Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Bulb and Root Rot in Lily (Lilium longiflorum) and Onion (Alliumcepa) in IsraelSara Lebiush-Mordechai1, Orly Erlich1, Marcel Maymon2, Stanley Freeman2, Tslila Ben-David3, Tal Ofek3,Eric Palevsky3 and Leah Tsror (Lahkin)1
1 Department of Plant Pathology and Weed Research, Agricultural Research Organization (ARO), Gilat Research Center, M.P. Negev, Israel
2 Department of Plant Pathology and Weed Research, Agricultural Research Organization (ARO), Beit Dagan, Israel
3 Department of Entomology, Agricultural Research Organization (ARO), Newe-Ya’ar Research Center, Ramat Yishay, Israel
Keywords
bulb diseases, Fusarium, Pythium, Rhizoctonia
Correspondence
L. Tsror (Lahkin), Department of Plant
Pathology and Weed Research, Agricultural
Research Organization (ARO), Gilat Research
Center, M.P. Negev, Israel.
E-mail: [email protected]
Received: June 26, 2013; accepted: November
9, 2013.
doi: 10.1111/jph.12214
Abstract
In the past 10 years, there has been a substantial increase in reports, from
growers and extension personnel, on bulb and root rots in lily (Lilium lon-
giflorum) in Israel. Rot in these plants, when grown as cut flowers, caused
serious economic damage expressed in reduction in yield and quality. In
lily, the fungal pathogens involved in the rot were characterized as binu-
cleate Rhizoctonia AG-A, Rhizoctonia solani, Pythium oligandrum, Fusarium
proliferatum (white and purple isolates) and F. oxysporum, using morpho-
logical and molecular criteria. These fungi were the prevalent pathogens
in diseased plants collected from commercial greenhouses. Pathogenicity
trials were conducted on lily bulbs and onion seedlings under controlled
conditions in a greenhouse to complete Koch’s postulates. Disease symp-
toms on lily were most severe in treatments inoculated with binucleate
Rhizoctonia AG-A, P. oligandrum and F. proliferatum. Plant height was lower
in the above treatments compared with the control plants. The least
aggressive fungus was R. solani. In artificial inoculations of onion, seedling
survival was significantly affected by all fungi. The most pathogenic fun-
gus was F. proliferatum w and the least were isolates of F. oxysporum (II
and III). All fungi were successfully re-isolated from the inoculated plants.
Introduction
Commercial production of lily in Israel, cultivated
mainly in the Upper Gallile, encompasses
approximately 20% of the total flower-growing area
(Luria, Min. Agric., personal communication). Lilium
longiflorum comprises about 90% of the different vari-
eties and is grown in greenhouses (80 ha of nursery,
120 ha of cut flowers) (Luria, Min. Agric., personal
communication). During 2001–2005, there was a sub-
stantial increase in reports from growers and exten-
sion personnel regarding bulb and root rots of lily in
Israel, causing serious economic damage resulting in
reduction in yield and quality of the cut flowers.
The optimal conditions for lily production in green-
houses are also suitable for the development of vari-
ous phytopathogenic agents causing root and bulb
rot, which affect development of plants, flower qual-
ity and total yield. Among the fungal diseases of lily,
basal decay (bulb root) caused by a single infection of
Fusarium oxysporum or compound infections of F. oxy-
sporum with other pathogens is common and the most
serious observed (L€offler et al. 1995; Lawson and Hus
1996). The soil-borne fungi Rhizoctonia, Pythium and
Fusarium were previously reported as pathogens that
occur worldwide and cause serious diseases in Asiatic
lilies (Lawson and Hus 1996). The predominantly
reported symptoms include brownish rot at the base
of the bulb scales, which spreads over the whole bulb.
Plants from field-grown bulbs often continue to
express the disease complex rather than any one sin-
gle disease. If replanted and grown again under warm
conditions, the bulbs may be damaged and rotten by
Rhizoctonia, Pythium and Fusarium (Bald et al. 1971).
J Phytopathol 162 (2014) 466–471 � 2014 Blackwell Verlag GmbH466
J Phytopathol

In Israel, onion is produced in open fields on area of
ca 1200 ha all over the country, from the Golan
Heights in the north to Arava valley in the south.
