Embed Size (px)
DESCRIPTION
Biosinteza
Citation preview

19.04.23 1
BIOSINTEZA ADNTRANSLAŢIA
Dana Liana DavidCatedra Biochimie
UMF “V. Babeş“Timişoara

19.04.23 2
Biosinteza ADN
Elementele procesului de translaţieARNm
– transportă informaţia de la genă la ribozomi participând direct la procesul de sinteză proteică.
– Informaţia transportată este codificată sub forma codonilor .
– Fiecare codon va dirija locul şi felul fiecărui AA din viitoare proteină.
ARNt - recunoaşte, leagă, activează şi transportă AA din citoplasmă
la complexul de sinteză. − transportă un singur tip de AA, dar un singur AA poate fi
transportat de mai multe tipuri de ARNt. La om s-au identificat 32 ARNt.
− Anticodonul - complementar cu codonii din ARNm. ARNr- orientarea moleculelor ARNt, - fixare şi derularea ARNm.

19.04.23 3
Ribozomii
* sediul sintezei proteice. - independenţi – sintetizează proteine de structură- legaţi de RE, formează RER – sintetizează proteine de
secreţie.- constituit din 2 subunităţi : - subunitatea mică (30 S la procariote, 40 S la
eucariote) - subunitatea mare (50 S la procariote, 60 S la
eucariote).Subunitatea mare prezintă 3 loci:
- aminoacil (A), - peptidil (P) - eliminare (E), ataşaţi ARNt. - Peptidiltranferaza - tranfera un ARNt dintr-un
locus în altul. Subunitatea mică − fixează ARNm− fixarea este laxă, permiţând derularea acestuia şi
incadrarea codonilor din secvenţa ARNm în locii subunitatii mari.

19.04.23 4
Etapele biosintezei
- aceleaşi etape la procariote cât şi la eucariote.• Activarea aminoacizilor din citoplasmă şi formarea
complexelor aminoacil –ARNt• Iniţierea sintezei proteice• Alungirea catenei polipeptidice• Terminarea sintezei şi eliberare proteinei

19.04.23 5
1. Activarea AA
Citoplasmă, • aminoacil-ARNt- sintetazaFormarea complexului aminoacil-ARNt: - proces endergonic, energie furnizată de hidroliza a 2
legături macroergice ATP. - Legătura ARNt – AA este macroergică (7kcal/mol). - Pentru fiecare AA există un ARNt specific ce expune
un anticodon specific.

19.04.23 6
2. Iniţierea sintezei proteice
• AA ce iniţiază sinteza:• întotdeauna formil – metionină la procariote • metionină - la eucariote. • se leagă de un ARNt special (ARNt de iniţiere),
recunoscut de un factor de iniţiere specific, IF2. • În afara procesului de biosinteză ribozomii se găsesc
sub formă disociată a celor două subunităţi, reasocierea lor fiind realizată de către ARNm.

19.04.23 7
2. Iniţierea sintezei proteice - formarea complexului de preiniţiere
Factorii de iniţiere: • IF1 leagă subunitatea mare 50S;• IF2 recunoaşte şi leagă ARNt- formil-Met;• IF3 leagă subunitatea mică 30S de ARNm la capătul
secvenţei Shine-Dalgano

19.04.23 8
3. Elongarea • În locusul aminoacil (A) liber din complexul de
iniţiere se va fixa prin complementaritate complexul ARNt - AA 2, al cărui anticodon este complementar codonului ARNm încadrat de locul aminoacil.
• Fixarea - factor de elongare (EF-Tu) şi prin hidroliza 1 mol GTP.
• peptidiltransferaza hidrolizează AA din locusul peptidil (P) şi îl leagă de AA 2 din locusul aminoacil (A) printr-o legătură peptidică.
• ARNt liber în poziţia P şi un dipeptid (H2N - formil-Met – AA 2) în poziţia A.
• formil – Met va constitui capătul N terminal al viitoare proteine.
• sub acţiunea unui factor de elongare EF-G (translocază) şi cu consumul unei molecule GTP are loc mişcarea ribozomului (translocarea) de-a lungul ARNm pe direcţia 5’ → 3’, pe o distanţa de 3 nucleotide (un codon).
• ARNt liber din poziţia P ajunge în poziţia E de unde va fi eliberat.
• ARNt-dipeptid din poziţia A ajunge în poziţia P.

