Embed Size (px)
Citation preview

BIOSYNTHESIS OF ESTRADIOL Cloning and characterization of rodent 17β-hydroxysteroid dehydrogenase/17-ketosteroid reductase types 1 and 7
PASINOKELAINEN
Biocenter Oulu andWHO Collaborating Centre for
Research on Reproductive Health
OULU 2000

BIOSYNTHESIS OF ESTRADIOL Cloning and characterization of rodent 17β-hydroxysteroid dehydrogenase/17-ketosteroid reductase types 1 and 7
PASI NOKELAINEN
Academic Dissertation to be presented with the assent of the Faculty of Medicine, University of Oulu, for public discussion in Auditorium 9 of the University Hospital of Oulu, on September 29th, 2000, at 12 noon.
OULUN YLIOPISTO, OULU 2000

Copyright © 2000Oulu University Library, 2000
Manuscript received 16 August 2000Accepted 21 August 2000
Communicated by Professor Heikki Ruskoaho Professor Timo Ylikomi
ALSO AVAILABLE IN PRINTED FORMAT
ISBN 951-42-5751-0
ISBN 951-42-5750-2ISSN 0355-3221 (URL: http://herkules.oulu.fi/issn03553221/)
OULU UNIVERSITY LIBRARYOULU 2000

Nokelainen, Pasi, Biosynthesis of estradiol. Cloning and characterization of rodent17ββββ-hydroxysteroid dehydrogenase/17-ketosteroid reductase types 1 and 7Biocenter Oulu and WHO Collaborating Centre for Research on ReproductiveHealth, P.O.Box 5000, FIN-90014 University of Oulu, Finland2000Oulu, Finland(Manuscript received 16 August 2000)
Abstract
17β-Hydroxysteroid dehydrogenases (17HSDs)/17-ketosteroid reductases (17KSRs) modulate thebiological activity of certain estrogens and androgens by catalyzing dehydrogenase and reductasereactions between 17β-hydroxy and 17-ketosteroids.
In the present study, cDNAs encoding mouse and rat 17HSD/KSR1 were cloned in order tostudy the role of rodent type 1 enzyme in ovarian estradiol (E2) biosynthesis and its enzymaticcharacteristics. Both rat and mouse 17HSD/KSR1 were expressed in granulosa cells of developingfollicles, where diethylstilbestrol and follicle-stimulating hormone stimulated follicular maturationand up-regulated the expression of 17HSD/KSR1, whereas human chorionic gonadotropin causedluteinization of follicles and down-regulation of the enzyme. In line with this, the rodent type 1enzymes are not expressed in the corpus luteum (CL). Mouse 17HSD/KSR1 showed substratespecificity different from that of the human counterpart. The mouse type 1 enzyme catalyzed thereaction from androstenedione to testosterone at least as efficiently as estrone (E1) to E2, whilehuman 17HSD/KSR1 clearly preferred the E1 to E2 reaction.
A mouse mammary epithelial cell line was found to possess strong estrogenic 17KSR activity.A novel type of 17HSD/KSR responsible for this activity was expression-cloned on the basis of itsability to convert E1 to E2 and it was chronologically named 17HSD/KSR7. Interestingly, it showed89 % identity with a rat protein called prolactin receptor-associated protein (PRAP), which isexpressed in the CL. Enzymatic characterization showed that both mouse 17HSD/KSR7 and PRAPefficiently catalyzed the reaction from E1 to E2. The mouse type 7 enzyme was most abundantlyexpressed in the ovary and placenta. Similar primary structure, enzymatic characteristics, and tissuedistribution of mouse 17HSD/KSR7 and PRAP suggest that PRAP is rat 17HSD/KSR7.
Further studies showed that in rat ovaries 17HSD/KSR7 is primarily expressed in the middleand second half of pregnancy, in parallel with E2 secretion from the CL. Using in situ hybridization,cell-specific expression of 17HSD/KSR7 was studied in the mouse ovary, uterus and placenta. Inthe mouse ovary, the enzyme was expressed exclusively in the CL. In the uterus on day 5½ postcoitum (p.c.), the type 7 enzyme was expressed in the decidua, mostly in the inner zone ofantimesometrial decidua. Between day 8 and 9½ p.c. the enzyme was abundant in deciduacapsularis of the developing placenta, after which expression moved to the basal zone. On days12½ and 14½ p.c., mouse type 7 enzyme was abundantly expressed in the spongiotrophoblasts,where expression decreased towards parturition. Altogether, rodent 17HSD/KSR7 is a new17HSD/KSR which is involved in the biosynthesis of E2 in the ovaries. In addition, E2 producedlocally in the decidua and placenta by the type 7 enzyme may have a role in decidualization and/orimplantation and placentation.
Keywords: steroidogenesis, estrogen, ovary, placenta

Acknowledgements
The present study was carried out at the World Health Organization Collaborating Centre(WHOCCR) for Research on Reproductive Health, University of Oulu, Finland.
I wish to express my deepest gratitude to my supervisors, first to the President of theAcademy of Finland, Professor Reijo Vihko, M.D., Ph.D., who introduced me to thisinteresting field of molecular endocrinology and provided excellent research facilities,and thereafter to Professor Pirkko Vihko, M.D., Ph.D., who after Reijo had left hisposition has shared her knowledge and given support and encouragement. I also owe mygratitude to my additional supervisor, Docent Hellevi Peltoketo, Ph.D., for her devotedand patient tutoring and contribution to my project, for her friendship and for valuablecomments on the manuscript of this thesis. In addition, I would like to extend myappreciation to Mauri Orava, Ph.D., who introduced me to the secrets of clonings at thebeginning of this study.
I am grateful to Professor Heikki Ruskoaho, M.D., Ph.D., and Professor TimoYlikomi, M.D., Ph.D., for their constructive criticism on the manuscript of this thesis. Ialso wish to thank Docent Nicholas Bolton, Ph.D. for his careful revision of the languageof the manuscript and the original articles. I would like to extend my thanks to DocentVeli Isomaa, Ph.D., for his support in completing this thesis and for help in all practicalarrangements needed to attain the Ph.D. degree.
I am also indebted to all my co-authors Professor Matti Poutanen, Ph.D., ProfessorHannu Rajaniemi, M.D., Ph.D., Docent Helena Autio-Harmainen, M.D., Ph.D., SergioGhersevich, Ph.D., Mika Mustonen, Ph.D., and Terhi Puranen, Ph.D., for their valuablecontribution to this work.
I wish to extend my sincere thanks to all my co-workers at WHOCCR for sharing theirknowledge and expertise. Especially I am grateful to Ms. Minna Eskelinen, whoseoutstanding and invaluable technical assistance and sincere friendship have been of greatimportance. The skillful technical assistance of Ms. Eeva Holopainen, Ms. Liisa Kaarelaand Ms. Helmi Konola is also gratefully acknowledged. I am also grateful to AnjaRaatikainen and Sirpa Annanperä for their skillful secretarial assistance and help in manydaily problems. In addition, special thanks are reserved for Juhani and Jaakko Heikkiläfor their invaluable help with computers.

I wish to express my warmest thanks to my mother, my parents- and sister-in-law andall my friends, who have given solid support and encouragement throughout these yearsand shared the interesting life outside science.
Finally, I owe my sincerest appreciation to my wife, Minna, for her love, unfailingsupport and patience during these years. I am also grateful for her efficiency in finalizingher doctoral thesis and the practical arrangements, which gave me a specialencouragement to keep going toward my dissertation.
This study was mainly supported by Biocenter Oulu, Research Council for Health ofthe Academy of Finland, the Cancer Foundation of Northern Finland and the EmilAaltonen Foundation. The WHOCCR is supported by the Ministries of Education, SocialAffairs and Health, and Foreign Affairs, Finland.
Oulu, August 2000
Pasi Nokelainen

Abbreviations
Hormones and steroids:
16α-OH-DHEAS 16α-hydroxydehydroepiandrosterone sulfate17-OH-P 17α-hydroxy-4-pregnene-3,20-dione17-hydroxypregnenolone 3β,17α-dihydroxy-5-pregnen-20-one20-OH-P 20α-hydroxyprogesterone, 20α-hydroxy-4-pregnen-3-one3α-diol 5α-androstane-3α,17β-diol5α-A-dione 5α-androstane-3,17-dioneA-diol androstenediol, 5-androstene-3β,17β-diolA-dione androstenedione, 4-androstene-3,17-dioneADT androsterone, 3α-hydroxy-5α-androstan-17-oneapigenin 4,5,7-trihydroxyflavonecoumestrol 2-(2,4-dihydroxyphenyl)-6-hydroxy-3-benzofurancarboxylic
acid δ-lactoneDES diethylstilbestrolDHEA dehydroepiandrosterone, 3β-hydroxy-5-androsten-17-oneDHEAS dehydroepiandrosterone sulfateDHT dihydrotestosterone, 17β-hydroxy-5α-androstan-3-oneE1 estrone, 3-hydroxy-1,3,5(10)-estratriene-17-oneE2 estradiol, 1,3,5(10)-estratriene-3,17β-diolE3 estriol, 1,3,5(10)-estratriene-3,16α,17β-triolFSH follicle-stimulating hormonegenistein 4,5,7-trihydroxyisoflavonehCG human chorionic gonadotropinLH luteinizing hormoneP progesterone, 4-pregnene-3,20-dionePLP-B PRL-like protein-Bpregnenolone 3β-hydroxy-5-pregnen-20-onePRL prolactinT testosterone, 17β-hydroxy-4-androsten-3-one

Others:
17HSD/KSR 17β-hydroxysteroid dehydrogenase/17-ketosteroid reductase17HSD/KSR1–8 17HSD/KSR type 1–type 820α-HSD 20α-hydroxysteroid dehydrogenase3α-HSD 3α-hydroxysteroid dehydrogenase3β-HSD 3β-hydroxysteroid dehydrogenase/∆5-∆4isomeraseAF transcriptional activation functionAKR aldoketoreductaseATP adenosine-5’-triphosphateBSA bovine serum albumincAMP cyclic adenosine-3’,5’-monophosphatecDNA complementary DNACMV cytomegalovirusER estrogen receptorERE estrogen-responsive elementERKO estrogen receptor knockoutGAPDH glyceraldehyde-3-phosphate dehydrogenaseHEK human embryonic kidneyHPLC high performance liquid chromatographyHSD17B1–5 gene for 17HSD/KSR type 1–type 5HSD17BP1 17HSD/KSR1 pseudogenehsd17b1 rat 17HSD/KSR1 genekb kilobasekDa kilodaltonKe6 mouse 17HSD/KSR8 geneKm Michaelis-Menten constantMFP multifunctional proteinmRNA messenger RNANAD+ nicotinamide-adenine dinucleotideNADP+ nicotinamide-adenine dinucleotide phosphateP450arom cytochrome P450 aromataseP450c17 17α-hydroxylase/C17-20 lyaseP450scc cholesterol side chain cleavage enzymePBS phosphate-buffered salinep.c. post coitumPKA protein kinase APKC protein kinase CPR progesterone receptorPRAP PRL receptor-associated proteinRACE rapid amplification of cDNA endsRT-PCR reverse transcriptase PCRSDR short-chain dehydrogenase/reductaseSHBG sex hormone binding globulinVmax maximal velocityX any amino acid

List of original publications
This thesis is based on following articles, which are referred to in the text by their Romannumerals:
I Ghersevich S, Nokelainen P, Poutanen M, Orava M, Autio-Harmainen H,Rajaniemi H & Vihko R (1994) Rat 17β-hydroxysteroid dehydrogenasetype 1: Primary structure and regulation of enzyme expression in rat ovaryby diethylstilbestrol and gonadotropins in vivo. Endocrinology 135: 1477-1487.
II Nokelainen P, Puranen T, Peltoketo H, Orava M, Vihko P & Vihko R (1996)Molecular cloning of mouse 17β-hydroxysteroid dehydrogenase type 1 andcharacterization of enzyme activity. Eur J Biochem 236: 482-490.
III Nokelainen P, Peltoketo H, Vihko R & Vihko P (1998) Expression cloningof a novel estrogenic mouse 17β-hydroxysteroid dehydrogenase/17-ketosteroid reductase (m17HSD7), previously described as a prolactinreceptor associated protein (PRAP) in rat. Mol Endrocrinol 12: 1048-1059.
IV Nokelainen P, Peltoketo H, Mustonen M & Vihko P (2000) Expression ofmouse 17β-hydroxysteroid dehydrogenase/17-ketosteroid reductase type 7in the ovary, uterus and placenta: localization from implantation to latepregnancy. Endocrinology 141: 772-778.

Contents
AbstractAcknowledgementsAbbreviationsList of original publicationsContents1 Introduction................................................................................................................... 132 Review of the literature................................................................................................. 15
2.1 Physiological roles of estrogens ............................................................................. 152.2 Principles of estrogen action................................................................................... 16
2.2.1 Estrogen receptors ......................................................................................... 162.2.1.1 Domain structure of steroid hormone receptors ................................ 162.2.1.2 Estrogen receptors α and β ................................................................ 17
2.2.2 Molecular mechanisms of estrogen action..................................................... 182.3 Pathway of estrogen biosynthesis ........................................................................... 192.4 Ovarian physiology................................................................................................. 21
2.4.1 Follicle........................................................................................................... 212.4.1.1 Follicular maturation and luteinization.............................................. 212.4.1.2 Estradiol biosynthesis in the follicle and its role during
folliculogenesis ................................................................................. 222.4.2 Corpus luteum ............................................................................................... 23
2.4.2.1 Steroidogenesis and role of estradiol in the corpus luteum ............... 232.5 The fetoplacental unit as a sex hormone source ..................................................... 252.6 17β-Hydroxysteroid dehydrogenases (17HSDs)/17-ketosteroid reductases
(17KSRs)................................................................................................................ 262.6.1 The short-chain dehydrogenases/reductases family....................................... 292.6.2 17HSD/KSR type 1 ....................................................................................... 29
2.6.2.1 Structural and functional properties of human 17HSD/KSR1........... 292.6.2.2 Tissue distribution and role of human 17HSD/KSR1 in estradiol
biosynthesis....................................................................................... 312.6.2.3 Human 17HSD/KSR1 in normal mammary gland and in
breast cancer...................................................................................... 322.6.2.4 Structure of the human 17HSD/KSR1 gene ...................................... 32

2.6.2.5 Rat 17HSD/KSR1.............................................................................. 332.6.2.6 Regulation of 17HSD/KSR1 ............................................................. 34
2.6.3 17HSD/KSR type 2 ....................................................................................... 352.6.3.1 Human 17HSD/KSR2 ....................................................................... 352.6.3.2 Rodent 17HSD/KSR2........................................................................ 36
2.6.4 17HSD/KSR type 3 ....................................................................................... 372.6.5 17HSD/KSR type 4 ....................................................................................... 382.6.6 17HSD/KSR type 5 ....................................................................................... 402.6.7 17HSD/KSR type 6 ....................................................................................... 412.6.8 17HSD/KSR type 8 ....................................................................................... 412.6.9 Other 17HSD/KSRs....................................................................................... 42
3 Outlines of the present study......................................................................................... 444 Materials and methods .................................................................................................. 45
4.1 Isolation of RNAs (I-IV) ........................................................................................ 454.2 Cloning of rat and mouse 17HSD/KSR1 cDNAs (I, II) ......................................... 454.3 Cloning of mouse and rat 17HSD/KSR7 cDNAs (III) ........................................... 464.4 Sequencing and bioinformatics (I-IV) .................................................................... 464.5 Measurement of 17HSD/KSR activity of human and mouse 17HSD/KSR1 (II) ... 474.6 Measurement of 17HSD/KSR activity of mouse and rat 17HSD/KSR7 (III)......... 474.7 Northern blot analysis (I-III) .................................................................................. 484.8 Reverse transcriptase polymerase chain reaction (II, III) ....................................... 484.9 Animal treatments (I, II) ......................................................................................... 494.10 Tissue samples (I-IV) ........................................................................................... 494.11 Immunohistochemistry (I, II)................................................................................ 494.12 In situ hybridization (I, IV)................................................................................... 50
5 Results........................................................................................................................... 515.1 Cloning and characterization of rat and mouse 17HSD/KSR1 cDNAs (I, II) ........ 515.2 Expression of 17HSD/KSR1 in rat ovary and regulation by gonadotropins (I)...... 525.3 Characterization of 17HSD/KSR activity of mouse 17HSD/KSR1 and
comparison with the human enzyme (II)................................................................ 525.4 Tissue distribution of mouse 17HSD/KSR1 (II)..................................................... 535.5 Cloning and characterization of mouse 17HSD/KSR7 cDNA (III)........................ 535.6 Characterization of the enzymatic properties of mouse and rat 17HSD/KSR7 (III) .. 545.7 Tissue distribution of 17HSD/KSR7 and its expression pattern in the ovary
during gestation (III, IV) ........................................................................................ 545.8 Expression of mouse 17HSD/KSR7 in the uterus and placenta (IV)...................... 55
6 Discussion ..................................................................................................................... 566.1 Cloning and characterization of mouse and rat 17HSD/KSR1............................... 566.2 Expression and regulation of rat 17HSD/KSR1 in granulosa cells......................... 576.3 Cloning and characterization of mouse and rat 17HSD/KSR7, and its expression
pattern in the ovary................................................................................................. 576.4 17HSD/KSR7 in the mouse uterus and placenta .................................................... 59
7 Summary and conclusions............................................................................................. 608 References..................................................................................................................... 61

1 Introduction
Estrogens, female sex steroid hormones, not only affect reproduction but also participatein a large number of different functions outside the female reproductive system, such asbone and lipid metabolism and functions in the cardiovascular system. Estrogen action istransmitted through two nuclear estrogen receptors, estrogen receptor α (ERα) and ERβ,which are ligand-inducible transcription factors (Tsai & O'Malley 1994, Couse & Korach1999). They interact with transcription machinery to initiate transcription of target genesof estrogen action. Of natural human estrogens, estradiol (E2) is biologically the mostactive sex hormone, whereas estrone (E1) and estriol (E3) are less active, as a result oftheir lower affinity to ERs and decreased affinity of E1-ER and E3-ER complexes toestrogen-responsive elements (EREs) in target genes (Hisaw 1959, Martucci & Fishman1979, Kuiper et al. 1997, Melamed et al. 1997). In humans, E2 biosynthesis occurs at anumber of tissue sites. The principal sites are granulosa cells of the ovary inpremenopausal women and syncytiotrophoblasts of the placenta in pregnant women. Inaddition, estrogens are produced in peripheral tissues such as the mammary gland andadipose tissue, especially in postmenopausal women (Labrie 1991, Simpson et al. 1997,Peltoketo et al. 1999a). In rodents, E2 biosynthesis is thought not to take place in theplacenta but in the corpus luteum (CL) (Gibori et al. 1988 and references therein).
In the traditional endocrine system, estrogens are synthesized in the ovaries, releasedto the blood circulation and transported to target tissues (Speroff et al. 1994). In theblood, estrogens are bound with high affinity to sex hormone-binding globulin (SHBG)and with low affinity to albumin, as a consequence of which only 2 % of total E2 is free(Siiteri et al. 1982, Goldfien & Monroe 1991). Estrogens are released from their bindingproteins before they diffuse into the target cells. In addition to endocrine signaling, E2
produced in peripheral tissues can locally act in an autocrine, paracrine or intracrinemanner (Labrie 1991).
In the pathway of E2 biosynthesis, the last two reactions are catalyzed by cytochromeP450 aromatase (P450arom) and 17β-hydroxysteroid dehydrogenase/17-ketosteroidreductase (17HSD/KSR) enzymes. P450arom catalyzes conversion of androgens toestrogens (reviewed by Simpson et al. 1997), whereas 17HSD/KSRs catalyzeinterconversion between E1 and E2, and androstenedione (A-dione) and testosterone (T),for example (reviewed by Peltoketo et al. 1999a). When this work began, two

14
17HSD/KSRs were known, type 1 and type 2. Human 17HSD/KSR1 catalyzes thereaction from E1 to E2, while 17HSD/KSR2 mainly catalyzes reactions from E2 to E1, andT to A-dione. The type 2 enzyme is mostly expressed in the placenta, endometrium, liver,kidney and gastrointestinal tract, where it has been suggested to have a protective role,inactivating excess E2 (Miettinen et al. 1996a, Moghrabi et al. 1997, Mustonen et al.1998a, Mustonen et al. 1998b). Originally 17HSD/KSR1 was known to be expressed inthe placenta, ovary, mammary gland and endometrium, both normal and malignant.However, in the ovaries as well as in peripheral tissues, P450arom was considered to bethe important enzyme involved in E2 biosynthesis, while 17HSD/KSR1, though needed inE2 biosynthesis, was assumed to be of lesser importance and an enzyme whose expressionis not regulated.
The present study focuses on 17HSD/KSR enzymes participating in E2 biosynthesis.Role of 17HSD/KSR1 in E2 biosynthesis was studied by cloning rat and mouse17HSD/KSR1 cDNAs and characterizing expression and regulation of the enzyme inovaries. Enzymatic properties of the rodent 17HSD/KSR1 were determined using mouse17HSD/KSR1 expressed in insect cells and the results compared with the human enzyme.In further studies, a mouse mammary epithelial cell line was found to possess strongestrogenic 17KSR activity, profile of which did not match with mouse 17HSD/KSRsknown at that time. Thereby, expression cloning and characterization of this novel type of17HSD/KSR, type 7, and its cell-specific expression in the placenta and uterus came animportant part of this thesis.

2 Review of the literature
2.1 Physiological roles of estrogens
Sexual maturation and function of the female reproductive tract require estrogen actioneven though initial development and differentiation of the tract appear to be estrogen-independent (Couse & Korach 1999 and references therein). Estrogens, together withother hormones, particularly gonadotropins, coordinate the menstrual cycle and stimulateproliferation of granulosa cells and growth of follicles (Goldfien & Monroe 1991, Adashi1996a, Robker & Richards 1998). Estrogens also induce and maintain female secondarysexual characteristics, enhance stromal development and ductal growth in the mammarygland, and stimulate development of the endometrial lining (Goldfien & Monroe 1991).More specifically, in the endometrium E2 stimulates proliferation of luminal andglandular epithelium, and increases expression of progesterone receptor (PR) (Couse &Korach 1999). In addition, estrogens together with progesterone (P) are necessary formaintaining pregnancy (Pepe & Albrecht 1995, Albrecht et al. 2000).
In addition to these effects in the reproductive system, estrogens are involved in a widevariety of other functions in the human body, such as bone and lipid metabolism andfunctions in the cardiovascular system. Estrogens play an important role in maintainingbone mass in adult women by suppressing bone remodeling and maintaining a balancebetween osteoblastic and osteoclastic activity (Turner et al. 1994). Recent studies havealso shown the importance of estrogens in bone development and mineralization in men(Smith et al. 1994, Simpson 1998). Estrogens have cardiovascular-protective effects thatare reported to be mediated indirectly by an effect on lipoprotein metabolism and directlyby an effect on the vessel wall itself (Farhat et al. 1996, Mendelsohn & Karas 1999). Thecentral nervous system is also a target of estrogen action, since estrogens affect certaintypes of memory, mood and behavior and they may even have neuroprotective effects inthe brain as regards Alzheimer’s disease, for example (Panay & Studd 1998, McEwen &Alves 1999). In addition, estrogens have been suggested to influence the immune system(Olsen & Kovacs 1996).

16
At the molecular level, E2 is a mitogenic factor that in several target tissues stimulatescell proliferation by regulating the expression of enzymes, proteins and growth factorsinvolved in cell replication and nucleic acid synthesis (Cullen & Lippman 1989, Robker& Richards 1998). Hence, estrogens can also act as tumor-promoting agents bystimulating the growth of malignant cells transformed by other mechanisms. The bestknown estrogen-dependent malignancies are breast cancer and endometrial cancer (Benz& Lewis 1991). Peripheral or local estrogen biosynthesis, which is significant aftermenopause, in particular (Labrie 1991), was connected to breast cancer when it wasrealized that E2 accumulates in breast cancer tissue (Fishman et al. 1977, McNeill et al.1986, Vermeulen et al. 1986, Thijssen & Blankenstein 1994). In line with this, enzymescatalyzing the last steps of E2 biosynthesis (17HSD/KSR1 and P450arom) are expressedin peripheral tissues.
2.2 Principles of estrogen action
2.2.1 Estrogen receptors
2.2.1.1 Domain structure of steroid hormone receptors
The mediators of estrogen action, ERs, belong to the steroid/thyroid hormone receptorsuperfamily, represented by a large number of genes encoding these ligand-activatedtranscription factors. In addition to estrogen and thyroid hormone, the family includesreceptors for androgens, progesterone, glucocorticoids, mineralocorticoids, vitamin D andretinoic acids. Furthermore, a number of orphan receptors, for which ligands have not yetbeen identified, have been cloned (Tsai & O'Malley 1994, Mangelsdorf et al. 1995).These nuclear receptors can be aligned on the basis of their primary structure homology,which shows six distinct domains, named A to F (Green & Chambon 1988). The N-terminal region (A/B domain) is of variable length and composition and is poorlyconserved between different receptors. This domain contains a hormone-independenttranscriptional activation function, (AF)-1 (Lees et al. 1989, Tora et al. 1989, Tsai &O'Malley 1994). The C-domain, containing two zinc-fingers, is a DNA-binding region,which is the most conserved region in the receptor family. It is also involved in receptordimerization (Kumar & Chambon 1988, Tsai & O'Malley 1994). The D-domain, alsocalled the hinge region, often contains a nuclear localization signal (Picard et al. 1990,Tsai & O'Malley 1994). The ligand-binding domain (E-domain) is relatively large and isfunctionally complex. It not only harbors regions important for ligand binding but alsoregions involved in receptor dimerization, nuclear localization and interaction withtranscriptional coactivators and corepressors (Tsai & O'Malley 1994, Horwitz et al.1996). It also contains AF-2, which in contrast to AF-1, is ligand-dependent (Webster et

17
al. 1988, Lees et al. 1989). The F-domain has been suggested to modulateagonist/antagonist effects of ligands (Montano et al. 1995).
2.2.1.2 Estrogen receptors α and β
Two estrogen receptors, ERα and ERβ, have been cloned to date. They are similar only inthe C and E regions. Human ERβ shows a high degree of identity (96 %) in the DNA-binding domain and moderate identity (58 %) in the ligand-binding domain comparedwith human ERα, whereas other regions are weakly conserved (Mosselman et al. 1996).Both receptor types bind E2 with high affinity, although ERβ has a slightly lower affinity(Kd = 0.5–0.6 nM) compared with ERα (Kd = 0.2 nM) (Kuiper et al. 1996, Tremblay et al.1997). The binding affinity of the two ERs towards other ligands is similar overall,although there are certain differences, e.g. regarding phytoestrogens (Kuiper et al.1997). In addition to their capacity to bind to DNA as homodimers, ERα and ERβ canform heterodimers which bind to the ERE with an affinity similar to ERα homodimer,and they all are able to stimulate transcription (Cowley et al. 1997, Pettersson et al.1997). In addition, several ERβ isoforms have been cloned from rat and human tissues(Warner et al. 1999 and references therein), but wether these isoforms have significantbiological and physiological roles remains to be investigated.
ERs are rather widely distributed in mammals. Regarding reproduction, importanttissues in which ERα is expressed include uterus, mammary gland, ovary, pituitary andhypothalamus, whereas ERβ is mostly expressed in the granulosa cells of developingfollicles (Couse et al. 1997, Enmark et al. 1997, Kuiper et al. 1997). The biological roleof ERs has been elucidated by studying ER null mice: estrogen receptor α knockout(αERKO) and βERKO (reviewed by Couse & Korach 1999). Even though the uterusundergoes normal pre- and neonatal development in ER null mice, in αERKO adults it ishypoplastic and estrogen-insensitive. In addition, the mammary glands of αERKO miceexhibit impaired development. Female αERKO mice are unovulatory and thus infertile.The sterile phenotype is most probably due to defects in the negative feedback loop of E2
on the hypothalamic-pituitary unit, which leads to chronic exposure to abnormally highconcentrations of LH, as a result of which follicles either prematurely luteinize or becomeatretic. Adult βERKO mice are subfertile, having reduced litter size due to a decline incompleted folliculogenesis and therefore they scarcely ovulate (Couse & Korach 1999). Itis of interest that the βERKO ovarian phenotype has incomplete penetrance when takinginto account that ERβ is specifically expressed in the granulosa cells (Byers et al. 1997,Fitzpatrick et al. 1999). It has been suggested that follicle-stimulating hormone (FSH) isable to correct the defect to some extent since it stimulates proliferation of granulosa cells(Couse & Korach 1999).

