Embed Size (px)
Citation preview

BREEDING FOR RESISTANCE IN RICE
Gurdev S . Khush
The International Rice Research Institute Los Baiios, Laguna, Philippines
Rice is the host for more disease and insect organisms than any other cereal crop. More than 60 diseases and over 100 insect species are known to attack the rice crop. Some of these are of major international importance.
During the last 10 years major changes have occurred in the varietal composition and cultural practices for rice. High yielding varieties are now planted on approximately 25% of the 130 million hectares planted to rice all over the world. These varieties are characterized by early maturity, photo- period insensitivity, short stature, high tillering, and dark green erect leaves. Approximately 80 improved plant type varieties have replaced literally hundreds of tall traditional cultivars, thereby reducing the genetic variability of the crop.
In the wake of introduction of varieties with improved plant type, farmers have started to use improved cultural practices such as more fertilizer and higher plant populations per unit area. Development of irrigation facilities and availability of early maturing, photoperiod-insensitive varieties have enabled the farmers in tropical Asia to grow successive crops of rice throughout the year on large areas.
Reduced genetic variability, improved cultural practices, and continuous cropping with rice, intended for increased rice production, have increased the genetic vulnerability of the crop. Within the last few years serious outbreaks of diseases and insects on rice have occurred in several countries. Very little research has been done on the chemical control of rice diseases in the tropics. Several insecticides have been identified, but chemical control of high insect populations for prolonged periods in tropical climates where insect generations overlap throughout the year is very expensive. Social and economic conditions in the tropics present other obstacles to the chemical control of rice diseases and insects. The use of host resistance to control diseases and insects is the most logical approach to overcome these production constraints.
MAJOR DISEASES AND INSECTS OF RICE
In most of the rice growing areas, more than one disease or insect cause serious yield losses, In Latin America, for example, blast, hoja blanca, and Sogatodes oryzicola are the factors limiting to rice production. In Africa, blast and stemborers take a serious toll of rice yields. In Asia, where 92% of the rice is produced and consumed, more than a dozen diseases and insects cause losses of epidemic proportions. In tropical Asia, the disease and insect problem is most serious because of year round favorable climate and long history of rice cultivation with consequent evolution of many disease and pest organisms. One year there may be an epidemic of bacterial blight, the next year green leafhopper and tungro may cause serious damage and the following year an outbreak of brown planthopper and grassy stunt might occur. Therefore, to minimize the yield losses from diseases and insects, varieties with multiple
296

Khush: Breeding for Resistance in Rice 297
resistance to most of the major diseases and insects are required. Five diseases (blast, sheath blight, bacterial blight, tungro, and grassy stunt) and six insect species (brown planthopper, green leafhopper, stemborer, gall midge, white- backed planthopper, and whorl maggot) are of common occurrence in most of the countries of tropical and subtropical Asia. Therefore, the development and dissemination of improved germplasm with multiple resistance to these diseases and insects has been the major objective of the Genetic Evaluation and Utiliza- tion program at IRRI for several years.
SOURCES OF RESISTANCE
Sources of resistance to most of the major diseases and insects have been identified and are discussed in two other papers in this volume by my colleagues Drs. S. H. Ou and M. D. Pathak. In general, several parental sources of resistance are known for most of the diseases and insects except grassy stunt. Only one source of resistance to grassy stunt is known, however. For some of the diseases such as blast and bacterial leaf blight, and some insects such as brown planthopper and green leafhopper, more than a hundred resistant varie- ties are known. The sources of resistance are in poor agronomic backgrounds, however. Most of the varieties are tall, with droopy leaves and weak stems and consequently have a low yield potential.
IMPROVING THE SOURCES OF RESISTANCE
As a first step the sources of resistance were transferred to improved plant- type background (short stature, erect leaves, sturdy stems, high tillering). This “conversion” was carried out by crossing the tall donor parents with an im- proved plant-type parent, growing fairly large F, populations (2,000-5,000 plants) and selecting improved plant-type segregants with resistance to the disease or insect in question. These selected lines were evaluated for plant type, grain quality, and the resistance trait under study for two or three generations. Superior lines were evaluated for yield in replicated trials. Several donor parents, where available, were used as sources of resistance for each disease and insect. Between 1965 and 1969, IR 8, several selections of IR 262 (IR 262-43-8 in particular), and IR 24 were used as improved plant-type parents. In the case of primitive donor parents with an extremely poor plant type, such as Oryza nivara, one or two backcrosses were made using the improved plant-type lines as recurrent parent. By the end of 1970 a large number of lines with im- proved plant type resistant to one or two diseases and insects became available.