19.04.23 9
3. Elongarea
• În poziţia A (liberă) este încadrat următorul codon pe direcţia 5’ → 3’ din ARNm.
• Procesul se repetă, urmând aceleaşi etape, fiecărui codon din sectorul codant al ARNm fiindu-i ataşat câte un AA transportat de ARNt cu anticodon complementar.

19.04.23 10
4. Terminarea sintezei şi eliberarea proteinei
• apariţia în secvenţa ARNm ce ajunge la citire in locusul A a unor codoni non-sens (UAA, UAG, UGA).
• 3 factori de eliberare:• RF1 şi RF2 recunosc codonii stop • RF3 şi o moleculă GTP eliberează proteina şi
disociază complexul de iniţiere în elementele componente

19.04.23 11
Diferenţe ale procesului de translaţie la eucariote
• Primul AA, din complexul de iniţiere al translaţiei este metionina.
• ARNm nu are secvenţa de recunoaştere Shine-Dalgano pentru fixarea ribozomilor
• Ribozomul începe translaţia la codonul AUG, localizat într-o secvenţă specifică = secvenţă de consens Kozak (CAAAAUG).
• Factorii de iniţiere, eiF - 6.

19.04.23 12
Energetica procesului de translaţie
• Legarea unui AA printr-o legătură peptidică necesită consumul a 4 legături macroergice:
- 2 - activarea AA şi formarea complexului aminoacil-ARNt,
- 2 - în procesul de elongare.

19.04.23 13
Efectul Wobble
• Legarea complementară codon – ARNm – anticodon - ARNt nu este perfectă.
• obligatorie complementaritatea doar a primelor două nucleotide din codon, pentru cea de-a 3-a există mai multe variante.
• cuplat cu degenerarea codului genetic, având ca scop limitarea efectului mutaţiilor.
• dacă mutaţia afectează doar a 3-a nucleotidă dintr-un codon ARNm, ea nu va avea repercursiuni asupra structurii proteice.

19.04.23 14
Fidelitatea procesului de translatie
Rata de eroare:• măsurarea frecvenţei de incorporare a unui AA în
structura unei proteine ce în mod normal nu conţine acel AA în secvenţă.
• un AA greşit încorporat la 104 AA încorporaţi corect, conform secvenţei ARNm,
• 1 din 25 molecule proteice de mărime medie (400 AA) poate să conţină o eroare de sinteză.
Fidelitatea sintezei proteice depinde de acurateţea celor două mecanisme de adaptare:
• legarea fiecărui AA de molecula de ARNt corespunzătoare
• complementaritatea perechilor de baze din codonii ARNm cu anticodonii ARNt.
Celulele si-au dezvoltat mecanisme de reparare “proofreading” pentru a reduce numărul de erori din aceste două etape cruciale ale sintezei proteice.

19.04.23 15
Realizarea conformaţiei finale a proteinelor (folding)
- sinteza proteinelor mari - adoptarea conformaţiei - proces asistat.
- şocul termic - asupra celulelor cresc - proteinele de şoc termic (Hsp = Heat shock proteins).
- identice cu proteinele ce asistă adoptarea conformaţiei corecte la sinteza proteinelor mari.
- Hsp70 şi Hsp 60 au fost denumite Chaperone (proteine de asistenţă).

19.04.23 16
Realizarea conformaţiei finale a proteinelor (folding)
• Proteina în curs de sinteză, începând cu capătul N-terminal, interacţionează cu proteine citoplasmatice.
• Chaperonele fixează capătul N-terminal şi proteina în sinteză, ca într-o cuşcă de protecţie ferind proteina de interacţiuni,
• Proteina îşi dezvoltă conformaţia corectă dictată de structura primară.
Proces asistat de folding – 2 enzime:- Protein-disulfit–izomeraza(PDI) - rupe punţile S-S
incorecte şi le reface în locurile corecte. - Concentraţia crescută în RE - sintetizate
proteinele de secreţie.- Peptidil-prolin-izomeraza(PPI) - aranjează resturile de
prolină în poziţiile cis şi trans.

