Embed Size (px)
Citation preview

Analytical Biochemistry 412 (2011) 183–188
Contents lists available at ScienceDirect
Analytical Biochemistry
journal homepage: www.elsevier .com/locate /yabio
BioVyon Protein A, an alternative solid-phase affinity matrixfor chromatin immunoprecipitation
Igor Chernukhin a,1, Sung Yun Kang a,1, Sam Brown b, Svetlana Gretton a, Claudia Fabiola Mendez-Catala a,Dave Cowieson b, Elena Klenova a,⇑a Department of Biological Sciences, University of Essex, Colchester, Essex CO4 3SQ, UKb Porvair Technology, A Division of Porvair Filtration Group, Wrexham LL13 9XS, UK
a r t i c l e i n f o
Article history:Received 12 November 2010Received in revised form 25 January 2011Accepted 25 January 2011Available online 1 February 2011
Keywords:Chromatin immunoprecipitation (ChIP)BioVyon Protein AProtein A SepharoseDynabeads Protein ACCCTC-binding factor (CTCF)CTCF binding (target) sites (CTSs)RNA polymerase IITATA boxQuantitative real-time polymerase chainreaction (Q-PCR)
0003-2697/$ - see front matter Crown Copyright � 2doi:10.1016/j.ab.2011.01.036
⇑ Corresponding author. Fax: +44 1206 872592.E-mail address: [email protected] (E. Klenova)
1 These authors contributed equally to this work.2 Abbreviations used: ChIP, chromatin immunoprecip
tion; PCR, polymerase chain reaction; HDPE, high-dquantitative real-time PCR; IgG, immunoglobulin G;GAPDH, glyceraldehyde 3-phosphate dehydrogenase; LHS, high-salt; SDS, sodium dodecyl sulfate; CTS, CTCF b
a b s t r a c t
Chromatin immunoprecipitation (ChIP) is an important technique in the study of DNA/protein interac-tions. The ChIP procedure, however, has limitations in that it is lengthy, can be inconsistent, and is proneto nonspecific binding of DNA and proteins to the bead-based solid-phase matrices that are often used forthe immunoprecipitation step. In this investigation, we examined the utility of a new matrix for ChIPassays, BioVyon Protein A, a solid support based on porous polyethylene. In ChIP experiments carriedout using two antibodies and seven DNA loci, the performance of BioVyon Protein A was significantly bet-ter, with a greater percentage of DNA pull-down in all of the assays tested compared with bead-basedmatrices, Protein A Sepharose, and Dynabeads Protein A. Furthermore, the rigid porous disc format withina column made the BioVyon matrix much easier to use with fewer steps and less equipment require-ments, resulting in a significant reduction in the time taken to process the ChIP samples. In summary,BioVyon Protein A provides a column-based assay method for ChIP and other immunoprecipitation-basedprocedures; the rigid porous structure of BioVyon enables a fast and robust protocol with higher ChIPenrichment ratios.
Crown Copyright � 2011 Published by Elsevier Inc. All rights reserved.
The chromatin immunoprecipitation (ChIP)2 assay is an impor-tant research tool in modern molecular biology [1–5]. It allows thestudy and identification of DNA sequences that are specificallybound to particular proteins; they represent important regulatoryelements in transcriptional machinery. The ChIP assay is a complexprocedure that involves several steps: DNA/protein crosslinking,sonication, immunoprecipitation (IP) of the crosslinked DNA/protein(chromatin) complexes, capture of these complexes, DNA recoveryfrom the precipitated product, and DNA analysis. During the IP step,antibodies specific to the protein component are employed andcapture of the immunoglobulin/DNA/protein complexes is achievedby the specific binding of immunoglobulins to Protein A and/or Pro-tein G conjugated to a solid support [6]. DNA analysis can be carriedout by polymerase chain reaction (PCR), quantitative real-time PCR
011 Published by Elsevier Inc. All r
.
itation; IP, immunoprecipita-ensity polyethylene; Q-PCR,SPE, solid-phase extraction;
S, low-salt; MS, medium-salt;inding (target) site.
(Q-PCR), hybridization on microarrays (ChIP–ChIP) [7,8], or directsequencing (ChIP–seq) [9].
ChIP assays, however, suffer from inherent problems that canoften lead to misleading, or even erroneous, interpretation of theChIP data [4]. These problems arise during IP, the most crucial partin the ChIP assay. Two main components of IP determine the qual-ity and quantity of the ChIP DNA: the antibody and the solid sup-port used for binding the antigen/antibody complex. The ChIPantibody may contribute to nonspecific background signal becauseof their possible cross-reactivity with other antigens. The ChIPantibody may also be responsible for a low yield of the recoveredDNA due to a low affinity for the protein bound to the DNA. How-ever, the bulk of nonspecific binding to the solid support, in partic-ular to agarose-based matrices such as Protein A Sepharose beadscommonly used in IP and ChIP assays, is thought to be due to theDNA/protein complexes reacting with diverse chemical groups onthe surface of the Sepharose.
The Sepharose-based matrices are derived from a naturallyoccurring material (seaweed) and have a very chemically heteroge-neous base structure with a very high surface area (see Supple-mental Fig. 1A in supplementary material). The high surface areaof the Protein A Sepharose variants generally favors IP. However,
ights reserved.