Rotting and wilting of onion bulbs and plants are
common. Wilting of onion plants with rot of the
basal plate of the bulb has been related mostly to
F. oxysporum f. sp. cepae (Lacy and Roberts 1982).
However, F. proliferatum was recently reported as the
predominant fungal species isolated from root and
bulbs of onion and garlic plants (Stankovic et al.
2007).
The objectives of the current study were to identify
and characterize the causal agents of bulb and root rot
in lily plants grown under greenhouse conditions and
onions grown in open fields in Israel.
Materials and Methods
Isolation and identification of pathogens from diseased
lily and onion plants
Random samples of diseased lily plants including
bulbs were collected from 51 greenhouse locations
and examined for the presence of pathogens; each
sample consisted of 5–10 plants. Stems, roots and
bulbs were washed, surface-sterilized with 0.3%
sodium hypochlorite for 10 min and dried in a lami-
nar flow cabinet. Segments (5 mm long) from roots,
bulb basal plate, scale leaf and stems were sampled
using a scalpel and plated on potato dextrose agar
(PDA; Difco Laboratories, Detroit, MI, USA) medium
supplemented with 100 ppm streptomycin sulphate,
on a minimal medium containing 2 g/l sorbose,
18 g/l agar and 100 ppm streptomycin sulphate, and
cornmeal agar (CMA; Difco Laboratories). After
incubation of the Petri dishes for 4–5 days at 27°C in
the dark, the fungi were identified microscopically.
In addition, diseased onion plants including their
bulbs were sampled from two locations (open field)
and examined for the presence of pathogens as
described previously for lily; each sample consisted of
25–40 plants.
All fungal isolates (Table 1) were classified to the
genus level by means of taxonomic keys including
macroscopic characteristics of the colony, as well as
microscopic structures. Binucleate Rhizoctonia was
identified by the Identification Service Central bureau
voor Schimmel cultures, Utrecht, Netherlands. Fusari-
um spp. isolates were molecularly identified using
the translation elongation factor EF-la, nuclear ribo-
somal internal transcribed spacer (ITS), b-tubulin and
H3 histone genes. Portions of the genes were PCR-
amplified and sequenced.
Isolation and purification of fungal DNA
Mycelia from 25 ml of potato dextrose broth cultures
were collected by vacuum filtration and lyophilized
until dry. DNA was extracted and purified as previ-
ously described (Freeman et al. 1993). The purified
DNA was dissolved in 0.5 ml of Tris–EDTA buffer
(10 mM Tris–HCl, 1 mM EDTA at pH 8.0) to a concen-
tration of 200–500 lg/ml and diluted to a concentra-
tion of 100 ng/ll for PCR.
PCR amplification
Translation elongation factor (EF-1a) gene was ampli-
fied using the primers EF 1a (5′ATGGGTAAGGA(AG)GACAAGAC 3′) and EF 2a (5′GGA(AG)GTACCAGT(GC)ATCATGTT 3′) as previously described (O’Don-
nell et al.1998).
Universal PCR primers ITS5 (5′ GGAAGTAAAAGTCGTAACAAGG 3′) and ITS4 (5′ TCCTCCGCTTATTGATATGC 3′) were used for the amplification of ITS1
and ITS2 regions between the small and large nuclear
rDNA, including the 5.8S rDNA, as described by
White et al. (1990). The b-tubulin gene was amplified
using the primers T1 (5′ AACATGCGTGAGATTG
TAAGT 3′) and T22 (5′ TCTGGATGTTGTTGGGAATCC3′) (O’Donnell and Cigelnik 1997). The H3 histone
gene was amplified using the primers H3-1a (5′ACTAAGCAGACCGCCCGCAGG 3′) and H3-1b (5′ GCGGGCGAGCTGGATGTCCTT 3′) as previously described
(Steenkamp et al. 2000).
Nucleotide sequence accession numbers
DNA sequences determined in this study have been
deposited in GenBank under accession numbers
KF222553 to KF222576.