19.04.23 17
Dirijarea şi secreţia proteică
Când lungimea polipeptidului în curs de sintetizare in complexul ribozom - ARNm ajunge la aproximativ 30 AA, capătul său N-terminal:
1. Importul cotranslaţional. - caracteristic proteinelor de secreţie.
2. Importul posttranslaţional. - proteinelor de membrană. - dacă polipeptidul este

19.04.23 18
Modificări posttranslaţionale
1. Modificari chimice, de mărime şi conformaţie ale catenei polipeptidice.
Prokariote, - N-formil - MET înlăturată întotdeauna din proteina
matură. Eucariote, - MET şi AA N-terminali sunt îndepărtaţi din structura
proteinei finale.2. Scindări ale catenei polipeptidice, - se elimină
fragmente ce menţineau proteina în stare inactivă. ex. -sinteza insulinei: sintetizată iniţial - preproinsulină.
este scindată succesiv în – proinsulina, apoi insulina.3. Procesul de „splicing” proteic (analog cu splicing-ul
ARN), - înlătură inteinele (analogi proteici ai intronilor din
ARNm) - ataşează exteinele (analogi proteici ai exonilor din
ARNm) - realizarea secvenţa proteinei mature.

19.04.23 19
Modificări posttranslaţionale
4. Modificări chimice ale AA - activatoare sau inhibitorii ale activităţii biologice.
- enzimatice/ neenzimatice- reversibile / ireversibileModificări neenzimatice• Racemizarea AA, trecând din forma L în D• Glicarea (glicozilarea) - legarea neenzimatică printr-
o legătură ceto-amidică a glucozei de grupările amino ale proteinelor plasmatice.Ex. glicozilarea hemoglobinei .
• Carbamilarea - realizată de ureea din sânge asupra proteinelor plasmatice.
• Oxidarea catenelor laterale ale AA din proteine. afectează proteinele parţial distruse şi nefuncţionale de la persoanele în vârstă.

19.04.23 20
Modificări posttranslaţionale
Modificări enzimatice 1. Ataşarea de resturi glucidice → glicoproteine - La eucariote - mai ales proteinele de secreţie şi
proteinele membranare - asigură:• realizarea structurilor necesare pentru funcţia
biologică • protecţie împotriva proteazelor Rezultă 2 tipuri de glicozilări :a) N-glicolizarea – modificare cotranslaţionalăb) O-glicolizarea – modificare posttranlaţională2. Acilarea proteinelor - acilarea grupărilor amino cu
acizi graşi (miristic, palmitic).3. Formarea de legături intra sau inter – catenare -
punţi de S, punţi ditirozină, carboxilarea Glu, iodinarea Tir .

19.04.23 21
Expresia genică
• procesul prin care o genă duce la sinteza unei proteine. • o genă nu este tradusă direct într-o proteină ci este
exprimată prin formarea unui compus intermediar numit ARN mesager.
- regiune codantă = porţiunea din ARNm care corespunde secvenţei AA din proteină
- secvenţe adiţionale netranslatabile, la fiecare capăt 3’ si 5’, care nu codifică AA.
- mutaţiile regiunii terminale determină alterarea expresiei genice ceea ce demonstrează rolul lor indirect în exprimarea genei.
Proteinele sunt considerate produsul final al expresiei genelor.
Controlul sintezei proteice = controlul expresiei genelor. - nivelul tuturor etapelor procesului: genă ARNm
proteine. - majoritar la nivelul transcripţiei - procariote - tip negativ (inhibiţie) - eucariote - tip pozitiv (activare).