184 Solid-phase affinity matrix for ChIP / I. Chernukhin et al. / Anal. Biochem. 412 (2011) 183–188
in ChIP assays, nonspecific binding of chromatin tends to occur as aresult of ionic interactions between the differently charged sur-faces of Sepharose and DNA. To minimize this problem, an addi-tional DNA preblocking step is often recommended in ChIPprotocols. During this step, the Sepharose-based solid support ispreincubated with nonhomologous DNA/RNA prior to IP to blockany potential active binding sites. However, this procedure mightnot remove all of the nonspecific binding and adds an additionalstep to the process.
To improve the quality of the IP step in a ChIP assay, PorvairFiltration Group has developed an alternative matrix, BioVyonProtein A, which is based on porous high-density polyethylene(HDPE). BioVyon has very different polymer chemistry fromSepharose; it is a synthetic polymer made up of repeat units ofthe hydrocarbon ethylene and is chemically homogeneous(Supplemental Fig. 1B), this structure is less variable and muchmore inert. The surface of BioVyon has been chemically etched(by a proprietary oxidation method), providing a moderate in-crease in surface area and allowing the covalent attachment of alinker and a Protein A molecule (Supplemental Figs. 2 and 3). Theetching/oxidation process introduces a relatively low surfaceconcentration of oxidized species, relative to a polysaccharide(Sepharose)- or methacrylate (Dynabeads)-based material, leavingan inert surface with a concentration of Protein A sufficient for theChIP assay. The chemical structure of BioVyon Protein A, describedabove, is expected to exhibit reduced amounts of nonspecific bind-ing to DNA/protein complexes.
BioVyon is manufactured in the form of a solid but porous discthat is rigid and incompressible and is inserted into a column(Supplemental Fig. 4). This makes the BioVyon matrix easier tohandle than bead-based matrices and allows a column to beprepared with a precise amount of Protein A fixed at the base.
In this study, we carried out ChIP assays with two different anti-bodies and several DNA loci using BioVyon Protein A in microcol-umns. These results were compared with the results of ChIPassays obtained using other matrices under similar conditions(Protein A Sepharose and Dynabeads Protein A). Dynabeads aresuperparamagnetic particles coated with a polyacrylate polymerlayer. From these experiments, we concluded that the BioVyonProtein A matrix presents an attractive alternative to the existingmatrices in ChIP and, potentially, in other IP-based assays.
Materials and methods
Cells
NIH 3T3 mouse fibroblasts were maintained in Dulbecco’s mod-ified Eagle’s medium (DMEM, Lonza, Basel, Switzerland) supple-mented with 10% donor serum and 50 lg/ml gentamicin (bothfrom Invitrogen, Carlsbad, CA, USA). MCF7 human breast cancercells were cultured in RPMI-1640 with Ultraglutamine 1 (bothfrom Lonza) supplemented with 10% fetal bovine serum (FBS, Bio-sera, East Sussex, UK) and 50 lg/ml gentamicin.
Antibodies
Antibodies for this study were as follows: anti-RNA polymeraseII (8WG16) mouse monoclonal (Covance Research Products,Princeton, NJ, USA), previously used in ChIP assays [10]; anti-CTCF(CCCTC-binding factor) rabbit polyclonal (Upstate (Millipore),Billerica, MA, USA), employed for genome-wide ChIP analyses(e.g., Refs. [11,12]); anti-His-tag rabbit polyclonal (Abcam,Cambridge, UK); anti-b-actin mouse monoclonal (Sigma–Aldrich,St. Louis, MO, USA); mouse and rabbit immunoglobulin G (IgG,Santa Cruz Biotechnology, Santa Cruz, CA, USA).
Q-PCR
The primers and conditions for Q-PCR are described inSupplemental Table 1 in the supplementary material. Q-PCRs wereperformed as reported previously with modifications [13]. In brief,the reaction components were assembled in a 25-ll mixture con-taining 3 ll of sample, 200 nM of each primer, and 12.5 ll of Sensi-Mix Plus SYBR Green PCR Kit (Quantace, London, UK). Amplification,data acquisition, and analysis were conducted using the Chromo4Real Time PCR (Bio-Rad Laboratories, Hercules, CA, USA). Dissocia-tion curve analysis was performed for each sample after PCR to ver-ify that a single amplicon of the expected melt curve characteristicswas obtained. The amount of precipitated DNA was calculated rel-ative to the total input chromatin and expressed as a percentage ofthe total according to the formula% input = 2DCt � 100%, whereDCt = Ct (input) � Ct (IP) and Ct is the mean threshold cycle of thecorresponding PCR reaction [14]; this method has been widely usedin the scientific community (for examples, see Refs. [13,15]). Theseexperiments were carried out in triplicate, and the average was ob-tained from the percentage input.
BioVyon Protein A columns
BioVyon Protein A columns were developed and manufacturedby Porvair Filtration Group and provided by the company for thisstudy. The columns have the same dimensions as a standard 1-ml solid-phase extraction (SPE) tube and contain a rigid porousHDPE BioVyon Protein A frit approximately 6 mm in diameterand 2 mm long (Supplemental Fig. 4). The frits have been chemi-cally treated to increase surface area using a proprietary selectiveoxidation method, which preferentially etched the surface and pro-vided carboxylic acid anchor groups, for further covalent attach-ment. The pits caused by the etching process can be clearly seenin Supplemental Fig. 3, which compares the microstructure of thesurface before and after etching. The pitting was responsible forthe increased surface area, and further details regarding this areavailable in Supplemental Fig. 2. The surface has been tailored bythis process to provide sufficient functionality for the IP stage ofthe ChIP assay. The anchor groups formed on the surface of theHDPE were then covalently coupled to the Protein A via a linkerto form the BioVyon Protein A solid phase.