Table 1 Collection sites and plant hosts for the fungi isolates used in
this study
Fungi species
Isolate
number
Isolate
colour Host Site
Fusarium oxysporum I White Onion Beit She’an
Fusarium oxysporum II Purple Lilly Be’erotayim
Fusarium oxysporum III Dark purple Onion Beit She’an
Fusarium proliferatum W White Onion Beit She’an
Fusarium proliferatum W White Lilly SdeYitzhak
Fusarium proliferatum P Purple Lilly Ahituv
Rhizoctonia solani Lilly Ein Habesor
Binucleate
Rhizoctonia AG-A
Lilly Kfar ha-Ro’e
Pythium oligandrum Lilly Kfar ha-Ro’e
J Phytopathol 162 (2014) 466–471 � 2014 Blackwell Verlag GmbH 467
S. Lebiush-Mordechai et al. Bulb and root rot in lily and onion in Israel

Pathogenicity tests on lily
Two pathogenicity tests (experiments A and B) to
complete Koch’s postulates were conducted under
controlled conditions in a greenhouse (25 � 3°C).Many disease-free lily bulbs, variety ‘Osnat’, obtained
from a commercial farm, were used for the experi-
ments after testing a sample of 10 bulbs of 150. The
bulbs were planted in 1.5-L pots containing per liter,
and 1 month later, inoculation with the tested
pathogens (Table 1) was carried out by adding coni-
dia (for Fusarium) or mycelium (for Rhizoctonia and
Pythium) at a concentration of 104–106 cfu/ml for
each fungus. The experimental design was a com-
plete randomized block with nine replicates for each
pathogen. Height and foliar disease symptoms of the
plants were monitored during 75 days. At the end of
the experiment, disease symptoms that developed on
bulbs and roots were assessed visually on an ordinal
scale from 0 to 10 (0 = healthy; 10 = completely rot-
ten), and pathogens were re-isolated. In an addi-
tional pathogenicity test (experiment C), using the
same experimental design, pathogen-free lily bulbs
from tissue culture were used. Inoculation was
conducted by placing 0.5-cm plugs of freshly grown
5-day-old mycelial PDA cultures on the wounded
bulb base, 16 bulbs for each pathogen. The bulbs
were incubated in a moist chamber (25 � 1°C, RH
95%) for 3 weeks. The experimental design was a
randomized block with four replications. Experi-
ments were conducted twice.
Pathogenicity tests on onion
Pathogenicity tests on onion seedlings were carried
out, in eight replicates, with the following fungi:
isolate of Fusarium proliferatum (white), Pythium
oligandrum, three isolates of F. oxysporum (I-III) and
binucleate Rhizoctonia AG-A (Table 1). Twelve seeds
of onion (Cv. ‘Ada 781’) were sown in a soil mix of
70% peat and 30% perlite (Shacham Givat Ada,
Israel) containing a slow release fertilizer 0.06 g/l
Osmocote (Scotts Miracle-Gro, Marysville, OH, USA)
in pots (360 ml). For the Fusarium isolates, 4 ml of
conidial suspension was added to each pot (4 9 105
conidia/pot), whereas for P. oligandrum, PDA plugs
of mycelium from 3-day-old colonies (1.8 9 103 per
pot) were used, covered by a thin layer of soil mix.
The experiment was conducted in a temperature-
controlled greenhouse set at 25°C �2°C. Forty days
later, the number of seedlings per pot was counted.
Surface-sterilized plant material was then placed on
PDA to assess fungal re-isolation. Experiments were
conducted twice.
Statistical analysis
Disease symptoms were analysed by analysis of vari-
ance (ANOVA). Means were compared with Student’s
multiple-range test at a significance level of P < 0.05.
Percentages were arcsine-transformed before analysis.
Results
Isolation and identification of pathogens from diseased
lily and onion plants
Symptoms on lily plants collected from commercial
greenhouses included chlorosis of leaves, wilting,
brown lesions on stem bases, bulbs and roots followed
by rotting of the roots and bulbs. Several fungi were
isolated from diseased plants (Table 2). In general,
the most prevalent fungi were F. proliferatum p and
Rhizoctonia spp (98% and 78% of samples,
Table 2 Prevalence of isolated fungi from lily plants sampled from commercial greenhouses in Israel (2001–2005)
Sampling
Region
Number of
samples
Incidence of infected bulbs (%)
Rhizoctonia solani
and Rhizoctonia
binucleate AG-Aa
Pythium
oligandruma
Fusarium
oxysporumb
Fusarium
proliferatum wb
Fusarium
proliferatum pbRhizoglyphus
robinic
North 30 97 37 60 25 93 70
Centre 16 56 7 20 56 100 38
South 5 80 40 20 20 100 80
78 28 33 34 98 63
aRhizoctonia spp. and P. oligandrm were isolated from root and basal plate.bFusarium spp. were isolated from root and basal plate and also from scale leaf, except F. proliferatum white, which was isolated from a basal plate
and scale leaf only.cPresence of Rhizoglyphus robini was visually evaluated.
