Embed Size (px)
Citation preview

CAROHYDRATE METABOLISM
PART 2
BY
PROF.DR. SOUAD ABOAZMA

OXIDATION OF GLUCOSE
The pathways for oxidation of glucose are classified into two main
groups:
a- The major pathways for complete oxidation of glucose
into CO2, H2O and energy are:
1- Glycolysis → convert one molecule of glucose into 2 mol of
pyruvic acid + 2 NADH.H+.
2- Oxidative decarboxylation of pyruvic to acetyl CoA +
NADH.H++CO2
3- Complete oxidation of acetyl CoA in Kerb’s cycle into CO2, H2O
and energy .
b- The minor pathways for oxidation, which are not for energy
production.
1- Hexose monophosphate pathway (HMP).
2- Uronic acid pathway.

GLYCOLYSIS
EMBDEN-MEYERHOF PATHWAY
acid in presence of pyruvicoxidation of glucose to give Def.:O2 and lactic acid in absence of mitochondria (RBCs) and in
absence of O2 .
Site: Cytoplasm of all cells especially muscles and RBCs.
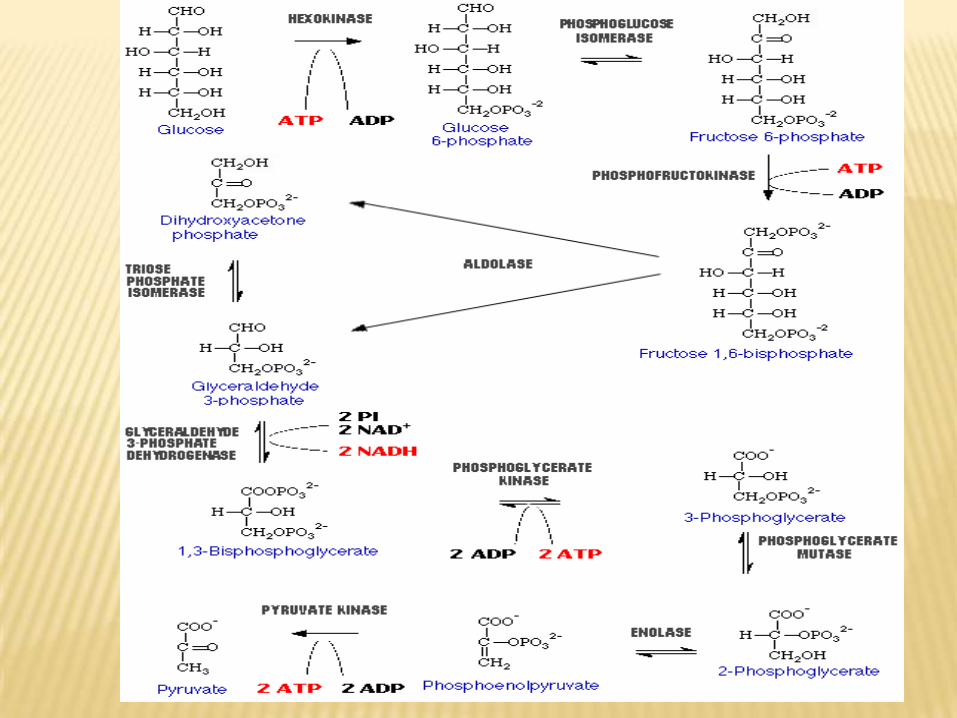
Steps:H – C = O
H C – OH
OH – C – H
H – C – OH
H – C – OH
CH2OH
H – C = O
H – C – OH
OH – C – H
H – C – OH
H – C – OH
CH2O-P
Mg
ATP ADP
Hexokinase, glucokinase
G-6-PD-Glucose
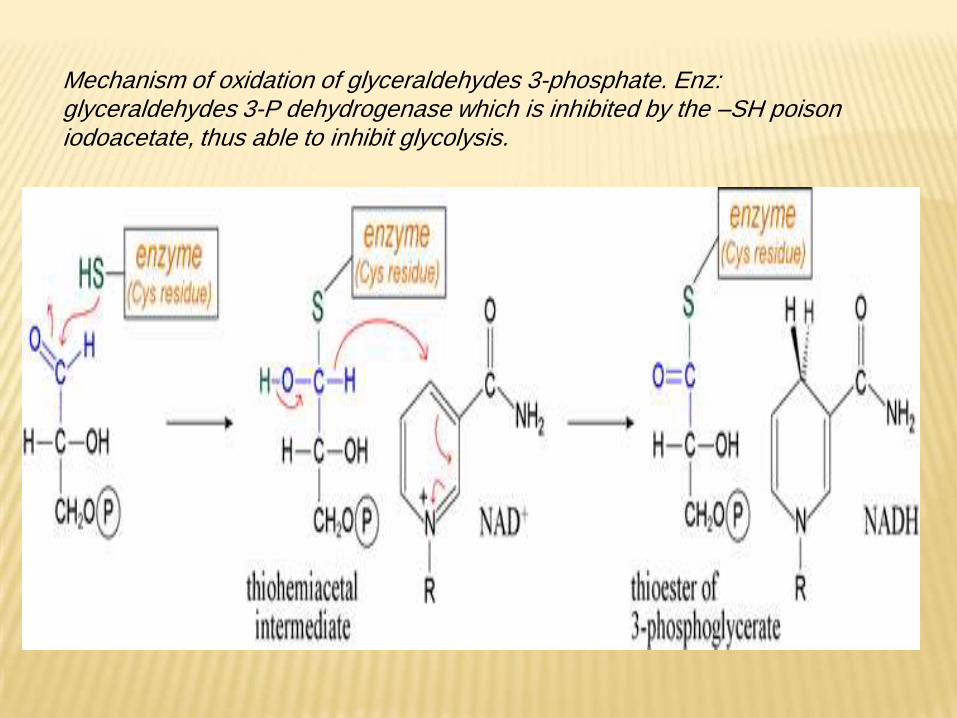
Mechanism of oxidation of glyceraldehydes 3-phosphate. Enz: glyceraldehydes 3-P dehydrogenase which is inhibited by the –SH poison iodoacetate, thus able to inhibit glycolysis.

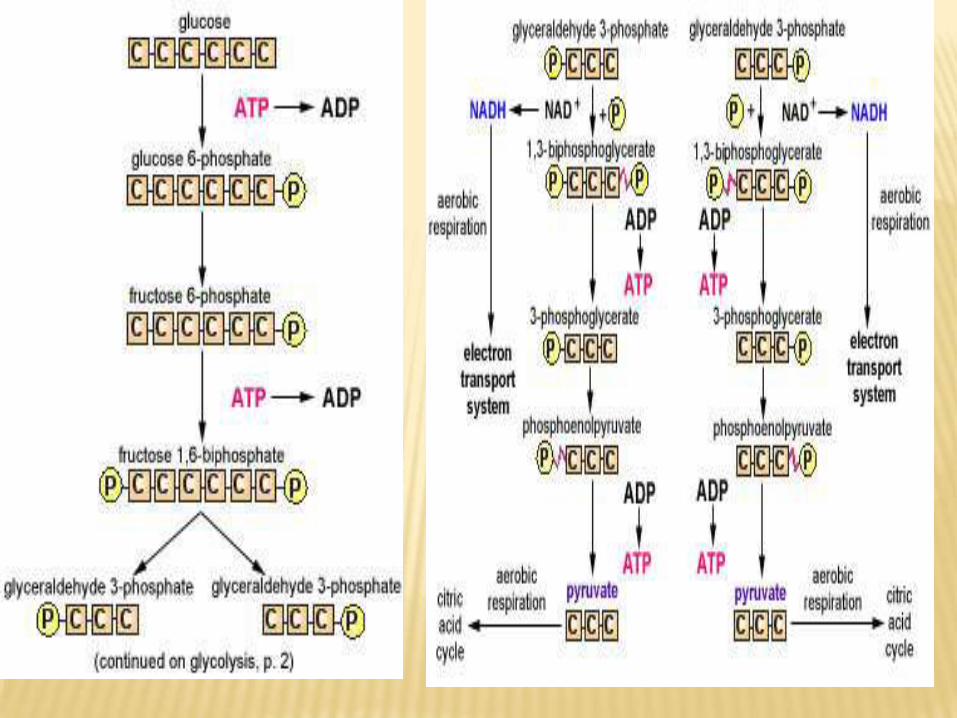
:GLYCOLYSISENERGY PRODUCTION FROM
A. glycolysis in presence of O2 (Aerobic glycolysis):
Reaction catalyzed by ATP production
Stage I
1. Hexokinase/Glucokinase reaction (for
phosphorylation)
-1 ATP
2. Phosphofrutokinase-1 (for phosphorylation) -1 ATP
Stage III
3. Glyceraldehyde-3-P dehydrogenase (oxidation of
2 NADH in electron transport chain)
+ 6 or +4 ATP
4. Phosphoglycerate kinase (substrate level
phosphorylation)
+2 ATP
Stage IV
5. Pyruvate kinase (substrate level phosphorlyation) +2 ATP
Net gain = 10 or 8 - 2 =
8 or 6ATP

):glycolysis(Anaerobic 2in Absence of OGlycolysisB.
•In absence of O2 re-oxidation of NADH at glyceraldehyde-3-P-
dehydrogenase stage cannot take place in electron-transport
chain.
But the cells have limited coenzyme. Hence to continue the
glycolytic pathway NADH must be oxidized to NAD+. This is
achieved by reoxidation of NADH by conversion of pyruvate
to lactate (without producing ATP) by the enzyme lactate
dehydrogenase. Occurs in cells with no mitochondria as RBCs
(mature) ,or under low O2 supply as intensive muscular exercise.
In anaerobic glycolysis per molecule of glucose oxidation 4 - 2 = 2 ATP
will be produced.