ChIP assays
ChIP assays were conducted according to the manufacturer’smanual for the ChIP Kit (Upstate (Millipore)) for Protein A Sephar-ose (Sigma–Aldrich) with some modifications. In ChIP experimentsinvolving BioVyon Protein A, the above protocol with some modi-fications was applied; the Protein A Sepharose slurry there was re-placed by the BioVyon Protein A columns. In the ChIP assays withDynabeads Protein A (Invitrogen), we followed the manufacturer’sprotocol. The detailed protocols of the ChIP experiments with thethree different matrices are provided in the Supplemental Materi-als and Methods Section of the supplementary material.
Statistical analysis
Statistical analysis was carried out using unpaired Student’s ttest. A significant value was detected when the probability was be-low the 5% confidence level (P < 0.05).
Results and discussion
In the ChIP protocol developed for BioVyon Protein A, the IP steprequired optimization for use with this solid support. As a model
Solid-phase affinity matrix for ChIP / I. Chernukhin et al. / Anal. Biochem. 412 (2011) 183–188 185
test system for ChIP in this study, we chose a previously describedassay for detection of binding of RNA polymerase II to a TATA boxwithin the promoter region of the mouse glyceraldehyde 3-phos-phate dehydrogenase (GAPDH) gene [10,16].
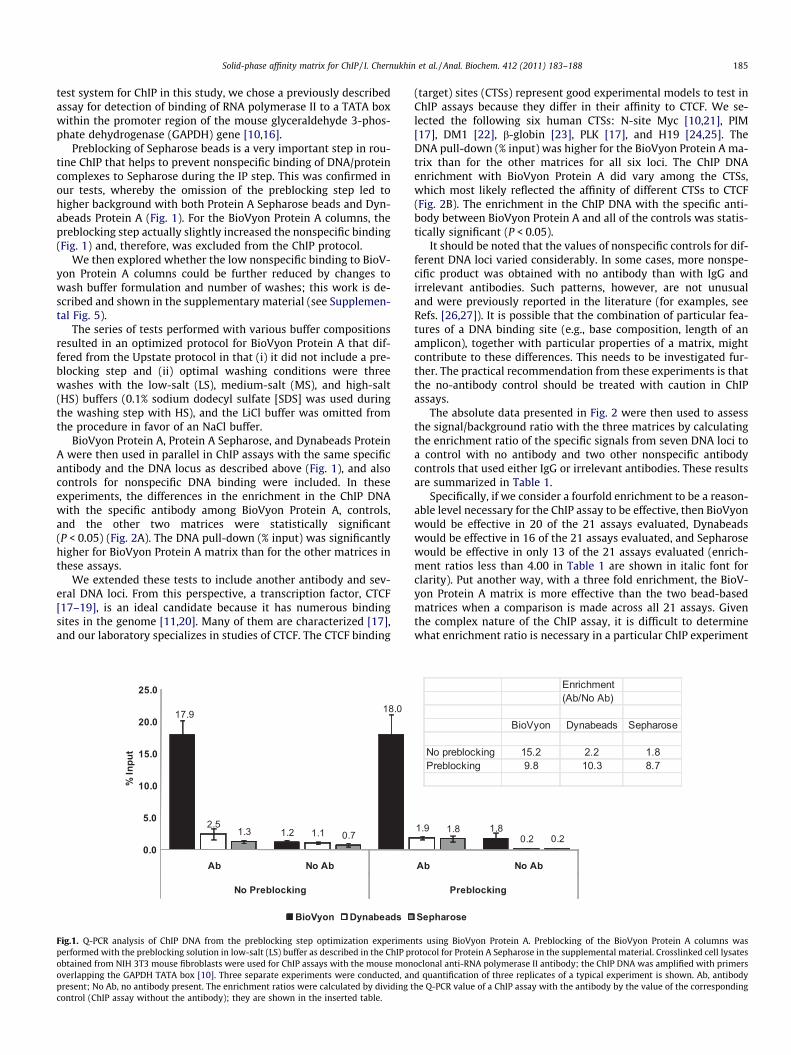
Preblocking of Sepharose beads is a very important step in rou-tine ChIP that helps to prevent nonspecific binding of DNA/proteincomplexes to Sepharose during the IP step. This was confirmed inour tests, whereby the omission of the preblocking step led tohigher background with both Protein A Sepharose beads and Dyn-abeads Protein A (Fig. 1). For the BioVyon Protein A columns, thepreblocking step actually slightly increased the nonspecific binding(Fig. 1) and, therefore, was excluded from the ChIP protocol.
We then explored whether the low nonspecific binding to BioV-yon Protein A columns could be further reduced by changes towash buffer formulation and number of washes; this work is de-scribed and shown in the supplementary material (see Supplemen-tal Fig. 5).