18
2.2.2 Molecular mechanisms of estrogen action
In the general estrogen action mode (Fig. 1), estrogens as lipophilic molecules passivelydiffuse into cells (MacLusky 1996), where they either bind to their specific receptors orare metabolized. According to the current concept, ERs when not occupied by ligand,form large heterocomplexes that contain heat shock proteins and immunophilins, whichhave been suggested to have chaperone functions (Pratt & Toft 1997 and referencestherein). In the case of steroid receptors, heat shock proteins have been suggested tofacilitate folding of the hormone-binding domain of the receptors into a high-affinitysteroid-binding conformation (Pratt & Toft 1997). There is also contradictory butconvincing data suggesting that at least in the case of PR and heat shock protein-90,which is a major constituent of heterocomplexes of steroid receptors, hetero-oligomerization of the two is an artifact taking place only in vitro (reviewed by Ylikomiet al. 1998).
Fig. 1. Simplified mechanism of classical estrogen action. In the blood circulation estrogens(E) are mostly bound to SHBG and albumin (= 98 %), from which they are released beforediffusing into a target cell. Most ERs are present in the nucleus, where they are complexedwith several proteins such as heat shock proteins (Hsp). When estrogen binds to ER,receptors are released from heterocomplexes, phosphorylated and dimerized. Activated ER-dimer is then able to bind basal transcriptional machinery and coactivators in a way thatinitiates transcription of an estrogen target gene.

19
Similarly to PR, ER is mainly localized in the nucleus (King & Greene 1984,Guiochon-Mantel et al. 1996), where E2 then binds to ER. This is believed to cause aconformational change in the receptor, as a result of which the receptor dissociates fromthe heterocomplex, dimerizes and binds to the palindromic ERE sequence (Pratt & Toft1997). Binding of the receptor dimer to DNA takes place via the zinc-fingers of the DNA-binding domain (Tsai & O'Malley 1994). Upon binding to ERE the agonist-boundreceptor interacts directly or indirectly with basal transcription machinery to enhancetranscription (Tsai & O'Malley 1994, Horwitz et al. 1996, Shibata et al. 1997). Duringthis binding and activation process, ER is also phosphorylated, which is believed tostimulate the binding of ER to DNA and/or enhance the gene transactivation capacity ofER (Tsai & O'Malley 1994, MacLusky 1996, Weigel 1996). The efficiency oftranscription is additionally modulated by coactivators and/or corepressors that interactwith nuclear receptors to enhance or suppress, respectively, the transcription rate atestrogen target genes (Horwitz et al. 1996, McKenna et al. 1998).
ERs also mediate gene transcription from an AP1 response element via binding ofAP1 transcription factors Fos and Jun (Gaub et al. 1990, Umayahara et al. 1994).Interestingly, at AP1 sites E2 mediates transcription only with ERα, whereas with ERβ itinhibits transcription. Antiestrogens mediate transcription at AP1 sites with both ERαand ERβ (Paech et al. 1997). Similarly to the AP1 element, ER mediates transactivationvia binding to Sp1 transcription factor, which binds to a GC-box and an ERE half-site(Batistuzzo de Medeiros et al. 1997, Porter et al. 1997). ER can also be activated in theabsence of E2 by agents such as growth factors, dopamine, cyclic adenosine-3’,5’-monophosphate (cAMP) and cyclins (Cenni & Picard 1999 and references therein).
In addition to the genomic action of steroids directly on target genes, estrogens canexert their effects in a nonclassic or nongenomic way through receptors on the cellsurface (Revelli et al. 1998). For instance, E2 has been shown to signal by way of theSHBG receptor (Nakhla et al. 1997, Rosner et al. 1999). Furthermore, a fraction of thepopulation of both ERα and ERβ has recently been shown to reside on the cell membraneand activate G-proteins and cell proliferation (Razandi et al. 1999). Most nongenomicactions of steroids so far described involve Ca2+ as a second messenger, preceded by anincrease in intracellular Ca2+ concentration which takes place via release of intracellularcalcium stores or stimulation of Ca2+ influx (Revelli et al. 1998). In addition, there is mostprobably cross-talk between the genomic and nongenomic actions of estrogens (Revelli etal. 1998).
2.3 Pathway of estrogen biosynthesis
Biosynthesis of sex steroids starts from cholesterol, which is a precursor of all steroidhormones (Fig. 2). Cholesterol can be synthesized from acetate in most steroidogenictissues, but the favored route is to obtain it from circulating LDL cholesterol or HDLcholesterol, at least in rats (Albrecht & Pepe 1990, Gibori 1993, Adashi 1996a, Miller1999). Cholesterol resides in LDL as esters which are hydrolyzed and transported to theouter membrane of mitochondria by a process not completely clear but which most

20
probably involves sterol carrier protein 2 (Miller 1995). The movement of cholesterolfrom the outer to inner mitochondrial membranes is facilitated by the steroidogenic acuteregulatory protein (Stocco 1999). Thereafter, cholesterol is converted to 3β-hydroxy-5-pregnen-20-one (pregnenolone) in three sequential steps, 20α-hydroxylation, 22-hydroxylation and C20,22-bond scission catalyzed by cholesterol side-chain cleavageenzyme (P450scc) at a single active site (Duque et al. 1978). This reaction has beenconsidered to be the rate-limiting step in steroidogenesis but the availability of cholesterolin the inner mitochondrial membrane is actually the limiting factor in vivo (Miller 1995,Stocco 1999).
Fig. 2. Pathway of E2 biosynthesis.
In the endoplasmic reticulum, pregnenolone is then converted either to 3β,17α-dihydroxy-5-pregnen-20-one (17-hydroxypregnenolone) by 17α-hydroxylase activity of17α-hydroxylase/C17-20 lyase (P450c17) (Conley & Bird 1997, Miller et al. 1997) or toP by 3β-hydroxysteroid dehydrogenase/∆5-∆4isomerase (3β-HSD) (Simard et al. 1996).Similarly to pregnenolone, P can be converted to a corresponding 17α-hydroxysteroid,17α-hydroxy-4-pregnene-3,20-dione (17-OH-P) by P450c17. Thereafter, 17-hydroxypregnenolone and 17-OH-P can be catalyzed to dehydroepiandrosterone (DHEA)and A-dione, respectively, by the 17,20-lyase activity of P450c17 (Conley & Bird 1997,Miller et al. 1997).
Reactions from pregnenolone to DHEA, and further to A-dione, are those of the ∆5-pathway, whereas reactions from pregnenolone to A-dione via P are those of the ∆4-pathway (Conley & Bird 1997). While rat P450c17 is able to utilize both pathways, thehuman enzyme almost exclusively uses the ∆5-pathway (Conley & Bird 1997). Whetheror not a human endocrine cell synthesizes DHEA or P is thought to depend on the 3β-HSD to P450c17 ratio, in addition to which DHEA synthesis can be controlled byregulating the 17,20-lyase activity of P450c17 by phosphorylation (Conley & Bird 1997,Miller et al. 1997).

21
DHEA, a general precursor of both androgens and estrogens, is converted to A-dioneby 3β-HSD (Simard et al. 1996). A-dione is then converted to T by 17HSD/KSR3(Geissler et al. 1994) or by 17HSD/KSR5 (Dufort et al. 1999, Pelletier et al. 1999). Thefinal steps of E2 biosynthesis are catalyzed by P450arom, which catalyzes aromatizationof androgens to estrogens (Simpson et al. 1997), and by 17HSD/KSR1, which converts E1
to E2 (Peltoketo et al. 1999a and references therein).
2.4 Ovarian physiology
2.4.1 Follicle
The ovaries are responsible for the maturation and release of fertilizable ova, and for thebiosynthesis of steroid hormones, mainly E2, A-dione and P (Adashi 1996a). Ovarianfunctions are controlled by complex feedback mechanisms that involve gonadotropin-releasing hormone from the hypothalamus, gonadotropins from the pituitary, and steroidhormones and protein hormones secreted locally in the ovary (Adashi 1996a, Ferin 1996).E2 is synthesized in the granulosa cells of follicles, where its synthesis is in parallel withfollicular maturation, so that preovulatory follicles are the most abundant E2 source(Gougeon 1996).
2.4.1.1 Follicular maturation and luteinization
Follicular maturation starts from primordial follicles, where a single layer of granulosacells surrounding the oocyte exit the G0 phase of the cell cycle and start slow proliferationunder circumstances that are still unclear (Gougeon 1996, Robker & Richards 1998).Once follicles have begun to grow, basal concentrations of the gonadotropins, FSH andLH, maintain growth from primary follicles to the small antral stage (Richards 1994,Gougeon 1996). During this maturation process the follicles acquire more gonadotropinreceptors, the expression of which is stimulated by FSH and E2. Follicles thereby gainenhanced responsiveness to FSH and LH, and begin producing E2, which leads tomaturation of the follicles to the antral stage (Adashi 1996a, Adashi 1996b, Gougeon1996). At the final phase, exposure to gonadotropins and estrogen triggers a rapid burst ofproliferation that results in the formation of large preovulatory follicles (Adashi 1996b,Gougeon 1996, Robker & Richards 1998). The mitogenic effect of FSH and E2 on thegrowth of granulosa cells is transduced via increasing expression of cyclin D2 and E,which are regulatory proteins of the cell cycle (Robker & Richards 1998). It ischaracteristic of primate folliculogenesis that usually only one follicle reaches thepreovulatory stage, while the rest of the follicles become atretic through the process ofapoptosis (Adashi 1996b, Ferin 1996, Gougeon 1996). Granulosa cells of preovulatory

22
follicles are not only highly proliferative but also differentiating and they acquire LHreceptors, whose expression is stimulated by LH itself (Richards 1994, Adashi 1996a,Gougeon 1996, Robker & Richards 1998). At the end of the follicular phase, the serumconcentration of E2 rises to a level able to stimulate the gonadotropin surge, an LH peakparticularly, that triggers changes leading to ovulation (Ferin 1996). During this processgranulosa cells exit the cell cycle, cease to divide and terminally differentiate, i.e.luteinize to form mature CL (Robker & Richards 1998 and references therein). Uponluteinization, the expression of cyclin D2 and E is down-regulated, whereas expression ofthe negative regulatory protein of the cell cycle, p27KIP1, is enhanced (Robker & Richards1998). Other proteins whose expression is changed during luteinization includeP450arom, P450c17 and ERβ. Their expression is down-regulated, whereas expression ofPR, P450scc, proteolytic enzymes and prostaglandin endoperoxide synthase-2 is up-regulated (Richards 1994, Tsafriri & Chun 1996, Byers et al. 1997, Richards et al. 1998,Fitzpatrick et al. 1999).
2.4.1.2 Estradiol biosynthesis in the follicle and its role duringfolliculogenesis
The pathway of E2 biosynthesis in follicles is compartmentalized, which is known as thetwo-cell, two-gonadotropin model (Hillier et al. 1994, Adashi 1996a). LH stimulatesbiosynthesis of androgens, mainly A-dione (Bergh et al. 1993), a precursor of E2
biosynthesis, in theca cells, which express LH receptor. One of the target genes of LHaction in theca cells is that for P450c17. From theca cells, androgens traverse thebasement membrane to the neighboring granulosa cells, which express FSH receptor. Ingranulosa cells A-dione is first catalyzed to E1 by P450arom and then to E2 by17HSD/KSR1, the expression of both of these enzymes being stimulated by FSH(Ghersevich et al. 1994a, Hillier et al. 1994, Adashi 1996a, Kaminski et al. 1997,Peltoketo et al. 1999a and references therein). In other words, theca cells do notsynthesize estrogens since they do not express P450arom or 17HSD/KSR1 (Hillier et al.1994, Sawetawan et al. 1994). Granulosa cells in turn are not able to produce androgenssince they do not express P450c17. However, both cell types express P450scc and 3β-HSD (Hillier et al. 1994, Richards 1994).
In addition to its multiple systemic effects, E2 exerts a variety of critical actions at thelevel of the ovary, where it affects the growth and development of ovarian follicles bystimulating the proliferation of granulosa cells (Robker & Richards 1998), stimulatingantrum formation, increasing, together with FSH, gonadotropin receptor levels in thegranulosa cells, enhancing gonadotropin-stimulated P450arom activity, and stimulatinggap junction formation among granulosa cells (Adashi 1996a, Shoham & Schachter 1996and references therein,). Furthermore, E2 plays a major role in feedback communicationbetween the ovary and the hypothalamic-pituitary unit. The first of two feedback loops isthe negative loop, whereby E2 feeds back to adjust gonadotropin-releasing hormoneand/or gonadotropin secretion (Ferin 1996). Positive feedback occurs as a stimulus toovulation, as described in section 2.4.1.1. In line with these estrogen effects, both ER

23
types are expressed in the ovary, ERβ being the dominant form in granulosa cells (Byerset al. 1997, Enmark et al. 1997, Fitzpatrick et al. 1999). In addition, recent studies withERKO mice suggests that both ERα and ERβ are involved in the negative feedback loopof E2 on the hypothalamic-pituitary unit, suppressing gonadotropin secretion (Couse &Korach 1999, Couse et al. 1999).
2.4.2 Corpus luteum
As a result of gonadotropin surge-stimulated follicular rupture, ovulation, the folliclebecomes the CL. The main role of the CL is the production of P, in addition to which itsecretes estrogens, and in humans, peptide/protein hormones such as relaxin, oxytocinand inhibin-like factors. The physiological role of P is related to preparation of thereproductive tract and gametes for fertilization, implantation, and the maintenance of theuterine environment during gestation (Gibori 1993, Pfeifer & Strauss 1996). The humanCL is an active endocrine gland during the luteal phase, while in rodents, secretion of Pafter ovulation is short-lived and therefore the CL of the cycle is not believed to have aclearly defined role. In both humans and rodents, the CL is a transient organ if nothormonally stimulated as a result of pregnancy, or as a result of mating in rodents. Thehuman CL is rescued by human chorionic gonadotropin (hCG), while in rodents PRL isneeded for this purpose. In rodents, function of the CL is indispensable throughoutgestation, whereas in humans, the placenta replaces luteal functions in early pregnancy(luteal-placental shift), after which the CL is luteolyzed (Taya & Greenwald 1981, Giboriet al. 1988, Gibori 1993, Stouffer 1996).
2.4.2.1 Steroidogenesis and role of estradiol in the corpus luteum
Although there is a transient decline in androgen and estrogen synthesis around the timeof ovulation, P450c17 and P450arom mRNAs and activity remain elevated in humanluteal tissue throughout the luteal phase as well as in the CL of pregnancy until the luteal-placental shift (Stouffer 1996). In rat CL of pregnancy, P450scc and 3β-HSD areexpressed more or less in a constitutive manner (Gibori 1993), whereas expression ofP450c17 and P450arom are regulated. Expression of P450c17 is stimulated by LH untilmid-pregnancy, when LH levels drop (Morishige et al. 1973, Khan et al. 1987), whichresults in a decline of androgen production. At this stage, the rodent placenta becomes theprimary source of androgen (Fig. 3) which is necessary for maximal ovarian E2
biosynthesis (Warshaw et al. 1986, Gibori et al. 1988). The amount of P450arom is lowin the first half of pregnancy, it increases from mid-pregnancy, and declines at the end ofpregnancy (Hickey et al. 1988). In contrast to granulosa cells, P450arom expression inthe rat CL is not affected by FSH, LH or cAMP but is regulated by PRL, placentallactogen and luteal E2 itself, for example (Gibori 1993). No specific 17HSD/KSR has

24
been demonstrated in the CL of rodents, whereas in humans, 17HSD/KSR1 is expressedat least to some extent in the CL (Sawetawan et al. 1994).
Fig. 3. Estrogen biosynthesis during pregnancy: primates vs. rodents. In primates estrogensare predominantly synthesized from fetal and maternal precursors in the placenta, whereasin rodents synthesis mainly takes place in the CL. Estrogen biosynthesis in the luteal cells ofrodent CL relies on luteal androgen precursors in the first half of gestation, while in thesecond half of pregnancy the placenta supplies the CL with androgen precursors. In rodents,cooperation between the CL and placenta is also evident in the source of P. The placentautilizes luteal P as an additional source for androgen biosynthesis since P production islimited in the rodent placenta (Gibori et al. 1988, Gibori 1993, Kuss 1994, Wilson & Parson1996).
In rodents, E2 acts by an intracrine mechanism to affect steroidogenesis,vascularization and protein synthesis of the CL (Gibori et al. 1988, Gibori 1993). Duringthe first week of pregnancy, PRL secreted by the pituitary is able to sustain basal levels ofP, but E2 is necessary for maximal P production (Gibori & Keyes 1980). In the secondweek, E2 becomes essential for normal production of P. Between days 12-21, E2 is

25
responsible together with placental lactogen, a PRL-related protein, for the remarkableincrease in CL weight, vascularization and steroidogenic capacity (Gibori et al. 1988,Gibori 1993). In line with estrogen’s local effects in the CL, luteal cells express both ERαand ERβ (Telleria et al. 1998, Misao et al. 1999). A prerequisite for E2 action is previousexposure of the CL to PRL and PRL-related hormones, an important role of which is tosustain high levels of ER (Gibori & Keyes 1980, Gibori et al. 1988). In fact, expression ofboth ERs has been shown to be up-regulated by PRL and placental lactogens (Telleria etal. 1998). The main action of E2 in steroidogenesis is to increase the supply of cholesterolsubstrate by mobilizing cholesterol storage, stimulating cholesterol synthesis andenhancing luteal cell content of lipoprotein receptors and thus uptake of circulatingcholesterol. It also stimulates the transport of cholesterol to mitochondrial P450scc. Inaddition, E2 enhances luteal cell hypertrophy by increasing protein content throughstimulation of expression of elongation factor 2. E2 stimulation of luteal cell hypertrophyis accompanied by a remarkable proliferation of vascular endothelial cells (Gibori et al.1988, Gibori 1993).
2.5 The fetoplacental unit as a sex hormone source
The placenta has several important functions during pregnancy: it physically anchors thefetus to the uterus, it transports nutrients from the maternal circulation to the fetus, itexcretes fetal metabolites into the maternal compartment, it has an immunomodulatoryrole in the maternal acceptance of the fetus, and it produces a wide range of peptide andsteroid hormones (Strauss et al. 1996, Wilson & Parson 1996). Estrogens (E1, E2 and E3)and P are the principal steroid hormones produced by the placenta during primatepregnancy (Albrecht & Pepe 1990, Miller 1999). At the beginning of human pregnancythese hormones are produced by the CL, whose functional life span is only ten weeks.From approximately the eighth week, the placenta acts as a significant sex hormonesource (Albrecht & Pepe 1990, Strauss et al. 1996).
The main role of estrogen during pregnancy is to stimulate key steps of P biosynthesisin syncytiotrophoblasts by enhancing cholesterol formation in the fetal liver, LDL uptakeand P450scc activity (Pepe & Albrecht 1995). In addition, E2 enhances uteroplacentalblood flow and possibly placental neovascularization to provide optimal gas exchangeand nutrients, and it is also involved in the regulation of mammary gland developmentduring pregnancy. The key role of P is to maintain pregnancy by promoting uterinemyometrial quiescence, in addition to which it has a suppressive action on the immunesystem to prevent rejection of the developing fetus and placenta (Pepe & Albrecht 1995,Wilson & Parson 1996).
Human placental villi consist of two types of trophoblast cells: an inner layer ofcytotrophoblasts, which are mitotically active mononuclear cells prominent early inpregnancy, and an outer layer of syncytiotrophoblasts, which are fused nondividingmultinuclear cells that become predominant later in pregnancy. Cytotrophoblasts andsyncytiotrophoblasts produce peptide and protein hormones, whereas thesyncytiotrophoblasts produce all of the steroid hormones (Ringler & Strauss 1990,

26
Wilson & Parson 1996). Biosynthesis of P starts from the uptake of LDL cholesterol,since de novo synthesis of cholesterol is believed to be limited in the placenta (Albrecht& Pepe 1990). Because human placenta does not express P450c17 (Albrecht & Pepe1990, Strauss et al. 1996), estrogen biosynthesis depends on fetal and maternal adrenalprecursors, DHEA and dehydroepiandrosterone sulfate (DHEAS), and 16α-hydroxydehydroepiandrosterone sulfate (16α-OH-DHEAS) produced by the fetal liver(Fig. 3) (Albrecht & Pepe 1990, Miller 1999). In the placenta, these steroids are taken upby syncytiotrophoblasts, where they are desulfonated by steroid sulfatase (Salido et al.1990) and thereafter converted to E1, E2 and E3 by sequential reactions catalyzed by 3β-HSD type 1, P450arom and 17HSD/KSR1 (Albrecht & Pepe 1990 and references therein,Miller 1999, Peltoketo et al. 1999a). The normal ratio of E1, E2 and E3 in term cord bloodis 14:5:81 (Miller 1999).
The rodent chorioallantoic placenta is structurally different from the primate placentaand consists of four types of differentiated trophoblast cells: trophoblast giant cells,spongiotrophoblasts, glycogen cells and syncytial trophoblast cells (Soares et al. 1996).They contribute to the formation of two morphologically and functionally distinct regionsof the chorioallantoic placenta, the junctional zone and the labyrinth zone. Trophoblastgiant cells and spongiotrophoblasts exhibit endocrine activities by producing a variety ofpeptide hormones and steroid hormones, whereas syncytial trophoblast cells in thelabyrinth zone carry out transport functions and the glycogen cells serve as a potentialenergy reserve (Sherman 1983, Soares et al. 1996). The biosynthesis of sex steroidsrepresents another major difference between the human and rodent placenta. In contrastto human placenta, rodent placenta is believed to be unable to synthesize E2 due to its lackof P450arom (Townsend & Ryan 1970, Rembiesa et al. 1971). In addition, only a limitedamount of P is secreted in the rodent placenta (Matt & MacDonald 1984). An uniquefeature of the rodent placenta is synthesis of androgen precursors (Fig. 3), which are thenused in E2 biosynthesis in the CL (Warshaw et al. 1986, Gibori et al. 1988).
2.6 17ββββ-Hydroxysteroid dehydrogenases (17HSDs)/17-ketosteroid
reductases (17KSRs)
The 17HSD/KSRs are nicotinamide adenine dinucleotide [NAD(H)]- and/or its phosphateform [NADP(H)]-dependent enzymes that in a positional and stereospecific mannercatalyze hydride transfer from NAD(P)H to 17-ketosteroids (17KSR activity) or from17β-hydroxysteroids to NAD(P)+ (17HSD activity) (Peltoketo et al. 1999a). Since bothestrogens and androgens have their highest activity in the 17β-form, 17HSD/KSRsregulate the biological activity of these sex steroids. The 17HSD/KSRs primarily convertthe relatively inactive sex steroids E1, A-dione and 5α-androstane-3,17-dione (5α-A-dione) to their more potent forms: E2, T and dihydrotestosterone (DHT), and vice versa.Therefore, reductive 17HSD/KSRs are indispensable for E2 and T biosynthesis in theovary and testis, respectively. In addition to the gonads, these 17HSD/KSRs in primatesand rodents are also expressed in certain peripheral tissues (Martel et al. 1992, Labrie etal. 1997), where they participate in regulation of local steroid hormone concentrations. In