REGULATION OF GLYCOLYSIS
There are 3 types of mechanisms responsible for regulation of
the enzyme activity which are:
1- Changed in rate of enzyme synthesis that affect the
quantity of enzyme as:
* Induction →↑ rate of enzyme synthesis at gene expression
→↑ mRNA synthesis →increase enzyme concentration.
* Repression →↓ rate of enzyme synthesis at gene expression
→↓ mRNA synthesis →decrease enzyme concentration.
2- Covalent modification by reversible phosphorylation
dephosphorylation.
3- Allosteric regulation by allosteric activator or inhibitor that
affect the quality of the enzymes.
A- Allosteric regulation of glycolysis:

irreversible reaction in 3 regulatory enzymes which responsible for 4 There are
.glycolysis
Hexokinase1.It is found in most tissues to give G-6-P when blood glucose level is low.
2.Acts on glucose and other hexoses to give hexose-6-P.
3.It has low km and Vmax→ acts maximally at fasting bl. glucose level.
4.It is inhibited by its products, which is G-6-P → allosteric feedback inhibition.
Glucokinase1.It is found in liver and acts maximally after meal.
2.Acts only on glucose.
3.It has a high km and high Vmax → so it is active when bl. glucose level is high (after meal).
4.It is induced (↑its rate of synthesis) by insulin.
5.It is not inhibited by G-6-P.
Phosphofructokinase1.It is the major regulatory enzyme in most tissues.
2.It is allosterically activated by F-6-P, AMP and inhibited by ATP, citrate, and H+.
Pyruvate kinase1.It is allosterically inhibited by ATP, fatty acids, alanine, and acetyl CoA. And activated by
F-1-6 diphosphate.
2. It is phosphorylated by cAMP dependent protein kinase, which becomes inactive
and dephosphorylated by phosphatase enzyme, which becomes active.

•Hormonal regulation:-B
Insulin/glucagons ratio is the main hormonal regulation of glucose
utilization; it increases during glucose feeding and decreases during
fasting.
A.Glucagons: it is secreted in case of carbohydrates deficiency or in
response to low blood glucose level (hypoglycemia). It affects liver
cells mainly as follows:
1.It acts as repressor of glycolytic key enzymes except hexokinase.
2.Through cAMP-dependent protein kinase A, it produces
phosphorylation of specific protein enzymes that lead to
inactivation of glycolytic key enzymes( only for pyruvate kinase).
B.Insulin: it is secreted after feeding of carbohydrate or in response to
high blood glucose level (hyperglycemia). It stimulates all pathways of
glucose utilization. Insulin binds to a specific cell membrane receptors
and produces certain signal cascade, which results in the following:
1.It acts as inducer for glycolytic key enzymes.
2.It activats phosphodiesterase enzyme(decreases cAMP that
leads to inhibition of protein kinase A).
3.It activats protein phosphatase-1 that produces
dephosphorylation of glycolytic key enzymes and their activation.

INHIBITORS OF GLYCOLYSIS:
1- Aresnate : which used in oxidative step
insted of Pi→ so glycolysis proceeds in
presence of arsenate but ATP, which formed
from 1-3 diphosphoglycerate is lost.
2- Iodoacetate produces inhibition of
glyceraldehydes-3-P dehydrogenase (inhibitor
of SH group).
3- Flouride inhibits enolase →↓↓ glycolysis in
bacteria →no production of lactic acid produced
by bacteria, which cause dental caries. It used
as anticoagulant in blood sample used for
estimation of blood glucose →↓↓ glycolysis in
RBCs .
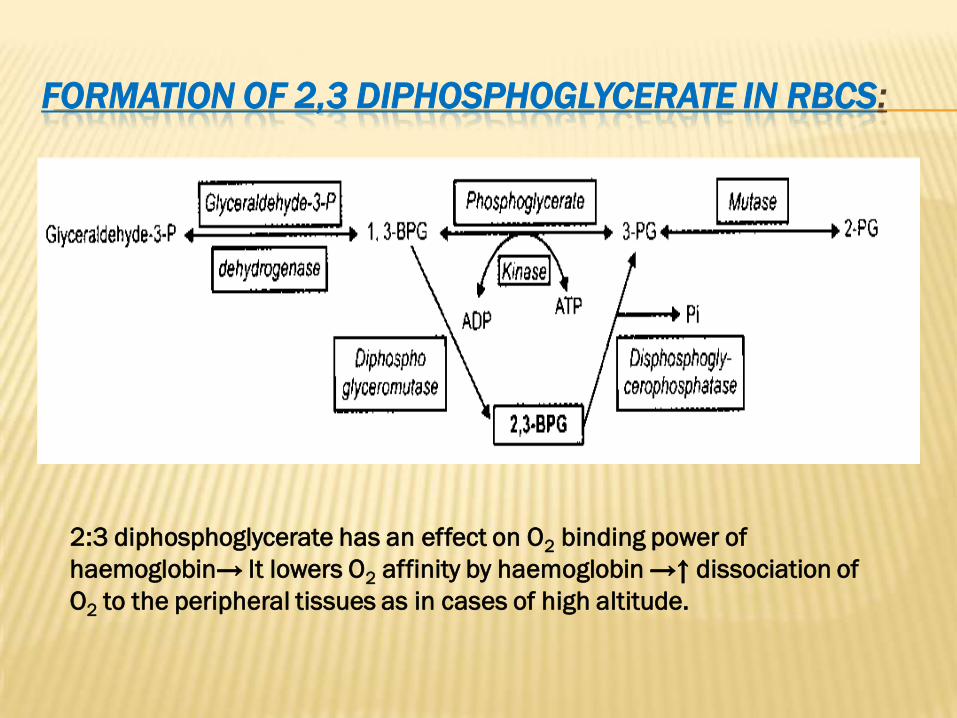
:IN RBCSDIPHOSPHOGLYCERATE3 ,2FORMATION OF
2:3 diphosphoglycerate has an effect on O2 binding power of
haemoglobin→ It lowers O2 affinity by haemoglobin →↑ dissociation of
O2 to the peripheral tissues as in cases of high altitude.

CLINICAL SIGNIFICANCE OF 2,3 DIPHSOPHOGLYCERATE:
1- Persons who live at high altitude undergo state of low
O2 affinity for HB due to simultaneous increase of 2,3
diphosphoglycerate. This increase can be reversed on
returning to sea level.
2- Fetal HB has less 2,3 diphosphoglycerate than adult
HB, so fetal HB has high O2 affinity.
3- During storage of blood in blood banks, there is
decrease in 2,3 diphosphoglycerate so, stored blood has
high O2 affinity, which is not suitable for blood transfusion
especially to ill patients. If 2,3 diphosphoglycerate is
added to stored blood, it can’t penetrate RBCs wall. So, it
is advisable to add insoine, which is a substance that can
penetrate RBCs wall and change it into 2,3
diphosphoglycerate through HMP shunt.
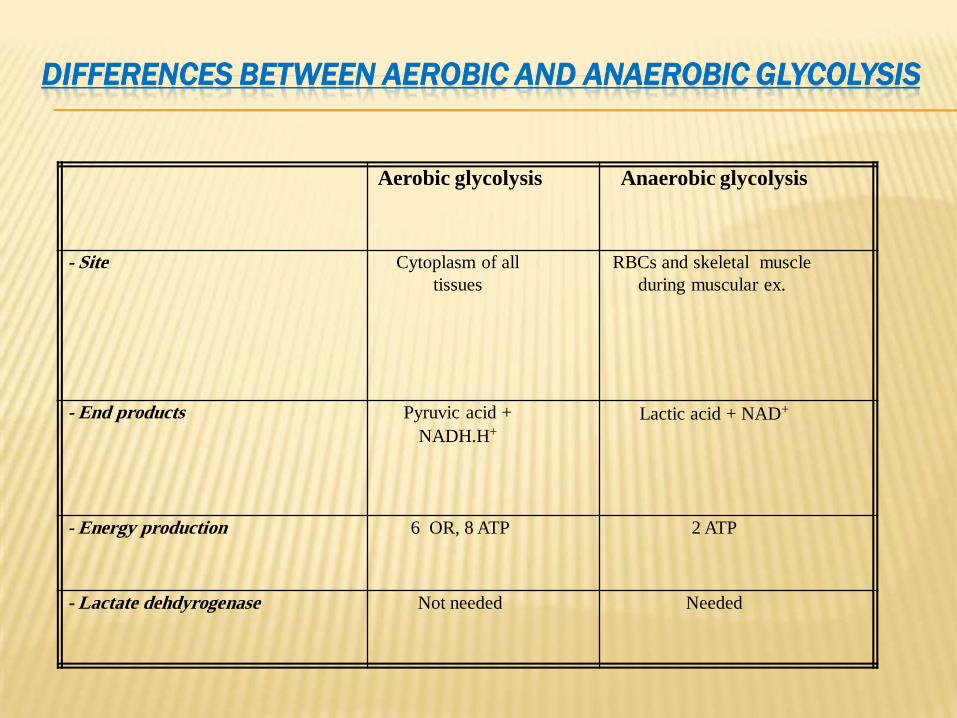
GLYCOLYSISDIFFERENCES BETWEEN AEROBIC AND ANAEROBIC
Aerobic glycolysis Anaerobic glycolysis
- Site Cytoplasm of all
tissues
RBCs and skeletal muscle
during muscular ex.
- End products Pyruvic acid +
NADH.H+
Lactic acid + NAD+
- Energy production 6 OR, 8 ATP 2 ATP
- Lactate dehdyrogenase Not needed Needed