J Phytopathol 162 (2014) 466–471 � 2014 Blackwell Verlag GmbH468
Bulb and root rot in lily and onion in Israel S. Lebiush-Mordechai et al.
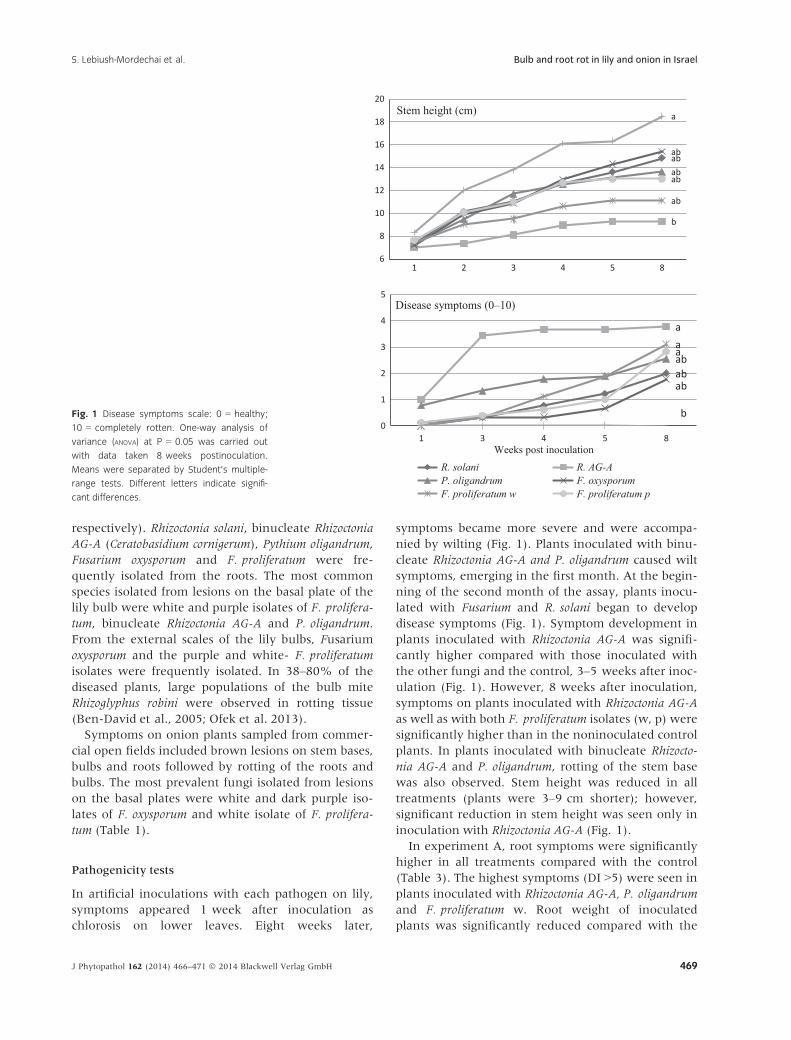
respectively). Rhizoctonia solani, binucleate Rhizoctonia
AG-A (Ceratobasidium cornigerum), Pythium oligandrum,
Fusarium oxysporum and F. proliferatum were fre-
quently isolated from the roots. The most common
species isolated from lesions on the basal plate of the
lily bulb were white and purple isolates of F. prolifera-
tum, binucleate Rhizoctonia AG-A and P. oligandrum.
From the external scales of the lily bulbs, Fusarium
oxysporum and the purple and white- F. proliferatum
isolates were frequently isolated. In 38–80% of the
diseased plants, large populations of the bulb mite
Rhizoglyphus robini were observed in rotting tissue
(Ben-David et al., 2005; Ofek et al. 2013).