Programs on breeding for resistance to blast and bacterial blight were started in 1965, to tungro and stemborers in 1966, to brown planthopper and green leafhopper in 1968, to grassy stunt in 1969, to gall midge in 1971, to sheath blight in 1974, and to whitebacked planthopper and whorl maggot in 1975. Progress made in developing improved germ plasm with resistance to blast, bacterial blight, tungro, grassy stunt, brown planthopper, and green leafhopper is shown in FIGURE 1 . Whereas only 12% of the entries in our replicated yield trials planted in the wet season of 1969 were resistant to blast, this proportion rose to 60% in the replicated yield trials planted in the wet season of 1974. Similarly, only 17% of the entries in the replicated yield trials of the 1969 wet
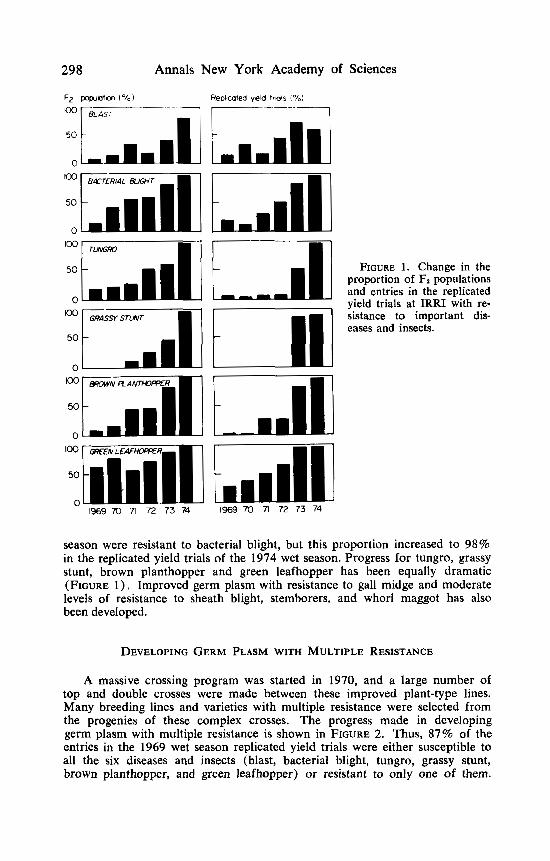
298 Annals New York Academy of Sciences
I00
50
0 I00
50
0
111 FIGURE 1. Change in the
proportion of F2 populations and entries in the replicated yield trials at IRRI with re- sistance to important dis- eases and insects.
season were resistant to bacterial blight, but this proportion increased to 98% in the replicated yield trials of the 1974 wet season. Progress for tungro, grassy stunt, brown planthopper and green leafhopper has been equally dramatic (FIGURE 1). Improved germ plasm with resistance to gall midge and moderate levels of resistance to sheath blight, stemborers, and whorl maggot has also been developed.
DEVELOPING GERM PLASM WITH MULTIPLE RESISTANCE
A massive crossing program was started in 1970, and a large number of top and double crosses were made between these improved plant-type lines. Many breeding lines and varieties with multiple resistance were selected from the progenies of these complex crosses. The progress made in developing germ plasm with multiple resistance is shown in FIGURE 2. Thus, 87% of the entries in the 1969 wet season replicated yield trials were either susceptible to all the six diseases and insects (blast, bacterial blight, tungro, grassy stunt, brown planthopper, and green leafhopper) or resistant to only one of them.