The series of tests performed with various buffer compositionsresulted in an optimized protocol for BioVyon Protein A that dif-fered from the Upstate protocol in that (i) it did not include a pre-blocking step and (ii) optimal washing conditions were threewashes with the low-salt (LS), medium-salt (MS), and high-salt(HS) buffers (0.1% sodium dodecyl sulfate [SDS] was used duringthe washing step with HS), and the LiCl buffer was omitted fromthe procedure in favor of an NaCl buffer.
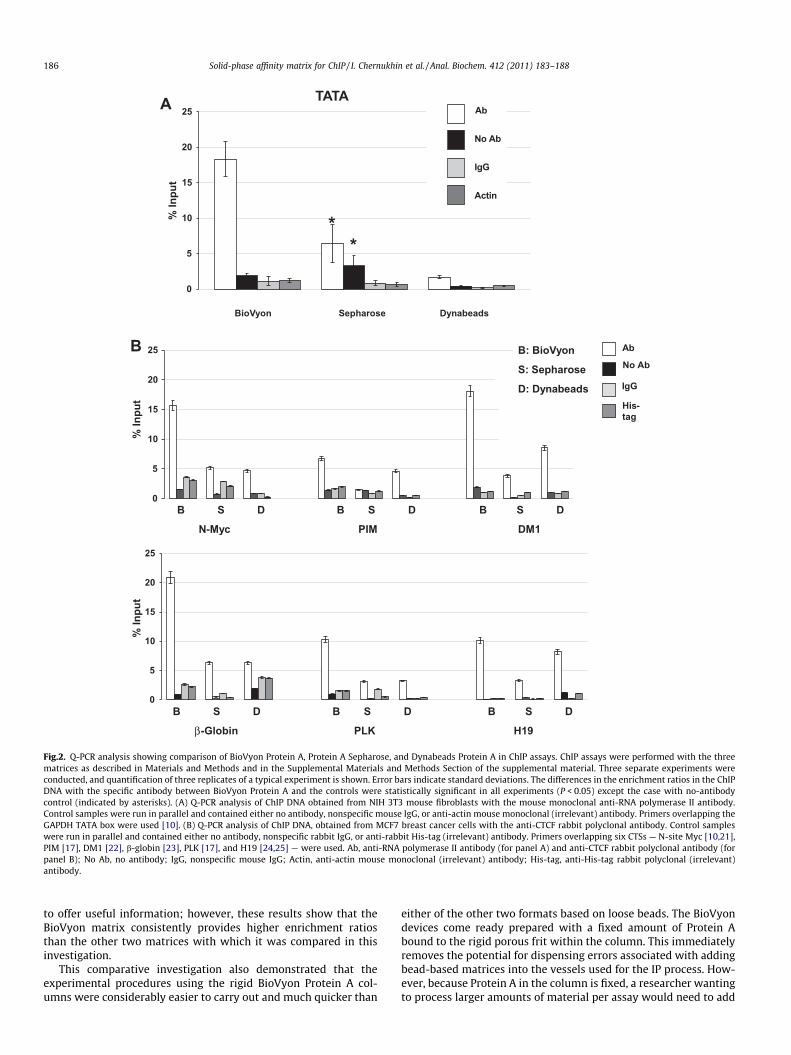
BioVyon Protein A, Protein A Sepharose, and Dynabeads ProteinA were then used in parallel in ChIP assays with the same specificantibody and the DNA locus as described above (Fig. 1), and alsocontrols for nonspecific DNA binding were included. In theseexperiments, the differences in the enrichment in the ChIP DNAwith the specific antibody among BioVyon Protein A, controls,and the other two matrices were statistically significant(P < 0.05) (Fig. 2A). The DNA pull-down (% input) was significantlyhigher for BioVyon Protein A matrix than for the other matrices inthese assays.
We extended these tests to include another antibody and sev-eral DNA loci. From this perspective, a transcription factor, CTCF[17–19], is an ideal candidate because it has numerous bindingsites in the genome [11,20]. Many of them are characterized [17],and our laboratory specializes in studies of CTCF. The CTCF binding
17.9
1.2
18.0
2.51.11.3 0.7
0.0
5.0
10.0
15.0
20.0
25.0
Ab No Ab
No Preblocking
% In
put
BioVyon Dynabeads
Fig.1. Q-PCR analysis of ChIP DNA from the preblocking step optimization experimenperformed with the preblocking solution in low-salt (LS) buffer as described in the ChIP probtained from NIH 3T3 mouse fibroblasts were used for ChIP assays with the mouse monoverlapping the GAPDH TATA box [10]. Three separate experiments were conducted, anpresent; No Ab, no antibody present. The enrichment ratios were calculated by dividing tcontrol (ChIP assay without the antibody); they are shown in the inserted table.
(target) sites (CTSs) represent good experimental models to test inChIP assays because they differ in their affinity to CTCF. We se-lected the following six human CTSs: N-site Myc [10,21], PIM[17], DM1 [22], b-globin [23], PLK [17], and H19 [24,25]. TheDNA pull-down (% input) was higher for the BioVyon Protein A ma-trix than for the other matrices for all six loci. The ChIP DNAenrichment with BioVyon Protein A did vary among the CTSs,which most likely reflected the affinity of different CTSs to CTCF(Fig. 2B). The enrichment in the ChIP DNA with the specific anti-body between BioVyon Protein A and all of the controls was statis-tically significant (P < 0.05).