27
turn, oxidative 17HSD/KSRs, preferring inactivation of sex hormones bydehydrogenation, are generally more widely expressed and they are likely to protecttissues from excessive hormone action (Moghrabi et al. 1997, Mustonen et al. 1998b,Peltoketo et al. 1999a, Peltoketo et al. 1999b).
At least eight different 17HSD/KSRs have been cloned to date, and they have beennamed chronologically types 1–8 (Peltoketo et al. 1999b and references therein). Despitethe same reaction type catalyzed, they share an amino acid sequence identity of less than30 %. In addition to their distinct primary structures, they are dissimilarly distributed,they differ in substrate and cofactor specificities, and they consequently have apparentlyseparate physiological functions. In particular, 17HSD/KSR types 1, 3 and 5 mainlycatalyze reductive reactions of estrogens and androgens (Poutanen et al. 1993, Geissler etal. 1994, Miettinen et al. 1996a, Dufort et al. 1999), whereas types 2, 4, 6 and 8 can beconsidered as oxidative enzymes (Wu et al. 1993, Adamski et al. 1995, Biswas & Russell1997, Fomitcheva et al. 1998). Several 17HSD/KSR enzymes, even counteracting ones,may coexist in the same tissue and even in the same cell type (Miettinen et al. 1996a),where steady-state concentrations of sex steroids are a result of the sum of action ofdifferent 17HSD/KSRs.
In addition, some of the enzymes named 17HSD/KSR have also been reported tocatalyze several other reactions, such as 20α-HSD, 3α-HSD and 3β-HSD reactions, β-oxidation of fatty acids, oxidation of xenobiotics and reactions between retinoic acids(Wu et al. 1993, Deyashiki et al. 1995, Dieuaide-Noubhani et al. 1996, Leenders et al.1996, Qin et al. 1997a, Dufort et al. 1999, Su et al. 1999). The properties of different17HSD/KSRs are summarized in Table 1.
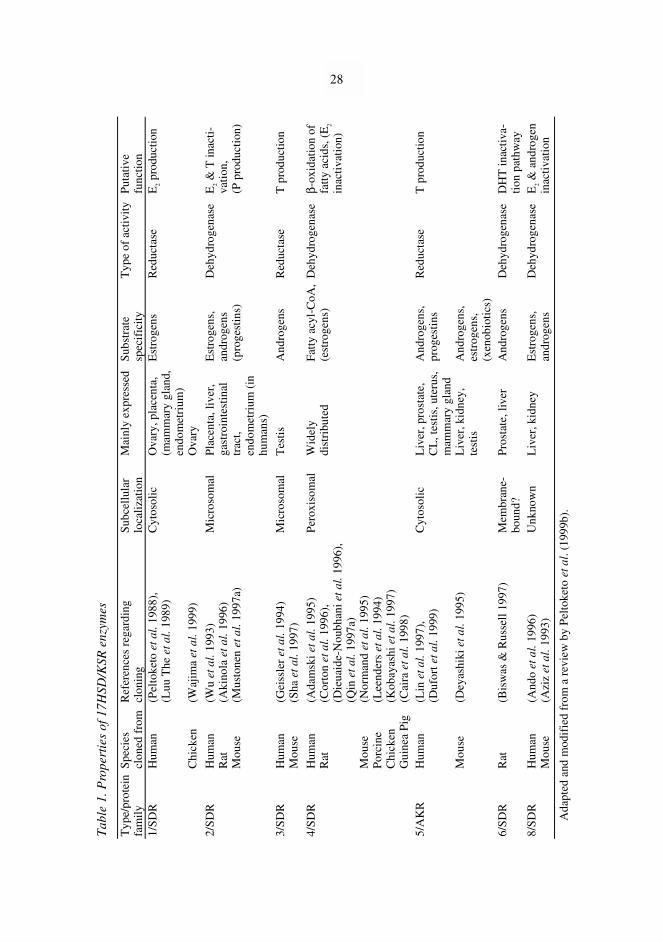
Tab
le 1
. Pro
pert
ies
of 1
7HSD
/KSR
enz
ymes
Typ
e/pr
otei
nfa
mil
yS
peci
escl
oned
fro
mR
efer
ence
s re
gard
ing
clon
ing
Sub
cell
ular
loca
liza
tion
Mai
nly
expr
esse
dS
ubst
rate
spec
ific
ity
Typ
e of
act
ivity
Put
ativ
efu
nctio
n1/
SD
RH
uman
Chi
cken
(Pel
toke
to e
t al.
1988
),(L
uu T
he e
t al.
1989
)
(Waj
ima
et a
l. 19
99)
Cyt
osol
icO
vary
, pla
cent
a,(m
amm
ary
glan
d,en
dom
etri
um)
Ova
ry
Est
roge
nsR
educ
tase
E2 p
rodu
ctio
n
2/S
DR
Hum
anR
atM
ouse
(Wu
et a
l. 19
93)
(Aki
nola
et a
l. 19
96)
(Mus
tone
n et
al.
1997
a)
Mic
roso
mal
Pla
cent
a, li
ver,
gast
roin
test
inal
trac
t,en
dom
etri
um (
inhu
man
s)
Est
roge
ns,
andr
ogen
s(p
roge
stin
s)
Deh
ydro
gena
seE
2 & T
inac
ti-
vati
on,
(P p
rodu
ctio
n)
3/S
DR
Hum
anM
ouse
(Gei
ssle
r et
al.
1994
)(S
ha e
t al.
1997
)M
icro
som
alT
esti
sA
ndro
gens
Red
ucta
seT
pro
duct
ion
4/S
DR
Hum
anR
at
Mou
seP
orci
neC
hick
enG
uine
a P
ig
(Ada
msk
i et a
l. 19
95)
(Cor
ton
et a
l. 19
96),
(Die
uaid
e-N
oubh
ani e
t al.
1996
),(Q
in e
t al.
1997
a)(N
orm
and
et a
l. 19
95)
(Lee
nder
s et
al.
1994
)(K
obay
ashi
et a
l. 19
97)
(Cai
ra e
t al.
1998
)
Per
oxis
omal
Wid
ely
dist
ribu
ted
Fat
ty a
cyl-
CoA
,(e
stro
gens
)D
ehyd
roge
nase
β-ox
idat
ion
offa
tty a
cids
, (E
2
inac
tiva
tion
)
5/A
KR
Hum
an
Mou
se
(Lin
et a
l. 19
97),
(Duf
ort e
t al.
1999
)
(Dey
ashi
ki e
t al.
1995
)
Cyt
osol
icL
iver
, pro
stat
e,C
L, t
esti
s, u
teru
s,m
amm
ary
glan
dL
iver
, kid
ney,
test
is
And
roge
ns,
prog
esti
ns
And
roge
ns,
estr
ogen
s,(x
enob
ioti
cs)
Red
ucta
seT
pro
duct
ion
6/S
DR
Rat
(Bis
was
& R
usse
ll 1
997)
Mem
bran
e-bo
und?
Pro
stat
e, li
ver
And
roge
nsD
ehyd
roge
nase
DH
T in
acti
va-
tion
pat
hway
8/S
DR
Hum
anM
ouse
(And
o et
al.
1996
)(A
ziz
et a
l. 19
93)
Unk
now
nL
iver
, kid
ney
Est
roge
ns,
andr
ogen
sD
ehyd
roge
nase
E2 &
and
roge
nin
acti
vati
on
Ada
pted
and
mod
ifie
d fr
om a
rev
iew
by
Pel
toke
to e
t al .
(199
9b).
28

29
2.6.1 The short-chain dehydrogenases/reductases family
Apart from 17HSD/KSR5, all 17HSD/KSRs cloned so far belong to a large proteinfamily of short-chain dehydrogenases/reductases (SDRs) which are also called short-chain alcohol dehydrogenases. 17HSD/KSR5 belongs to the aldoketoreductase (AKR)protein family. The SDR family includes enzymes from humans, animals, plants, insectsand bacteria that use steroids, sugars, prostaglandins, antibiotics, aromatic hydrocarbonsand compounds involved in nitrogen metabolism as substrates (Krozowski 1992,Krozowski 1994, Jörnvall et al. 1995). These enzymes have an average length of 250amino acids and they act independently of metal cofactors, in contrast to medium-chaindehydrogenases/reductases that need zinc (Persson et al. 1991). Even though SDRs sharea low overall amino acid identity (15–30 %), their three-dimensional folding andhydrophobic core are similar, and most SDRs are dimers or tetramers (Jörnvall et al.1995). In addition, overall comparisons of SDR enzymes suggest the possibility of relatedreaction mechanisms and domain properties within the family (Persson et al. 1991). Ofthe family, bacterial 3α,20β-hydroxysteroid dehydrogenase was the first SDR whosetertiary structure was determined using crystallographic analyses (Ghosh et al. 1991,Ghosh et al. 1994).
The only amino acid residue totally conserved in the entire family is the tyrosineresidue in the highly conserved Tyr-X-X-X-Lys motif (X, any amino acid) (Jörnvall et al.1995), which forms part of a catalytic triad involved in the transfer of hydride betweencofactor and substrate (Ghosh et al. 1995, Jörnvall et al. 1995). A largely conservedserine residue, 13 residues upstream from the tyrosine motif (Jörnvall et al. 1995) is alsopart of the catalytic triad (Ghosh et al. 1995). An additional conserved region is aglycine-rich motif (Gly-X-X-X-Gly-X-Gly) in the N-terminal part of the molecule thatbinds cofactor, NAD(H) or NADP(H) (Krozowski 1992, Jörnvall et al. 1995). As aphylogenetic study has shown, 17HSD/KSRs do not form a subclass within the SDRfamily but are in different branches (Baker 1995). It is not known, however, if different17HSD/KSRs are a result of gene duplication followed by sequence divergence orfunctional convergence (Baker 1995).
2.6.2 17HSD/KSR type 1
2.6.2.1 Structural and functional properties of human 17HSD/KSR1
Human 17HSD/KSR1 was partially purified from human placenta as early as in 1958(Langer & Engel 1958). The complete amino acid sequence based on cloned cDNA wasdescribed 30 years later by Peltoketo et al. (1988) and Luu-The et al. (1989). The yearsbetween the 60s and the late 80s were the time of development of purification methodsand analyses of enzymatic properties. Human 17HSD/KSR1 is a cytosolic enzyme thatwas initially found to catalyze mainly the interconversion between E1 and E2 using

30
NAD(H) or NADP(H) as a cofactor (Jarabak 1969, Jarabak & Sack 1969, Karavolas &Engel 1971, Jin & Lin 1999). Purified native 17HSD/KSR1 and recombinant type 1enzyme, produced in Sf9 cells, catalyze more efficiently the reaction from E1 to E2 thanthe opposite reaction (Puranen et al. 1994, Puranen et al. 1997a, Jin & Lin 1999). Inaddition, when the activity of 17HSD/KSR1 is measured in intact cultured cells, which isbelieved to correspond well to conditions in vivo, the enzyme almost exclusivelycatalyzes the reaction from E1 to E2 (Poutanen et al. 1993). Furthermore, there is acorrelation between E1 to E2 17KSR activity and type 1 enzyme expression in several celllines (Miettinen et al. 1996a).
In addition to estrogenic reactions, 17HSD/KSR1 has been detected to catalyzeinterconversions between A-dione and T, DHEA and androstenediol (A-diol), 5α-A-dione and DHT, and between P and 20α-hydroxyprogesterone (20-OH-P) (Purdy et al.1964, Jarabak & Sack 1969, Strickler et al. 1981, Mendoza-Hernández et al. 1984,Puranen et al. 1997a). However, the results of kinetic studies have shown that human17HSD/KSR1 prefers C18 steroids over C19 and C20 steroids as substrates (Langer et al.1959, Adams et al. 1962, Purdy et al. 1964, Jarabak & Sack 1969, Mendoza-Hernándezet al. 1984, Puranen et al. 1997a).
Native human 17HSD/KSR1 is a dimer composed of two identical subunits which arenon-covalently bound, but strongly associated with each other (Burns et al. 1971, Burnset al. 1972, Lin et al. 1992, Puranen et al. 1997b). According to the results ofpolyacrylamide gel electrophoresis the size of a subunit is approximately 34 kilodaltons(kDa), which corresponds well with the calculated molecular mass (34.9 kDa) based onthe cDNA (Peltoketo et al. 1988, Luu The et al. 1989). The native enzyme has beenreported to show microheterogeneity, with several isoelectric points (Engel & Groman1974) that could be due to post-translational modifications. The absence of carbohydratesin the purified protein (Burns et al. 1972) and the absence of N-linked glycosylation sitesin the primary structure (Peltoketo et al. 1988) rules out this source of heterogeneity.Recent studies have shown that the enzyme possesses a serine residue, Ser134, which isphosphorylated by protein kinase A (PKA) in vitro (Barbieri et al. 1994, Puranen et al.1997b). Phosphorylation, however, has no significant effect on the catalytic properties ormicroheterogeneity of the enzyme (Puranen et al. 1997b).
Using X-ray crystallographic techniques, the three-dimensional structure of human17HSD/KSR1 has recently been resolved by Ghosh et al. (1995), utilizing the structure of3α,20β-hydroxysteroid dehydrogenase. The structure and reaction mechanism have beenfurther resolved in subsequent studies (Gibori et al. 1988, Azzi et al. 1996). Human17HSD/KSR1 has a secondary structure with alternating α-helices and β-strandsresulting in a similar overall fold as in other SDRs (Ghosh et al. 1995). At the catalyticsite, Tyr155 is essential for catalysis (Puranen et al. 1994, Puranen et al. 1997b). Thistyrosine residue is a proton donor in the hydride transfer chain. According to the currentmodel, the hydroxyl groups at Tyr155 and Ser142 and the steroid O17 atom form a triangularhydrogen bond network enabling hydride transfer and reduction of E1 to E2. Ser142 hasbeen suggested to stabilize the intermediate transition states. In addition, a third aminoacid, Lys159, has been suggested to be directly involved in catalysis by forming hydrogenbonds with the cofactor and apparently lowering the pKa of the hydroxyl proton in Tyr155
to facilitate proton transfer (Ghosh et al. 1995, Azzi et al. 1996, Breton et al. 1996,Peltoketo et al. 1998). The difference in substrate specificity between human and rat

31
17HSD/KSR1 (see section 2.6.2.5.) has been suggested to be due to Asn152, Pro187 andamino acids between residues 190–230, and membrane association near the active site ofthe enzyme (Puranen et al. 1997b, Peltoketo et al. 1998). Detailed knowledge of thestructure of 17HSD/KSR1 is being utilized to design and generate specific and potentinhibitors of 17HSD/KSR1 for clinical use.
2.6.2.2 Tissue distribution and role of human 17HSD/KSR1in estradiol biosynthesis
A 1.3-kilobase (kb) mRNA encoding human 17HSD/KSR1 is mainly expressed in theovary and placenta, in addition to which it is also expressed in the mammary gland,endometrium, and certain breast cancer and choriocarcinoma cell lines (Luu-The et al.1990, Poutanen et al. 1990, Miettinen et al. 1996a, Zeitoun et al. 1998, Miettinen et al.1999). In line with this, 17HSD/KSR1 protein is expressed in the ovary, placenta, inchoriocarcinoma cell lines, normal endometrium (Mäentausta et al. 1990, Mäentausta etal. 1991, Sawetawan et al. 1994, Miettinen et al. 1996a), endometriotic lesions (Zeitounet al. 1998) and in around 50 % of endometrial adenocarcinoma specimens (Mäentaustaet al. 1992). The protein has also been detected in some samples of normal mammarygland (Söderqvist et al. 1998), and in about 70 % of benign and 50 % of malignant breastspecimens, where it is present in the epithelial cells, although in highly variable amounts(Poutanen et al. 1992a, Sasano et al. 1996).
In the ovary 17HSD/KSR1 is expressed in the granulosa cells of developing follicles,ranging from primary to tertiary follicles (Sawetawan et al. 1994). The associationbetween 17HSD/KSR1 expression and the maturation stage of follicles (Ghersevich et al.1994a, Sawetawan et al. 1994) is related to their capacity to produce E2 (Gougeon 1996).The correlation of 17HSD/KSR1 expression with 17KSR activity and with E2 productionin granulosa cells further confirms that the type 1 enzyme is the major 17HSD/KSRinvolved in E2 biosynthesis during follicle maturation (Ghersevich et al. 1994a). Human17HSD/KSR1 is also expressed in the granulosa-luteal cells of the CL (Ghersevich et al.1994a, Sawetawan et al. 1994).
In the placenta 17HSD/KSR1 is expressed in the syncytiotrophoblasts (Fournet-Dulguerov et al. 1987, Mäentausta et al. 1991, Sawetawan et al. 1994, Mustonen et al.1998a, Takeyama et al. 1998), which have a high capacity to produce E2 from androgenprecursors of fetal and maternal origin, since the cells also abundantly express P450arom(Fournet-Dulguerov et al. 1987, Simpson et al. 1995). In the endometrium the type 1enzyme is present in glandular and surface epithelial cells (Mäentausta et al. 1991),where its concentration is highest during the early to mid-secretory phase (Mäentausta etal. 1990, Mäentausta et al. 1991). E2 stimulates proliferation of the endometrialepithelium in the proliferative phase, but the role of 17HSD/KSR1 and E2 biosynthesis insecretory endometrium is obscure since ER concentrations decrease in the secretoryphase (Fleming et al. 1980) and it is oxidative 17HSD/KSR2 that predominates in theendometrium (Casey et al. 1994).

32
2.6.2.3 Human 17HSD/KSR1 in normal mammary gland andin breast cancer
During adolescence the duct system of the mammary gland proliferates extensively, andthis is under the control of estrogen and growth hormones (Couse & Korach 1999 andreferences therein). It is thus not surprising that estrogen-induced proliferation is assumedto play a central role in the promotion of breast cancer and tumor growth (Cullen &Lippman 1989 and references therein). Estrogen metabolism in the mammary gland andits influence on the risk of breast cancer have been subjects of interest lately (Labrie1991, Santen 1996, Bulun et al. 1997), since it was realized that E2 accumulates in breastcancer tissue (Fishman et al. 1977, McNeill et al. 1986, Vermeulen et al. 1986, Thijssen& Blankenstein 1994), and 17HSD/KSR1, P450arom and steroid sulfatase are expressedin the mammary gland (Santner et al. 1984, Sasano et al. 1996, Bulun et al. 1997). Theseenzymes are needed to synthesize E2 in situ from the circulating precursors A-dione, E1
and E1-sulfate. After menopause, in particular, peripheral estrogen biosynthesis plays amajor role in estrogen action (Labrie 1991).
The role of human 17HSD/KSR1 in local E2 biosynthesis and its impact onproliferation have been demonstrated in studies where the enzyme was expressed incultured breast cancer cells to the same degree as in certain breast tumors. E1 increasedthe growth of cells in a similar manner as those treated with E2, showing the activity andgrowth-promoting role of 17HSD/KSR1. A growth response was not achieved with E1 inbreast cancer cells not expressing 17HSD/KSR1 (Miettinen et al. 1996b). Hence,inhibitors of the type 1 enzyme could be feasible as regards additional breast cancertreatment, since some breast cancer patients initially treated with antiestrogens relapse,probably due to enhanced estrogen sensitivity, indicating that inhibitors that blockestrogen biosynthesis completely are needed (Santen et al. 1990, Masamura et al. 1995).Antiestrogens and inhibitors of E2 biosynthesis, mainly inhibitors of P450arom, havealready been utilized in the treatment of breast cancer for over two decades (Santen et al.1990, Miller 1996). Designing of inhibitors of 17HSD/KSR1 and steroid sulfatase is alsoin progress (Penning 1996, Reed et al. 1996). Interestingly, several phytoestrogens,especially apigenin, coumestrol and genistein, have inhibitory effects on 17HSD/KSR1activity in vitro and in cultured breast cancer cells (Mäkelä et al. 1995, Mäkelä et al.1998). Flavonoids are considered to be the least estrogenic of phytoestrogens. In line withthis, higher concentrations of the flavonoid apigenin are needed to bring about estrogenaction than to inhibit 17HSD/KSR1 (Mäkelä et al. 1995, Mäkelä et al. 1998).
2.6.2.4 Structure of the human 17HSD/KSR1 gene
The gene encoding human 17HSD/KSR1, HSD17B1 (previously also called EDH17B2)is located in tandem with the HSD17BP1 gene in chromosome region 17q12-21(Winqvist et al. 1990, Luu-The et al. 1990, Peltoketo et al. 1992, Simard et al. 1993).HSD17B1 is a small gene consisting of six exons and five short introns. It shares 89 %identity with HSD17BP1, which is organized in a similar manner. HSD17BP1 is

33
considered to be a pseudogene, since it contains a premature in-frame stop codon at theposition coding for amino acid 214 (Luu-The et al. 1990). In addition, there is anucleotide change (A→C) at the putative TATA box of HSD17BP1 which significantlydecreases promoter activity in vitro (Peltoketo et al. 1994). However, Touitou et al.(1994) reported that by using the reverse transcriptase polymerase chain reaction (RT-PCR) a transcript from HSD17BP1 can be detected in several cancer cell lines and biopsysamples from the mammary gland, uterus and placenta.
Human HSD17B1 is transcribed into two mRNAs, 1.3 and 2.3 kb in size, using twomajor transcription start sites located 9–10 and 971 nucleotides upstream of thetranslation initiation codon (Luu The et al. 1989, Luu-The et al. 1990). The shortertranscript is expressed in cells containing 17HSD/KSR1 enzyme, and its amountcorrelates well with the concentration of the protein and 17KSR activity (Miettinen et al.1996a). So far, three elements regulating the efficiency of transcription of the 1.3-kbmRNA have been identified in the 5’-region of human HSD17B1. First, the fragmentbetween -78 to +9, with respect to the cap site for the 1.3-kb transcript, has beencharacterized as a proximal promoter. Second, the region from -661 to -392 is a cell-specific enhancer and third, the fragment between these two elements, from -392 to -78,acts as a silencer (Piao et al. 1995, Peltoketo et al. 1999a). The role of the 2.3-kbtranscript is less clear than that of the shorter type, since 2.3-kb mRNA is constitutivelyexpressed in several tissues, and its concentration does not correlate with 17HSD/KSR1protein levels or 17KSR activity (Miettinen et al. 1996a). In addition, no typical promoterelements proximal to the transcription start site of the 2.3-kb mRNA have been detected(Peltoketo et al. 1992).
Fourteen single base allelic variations characterized as polymorphisms have beendetected in human HSD17B1 (Normand et al. 1993, Simard et al. 1993, Mannermaa et al.1994, Peltoketo et al. 1994). Of these, only two resulted in a change of amino acid,namely Ala237→Val and Ser312→Gly. However, these amino acid changes do not affect theactivity of 17HSD/KSR1 (Puranen et al. 1994). Interestingly, one single base allelicvariation located in the putative TATA box decreases promoter activity by 45 % in vitro(Peltoketo et al. 1994). However, its frequency is the same among breast cancer patientsand control individuals.
2.6.2.5 Rat 17HSD/KSR1
The rat hsd17b1 gene is transcribed into two mRNAs, 1.4 and 1.7 kb in size, as a result ofalternative polyadenylation (Akinola et al. 1998). Cis-acting elements known to modulateexpression of the human HSD17B1 gene are not conserved in the rat gene, indicating thatthe two genes are partly differentially regulated (Akinola et al. 1998). The rat17HSD/KSR1 mRNAs are almost exclusively expressed in the ovary, and only very lowamounts of the transcripts have been found in the kidney (Akinola et al. 1996). Inaddition, using human 17HSD/KSR1 antibody, type 1 enzyme has been detected in ratbrain (Pelletier et al. 1995).

34
Unlike the human enzyme, rat 17HSD/KSR1 catalyzes the reaction from A-dione to Tas efficiently as that of E1 to E2, both in cultured cells and in vitro (Akinola et al. 1996,Puranen et al. 1997a). Of the estrogens, the rat enzyme strongly prefers E1 over E2 as asubstrate (Puranen et al. 1997a). In contrast to human type 1 enzyme, which is a cytosolicprotein, rat 17HSD/KSR1 is inactive in cell homogenates without a nonionic detergent.Therefore, membrane interaction has been suggested to be important in maintaining itsfull activity (Puranen et al. 1997a).
Rat 17HSD/KSR1 is expressed throughout the rat estrous cycle in the granulosa cellsof developing follicles of different stages. The expression of rat type 1 enzyme isconstitutive in whole ovaries in different phases of the estrous cycle (Akinola et al. 1997).However, studies by Ghersevich et al. (1994b) have shown that treatment with pregnantmare serum gonadotropin for 1 day up-regulates the expression of 17HSD/KSR1 in thefollicles of rat ovaries, whereas after 2-day treatment the expression is down-regulated.The expression is further decreased by 1-day treatment with hCG (Ghersevich et al.1994b). Therefore, the expression of rat 17HSD/KSR1 in granulosa cells is suggested tobe related to the maturation and luteinization stage of the follicles (Ghersevich et al.1994b, Akinola et al. 1997). In addition, rat type 1 enzyme is more or less constitutivelyexpressed in whole ovaries during gestation, though a slight increase in the mRNA levelhas been detected from day 5 to day 15 (Akinola et al. 1997).
2.6.2.6 Regulation of 17HSD/KSR1
Regulation of 17HSD/KSR1 is best characterized in the ovaries and in cells of placentalorigin. Pituitary gonadotropins (FSH and LH) are the main regulators of the maturation offollicles and E2 secretion, and this process includes regulation of P450arom and17HSD/KSR1 (Hickey et al. 1988, Fitzpatrick & Richards 1991, Peltoketo et al. 1999aand references therein). As mentioned above, in vivo studies using pregnant mare serumgonadotropin-treatment showed that 17HSD/KSR1 expression is related to the maturationand luteinization of follicles, suggesting that FSH may up-regulate expression of theenzyme whereas LH may down-regulate it, at least in rodents (Ghersevich et al. 1994b).The effect of FSH is supported by the results of in vitro studies showing that FSHpartially prevents the decrease in expression of 17HSD/KSR1 in cultured rat granulosacells (Ghersevich et al. 1994c). The stimulatory effect of FSH appears to be mediated bya cAMP-dependent pathway, since a cAMP analog and forskolin mimic the affect of FSH(Ghersevich et al. 1994c). The PKA-dependent up-regulation of 17HSD/KSR1expression is further modulated by protein kinase C (PKC)-dependent inhibition,estrogens, androgens and autocrine/paracrine growth factors present in the ovary(Ghersevich et al. 1994c, Kaminski et al. 1997).
While granulosa cells have a unigue gonadotropin-dependent system for regulation of17HSD/KSR1, the synergy of retinoic acid action with the influence of epidermal growthfactor and PKA and PKC activators results in an increase of 17HSD/KSR1 concentrationin human trophoblast-like cells (Piao et al. 1997, Peltoketo et al. 1999a and referencestherein). In T-47D breast cancer cells, epidermal growth factor and PKA and PKC