Symptoms on onion plants sampled from commer-
cial open fields included brown lesions on stem bases,
bulbs and roots followed by rotting of the roots and
bulbs. The most prevalent fungi isolated from lesions
on the basal plates were white and dark purple iso-
lates of F. oxysporum and white isolate of F. prolifera-
tum (Table 1).
Pathogenicity tests
In artificial inoculations with each pathogen on lily,
symptoms appeared 1 week after inoculation as
chlorosis on lower leaves. Eight weeks later,
symptoms became more severe and were accompa-
nied by wilting (Fig. 1). Plants inoculated with binu-
cleate Rhizoctonia AG-A and P. oligandrum caused wilt
symptoms, emerging in the first month. At the begin-
ning of the second month of the assay, plants inocu-
lated with Fusarium and R. solani began to develop
disease symptoms (Fig. 1). Symptom development in
plants inoculated with Rhizoctonia AG-A was signifi-
cantly higher compared with those inoculated with
the other fungi and the control, 3–5 weeks after inoc-
ulation (Fig. 1). However, 8 weeks after inoculation,
symptoms on plants inoculated with Rhizoctonia AG-A
as well as with both F. proliferatum isolates (w, p) were
significantly higher than in the noninoculated control
plants. In plants inoculated with binucleate Rhizocto-
nia AG-A and P. oligandrum, rotting of the stem base
was also observed. Stem height was reduced in all
treatments (plants were 3–9 cm shorter); however,
significant reduction in stem height was seen only in
inoculation with Rhizoctonia AG-A (Fig. 1).
In experiment A, root symptoms were significantly
higher in all treatments compared with the control
(Table 3). The highest symptoms (DI >5) were seen in
plants inoculated with Rhizoctonia AG-A, P. oligandrum
and F. proliferatum w. Root weight of inoculated
plants was significantly reduced compared with the
ab
b
ab
ab
ab
ab
a
6
8
10
12
14
16
18
20
1 2 3 4 5 8
Stem height (cm)
ab
a
ab
ab
aa
b0
1
2
3
4
5
1 3 4 5 8
Disease symptoms (0–10)
R. solani R. AG-AP. oligandrum F. oxysporumF. proliferatum w F. proliferatum p
Weeks post inoculation
Fig. 1 Disease symptoms scale: 0 = healthy;
10 = completely rotten. One-way analysis of
variance (ANOVA) at P = 0.05 was carried out
with data taken 8 weeks postinoculation.
Means were separated by Student’s multiple-
range tests. Different letters indicate signifi-
cant differences.
J Phytopathol 162 (2014) 466–471 � 2014 Blackwell Verlag GmbH 469
S. Lebiush-Mordechai et al. Bulb and root rot in lily and onion in Israel
control, except for those inoculated with F. prolifera-
tum p. Bulb symptoms were significantly higher in
plants inoculated with Rhizoctonia AG-A, P. oligandrum
and both F. proliferatum isolates as compared to the
control. The highest symptoms were observed in inoc-
ulations with Rhizoctonia AG-A and F. proliferatum w
(Table 3). Stem symptoms in plants inoculated with
all fungi, except R. solani, were significantly higher
than in the control, with the highest levels in F. prolif-
eratum w.
In experiment B, the overall disease symptoms
were lower than in experiment A. Root symptoms
were the highest in plants inoculated with P. oligan-
drum and F. proliferatum w, bulb symptoms were the
highest in F. oxysporum inoculation, whereas stem
symptoms were the highest in P. oligandrum and
F. proliferatum p inoculations (Table 3).
In the pathogenicity test on tissue-cultured patho-
gen-free lily bulbs, symptoms of rotting first appeared
at the base of the bulb scales, spreading later to the
whole scales 1 week after inoculation (Table 4). All
inoculated bulbs were significantly different from the
noninoculated control, regardless of the inoculated
pathogen. The levels of rotten tissue in bulbs inocu-
lated with Rhizoctonia AG-A and P. oligandrum were
higher than those inoculated with R. solani (Table 4).
In artificial inoculations of onion, seedling survival
was significantly affected by all fungi (Table 5). The
most pathogenic fungus was F. proliferatum w,
whereas the least pathogenic fungi were F. oxysporum
II and III.