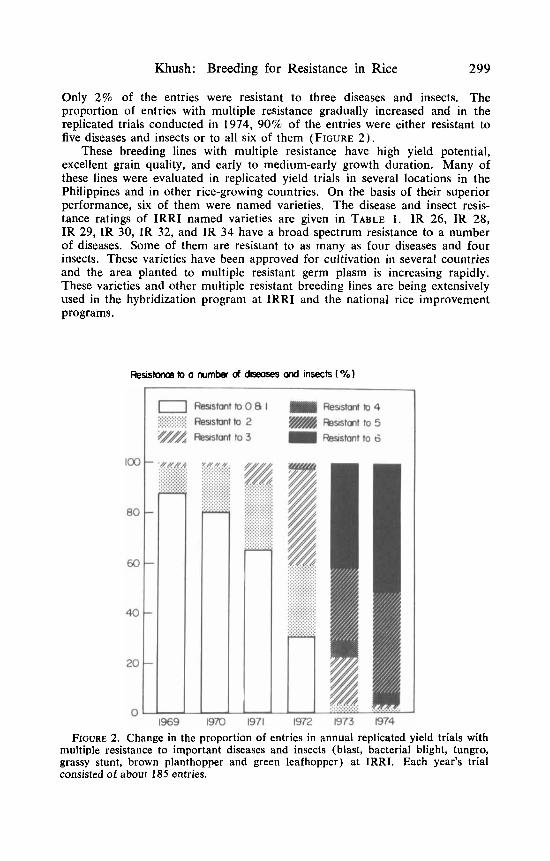
Khush: Breeding for Resistance in Rice 299
Only 2% of the entries were resistant to three diseases and insects. The proportion of entries with multiple resistance gradually increased and in the replicated trials conducted in 1974, 90% of the entries were either resistant to five diseases and insects or to all six of them (FIGURE 2) .
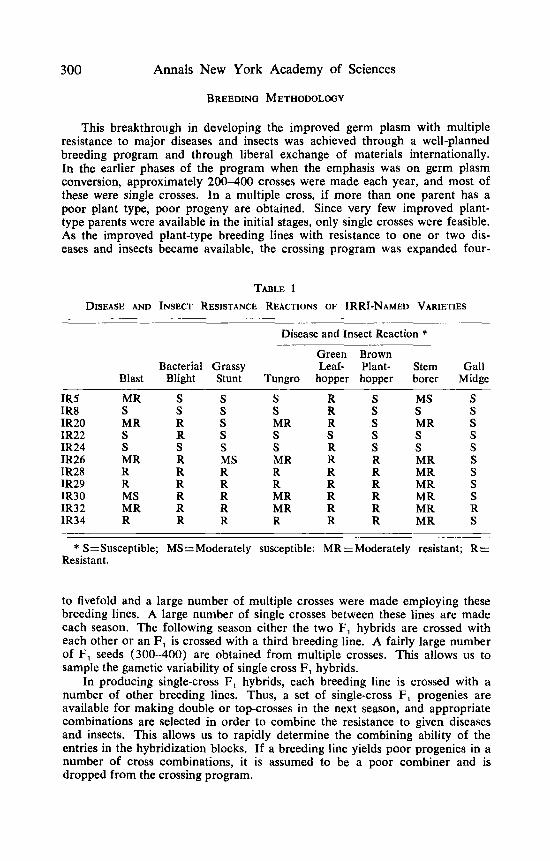
These breeding lines with multiple resistance have high yield potential, excellent grain quality, and early to medium-early growth duration. Many of these lines were evaluated in replicated yield trials in several locations in the Philippines and in other rice-growing countries. On the basis of their superior performance, six of them were named varieties. The disease and insect resis- tance ratings of IRRI named varieties are given in TABLE 1. IR 26, IR 28, IR 29, IR 30, IR 32, and IR 34 have a broad spectrum resistance to a number of diseases. Some of them are resistant to as many as four diseases and four insects. These varieties have been approved for cultivation in several countries and the area planted to multiple resistant germ plasm is increasing rapidly. These varieties and other multiple resistant breeding lines are being extensively used in the hybridization program at IRRI and the national rice improvement programs.
FIGURE 2. Change in the proportion of entries in annual replicated yield trials with multiple resistance to important diseases and insects (blast, bacterial blight, tungro, grassy stunt, brown planthopper and green leafhopper) at IRRI. Each year’s trial consisted of about 185 entries.