35
activators decrease or lack influence on retinoic acid-stimulated 17HSD/KSR1expression, further pointing to tissue-specific regulation of 17HSD/KSR1 expression(Piao et al. 1997). According to Poutanen et al. (1990, 1992b), expression of the type 1enzyme in the same cell line is up-regulated by progestins and the effect is mediatedthrough PR. Similarly to the breast cancer cell line, expression of 17HSD/KSR1 in theendometrium has been suggested to be up-regulated by P, since the concentration of theenzyme is higher in the secretory than in the proliferative phase (Mäentausta et al. 1990,Mäentausta et al. 1991). This is supported by the results of a study showing that anantiprogestin, mifepristone, prevented expression of 17HSD/KSR1 in the endometrium,in 8 out of 10 subjects (Mäentausta et al. 1993).
2.6.3 17HSD/KSR type 2
2.6.3.1 Human 17HSD/KSR2
The human HSD17B2 gene has been mapped to chromosome region 16q24 (Casey et al.1994) and it encodes two alternatively spliced mRNAs. The mRNAs give rise to activehuman 17HSD/KSR2, a protein of 387 amino acids, as well as to a related 291-aminoacid protein of unknown function (Labrie et al. 1995). Human 17HSD/KSR2 is anoxidative enzyme that prefers NAD+ as a cofactor and it catalyzes C18 and C19 steroidsequally well. More specifically, the type 2 enzyme converts E2, T, DHT and A-diol to E1,A-dione, 5α-A-dione and DHEA, respectively. In addition, it catalyzes reduction of 20-OH-P to P, but less efficiently than the other substrates (Wu et al. 1993, Puranen et al.1999). 17HSD/KSR2 protein is bound to the microsomal fraction in vitro (Wu et al.1993). In line with this, the primary structure of human 17HSD/KSR2 has been found tocontain a putative N-terminal signal sequence followed by a hydrophobic region that isapparently a transmembrane domain. The C-terminus contains a putative motif forendoplasmic reticulum retention (Wu et al. 1993). In addition, the protein has beenlocalized in the endoplasmic reticulum, by using a double immunofluorescence labelingtechnique (Puranen et al. 1999). The study also confirmed that the putative N-terminalsignal sequence is not cleaved.
Human 17HSD/KSR2 mRNA, 1.5 kb in size, is mostly expressed in the placenta,endometrium, liver and small intestine. Using Northern blot, faint to moderate signals for17HSD/KSR2 have also been detected in the kidney, pancreas, colon, normal prostate,benign prostatic hyperplasia and in prostatic carcinoma (Casey et al. 1994, Elo et al.1996, Miettinen et al. 1996a). In addition, human 17HSD/KSR2 has been detected insebaceous glands (Thiboutot et al. 1998). In endometrium, expression of the type 2enzyme has been localized to the glandular epithelium (Casey et al. 1994, Mustonen etal. 1998a), and in the intestine, to the surface epithelium of the villi (Mustonen et al.1998a). In the placenta, it is expressed in cytotrophoblasts (Mustonen et al. 1998a), or,according to a contradictory report, in endothelial cells of certain blood vessels

36
(Takeyama et al. 1998). In the liver, it has been reported to be expressed in hepatocytes(Takeyama et al. 1998).
The 17HSD/KSR2 enzyme may primarily be involved in decreasing the stimulatoryeffects of highly active 17β-hydroxysteroids, even in tissues not classified as traditionalsex steroid target tissues. The presence of the type 2 enzyme in the placenta may serve toinactivate both E2 and T in order to protect the fetus from excess estrogen and androgen(Moghrabi et al. 1997, Mustonen et al. 1997b, Mustonen et al. 1998a). The capability of17HSD/KSR2 to convert 20α-OH-P to P, and the role of this activity in the maintenanceof pregnancy and P concentrations in the maternal circulation remain to be studied.
In the endometrium the expression of 17HSD/KSR2 mRNA is regulated according tothe menstrual cycle. The concentration of the mRNA is greater in the secretory than in theproliferative phase and is highest during the mid- to late secretory phase, when plasma Plevels are at a maximum, suggesting that the expression is stimulated by P (Casey et al.1994). The predominant oxidative 17HSD activity in the endometrium (Tseng 1980) andlower expression of 17HSD/KSR1 than of 17HSD/KSR2 indicate that the type 2 enzymeis the principal 17HSD/KSR in the endometrium (Casey et al. 1994). Thus,17HSD/KSR2 together with decreased ER concentrations (Fleming et al. 1980) andincreased steroid sulfotransferase activity, may participate in the progestin-induced down-regulation of estrogen action during the secretory phase (Tseng 1980 and referencestherein). Interestingly, inactivation of E2 has been assumed to be impaired inendometriotic tissues due to deficient expression of 17HSD/KSR2 (Zeitoun et al. 1998).Greater expression of 17HSD/KSR2 in benign prostatic hyperplasia than in prostaticcarcinoma also indicates a protective role of the enzyme (Elo et al. 1996). Moreover, thewide expression of the type 2 enzyme in the gastrointestinal and urinary tracts (seesection 2.6.3.2) indicates that the protein may be one of the enzymes in the intestine andliver that is involved in the rapid degradation and excretion of steroids and steroid-likecompounds originating from nutriments, for example (Mustonen et al. 1998b, Peltoketoet al. 1999a).
2.6.3.2 Rodent 17HSD/KSR2
Mouse and rat 17HSD/KSR2, consisting of 381 amino acids and having a molecular massof approximately 42.0 kDa, have been cloned from liver and placenta, respectively(Akinola et al. 1996, Mustonen et al. 1997a). In contrast to the human type 2 enzyme, theprimary structure of rodent 17HSD/KSR2 lacks an N-terminal lysine-rich putative signalsequence and a C-terminal lysine-rich endoplasmic reticulum retention signal (Akinola etal. 1996, Mustonen et al. 1997a). However, both rodent and human type 2 enzymes havean arginine-rich motif in the C-terminus which is believed to play a role as a retrievalsignal for endoplasmic reticulum-resident membrane proteins (Mustonen et al. 1997a).The activity of rodent 17HSD/KSR2 has been characterized in cultured cells, where itcatalyzes oxidation of E2 and T with an efficacy similar to that of the human enzyme(Akinola et al. 1996, Mustonen et al. 1997a).

37
The expression pattern of 17HSD/KSR2 in rodent tissues is similar to that in humantissues, being highest in the placenta, small intestine and liver. An exception is the uterus,where the rodent enzyme has not been detected (Akinola et al. 1996, Mustonen et al.1997a, Mustonen et al. 1998b). Expression of the type 2 enzyme in the liver and smallintestine is constitutive throughout the life span of rats, including gestation. In theplacenta, the enzyme is modestly expressed in early and mid-pregnancy but it is up-regulated toward the end of pregnancy (Akinola et al. 1997).
Cell-specific expression of 17HSD/KSR2 has been studied in more detail in mice,where the enzyme is expressed in epithelial cells of various types, such as stratifiedsquamous epithelium of the esophagus, the surface epithelial cells of the stomach, smallintestine, and colon, and the epithelium of the urinary bladder (Mustonen et al. 1998b).Mouse type 2 enzyme is also abundantly expressed in the hepatocytes of the liver andthick limbs of the loops of Henle in the kidneys. In addition, the transcript can bedetected in the seminiferous tubules, and in the sebaceous glands of the skin (Mustonen etal. 1998b). Another cell-specific expression study has been conducted with the mousefetus and developing placenta (Mustonen et al. 1997b). In the choriovitelline placenta, theenzyme is expressed both in mural and polar giant cells. In mid-gestation it is present atthe junctional zone, whereas towards the end of pregnancy the expression shifts mainly tothe labyrinth region of the placenta. In addition, the transcript can be found in the visceralyolk sac throughout pregnancy. In the fetus the expression is congruent with that in adultmice. It is expressed in the liver from day 11, in epithelial cell layers of the esophagusand intestine from day 12, in the stomach and kidney from day 15, and in the oropharynx,nasopharynx and tongue from day 16. In conclusion, the expression pattern of mouse17HSD/KSR2 in the developing placenta and fetus suggests a role for the enzyme inmaintaining a barrier to the highly active 17β-hydroxy forms of sex steroids betweenfetus and mother. This in turn could be vital for the normal development of the fetus aswell as for the normal progression of pregnancy (Mustonen et al. 1997b).
2.6.4 17HSD/KSR type 3
Human 17HSD/KSR3 is a 310-amino acid microsomal enzyme which catalyzes mostefficiently the reaction from A-dione to T (Geissler et al. 1994). In addition, it is to alesser extent able to catalyze reduction of E1 and DHEA to E2 and A-diol, respectively. Asa cofactor the enzyme prefers NADP(H) over NAD(H). The human HSD17B3 gene,consisting of 11 exons, is located at chromosome region 9q22. A 1.3-kb transcript of thetype 3 enzyme is almost exclusively expressed in the testis (Geissler et al. 1994,Miettinen et al. 1996a). Using RT-PCR, 17HSD/KSR3 mRNA has been found in aSertoli-Leydig cell tumor adjacent theca lutein ovarian tissue (Barbieri & Gao 1997) andin adipose tissue (Corbould et al. 1998). Expression of the type 3 enzyme has not beenfound in other tissues having androgenic 17HSD/KSR activity such as the placenta andliver (Miettinen et al. 1996a, Labrie et al. 1997).
The physiological role of 17HSD/KSR3 is biosynthesis of T (Geissler et al. 1994),which is needed with Müllerian inhibiting substance and DHT to induce male sexual

38
differentiation during fetal development (Conte & Grumbach 1991, Wilson et al. 1993).This is evidenced by mutations found in the HSD17B3 gene which lead to decreased Tformation in the fetal testis and consequently to a human intersex disorder termed malepseudohermaphroditism (Geissler et al. 1994). The disorder can also be caused by otherdefects in androgen action, mainly mutations in the androgen receptor gene (McPhaul etal. 1993) and in the steroid 5α-reductase 2 gene (Wilson et al. 1993), which encodes anenzyme responsible for the conversion of T to DHT in the urogenital tract. 17HSD/KSR3deficiency is an autosomal and recessive disorder affecting males, whereas females areasymptomatic (Rösler et al. 1996, Mendonca et al. 1999).
The characteristic phenotype of pseudohermaphroditism is a 46,XY individual withtestes and male Wolffian duct-derived urogenital structures such as epididymides, vasdeferens, and seminal vesicles, but with female external genitalia (Conte & Grumbach1991, Geissler et al. 1994). At puberty, these individuals often start to show signs ofvirilization. At that time the concentration of A-dione in the plasma is characteristicallytenfold elevated. The plasma T levels are higher than in 46,XX women but may be in thenormal male range. Interestingly, virilization at puberty and almost normal developmentof Wolffian ducts suggests residual A-dione to T activity, possibly catalyzed by another17HSD/KSR (Andersson et al. 1996 and references therein). In studies of 17HSD/KSR3deficiency, several mutations have been found in the 17HSD/KSR3 gene. These includemissense mutations and mutations causing a splicing error which inactivate17HSD/KSR3 either severely or completely (Geissler et al. 1994, Moghrabi & Andersson1998).
17HSD/KSR3 has been cloned from mouse (Sha et al. 1997) and rat tissues (Tsai-Morris et al. 1999). In mice, the type 3 enzyme is expressed in fetal, postnatal and adulttestes, as shown by RT-PCR analysis (Sha et al. 1996). In the adult testis, mouse17HSD/KSR3 transcripts have been localized in the interstitial tissue, most likely inLeydig cells (Baker et al. 1997). Using RT-PCR, mouse 17HSD/KSR3 has also beendetected in many other tissues, such as the ovary and prostate (Sha et al. 1997). Similar tothe human enzyme, rat 17HSD/KSR3 is almost exclusively expressed in the testis. The ratenzyme catalyzes most efficiently the reaction from A-dione to T, and to a lesser extent,DHEA to A-diol (Tsai-Morris et al. 1999). In contrast to the human enzyme, the rat type3 enzyme does not possess estrogenic 17KSR activity. Interestingly, 17KSR activity of rat17HSD/KSR3 is increased by glucose and its expression is down-regulated by hCG inLeydig cells (Tsai-Morris et al. 1999). It has been shown that activation of the glycolyticpathway is needed to meet ATP requirements for maximal A-dione to T 17KSR activity(Khanum et al. 1997).
2.6.5 17HSD/KSR type 4
17HSD/KSR4 is peroxisomal multifunctional protein II (MFP-II), with several differentnames (see review de Launoit & Adamski 1999). It was first purified and cloned fromporcine tissues (Adamski et al. 1992, Leenders et al. 1994). 17HSD/KSR4/MFP-II isprimarily translated as an 80-kDa protein from a 3-kb transcript (Leenders et al. 1994).

39
The protein is later cleaved to approximately 32-kDa and 45-kDa fragments havingdistinct activities (Adamski et al. 1992, Dieuaide-Noubhani et al. 1996, Dieuaide-Noubhani et al. 1997). In addition to porcine tissues, this multifunctional and widelyexpressed protein has been cloned from human (Adamski et al. 1995), mouse (Normandet al. 1995) and rat tissues (Corton et al. 1996, Dieuaide-Noubhani et al. 1996, Qin et al.1997a), for example. The similarity of the protein between different species is very high,around 80 %, and they all have the same multidomain structure (de Launoit & Adamski1999).
17HSD/KSR4/MFP-II has been found to be expressed in almost all tissues examined,being highest in the liver and kidney. In addition, its 3-kb mRNA is expressed in severalcancer cell lines (Adamski et al. 1995, Normand et al. 1995, Qin et al. 1997a, Möller etal. 1999). Expression of the rat enzyme has been shown to be induced by peroxisomalproliferator chemicals in the liver and kidney and this induction was dependent onperoxisome proliferator-activated receptor α (Corton et al. 1996, Fan et al. 1998). Inporcine endometrium, the enzyme has been suggested to be up-regulated by P (Kaufmannet al. 1995, Markus et al. 1995).
Based on homology and the results of functional studies, the 17HSD/KSR4/MFP-IIpolypeptide can be divided into three different regions. The N-terminal domain, includingthe first 300 amino acids, is similar to that of several members of the SDR family,especially two dehydrogenase domains of a yeast enzyme participating in peroxisomal β-oxidation of fatty acids (Adamski et al. 1995, Normand et al. 1995, Leenders et al. 1996,Qin et al. 1997a). In line with sequence identity, the N-terminal domain of17HSD/KSR4/MFP-II catalyzes dehydrogenase reactions of fatty acids, 3-hydroxyacyl-CoA esters. Therefore, a role in the β-oxidation of fatty acids has been suggested forMFP-II (Dieuaide-Noubhani et al. 1996, Leenders et al. 1996, Qin et al. 1997a). The 3-hydroxyacyl-CoA dehydrogenase activity of 17HSD/KSR4/MFP-II is D-specific whilethat of MFP-I is L-specific (Qin et al. 1997a). The central region, corresponding to aminoacids 343–607 in the human protein, shows moderate identity with the C-terminalhydratase domain of the same yeast protein (Adamski et al. 1995, Normand et al. 1995,Leenders et al. 1996, Qin et al. 1997a). In agreement with this, the central/2-enoyl-CoAhydratase domain catalyzes hydration of intermediates of bile acid synthesis (Qin et al.1997b). The least characterized domain is the C-terminal region of 17HSD/KSR4/MFP-II(residues 596–736 in the human protein). It shares 39 % identity with human sterolcarrier protein 2 which is assumed to participate in the intracellular transport of sterolsand lipids (Adamski et al. 1995, Normand et al. 1995, Leenders et al. 1996, Qin et al.1997a). The C-terminal domain of the porcine protein has been shown in vitro to transfer7-dehydrocholesterol and phosphatidylcholine between membranes, similarly to humansterol carrier protein 2 (Leenders et al. 1996).
The 17HSD activity of 17HSD/KSR4/MFP-II is an interesting enigma.17HSD/KSR4/MFP-II has been shown to be able to convert E2 to E1 to some extent(Adamski et al. 1995, Carstensen et al. 1996), but studies with the rat protein have shownthat 17HSD activity is remarkably lower than 3-hydroxyacyl-CoA dehydrogenase activity(Dieuaide-Noubhani et al. 1996, Qin et al. 1997a). In spite of that, the Michaelis-Mentenconstant (Km) for E2 is similar to those which other 17HSD/KSRs have for their preferredsubstrates (de Launoit & Adamski 1999). However, in a recent study no correlationbetween 17HSD/KSR4/MFP-II expression and 17HSD activity (E2→E1) was seen in

40
various cell lines (Miettinen et al. 1999). Additional studies are needed to find out if17HSD/KSR4/MFP-II has any role in estrogen metabolism.
Direct evidence concerning the role of 17HSD/KSR4/MFP-II in peroxisomal β-oxidation has come from the discovery that mutations in the HSD17B4 gene encodingMFP-II cause serious peroxisomal disorders. Mutations have been found both in D-hydroxyacyl-CoA dehydrogenase (van Grunsven et al. 1998, van Grunsven et al. 1999a)and enoyl-CoA hydratase regions (Suzuki et al. 1997, van Grunsven et al. 1999b).Mutations lead to accumulation of intermediates of bile acid metabolism and/or very-long-chain fatty acid metabolism which influence renal function and neuronaldevelopment. A D-hydroxyacyl-CoA dehydrogenase deficiency patient has a Gly16→Sermutation which resides in an important loop of Rossman fold forming the NAD+-bindingsite (van Grunsven et al. 1998). It is thus not surprising that the mutation also inactivates17HSD activity of the protein (Möller et al. 1999).
2.6.6 17HSD/KSR type 5
Human 17HSD/KSR5 belongs to the aldoketoreductase family and is highly homologouswith type 1 (84 %) and type 3 (86 %) 3α-HSD as well as with 20α-HSD (88 %) (Dufortet al. 1999). 17HSD/KSR5 has also been cloned from mouse tissues (Deyashiki et al.1995). The human HSD17B5 gene is located at chromosome region 10p14-15 and thecorresponding mouse gene at chromosome 13, region A2 (Rheault et al. 1999a). Thehuman enzyme has been cloned and characterized as 3α-HSD type 2 (Khanna et al. 1995,Lin et al. 1997), but activity measurements in intact cells showed that both mouse andhuman 17HSD/KSR5 more efficiently catalyze conversion of A-dione to T thanconversion of DHT to 5α-androstane-3α,17β-diol (3α-diol), which represents 3α-HSDactivity (Dufort et al. 1999). The sequence of the human type 5 enzyme reported byKhanna et al. (1995) shows a two-amino acid difference from the one cloned by Dufort etal. (1999). However, change of these two amino acids did not significantly affect17HSD/KSR activity (Dufort et al. 1999). The 17HSD/KSR activity of the human type 5enzyme, but not that of the mouse counterpart, is very labile, since the activity in cellhomogenates is only 10 % of that observed in intact cells. However, 3α-HSD activitydoes not decrease that much upon homogenization. Therefore, if enzymatic properties of17HSD/KSR5 were measured only in vitro, the protein would erratically be considered as3α-HSD and not 17HSD/KSR (Dufort et al. 1999).
Human and mouse 17HSD/KSR5 show differences in their substrate specificitypatterns. The mouse type 5 enzyme was originally reported to catalyze the reaction fromE2 to E1 (Deyashiki et al. 1995), but in a recent study using cultured cells it was shownthat transformation of E2 to E1 represents only 1 % of the rate of conversion of A-dione toT, which is the highest activity (Dufort et al. 1999, Rheault et al. 1999b). The mouseenzyme is also able to catalyze conversions of E1, 5α-A-dione, androsterone (ADT),DHEA and P to E2, DHT, 3α-diol, A-diol and 20-OH-P respectively (Dufort et al. 1999,Rheault et al. 1999b). In addition, mouse 17HSD/KSR5 has been reported to be activetoward xenobiotic substrates in vitro (Deyashiki et al. 1995). Human 17HSD/KSR5 also

41
efficiently converts A-dione to T, but most efficiently it catalyzes P to 20-OH-P. Inaddition, it is able to catalyze conversion of T to A-dione and interconversion betweenADT and 3α-diol (Dufort et al. 1999). Furthermore, it has 3α-HSD activity towards ADTand 3α-diol (Lin et al. 1997, Dufort et al. 1999).
Results of RNase protection assays show that human 17HSD/KSR5 is abundantlyexpressed in the liver, prostatic cancer cells and osteocarcinoma cells, in addition towhich it is modestly expressed in normal prostate and in the adrenal (Dufort et al. 1999).Mouse 17HSD/KSR5 is also abundantly expressed in the liver (Deyashiki et al. 1995). Inthe human prostate 17HSD/KSR5 is mainly expressed in the basal cells of glandularepithelium, in stromal fibroblasts and in endothelial cells (El-Alfy et al. 1999). Animmunohistochemical study has shown that human 17HSD/KSR5 is also expressed in theCL, in the epithelial cells of the endometrium and mammary gland, in Leydig cells andoccasionally in Sertoli and germ cells (Pelletier et al. 1999). This relatively wide tissuedistribution is in line with a previous report showing that several human and rhesusmonkey tissues have androgenic 17KSR activity (Labrie et al. 1997).
2.6.7 17HSD/KSR type 6
17HSD/KSR6 cDNA has so far been cloned only from rat prostate (Biswas & Russell1997). Rat 17HSD/KSR6 is a protein of 327 or 317 amino acids depending on whichAUG codon is used to encode for the first methionine. In vitro, the enzyme mostefficiently catalyzes oxidation of 3α-diol to ADT, which is part of the inactivation path ofDHT. DHT is first reduced at the 3-keto group by 3α-HSD to yield 3α-diol which is thenoxidized at the 17β-hydroxy group to ADT by 17HSD/KSR6. As a cofactor the enzymeuses NAD+. The enzyme is mainly expressed in the ventral prostate and liver, in additionto which a 1.5-kb transcript is modestly expressed in the kidney. In castrated rats, thelevel of 17HSD/KSR6 mRNA was decreased. 17HSD/KSR6 belongs to the SDR familyand has its greatest sequence identity (65 %) with retinol dehydrogenase 1. Retinoic acidshave been found to inhibit the reaction from 3α-diol to ADT, suggesting that they couldbe potential substrates. However, the concentrations of retinoic acids used in theexperiment were above physiological levels, i.e. µM vs. nM (Biswas & Russell 1997).
2.6.8 17HSD/KSR type 8
17HSD/KSR8 cDNA was cloned from a mouse kidney cDNA library and initially giventhe name ke 6 (Aziz et al. 1993). Five years later ke 6 was reported to be a 17HSD/KSR,with the highest identity (35 %) to 17HSD/KSR4 (Fomitcheva et al. 1998). 17HSD/KSR8 is slightly smaller than other 17HSD/KSRs, consisting of 260 amino acids andhaving a molecular mass of 34 kDa (Aziz et al. 1993, Aziz et al. 1996, Fomitcheva et al.1998). In vitro characterization of the enzymatic nature of 17HSD/KSR8 has shown thatit catalyzes oxidation of E2, T and DHT, and reduction of E1, using NAD(H) as a cofactor

42
(Fomitcheva et al. 1998). The enzyme has the highest affinity for E2 (Km = 0.11 µM). Theresults of kinetic studies thus suggest that E2 inactivation may be the main physiologicalfunction of the enzyme (Fomitcheva et al. 1998).
The Ke6 gene encoding mouse 17HSD/KSR8 consists of nine exons and it resides inmouse chromosome 17, among genes for a major histocompatibility complex (Maxwellet al. 1995). The gene is transcribed into two mRNAs, 1.0 kb and 1.2 kb in size. The 1.0-kb transcript is most abundantly expressed in the kidney and liver, in addition to which itis moderately expressed in the ovary, testis, spleen, brain, heart and intestine (Aziz et al.1993, Maxwell et al. 1995, Fomitcheva et al. 1998). An immunohistochemical study hasshown that in the ovary 17HSD/KSR8 is expressed in the cumulus cells and in theendothelial cells of blood vessels (Fomitcheva et al. 1998). The 1.2-kb transcript isexpressed in the spleen (Aziz et al. 1993, Maxwell et al. 1995). This second transcript isa splicing variant, where exon 9 is replaced by intron 8, leading to an altered amino acidsequence in the C-terminus of the protein (Maxwell et al. 1995). The biological impact ofthe 1.2-kb transcript is unknown. It is of interest that compared with normal mice thereare decreased 17HSD/KSR8 transcript and protein concentrations in the kidneys of micehaving various cystic kidney diseases which are considered to be models of humanheritable polycystic kidney disease (Aziz et al. 1993, Maxwell et al. 1995, Aziz et al.1996). Down-regulation of 17HSD/KSR8 expression has recently been suggested to bedue to impaired gene regulation and not because of mutation in the Ke6 gene (Ramirez etal. 1998).
Human 17HSD/KSR8 cDNA has also been cloned from kidney tissue (Ando et al.1996). Its gene consists of eight exons and it is located at chromosome region 6p21(Kikuti et al. 1997), where the gene for human autosomal recessive polycystic kidneydisease has been mapped. Characterization of the enzymatic nature of human17HSD/KSR8 has not yet been reported. A 1-kb transcript encoding human type 8enzyme is well expressed in the liver and pancreas, and it is expressed at moderate levelsin the kidney and skeletal muscle. The enzyme is also weakly expressed in the heart,placenta and lung (Kikuti et al. 1997).
The physiological role of 17HSD/KSR8 is unknown, but a protective role as aregulator of sex steroid levels, especially in the kidney, which is a highly steroid-sensitiveorgan, has been suggested (Fomitcheva et al. 1998). This is supported by the results of astudy showing that antisense oligonucleotide-mediated inhibition of expression of theenzyme caused formation of renal cysts in embryonic kidney organ cultures (Aziz et al.1995).
2.6.9 Other 17HSD/KSRs
Recently, 17β/3α-hydroxysteroid/retinoid short chain dehydrogenase/reductase, a novelprotein having 17HSD/KSR activity, has been cloned from mouse tissues (Su et al. 1999).It has greatest identity (88 %) with rat 17HSD/KSR6, but the two proteins differ insubstrate specificity. Using NAD+ as a cofactor the enzyme most efficiently catalyzes thereactions from E2 to E1 and 3α-diol to ADT, and then ADT to 5α-A-dione, thus showing

43
3α-HSD activity. In addition, the enzyme is able to convert all-trans-retinol to all-trans-retinal. The enzyme, however, is not believed to function in the pathway of retinoic acidbiosynthesis, while its physiological role has been suggested to be inactivation ofestrogen and a variety of androgens in the liver, where it has been reported to beexclusively expressed (Su et al. 1999).
Other examples of proteins having 17HSD/KSR activity are a protein from fungus(Lanisnik Rizner et al. 1999) and human brain short chain L-3-hydroxyacyl coenzyme Adehydrogenase, which has been shown to convert E2 to E1 (He et al. 1999). Whether E2 isthe physiological substrate for these enzymes is not yet known.

3 Outlines of the present study
17HSD/KSR1 catalyzes conversion of E1 to E2, which is biologically the most activenatural estrogen and which is mainly synthesized in the ovary and placenta. E2 has animportant role in female reproduction, including folliculogenesis. This study was aimedto characterize enzymatic properties of rodent 17HSD/KSR1 and its physiological roleand regulation in ovaries. In addition, an expression cloning strategy was used todemonstrate that rodents possess a novel type of 17HSD/KSR, which is involved in E2
biosynthesis in ovaries as well as in other tissues, as determined by studying cell-specificexpression of the enzyme. Specific aims of the study were:
1. to clone rat 17HSD/KSR1 cDNA and use it in studies concerning expression andregulation of the rat type 1 enzyme in the ovary,
2. to clone mouse 17HSD/KSR1 cDNA, and characterize the mouse type 1 enzyme andits tissue distribution,
3. to expression-clone a novel type of mouse 17HSD/KSR cDNA and study thecatalytic properties of the enzyme encoded by the cloned cDNA, named17HSD/KSR7, and
4. to characterize the tissue distribution and cell-specific expression of 17HSD/KSR7 inrodents and thereby elucidate the role of the type 7 enzyme in estrogen biosynthesis.