It should be noted that the values of nonspecific controls for dif-ferent DNA loci varied considerably. In some cases, more nonspe-cific product was obtained with no antibody than with IgG andirrelevant antibodies. Such patterns, however, are not unusualand were previously reported in the literature (for examples, seeRefs. [26,27]). It is possible that the combination of particular fea-tures of a DNA binding site (e.g., base composition, length of anamplicon), together with particular properties of a matrix, mightcontribute to these differences. This needs to be investigated fur-ther. The practical recommendation from these experiments is thatthe no-antibody control should be treated with caution in ChIPassays.
The absolute data presented in Fig. 2 were then used to assessthe signal/background ratio with the three matrices by calculatingthe enrichment ratio of the specific signals from seven DNA loci toa control with no antibody and two other nonspecific antibodycontrols that used either IgG or irrelevant antibodies. These resultsare summarized in Table 1.
Specifically, if we consider a fourfold enrichment to be a reason-able level necessary for the ChIP assay to be effective, then BioVyonwould be effective in 20 of the 21 assays evaluated, Dynabeadswould be effective in 16 of the 21 assays evaluated, and Sepharosewould be effective in only 13 of the 21 assays evaluated (enrich-ment ratios less than 4.00 in Table 1 are shown in italic font forclarity). Put another way, with a three fold enrichment, the BioV-yon Protein A matrix is more effective than the two bead-basedmatrices when a comparison is made across all 21 assays. Giventhe complex nature of the ChIP assay, it is difficult to determinewhat enrichment ratio is necessary in a particular ChIP experiment
1.81.90.2
1.80.2
Ab No Ab
Preblocking
Sepharose
Enrichment(Ab/No Ab)
BioVyon Dynabeads Sepharose
No preblocking 15.2 2.2 1.8Preblocking 9.8 10.3 8.7
ts using BioVyon Protein A. Preblocking of the BioVyon Protein A columns wasotocol for Protein A Sepharose in the supplemental material. Crosslinked cell lysatesoclonal anti-RNA polymerase II antibody; the ChIP DNA was amplified with primersd quantification of three replicates of a typical experiment is shown. Ab, antibodyhe Q-PCR value of a ChIP assay with the antibody by the value of the corresponding
0
5
10
15
20
25 Ab
No Ab
IgG
His-tag
B: BioVyon
S: Sepharose
D: Dynabeads
0
5
10
15
20
25
N-Myc PIM DM1
B S D B S D B S D
β-Globin PLK H19
B S D B S D B S D
BioVyon Sepharose Dynabeads
0
5
10
15
20
25TATA
Ab
No Ab
Actin
IgG
**%
Inpu
t
% In
put
% In
put
A
B
Fig.2. Q-PCR analysis showing comparison of BioVyon Protein A, Protein A Sepharose, and Dynabeads Protein A in ChIP assays. ChIP assays were performed with the threematrices as described in Materials and Methods and in the Supplemental Materials and Methods Section of the supplemental material. Three separate experiments wereconducted, and quantification of three replicates of a typical experiment is shown. Error bars indicate standard deviations. The differences in the enrichment ratios in the ChIPDNA with the specific antibody between BioVyon Protein A and the controls were statistically significant in all experiments (P < 0.05) except the case with no-antibodycontrol (indicated by asterisks). (A) Q-PCR analysis of ChIP DNA obtained from NIH 3T3 mouse fibroblasts with the mouse monoclonal anti-RNA polymerase II antibody.Control samples were run in parallel and contained either no antibody, nonspecific mouse IgG, or anti-actin mouse monoclonal (irrelevant) antibody. Primers overlapping theGAPDH TATA box were used [10]. (B) Q-PCR analysis of ChIP DNA, obtained from MCF7 breast cancer cells with the anti-CTCF rabbit polyclonal antibody. Control sampleswere run in parallel and contained either no antibody, nonspecific rabbit IgG, or anti-rabbit His-tag (irrelevant) antibody. Primers overlapping six CTSs — N-site Myc [10,21],PIM [17], DM1 [22], b-globin [23], PLK [17], and H19 [24,25] — were used. Ab, anti-RNA polymerase II antibody (for panel A) and anti-CTCF rabbit polyclonal antibody (forpanel B); No Ab, no antibody; IgG, nonspecific mouse IgG; Actin, anti-actin mouse monoclonal (irrelevant) antibody; His-tag, anti-His-tag rabbit polyclonal (irrelevant)antibody.
186 Solid-phase affinity matrix for ChIP / I. Chernukhin et al. / Anal. Biochem. 412 (2011) 183–188
to offer useful information; however, these results show that theBioVyon matrix consistently provides higher enrichment ratiosthan the other two matrices with which it was compared in thisinvestigation.