Discussion
Leaf chlorosis, wilting and root and bulb rot of lily
plants grown in commercial greenhouses in Israel are
predominantly caused by binucleate Rhizoctonia AG-A,
P. oligandrum and F. proliferatum. Pathogenicity tests
completing Koch’s postulates on lily and onion were
successfully demonstrated. All fungi were re-isolated
from inoculated plants. Based on the data collected
from these trials, it can be concluded that binucleate
Rhizoctonia AG-A, P. oligandrum and F. proliferatum
were the most pathogenic in lily (Lilium longiflorum)
and the white strain of F. proliferatum in onion seed-
ling. Fusarium proliferatum affecting Lilium crops was
recently reported in Spain (Prados-Ligero et al. 2008).
Table 3 Effect of artificial inoculation with fungal pathogens on disease
and dry root weight of lily, 8 weeks postinoculation
Disease symptoms (0–10)Root Weight
(mg)Root Bulb Stem
Experiment A
Rhizoctonia solani 3.9 ab 2.7 bc 2.0 bc 642.5 b
Rhizoctonia AG-A 5.9 a 6.9 a 3.6 ab 523.8 bc
Pythium oligandrum 5.8 a 4.0 b 4.4 ab 358.6 c
Fusarium oxysporum p 4.2 ab 2.7 bc 3.3 ab 633.8 b
Fusarium proliferatum w 5.2 ab 5.0 ab 5.0 a 511.3 bc
Fusarium proliferatum p 3.4 ab 3.9 b 3.2 ab 680.6 ab
Control 0.3 c 0.6 c 0.6 c 890.0 a
Experiment B
Rhizoctonia solani 1.8 bc 0.6 ab 1.3 c 699.2 ab
Rhizoctonia AG-A 2.3 bc 0.6 ab 1.2 bc 605.8 b
Pythium oligandrum 3.0 ab 0.9 ab 3.1 a 465.8 b
Fusarium oxysporum p 1.7 c 1.2 a 2.2 abc 907.5 a
Fusarium proliferatum w 4.2 a 0.6 ab 1.3 c 684.2 ab
Fusarium proliferatum p 1.8 bc 1.1 ab 2.6 ab 676.7 ab
Control 1.2 c 0.4 b 0.4 c 912.5 a
Disease symptoms scale: 0 = healthy; 10 = completely rotten. One-way
analysis of variance (ANOVA) was carried out at P = 0.05. Means were
separated by Student’s multiple-range tests. Different letters within a
column indicate significant differences.
Table 4 Effect of artificial inoculation on tissue-cultured pathogen-free
lily bulbs with fungal pathogens on disease
Experiment C
Disease symptoms (0–10)
Bulb Scale leaf
Rhizoctonia solani 5.8 b 2.3 b
Rhizoctonia AG-A 8.1 a 3.9 ab
Pythium oligandrum 8.4 a 4.9 a
Fusarium oxysporum p 7.3 ab 2.9 b
Fusarium proliferatum w 7.4 ab 5.1 a
Fusarium proliferatum p 7.3 ab 3.9 ab
Control 0.5 c 0.3 c
Disease symptoms scale: 0 = healthy; 10 = completely rotten. One-way
analysis of variance (ANOVA) was carried out at P = 0.05. Means were
separated by Student’s multiple-range tests. Different letters within a
column indicate significant differences.
Table 5 Onion seedling survival (%), 40 days postinoculation with fungal
pathogens
Experiment A Experiment B
Pythium oligandrum 52.3 b 66.7 b
Binucleate Rhizoctonia AG-A 59.1 b nd
Fusarium oxysporum w I 65.2 b 57.3 b
Fusarium oxysporum p II nd 89.6 a
Fusarium oxysporum dp III nd 83.3 a
Fusarium proliferatum w 48.0 b 9.4 c
Control 91.8 a 87.5 a
Percentages were arcsine-transformed before analysis. One-way analy-
sis of variance (ANOVA) was carried out at P = 0.05. Means were sepa-
rated by Student’s multiple-range tests. Different letters within a column
indicate significant differences.
J Phytopathol 162 (2014) 466–471 � 2014 Blackwell Verlag GmbH470
Bulb and root rot in lily and onion in Israel S. Lebiush-Mordechai et al.