300 Annals New York Academy of Sciences
BREEDING METHODOLOGY
This breakthrough in developing the improved germ plasm with multiple resistance to major diseases and insects was achieved through a well-planned breeding program and through liberal exchange of materials internationally. In the earlier phases of the program when the emphasis was on germ plasm conversion, approximately 200-400 crosses were made each year, and most of these were single crosses. In a multiple cross, if more than one parent has a poor plant type, poor progeny are obtained. Since very few improved plant- type parents were available in the initial stages, only single crosses were feasible. As the improved plant-type breeding lines with resistance to one or two dis- eases and insects became available, the crossing program was expanded four-
TABLE 1 DISEASE AND INSECT RESISTANCE REACTIONS OF IRRI-NAMED VARIETIES
Disease and Insect Reaction * Green Brown
Bacterial Grassy Leaf- Plant- Stem Gall Blast Blight Stunt Tungro hopper hopper borer Midge
IR5 IR8 IR20 IR22 IR24 IR26 IR28 IR29 IR30 IR32 IR34
MR S S S R S S S S S R S MR R S MR R S S R S S S S S S S S R S MR R MS MR R R R R R R R R R R R R R R MS R R MR R R MR R R MR R R R R R R R R
MS S MR S S MR MR MR MR MR MR
S S S S S S S S S R S
* S=Susceptible; MS=Moderately susceptible; MRZModerately resistant; R = Resistant.
to fivefold and a large number of multiple crosses were made employing these breeding lines. A large number of single crosses between these lines are made each season. The following season either the two F, hybrids are crossed with each other or an F, is crossed with a third breeding line. A fairly large number of F, seeds (300-400) are obtained from multiple crosses. This allows us to sample the gametic variability of single cross F, hybrids.
In producing single-cross F, hybrids, each breeding line is crossed with a number of other breeding lines. Thus, a set of single-cross F, progenies are available for making double or top-crosses in the next season, and appropriate combinations are selected in order to combine the resistance to given diseases and insects. This allows us to rapidly determine the combining ability of the entries in the hybridization blocks. If a breeding line yields poor progenies in a number of cross combinations, it is assumed to be a poor combiner and is dropped from the crossing program.

Khush: Breeding for Resistance in Rice 30 1
The pedigree method is employed almost exclusively in handling the segre- gating materials. Thus the selection is based on comprehensive records on the disease and insect reactions of each line, and, in the case of F, and advanced generation lines, on the reaction of ancestral lines as well. The bulk method is not used as it does not permit the concurrent screening for a number of diseases and insects. The backcross method has not been employed either because of a lack of suitable recurrent parents. A few backcrosses were made in the crosses with 0. nivara for grassy stunt resistance; IR 8, IR 20, and IR 24 were used as recurrent parents. After three or four backcrosses, we obtained grassy-stunt-resistant breeding lines similar to the recurrent parent, but they lacked resistance to other important diseases and insects such as tungro and brown planthopper and were again entered in the crossing program. However, suitable recurrent parents with multiple resistance are now available and are being employed in the backcrossing program for incorporating resis- tance to whitebacked planthopper.
The success of the breeding program for disease and insect resistance depends to a large measure upon the fidelity, practicability, and volume of materials that can be handled by a screening technique, Various greenhouse and field screening methods employed at IRRI have been adapted to handle a large volume of breeding materials. On the average we grow about 50,000 pedigree rows each year. A great majority of them are screened for reaction to all the major diseases and insects. If possible, we try to subject the breeding materials to artificially created or naturally occurring disease and insect epi- demics. Most of the nurseries are grown without any insecticide treatment. This allows for the build-up of huge populations of planthopper, leafhoppers, and stemborers. Presence of planthoppers, and leafhoppers in large numbers also ensures the spread of virus diseases in the nurseries. Sometimes we plant artificially virus inoculated plants around the borders of the nurseries to provide the inoculum. The insect populations are manipulated by applying selective insecticides. For example, at the IRRI farm, diazinon has no toxic effect on the brown planthopper but it kills all the predators and other natural enemies of this insect. Thus, by judicious application of diazinon we have been inducing outbreaks of brown planthopper in our nurseries. This leads to increased incidence of grassy stunt as well.