This comparative investigation also demonstrated that theexperimental procedures using the rigid BioVyon Protein A col-umns were considerably easier to carry out and much quicker than
either of the other two formats based on loose beads. The BioVyondevices come ready prepared with a fixed amount of Protein Abound to the rigid porous frit within the column. This immediatelyremoves the potential for dispensing errors associated with addingbead-based matrices into the vessels used for the IP process. How-ever, because Protein A in the column is fixed, a researcher wantingto process larger amounts of material per assay would need to add
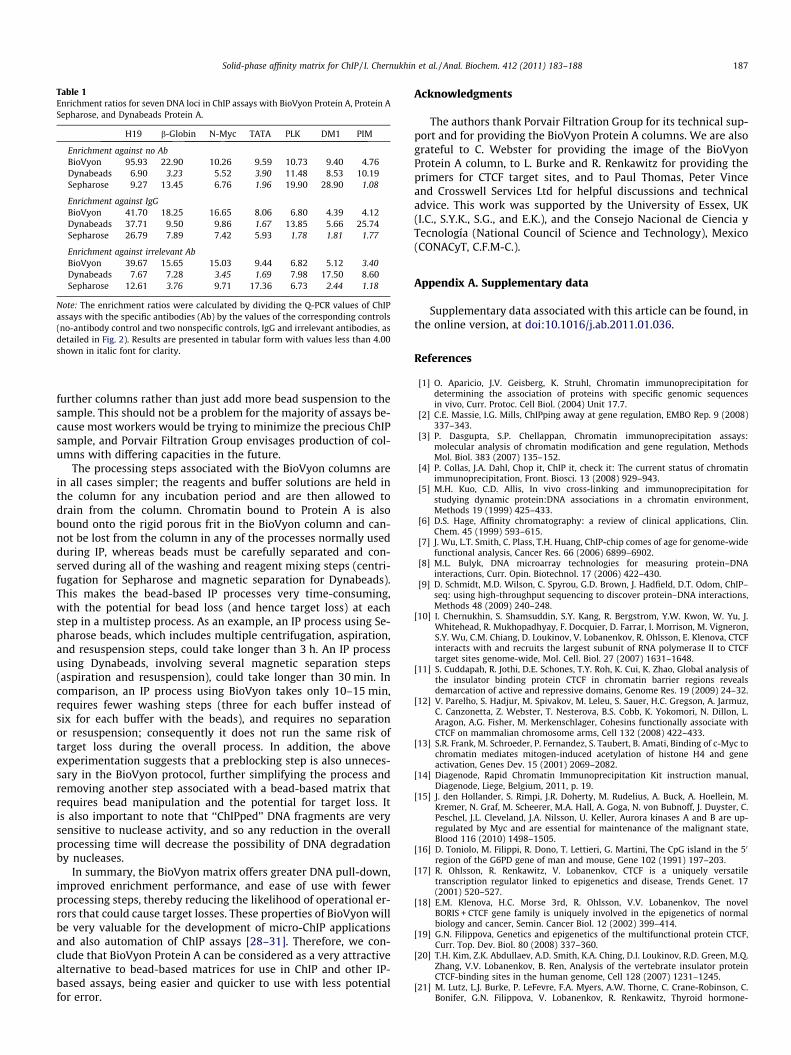
Table 1Enrichment ratios for seven DNA loci in ChIP assays with BioVyon Protein A, Protein ASepharose, and Dynabeads Protein A.
H19 b-Globin N-Myc TATA PLK DM1 PIM
Enrichment against no AbBioVyon 95.93 22.90 10.26 9.59 10.73 9.40 4.76Dynabeads 6.90 3.23 5.52 3.90 11.48 8.53 10.19Sepharose 9.27 13.45 6.76 1.96 19.90 28.90 1.08
Enrichment against IgGBioVyon 41.70 18.25 16.65 8.06 6.80 4.39 4.12Dynabeads 37.71 9.50 9.86 1.67 13.85 5.66 25.74Sepharose 26.79 7.89 7.42 5.93 1.78 1.81 1.77
Enrichment against irrelevant AbBioVyon 39.67 15.65 15.03 9.44 6.82 5.12 3.40Dynabeads 7.67 7.28 3.45 1.69 7.98 17.50 8.60Sepharose 12.61 3.76 9.71 17.36 6.73 2.44 1.18
Note: The enrichment ratios were calculated by dividing the Q-PCR values of ChIPassays with the specific antibodies (Ab) by the values of the corresponding controls(no-antibody control and two nonspecific controls, IgG and irrelevant antibodies, asdetailed in Fig. 2). Results are presented in tabular form with values less than 4.00shown in italic font for clarity.
Solid-phase affinity matrix for ChIP / I. Chernukhin et al. / Anal. Biochem. 412 (2011) 183–188 187
further columns rather than just add more bead suspension to thesample. This should not be a problem for the majority of assays be-cause most workers would be trying to minimize the precious ChIPsample, and Porvair Filtration Group envisages production of col-umns with differing capacities in the future.
The processing steps associated with the BioVyon columns arein all cases simpler; the reagents and buffer solutions are held inthe column for any incubation period and are then allowed todrain from the column. Chromatin bound to Protein A is alsobound onto the rigid porous frit in the BioVyon column and can-not be lost from the column in any of the processes normally usedduring IP, whereas beads must be carefully separated and con-served during all of the washing and reagent mixing steps (centri-fugation for Sepharose and magnetic separation for Dynabeads).This makes the bead-based IP processes very time-consuming,with the potential for bead loss (and hence target loss) at eachstep in a multistep process. As an example, an IP process using Se-pharose beads, which includes multiple centrifugation, aspiration,and resuspension steps, could take longer than 3 h. An IP processusing Dynabeads, involving several magnetic separation steps(aspiration and resuspension), could take longer than 30 min. Incomparison, an IP process using BioVyon takes only 10–15 min,requires fewer washing steps (three for each buffer instead ofsix for each buffer with the beads), and requires no separationor resuspension; consequently it does not run the same risk oftarget loss during the overall process. In addition, the aboveexperimentation suggests that a preblocking step is also unneces-sary in the BioVyon protocol, further simplifying the process andremoving another step associated with a bead-based matrix thatrequires bead manipulation and the potential for target loss. Itis also important to note that ‘‘ChIPped’’ DNA fragments are verysensitive to nuclease activity, and so any reduction in the overallprocessing time will decrease the possibility of DNA degradationby nucleases.
