Embed Size (px)
Citation preview

Molecular and Cellular Endocrinology, 75 (1991) 101-110 101 © 1991 Elsevier Scientific Publishers Ireland, Ltd. 0303-7207/91/$03.50
MOLCEL 02422
Character izat ion of m acaque 3f l -hydroxy-5-ene steroid dehydrogenase /AS-A 4 i somerase: structure and express ion in s teroidogenic and peripheral t issues
in pr imate
Jacques Simard 1, Michael H. Melner 2, Nathalie Breton 1, Kenneth G. Low 2, Hui-Fen Zhao 1, Laura M. Periman 2 and Fernand Labrie 1
I Medical Research Council Group in Molecular Endocrinology, CHUL Research Centre and Laval University, Quebec, Que. G1 V 4G2, Canada, and 2 Divisions of Reproductive Biology and Neuroscience, Oregon Regional Primate Research Center, Beaverton,
OR 97006, U.S.A.
(Received 13 September 1990; accepted 18 October 1990)
Key words: Steroidogenesis; Cloning; Gonads; Adrenal; 3fl-Hydroxysteroid dehydrogenase/aS-A a isomerase; Intracrine activity; (Rhesus monkey)
Summary
The conversion of 3fl-hydroxy-5-ene steroids by the enzyme complex 3fl-hydroxysteroid dehydro- genase/AS-A 4 isomerase (3fl-HSD) is an obligatory step in the biosynthesis of all classes of hormonal steroids in classical steroidogenic as well as in peripheral tissues. To develop a model more closely related to the human, we have isolated and characterized cDNA clones encoding macaque 3fl-HSD by screening a rhesus monkey ovary ~gt l l cDNA library using a human 3B-HSD cDNA probe. Nucleotide sequence of 1629 bp from overlapping cDNA clones predicts a protein of 372 amino acids with a calculated molecular mass of 41,874 (excluding the first Met). The deduced amino acid sequence of macaque 3fl-HSD displays 79.4% and 93.9% similarity with that of bovine and human 3fl-HSD, respectively. RNA blot analysis performed under high stringency conditions of macaque poly(A) ÷ RNA samples using full-length 32p-labeled macaque 3fl-HSD cDNA revealed the presence of an approximately 1.7 kb mRNA species in classical steroidogenic tissues, namely the ovary, testis and adrenal glands as well as in several peripheral tissues including the liver, kidney and epididymis. Computer analysis of the deduced macaque 3fl-HSD protein sequence predicts the presence of an NH2-terminal membrane-associated segment as well as four additional membrane-spanning segments, thus suggesting that 3fl-HSD is an integral protein. The availability of macaque cDNA should permit detailed studies concerning the tissue-specific expression as well as the hormonal regulation of 3fl-HSD mRNA in classical steroidogenic glands as well as in peripheral tissues which are an important site of steroidogenesis in primates.
Introduction
Steroid hormones are well recognized to play a crucial role in the development, growth and physi-
Address for correspondence: Dr. Jacques Simard, MRC Group in Molecular Endocrinology, CHUL Research Centre, 2705 Laurier Boulevard, Quebec, Que. G1V 4G2, Canada.
ological function of almost all mammalian tissues. Following cleavage of the aliphatic side-chain of cholesterol and the resulting formation of preg- nenolone and related As-3/3-hydroxysteroids, the next key step in the formation of all classes of steroid hormones, namely progesterone, mineralo- corticoids, ghicocorticoids, androgens and estro- gens, requires the obligatory oxidation and iso- merization by the enzyme AS-3fl-hydroxysteroid

102
dehydrogenase/AS-A 4 isomerase, hereafter called 3]~-HSD. This membrane-bound enzymatic system is found in classical steroidogenic tissues such as the placenta (Luu-The et al., 1989, 1990a), adrenal cortex (Ishii-Ohba et al., 1987), testis (Ishii-Ohba et al., 1986) and ovary (Zhao et al., 1989; Dupont et al., 1990a, b) as well as in several peripheral tissues including the prostate (Abalain et al., 1989; Lacoste et al., 1990), breast (Abul-Hajj, 1975), skin (Cameron et al., 1966), brain (Bauer and Bauer, 1989; Jung-Testas et al., 1989), liver (Lax and Schreifers, 1981) and kidney (Devine et al., 1986).