GLYCOLYSISDISEASES ASSOCIATED WITH IMPAIRED
:deficiency Hexokinase-1•In patients with inherited defects of hexokinase activity, the red blood cells contain low
concentrations of the glycolytic intermediates including the precursor of 2,3-DPG.
•In consequence, the hemoglobin of these patients has an abnormally high oxygen affinity.
•The oxygen saturation curves of red blood cells from a patient with hexokinase deficiency
are shifted to the left, which indicates that oxygen is less available for the tissues.
2- Pyruvate kinase deficiency (hemolytic anemia):•All red blood cells are completely dependent upon glycolytic activity for ATP production.
•Failure of the pyruvate kinase reaction, the production of ATP will decrease leading to
hemolysis of red cells.
•Inadequate production of ATP reduces the activity of the Na+ - and K+ -stimulated ATPase
ion pump.
3- Lactic acidosis:-•Blood levels of lactic acid are normally less than 1.2 mM. In lactic acidosis, the values for
blood lactate may be 5 mM or more.
•The high concentration of lactate results in lowered blood pH and bicarbonate levels.
•High blood lactate levels can result from increased formation or decreased utilization of
lactate.
•Common cause of hyperlacticidemia is anoxia.
•Tissue anoxia may occur in shock and other conditions that impair blood flow, in respiratory
disorders, and in severe anemia.

AEROBIC AND ANAEROBIC EXERCISE USE DIFFERENT FUELS
distance running, while -Aerobic exercise is exemplified by longanaerobic exercise by sprinting or weight lifting.
organ -there is really very little interanaerobic exercise During
cooperation. The vessels within the muscles are compressed during peak
contraction, thus their cells are isolated from the rest of the body. Muscle
largely relies on its own stored glycogen and phosphocreatine.
energy phosphate for ATP -serves as a source of highPhosphocreatineare glycolysisand glycogenolysisuntil seconds 5 -4for first synthesis
becomes the primary source of ATP for want of Glycolysis. stimulatedoxygen.
is metabolically more interesting. For moderate Aerobic exercise . of muscle glycogen.glycolysisenergy is derived from thetexercise, much of
However, a well-fed individual doesn't store enough glucose and glycogen to
provide the energy needed for running long distances. The respiratory
quotient, the ratio of carbon dioxide exhaled to oxygen consumed, falls
progressive switch from glycogen during distance running. This indicates the
gradually increases as glucose Lipolysisto fatty acid oxidation during a race. stores are exhausted, and, as in the fast state, muscles oxidize fatty acids in
. preference to glucose as the former become available

Marathon runner (42.2 KM or 26 Miles)
•In the marathon runner predominantly red fibres (oxidative)
are used.
•Red fibres contain myoglobin and mitochondria.
•The major sources of energy in marathon runner are:-
•Aerobic metabolism is the principal source of ATP.
•Blood glucose.
•Hepatic glycogen is degraded to maintain the level of blood
glucose.
•Muscle glycogen is also a source of fuel but it is degraded
slowly than in sprinter .
•Free fatty acids derived from the triacylglycerol of adipose
tissue. Sources of fuel used by marathon runner and there duration of action
Source of fuel Duration of action by minutes
Blood glucose
Liver glycogen
Muscle glycogen
Triacylglycerol
4
18
70
4000

MAJOR FEATURES OF SKELETAL MUSCLE S METABOLISM
1.Skeletal muscle functions under both aerobic (resting) and anaerobic (eg, sprinting)
conditions, so both aerobic and anaerobic glycolysis operate, depending on conditions.
2.Skeletal muscle contains myoglobin as a reservoir of oxygen.
3.Insulin acts on skeletal muscle to increase uptake of glucose.
4.In the fed state, most glucose is used to synthesize glycogen, which acts as a store of glucose for
use in exercise, 'preloading' with glucose is used by some long-distance athletes to build up stores
of glycogen.
5.Epinephrine stimulates glycogenolysis in skeletal muscle, whereas glucagon does not because of
absence of its receptors.
6.Skeletal muscle cannot contribute directly to blood glucose because it does not contain glucose-6-
phosphatase.
7.Lactate produced by anaerobic metabolism in skeletal muscle passes to liver, which uses it to
synthesize glucose, which can then return to muscle, (the cori cycle).
8.Skeletal muscle contains phosphocreatine, which acts as an energy store for short-term (seconds)
demands.
9.Free fatty acids in plasma are a major source of energy, particularly under marathon conditions
and in prolonged starvation.
10.Skeletal muscle can utilize ketone bodies during starvation.
11.Skeletal muscle is the principle site of metabolism of branched chain amino acids, which are
used as energy source.
12.Proteolysis of muscle during starvation supplies amino acids for gluconeogenesis.
13.Major amino acids emanating from muscle are alanine (destined mainly for gluconeogenesis in
liver and forming part of the glucose-alanine cycle) and glutamine (destined mainly for the gut and
kidneys).

Significance of glycolysis•Glycolysis is the principle route for glucose
metabolism for the production of ATP molecules
•It also provide pathway for the metabolism of
fructose and galactose derived from diet.
•It represent the only source of energy for RBCs and
contracting muscle.
•It provide mitochondria with pyruvic which give
acetyle CoA (Kreb's Cycle)
•Glycolysis give DHAP which reduced to α
glycerophosphate which form backbone of
triacylglycerol in lipogenesis
•In erythrocytes glycolysis supplies 2,3 DPG which
is required for haemoglobin function in transport of
oxygen.

ACIDPYRUVICOF DECARBOXYLATIONOXIDATIVE
keto-αacid and other pyruvicIt is conversion of :Def.acids into CoA derivatives, CO2 & NADH.H+.
RBCsIn mitochondrial matrix of all tissues except : SiteCoAacid into acetyl pyruvicThe conversion of : Steps
is catalyzed by pyruvate dehydrogenase complex, which
composed of 3 enzymes act cooperative with each other
in presence of 5 co-enzymes: TPP, lipoic acid, FAD,
NAD+, and CoASH.
Pyruvate +TTP + Lipoic acid + CoA +FAD+ NAD+ --→ CO2 + Acetyl-CoA + NADH + H+
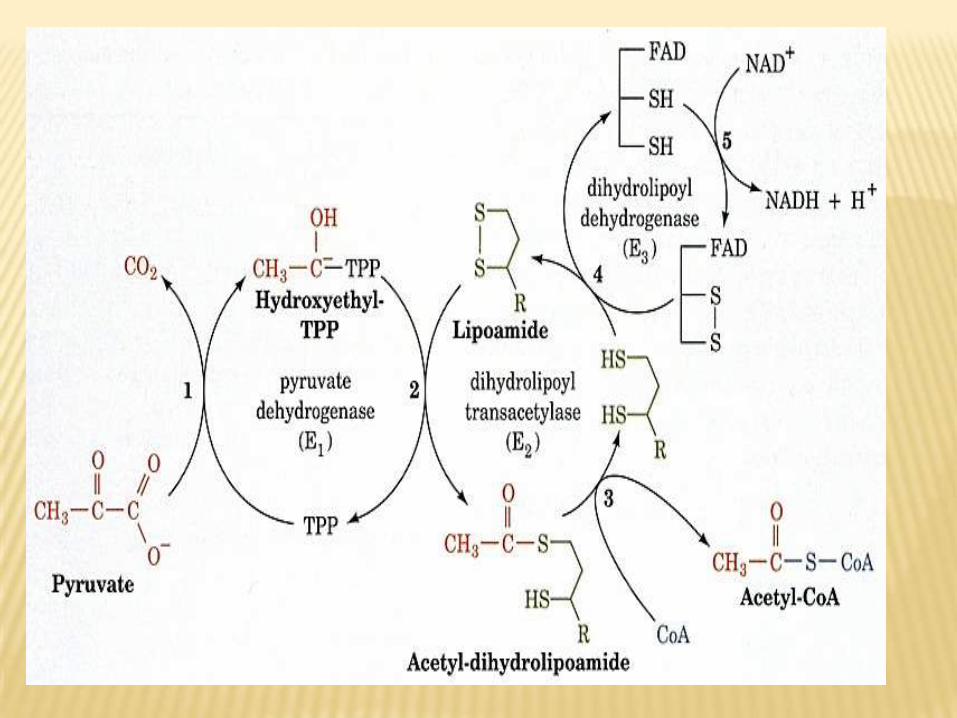

acid:pyruvicof decarboxylationSteps of oxidative •Pyruvate is decarboxylated to form a hydroxyethyl
derivative bound to the reactive carbon of thiamine
pyrophosphate, the coenzyme of pyruvate decarboxylase.•The hydroxyethyl intermediate is oxidized by transfer to the
disulfide form of lipoic acid covalently bound to
dithydrolipoyl transacetylase.•The acetyl group, bound as a thioester to the side chain of
lipoic acid, is transferred to CoA.
•The sulfhydryl form of lipoic acid is oxidized by FAD-dependent dihydrolipoyl dehydrogenase, leading to the
regeneration of oxidized lipoic acid.
•Reduced flavoprotein is reoxidized to FAD by dihydrolipoyl
dehydrogenase and NAD+.