Every effort is made to eliminate the susceptible materials in the early generations. The screening starts even in the F, generation of a cross. Let US consider a double cross between four parents of which A is resistant to bac- terial blight, B is resistant to grassy stunt, C is resistant to brown planthopper, and D is resistant to green leafhopper. All these traits are under the control of single dominant genes that segregate independently of each other. About 400 seeds from the double cross A / B / / C / D are obtained. These seeds are germinated and inoculated with grassy stunt virus in the greenhouse. Approxi- mately 50% of the seedlings are susceptible and are eliminated. The remaining 200 are transplanted in the field and inoculated with bacterial blight. About 50% of these are susceptible and are rooted out. The remaining 100 plants are harvested separately. Two small seed samples are taken from each, and the progeny are tested for resistance to brown planthopper and green leaf- hopper. Those carrying the brown planthopper resistance gene (50% ) and the ones carrying the green leafhopper resistance gene ( 5 0 % ) are identified. The F, populations are grown only from those carrying both the genes (25-30 plants). Thus, by judicious and timely screening the original F, sample of 400

302 Annals New York Academy of Sciences
111561 111737
11833 112042 BL 1 QLH
EL BET GSBPH GLH
FIGURE 3. Pedigree of IR 28, IR 29, and IR 34. The progress in combining to- gether the resistance to six major diseases and insects from several parents is indi- cated. Disease and insect resistance: BL=blast disease; BBzbacterial blight disease; T=tungro virus disease; GS= grassy stunt virus disease; BPH=brown planthopper; and GLH=green leafhopper.
is reduced to 25-30 plants. The F2 populations grown from these plants segregate for the four resistance genes. The populations are subjected to appropriate disease and insect pressures. Agronomically desirable plants with multiple resistance are selected and rescreened in the F, and F, generations to obtain true breeding lines. The complexity of crossing and screening procedures adopted in developing germ plasm with multiple resistance is exemplified by the pedigree of IR 28, IR 29 and IR 34 (FIGURE 3 ) . Eight different varieties contributed to their ancestry, and the final germ plasm developed from 15 different crossing steps.
A large number of multiple crosses between breeding lines of improved plant type, known combining ability, and resistance to a number of diseases and insects are made each season and screened according to the procedures briefly outlined above. New parents with different sources of resistance are constantly included in the crossing program. This integrated varietal development pro- gram has resulted in superior germ plasm with resistance to all the major diseases and insects.
INHERITANCE OF RESISTANCE
We are constantly concerned about the stability of resistance in the newly developed germ plasm. There is growing support for the contention that the resistance governed by polygenes (also referred to as general resistance or horizontal resistance) is longer lasting than resistance governed by major genes (specific or vertical resistance). When we initiated the program on breeding for disease and insect resistance in rice, very little was known about the genetics of resistance. We started using whatever donor parents were available as sources of resistance and developed improved germ plasm with multiple resis-

Khush: Breeding for Resistance in Rice 303
tance to as many as four diseases and four insects within a short period of 7-8 years. During this period we concurrently investigated the mode of inheritance of resistance to some of the diseases and insects.
Resistance to bacterial blight is under monogenic control. Six different loci for resistance have been identified. Three dominant genes for resistance to Japanese isolates of the bacterium were identified in Japan. X a l and Xu2 are closely linked,l8 whereas Xa3 , formerly called X u w, segregates independently of X a l and Xu2:' Three genes for resistance to the Philippine isolates are
11, l2 The dominant gene, X a 4 , is widely distributed, whereas the recessive gene xa5 is restricted in its distribution to Bangladesh and the ad- joining states of India. Another, as yet undesignated, gene has been found in the varieties Zenith and Malagkit Sungsong (TABLE 2). We are analyzing a large number of resistant varieties to find new genes for resistance.
A considerable amount of work has been done on the genetics of resistance to blast in Japan, and 10 different loci for resistance to the Japanese races of blast have been identified (Kiyosawa and the paper by Kiyosawa in this volume). Very little work has been done on the genetics of resistance to tropical races and specific genes for resistance have not been identified.
The resistance of 0. nivara, the only known source of resistance to grassy stunt, is controlled by a single dominant gene.6 A thorough genetic analysis of tungro resistance has not been done, but it appears to be under major gene control. Differences in resistance to sheath blight are of a quantitative nature. The inheritance of resistance to brown planthopper has been studied in 40 resistant varieties (TABLE 3) , and four genes for resistance, two of them
TABLE 2 SOME BACTERIAL-BLIGHT-RESISTANT VARIETIES AND THE RESISTANCE GENES
POSSESSED BY THEM
Variety Resistance Gene Reference
Kogyoku Zensho 26 Norin 27 Asakaze Bornba Rantaj-emas Tadukan Tetep Wase Aikoku 3 TKM6 Sigadis IR 22 Syntha MTUlS Horn Thong BJ 1 DZ 192 Dular Hashikalmi Zenith Malagkit Sungsong
X a l X a l X u 1 X u I X a l X a l , Xu2 X a l , Xu2 X a l , Xu2 X a 3 , ( X a W ) Xu4 Xu4 Xu4 Xu4 Xu4 Xu4 X U 5
xu5 xu5 xu5 ? ?