We have recently characterized human (Luu- The et al., 1989) and bovine (Zhao et al., 1989) 3]3-HSD cDNAs, both of which encode a deduced protein of 372 amino acids. The human 42 kDa 3/~-HSD protein possesses 3/3-hydroxysteroid de- hydrogenase as well as AS-a 4 isomerase activities as demonstrated by expression of the correspond- ing cDNA (Lachance et al., 1990). Moreover, a human 3~8-HSD gene was characterized (Lachance et al., 1990) and assigned by in situ hybridization to the p l l - p 1 3 region of the short arm of chro- mosome 1 (Brrub6 et al., 1989).
In contrast to animal models, namely rats, mice and domestic animals where the secretion of sex steroids takes place exclusively in the gonads (Cut- ler et al., 1978; Labrie et al., 1985; Brlanger et al., 1989), human adrenals secrete large amounts of the inactive precursor steroids dehydroepi- androsterone (DHEA) and especially its sulfate (DHEA-S) which are converted into potent androgens and estrogens in peripheral tissues. Since non-human primates also possess high plasma levels of adrenal precursor androgens (Cutler et al., 1978; Pepe and Albrecht, 1990), thus offering a model close to the human situa- tion, we have cloned and characterized the struc- ture of macaque 3fl-HSD cDNA as well as the tissue-specific expression of the corresponding mRNA species in steroidogenic and peripheral tissues.
Materials and methods
Construction and screening of the macaque ovary Xgtll eDNA #brary
Total RNA was isolated from three pairs of rhesus macaque (Macaea mulata) ovary by ho-
mogenizing tissue in guanidinium isothiocyanate followed by centrifugation through a cushion of 5.7 M CsCI as described (Melner et al., 1986; Low et al., 1989). Poly(A) + RNA was purified by using oligo(dT)-cellulose. A macaque ovary cDNA library was constructed in ?~gtll vector using both the oligo-dT-primed and random-primed methods by Clontech Laboratories (Palo Alto, CA, U.S.A.).
Approximately 4 x 10 s recombinant plaques from the macaque ovary ?~gtll cDNA expression library were screened with 32p-labeled human 3]3- HSD cDNA hp313-HSD63 (Luu-The et al., 1989) according to standard procedures (Zhao et al., 1989; Sambrook et al., 1989). Prehybridization was performed for 3 h at 37 °C in 30% formamide, 5 x SSPE (1 x SSPE being 0.18 M NaC1, 10 mM NaH2PO4, pH 7.4, 1 mM EDTA), 0.1% sodium dodecyl sulfate (SDS), 5 x Denhardt 's (0.1% bovine serum albumin (BSA), 0.1% Ficoll, 0.1% polyvinylpyrrolidone) and 100 # g / m l denatured salmon testis DNA. Thereafter, the [3zp]hp3/~- HSD63 cDNA probe (1 x 10 6 cpm/ml ) was add- ed to fresh prehybridization buffer. After 20 h of hybridization at 37°C, the filters were washed twice (20 min each) in 2 x SSC (1 x SSC being 0.15 M NaCI, 0.015 M sodium citrate, pH 7.0)/0.1% SDS at 25°C, once for 20 min in 2 x SSC/0.1% SDS at 4 0 ° C followed by 1 h in 1 X SSC/0.1% SDS at 40°C. Nitrocellulose filters were then exposed for 5 h at - 8 0 ° C . Seven plaques hybridized strongly with the probe. DNA from putative positive k clones was then isolated and first characterized by restriction endonuclease digestion and hybridization analysis with 32p_
labeled human 3AS-HSD cDNA. Inserts were ex- cised by EcoRI digestion and then subcloned into Bluescript KS vector (Stratagene).