4 Materials and methods
Detailed descriptions of the materials and methods are presented in the original articles, I-IV.
4.1 Isolation of RNAs (I-IV)
Total RNA was isolated by homogenization of samples in a guanidine-isothiocyanatebuffer, followed by CsCl density ultracentrifugation (Davis et al. 1986). For RT-PCR,total RNA was alternatively isolated using an RNeasy total RNA isolation kit (Qiagen).Poly(A)-enriched RNA was extracted from total RNA using oligo-dT-cellulosechromatography (Sambrook et al. 1989). For a cDNA expression library, poly(A)-enriched RNA was isolated using a FastTrack 2.0 kit (Invitrogen).
4.2 Cloning of rat and mouse 17HSD/KSR1 cDNAs (I, II)
A cDNA library, constructed from FSH- and E2-stimulated ovaries of Sprague-Dawleyrats (Clontech), was screened with a [32P]-labeled fragment (nucleotides 1–969) of human17HSD/KSR1 cDNA (Peltoketo et al. 1988) and a rat 17HSD/KSR1 cDNA fragment(nucleotides 374–767) was obtained. To clone cDNA containing the whole coding region,the same cDNA library was screened with the rat 17HSD/KSR1 cDNA fragment as aprobe. The cDNA was then cloned into pBluescript KS+ vector (Stratagene) andsequenced.
To clone mouse 17HSD/KSR1 cDNA, a cDNA library was constructed from poly(A)-enriched RNA extracted from the ovaries of diethylstilbestrol- (DES-) treated mice, usinga TimeSaver cDNA synthesis kit (Pharmacia). The library was screened with a [32P]-labeled fragment (nucleotides 374–767) of rat 17HSD/KSR1 cDNA to obtain a firstcDNA fragment (nucleotides 167–1317). The missing 5’-fragment was cloned using a 5’-

46
AmpliFINDER RACE kit (Clontech), from the same RNA fraction. Mouse 17HSD/KSR1cDNA containing the whole coding region was cloned into pBluescript KS+ vector byligating the two sequenced cDNA fragments simultaneously.
4.3 Cloning of mouse and rat 17HSD/KSR7 cDNAs (III)
Mouse 17HSD/KSR7 cDNA was cloned by expression cloning. A cDNA expressionlibrary was generated by Invitrogen in cDNA3.1 plasmids containing cytomegalovirus(CMV) promoter. Poly(A)-enriched RNA extracted from the HC11 cell line (Ball et al.1988) was used for generation of the library. The cDNA library was divided into pools of2500 clones/colonies from which plasmid DNAs were isolated and transfected intohuman embryonic kidney (HEK) 293 cells grown on six-well plates. After transfectionthe medium was replaced with one containing 100 nM unlabeled E1 + 200000 cpm of 3H-labeled E1/ml. After 18 h the media were collected, and the substrates and products wereseparated in a Symmetry C18 reverse-phase chromatography column connected to a highperformance liquid chromatography (HPLC) system (Waters) as described by Puranen etal. (1997b). Radioactivity of steroids was measured by an on-line β-counter and poolsshowing conversion of E1 to E2 were identified. Thereafter, plasmids from positive poolswere retransformed to generate 20 pools of 1000 transformants on the second round ofscreening and 20 pools of 100 transformants on the third round of screening. Plasmidsfrom new pools were transfected into HEK-293 cells to identify a pool where the positiveclone was enriched. At the final stage, positive plasmid pools were retransformed, and100 single colonies were picked up for plasmid isolation on a miniprep scale. Aftertransfection of these 100 minipreps, a pure enriched plasmid encoding 17HSD/KSRactivity was identified and sequenced.
Rat 17HSD/KSR7 cDNA was cloned by RT-PCR on the basis of the sequencepublished as a PRL receptor-associated protein (PRAP) cDNA at that time (Duan et al.1996). A 1013 bp fragment (nucleotides 9–1021, containing the whole coding region)was reverse-transcribed and amplified from total RNA isolated from the ovaries ofpregnant rats (12-day p.c.) using SuperScriptII RT (GIBCO BRL) and Pyrococcusfuriosus DNA polymerase (Stratagene). The cDNA fragment was then cloned intoBluescript KS+ and CMV6 plasmid vectors and sequenced.
4.4 Sequencing and bioinformatics (I-IV)
Rat and mouse 17HSD/KSR1 cDNAs were sequenced from both their strands by thedideoxy termination method described by Sanger et al. (1977), using a T7Sequencing kitand Deaza G/A T7Sequencing mixes (Pharmacia). Other sequencings thereafter wereperformed using an automatic DNA sequencer (ABI Prism 377 DNA sequencer, Perkin-Elmer). Sequence analyses were carried out with a GCG package (Genetics Computer

47
Group, Inc.) on a Unix computer at CSC (Center for Scientific Computing, Espoo) runthrough the internet.
4.5 Measurement of 17HSD/KSR activity of human and mouse
17HSD/KSR1 (II)
17HSD/KSR activity in cultured Sf9 cells was measured in six-well plates. The cellswere infected with baculoviruses (AcNPVs) containing a fragment coding for17HSD/KSR1. After 50 h infection, both reductive (E1→E2, A-dione→T) and oxidative(E2→E1, T→A-dione) 17HSD/KSR activities were measured by adding 2 ml serum-freeTNF-FH medium, containing 10 µM unlabeled and 300000 cpm 3H-labeled substrate. Atfour time points (10, 20, 30, 40 min), the media were collected, and the substrates andproducts were separated in a Sephasil C18 reverse-phase chromatography columnconnected to a SMARTTM System (Pharmacia) (Jantus Lewintre et al. 1994), and theamount of substrate converted was calculated.
In addition, substrate specificities of mouse and human 17HSD/KSR1 were furthercompared in the conditions described above. In these experiments, the medium containedeither 10 µM E1 and different amounts (5–100 µM) of A-dione or 10 µM A-dione anddifferent amounts (5–100 µM) of E1.
4.6 Measurement of 17HSD/KSR activity of mouse and rat
17HSD/KSR7 (III)
The 17HSD/KSR activity of mouse and rat 17HSD/KSR7 was measured under CMV-promoter after transient transfection in HEK-293 cells grown on six-well plates. Aftertransfection the medium was replaced with one containing 100 nM or 1 nM unlabeledsubstrate + 200000 cpm of 3H-labeled substrate/ml. After incubation (2–24 h), the mediawere collected, and the substrates and products were separated as described in section4.3, and the amount of substrate converted was calculated.
For activity measurements in vitro, transfected HEK-293 cells were lysed in 10 mMpotassium phosphate buffer, pH 8, containing 1 mM EDTA, 20 % glycerol, 0.08 % N,N-bis-(3-D-gluconamidopropyl)cholamide (Big Chap, Calbiochem-NovabiochemInternational), 0.05 % bovine serum albumin (BSA), 0.02 % NaN3 andphenylmethylsulfonyl fluoride (87 ng/ml). For determination of Km and maximal velocity(Vmax) values, cell extracts were diluted in 10 mM potassium phosphate buffer, pH 8,containing 1 mM EDTA and 0.05 % BSA, and the samples were then mixed withdifferent amounts of substrate. The enzymatic reactions were started by adding a cofactor(NADP+/NADPH, Boehringer Mannheim) to a final concentration of 1 mmol/l, and thesamples were incubated for an appropriate time at 37 °C. The reactions were stopped byfreezing the reaction mixture quickly in an ethanol-dry ice bath. Steroids were extracted,

48
separated from each other (see section 4.3) and the amounts of substrate formed per minand per total amount of protein were calculated. Kinetic parameters (Km and Vmax) werecalculated by using a GraFit program (Erithacus Software Ltd). The program fits data tothe Michaelis-Menten equation using nonlinear regression analysis.
4.7 Northern blot analysis (I-III)
Samples of RNA were resolved by 1 % gel electrophoresis and transferred overnight tonylon membranes (Hybond N, Amersham) by capillary force. The RNAs were fixed tothe membrane by UV cross-linking. The membranes were prehybridized for 2–4 h in asolution containing 5 x SSPE [0.75 M NaCl, 50 mM sodium phosphate (pH 7.4), 5 mMEDTA], 50 % formamide, 0.1 % BSA, 0.1 % Ficoll, 0.1 % polyvinyl pyrrolidone, 0.5 %SDS and 20 µg or 100 µg salmon sperm DNA/ml at the same temperature as used inhybridization. The membranes were then hybridized overnight with one of the following[32P]-labeled cDNA probes: rat 17HSD/KSR1 (nucleotides 374–767) at 48 °C, ratP450arom at 45 °C, mouse 17HSD/KSR1 (nucleotides 1–1059) at 50 °C, mouse17HSD/KSR7 (nucleotides 33–849) at 42 °C, or rat 17HSD/KSR7 (nucleotides 9–897) at42 °C. After hybridization, the membranes were washed in decreasing salt concentrationsand exposed to Kodak XAR films. In order to estimate the amount and quality of RNAapplied to the gels, the membranes were hybridized at 42 °C with a glyceraldehyde-3-phosphate dehydrogenase (GAPDH) cDNA probe.
4.8 Reverse transcriptase polymerase chain reaction (II, III)
RT-PCR was carried out on 250 ng of total RNA using a Thermostable recombinantThermus thermophilus (rTth) Reverse Transcriptase RNA PCR kit (Perkin Elmer).Amplified fragments corresponded to mouse 17HSD/KSR1 cDNA (nucleotides 45–492)or mouse 17HSD/KSR7 cDNA (nucleotides 351–849). A fraction of each PCRs wasresolved electrophoretically and transferred to nylon membranes. The membranes werehybridized with mouse 17HSD/KSR1 or 17HSD/KSR7 cDNA probes as described insection 4.7. The quality of the RNA used was tested by checking the integrity ofribosomal bands after electrophoresis and/or by performing RT-PCR with mouse GAPDHprimers.

49
4.9 Animal treatments (I, II)
For preparation of a mouse ovarian cDNA library, 75 immature BALB/c mice wereimplanted subcutaneously with DES pellets (Innovative Research of America), which for5 days released 0.25 mg diethylstilbestrol/day.
To study regulation of 17HSD/KSR1 in ovaries, female Sprague-Dawley rats, 26-28days old, were used 2 days after hypophysectomy. One group of rats was implantedsubcutaneously with DES pellets (Innovative Research of America) releasing 1.2 mgDES/day. Rats in this group were then further treated subcutaneously with placebo, FSH,FSH followed by hCG, or hCG alone. Another group of animals were not primed withDES but were injected with vehicle alone [1 % BSA in phosphate-buffered saline (PBS)]or gonadotropins as above.
4.10 Tissue samples (I-IV)
Tissue specimens for RNA isolation were frozen in liquid nitrogen and stored at -70 °Cuntil processed. For immunohistochemistry, tissues excised from animals wereimmediately fixed in 4 % paraformaldehyde-0.5 % glutaraldehyde-PBS at 4 °C for 3 h, in4 % paraformaldehyde-5 % sucrose-PBS at 4 °C for 6 h and in 20 % sucrose-PBSovernight. Samples were thoroughly washed with PBS after each treatment. The fixedsamples were then frozen in isopentane cooled with liquid nitrogen. Sections of 7 µmwere cut and mounted on polylysine-coated glass slides.
For in situ hybridization using rat 17HSD/KSR1 riboprobe, 8–10 µm cryosectionswere cut and mounted on slides coated with aminopropyltriethoxysilane and fixed with4 % paraformaldehyde (Autio-Harmainen et al. 1992). For in situ hybridization usingother probes, tissues excised were immediately fixed overnight in 4 % paraformaldehyde-PBS solution at 4 °C. Next day they were dehydrated with ethanol, and embedded inparaffin (Merck) overnight at 58 °C. Thereafter, 7 µm sections were cut and collected onSuper Frost+ glass slides (Menzel-Gläser).
4.11 Immunohistochemistry (I, II)
Non-specific binding of antibodies was blocked by incubating the slides in 10 % goatserum-PBS at room temperature for 30 min. Immunoaffinity-purified human17HSD/KSR1 antibodies (Mäentausta et al. 1990) in 10 % goat serum-PBS were addedto the tissue sections and incubated at 4 °C overnight. Thereafter, all steps wereperformed at room temperature. The slides were washed 3 x 5 min with PBS, andincubated for 2 h in a 1:50 dilution of biotinylated anti-rabbit antibody (Dakopatts) in10 % goat serum-PBS. After washing, the slides were further treated for 1 h in a 1:50dilution of streptavidin-conjugated FITC (Dakopatts) in 10 % goat serum-PBS. The

50
preparations were then washed 5 x 5 min with PBS, and mounted with a drop of 5 %(w/w) n-propylgallate-glycerol.
4.12 In situ hybridization (I, IV)
A 394-bp fragment (nucleotides 374–767) of rat 17HSD/KSR1 was cloned into pGEM-4Z vector (Promega) and a 499-bp fragment (nucleotides 351–849) of mouse17HSD/KSR7 was cloned into pBluescript KS+ vector. In addition, a 509-bp fragment(nucleotides 185–693) of prolactin-like protein B (PLP-B) cDNA (Müller et al. 1998)was cloned from poly(A)-enriched RNA from placentas of pregnant mice (day 11–13) byRT-PCR and was cloned into pBluescript KS+. The rat 17HSD/KSR1 fragment inpGEM-4Z and the mouse 17HSD/KSR7 fragment in pBluescript KS+, the mouse PLP-BcDNA fragment in pBluescript KS+, a 737-bp fragment (nucleotides 584–1320) of mouse17HSD/KSR2 in SP72 plasmid (Mustonen et al. 1998b) and a 403-bp fragment(nucleotides 1–403) of mouse 17HSD/KSR1 in pBluescript KS+ were used as templatesto transcribe rat 17HSD/KSR1, mouse 17HSD/KSR7, mouse PLP-B, mouse17HSD/KSR2 and mouse 17HSD/KSR1 [α-35S]-labeled riboprobes, respectively.
In situ hybridization using rat 17HSD/KSR1 riboprobe was performed in cryosectionsas previously described (Autio-Harmainen et al. 1992). The in situ hybridization protocolusing other probes and paraffin sections was based on that described by Quéva et al.(1992) with minor modifications described by Mustonen et al. (1998b). Each section pairwas hybridized with an antisense and a sense probe. To visualize cells, sections werestained with Hoechst 33258 or hematoxylin and eosin.

5 Results
5.1 Cloning and characterization of rat and mouse 17HSD/KSR1
cDNAs (I, II)
A cDNA encoding rat 17HSD/KSR1 was cloned from a cDNA library which wasconstructed from FSH- and E2-stimulated rat ovaries. The cloned cDNA, 1.2 kb in size,encodes a protein of 344 amino acids with a calculated molecular mass of 37.0 kDa.Similarly to rat ovary, DES was found to increase the number of secondary follicles andgranulosa cells expressing 17HSD/KSR1 in the mouse ovary. Therefore, to clone mouse17HSD/KSR1 cDNA, a library was prepared from mRNA extracted from the ovaries ofmice treated with DES. A 1.2-kb cDNA clone for mouse 17HSD/KSR1 obtained from thelibrary was not full-length since it lacked the region coding 45 N-terminal amino acids.The missing region was cloned using the RACE technique. The whole cDNA, 1317 bp insize, similarly to the rat, encoded a protein of 344 amino acids with a calculatedmolecular mass of 36.8 kDa.
The overall amino acid identity between rat and human 17HSD/KSR1 was 68 %. Theidentity of rat 17HSD/KSR1 was further proved by comparing hydropathy plots of thehuman and rat enzymes. These were very similar apart from the C-terminal region.Between the mouse and rat enzyme, the identity was 93 % and between the threeproteins, 63 %. The lowest identity was localized to the C-terminus, which was 17 aminoacids longer in the rodent enzymes than in the human counterpart. Like human17HSD/KSR1, both rodent enzymes belong to the SDR family and they contain fiveconserved regions (A-E) typical of the SDRs characterized at that time (Krozowski 1992,Krozowski 1994) and the consensus sequence of SDR family members in the PROSITEdata bank. The consensus sequence includes a strictly conserved Tyr-X-X-X-Lyssegment, in addition to which there is a Gly-X-X-X-Gly-X-Gly segment near the N-terminus, also typical of the SDRs. The former segment is part of the catalytic center,while the latter binds cofactor. Furthermore, a potential cAMP-dependentphosphorylation site, Ser134, shown to be phosphorylated in the human enzyme (Barbieriet al. 1994, Puranen et al. 1997b), is also present in mouse and rat 17HSD/KSR1. Like

52
rat 17HSD/KSR1 cDNA, the mouse cDNA contains an Alu-like or B1-like repetitivesequence in the antisense strand of the 3’-noncoding region.
5.2 Expression of 17HSD/KSR1 in rat ovary and regulation by
gonadotropins (I)
Expression and regulation of 17HSD/KSR1 in the ovaries of immature rats were analyzedusing immunohistochemistry, Northern blots and in situ hybridization. In the ovaries ofhypophysectomized but otherwise untreated animals, 17HSD/KSR1 was modestly butexclusively expressed in the granulosa cells of a few small antral follicles. One to twodays of FSH treatment clearly induced 17HSD/KSR1 expression in the granulosa cells ofgrowing antral follicles, whereas one-day treatment of FSH-primed animals with hCGcaused luteinization of follicles and a lower expression level of 17HSD/KSR1. Similarlyto FSH, DES strongly up-regulated expression of the type 1 enzyme. In contrast tohypophysectomized but otherwise untreated animals, in DES-primed rats FSH causedluteinization of follicles and a decrease of enzyme expression, starting from the basallayer of granulosa cells. In line with this, 17HSD/KSR1 was not expressed in the rat CL.In addition, 17HSD/KSR1 was not detected in either theca cells or stromal tissues.
Expression of P450arom in the ovaries was also followed. Similar to the effect on17HSD/KSR1, FSH stimulated the expression of P450arom in hypophysectomizedimmature rats. However, in contrast to the down-regulation of 17HSD/KSR1 by hCG,expression of P450arom was strongly increased after one-day treatment with hCG. DESalone did not stimulate P450arom expression but it enhanced the effect of FSH onP450arom expression.
5.3 Characterization of 17HSD/KSR activity of mouse 17HSD/KSR1
and comparison with the human enzyme (II)
Mouse and human recombinant 17HSD/KSR1 in cultured Sf9 insect cells mainlycatalyzed reductive reactions from E1 to E2 and from A-dione to T. Both enzymescatalyzed E1 to E2 similarly (Fig. 4). However, mouse 17HSD/KSR1 converted A-dione toT at least as efficiently as E1 to E2, while the production rate of T from A-dione catalyzedby the human enzyme was less than 10 % of that catalyzed by the mouse enzyme underthe conditions used. Oxidative activities (E2→E1 and T→A-dione) were minor with bothenzymes. The difference in substrate specificity towards androgens was further confirmedby inhibiting conversion of E1 to E2 with increasing amounts of A-dione and, vice versa.For example, a 10-fold excess of A-dione did not decrease E2 formation catalyzed byhuman 17HSD/KSR1, while E2 formation catalyzed by the mouse enzyme was reducedby 43 % with a fivefold excess of A-dione. In line with this, the minor formation of Tcatalyzed by the human enzyme was decreased close to baseline level by E1 at a

53
concentration of only 50 % that of A-dione. In contrast, the androgenic activity of mouse17HSD/KSR1 was inhibited by 43 % with a fivefold excess of E1.
Fig. 4. Comparison of substrate specificity of mouse and human 17HSD/KSR1 in cultured Sf9cells. Medium containing 10 µM E1 or A-dione was applied to the cells grown on six-wellplates after infection with baculoviruses containing a fragment coding mouse or human type1 enzyme. After 10 min, the media were collected and the amount of product formed wasmeasured.
5.4 Tissue distribution of mouse 17HSD/KSR1 (II)
The expression pattern of mouse 17HSD/KSR1 was studied using Northern blot and RT-PCR analyses. In Northern blots, the probe used recognized in ovarian samples a 1.4-kbmRNA as the main transcript and in addition, low amounts of 2.0-kb and 2.3-kbtranscripts were detected. A very faint signal for the 1.4-kb mRNA was also observed insamples from the uterus and adrenal gland. No expression was detected in other tissuesstudied, e.g. the placenta and mammary gland. Expression of mouse 17HSD/KSR1 in theuterus and adrenal gland was confirmed by RT-PCR, which showed a moderate level ofamplification in RNA samples from the two tissues. In addition, modest amplificationtook place in samples from male liver, testis and female brain.
5.5 Cloning and characterization of mouse 17HSD/KSR7 cDNA (III)
The HC11 cell line, originating from the epithelial cells of the mammary gland of apregnant mouse, was found to possess strong estrogenic 17KSR activity. E1 wasefficiently converted to E2 in cultured cells, whereas A-dione was poorly converted to T.Because the activity profile did not match with mouse 17HSD/KSRs known at that time,

54
and because mouse 17HSD/KSR probes did not recognize any transcripts, the cell linewas assumed to express a novel type of 17HSD/KSR. To clone a corresponding cDNA,an expression cDNA library was prepared from HC11 cells. Based on the ability ofclones to convert E1 to E2 after transfection into cultured HEK-293 cells, two cDNAclones encoding mouse 17HSD/KSR7 were obtained. The longer cDNA, 1756 bp in size,encodes a protein of 334 amino acids with a predicted molecular mass of 37.3 kDa. Thededuced amino acid sequence of the protein was compared with those in a sequencedatabase, which showed that the protein had a 89 % identity to a rat protein called PRLreceptor-associated protein, PRAP. Analysis using the PROSITE databank showed thatmouse 17HSD/KSR7 and PRAP belong to the SDR family and they possess theconserved glycine pattern for cofactor-binding and the three most important catalyticamino acids within the SDR consensus sequence (Ser180, Tyr193 and Lys197).
5.6 Characterization of the enzymatic properties of mouse and rat
17HSD/KSR7 (III)
To characterize the enzymatic activities of mouse 17HSD/KSR7, the cDNA wasexpressed under the CMV promoter in cultured HEK-293 cells. The mouse enzymeefficiently catalyzed only the reductive reaction from E1 to E2. No significant17HSD/KSR or 20α-HSD activity was detected when E2, A-dione, T, P or 20-OH-P wasused as a substrate. Further characterization of the activity was carried out in vitro using ahomogenate of transfected HEK-293 cells. Kinetic parameters (Km and Vmax) weredetermined only for E1 and E2, since activity with A-dione, T, DHEA, A-diol, P or 20-OH-P was almost negligible. Vmax values for E1 and E2 were 556.5 and 616.3 pmolproduct/min/mg total protein, and Km values were 2.28 µM and 12.87 µM, respectively.The calculated catalytic efficiency (Vmax/Km) for E1 was 244 whereas for E2 it was 48. Thecatalytic efficiency values and Km clearly showed that E1 is the preferred substrate for theenzyme. In addition, it was shown that the mouse type 7 enzyme preferred NADPH overNADH as a cofactor. In order to characterize the enzymatic activity of rat17HSD/KSR7/PRAP, its cDNA was cloned by RT-PCR from the ovaries of pregnant ratsand expressed under the CMV promoter in cultured HEK-293 cells. Similarly to themouse enzyme, the rat protein catalyzed only the reaction from E1 to E2 and no activitywas detected when E2, A-dione, T, P or 20-OH-P was used as a substrate.
5.7 Tissue distribution of 17HSD/KSR7 and its expression pattern in
the ovary during gestation (III, IV)
In Northern blot analysis, the mouse 17HSD/KSR7 probe recognized several transcripts,a 4.6-kb mRNA being the dominant form, and others (3.6, 1.7 and 1.2 kb) being weaklydetected. The 4.6-kb transcript was most abundantly expressed in the ovaries of pregnant

55
mice and in the placenta. 17HSD/KSR7 was weakly expressed in the ovaries ofnonpregnant mice, in the mammary gland, liver, kidney, testis and in the HC11 cell line.Because of the remarkable size difference between the major mRNA and the clonedcDNA, and because of the fact that the size of the mRNA was about the same as that of28S ribosomal RNA (4.7 kb), the authenticity of the weak signals in the Northern blotwas confirmed by RT-PCR, which showed that the enzyme was expressed in the tissuesidentified in the Northern blot.
Expression of 17HSD/KSR7 in the ovaries during pregnancy was followed in Northernblots, using RNA samples from rat ovaries. The rat 17HSD/KSR7/PRAP proberecognized two main transcripts, 4.6 kb and 1.8 kb in size, of which the longer transcriptwas the dominant form. 17HSD/KSR7/PRAP was expressed throughout pregnancy butwas remarkably up-regulated at day 8 p.c. and strongly decreased close to parturition, onday 21 p.c.
Expression of the type 7 enzyme in mouse ovary was studied in more detail using insitu hybridization, which showed that the enzyme was exclusively and highly expressedin the CLs of both nonpregnant and pregnant animals. The 17HSD/KSR7 transcript waspresent only in fresh CLs, i.e. in those of the cycle and those of the pregnancy, but not inthose of previous cycle(s), which were regressing. In agreement with the Northern blotresults, the expression level of the type 7 enzyme in the CLs was decreased close toparturition, when they showed clear signs of luteolysis.
5.8 Expression of mouse 17HSD/KSR7 in the uterus and placenta (IV)
Using in situ hybridization, expression of the mouse type 7 enzyme was studied in theuterus and placenta, where cell-specific expression was followed from implantation tolate pregnancy. In the uterus, 17HSD/KSR7 was first detected on day 5½ p.c., whenexpression surrounded the implantation site on the antimesometrial side. Expression wasmore abundant in the inner zone than in the outer zone of the decidua. Expression ofmouse 17HSD/KSR7 was congruent with that of PLP-B, which was used as a marker of adecidual reaction. PLP-B was expressed in the inner zone of the decidua on theantimesometrial side, but not in the outer zone. As gestation progressed, the type 7enzyme was abundantly expressed in the decidua capsularis on day 8 and 9½ p.c., anddisappeared thereafter from declining antimesometrial decidua. On day 9 p.c. onward,expression of 17HSD/KSR7 was observed in the basal layer of the developingchorioallantoic placenta. On days 12½ and 14½ p.c., the enzyme was abundantlyexpressed in the spongiotrophoblasts, where expression gradually declined towardparturition. Similarly, PLP-B, a marker of spongiotrophoblasts, was abundantly expressedin the spongy layer on days 12½ and 14½ p.c., after which its expression declinedsomewhat faster than that of 17HSD/KSR7, being almost undetectable on day 17½ p.c.No specific signals for mouse 17HSD/KSR7 or PLP-B were detected in the giant cells atany stage of pregnancy.