In summary, the BioVyon matrix offers greater DNA pull-down,improved enrichment performance, and ease of use with fewerprocessing steps, thereby reducing the likelihood of operational er-rors that could cause target losses. These properties of BioVyon willbe very valuable for the development of micro-ChIP applicationsand also automation of ChIP assays [28–31]. Therefore, we con-clude that BioVyon Protein A can be considered as a very attractivealternative to bead-based matrices for use in ChIP and other IP-based assays, being easier and quicker to use with less potentialfor error.
Acknowledgments
The authors thank Porvair Filtration Group for its technical sup-port and for providing the BioVyon Protein A columns. We are alsograteful to C. Webster for providing the image of the BioVyonProtein A column, to L. Burke and R. Renkawitz for providing theprimers for CTCF target sites, and to Paul Thomas, Peter Vinceand Crosswell Services Ltd for helpful discussions and technicaladvice. This work was supported by the University of Essex, UK(I.C., S.Y.K., S.G., and E.K.), and the Consejo Nacional de Ciencia yTecnología (National Council of Science and Technology), Mexico(CONACyT, C.F.M-C.).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ab.2011.01.036.
References
[1] O. Aparicio, J.V. Geisberg, K. Struhl, Chromatin immunoprecipitation fordetermining the association of proteins with specific genomic sequencesin vivo, Curr. Protoc. Cell Biol. (2004) Unit 17.7.
[2] C.E. Massie, I.G. Mills, ChIPping away at gene regulation, EMBO Rep. 9 (2008)337–343.
[3] P. Dasgupta, S.P. Chellappan, Chromatin immunoprecipitation assays:molecular analysis of chromatin modification and gene regulation, MethodsMol. Biol. 383 (2007) 135–152.
[4] P. Collas, J.A. Dahl, Chop it, ChIP it, check it: The current status of chromatinimmunoprecipitation, Front. Biosci. 13 (2008) 929–943.
[5] M.H. Kuo, C.D. Allis, In vivo cross-linking and immunoprecipitation forstudying dynamic protein:DNA associations in a chromatin environment,Methods 19 (1999) 425–433.
[6] D.S. Hage, Affinity chromatography: a review of clinical applications, Clin.Chem. 45 (1999) 593–615.
[7] J. Wu, L.T. Smith, C. Plass, T.H. Huang, ChIP-chip comes of age for genome-widefunctional analysis, Cancer Res. 66 (2006) 6899–6902.
[8] M.L. Bulyk, DNA microarray technologies for measuring protein–DNAinteractions, Curr. Opin. Biotechnol. 17 (2006) 422–430.
[9] D. Schmidt, M.D. Wilson, C. Spyrou, G.D. Brown, J. Hadfield, D.T. Odom, ChIP–seq: using high-throughput sequencing to discover protein–DNA interactions,Methods 48 (2009) 240–248.
[10] I. Chernukhin, S. Shamsuddin, S.Y. Kang, R. Bergstrom, Y.W. Kwon, W. Yu, J.Whitehead, R. Mukhopadhyay, F. Docquier, D. Farrar, I. Morrison, M. Vigneron,S.Y. Wu, C.M. Chiang, D. Loukinov, V. Lobanenkov, R. Ohlsson, E. Klenova, CTCFinteracts with and recruits the largest subunit of RNA polymerase II to CTCFtarget sites genome-wide, Mol. Cell. Biol. 27 (2007) 1631–1648.
[11] S. Cuddapah, R. Jothi, D.E. Schones, T.Y. Roh, K. Cui, K. Zhao, Global analysis ofthe insulator binding protein CTCF in chromatin barrier regions revealsdemarcation of active and repressive domains, Genome Res. 19 (2009) 24–32.
[12] V. Parelho, S. Hadjur, M. Spivakov, M. Leleu, S. Sauer, H.C. Gregson, A. Jarmuz,C. Canzonetta, Z. Webster, T. Nesterova, B.S. Cobb, K. Yokomori, N. Dillon, L.Aragon, A.G. Fisher, M. Merkenschlager, Cohesins functionally associate withCTCF on mammalian chromosome arms, Cell 132 (2008) 422–433.
[13] S.R. Frank, M. Schroeder, P. Fernandez, S. Taubert, B. Amati, Binding of c-Myc tochromatin mediates mitogen-induced acetylation of histone H4 and geneactivation, Genes Dev. 15 (2001) 2069–2082.
[14] Diagenode, Rapid Chromatin Immunoprecipitation Kit instruction manual,Diagenode, Liege, Belgium, 2011, p. 19.