DNA sequencing and computer analysis Sequencing of double-stranded plasmid DNA
was performed according to the dideoxy chain termination method (Sanger et at., 1977) using modified T 7 DNA polymerase (Tabor and Richardson, 1987) (Sequenase Kit, United States Biochemical Corp.). Oligonucleotides synthesized with a Biosearch DNA synthesizer as well as T 7 or T 3 vector primers were used as sequencing primers as shown in Fig. 1. In all cases, both DNA strands were sequenced. Analysis of the sequencing data

was performed using the Microgenie software (Beckman). Hydropathy profiles calculated ac- cording to Kyte and Doolittle (1982) were ob- tained on a VAX computer using the program of the University of Wisconsin Genetics Computer Group (Devereux et al., 1984). Analysis of trans- membrane segments was performed using the pro- grams RAOARGOS (Rao and Argos, 1986), HELIXMEM (Eisenberg et al., 1984) and SOAP (Klein et al., 1985) of the P C / G E N E software (Release 6.01; InteUi Genet ics /Genofi t ; Mountain View, CA, U.S.A.).
Messenger RNA isolation and RNA blot analysis RNA was isolated from a 7-year-old male and
a 7-year-old female macaque (Macaca fuscata) by the method of Chirgwin et al. (1979) essentially as described (Melner et al., 1986; Low et al., 1989). Immediately after dissection, the tissue was ho- mogenized in a 1 :10 dilution of 4 M guanidine thiocyanate, 0.5% sodium N-laurylsarcosine, 25 mM sodium citrate, pH 7.0, and 0.1 M 2- mercaptoethanol. Cesium chloride (Accurate Chemical Co., Hicksville, NY, U.S.A.) was added to the homogenate at 1 g/2.5 ml and layered onto a cushion of 5.7 M CsC1 in 0.1 M EDTA. The poly(A) + RNA was selected using a batch elution modification of the oligo d(T) affinity method previously described (Aviv and Leder, 1972). Iden- tical amounts of poly(A) + RNA were denatured in 50% formamide, 2.2 M formaldehyde, 40 mM 3-[N-morpholino]propanesulfonic acid, pH 7.0, at 55°C for 15 min and then loaded onto 1.5% agarose 2.2 M formaldehyde gels and electro- phoresed for 16 h at 20 V. After eleetrophoresis, the RNA was transferred to 0.1 /~m pore Nytran sheets (Schleicher & Schuell, Keene, NH, U.S.A.) by capillary action using 3 M NaC1, 0.3 M tri- sodium citrate. The blots were washed for 30 s in 1 M ammonium acetate and baked at 80 °C for 2 h.
After a 4 h prehybridization in 50% formamide, 5 x SSPE, 5 x Denhardt's, 0.1% SDS, 200 ~tg/ml yeast tRNA, 200 /xg/ml denatured salmon testis DNA and 2 ~ g / m l poly(A) ÷, RNA blots were hybridized with cDNA probes corresponding to cDNA inserts from mo3fl-HSD1 and mo3fl- HSD10 clones labeled by the random primer method (Feinberg and Vogelstein, 1983) added at
103
a concentration of 2 × 10 6 c p m / m l in fresh buffer containing 4% dextran sulfate. Hybridization was performed at 42°C for 16 h. Following hybridiza- tion, the filters were washed in 2 x SSC/0.1% SDS at 25°C (2 times, 10 rain), 1 x SSC/0.1% SDS at 65°C (2 times, 30 min) and 0.5 SSC/0.1% SDS at 65 °c for 30 min. The filters were autora- diographed at - 8 0 °C using Kodak XAR-5 films with intensifying screens.