19.04.23 22
Controlul expresiei genelor la procarioteteoria operon
nivelul transcripţiei. • Jacob şi Monod (1962) - teoria operon - model de
reglare a biosintezei proteice - represia enzimatică - porneşte de la premiza existenţei mai multor tipuri de gene grupate intr-un cluster numit operon.
• mecanism de reglare : inducţia şi represia.Modelul admite:• Э genei reglator – controlează gena structurală prin
intermediul unei proteine, a cărui sinteză o specifică = represor.
• Activarea represorului inhibă sinteza ARNm → inhibarea translaţiei
• Inactivarea represorului permite transcripţia genei structurale controlate → sinteză proteică

19.04.23 23
Controlul expresiei genelor la procarioteteoria operon
- Legarea represor – operon → bloc. ARN-polimerazei la promotor → suprimarea transcripţiei genei structurale
- Abilitatea represorului de a se lega ↔ operator/inductor este responsabilă de represia sau inducţia enzimatică.
- culturi de E.coli crescute într-un mediu fără lactoză nu secretă lactază

19.04.23 24
Controlul expresiei genelor la procarioteteoria operon
Concluzii: • informaţia (gena) pentru lactază există în genomul E.
coli, dar în lipsa lactozei expresia ei este inhibată.• În cromozomul procariot, genele ce contribuie la
aceiaşi cale metabolică sunt situate împreună, fiind reglate împreună.
• segmentul de genă corespunzător sintezei unei catene polipeptidice se numeşte cistron.
ex. operonul pentru TRP - conţine 5 gene structurale necesare sintezei a trei enzime implicate în sinteza TRP.
• Operonul este controlat de o proteină supresor, ce devine activă doar la legarea de TRP şi blochează transcripţia prin fixarea de situsul operator.

19.04.23 25
Controlul expresiei genelor la eucariote
complex:- cantităţii mult mai mari de ADN, organizării superioare,
localizare nucleara , T ½ al ARNm mult mai lung. - doar 2-5% din ADN este transcris sub formă ARNm. Reglarea:• activarea genelor cu proteine activatoare• reglare pozitiva - mai avantajos să activezi 2% din
materialul genetic decât să inhibi 98%.Diversitatea celulară nu este produsul pierderii unor gene
în cursul diferenţierii celulare ci al modificărilor expresiei genelor.
Proteinele = reflexia finală expresiei genelor, clasificate:• proteine comune tuturor celulelor din organism: proteine
structurale din cromozomi ARN polimeraze, enzime de reparaţie ale ADN, proteinele ribozomale, proteine din citoschelet,etc.
• proteine caracteristice unor anumite ţesuturi. Ex. Hb se găseşte doar în seria eritrocitară.
Modificările posttranscripţionale şi posttranslaţionale: - 1 genă să genereze o întreagă familie de proteine.

19.04.23 26
Controlul expresiei genelor la eucariote
Reglarea - 2 tipuri de mecanisme:1) genetic pe termen scurt (reversibil) - se modifică
activitatea unor gene fapt exprimat prin fluctuaţii în sinteza de ADN, ARN şi proteine.
2) genetic pe termen lung (ireversibil) - implică mecanisme legate de diferenţierea celulară şi care au loc în cursul dezvoltării ontogenice.
- codificat în secvenţa ADN.Nivele de control ale expresiei genelor:• controlul transcripţional - reglează unde, când şi cât
este realizată transcrierea genei.• controlul procesării ARN• controlul transportului şi localizării ARN• controlul translaţional.• controlul degradării ARNm.• controlul activităţii proteinelor.
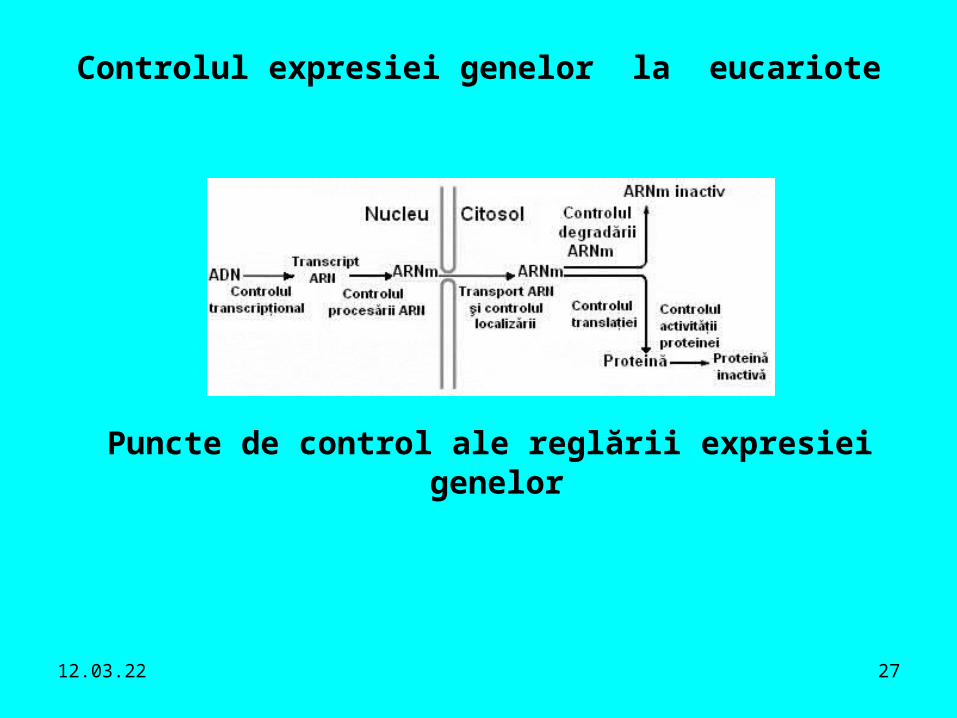
19.04.23 27
Controlul expresiei genelor la eucariote
Puncte de control ale reglării expresiei genelor