Rhizoctonia solani had less effect than the other fungi,
as previously reported by Smith and Maginnes
(1996), who observed that although R. solani was
common on upper parts of the scales, it appeared to
cause minimal damage.
In conclusion, lily bulb rots are most serious as they
can result in total crop loss. The F. proliferatum purple
isolates were the most common species isolated from
lesions, mainly from the basal plate, but were not as
virulent compared to Rhizoctonia AG-A, P. oligandrum
and the white isolates of F. proliferatum.
Acknowledgements
The research was supported by the Chief Scientist of
The Ministry of Agriculture and Rural Development,
Israel. This paper is a contribution of the Agricultural
Research Organization, Institute of Plant Protection,
Bet Dagan, Israel.
References
Bald JG, Suzuki T, Doyle A. (1971) Pathogenicity of Fusari-
um oxysporum to Easter lily, narcissus and gladiolus. Ann
Appl Biol 67:331–342.
Ben-David T, Tsror L, Palevsky E. (2005) Developing an
action threshold for the bulb mite, Rhizoglyphus robini on
lily, onion and garlic. IOBC/wprs Bull 28:11–14.
Freeman S, Pham M, Rodriguez RJ. (1993) Molecular
genotyping of Colletotrichum species based on arbitrarily
primed PCR, A+T-rich DNA, and nuclear DNA analyses.
Exp Mycol 17:309–322.
Lacy ML, Roberts DL. (1982) Yields of onion cultivars in
midwestern organic soils infested with Fusarium oxyspo-
rum f. sp. cepae and Pyrenochaeta terrestris. Plant Dis
66:1003–1006.
Lawson RH, Hus HT. (1996) Lily diseases and their control.
Acta Hort 414:175–187.
L€offler HJM, Straathof TP, Mouris JR, Baayen RP.
(1995) Durability of resistance in lily to basal rot:
evaluation of virulence and aggressiveness among
isolates of Fusarium oxysporum f. sp.lilii. Eur J Plant
Pathol 102:261–271.
O’Donnell K, Cigelnik E. (1997) Two divergent intrage-
nomic rDNA ITS2 types within a monophyletic lineage
of the fungus Fusarium are nonorthologous. Mol Phylo-
gen Evol 7:103–106.
O’Donnell K, Kistler HC, Cigelnik E, Ploetz RC. (1998)
Multiple evolutionary origins of the fungus causing
Panama disease of banana: concordant evidence from
nuclear and mitochondrial gene genealogies. Proc Natl
Acad Sci USA 95:2044–2049.
Ofek T, Gal S, Inbar M, Lebiush-Mordechai S, Tsror L,
Palevsky E. (2013) The role of onion-associated
fungi in bulb mite infestation and damage to onion
seedlings. Exp Appl Acarol 61. doi: 10.1007/
s10493-013-9750-2.
Prados-Ligero AM, Basallote-Ureba MJ, Melero-Vara JM.
(2008) First report of Fusarium oxysporum f. sp. lilii and
F. proliferatum affecting Lilium crops in Spain. Trop Plant
Pathol 33:235–236.
Smith JD, Maginnes EA. (1996) Scale rot test for hybrid
lilies. Can Plant Dis Surv 46:92–94.
Stankovic S, Levic J, Petrovic T, Logrieco A, Moretti A.
(2007) Pathogenicity and mycotoxin production by
Fusarium proliferatum isolated from onion and garlic in
Serbia. Eur J Plant Pathol 118:165–172.
Steenkamp E, Britz H, Coutinho T, Wingfield B, Marasas
W, Wingfield M. (2000) Molecular characterization of
Fusarium subglutinans associated with mango malforma-
tion. Mol Plant Pathol 1:187–193.
White TJ, Bruns T, Lee S, Taylor J. (1990) Amplification
and direct sequencing of fungal ribosomal RNA genes
for phylogenetics. In: Innis MA, Gelfand DH, Sninsky
JJ (eds) PCR Protocols, A Guide to Methods and
Applications. San Diego, CA, Academic Press,
pp 315–322.
J Phytopathol 162 (2014) 466–471 � 2014 Blackwell Verlag GmbH 471
S. Lebiush-Mordechai et al. Bulb and root rot in lily and onion in Israel





