OF DECARBOXYLATIONREGULATION OF OXIDATIVE
ACID :PYRUVIC
1- Product inhibition : The enzyme complex is inhibited by acetyl CoA, which
accumulates when it is produced faster than it can be oxidized by
citric acid cycle. The enzyme is also inhibited by elevated levels of
NADH+.H, which occure when the electron transport chain is
overloaded with substrate and oxygen is limited.
2- Covalent modification:The pyruvate dahydrogenase complex exists in two forms: an active
nonphosphorylated form and an inactive phosphorylated form.Phosphorylated
and nonphosphorylated pyruvate dehydrogenase can be interconverted by
two separate enzymes, a kinase and a phosphatase. The kinase is activated
by increase in the ratio of acetylCoA/ CoA or NADH/ NAD+. An increase in the
ratio of ADP/ATP, which signals increased demand for energy production ,
inhibits the kinase and allows the phosphatase to produce more of the active
,nonphosphorylated enzyme.
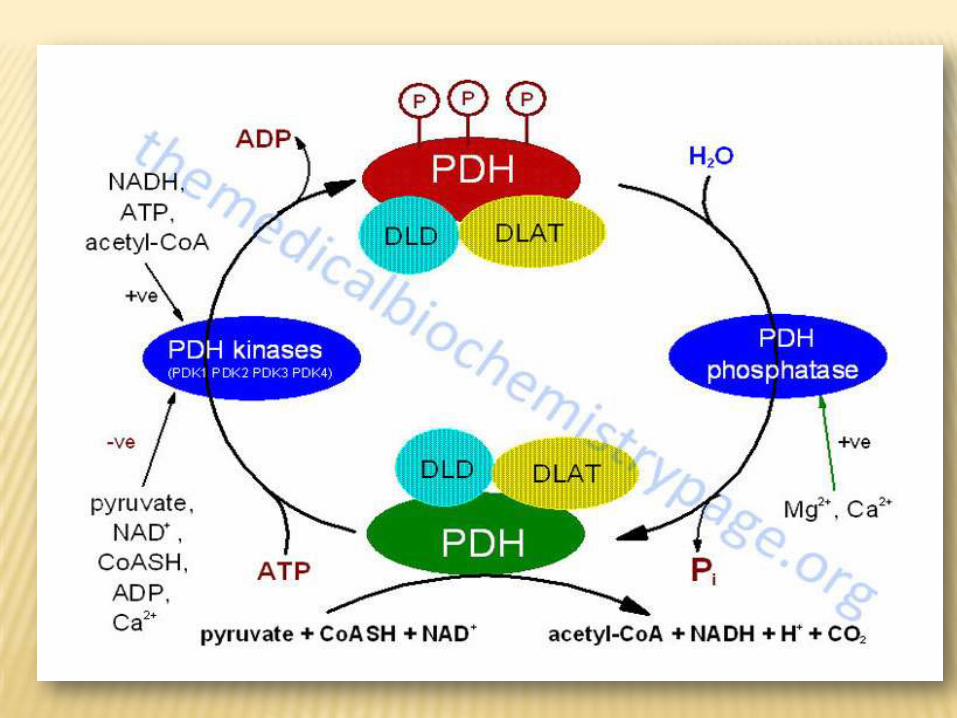

:METABOLISMPYRUVATECLINICAL ASPECTS OF
metabolism leads to lactic pyruvateInhibition of :acidosis, which may be due to
1- Arsenite or mercuric ions complex the –SH
group of lipoic acid.
2--Dietary deficiency of thiamin as in alcoholics.
These two factors lead to inhibition of pyruvate
dehydrogenase complex.
3- Inherited pyruvate dehydrogenase deficiency,
which may be due to defects in one or more of
the components of the enzyme complex.

CITRIC ACID CYCLE
CYCLE) KREB’SACID CYCLE (TRICARBOXYLIC
Def.:
It is the series of reactions in mitochondria, which
oxidized acetyl CoA to CO2, H2O & reduced H2 carriers
(as NADH.H+ & FADH2) that oxidized through
respiratory chains for ATP synthesis.
Site:
Mitochondria of all tissue cells except RBCs, which
not contain mitochondria. The enzymes of the cycle
are present in mitochondrial matrix except succinate
dehydrogenase, which is tightly bound to inner
mitochondrial membrane.
Steps:
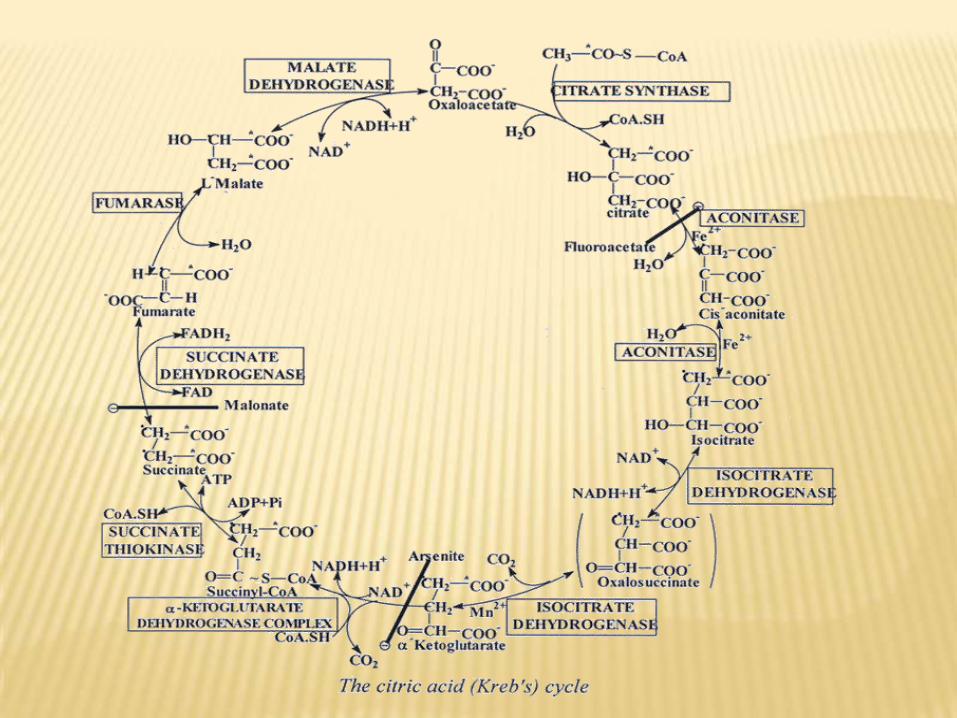
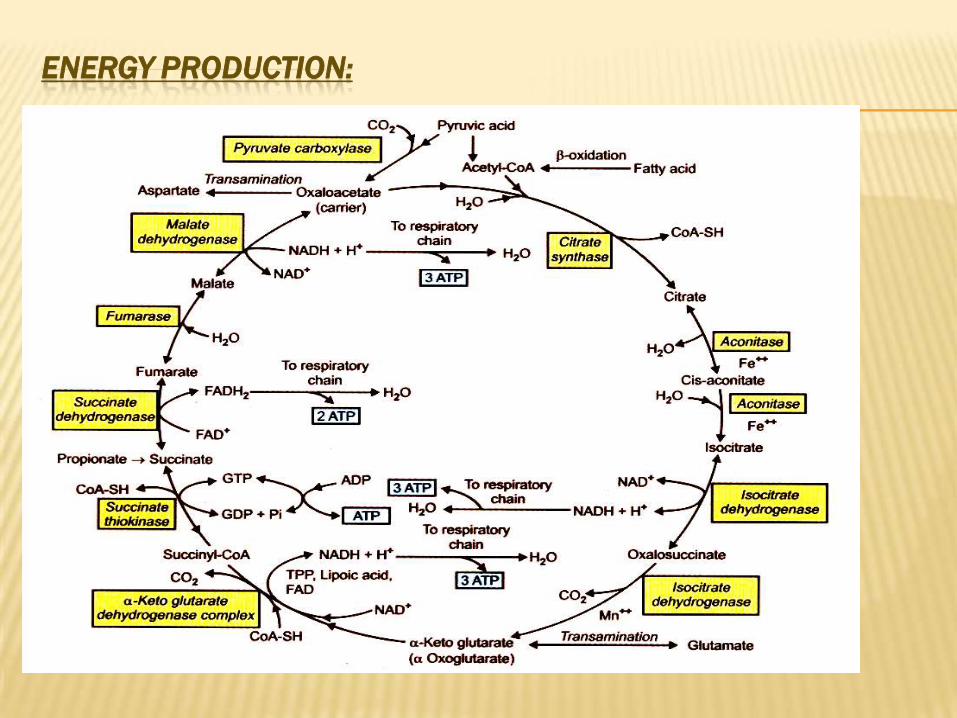
ENERGY PRODUCTION:

:ENERGY PRODUCTION FROM OXIDATION OF ONE MOLECULE OF GLUCOSE

REGULATION OF KREB’S CYCLE:
1- As the primary function of TCA cycle is to provide energy, respiratory
control via the E.T.C and oxidative phosphorylation exerts the main control.
2- In addition to this overall and coarse control, several enzymes of TCA
cycle are also important in the regulation.
Three key enzymes are:
(a)Citrate synthase. (b)Mitochondrial isocitrate dehydrogenase.
(c)α-ketoglutarate dehydrogenase.
These enzymes are responsive to the energy status as expressed by the [ATP]/[ADP] ratio and [NADH]/[NAD+] ratio.
(a)Citrate synthase enzymes is allosterically inhibited by ATP and long-chain
acyl CoA.
(b)NAD+-dependent mitochondrial iso-citrate dehydrogenase (ICD) is activated
allosterically by ADP and is inhibited by ATP and NADH.
(c)α-ketoglutarate dehyrogenase complex which allosterically inhibited by succinyl
CoA, NADH-H+ and ATP.
3- In addition to above succinate dehydrogenase enzyme is inhibited by oxaloacetate
(OAA) and the avability of OAA is controlled by malate dehydrogenase, which
depends on [NADH]/[NAD+] ratio.