6 Discussion
6.1 Cloning and characterization of mouse and rat 17HSD/KSR1
17HSD/KSR1 was cloned from mouse and rat tissues, and as expected, rodent type 1enzymes have similar amino acid sequences and they belong to the SDR family,containing the glycine pattern for cofactor-binding in the N-terminus, and the mostconserved and important catalytic amino acids. The greatest identity between rodent andhuman 17HSD/KSR1 is in the first 200 amino acids (81 %), which also is typical ofSDRs (Jörnvall et al. 1995). The most striking difference between the human and rodenttype 1 enzymes is their substrate specificity. Human 17HSD/KSR1 mainly catalyzesconversion of E1 to E2, while mouse 17HSD/KSR1 converts A-dione to T as efficiently asE1 to E2. In line with the mouse enzyme, rat 17HSD/KSR1 equally well catalyzesconversion of both E1 and A-dione (Puranen et al. 1997a). Low homology (32 %) in theamino acid region 197–230 between human and rodent 17HSD/KSR1 has been suggestedto partly explain why the rodent type 1 enzyme has broader substrate specificity (Puranenet al. 1997a). The biological significance of this broader substrate acceptance is notknown, but at least it shows that two routes are possible for E2 biosynthesis in the rodentovary: 1) from A-dione to E1 and then to E2, and/or 2) from A-dione to T and then to E2.In the human ovary, the favored route is the former pathway, since theca cells mainlyproduce A-dione (Bergh et al. 1993) and P450arom catalyzes A-dione as efficiently as T(Kellis & Vickery 1987).
Rodent 17HSD/KSR1 is most abundantly expressed in the granulosa cells ofdeveloping follicles (present study, Akinola et al. 1996). According to the Northern blotand RT-PCR analyses, the mouse type 1 enzyme is modestly expressed in the uterus andadrenals. The human type 1 enzyme is also expressed in the endometrium (Mäentausta etal. 1991, Miettinen et al. 1996a, Zeitoun et al. 1998), whereas expression in the adrenalshas not been reported. In contrast to humans, mouse 17HSD/KSR1 is not expressed in themammary gland or placenta, where the human counterpart is expressed (Mäentausta et al.1990, Miettinen et al. 1996a, Söderqvist et al. 1998). Furthermore, using RT-PCR, mouse17HSD/KSR1 was found to be modestly expressed in the liver, testis and brain, which,excluding the liver, has been confirmed by Sha et al., who also used RT-PCR (1997). In

57
addition, Sha et al. (1996, 1997) detected the transcript in several other tissues such as theprostate, fetal and young postnatal testes, heart and kidney, and female kidney and malespleen.
On the basis of these studies and other studies using rodent and human samples(reviewed in Peltoketo et al. 1999a), it can be concluded that in the mouse, 17HSD/KSR1is the main 17HSD/KSR which in the granulosa cells is involved in biosynthesis of E2.The physiological significance, if any, of the low expression of 17HSD/KSR1 in othermouse tissues, as detected by RT-PCR, remains open.
6.2 Expression and regulation of rat 17HSD/KSR1 in granulosa cells
Rat 17HSD/KSR1 is exclusively expressed in the granulosa cells of growing follicles,where FSH stimulates expression of the enzyme, as is the case for P450arom (Adashi1996a, Fitzpatrick et al. 1997). Thus, FSH stimulation of E2 biosynthesis occurs viaenhancement of both enzymes catalyzing the last two reactions of E2 biosynthesis. Inaddition, the maturation stage of granulosa cells most likely affects the response of thecells to gonadotropins, since in hypophysectomized but otherwise untreated animals FSHstimulates expression of 17HSD/KSR1 and P450arom, while in DES-primed animals itcauses down-regulation of the type 1 enzyme. hCG, in turn, stimulated luteinization inFSH-treated granulosa cells, while in DES-treated animals it caused follicular atresia.
Strong up-regulation of 17HSD/KSR1 expression by DES suggests an autocrine rolefor estrogens, establishing a short positive feedback loop that could maintain increasingE2 biosynthesis during folliculogenesis. Interestingly, DES has been found to beineffective as regards 17HSD/KSR1 expression in granulosa cells in vitro, which suggeststhat the estrogen effect may be indirect and may require a paracrine factor or factorsmissing in cell culture conditions (Ghersevich et al. 1994c). Unlike the expression of17HSD/KSR1, that of P450arom is not induced by estrogen alone (Hickey et al. 1988,Fitzpatrick & Richards 1991, present study), but estrogen is able to enhance the FSH-induced expression of P450arom (Fitzpatrick & Richards 1991, present study).
In agreement with decreased E2 biosynthesis taking place upon luteinization, theresults indicate that the ovulatory surge of LH down-regulates expression of17HSD/KSR1 in granulosa cells.
6.3 Cloning and characterization of mouse and rat 17HSD/KSR7, and
its expression pattern in the ovary
Mouse 17HSD/KSR7 shows a 89 % identity with rat PRAP, which has beendemonstrated to be associated with a short form of PRL receptor (Duan et al. 1996).Sequence analyses indicated that both proteins belong to the SDR family and containcatalytic amino acids needed in the 17KSR reaction (present study). Both mouse

58
17HSD/KSR7 and rat PRAP catalyze the reaction from E1 to E2 in vitro and in intactcultured cells. The enzyme was also able to catalyze the reverse reaction in vitro, which isnot surprising, since as in the case of human 17HSD/KSR1 (Puranen et al. 1997a), thedirection of reaction may be reversed by excess of cofactor. Kinetic parameters showedthat the enzyme prefers E1 over E2 as a substrate. As suggested to be typical of reductiveenzymes (Luu-The et al. 1995), mouse 17HSD/KSR7 also more efficiently utilizes thephosphorylated forms of cofactor, NADP(H), than the nonphosphorylatedforms. Altogether, similar primary structures, enzymatic characteristics, and the tissuedistribution of mouse 17HSD/KSR7 and rat PRAP suggest that PRAP is rat17HSD/KSR7.
Recently, a cDNA encoding human 17HSD/KSR7 and its gene located at chromosomeregion 10p11.2 have been cloned (Krazeisen et al. 1999). This uncharacterized protein isreported to be clearly expressed in the placenta and a liver cell line. In addition, it isexpressed in the ovary, mammary gland and testis. Furthermore, based on ExpressedSequence Tag clones in the GenBank, the protein is assumed to be expressed in severaladditional tissues such as fetal liver and spleen (Krazeisen et al. 1999).
Similarly to mouse 17HSD/KSR7, a cDNA probe for rat type 7 enzyme recognizedtwo transcripts, 1.8 kb and 4.3 kb in size, and in both species the longer mRNA is thedominant form. In addition to these two transcripts, a third rat 17HSD/KSR7 mRNA, 5.5kb in size, has been detected by Duan et al. (1997). At least in the case of the mouse, thelong transcript is mostly comprised of a long 3’-noncoding region, probably a result ofalternative polyadenylation (Nokelainen, P., unpublished data). Nevertheless, the resultsof functional studies, and an in-frame stop codon in the 5’-noncoding region, haveconfirmed that the cDNA contains the whole coding area.
In agreement with the results of recent studies on rats (Parmer et al. 1992, Duan et al.1997), this study (III, IV) shows that 17HSD/KSR7 is most abundantly expressed in theovaries, more specifically in the CL of both pregnant and nonpregnant animals. A markedincrease of 17HSD/KSR7 expression around gestation day 8 p.c., and sustained abundantexpression until close to parturition coincide with enhanced androgen secretion from theplacenta (Warshaw et al. 1986), expression of P450arom in the CL (Hickey et al. 1988)and elevated serum E2 concentrations (Taya & Greenwald 1981). This indicates that17HSD/KSR7, not 17HSD/KSR1, is the 17HSD/KSR which is necessary for E2
biosynthesis in the rodent CL. However, an additional role for 17HSD/KSR7 in PRLsignaling cannot be excluded, since rat 17HSD/KSR7 has been found to be associatedwith a short form of PRL receptor, and PRL up-regulates expression of17HSD/KSR7/PRAP (Duan et al. 1997). Furthermore, E2 has been reported to increaseexpression of 17HSD/KSR7 in hypophysectomized rats in vivo (Duan et al. 1997) andPRL up-regulates expression of ERs in the CL (Telleria et al. 1998). It could therefore bepossible that there is cross-talk between E2 and PRL signaling via 17HSD/KSR7.

59
6.4 17HSD/KSR7 in the mouse uterus and placenta
In addition to the CL, mouse 17HSD/KSR7 is expressed in the decidua, more abundantlyin the inner zone of the antimesometrial layer of the decidua. Estrogen is known to play acrucial role in implantation, particularly in rodents (Cross et al. 1994). Together with P, itprimes the uterus for implantation, nidatory estrogen triggers implantation, and inaddition it augments several effects of P on the decidual cell layer after implantation.Since ovariectomy prevents implantation (McLaren 1971, Paria et al. 1993), local E2
biosynthesis possibly occurring in the uterus is not able to replace the ablation of ovarianestrogen. However, E2 produced in the uterus may have a role in stromal proliferation anddifferentiation into decidual cells when E2 produced in the ovaries is not maximal. Indeed,E2-induced cell proliferation is increased on the antimesometrial side of the implantationsite in particular (Yamada & Nagata 1992, Weitlauf 1994), concurrently with theexpression of 17HSD/KSR7. In decidual cells, E2 together with P is known to modulatethe expression of several genes suggested to have a role in decidualization and muraltrophoblast giant cell invasion (Schatz et al. 1994, Schatz et al. 1995, Das et al. 1999).
Expression of the enzyme in the placenta is interesting, since rodent placenta, incontrast to the human placenta, has been assumed to be unable to synthesize E2 as a resultof its lack of P450arom (Rembiesa et al. 1971) and 17HSD/KSR1 (present study). Themouse placenta is rich in 17HSD/KSR2, which is suggested to prevent the transfer ofactive 17β-hydroxy forms of sex steroids between the fetus and the maternal circulation,by converting E2 to E1 (Mustonen et al. 1997a, Mustonen et al. 1997b). However, it hasalso previously been proposed that mouse placenta may express multiple forms of17HSD/KSR (Blomquist et al. 1993). It is therefore possible that 17HSD/KSR7 catalyzesconversion of circulating E1 to E2 in the placenta for local needs. Accordingly, mouseplacenta may not secrete estrogens in the classical endocrine manner, since 17HSD/KSR2can limit the entrance of E2 to the circulation (Mustonen et al. 1997b).
As shown in this study, E2 synthesized by 17HSD/KSR7 in situ, may exert paracrine,autocrine or even intracrine effects in the placenta. This is supported by a report showingthat mouse trophoblasts contain estrogen receptors (Iguchi et al. 1993). In addition,treatment of pregnant rats on day 9 to 11 p.c. with an antiestrogen, tamoxifen, has beendemonstrated to cause severe impairment of decidual development, which is associatedwith altered placental bed vascularization, deteriorated fetoplacental development andincreased incidences of growth-retarded fetuses and fetal death (Sadek & Bell 1996).Hence, estrogen action could be needed to carry rodent placentation successfully tocompletion, and 17HSD/KSR2 and 17HSD/KSR7, which show precise cell-specificexpression, may be needed in the placenta for appropriate regulation of estrogen action.Altogether, the results have shown cell-specific expression of 17HSD/KSR7 in theovaries, placenta and uterus. In rodent ovaries, 17HSD/KSR1 and 17HSD/KSR7 areexpressed in the follicles and CLs, respectively. In the placenta and uterus, 17HSD/KSR7and 17HSD/KSR2 are expressed in adjacent cells. This may reflect part of a system bywhich estrogen action is cell-specifically regulated in these tissues.

7 Summary and conclusions
Overall, 17HSD/KSR1 and -7 were cloned from mouse and rat. Both enzymes catalyzethe reaction from E1 to E2 and are important enzymes in E2 biosynthesis taking place inthe ovary. In detail, the findings of the present study are summarized as follows:
1. This study showed that both rat and mouse 17HSD/KSR1 are expressed in thegranulosa cells of developing follicles, where the expression and thereby E2
biosynthesis increases along with follicular maturation. Estrogen and FSH up-regulate both the maturation of follicles and the expression of 17HSD/KSR1 in them,whereas LH causes luteinization of follicles and down-regulation of the enzyme.Consequently, the rodent type 1 enzyme is not expressed in the CL, indicating thatanother 17HSD/KSR catalyzes the E1 to E2 reaction in luteal cells.
2. A comparison of the enzymatic properties of mouse and human 17HSD/KSR1showed that the mouse type 1 enzyme catalyzes the reaction from A-dione to T atleast equally efficiently as the reaction E1 to E2, while human 17HSD/KSR1 clearlyprefers the E1 to E2 reaction, suggesting that in rodent ovaries E2 can be synthesizedequally well from E1 and T.
3. A novel type of 17HSD/KSR was expression-cloned from a mouse mammaryepithelial cell line and was chronologically named 17HSD/KSR7. Both mouse andrat 17HSD/KSR7 efficiently catalyze the reaction from E1 to E2. 17HSD/KSR7 ismost abundantly expressed in the CL, where the expression parallels E2 secretionfrom the CL. Therefore, it is 17HSD/KSR7 and not 17HSD/KSR1 that participates inE2 biosynthesis in the rodent CL.
4. A cell-specific expression study showed that the mouse type 7 enzyme is alsoexpressed in the decidua surrounding the implantation site, in the decidua capsularisof the developing placenta and in the spongiotrophoblasts of the mature placenta,where the expression was particularly high on days 12½ and 14½ p.c. E2 synthesizedlocally by 17HSD/KSR7 is thus suggested to have a role in decidualization and/orimplantation and placentation.

8 References
Adams JA, Jarabak J & Talalay P (1962) The steroid specificity of the 17β-hydroxysteroiddehydrogenase of human placenta. J Biol Chem 237: 3069-3073.
Adamski J, Husen B, Marks F & Jungblut PW (1992) Purification and properties of estradiol 17β-dehydrogenase extracted from cytoplasmic vesicles of porcine endometrial cells. Biochem J288: 375-381.
Adamski J, Normand T, Leenders F, Monté D, Begue A, Stéhelin D, Jungblut PW & de Launoit Y(1995) Molecular cloning of a novel widely expressed human 80 kDa 17β-hydroxysteroiddehydrogenase IV. Biochem J 311: 437-443.
Adashi EY (1996a) The ovarian follicle: life cycle of a pelvic clock. In: Adashi EY, Rock JA, &Rosenwaks Z (eds) Reproductive Endocrinology, Surgery, and Technology. Lippincott-RavenPublishers, Philadelphia, p 211-234.
Adashi EY (1996b) The ovarian follicular apparatus. In: Adashi EY, Rock JA, & Rosenwaks Z(eds) Reproductive Endocrinology, Surgery, and Technology. Lippincott-Raven Publishers,Philadelphia, p 17-40.
Akinola LA, Poutanen M, Peltoketo H, Vihko R & Vihko P (1998) Characterization of rat 17β-hydroxysteroid dehydrogenase type 1 gene and mRNA transcripts. Gene 208: 229-238.
Akinola LA, Poutanen M & Vihko R (1996) Cloning of rat 17β-hydroxysteroid dehydrogenasetype 2 and characterization of tissue distribution and catalytic activity of rat type 1 and type 2enzymes. Endocrinology 137: 1572-1579.
Akinola LA, Poutanen M, Vihko R & Vihko P (1997) Expression of 17β-hydroxysteroiddehydrogenase type 1 and type 2, P450 aromatase, and 20α-hydroxysteroid dehydrogenaseenzymes in immature, mature, and pregnant rats. Endocrinology 138: 2886-2892.
Albrecht ED, Aberdeen GW & Pepe GJ (2000) The role of estrogen in the maintenance of primatepregnancy. Am J Obstet Gynecol 182: 432-438.
Albrecht ED & Pepe GJ (1990) Placental steroid hormone biosynthesis in primate pregnancy.Endocr Rev 11: 124-150.
Andersson S, Russell DW & Wilson JD (1996) 17β-Hydroxysteroid dehydrogenase 3 deficiency.Trends Endrocrinol Metab 7: 121-126.
Ando A, Kikuti YY, Shigenari A, Kawata H, Okamoto N, Shiina T, Chen, L, Ikemura T, Abe K,Kimura M & Inoko H (1996) cDNA cloning of the human homologues of the mouse Ke4 andKe6 genes at the centromeric end of the human MHC region. Genomics 35: 600-602.
Autio-Harmainen H, Hurskainen T, Niskasaari K, Höyhtyä M & Tryggvason K (1992)Simultaneous expression of 70 kilodalton type IV collagenase and type IV collagen α1 (IV)chain genes by cells of early human placenta and gestational endometrium. Lab Invest 67: 191-200.

62
Aziz N, Brown D, Lee W-S & Naray-Fejes-Toth A (1996) Aberrant 11β-hydroxysteroiddehydrogenase-1 activity in the cpk mouse: implications for regulation by the Ke 6 gene.Endocrinology 137: 5581-5588.
Aziz N, Maxwell MM, St.-Jacques B & Brenner BM (1993) Downregulation of Ke 6, a novel geneencoded within the major histocompatibility complex, in murine polycystic kidney disease. MolCell Biol 13: 1847-1853.
Aziz N, Nearing J, Kriedberg J & Donovan M (1995) Antisense oligonucleotide mediatedinhibition of ke 6 gene expression induces cysts in embryonic kidneys in organ cultures(abstract). J Am Soc Nephrol 6: 690.
Azzi A, Rehse PH, Zhu D-W, Campbell RL, Labrie F & Lin S-X (1996) Crystal structure of humanestrogenic 17β-hydroxysteroid dehydrogenase complexed with 17β-estradiol. Nat Struct Biol 3:665-668.
Baker ME (1995) Unusual evolution of 11β- and 17β-hydroxysteroid and retinol dehydrogenases.Bioessays 18: 63-70.
Baker PJ, Sha JH & O'Shaughnessy PJ (1997) Localisation and regulation of 17β-hydroxysteroiddehydrogenase type 3 mRNA during development in the mouse testis. Mol Cell Endocrinol 133:127-133.
Ball RK, Friis RR, Schoenenberger CA, Doppler W & Groner B (1988) Prolactin regulation of β-casein gene expression and of a cytosolic 120-kd protein in a cloned mouse mammary epithelialcell line. EMBO J 7: 2089-2095.
Barbieri RL & Gao X (1997) Presence of 17β-hydroxysteroid dehydrogenase type 3 messengerribonucleic acid transcript in an ovarian Sertoli-Leydig cell tumor. Fertil Steril 68: 534-537.
Barbieri RL, Gao X & Frost RA (1994) Phosphorylation of 17β-hydroxysteroid dehydrogenase inBeWo choriocarcinoma cells. Am J Obstet Gynecol 171: 223-230.
Batistuzzo de Medeiros SR, Krey G, Hihi AK & Wahli W (1997) Functional interactions betweenthe estrogen receptor and the transcription activator Sp1 regulate the estrogen-dependenttranscriptional activity of the vitellogenin A1 io promoter. J Biol Chem 272: 18250-18260.
Benz CC & Lewis BJ (1991) Hormones & Cancer. In: Greenspan FS (ed) Basic and ClinicalEndocrinology. Prentice-Hall International Inc., East Norwalk, USA, p 701-714.
Bergh C, Olsson J-H, Selleskog U & Hillensjö T (1993) Steroid production in cultured thecal cellsobtained from human ovarian follicles. Hum Reprod 8: 519-524.
Biswas MG & Russell DW (1997) Expression cloning and characterization of oxidative 17β- and3α-hydroxysteroid dehydrogenases from rat and human prostate. J Biol Chem 272: 15959-15966.
Blomquist CH, Hensleigh HC, Block DL & Feeney LA (1993) Placental 17β-hydroxysteroidoxidoreductase, lactate dehydrogenase and malate dehydrogenase during the latter half ofpregnancy in the mouse. J Steroid Biochem Mol Biol 46: 61-67.
Breton R, Housset D, Mazza C & Fontecilla-Camps JC (1996) The structure of a complex ofhuman 17β-hydroxysteroid dehydrogenase with estradiol and NADP+ identifies two principaltargets for the design of inhibitors. Structure 4: 905-915.
Bulun SE, Noble LS, Takayama K, Michael MD, Agarwal V, Fisher C, Zhao Y, Hinshelwood MM,Ito Y & Simpson ER (1997) Endocrine disorders associated with inappropriately high aromataseexpression. J Steroid Biochem Mol Biol 61: 133-139.
Burns DJW, Engel LL & Bethune JL (1971) The subunit structure of human placental 17β-estradiol dehydrogenase. Biochem Biophys Res Commun 44: 786-792.
Burns DJW, Engel LL & Bethune JL (1972) Amino acid composition and subunit structure. Humanplacental 17β-estradiol dehydrogenase. Biochemistry 11: 2699-2703.
Byers M, Kuiper GGJM, Gustafsson J-Å & Park-Sarge O-K (1997) Estrogen receptor-β mRNAexpression in rat ovary: down-regulation by gonadotropins. Mol Endocrinol 11: 172-182.
Caira F, Clémencet M-C, Cherkaoui-Malki M, Dieuaide-Noubhani M, Pacot, C, van Veldhoven PP& Latruffe N (1998) Differential regulation by a peroxisome proliferator of the differentmultifunctional proteins in guinea pig: cDNA cloning of the guinea pig D-specificmultifunctional protein 2. Biochem J 330: 1361-1368.

63
Carstensen JF, Tesdorpf JG, Kaufmann M, Markus MM, Husen B, Leenders F, Jakob F, de LaunoitY & Adamski J (1996) Characterization of 17β-hydroxysteroid dehydrogenase IV. J Endocrinol150: S3-S12.
Casey ML, MacDonald PC & Andersson S (1994) 17β-Hydroxysteroid dehydrogenase type 2:chromosomal assignment and progestin regulation of gene expression in human endometrium. JClin Invest 94: 2135-2141.
Cenni B & Picard D (1999) Ligand-independent activation of steroid receptors: new roles for oldplayers. Trends Endrocrinol Metab 10: 41-46.
Conley AJ & Bird IM (1997) The role of cytochrome P450 17α-hydroxylase and 3β-hydroxysteroid dehydrogenase in the integration of gonadal and adrenal steroidogenesis via the∆5 and ∆4 pathways of steroidogenesis in mammals. Biol Reprod 56: 789-799.
Conte FA & Grumbach MM (1991) Abnormalities of sexual differentiation. In: Greenspan FS (ed)Basic and Clinical Endocrinology. Prentice-Hall International Inc., East Norwalk, USA, p 491-518.
Corbould AM, Judd SJ & Rodgers RJ (1998) Expression of types 1, 2, and 3 17β-hydroxysteroiddehydrogenase in subcutaneous abdominal and intra-abdominal adipose tissue of women. J ClinEndocrinol Metab 83: 187-194.
Corton JC, Bocos C, Moreno ES, Merritt A, Marsman DS, Sausen PJ, Cattley RC & Gustafsson J-Å (1996) Rat 17β-hydroxysteroid dehydrogenase type IV is a novel peroxisome proliferator-inducible gene. Mol Pharmacol 50: 1157-1166.
Couse JF, Hewitt SC, Bunch DO, Sar M, Walker VR, Davis BJ & Korach KS (1999) Postnatal sexreversal of the ovaries in mice lacking estrogen receptors α and β. Science 286: 2328-2331.
Couse JF & Korach KS (1999) Estrogen receptor null mice: what have we learned and where willthey lead us? Endocr Rev 20: 358-417.
Couse JF, Lindzey J, Grandien K, Gustafsson J-Å & Korach KS (1997) Tissue distribution andquantitative analysis of estrogen receptor-α (ERα) and estrogen receptor-β (ERβ) messengerribonucleic acid in the wild-type and ERα-knockout mouse. Endocrinology 138: 4613-4621.
Cowley SM, Hoare S, Mosselman S & Parker MG (1997) Estrogen receptors α and β formheterodimers on DNA. J Biol Chem 272: 19858-19862.
Cross JC, Werb Z & Fisher SJ (1994) Implantation and the placenta: key pieces of the developmentpuzzle. Science 266: 1508-1518.
Cullen KJ & Lippman ME (1989) Estrogen regulation of protein synthesis and cell growth inhuman breast cancer. Vitam Horm 45: 127-172.
Das SK, Lim H, Paria BC & Dey SK (1999) Cyclin D3 in the mouse uterus is associated with thedecidualization process during early pregnancy. J Mol Endocrinol 22: 91-101.
Davis LG, Dibner MD & Battey JF (1986) Preparation and analysis of RNA from eukaryotic cells.In: Basic Methods in Molecular Biology. Elsevier, New York, p 130-135.
Deyashiki Y, Ohshima K, Nakanishi M, Sato K, Matsuura K & Hara A (1995) Molecular cloningand characterization of mouse estradiol 17β-dehydrogenase (A-specific), a member of thealdoketoreductase family. J Biol Chem 270: 10461-10467.
Dieuaide-Noubhani M, Novikov D, Baumgart E, Vanhooren JCT, Fransen M, Goethals M,Vandekerckhove J, van Veldhoven PP & Mannaerts GP (1996) Further characterization of theperoxisomal 3-hydroxyacyl-CoA dehydrogenases from rat liver. Relationship between thedifferent dehydrogenases and evidence that fatty acids and the C27 bile acids di- and tri-hydroxycoprostanic acids are metabolized by separate multifunctional proteins. Eur J Biochem240: 660-666.
Dieuaide-Noubhani M, Novikov D, Vandekerckhove J, van Veldhoven PP & Mannaerts GP (1997)Identification and characterization of the 2-enoyl-CoA hydratases involved in peroxisomal β-oxidation in rat liver. Biochem J 321: 253-259.
Duan WR, Linzer DIH & Gibori G (1996) Cloning and characterization of an ovarian-specificprotein that associates with the short form of the prolactin receptor. J Biol Chem 271: 15602-15607.
Duan WR, Parmer TG, Albarracin CT, Zhong L & Gibori G (1997) PRAP, a prolactin receptorassociated protein: its gene expression and regulation in the corpus luteum. Endocrinology 138:3216-3221.