[15] J. den Hollander, S. Rimpi, J.R. Doherty, M. Rudelius, A. Buck, A. Hoellein, M.Kremer, N. Graf, M. Scheerer, M.A. Hall, A. Goga, N. von Bubnoff, J. Duyster, C.Peschel, J.L. Cleveland, J.A. Nilsson, U. Keller, Aurora kinases A and B are up-regulated by Myc and are essential for maintenance of the malignant state,Blood 116 (2010) 1498–1505.
[16] D. Toniolo, M. Filippi, R. Dono, T. Lettieri, G. Martini, The CpG island in the 50
region of the G6PD gene of man and mouse, Gene 102 (1991) 197–203.[17] R. Ohlsson, R. Renkawitz, V. Lobanenkov, CTCF is a uniquely versatile
transcription regulator linked to epigenetics and disease, Trends Genet. 17(2001) 520–527.
[18] E.M. Klenova, H.C. Morse 3rd, R. Ohlsson, V.V. Lobanenkov, The novelBORIS + CTCF gene family is uniquely involved in the epigenetics of normalbiology and cancer, Semin. Cancer Biol. 12 (2002) 399–414.
[19] G.N. Filippova, Genetics and epigenetics of the multifunctional protein CTCF,Curr. Top. Dev. Biol. 80 (2008) 337–360.
[20] T.H. Kim, Z.K. Abdullaev, A.D. Smith, K.A. Ching, D.I. Loukinov, R.D. Green, M.Q.Zhang, V.V. Lobanenkov, B. Ren, Analysis of the vertebrate insulator proteinCTCF-binding sites in the human genome, Cell 128 (2007) 1231–1245.
[21] M. Lutz, L.J. Burke, P. LeFevre, F.A. Myers, A.W. Thorne, C. Crane-Robinson, C.Bonifer, G.N. Filippova, V. Lobanenkov, R. Renkawitz, Thyroid hormone-

188 Solid-phase affinity matrix for ChIP / I. Chernukhin et al. / Anal. Biochem. 412 (2011) 183–188
regulated enhancer blocking: cooperation of CTCF and thyroid hormonereceptor, EMBO J. 22 (2003) 1579–1587.
[22] G.N. Filippova, C.P. Thienes, B.H. Penn, D.H. Cho, Y.J. Hu, J.M. Moore, T.R.Klesert, V.V. Lobanenkov, S.J. Tapscott, CTCF-binding sites flank CTG/CAGrepeats and form a methylation-sensitive insulator at the DM1 locus, Nat.Genet. 28 (2001) 335–343.
[23] C.M. Farrell, A.G. West, G. Felsenfeld, Conserved CTCF insulator elements flankthe mouse and human b-globin loci, Mol. Cell. Biol. 22 (2002) 3820–3831.
[24] A.C. Bell, G. Felsenfeld, Methylation of a CTCF-dependent boundary controlsimprinted expression of the Igf2 gene, Nature 405 (2000) 482–485.
[25] D. Takai, F.A. Gonzales, Y.C. Tsai, M.J. Thayer, P.A. Jones, Large scale mapping ofmethylcytosines in CTCF-binding sites in the human H19 promoter andaberrant hypomethylation in human bladder cancer, Hum. Mol. Genet. 10(2001) 2619–2626.
[26] A.J. Wilson, D.S. Byun, S. Nasser, L.B. Murray, K. Ayyanar, D. Arango, M.Figueroa, A. Melnick, G.D. Kao, L.H. Augenlicht, J.M. Mariadason, HDAC4
promotes growth of colon cancer cells via repression of p21, Mol. Biol. Cell 19(2008) 4062–4075.
[27] J.W. Kim, P. Gao, Y.C. Liu, G.L. Semenza, C.V. Dang, Hypoxia-inducible factor 1and dysregulated c-Myc cooperatively induce vascular endothelial growthfactor and metabolic switches hexokinase 2 and pyruvate dehydrogenasekinase 1, Mol. Cell. Biol. 27 (2007) 7381–7393.
[28] J.A. Dahl, P. Collas, MicroChIP: chromatin immunoprecipitation for small cellnumbers, Methods Mol. Biol. 567 (2009) 59–74.
[29] J.A. Dahl, A.H. Reiner, P. Collas, Fast genomic muChIP–chip from 1000 cells,Genome Biol. 10 (2009) R13.
[30] S. Flanagin, J.D. Nelson, D.G. Castner, O. Denisenko, K. Bomsztyk, Microplate-based chromatin immunoprecipitation method matrix ChIP: a platform tostudy signaling of complex genomic events, Nucleic Acids Res. 36 (2008) e17.
[31] A.R. Wu, J.B. Hiatt, R. Lu, J.L. Attema, N.A. Lobo, I.L. Weissman, M.F. Clarke, S.R.Quake, Automated microfluidic chromatin immunoprecipitation from 2000cells, Lab Chip 9 (2009) 1365–1370.























![Research Paper JNK/AP1 Pathway Regulates MYC ...Chromatin immunoprecipitation assays (ChIP) ChIP analysis was performed as previously described [11]. Chromatin solutions were precipitated](https://img.pdfslide.net/doc/110x75/608625bcea8a6a2e9165f1fb/research-paper-jnkap1-pathway-regulates-myc-chromatin-immunoprecipitation-assays.jpg)