19.04.23 28
I. Controlul la nivelul transcriptional – expresie genica
• forma predominantă 1. Represia transcriptiei - esentială - proteine represoare actioneaza asupra genelor ţintă:• direct, prin domeniile de legare a ADN, • indirect prin interactiunea cu proteinele legate de
ADN . - mascarea unui domeniu de activare a
transcriptiei, - blochează interacţiunea dintre un activator si
alte elemente ale complexului de transcripţie, - îndepartează un activator de ADN.

19.04.23 29
I. Controlul la nivelul transcriptional
Metilarea ADN interfera cu procesul de transcriptie la nivelul rest CIT din
segmentul promotor, necesare legarii factorilor de transcriptie.
Cromatina activă transcriptional este nemetilată. Factorii de transcriptie cis au situsuri de legare bogate in
GC - metilarea resturilor de CIT va anula capacitatea de legare a acestora si implicit va bloca transcriptia.
• ADN metiltransferaze .• predominant in regiunile repetitive, cum ar fi ADN satelit
sau elemente transpozabile (LINES, SINES). • Promotorii genelor sunt nemetilate asigurând
transcriptia acestora.• Reducerea metilarii regiunilor repetitive creează
instabilitate genomică favorizând aparitia de mutatii, in timp ce metilarea din promotorii genelor inhiba transcripţia.
• Daca acest proces inhiba transcriptia genelor supresoare de tumori, atunci se creează condiţii de dezvoltare a procesului neoplazic.

19.04.23 30
I. Controlul la nivelul transcriptional
Metilarea ADN şi cancerul metilarea – demetilarea ADN, - genomul cc – hipometilat → instabilitate
genomică → reactivarea promotorilor transpozomilor → reglare aberantă prin interferenţă transcriptională/transcripţii antisens.
- demetilarea AND - marker precoce severitate / stadializare / metastaze
Hipermetilarea aberantă a genomului → modificări de structură a cromatinei → inhibarea transcripţiei genei respective.

19.04.23 31
Controlul vitezei de iniţiere al transcripţiei
• factorii amplificatori sau inhibitori ce cresc/reduc viteza de transcripţie prin formare complexului de iniţiere.
• reglarea prin fosforilare - defosforilare a unor factori de iniţiere a translaţiei , IF-2.
• acţiune de răspuns faţă de diferiţi factori de mediu (temperatură , factori de creştere sau alimentari, infecţii etc.)

19.04.23 32
Splicing alternativ
• Cel puţin 1/3 genele umane produc proteine multiple utilizând acest mecanism.
• Reglarea splicing-ului ARN generează diferite versiuni ale aceleiaşi proteine în diferite ţesuturi.
• Reglarea se face pe baza competivităţii situsurilor de splicing obţinută prin acţiunea proteinelor reglatoare.