64
Dufort I, Rheault P, Huang X-F, Soucy P & Luu-The V (1999) Characteristics of a highly labilehuman type 5 17β-hydroxysteroid dehydrogenase. Endocrinology 140: 568-574.
Duque C, Morisaki M & Ikekawa N (1978) The enzyme activity of bovine adrenocorticalcytochrome P-450 producing pregnenolone from cholesterol: kinetic and electrophoretic studieson the reactivity of hydroxycholesterol intermediates. Biochem Biophys Res Commun 82: 179-187.
El-Alfy M, Luu-The V, Huang X-F, Berger L, Labrie F & Pelletier G (1999) Localization of type 517β-hydroxysteroid dehydrogenase, 3β-hydroxysteroid dehydrogenase, and androgen receptorin the human prostate by in situ hybridization and immunocytochemistry. Endocrinology 140:1481-1491.
Elo JP, Akinola LA, Poutanen M, Vihko P, Kyllönen AP, Lukkarinen O & Vihko R (1996)Characterization of 17β-hydroxysteroid dehydrogenase isoenzyme expression in benign andmalignant human prostate. Int J Cancer 66: 37-41.
Engel LL & Groman EV (1974) Human placental 17β-estradiol dehydrogenase: characterizationand structural studies. Recent Prog Horm Res 30: 139-169.
Enmark E, Pelto-Huikko M, Grandien K, Lagercrantz S, Lagercrantz J, Fried G, Nordenskjöld M &Gustafsson J-Å (1997) Human estrogen receptor β-gene structure, chromosomal localization,and expression pattern. J Clin Endocrinol Metab 82: 4258-4265.
Fan L-Q, Cattley RC & Corton JC (1998) Tissue-specific induction of 17β-hydroxysteroiddehydrogenase type IV by peroxisome proliferator chemicals is dependent on the peroxisomeproliferator-activated receptor α. J Endocrinol 158: 237-246.
Farhat MY, Lavigne MC & Ramwell PW (1996) The vascular protective effects of estrogen.FASEB J 10: 615-624.
Ferin MJ (1996) The menstrual cycle: an integrative view. In: Adashi EY, Rock JA, & RosenwaksZ (eds) Reproductive Endocrinology, Surgery, and Technology. Lippincott-Raven Publishers,Philadelphia, p 103-121.
Fishman J, Nisselbaum JS, Menendez-Botet CJ & Schwartz MK (1977) Estrone and estradiolcontent in human breast tumors: relationship to estradiol receptors. J Steroid Biochem 8: 893-896.
Fitzpatrick SL, Carlone DL, Robker RL & Richards JS (1997) Expression of aromatase in theovary: down-regulation of mRNA by the ovulatory luteinizing hormone surge. Steroids 62: 197-206.
Fitzpatrick SL, Funkhouser JM, Sindoni DM, Stevis PE, Deecher DC, Bapat AR, Merchenthaler I& Frail DE (1999) Expression of estrogen receptor-β protein in rodent ovary. Endocrinology140: 2581-2591.
Fitzpatrick SL & Richards JS (1991) Regulation of cytochrome P450 aromatase messengerribonucleic acid and activity by steroids and gonadotropins in rat granulosa cells. Endocrinology129: 1452-1462.
Fleming H, Namit C & Gurpide E (1980) Estrogen receptors in epithelial and stromal cells ofhuman endometrium in culture. J Steroid Biochem 12: 169-174.
Fomitcheva J, Baker ME, Anderson E, Lee GY & Aziz N (1998) Characterization of Ke 6, a new17β-hydroxysteroid dehydrogenase, and its expression in gonadal tissues. J Biol Chem 273:22664-22671.
Fournet-Dulguerov N, MacLusky NJ, Leranth CZ, Todd R, Mendelson CR, Simpson ER &Naftolin F (1987) Immunohistochemical localization of aromatase cytochrome P-450 andestradiol dehydrogenase in the syncytiotrophoblast of the human placenta. J Clin EndocrinolMetab 65: 757-764.
Gaub M-P, Bellard M, Scheuer I, Chambon P & Sassone-Corsi P (1990) Activation of theovalbumin gene by the estrogen receptor involves the fos-jun complex. Cell 63: 1267-1276.
Geissler WM, Davis DL, Wu L, Bradshaw KD, Patel S, Mendonca BB, Elliston KO, Wilson JD,Russell DW & Andersson S (1994) Male pseudohermaphroditism caused by mutations oftesticular 17β-hydroxysteroid dehydrogenase 3. Nat Genet 7: 34-39.
Ghersevich SA, Poutanen MH, Martikainen HK & Vihko RK (1994a) Expression of 17β-hydroxysteroid dehydrogenase in human granulosa cells: correlation with follicular size,cytochrome P450 aromatase activity and oestradiol production. J Endocrinol 143: 139-150.

65
Ghersevich SA, Poutanen MH, Rajaniemi HJ & Vihko RK (1994b) Expression of 17β-hydroxysteroid dehydrogenase in the rat ovary during follicular development and luteinizationinduced with pregnant mare serum gonadotrophin and human chorionic gonadotrophin. JEndocrinol 140: 409-417.
Ghersevich S, Poutanen M, Tapanainen J & Vihko R (1994c) Hormonal regulation of rat 17β-hydroxysteroid dehydrogenase type 1 in cultured rat granulosa cells: effects of recombinantfollicle-stimulating hormone, estrogens, androgens, and epidermal growth factor. Endocrinology135: 1963-1971.
Ghosh D, Pletnev VZ, Zhu D-W, Wawrzak Z, Duax WL, Pangborn W, Labrie F & Lin S-X (1995)Structure of human estrogenic 17β-hydroxysteroid dehydrogenase at 2.20 Å resolution.Structure 3: 503-513.
Ghosh D, Wawrzak Z, Weeks CM, Duax WL & Erman M (1994) The refined three-dimensionalstructure of 3α,20β-hydroxysteroid dehydrogenase and possible roles of the residues conservedin short-chain dehydrogenases. Structure 2: 629-640.
Ghosh D, Weeks CM, Grochulski P, Duax WL, Erman M, Rimsay RL & Orr JC (1991) Three-dimensional structure of holo 3α,20β-hydroxysteroid dehydrogenase: a member of a short-chaindehydrogenase family. Proc Natl Acad Sci USA 88: 10064-10068.
Gibori G (1993) The corpus luteum of pregnancy. In: Adashi EY & Leung PCK (eds) The Ovary.Raven Press, New York, p 261-317.
Gibori G & Keyes PL (1980) Luteotropic role of estrogen in early pregnancy in the rat.Endocrinology 106: 1584-1588.
Gibori G, Khan I, Warshaw ML, McLean MP, Puryear TK, Nelson S, Durkee TJ, Azhar S,Steinschneider A & Rao MC (1988) Placental-derived regulators and the complex control ofluteal cell function. Recent Prog Horm Res 44: 377-429.
Goldfien AG & Monroe SE (1991) Ovaries. In: Greenspan FS (ed) Basic and ClinicalEndocrinology. Prentice-Hall International Inc., East Norwalk, USA, p 442-490.
Gougeon A (1996) Regulation of ovarian follicular development in primates: facts and hypotheses.Endocr Rev 17: 121-155.
Green S & Chambon P (1988) Nuclear receptors enhance our understanding of transcriptionregulation. Trends Genet 4: 309-314.
van Grunsven EG, van Berkel E, Ijlst L, Vreken P, de Klerk JBC, Adamski J, Lemonde H, ClaytonPT, Cuebas DA & Wanders RJA (1998) Peroxisomal D-hydroxyacyl-CoA dehydrogenasedeficiency: resolution of the enzyme defect and its molecular basis in bifunctional proteindeficiency. Proc Natl Acad Sci USA 95: 2128-2133.
van Grunsven EG, van Berkel E, Mooijer PA, Watkins PA, Moser HW, Suzuki Y, Jiang LL,Hashimoto T, Hoefler G, Adamski J & Wanders RJ (1999a) Peroxisomal bifunctional proteindeficiency revisited: resolution of its true enzymatic and molecular basis. Am J Hum Genet 64:99-107.
van Grunsven EG, Mooijer PA, Aubourg P & Wanders RJ (1999b) Enoyl-CoA hydratasedeficiency: identification of a new type of D-bifunctional protein deficiency. Hum Mol Genet 8:1509-1516.
Guiochon-Mantel A, Delabre K, Lescop P & Milgrom E (1996) Intracellular traffic of steroidhormone receptors. J Steroid Biochem Mol Biol 56: 3-9.
He X-Y, Merz G, Mehta P, Schulz H & Yang S-Y (1999) Human brain short chain L-3-hydroxyacyl coenzyme A dehydrogenase is a single-domain multifunctional enzyme.Characterization of a novel 17β-hydroxysteroid dehydrogenase. J Biol Chem 274: 15014-15019.
Hickey GJ, Chen S, Besman MJ, Shively JE, Hall PF, Gaddy-Kurten D & Richards JS (1988)Hormonal regulation, tissue distribution, and content of aromatase cytochrome P450 messengerribonucleic acid and enzyme in rat ovarian follicles and corpora lutea: relationship to estradiolbiosynthesis. Endocrinology 122: 1426-1436.
Hillier SG, Whitelaw PF & Smyth CD (1994) Follicular oestrogen synthesis: the 'two-cell, two-gonadotropin' model revisited. Mol Cell Endocrinol 100: 51-54.
Hisaw FL (1959) Comparative effectiveness of estrogens on fluid imbibition and growth of the rat'suterus. Endocrinology 64: 276-289.

66
Horwitz KB, Jackson TA, Bain DL, Richer JK, Takimoto GS & Tung L (1996) Nuclear receptorcoactivators and corepressors. Mol Endocrinol 10: 1167-1177.
Iguchi T, Tani N, Sato T, Fukatsu N & Ohta Y (1993) Developmental changes in mouse placentalcells from several stages of pregnancy in vivo and in vitro. Biol Reprod 48: 188-196.
Jantus Lewintre E, Orava M, Peltoketo H & Vihko R (1994) Characterization of 17β-hydroxysteroid dehydrogenase type 1 in choriocarcinoma cells: regulation by basic fibroblastgrowth factor. Mol Cell Endocrinol 104: 1-9.
Jarabak J (1969) Soluble 17β-hydroxysteroid dehydrogenase of human placenta. In: MethodsEnzymol. Academic Press, New York-London, p 746-752.
Jarabak J & Sack GH (1969) A soluble 17β-hydroxysteroid dehydrogenase from human placenta.The binding of pyridine nucleotides and steroids. Biochemistry 8: 2203-2212.
Jin J-Z & Lin S-X (1999) Human estrogenic 17β-hydroxysteroid dehydrogenase: predominance ofestrone reduction and its induction by NADPH. Biochem Biophys Res Commun 259: 489-493.
Jörnvall H, Persson B, Krook M, Atrian S, Gonzàlez-Duarte R, Jeffery J & Ghosh D (1995) Short-chain dehydrogenases/reductases (SDR). Biochemistry 34: 6003-6013.
Kaminski T, Akinola L, Poutanen M, Vihko R & Vihko P (1997) Growth factors and phorbol-12-myristate-13-acetate modulate the follicle-stimulating hormone- and cyclic adenosine-3',5'-monophosphate-dependent regulation of 17β-hydroxysteroid dehydrogenase type 1 expressionin rat granulosa cells. Mol Cell Endocrinol 136: 47-56.
Karavolas HJ & Engel LL (1971) Human placental 17β-estradiol dehydrogenase. VI. Substratespecificity of the diphosphopyridine nucleotide (triphosphopyridine nucleotide)-linked enzyme.Endocrinology 88: 1165-1169.
Kaufmann M, Carstensen J, Husen B & Adamski J (1995) The tissue distribution of porcine 17β-estradiol dehydrogenase and its induction by progesterone. J Steroid Biochem Mol Biol 55: 535-539.
Kellis JT, & Vickery LE (1987) Purification and characterization of human placental aromatasecytochrome P-450. J Biol Chem 262: 4413-4420.
Khan I, Sridaran R, Johnson DC & Gibori G (1987) Selective stimulation of luteal androgenbiosynthesis by luteinizing hormone: comparison of hormonal regulation of P45017α activity incorpora lutea and follicles. Endocrinology 121: 1312-1319.
Khanna M, Qin K-N, Wang RW & Cheng K-C (1995) Substrate specificity, gene structure, andtissue-specific distribution of multiple human 3α-hydroxysteroid dehydrogenases. J Biol Chem270: 20162-20168.
Khanum A, Buczko E & Dufau ML (1997) Essential role of adenosine triphosphate in activation of17β-hydroxysteroid dehydrogenase in the rat Leydig cell. Endocrinology 138: 1612-1620.
Kikuti YY, Tamiya G, Ando A, Chen L, Kimura M, Ferreira E, Tsuji K, Trowsdale J & Inoko H(1997) Physical mapping 220 kb centromeric of the human MHC and DNA sequence analysisof the 43-kb segment including the RING1, HKE6, and HKE4 genes. Genomics 42: 422-435.
King WJ & Greene GL (1984) Monoclonal antibodies localize oestrogen receptor in the nuclei oftarget cells. Nature 307: 745-747.
Kobayashi K, Kobayashi H, Ueda M & Honda Y (1997) Expression of 17β-hydroxysteroiddehydrogenase type IV in chick retinal pigment epithelium. Exp Eye Res 64: 719-726.
Krazeisen A, Breitling R, Imai K, Fritz S, Möller G & Adamski J (1999) Determination of cDNA,gene structure and chromosomal localization of the novel human 17β-hydroxysteroiddehydrogenase type 7. FEBS Lett 460: 373-379.
Krozowski Z (1992) At the cutting edge: 11β-hydroxysteroid dehydrogenase and the short-chainalcohol dehydrogenase (SCAD) superfamily. Mol Cell Endocrinol 84: C25-C31.
Krozowski Z (1994) The short-chain alcohol dehydrogenase superfamily: variations on a commontheme. J Steroid Biochem Mol Biol 51: 125-130.
Kuiper GGJM, Carlsson B, Grandien K, Enmark E, Häggblad J, Nilsson S & Gustafsson J-Å(1997) Comparison of the ligand binding specificity and transcript tissue distribution of estrogenreceptors α and β. Endocrinology 138: 863-870.
Kuiper GGJM, Enmark E, Pelto-Huikko M, Nilsson S & Gustafsson J-Å (1996) Cloning of a novelestrogen receptor expressed in rat prostate and ovary. Proc Natl Acad Sci USA 93: 5925-5930.

67
Kumar V & Chambon P (1988) The estrogen receptor binds tightly to its responsive element as aligand-inducer homodimer. Cell 55: 145-156.
Kuss E (1994) The fetoplacental unit of primates. Exp Clin Endocrinol 102: 135-165.Labrie F (1991) Intracrinology. Mol Cell Endocrinol 78: C113-C118.Labrie F, Luu-The V, Lin S-X, Labrie C, Simard J, Breton R & Bélanger A (1997) The key role of
17β-hydroxysteroid dehydrogenases in sex steroid biology. Steroids 62: 148-158.Labrie Y, Durocher F, Lachance Y, Turgeon C, Simard J, Labrie C & Labrie F (1995) The human
type II 17β-hydroxysteroid dehydrogenase gene encodes two alternatively spliced mRNAspecies. DNA Cell Biol 14: 849-861.
Langer LJ, Alexander JA & Engel LL (1959) Human placental estradiol-17β dehydrogenase.Kinetics and substrate specificities. J Biol Chem 234: 2609-2614.
Langer LJ & Engel LL (1958) Human placental estradiol-17β dehydrogenase: concentration,characterization and assay. J Biol Chem 233: 583-588.
Lanisnik Rizner T, Moeller G, Thole HH, Zakelj-Mavric M & Adamski J (1999) A novel 17β-hydroxysteroid dehydrogenase in the fungus Cochliobolus lunatus: new insights into theevolution of steroid-hormone signalling. Biochem J 337: 425-431.
de Launoit Y & Adamski J (1999) Unique multifunctional HSD17B4 gene product: 17β-hydroxysteroid dehydrogenase 4 and D-3-hydroxyacyl-coenzyme A dehydrogenase/hydrataseinvolved in Zellweger syndrome. J Mol Endocrinol 22: 227-240.
Leenders F, Adamski J, Husen B, Thole HH & Jungblut PW (1994) Molecular cloning and aminoacid sequence of the porcine 17β-estradiol dehydrogenase. Eur J Biochem 222: 221-227.
Leenders F, Tesdorpf JG, Markus M, Engel T, Seedorf U & Adamski J (1996) Porcine 80-kDaprotein reveals intrinsic 17β-hydroxysteroid dehydrogenase, fatty acyl-CoA-hydratase/dehydrogenase, and sterol transfer activities. J Biol Chem 271: 5438-5442.
Lees JA, Fawell SE & Parker MG (1989) Identification of constitutive and steroid-dependenttransactivation domains in the mouse estrogen receptor. J Steroid Biochem 34: 33-39.
Lin H-K, Jez JM, Schlegel BP, Peehl DM, Pachter JA & Penning TM (1997) Expression andcharacterization of recombinant type 2 3α-hydroxysteroid dehydrogenase (HSD) from humanprostate: demonstration of bifunctional 3α/17β-HSD activity and cellular distribution. MolEndocrinol 11: 1971-1984.
Lin S-X, Yang F, Jin J-Z, Breton R, Zhu D-W, Luu-The V & Labrie F (1992) Subunit identity ofthe dimeric 17β-hydroxysteroid dehydrogenase from human placenta. J Biol Chem 267: 16182-16187.
Luu The V, Labrie C, Zhao HF, Couët J, Lachance Y, Simard J, Leblanc G, Côté J, Bérubé D,Gagné R & Labrie F (1989) Characterization of cDNAs for human estradiol 17β-dehydrogenaseand assignment of the gene to chromosome 17: evidence of two mRNA species with distinct 5'-termini in human placenta. Mol Endocrinol 3: 1301-1309.
Luu-The V, Labrie C, Simard J, Lachance Y, Zhao H-F, Couët J, Leblanc G & Labrie F (1990)Structure of two in tandem human 17β-hydroxysteroid dehydrogenase genes. Mol Endocrinol 4:268-275.
Luu-The V, Zhang Y, Poirier D & Labrie F (1995) Characteristics of human types 1, 2 and 3 17β-hydroxysteroid dehydrogenase activities: oxidation/reduction and inhibition. J Steroid BiochemMol Biol 55: 581-587.
MacLusky NJ (1996) Sex steroid receptors. In: Adashi EY, Rock JA, & Rosenwaks Z (eds)Reproductive Endocrinology, Surgery, and Technology. Lippincott-Raven Publishers,Philadelphia, p 627-663.
Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schütz G, Umesono K, Blumberg B, Kastner P,Mark M, Chambon P & Evans RM (1995) The nuclear receptor superfamily: the second decade.Cell 83: 835-839.
Mannermaa A, Peltoketo H, Winqvist R, Ponder BAJ, Kiviniemi H, Easton DF, Poutanen M,Isomaa V & Vihko R (1994) Human familial and sporadic breast cancer: analysis of the codingregions of the 17β-hydroxysteroid dehydrogenase 2 gene (EDH17B2) using a single-strandconformation polymorphism assay. Hum Genet 93: 319-324.
Markus M, Husen B & Adamski J (1995) The subcellular localization of 17β-hydroxysteroiddehydrogenase type 4 and its interaction with actin. J Steroid Biochem Mol Biol 55: 617-621.

68
Martel C, Rhéaume E, Takahashi M, Trudel C, Couët J, Luu-The V, Simard J & Labrie F (1992)Distribution of 17β-hydroxysteroid dehydrogenase gene expression and activity in rat andhuman tissues. J Steroid Biochem Mol Biol 41: 597-603.
Martucci CP & Fishman J (1979) Impact of continuously administered catechol estrogens onuterine growth and luteinizing hormone secretion. Endocrinology 105: 1288-1292.
Masamura S, Santner SJ, Heitjan DF & Santen RJ (1995) Estrogen deprivation causes estradiolhypersensitivity in human breast cancer cells. J Clin Endocrinol Metab 80: 2918-2925.
Matt DW & MacDonald GJ (1984) In vitro progesterone and testosterone production by the ratplacenta during pregnancy. Endocrinology 115: 741-747.
Maxwell MM, Nearing J & Aziz N (1995) Ke 6 gene: sequence and organization and aberrantregulation in murine polycystic kidney disease. J Biol Chem 270: 25213-25219.
McEwen BS & Alves SE (1999) Estrogen actions in the central nervous system. Endocr Rev 20:279-307.
McKenna NJ, Lanz RB & O'Malley BW (1998) Nuclear receptor coregulators: cellular andmolecular biology. Endocr Rev 20: 321-344.
McLaren A (1971) Blastocysts in the mouse uterus: the effect of ovariectomy, progesterone andoestrogen. J Endocrinol 50: 515-526.
McNeill JM, Reed MJ, Beranek PA, Bonney RC, Ghilchik MW, Robinson DJ & James VHT(1986) A comparison of the in vivo uptake and metabolism of 3H-oestrone and 3H-oestradiol bynormal breast and breast tumour tissues in post-menopausal women. Int J Cancer 38: 193-196.
McPhaul MJ, Marcelli M, Zoppi S, Griffin JE & Wilson JD (1993) Genetic basis of endocrinedisease 4. The spectrum of mutations in the androgen receptor gene that causes androgenresistance. J Clin Endocrinol Metab 76: 17-23.
Melamed M, Castaño E, Notides AC & Sasson S (1997) Molecular and kinetic basis for the mixedagonist/antagonist activity of estriol. Mol Endocrinol 11: 1868-1878.
Mendelsohn ME & Karas RH (1999) The protective effects of estrogen on the cardiovascularsystem. N Engl J Med 340: 1801-1811.
Mendonca BB, Arnhold IJP, Bloise W, Andersson S, Russell DW & Wilson JD (1999) 17β-hydroxysteroid dehydrogenase 3 deficiency in women. J Clin Endocrinol Metab 84: 802-804.
Mendoza-Hernández G, Calcagno M, Sánchez-Nuncio HR & Diaz-Zagoya JC (1984)Dehydroepiandrosterone is a substrate for estradiol 17β-dehydrogenase from human placenta.Biochem Biophys Res Commun 119: 83-87.
Miettinen M, Mustonen M, Poutanen M, Isomaa V, Wickman M, Söderqvist G, Vihko R & VihkoP (1999) 17β-hydroxysteroid dehydrogenases in normal human mammary epithelial cells andbreast tissue. Breast Cancer Res Treat 57: 175-182.
Miettinen MM, Mustonen MVJ, Poutanen MH, Isomaa VV & Vihko RK (1996a) Human 17β-hydroxysteroid dehydrogenase type 1 and type 2 isoenzymes have opposite activities in culturedcells and characteristic cell- and tissue-specific expression. Biochem J 314: 839-845.
Miettinen MM, Poutanen MH & Vihko RK (1996b) Characterization of estrogen-dependent growthof cultured MCF-7 human breast-cancer cells expressing 17β-hydroxysteroid dehydrogenasetype 1. Int J Cancer 68: 600-604.
Miller WL (1995) Mitochondrial specificity of the early steps in steroidogenesis. J Steroid BiochemMol Biol 55: 607-616.
Miller WL (1999) Steroid hormone biosynthesis and actions in the materno-feto-placental unit.Clin Perinatol 25: 799-817.
Miller WL, Auchus RJ & Geller DH (1997) The regulation of 17,20 lyase activity. Steroids 62:133-142.
Miller WR (1996) Aromatase inhibitors. Endocr-Relat Cancer 3: 65-79.Misao R, Nakanishi Y, Sun WS, Fujimoto J, Iwagaki S, Hirose R & Tamaya T (1999) Expression
of oestrogen receptor α and β mRNA in corpus luteum of human subjects. Mol Hum Reprod 5:17-21.
Moghrabi N & Andersson S (1998) 17β-Hydroxysteroid dehydrogenases: physiological roles inhealth and disease. Trends Endrocrinol Metab 9: 265-270.

69
Moghrabi N, Head JR & Andersson S (1997) Cell type-specific expression of 17β-hydroxysteroiddehydrogenase type 2 in human placenta and fetal liver. J Clin Endocrinol Metab 82: 3872-3878.
Montano MM, Müller V, Trobaugh A & Katzenellenbogen BS (1995) The carboxy-terminal Fdomain of the human estrogen receptor: role in the transcriptional activity of the receptor andthe effectiveness of antiestrogens as estrogen antagonists. Mol Endocrinol 9: 814-825.
Morishige WK, Pepe GJ & Rothchild I (1973) Serum luteinizing hormone, prolactin andprogesterone levels during pregnancy in the rat. Endocrinology 92: 1527-1530.
Mosselman S, Polman J & Dijkema R (1996) ERβ: Identification and characterization of a novelhuman estrogen receptor. FEBS Lett 392: 49-53.
Mustonen M, Poutanen M, Chotteau-Lelievre A, de Launoit Y, Isomaa V, Vainio S, Vihko R &Vihko P (1997b) Ontogeny of 17β-hydroxysteroid dehydrogenase type 2 mRNA expression inthe developing mouse placenta and fetus. Mol Cell Endocrinol 134: 33-40.
Mustonen MV, Isomaa VV, Vaskivuo T, Tapanainen J, Poutanen MH, Stenbäck F, Vihko RK &Vihko PT (1998a) Human 17β-hydroxysteroid dehydrogenase type 2 messenger ribonucleicacid expression and localization in term placenta and in endometrium during the menstrualcycle. J Clin Endocrinol Metab 83: 1319-1324.
Mustonen MVJ, Poutanen MH, Isomaa VV, Vihko PT & Vihko RK (1997a) Cloning of mouse17β-hydroxysteroid dehydrogenase type 2, and analysing expression of the mRNAs for types 1,2, 3, 4 and 5 in mouse embryos and adult tissues. Biochem J 325: 199-205.
Mustonen MVJ, Poutanen MH, Kellokumpu S, de Launoit Y, Isomaa VV, Vihko RK & Vihko PT(1998b) Mouse 17β-hydroxysteroid dehydrogenase type 2 mRNA is predominantly expressed inhepatocytes and in surface epithelial cells of the gastrointestinal and urinary tracts. J MolEndocrinol 20: 67-74.
Müller H, Ishimura R, Orwig KE, Liu B & Soares MJ (1998) Homologues for prolactin-likeproteins A and B are present in the mouse. Biol Reprod 58: 45-51.
Mäentausta O, Boman K, Isomaa V, Stendahl U, Bäckström T & Vihko R (1992)Immunohistochemical study of the human 17β-hydroxysteroid dehydrogenase and steroidreceptors in endometrial adenocarcinoma. Cancer 70: 1551-1555.
Mäentausta O, Peltoketo H, Isomaa V, Jouppila P & Vihko R (1990) Immunological measurementof human 17β-hydroxysteroid dehydrogenase. J Steroid Biochem 36: 673-680.
Mäentausta O, Sormunen R, Isomaa V, Lehto V-P, Jouppila P & Vihko R (1991)Immunohistochemical localization of 17β-hydroxysteroid dehydrogenase in the humanendometrium during the menstrual cycle. Lab Invest 65: 582-587.
Mäentausta O, Svalander P, Danielsson KG, Bygdeman M & Vihko R (1993) The effects of anantiprogestin, mifepristone, and an antiestrogen, tamoxifen, on endometrial 17β-hydroxysteroiddehydrogenase and progestin and estrogen receptors during the luteal phase of the menstrualcycle: an immunohistochemical study. J Clin Endocrinol Metab 77: 913-918.
Mäkelä S, Poutanen M, Kostian ML, Lehtimäki N, Strauss L, Santti R & Vihko R (1998) Inhibitionof 17β-hydroxysteroid oxidoreductase by flavonoids in breast and prostate cancer cells. ProcSoc Exp Biol Med 217: 310-316.
Mäkelä S, Poutanen M, Lehtimäki J, Kostian M-L, Santti R & Vihko R (1995) Estrogen-specific17β-hydroxysteroid oxidoreductase type 1 (E.C. 1.1.1.62) as a possible target for the action ofphytoestrogens. Proc Soc Exp Biol Med 208: 51-59.
Möller G, Leenders F, van Grunsven EG, Dolez V, Qualmann B, Kessels MM, Markus M,Krazeisen A, Husen B, Wanders RJA, de Launoit Y & Adamski J (1999) Characterization of theHSD17B4 gene: D-specific multifunctional protein 2/17β-hydroxysteroid dehydrogenase IV. JSteroid Biochem Mol Biol 69: 441-446.
Nakhla AM, Romas NA & Rosner W (1997) Estradiol activates the prostate androgen receptor andprostate-specific antigen secretion through the intermediacy of sex hormone-binding globulin. JBiol Chem 272: 6838-6841.
Normand T, Husen B, Leenders F, Pelczar H, Baert J-L, Begue A, Flourens A-C, Adamski J & deLaunoit Y (1995) Molecular characterization of mouse 17β-hydroxysteroid dehydrogenase IV. JSteroid Biochem Mol Biol 55: 541-548.