Results
Isolation and characterization of cDNA clones To identify cDNAs encoding macaque 3fl-HSD,
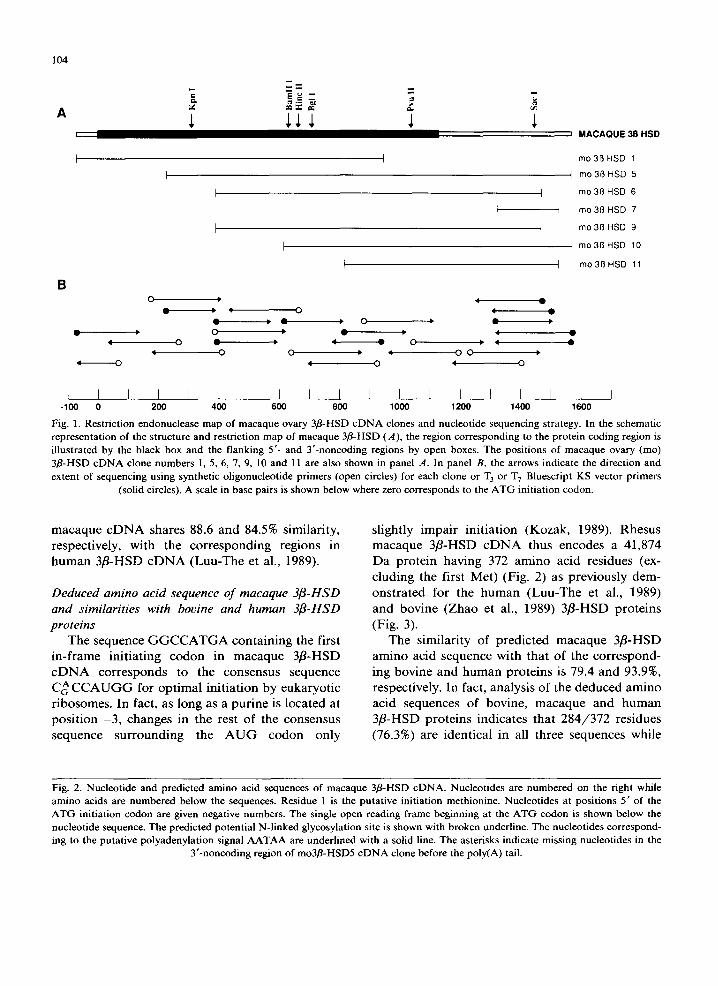
the 32p-labeled human hp3fl-HSD63 cDNA insert was used to screen 4 x 105 plaques from a rhesus macaque ovary Xgtl l library. Seven putative posi- tive clones showing a strong hybridization signal were detected. After subcloning in Bluescripts KS vector, the seven clones were then sequenced in both orientations by the dideoxy-chain termina- tion method. Restriction maps and lengths of these seven clones as well as the sequencing strategy are shown in Fig. 1.
All clones share identical sequences in their overlapping regions. The first in-frame ATG codon in the cDNA sequence is designated as position 1 and is preceded by an in-frame terminator codon TAG situated 63 nucleotides upstream. The nueleotide sequence of macaque eDNA has an open reading frame of 1119 nucleotides (Fig. 2). The mo3fl-HSD1 cDNA clone contains a 68 bp 5 untranslated region. The mo3fl-HSD5 and mo3fl- HSD10 cDNAs possess 420 and 425 bp, respec- tively, in their 3' untranslated regions before the poly(A) tail. Such heterogeneity in the site of polyadenylation has been previously demonstrated in human (Luu-The et al., 1989; Lachance et al., 1990; Lorence et al., 1990). The polyadenylation consensus signal (Proudfoot and Brownlee, 1976) is located 13 bp and 20 bp upstream from the poly(A) tail in cDNA clones mo3fl-HSD5 and mo3fl-HSD10, respectively. The nucleotide se- quence of the expected coding region of macaque cDNA shares 82.4 and 94.4% similarity with the corresponding sequence in bovine (Zhao et al., 1989) and human (Luu-The et al., 1989) 3fl-HSD cDNAs, respectively. Moreover, the nueleotide se- quence of the 5' and 3' untranslated regions of
104
A
B
I I I I
F
O > 4 ~.
0 0 u, q O 0 4 0 q 0 ,I 0
; M A C A Q U E 3B H S D
mo 3B HSD 1
4 mo 3~ HSD 5
mo 3B HSD 6
mo 3~ HSD 7
mo 3B HSD 9
I mo3BHSD 10
mo3~3HSD 11
L I I I I I I I I I 1 1 [ I 1 I I 1 I -100 0 200 400 600 800 1000 1200 1400 1600
Fig. 1. Restriction endonuclease map of macaque ovary 3fl-HSD c D N A clones and nucleotide sequencing strategy. In the schematic representation of the structure and restriction map of macaque 3fl-HSD (A), the region corresponding to the protein coding region is illustrated by the black box and the flanking 5'- and Y-noncoding regions by open boxes. The positions of macaque ovary (mo) 3fl-HSD cDNA clone numbers 1, 5, 6, 7, 9, 10 and 11 are also shown in panel A. In panel B, the arrows indicate the direction and extent of sequencing using synthetic oligonucleotide primers (open circles) for each clone or T 3 or T 7 Bluescript KS vector primers
(solid circles). A scale in base pairs is shown below where zero corresponds to the A T G initiation codon.