CYCLEKREB’SFUNCTIONS OF •lipids CHO,It is the final pathway for complete oxidation of all foodstuffs -1
and protein, which are converted to acetyl CoA.
2- It is the major source of energy for cells except cells without mitochondria
as RBCs.
3- It is the major source of succinyl CoA, which used for:
1.Porphyrine and HB synthesis.
2.Ketone bodies activation.
3.Converted to OAA → glucose.
4. Detoxication by conjugation
•4- Synthetic functions of Kreb’s cycle:
a- Amphibolic reactions.
Some components of Kreb’s cycle are used in synthesis of other
substances as:
In fasting state, oxaloacetic acid is used for synthesis of glucose
by gluconeogenesis.
In fed state, citric acid is used for synthesis of fatty acids.
Reactions of Kreb’s cycle are used for synthesis of amino acid
(transamination into non essential amino acids) eg:
-OAA + glutamic acid aspartic acid + α-ketoglutarate.
-Pyruvic acid + glutamic acid alanine + α-ketoglutarate.
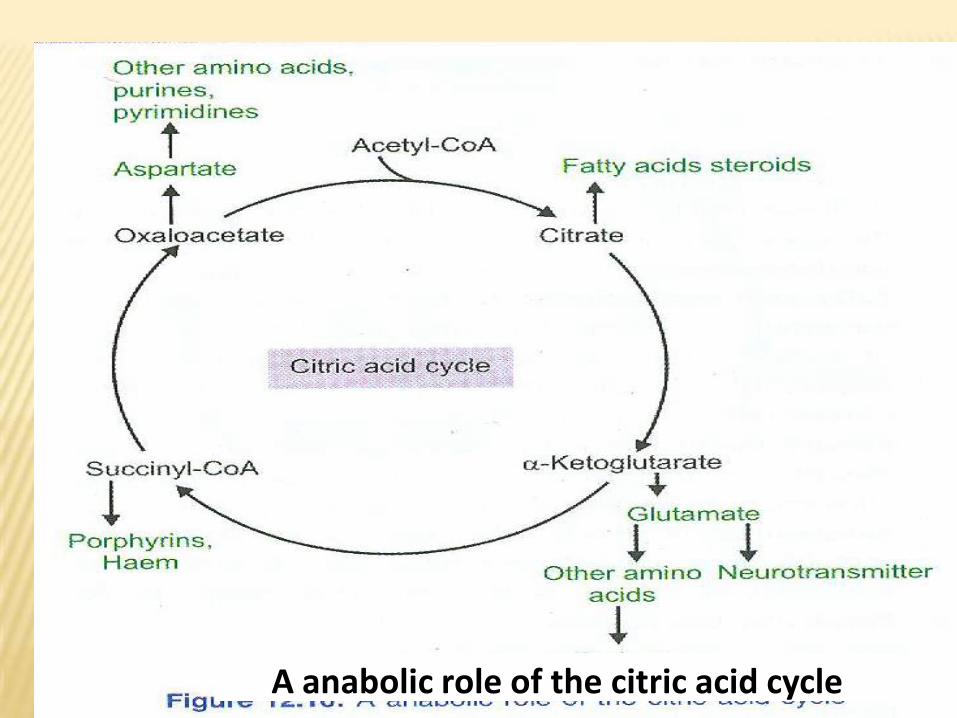
A anabolic role of the citric acid cycle

•b- Anaplerotic reactions.
Synthesis of one or more component of Kreb’s cycle from outside
the cycle:
O.A.A. can be synthesized from pyruvic acid by pyruvate
carboxylase, and from aspartic acid by transamination.
Fumarate can be synthesized from phenylalanine and tyrosine.
Succinyl CoA can be synthesized from valine, isoleucine,
methionine, and threonine.
α-ketogluterate can be synthesized from glutamic acid by
transamination.
Inhibitors of Citric Acid Cycle
1-Flouro-acetate reacts with oxalacetate forming
flourocitrate, which inhibits the aconitase enzyme.
2-Arsenite inhibits α-ketogluterate dehydrogenase.
3-Malonate acts as competitive inhibitor for succinate
dehydrogenase.
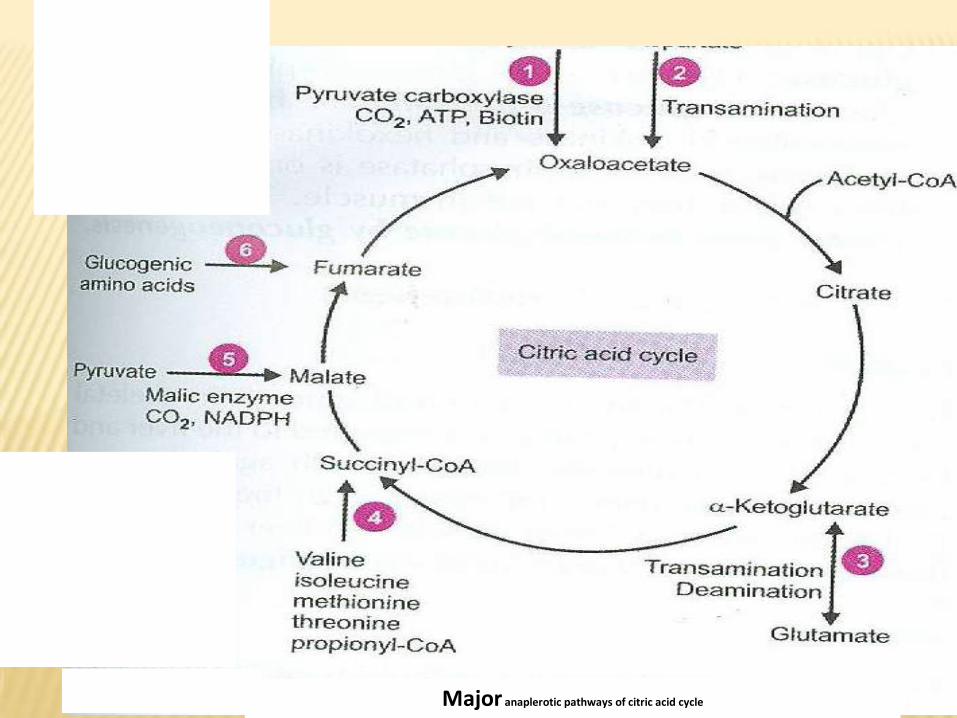
Major anaplerotic pathways of citric acid cycle