70
Normand T, Narod S, Labrie F & Simard J (1993) Detection of polymorphisms in the estradiol17β-hydroxysteroid dehydrogenase II gene at the EDH17B2 locus on 17q11-q21. Hum MolGenet 2: 479-483.
Olsen NJ & Kovacs WJ (1996) Gonadal steroids and immunity. Endocr Rev 17: 369-384.Paech K, Webb P, Kuiper GGJM, Nilsson S, Gustafsson J-Å, Kushner PJ & Scanlan TS (1997)
Differential ligand activation of estrogen receptors ERα and ERβ at AP1 sites. Science 277:1508-1510.
Panay N & Studd JWW (1998) The psychotherapeutic effects of estrogens. Gynecol Endocrinol 12:353-365.
Paria BC, Huet-Hudson YM & Dey SK (1993) Blastocyst's state of activity determines the"window" of implantation in the receptive mouse uterus. Proc Natl Acad Sci USA 90: 10159-10162.
Parmer TG, McLean MP, Duan WR, Nelson SE, Albarracin CT, Khan I & Gibori G (1992)Hormonal and immunological characterization of the 32 kilodalton ovarian-specific protein.Endocrinology 131: 2213-2221.
Pelletier G, Luu-The V & Labrie F (1995) Immunocytochemical localization of type I 17β-hydroxysteroid dehydrogenase in the rat brain. Brain Res 704: 233-239.
Pelletier G, Luu-The V, Têtu B & Labrie F (1999) Immunocytochemical localization of type 5 17β-hydroxysteroid dehydrogenase in human reproductive tissues. J Histochem Cytochem 47: 731-737.
Peltoketo H, Ghosh D & Vihko P (1998) 17β-hydroxysteroid dehydrogenase type 1: towardsstructure and function-based drug design. Curr Top Ster Res 1: 111-117.
Peltoketo H, Isomaa V, Mäentausta O & Vihko R (1988) Complete amino acid sequence of humanplacental 17β-hydroxysteroid dehydrogenase deduced from cDNA. FEBS Lett 239: 73-77.
Peltoketo H, Isomaa V & Vihko R (1992) Genomic organization and DNA sequences of human17β-hydroxysteroid dehydrogenase genes and flanking regions. Localization of multiple Alusequences and putative cis-acting elements. Eur J Biochem 209: 459-466.
Peltoketo H, Luu-The V, Simard J & Adamski J (1999b) 17β-Hydroxysteroid dehydrogenase(HSD)/17-ketosteroid reductase (KSR) family; nomenclature and main characteristics of the17HSD/KSR enzymes. J Mol Endocrinol 23: 1-11.
Peltoketo H, Piao Y, Mannermaa A, Ponder BAJ, Isomaa V, Poutanen M, Winqvist R & Vihko R(1994) A point mutation in the putative TATA box, detected in nondiseased individuals andpatients with hereditary breast cancer, decreases promoter activity of the 17β-hydroxysteroiddehydrogenase type 1 gene 2 (EDH17B2) in vitro. Genomics 23: 250-252.
Peltoketo H, Vihko P & Vihko R (1999a) Regulation of estrogen action: role of 17β-hydroxysteroid dehydrogenases. Vitam Horm 55: 353-398.
Penning TM (1996) 17β-Hydroxysteroid dehydrogenase: inhibitors and inhibitor design. Endocr-Relat Cancer 3: 41-56.
Pepe GJ & Albrecht ED (1995) Actions of placental and fetal adrenal steroid hormones in primatepregnancy. Endocr Rev 16: 608-648.
Persson B, Krook M & Jörnvall H (1991) Characteristics of short-chain alcohol dehydrogenasesand related enzymes. Eur J Biochem 200: 537-543.
Pettersson K, Grandien K, Kuiper GGJM & Gustafsson J-Å (1997) Mouse estrogen receptor βforms estrogen response element-binding heterodimers with estrogen receptor α. MolEndocrinol 11: 1486-1496.
Pfeifer SM & Strauss JF (1996) Progestins. In: Adashi EY, Rock JA, & Rosenwaks Z (eds)Reproductive Endocrinology, Surgery, and Technology. Lippincott-Raven Publishers,Philadelphia, p 493-504.
Piao Y-S, Peltoketo H, Jouppila A & Vihko R (1997) Retinoic acids increase 17β-hydroxysteroiddehydrogenase type 1 expression in JEG-3 and T47D cells, but the stimulation is potentiated byepidermal growth factor, 12-O-tetradecanoylphorbol-13-acetate, and cyclic adenosine 3',5'-monophosphate only in JEG-3 cells. Endocrinology 138: 898-904.
Piao Y-S, Peltoketo H, Oikarinen J & Vihko R (1995) Coordination of transcription of the human17β-hydroxysteroid dehydrogenase type 1 gene (EDH17B2) by a cell-specific enhancer and asilencer: identification of a retinoic acid response element. Mol Endocrinol 9: 1633-1644.

71
Picard D, Kumar V, Chambon P & Yamamoto KR (1990) Signal transduction by steroid hormones:nuclear localization is differentially regulated in estrogen and glucocorticoid receptors. CellRegul 1: 291-299.
Porter W, Saville B, Hoivik D & Safe S (1997) Functional synergy between the transcription factorSp1 and the estrogen receptor. Mol Endocrinol 11: 1569-1580.
Poutanen M, Isomaa V, Kainulainen K & Vihko R (1990) Progestin induction of 17β-hydroxysteroid dehydrogenase enzyme protein in the T-47D human breast-cancer cell line. Int JCancer 46: 897-901.
Poutanen M, Isomaa V, Lehto V-P & Vihko R (1992a) Immunological analysis of 17β-hydroxysteroid dehydrogenase in benign and malignant human breast tissue. Int J Cancer 50:386-390.
Poutanen M, Miettinen M & Vihko R (1993) Differential estrogen substrate specificities fortransiently expressed human placental 17β-hydroxysteroid dehydrogenase and an endogenousenzyme expressed in cultured COS-m6 cells. Endocrinology 133: 2639-2644.
Poutanen M, Moncharmont B & Vihko R (1992b) 17β-hydroxysteroid dehydrogenase geneexpression in human breast cancer cells: regulation of expression by a progestin. Cancer Res 52:290-294.
Pratt WB & Toft DO (1997) Steroid receptor interactions with heat shock protein andimmunophilin chaperones. Endocr Rev 18: 306-360.
Puranen T, Poutanen M, Ghosh D, Vihko R & Vihko P (1997a) Origin of substrate specificity ofhuman and rat 17β-hydroxysteroid dehydrogenase type 1, using chimeric enzymes and site-directed substitutions. Endocrinology 138: 3532-3539.
Puranen T, Poutanen M, Ghosh D, Vihko P & Vihko R (1997b) Characterization of structural andfunctional properties of human 17β-hydroxysteroid dehydrogenase type 1 using recombinantenzymes and site-directed mutagenesis. Mol Endocrinol 11: 77-86.
Puranen TJ, Kurkela RM, Lakkakorpi JT, Poutanen MH, Itäranta PV, Melis JP, Ghosh D, VihkoRK & Vihko PT (1999) Characterization of molecular and catalytic properties of intact andtruncated human 17β-hydroxysteroid dehydrogenase type 2 enzymes: intracellular localizationof the wild-type enzyme in the endoplasmic reticulum. Endocrinology 140: 3334-3341.
Puranen TJ, Poutanen MH, Peltoketo HE, Vihko PT & Vihko RK (1994) Site-directed mutagenesisof the putative active site of human 17β-hydroxysteroid dehydrogenase type 1. Biochem J 304:289-293.
Purdy RH, Halla M & Little B (1964) 20α-Hydroxysteroid dehydrogenase activity a function ofhuman placental 17β-hydroxysteroid dehydrogenase. Biochim Biophys Acta 89: 557-560.
Qin Y-M, Haapalainen AM, Conry D, Cuebas DA, Hiltunen JK & Novikov DK (1997b)Recombinant 2-enoyl-CoA hydratase derived from rat peroxisomal multifunctional enzyme 2:role of the hydratase reaction in bile acid synthesis. Biochem J 328: 377-382.
Qin Y-M, Poutanen MH, Helander HM, Kvist A-P, Siivari KM, Schmitz W, Conzelmann E,Hellman U & Hiltunen JK (1997a) Peroxisomal multifunctional enzyme of β-oxidationmetabolizing D-3-hydroxyacyl-CoA esters in rat liver: molecular cloning, expression andcharacterization. Biochem J 321: 21-28.
Quéva C, Ness SA, Grässer FA, Graf T, Vandenbunder B & Stéhelin D (1992) Expression patternsof c-myb and of v-myb induced myeloid-1 (mim-1) gene during the development of the chickembryo. Development 114: 125-133.
Ramirez S, Fomitcheva I & Aziz N (1998) Abnormal regulation of the Ke 6 gene, a new 17β-hydroxysteroid dehydrogenase in the cpk mouse kidney. Mol Cell Endocrinol 143: 9-22.
Razandi M, Pedram A, Greene GL & Levin ER (1999) Cell membrane and nuclear estrogenreceptors (ERs) originate from a single transcript: studies of ERα and ERβ expressed in Chinesehamster ovary cells. Mol Endocrinol 13: 307-319.
Reed MJ, Purohit A, Woo LWL & Potter BVL (1996) The development of steroid sulphataseinhibitors. Endocr-Relat Cancer 3: 9-23.
Rembiesa R, Marchut M & Warchol A (1971) Formation and metabolism of progesterone by themouse placenta in vitro. J Steroid Biochem 2: 111-119.
Revelli A, Massobrio M & Tesarik J (1998) Nongenomic actions of steroid hormones inreproductive tissues. Endocr Rev 19: 3-17.

72
Rheault P, Charbonneau A & Luu-The V (1999b) Structure and activity of the murine type 5 17β-hydroxysteroid dehydrogenase gene. Biochim Biophys Acta 1447: 17-24.
Rheault P, Dufort I, Soucy P & Luu-The V (1999a) Assignment of HSD17B5 encoding type 517beta-hydroxysteroid dehydrogenase to human chromosome bands 10p15–>p14 and mousechromosome 13 region A2 by in situ hybridization: identification of a new syntenic relationship.Cytogenet Cell Genet 84: 241-242.
Richards JS (1994) Hormonal control of gene expression in the ovary. Endocr Rev 15: 725-751.Richards JS, Russell DL, Robker RL, Dajee M & Alliston TN (1998) Molecular mechanisms of
ovulation and luteinization. Mol Cell Endocrinol 145: 47-54.Ringler GE & Strauss JF (1990) In vitro systems for the study of human placental endocrine
function. Endocr Rev 11: 105-123.Robker RL & Richards JS (1998) Hormonal control of the cell cycle in ovarian cells: proliferation
versus differentiation. Biol Reprod 59: 476-482.Rosner W, Hryb DJ, Khan MS, Nakhla AM & Romas NA (1999) Sex hormone-binding globulin
mediates steroid hormone signal transduction at the plasma membrane. J Steroid Biochem MolBiol 69: 481-485.
Rösler A, Silverstein S & Abeliovich D (1996) A (R80Q) mutation in 17β-hydroxysteroiddehydrogenase type 3 gene among Arabs of Israel is associated with pseudohermaphroditism inmales and normal asymptomatic females. J Clin Endocrinol Metab 81: 1827-1831.
Sadek S & Bell SC (1996) The effects of the antihormones RU486 and tamoxifen on fetoplacentaldevelopment and placental bed vascularisation in the rat: a model for intrauterine fetal growthretardation. Br J Obstet Gynaecol 103: 630-641.
Salido EC, Yen PH, Barajas L & Shapiro LJ (1990) Steroid sulfatase expression in human placenta:immunocytochemistry and in situ hybridization study. J Clin Endocrinol Metab 70: 1564-1567.
Sambrook J, Fritsch EF & Maniatis T (1989) Extraction, purification, and analysis of messengerRNA from eukaryotic cells. In: Molecular Cloning: A Laboratory Manual. Cold Spring HarborLaboratory Press, Cold Spring Harbor, p 7.1-7.84.
Sanger F, Nicklen S & Coulson AR (1977) DNA sequencing with chain-terminating inhibitors.Proc Natl Acad Sci USA 74: 5463-5467.
Santen RJ (1996) Estrogen synthesis inhibitors for breast cancer: an introductory overview.Endocr-Relat Cancer 3: 1-8.
Santen RJ, Manni A, Harvey H & Redmond C (1990) Endocrine treatment of breast cancer inwomen. Endocr Rev 11: 221-265.
Santner SJ, Feil PD & Santen RJ (1984) In situ estrogen production via the estrone sulfatasepathway in breast tumors: relative importance versus the aromatase pathway. J Clin EndocrinolMetab 59: 29-33.
Sasano H, Frost AR, Saitoh R, Harada N, Poutanen M, Vihko R, Bulun SE, Silverberg SG &Nagura H (1996) Aromatase and 17β-hydroxysteroid dehydrogenase type 1 in human breastcarcinoma. J Clin Endocrinol Metab 81: 4042-4046.
Sawetawan C, Milewich L, Word RA, Carr BR & Rainey WE (1994) Compartmentalization oftype I 17β-hydroxysteroid oxidoreductase in the human ovary. Mol Cell Endocrinol 99: 161-168.
Schatz F, Aigner S, Papp C, Toth-Pal E, Hausknecht V & Lockwood CJ (1995) Plasminogenactivator activity during decidualization of human endometrial stromal cells is regulated byplasminogen activator inhibitor 1. J Clin Endocrinol Metab 80: 2504-2510.
Schatz F, Papp C, Toth-Pal E & Lockwood CJ (1994) Ovarian steroid-modulated stromelysin-1expression in human endometrial stromal and decidual cells. J Clin Endocrinol Metab 78: 1467-1472.
Sha J, Baker P & O'Shaughnessy PJ (1996) Both reductive forms of 17β-hydroxysteroiddehydrogenase (types 1 and 3) are expressed during development in the mouse testis. BiochemBiophys Res Commun 222: 90-94.
Sha JA, Dudley K, Rajapaksha WRAKJS & O'Shaughnessy PJ (1997) Sequence of mouse 17β-hydroxysteroid dehydrogenase type 3 cDNA and tissue distribution of the type 1 and type 3isoform mRNAs. J Steroid Biochem Mol Biol 60: 19-24.

73
Sherman MI (1983) Endocrinology of rodent trophoblast cells. In: Loke Y & Whyte A (eds)Biology of Trophoblast. Elsevier, Amsterdam, p 401-467.
Shibata H, Spencer TE, Oñate SA, Jenster G, Tsai SY, Tsai M-J & O'Malley BW (1997) Role ofco-activators and co-repressors in the mechanism of steroid/thyroid receptor action. Recent ProgHorm Res 52: 141-165.
Shoham Z & Schachter M (1996) Estrogen biosynthesis – regulation, action, remote effects, andvalue of monitoring in ovarian stimulation cycles. Fertil Steril 65: 687-701.
Siiteri PK, Murai JT, Hammond GL, Nisker JA, Raymoure WJ & Kuhn RW (1982) The serumtransport of steroid hormones. Recent Prog Horm Res 38: 457-510.
Simard J, Durocher F, Mébarki F, Turgeon C, Sanchez R, Labrie Y, Couet J, Trudel C, Rhéaume E,Morel Y, Luu-The V & Labrie F (1996) Molecular biology and genetics of the 3β-hydroxysteroid dehydrogenase/∆5-∆4 isomerase gene family. J Endocrinol 150: S189-S207.
Simard J, Feunteun J, Lenoir G, Tonin P, Normand T, Luu The V, Vivier A, Lasko D, Morgan K,Rouleau GA, Lynch H, Labrie F & Narod SA (1993) Genetic mapping of the breast-ovariancancer syndrome to a small interval on chromosome 17q12-21: exclusion of candidate genesEDH17B2 and RARA. Hum Mol Genet 2: 1193-1199.
Simpson ER (1998) Genetic mutations resulting in estrogen insufficiency in the male. Mol CellEndocrinol 145: 55-59.
Simpson ER, Michael MD, Agarwal VR, Hinshelwood MM, Bulun SE & Zhao Y (1997)Expression of the CYP19 (aromatase) gene: an unusual case of alternative promoter usage.FASEB J 11: 29-36.
Simpson ER, Zhao Y, Agarwal VR, Michael MD, Bulun SE, Hinshelwood MM, Graham-LorenceS, Sun T, Fisher CR, Qin K & Mendelson CR (1995) Aromatase expression in health anddisease. Recent Prog Horm Res 52: 185-214.
Smith EP, Boyd J, Frank GR, Takahashi H, Cohen RM, Specker B, Williams TC, Lubahn DB &Korach KS (1994) Estrogen resistance caused by a mutation in the estrogen-receptor gene in aman. N Engl J Med 331: 1056-1061.
Soares MJ, Chapman BM, Rasmussen CA, Dai G, Kamei T & Orwig KE (1996) Differentiation oftrophoblast endocrine cells. Placenta 17: 277-289.
Speroff L, Glass RH & Kase NG (1994) Hormone biosynthesis, metabolism, and mechanism ofaction. In: Clinical Gynecologic Endocrinology and Infertility. Williams & Wilkins, Baltimore,USA, p 31-107.
Stocco DM (1999) Steroidogenic acute regulatory (StAR) protein: what's new. Bioessays 21: 768-775.
Stouffer RL (1996) Corpus luteum formation and demise. In: Adashi EY, Rock JA, & RosenwaksZ (eds) Reproductive Endocrinology, Surgery, and Technology. Lippincott-Raven Publishers,Philadelphia, p 251-269.
Strauss JF, Martinez F & Kiriakidou M (1996) Placental steroid hormone synthesis: unique featuresand unanswered questions. Biol Reprod 54: 303-311.
Strickler RC, Tobias B & Covey DF (1981) Human placental 17β-estradiol dehydrogenase and20α-hydroxysteroid dehydrogenase. J Biol Chem 256: 316-321.
Su J, Lin M & Napoli JL (1999) Complementary deoxyribonucleic acid cloning and enzymaticcharacterization of a novel 17β/3α-hydroxysteroid/retinoid short chain dehydrogenase/reductase. Endocrinology 140: 5275-5284.
Suzuki Y, Jiang LL, Souri M, Miyazawa S, Fukuda S, Zhang Z, Une M, Shimozawa N, Kondo N,Orii T & Hashimoto T (1997) D-3-hydroxyacyl-CoA dehydratase/D-3-hydroxyacyl-CoAdehydrogenase bifunctional protein deficiency: a newly identified peroxisomal disorder. Am JHum Genet 61: 1153-1162.
Söderqvist G, Poutanen M, Wickman M, von Schoultz B, Skoog L, Vihko & R. (1998) 17β-hydroxysteroid dehydrogenase type 1 in normal breast tissue during the menstrual cycle andhormonal contraception. J Clin Endocrinol Metab 83: 1190-1193.
Takeyama J, Sasano H, Suzuki T, Iinuma K, Nagura H & Andersson S (1998) 17β-hydroxysteroiddehydrogenase types 1 and 2 in human placenta: an immunohistochemical study withcorrelation to placental development. J Clin Endocrinol Metab 83: 3710-3715.

74
Taya K & Greenwald GS (1981) In vivo and in vitro ovarian steroidogenesis in the pregnant rat.Biol Reprod 25: 683-691.
Telleria CM, Zhong L, Deb S, Srivastava RK, Park KS, Sugino N, Park-Sarge O-K & Gibori G(1998) Differential expression of the estrogen receptors α and β in the rat corpus luteum ofpregnancy: regulation by prolactin and placental lactogens. Endocrinology 139: 2432-2442.
Thiboutot D, Martin P, Volikos L & Gilliland K (1998) Oxidative activity of the type 2 isozyme of17β-hydroxysteroid dehydrogenase (17β-HSD) predominates in human sebaceous glands. JInvest Dermatol 111: 390-395.
Thijssen JHH & Blankenstein MA (1994) Steroid hormones in the human breast. Endocr-RelatCancer 2: 43-48.
Tora L, White J, Brou C, Tasset D, Webster N, Scheer E & Chambon P (1989) The human estrogenreceptor has two independent nonacidic transcriptional activation functions. Cell 59: 477-487.
Touitou I, Cai QQ & Rochefort H (1994) 17 beta hydroxysteroid dehydrogenase 1 "pseudogene" isdifferentially transcribed: still a candidate for the breast-ovarian cancer susceptibility gene(BRCA1). Biochem Biophys Res Commun 201: 1327-1332.
Townsend L & Ryan KJ (1970) In vitro metabolism of pregnenolone-7α-3H, progesterone-4-14Cand androstenedione-4- 14C by rat placental tissue. Endocrinology 87: 151-155.
Tremblay GB, Tremblay A, Copeland NG, Gilbert DJ, Jenkins NA, Labrie, F & Giguère V (1997)Cloning, chromosomal localization, and functional analysis of the murine estrogen receptor β.Mol Endocrinol 11: 353-365.
Tsafriri A & Chun S-Y (1996) Ovulation. In: Adashi EY, Rock JA, & Rosenwaks Z (eds)Reproductive Endocrinology, Surgery, and Technology. Lippincott-Raven Publishers,Philadelphia, p 235-249.
Tsai M-J & O'Malley BW (1994) Molecular mechanisms of action of steroid/thyroid receptorsuperfamily members. Annu Rev Biochem 63: 451-486.
Tsai-Morris CH, Khanum A, Tang P-Z & Dufau ML (1999) The rat 17β-hydroxysteroiddehydrogenase type III: molecular cloning and gonadotropin regulation. Endocrinology 140:3534-3542.
Tseng L (1980) Hormonal regulation of steroid metabolic enzymes in human endometrium. In:Thomas JA & Singhal L (eds) Advances in Sex Steroid Hormone Research. Urban &Schwarzenberg, Baltimore and Munich, p 329-361.
Turner RT, Riggs BL & Spelsberg TC (1994) Skeletal effects of estrogen. Endocr Rev 15: 275-300.Umayahara Y, Kawamori R, Watada H, Imano E, Iwama N, Morishima T, Yamasaki Y, Kajimoto
Y & Kamada T (1994) Estrogen regulation of the insulin-like growth factor I gene transcriptioninvolves an AP-1 enhancer. J Biol Chem 269: 16433-16442.
Vermeulen A, Deslypere JP, Paridaens R, Leclercq G, Roy F & Heuson JC (1986) Aromatase, 17β-hydroxysteroid dehydrogenase and intratissular sex hormone concentrations in cancerous andnormal glandular breast tissue in postmenopausal women. Eur J Cancer Clin Oncol 22: 515-525.
Wajima Y, Furusawa T, Kawauchi S, Wakabayashi N, Nakabayashi O, Nishimori K & Mizuno S(1999) The cDNA cloning and transient expression of an ovary-specific 17β-hydroxysteroiddehydrogenase of chickens. Gene 233: 75-82.
Warner M, Nilsson S & Gustafsson J-Å (1999) The estrogen receptor family. Curr Opin ObstetGynecol 11: 249-254.
Warshaw ML, Johnson DC, Khan I, Eckstein B & Gibori G (1986) Placental secretion ofandrogens in the rat. Endocrinology 119: 2642-2648.
Webster NJG, Green S, Jin JR & Chambon P (1988) The hormone-binding domains of the estrogenand glucocorticoid receptors contain an inducible transcription activation function. Cell 54: 199-207.
Weigel NL (1996) Steroid hormone receptors and their regulation by phosphorylation. Biochem J319: 657-667.
Weitlauf HM (1994) Biology of implantation. In: Knobil E & Neill JD (eds) The Physiology ofReproduction. Raven Press, New York, p 391-440.
Wilson J & Parson M (1996) Endocrinology of human gestation. In: Adashi EY, Rock JA, &Rosenwaks Z (eds) Reproductive Endocrinology, Surgery, and Technology. Lippincott-RavenPublishers, Philadelphia, p 451-475.

75
Wilson JD, Griffin JE & Russell DW (1993) Steroid 5α-reductase 2 deficiency. Endocr Rev 14:577-593.
Winqvist R, Peltoketo H, Isomaa V, Grzeschik K-H, Mannermaa A & Vihko R (1990) The gene for17β-hydroxysteroid dehydrogenase maps to human chromosome 17, bands q12-q21, and showsan RFLP with ScaI. Hum Genet 85: 473-476.
Wu L, Einstein M, Geissler WM, Chan HK, Elliston KO & Andersson S (1993) Expression cloningand characterization of human 17β-hydroxysteroid dehydrogenase type 2, a microsomal enzymepossessing 20α-hydroxysteroid dehydrogenase activity. J Biol Chem 268: 12964-12969.
Yamada AT & Nagata T (1992) Light and electron microscopic radioautography of DNA synthesisin the endometria of pregnant-ovariectomized mice during activation of implantation window.Cell Mol Biol 38: 763-774.
Ylikomi T, Wurtz J-M, Syvälä H, Passinen S, Pekki A, Haverinen M, Bläuer M, Tuohimaa P &Gronemeyer H (1998) Reappraisal of the role of heat shock proteins as regulators of steroidreceptor activity. Crit Rev Biochem Mol 33: 437-466.
Zeitoun K, Takayama K, Sasano H, Suzuki T, Moghrabi N, Andersson S, Johns A, Meng L,Putman M, Carr B & Bulun SE (1998) Deficient 17β-hydroxysteroid dehydrogenase type 2expression in endometriosis: failure to metabolize 17β-estradiol. J Clin Endocrinol Metab 83:4474-4480.





