macaque cDNA shares 88.6 and 84.5% similarity, respectively, with the corresponding regions in human 3fl-HSD cDNA (Luu-The et al., 1989).
Deduced amino acid sequence of macaque 3fl-HSD and similarities with bovine and human 3~-HSD proteins
The sequence GGCCATGA containing the first in-frame initiating codon in macaque 3fl-HSD cDNA corresponds to the consensus sequence CA CCAUGG for optimal initiation by eukaryotic ribosomes. In fact, as long as a purine is located at position -3, changes in the rest of the consensus sequence surrounding the AUG codon only
slightly impair initiation (Kozak, 1989). Rhesus macaque 3fl-HSD cDNA thus encodes a 41,874 Da protein having 372 amino acid residues (ex- cluding the first Met) (Fig. 2) as previously dem- onstrated for the human (Luu-The et al., 1989) and bovine (Zhao et al., 1989) 3fl-HSD proteins (Fig. 3).
The similarity of predicted macaque 3fl-HSD amino acid sequence with that of the correspond- ing bovine and human proteins is 79.4 and 93.9%, respectively. In fact, analysis of the deduced amino acid sequences of bovine, macaque and human 3fl-HSD proteins indicates that 284/372 residues (76.3%) are identical in all three sequences while
Fig. 2. Nucleotide and predicted amino acid sequences of macaque 3fl-HSD cDNA. Nucleotides are numbered on the right while amino acids are numbered below the sequences. Residue 1 is the putative initiation methionine. Nucleotides at positions 5 ' of the A T G initiation codon are given negative numbers. The single open reading frame beginning at the A T G codon is shown below the nucleotide sequence. The predicted potential N-linked glyeosylation site is shown with broken underline. The nucleotides correspond- ing to the putative polyadenylation signal A A T A A are underlined with a solid line. The asterisks indicate missing nucleotides in the
Y-noncoding region of mo3fl-HSD5 c D N A clone before the poly(A) tail.

TAAGTTAGATCTGCTCCCCAAGCATCTTCTGTTTCCTGGCGAGTGATTCCTGCTACCTTGGATTGGCC
ATG ACG GGC TGG A~C TGC CTT GTG ACA GGA
Met Thr Gly Trp Ser Cys Leu Val Thr Gly
i0
GAG ATC AGG GTC TTG GAC AAG GCC TTC AGA
Glu Ile Arg Val Leu Asp Lys Ala Phe Arg
40
GGA GAC ATT CTG GAT GAG CCA TTC CTG AAG
Giy Asp Ile Leu Asp Glu Pro Phe Leu Lys
7O
GTC ACT CAC AGA GAG TCC ATC ATG AAC GTC
Val Thr His Arg Glu Ser Ile Met Asn Val
i00
ATC TAC ACC AGT ACC CTA GAG GTA GCC GGG
Ile Tyr Thr Ser Thr Leu Glu Val Ala Gly
130
TGG CCT GCT CCA TAC CCA TAC AGC AAA AAG