Sakaguchi (1967)= Sakaguchi (1967)" Sakaguchi (1967)" Sakaguchi (1967)" Sakaguchi (1967)" Sakaguchi (1967)'' Sakaguchi (1967)'' Sakaguchi (1967)'' Ezuka et al. ( 1975)4 Petpsisit et al. (1976)Ia Petpsisit et al. ( 1976)Ia Petpsisit et a/. (1976)" Olufowote et al. (1976)" Olufowote ef al. (1976)'' Librojo et al. (1976)' Petpsisit et a/. ( 1976)" Petpsisit et a / . (1976)" Librojo et al. (1976)' Librojo et al. (1976)' Olufowote et al. (1976)" Olufowote et al. ( 1976)u

304 Annals New York Academy of Sciences
TABLE 3
POSSESSED BY THEM SOME BROWN-PLANTHOPPER-RESISTANT VARIETIES AND THE RESISTANCE GENES
Variety/ Resistance Selection Gene Reference
Mudgo MTU 15 co 22 ASD 7 MGL 2 PTB 18 H 105 IR1154-243 IR747B2-6 Anbaw C7 Tibiriwewa Balamawee co 10 Heenakkulama MTU 9 Sinnakayam SLO 12 Sudhubalawee Sudurvi 305 ASD 9 Dikwee 328 Hathiel Kosatawee Madayal Mahadikwee Malkora M.I. 329 Murungakayan302 Ovarkaruppan Palasithari 601
Seruvellai Sinna Karuppan Vellailangayan Rathuheenati Babawee
PK- 1
Bhp 1 Bhp 1 Bhp 1 bph 2 Bph 1 bph 2 bph 2 bph 2 Bph 1 bph 2 Bph 1 Bph 1 Bph 1 Bph 1 Bph 1 Bph 1 Bph 1 Bph 1 Bph 1 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 bph 2 Bph 3 buh 4
Athwal era / . (1971)' Athwal er a/ . ( 1971)' Athwal et al. ( 1971)' Athwal e! al. (1971)' Athwal & Pathak (1972)' Athwal & Pathak (1972)l Martinez & Khush (1974)" Martinez & Khush (1974)'' Martinez & Khush (1974)'O Lakshminara ana & Khush (1976)*
Ptb 21 H5
Kaosen IR9-60
? * Lakshminarayana & Khush (1976)" bph 2 Chang (1975)' bph 2 Chang (1975)'
-yu 12 bph 2 Chang (1975)'
* This variety has two genes for resistance but their allelic relationships are not known.

Khush: Breeding for Resistance in Rice 305
dominant and two recessive, have been identified. Bphl and bph2 are closely linked, but Bph3 and bph4 segregate independently of each other as well of Bphl and bph2. Bphl and bph2 have been incorporated into improved germ plasm. The resistance of IR 26, IR 28, IR 29, IR 30, and IR 34 is governed by Bphl, whereas bph2 conveys resistance in IR 32.
Five independent genes for resistance to green leafhopper have been iden- tified from the analysis of 13 resistant varieties (TABLE 4). Four of the genes are dominant, whereas one is recessive. Glhl, Glh2 and GIh3 have been incorporated into improved germ plasm. Resistance to gall midge is also con- veyed by a major gene," and several improved plant-type varieties with gall midge resistance have developed. Resistance to stemborers appears to be under polygenic control.
TABLE 4 SOME GREEN-LEAFHOPPER-RESISTANT VARIETIES AND THE RESISTANCE GENES
POSSESSED BY THEM
Variety/ Resistance Selection Gene Reference
Pankhari 203 G l h 1 Athwal et al. (1971y ASD 7 G l h 2 Athwal ef al. (1971y IR8 G l h 3 Athwal et al. ( 1971)2 Ptb 18 G l h l f ? " Athwal & Pathak (1972)' J hingasail G l h I Siwi & Khush ( 1976)15 Godalki G l h 2 Siwi & Khush (1976)'" Lien-tsan G l h 2 Siwi & Khush ( 1 9 7 6 y Palasithari 601 G l h 2 Siwi & Khush (1976)= Betong G l h 3 Siwi & Khush (1976)13 DNJ 97 G I h 3 Siwi & Khush ( 1976)15 H.5 G l h 3 Siwi & Khush ( 1976)15 Ptb 8 g l h 4 Siwi & Khush ( 1976)15 ASD 8 G l h 5 Siwi & Khush ( 1976)15
0 Ptb 18 has two genes for resistance. One of them is allelic to G I h I, but the allelic relationship of the second is not known.