ROLES OF VITAMINS IN CITRIC ACID CYCLE
Four of the soluble vitamins of B complex have
important roles in cirtic acid cycle. They are:
-αin the form of FAD, a cofactor in ,riboflavin-1
ketogluterate dehydrogenase complex and in
in the form of ,niacin-2; dehydrogenasesuccinate
NAD, the coenzyme for three dehydrogenases in the
cycle, isocitrate dehydrogenase, α-ketogluterate
thiamin-3; dehydrogenasemalateand dehydrogenase
(vitamin B1), as TPP, the coenzyme for decarboxylation
in α-ketogluterate dehyrdogenase reaction; and
as part of coenzyme A, which , pantothenic acid-4
present in the form of acetyl-CoA and succinyl-CoA.
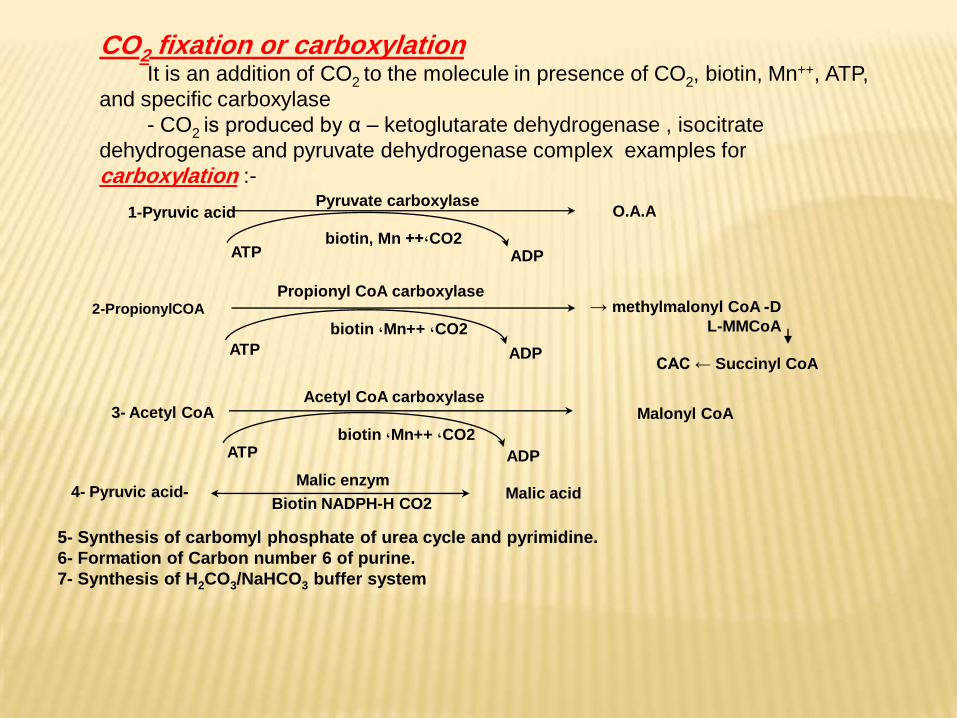
-4- Pyruvic acidMalic enzym
Biotin NADPH-H CO2Malic acid
1-Pyruvic acidPyruvate carboxylase
ATPCO2,++ biotin, Mn
ADP
O.A.A
3- Acetyl CoAAcetyl CoA carboxylase
ATPCO2, Mn++, biotin
ADP
Malonyl CoA
2-PropionylCOA
Propionyl CoA carboxylase
ATP
CO2, Mn++, biotin
ADP
D- methylmalonyl CoA → L-MMCoA
CAC ← Succinyl CoA
carboxylationfixation or 2COIt is an addition of CO2 to the molecule in presence of CO2, biotin, Mn++, ATP,
and specific carboxylase
- CO2 is produced by α – ketoglutarate dehydrogenase , isocitrate
dehydrogenase and pyruvate dehydrogenase complex examples for
carboxylation :-
5- Synthesis of carbomyl phosphate of urea cycle and pyrimidine.
6- Formation of Carbon number 6 of purine.
7- Synthesis of H2CO3/NaHCO3 buffer system

GLUCONEOGENESIS
It is the formation of glucose from non CHO sources. Its main function is to supply blood glucose in cases of carbohydrate deficiency (fasting, starvation, and low carbohydrate diet). Sites:Cytoplasm and mitochondria of liver and kidney due to presence of glucose-6-phophatase and fructose-1, 6-biphosphatase.

Steps:A.These mechanisms are concerned with conversion of glucogenic amino acids., lactate, glycerol, and propionic acid to glucose, which are reverse to glycolytic pathway (except for three irreversible kinases) to supply erythrocytes, skeletal muscles, nervous system, and mammary glands with their need of glucose.B.Energy barrier will obstruct reversal of glycolysis at the following sites:

1- Conversion of pyruvate to phosphoenol pyruvateThis can be overcomed by dicarboxylic acid shuttle, as the
following :-1.In mitochondria, pyruvic acid is converted to oxalacetatein presence of pyruvate carboxylase, ATP, biotin, Mn++ and CO2.2.Oxalacetate can not diffuse outside mitochondria →converted into compounds that can diffuse from mitochondria as malate, aspartate, and citrate → they are converted once again to oxalacetate in extramitochondriaportion of the cells.
-Oxalacetate reformed in extramitochondriacompartment is converted to phosphoenol pyruvatein presence of phosphoenol pyruvatecarboxykinase enzyme and GTP.
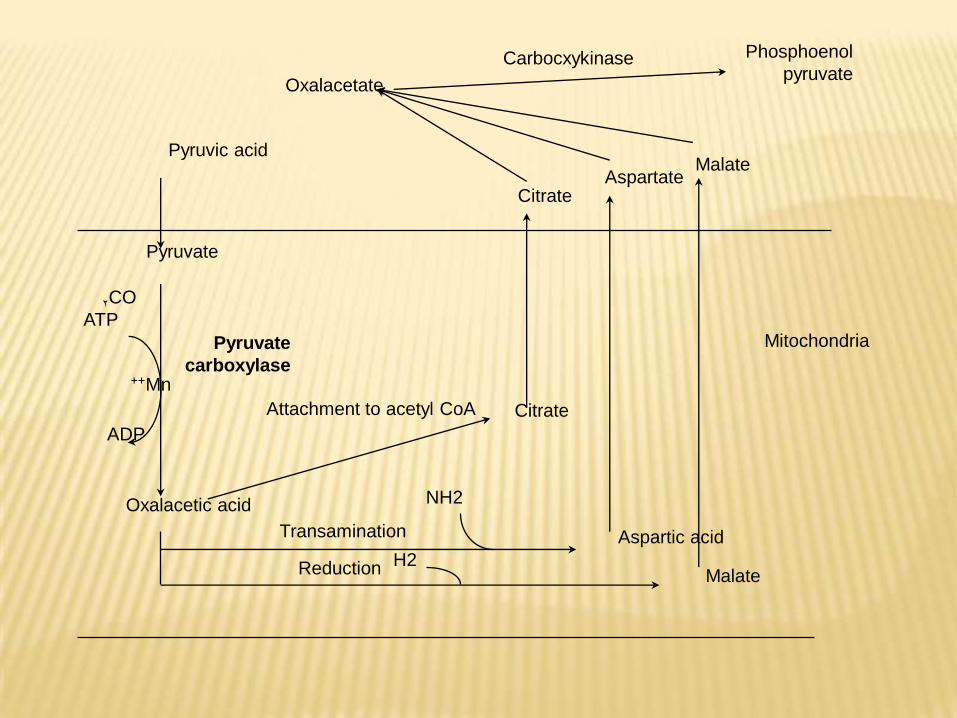
Citrate
Phosphoenol
pyruvateCarbocxykinase
Oxalacetate
MalateAspartate
Pyruvic acid
Pyruvate
Pyruvate
carboxylase
CO2
ATP
Mn++
ADP
Oxalacetic acid
Citrate
Aspartic acid
MalateReduction H2
NH2
Transamination
Attachment to acetyl CoA
Mitochondria

2- Conversion of fructose 1:6 biphosphate to F-6-P: -
This occurs by fructose 1:6 biphosphatase, which present in liver and kidneys.
3- Conversion of Glucose-6-P to glucose:This is catalysed by another enzyme, which is
G-6-Phosphatase that is present in liver, intestine, and kidney.
Carbon sources for glucoeogenesis:

1- Propionic acid:1.It is the product of odd number fatty acid oxidation by βoxidation.2.It is converted into succinyl CoA, which converted into oxalacetic acid → phosphoenol pyruvic →→Glucose.2- Glycerol:
glycerol-3-P converted into dihydroxy acetone-P, which can be converted by trio’s isomerase into glyceraldehydes-3-P→→→ glucose.
Glycerokinase is present in liver and kidney mainly3- Glucogenic amino acids:
Amino acids by deamination can be converted into α-keto acids as pyruvic, α-ketoglutaric and OAA → they can be converted into glucose. Proteins are considered as one of the main sources of blood glucose especially after 18 hours due to deplation of liver glycogen.

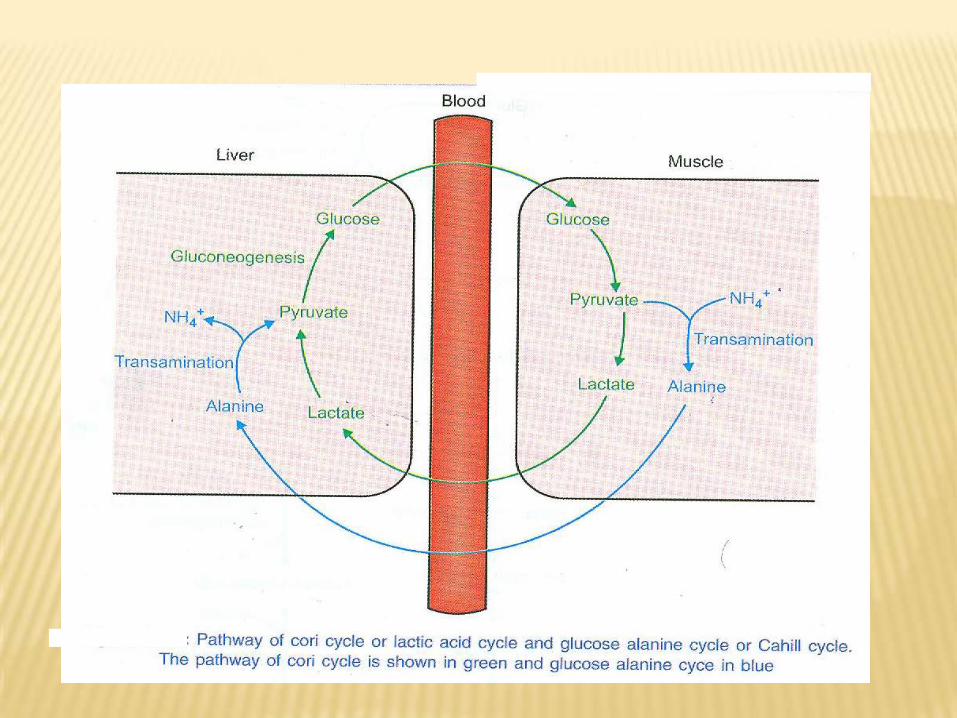
4- Lactic acid:In vigorous skeletal muscle activity, large amount of lactic acid produced → passes to the liver through blood stream → converted in liver into pyruvic acid and lastly to glucose → reach muscle once again through blood → this cycle called Cori cycle.Importance of Cori cycle:•It prevents loss of lactate as waste products in urine.•Oxidation of reduced NAD.•It supplies red cells and contracting muscles with glucose for reutilization and ATP production.•Prevent accumulation of lactic acid, which change pH of blood.

