting, Uppsala, Schweden. Vortrag Es wurde kein „Book of Abstracts“ veröffentlicht.
Jacobs J., Hemschemeier A. and Happe T. (2007) A novel anaerobically induced ferredoxin is involved in photosynthetic hydrogen production. 16. Photosynthese-Workshop Nord-West,
Mühlheim, Deutschland. Vortrag
Jacobs, J., Hemschemeier, A., and T. Happe (2006) Analysis of novel genes and proteins that are expressed under sulphur deprivation and anaerobic conditions. 12 th International
Conference on the Cell and Molecular Biology of Chlamydomonas, Portland, USA, Poster
Hemschemeier A., Jacobs J., Kamp C. and Happe T. (2006) [Fe]-hydrogenases and H2-metabolism in the unicellular green alga Chlamydomonas reinhardtii. Solar H Meeting,
Gelsenkirchen, Deutschland, Vortrag

87
Danksagungen
An dieser Stelle möchte ich mich herzlich bei allen Menschen bedanken, die mich im Laufe
meiner Promotion unterstützt haben.
Ein besonderer Dank gilt:
o Meinem akademischen Lehrer und Doktorvater Prof. Dr. Thomas Happe für die
interessante Themenstellung, das mir entgegengebrachte Vertrauen, sein stetiges
Interesse an dieser Arbeit und die damit verbundenen wertvollen wissenschaftlichen
Ratschläge.
o Prof. Dr. Elmar Weiler für die freundliche Übernahme des Korreferates.
o Dr. Anja Hemschemeier für die ständige Diskussionsbereitschaft und viele gute Ideen.
o Camilla Lambertz, mit der ich nicht nur viele wissenschafliche Gespräche geführt
habe, sondern auch ausserhalb des Labors viel Spaß hatte.
o Meinen Büronachbarn Sven Stripp und Miriam Pape.
o Allen weiteren Mitgliedern der AG Photobiotechnologie für das angenehme
Arbeitsklima, die tolle Zusammenarbeit, nette Gespräche und die vielen Feiern und
Grillabende.
o Natürlich auch meinen Eltern und Hubert, die immer für mich da waren.

88
Erklärung
Hiermit erkläre ich, dass ich die Arbeit selbständig verfasst und bei keiner anderen Fakultät
eingereicht habe und dass ich keine anderen als die angegebenen Hilfsmittel verwendet habe.
Es handelt sich bei der heute von mir eingereichten Dissertation um fünf in Wort und Bild
völlig übereinstimmende Exemplare.
Weiterhin erkläre ich, dass digitale Abbildungen nur die originalen Daten enthalten und in
keinem Fall inhaltsverändernde Bildbearbeitung vorgenommen wurde.
Bochum, den 01.07.2009
______________________________
Jessica Jacobs





