STABILITY OF RESISTANCE
Some information on the stability of vertical or major gene resistance in rice is available. Bacterial-blight-resistant varieties with the major gene X a l for resistance have been grown in Japan for the last 30 years. In most of the areas the resistance has held up quite well. A strain of the bacterium virulent to X a l did appear in the Beniya district of Kyushu province but apparently has remained confined to that area. Similarly, IR 20 and IR 26 having Xu4 for resistance to bacterial blight have been grown widely for several years in the tropics and resistance has held up except in a small area of the Philippines where a strain of bacterium moderately virulent to Xu4 has appeared. This strain has also remained localized, and even in the locality where this strain occurs, varieties with Xu4 suffer very little damage from bacterial blight. Similarly, several bacterial-blight-resistant varieties such as Benong, Sigadis,

306 Annals New York Academy of Sciences
Syntha, and Dewi Tara have been grown in Indonesia for 10-20 years and TKM6, MTU15 and Co 21 in India for many years. Occurrence of bacterial strains virulent to these varieties under the influence of host resistance has not been reported.
Several green-leafhopper-resistant varieties such as Peta, Intan, and Benga- wan were widely grown in Indonesia and the Philippines for 30-35 years. Several improved plant type varieties such as IR5, IR 8, IR 20, IR 26, and C4-63, which inherited GIW for resistance to green leafhopper from Peta, have also been grown for several years. No clear-cut evidence for the origin of green leafhopper biotypes virulent to Glh3 under the influence of host resistance has been found. For the brown planthopper, however, the situation is different. Within 18 months of their introduction into the British Solomon Islands, the brown-planthopper-resistant varieties became susceptible because of the appearance of a new biotype. Similarly, within two years of its large-scale cultivation in the Philippines, IR 26 was attacked by new biotypes of the insect in two widely separated localities. The germ plasm for resistance to brown planthopper comes from South India and Sri Lanka where biotypes of brown planthopper are virulent to these varieties. These biotypes probably originated under the influence of host resistance.
The influence of host resistance on the insect populations of brown plant- hopper and green leafhopper is obviously different. In my opinion, the differ- ence is due to the differential selection pressure exerted by the resistant varieties on insect populations. The level of resistance to brown planthopper conveyed by Bphl and bph2 is so high that the insect cannot perpetuate itself on resistant varieties. So it must change or be eliminated entirely. On the other hand, the level of resistance to green leafhopper conditioned by Glh3 is only moderate. The insect does some feeding on the resistant plant and is able to reproduce, although at a much lower rate than on susceptible varieties. Thus, it is able to perpetuate itself on these varieties and chances for the origin of more virulent biotypes are considerably lower than that for brown planthopper. Therefore, the useful life of Glh3 may be considerably longer than that of either BphI or bph2. GlhI and Glh2 confer a much higher level of resistance to green leafhopper, comparable to that conferred by Bphl for brown plant- hopper. When varieties having either one of these two genes are grown widely, new biotypes of green leafhopper might arise rapidly. Differences in the in- herent capacity of the two insect species to change under the influence of host resistance may also be responsible for the differences in longevity of resistance to these two insects.
PRESENT AND FUTURE STRATEGY OF BREEDING
Our present strategy is to incorporate the major genes for resistance into an improved plant-type background and combine each of them with major genes for resistance to other diseases and insects. If the germ plasm with a particular gene becomes susceptible, lines with a second gene would be avail- able. Thus, five brown-planthopper-resistant varieties with Bphl for resistance were released. After about two years of cultivation these varieties started showing susceptibility in some localities of the Philippines. However, by that time multiple disease- and insect-resistant germ plasm with bph2 for resistance to brown planthopper was ready and made available to farmers.

Khush: Breeding for Resistance in Rice 307
In the long run, we are endeavoring to identify several major genes for resistance to each disease and insect and transfer these genes into an isogenic background using recurrent parents with resistance to other diseases and insects. Such a program is now underway for resistance to brown planthopper. When several major genes for resistance and corresponding isogenic lines become available, we plan to develop multiline varieties for resistance to this insect. A similar approach merits consideration for resistance to blast.