5- Glucose-alanine cycle:During starvation there is muscle protein catabolism →in presence of NH3 and pyruvic acid (produced from glycolysis), alanine is formed →reach liver and converted into pyruvic acid, which give glucose through glucogenesis and NH3 which converted into urea →excreted in urine.Significant of glucose alanine cycle:•Disposal of NH3 produced from muscle protein catabolism through formation of urea, which excreted in urine.•Conserve NAD/NADH.H+ ratio.

REGULATION OF GLUCONEOGENESISKey enzymes which regulate gluconeogeneis are: Pyruvate carboxylase, Phospho-enol-Pyruvate carboxykinase (PEPCK), Fructose-1, 6-bi-phosphatase and Glucose-6-phosphatase.
A -short time regulation:-1- Effect of increased fatty acid oxidation on gluconeogenesis : Fasting,
low carbohydrates in diet, stress and sever muscular exercise stimulate
secretion of anti-insulin hormones which increase lipolysis in adipose
tissues. The increased fatty acids oxidation acts as a signal to inhibit
glycolysis and stimulate gluconeogenesis in the liver by the following
mechanisms:
*It increases the production of ATP which produce allosteric inhibition of
phosphofructokinase-1 (PFK-1) and pyruvate kinase.
*The excess acetyl-CoA, produced by fatty acid oxidation allosterically
stimulate pyruvate carboxylase and inhibites pyruvate dehydrogenase thus
direct pyruvate to gluconeongesis.
*Excess citrate (produced from acetyl CoA) produces allosteric inhibition of
PFK-1 and PFK-2.
.

2- Fructose 2,6-bisphosphate plays a unique role in the
regulation of glycolysis and gluconeogenesis in liver
Fructose 2,6 bisphosphate is formed by phosphorylation of fructose
6-phosphate by PFK-2, the same enzyme is also responsible for its
breakdown, since it has fructose 2,6 bisphosphatase activity.
Fructose 2,6 bisphosphatase (bifunctional enzyme ) is under the
allosteric control of fructose 6-phosphate which stimulate kinase and
inhibit phosphatase.
Carbohdrate feeding and insulin stimulate PFK-2 and inhibite
fructose 2,6 bisphosphatase, producing accumulation of fructose 2,6
bisphosphate which produce allosteric activation of glycolytic key
enzyme PFK-1 and allosteric inhibition of gluconeogenesis key
enzyme fructose 1,6 bisphosphatase.
In fasting state, glucagons stimulate production of cAMP which
activate cAMP dependent protein kinase-A which in turn inactivates
PFK-2 and activate fructose 2,6 bisphosphatase. Hence
gluconeogenesis is stimulated by decrease in the concentration of
fructose 2,6 bisphosphate which inactivate PFK-1 and relieves the
inhibition of Fructose 1,6 bisphosphate.
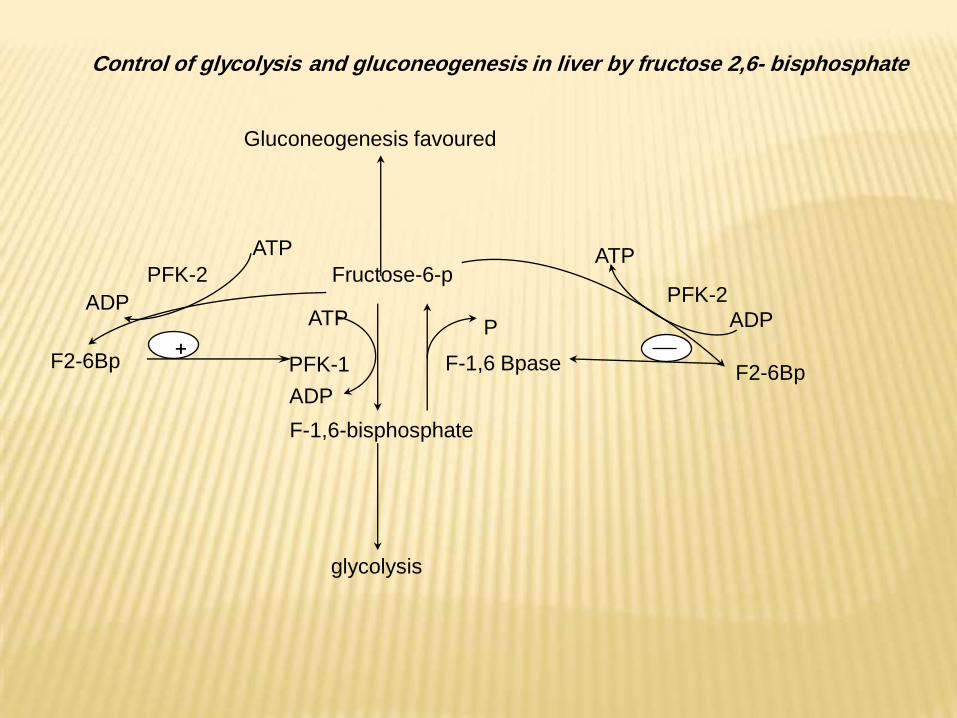
Fructose-6-p
F-1,6 BpasePFK-1F2-6Bp
ATP
ADP
ATP
ADP
PFK-2
+
F-1,6-bisphosphate
glycolysis
F2-6Bp
ATP
ADP
PFK-2
Gluconeogenesis favoured
P
Control of glycolysis and gluconeogenesis in liver by fructose 2,6- bisphosphate

-long time regulation:-B
Glucagon and insulin also have longterm effects on hepatic glycolysis and gluconeogenesis by induction and repression of key enzymes of both pathway. A high glucagon/insulin ratio in blood increases the capacity for gluconeogenesis and decreases that for glycolysis in liver. A low glucagon/insulin ratio has the opposite effects. Glucagon signals induction of greater quantities of PEP carboxykinase, Fructose 1,6 bisphosphatase and glucose 6-phosphatase.

Pentose phosphate pathway
Definition
•The penstose phosphate pathway is an alternative rout for
the metabolism of glucose, ATP neither produced nor utilized.
It is the pathway for formation of pentose phosphate.
•The pentose phosphate pathway is also described as a
shunt rather than pathway because it shunts (to move from
one trak to another) hexoses from glycolysis forming
pentoses, which may be cycled back into the pathway of
glycolysis by conversion into fructose 6-phosphate and
glycerldehde-3 phosphate and used for resynthesis of
glucose-6 phospahte so named hexose monophosphate
shunt is also phosphogluconate pathway.

• It is a multicyclic process in which three molecules of glucose 6-phosphate give rise to :
-Three molecules of Co2-Three molecules of pentoses-6 molecular of NADPH The pentoses are rearranged to generate two
molecules of glucose-6-phosphate and one molecule of glycerldehyde 3-phosphate .
Site: occurs in cytoplasm of many tissues including liver, adipose tissues, adrenal, ovary, testis, red cell and retina

The main functions of pentose shunt are:1-Production of NADPH-H+, which used in:Synthesis of fatty acids and cholesterol.Reduction of oxidized glutathione.Activation of folic acid.
2-Formation of pentoses in the form of ribose-5 P, which used in:Synthesis of nucleotides either free as ATP, or polynucleotides as DNA and RNA.Synthesis coenzymes as FAD, NAD+.