19.04.23 33
Reglarea transportului ARN din nucleu în
citoplasmă
• 1/20 din totalul ARN sintetizat în nucleu trece în citoplasmă.
• ARN este degradat în nucleu de un complex proteic numit exozom
• proces incomplet de splicing.

34
II. Controlul la nivel translational
Etape:1. Blocarea situsului de start al translaţiei de către
proteine supresoare, reglate de efectul fiziologic al proteinei translatate.
2. Centre interne multiple ale ARNm pentru iniţierea sintezei proteice.
- mecanism viral, blocheză sinteza proteică a gazdei, orientează sinteza proteinelor virale
3. Controlul stabilităţii ARNm. - degradarea ARN prin scurtarea cozii de poliA, - îndepărtarea protecţiei de la capătul 5’ - ARNm este
degradat rapid. - prelungirea vietii ARNm - lungirea poliA cu noi
unităţi. 4. Controlul factorilor de translaţie (factori de iniţiere,
elongare si terminare). - blocaţi/activaţi prin acţiunea unor inhibitori,
fosforilare/defosforilare.

19.04.23 35
III. Controlul expresiei genelor prin molecule ARN
• 2% din ADN-ul uman codifică proteine • 98% este implicat în reglarea genelor din cei 2%. • 98% din ARN obţinut prin transcripţie provine din
zona necodantă pentru proteine. Funcţiile acestor tipuri de ARN :• legare şi blocare a ARNm (inhibitori translaţie).• legare şi blocare a ADN (control transcripţie)• legare şi blocare proteine (blocare transcripţie)Aceste procese sunt incluse în termenul genetic de
interferenţă ARN. - 2 tipuri de ARN:• micro ARN• ARN scurt de interferenţă sau neutralizare a ARN
(SiRNA)

19.04.23 36
Inhibitori naturali si chemoterapeutici ai expresiei genelor
A. Inhibitori ai transcripţiei • inhibitori ai ARN polimerazei. Ex. ARN polimeraza de la procariote este inhibată de
rifampicină, care datorită acestei propietăţi este utilizată ca antibiotic selectiv, tratamentul TBC blocând dezvoltarea Mycobacterium tuberculosis. Avantaj- toxicitatea redusă.
-amanitina este o toxină din ciuperca Amantina phaloides, care blochează selectiv ARN polimeraza II (ce sintetizează ARNm) de la eucariote. Din acest motiv produce intoxicaţii grave la om.
B. Inhibitori ai translaţiei• inhibitori ai sintezei proteice la nivel ribozomal
(substanţe antibiotice) sunt utilizate microorganisme ca arme chimice împotriva competitorilor sau paraziţilor.
Ex- mucegaiul Penicilium produce inhibitori antibacterieni.
• Datorită diferenţelor structurale între ribozomii de la procariote şi eucariote, aceste antibiotice sunt selective fie pentru procariote (streptomicina, tetraciclină, eritomicină), fie pentru eucariote (cicloheximidă, puromicină).

19.04.23 37
Inhibitori naturali si chemoterapeutici ai expresiei genelor
• Aminoglicozidele (streptomicină, neomicină, tobramicină, gentamicină, amikacină) se leagă ireversibil de subunitatea 30 S a ribozomilor bacterieni, blocând activitatea peptidil transferazei
• Tetraciclinele (tetraciclină, doxiciclină, demeclociclină) se leagă reversibil la subunitatea 30 S, inducând modificări conformaţionale ce împiedică alinierea codonilor ARNm cu anticodonii ARNt.
• Macrolidele (eritromicină, azitromicină, claritromicină) se leagă reversibil de subunitatea 50 S şi inhibă activitatea peptidil transferazei.
• Streptograminele (quimpristină, dalfopristină) împiedică legarea ARNt de subunitatea 50 S.
• În cazul bacteriilor cu creştere rapidă blocarea sintezei proteice va bloca dezvoltare microorganismelor.
• toxina difterică, proteină produsă de microorganismul Corynebacterium difterie. blochează factorul de elongare EF-2 (translocază) de la eucariote, blocând astfel regenerare celulară.