We are also identifying germ plasm with field tolerance to brown plant- hopper. Such varieties suffer damage from the insect but take longer to be killed than susceptible controls. We believe these differences are governed by minor genes. As a first step these tolerance levels were transferred to a desir- able agronomic background. We are now trying to combine these polygenes for tolerance from several different sources by a process of recurrent selection. For this purpose, we are using a diallel selective mating system, originally proposed by Jensen.5 This method involves: ( 1 ) crossing a number of tolerant parents in all possible combinations; (2) intercrossing the F, populations so obtained in all the possible combinations; (3) screening the double cross F, progeny for resistance; and (4) intercrossing the selected plants found to have better tolerance than either of the parents. The crossing, screening, and selection process will be continued until minor genes from different sources are accumulated and the intensity of the trait is built up.
If the level of horizontal resistance obtained this way is adequate, the germ plasm would be usable directly on the farmers’ fields. However, if ade- quate levels of resistance are not obtained, the tolerant lines would be used as recurrent parents in developing isogenic lines incorporating major genes. Multiline varieties produced from these isogenic lines would utilize major as well as minor genes for resistance to brown planthopper.
REFERENCES
1. ATHWAL, D. S. & M. D. PATHAK. 1972. Genetics of resistance to rice insects. In Rice Breeding. International Rice Research Institute. Los Bafios, Philip- pines. : 375-386.
2. ATHWAL, D. S., M. D. PATHAK, E. H. BACALANGCO & C. D. PURA. 1971. Gene- tics of resistance to brown planthoppers and green leafhoppers in Oryza sativa L. Crop Sci. 11: 747-750.
3. CHANG, W. L. 1975. Inheritance of resistance to brown planthopper in rice. Sabrao J. 7: 53-60.
4. EZUKA, A., 0. HORINO, K. TORIYAMA, H. SHINODA & T. MORINAKA. 1975. In- heritance of resistance of rice variety Wase Aikoku 3 to Xanthornonas oryzae. Bull. Tokai-Kinki Nat. Agr. Expt. Sta. 28: 124-130.
5. JENSEN, N. F. 1970. A diallel selective mating system for cereal breeding. Crop Sci. 10: 629-635.
6. KHUSH, G. S . & K. C. LING. 1974. Inheritance of resistance to grassy stunt virus and its vector in rice. J. Hered. 65: 134-136.
7. KIYOSAWA, S . 1974. Studies on genetics and breeding of blast resistance in rice. Nat. Inst. Agr. Sci. Ser. D., Miscellaneous Publication No. 1. (58 pp.).
8. LAKSHMINARAYANA, A. & G. S. KHUSH. 1976. New genes for resistance to brown planthopper in rice. Crop Sci. 16: In press.
9. LIBROJO, V., H. E. KAUFFMAN & G. S. KHUSH. 1976. Genetic analysis of bac- terial blight resistance in four varieties of rice. Sabrao Jour. 8: In press.
10. MARTINEZ. C. R. & G. S. KHUSH. 1974. Sources and inheritance of resistance

308 Annals New York Academy of Sciences
to brown planthopper in some breeding lines of rice, Oryza sativa L. Crop Sci. 14: 264-267.
11. OLUFOWOTE, J. O., G. S. KHUSH & H. E. KAUFFMAN. 1976. Genetics of bacterial blight resistance in rice. Phytopathology 6 6 In press.
12. PETPISIT, V., G. S. KHUSH & H. E. KAUFFMAN. 1976. Inheritance of resistance to bacterial blight in rice. Crop Sci. 16: In press.
13. SAKAGUCHI, S. 1967. Linkage studies on the resistance to bacterial leaf blight, Xanfhornonas oryzae (Uyeda et Jshiyama) Dowson, in rice. Bull. Nat. Inst. Agr. Sci. Ser. D. 1 6 1-18.
14. SATYANARAYANAIAH, K. & M. V. REDDI. 1972. Inheritance of resistance to insect gall-midge (Pachydiplosis oryzae Wood Mason) in rice. Andhra Agric. J.
15. SIWI, B. H. & G. S. KHUSH. 1976. New genes for resistance to green leafhopper 19(1 & 2): 1-8.
in rice. Crop Sci. 1 6 In press.





