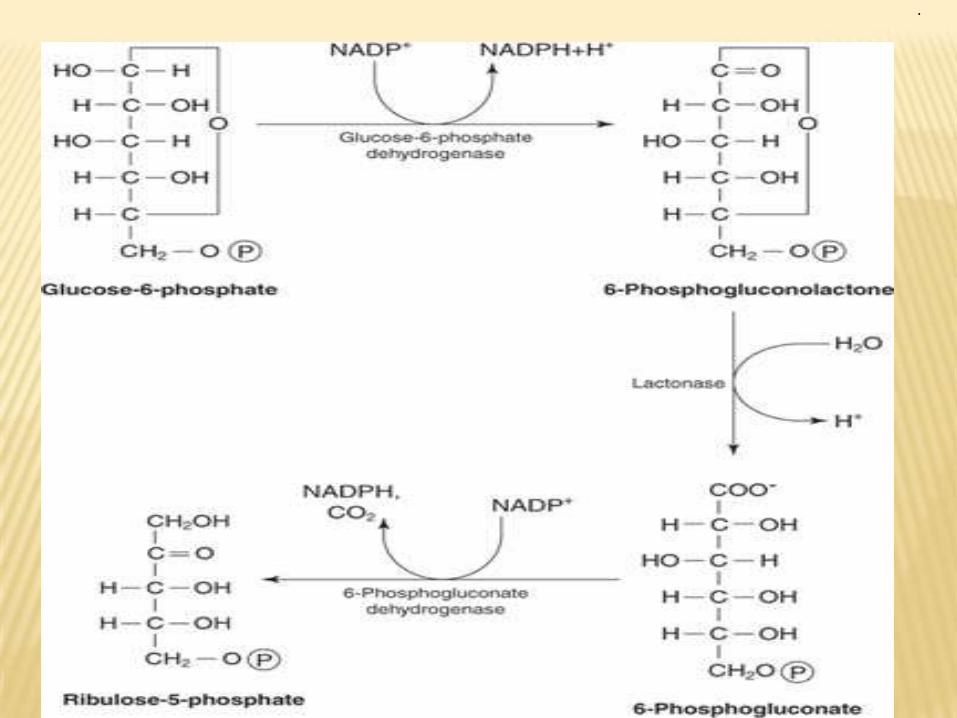
Steps: I- Oxidative irreversible reaction ( phase I) which include:•the oxidative portion of HMP consists of three reactions that lead to formation of ribulose-5-P, CO2 and two molecules of NADPH-H+ for each glucose-6 phosphate oxidized.2- Non oxidative revesible reaction which include:
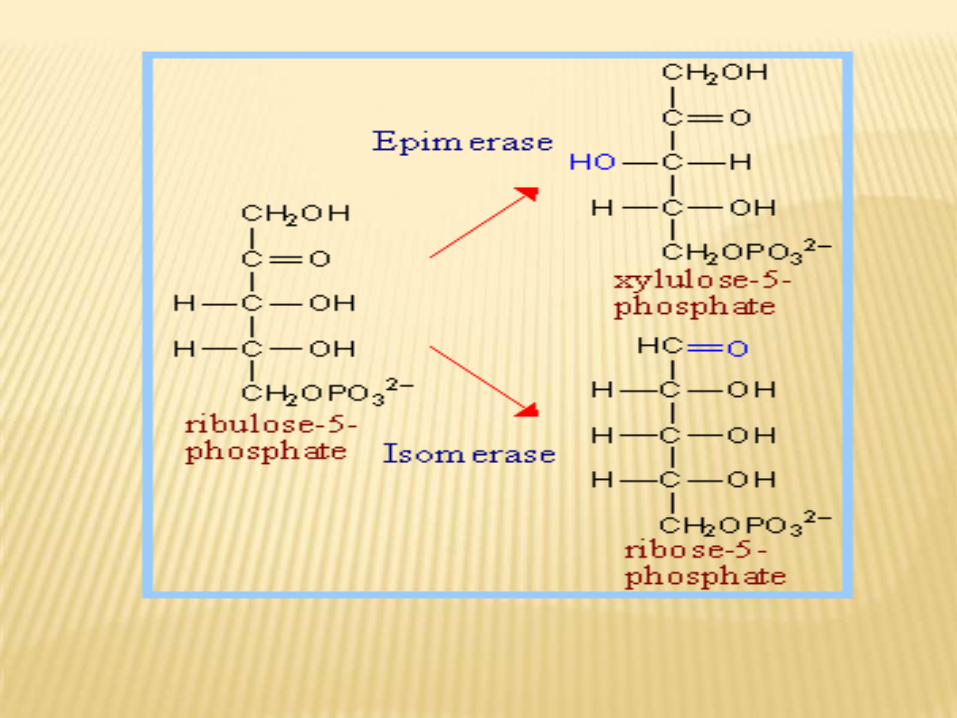
a- Phase II:Ribulose-5-P by ribulose-5-phospho epimerase is converted into xylulose-5-P while by ribulose-5 phosphe isomerase is converted into ribose-5-P.
b- phase III :-pentose-5-P is converted into glyceraldehydes-3-P and fructose-6-P by transketolase and transaldolase. These enzymes create a reversible link between pentose phosphate pathway and glycolysis by catalyzing these 3 reactions.
.
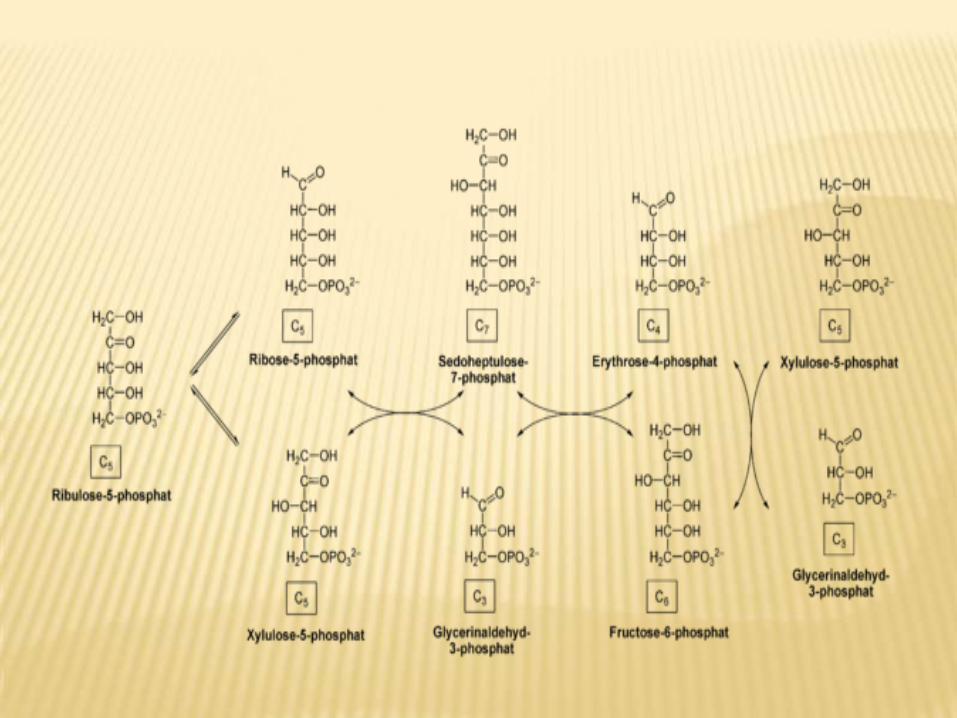

Function of non oxidative reversible phase : Formation of different pentoses 5-PIn skeletal muscles due to deficiency of G-6-pD, so muscle can synthesize ribose 5 phosphate from fructose-6-phosphate and glyceraldehydes 3-phosphate by pathway reversible to phase II.RBCs need more NADPH it than pentose-5 so convert three pentose-5-p into glycolytic intermediate.

REGULATION OF PENTOSE PHOSPHATE PATHWAY ( PPP):1- G-6-P dehydrogenase increased in amount when diet contains excess
carbohydrates.
2- The regulatory enzymes of PPP are g-6-P dehydrogenase and
lactonase enzymes.Insulin acts as inducer for their synthesis.
3- NADPH.H+ accumulation produces feedback inhibition of g-6-
dehydrogenase.
PPP is the major sources of NADPH.H+ but there are 2 other sources
for NADPH.H+ which present in cytoplasme:
1)cytoplasmic isocitrate dehydrogenase.
2)malic enzyme which convert malic acid into pyruvic acid ,CO2 and
NADPH.H+ .
4- Depending on the needs of the cell, conversion of pentose-5-P to
hexose-6-P can occure. The reaction favour the formation of
glyceraldehydes-3-P → glycerol-3-P which used in synthesis of
phosphoglycerides as triacylglycerol. Also glyceraldehydes-3-P can be
converted to 2,3 diphosphoglycerate in RBCs which ↓↓ O2 association
with HB→↑O2 dissociation to the tissues.

Role of NADPH in Red blood cell:•Red blood cells need NADPH to maintain glutathione in reduced form to protect themselves from oxidizing agents. •The red cell is exposed to large amount of molecular oxygen. Some of the molecular oxygen is converted to superoxide and H2O2 that can cause irreversible damage to the cell. •Reduced glutathione, a strong reducing agent protects against damage by reducing H2O2 to H2O.•The role of NADPH is to maintain glutathione in reduced form which is essentially required to preserve the integrity of RBCs. It also keeps iron of the haemoglobin in reduced ferrous state and prevents formation of methaemoglobin

Role of NADPH in disposal of H2O2

Deficiency of glucose 6-phosphate dehydrogenase.•Several types of X-linked inherited deficiency of the enzyme glucose-6-phosphate dehydrogenase have been recognized. •Enzyme deficient cells have a lower rate of NADPH production, resulting in a deficiency of reduced glutathione (GSH) which is essential to maintain the integrity of erythrocyte membrane and for keeping HB in the ferrous state. •Most individuals who have G-6-pD mutation are asymptomatic. Some individuals develop haemolytic anaemia if they are exposed to : •Certain oxidant drugs e.g. Antibiotic (sulphamethoxazole), antimalarial(primaquine), Antipyretic (acetoaminiphen).•Certain type of infection: The inflammatory response of infection generate free radicals which can damage the red cells easily if G6-PD is deficient. •Favism (due to ingestion of fava beans)•Favism is the haemolytic anaemia due to ingestion of fava beans (broad beans) in individuals with G-6-pD deficiency •Fava beans contain the purine glycosides these compound react with glutathione leading to reduced level of glutathione (GSH).•Favism is not observed in all individuals with G-6-PD deficiency but all patients with favism have G-6-PD deficiency.





