Trp Pro Ala Pro Tyr Pro Tyr Set Lys Lys
160
TAC ACT TGT GCC TTA AGA CCC ATG TAT ATC
Tyr Thr Cys Ala Leu Arg Pro Met Tyr Ile
190
ATC CTG TCA AGT GTT GGC AAG TTC TCC ACT
Ile Leu Ser Ser Val Gly Lys Phe Set Thr
220
CGG GAC CCC AAG AAG GCC CCA AGT GTC CAA
Arg Asp Pro Lys Lys Ala Pro Ser Val Gln
250
ATC CTG AGC AAA GAG TTT GGC CTC TGC CTT
Ile Leu Ser Lys G]u Phe GIy Leu Cys Leu
28O
GTG AGC TTC CTA CTC AGC CCA GTT TAC TCC
Val Ser Phe Leu Leu Set Pro Val Tyr Ser
310
TAT AAG AAG GCT CAG CGA GAT CTG GCG TAT
Tyr Lys Lys Ala Gln Arg Asp Leu Ala Tyr
340
GTG GAC CGG CAC AAG GAG ACC CTG AAG
Val Asp Arg His Lys G1u Thr Leu Lys
GCA GGA GGG TTT CTG GGT CAG AGG ATC GTC
Ala Gly Gly Phe Leu Gly Gln Arg Ile Val
2O
CCA GAA CTG AGG GAG GAA TTT TCC AAG
Pro GIu Leu Arg Glu Glu Phe Ser Lys
CGC CTC TTG GTG GAG GAG AAG GAG CTG AAG
Arg Leu Leu Val Glu Glu Lys Glu Leu Lys
30
CTC CAG AAT AAG ACC AAG CTG ACT GTA CTG GAA
Leu Gln Asn Lys Thr Lys Leu Thr Val Leu Glu
50 ...................
60
AGA GCC TGC CAG GAC GTC TCG GTC GTC ATC
Arg Ala Cys Gln Asp Val Ser Val Val Ile
8O
AAT GTG AAA GGT ACC CAG CTC CTG TTG GAG
Asn Val Lys Gly Thr Gln Leu Leu Leu Glu
ii0
CCC AAC TCC TAC AAG GAA ATC ATC CAG AAT
Pro ASh Ser Tyr Lys Glu Ile Ile Gln Asn
140
CTT GCT GAG AAG GCT GTG CTG GCA GCC AAT
Leu Ala Glu Lys Ala Val Leu Ala Ala Asn
170
TAT GGG GAA GGA GGC CCA TTC CTT TCT GCC
Tyr Gly Glu Gly Gly Pro Phe Leu Ser Ala
2O0
GTC AAC CCA GTC TAT GTT GGC AAC GTG GCC
Val Asn Pro Val Tyr Val Gly Asn Val Ala
230
GGA CAG TTC TAT TAC ATC TCA GAT GAC ACA
Gly Gln Phe Tyr Tyr Ile Set Asp Asp Thr
260
GAT TCC AGA TGG AGC CTT CCT TTA GCC CTG
Asp Ser Arg Trp Ser Leu Pro Leu Ala Leu
290
TAT CAA CCG CCC TTC AAC CGC CAC ACG GTG
Tyr Gln Pro Pro Phe Asn Arg His Thr Val
320
AAG CCG CTC TAC AGC TGG GAG GAA GCC
Lys Pro Leu Tyr Ser Trp Glu Glu Ala
CAC ACC GCC TGT ATC ATT GAT GTC TTT GGT
His Thr Ala Cys Ile Ile Asp Val Phe Gly
9O
GCC TGT GTC CAA GCC AGT GTG CCA GTC TTC
Ala Cys Val Gln Ala Ser Val Pro Val Phe
120
GGC CAT GAA GAA GAG CCT CTG GAG AAC ACA
Gly His Glu Glu Glu Pro Leu Glu ASh Thr
150
GGG TGG ACT CTG AAA AAT GGT GGC ACC TTG
Gly Trp Thr Leu Lys Asn Gly Gly Thr Leu
180
AGT ATA AAT GAG GCC CTG AAC AAC AAT GGG
Ser Ile Asn Glu Ala Leu Asn ASh ASh Gly
210
TGG GCT CAC ATT CTG GCC TTG AGG GCC CTG
Trp Ala His Ile Leu Ala Leu Arg Ala Leu
240
CCT CAC CAA AGC TAT GAT AAC CTT AAT TAC
Pro His Gln Set Tyr Asp Asn Leu Asn Tyr
270
ATG TAC TGG ATT GGC TTC CTG CTG GAA GTA
Met Tyr Trp Ile Gly Phe Leu Leu Glu Val
30O
ACA TTG TCA AAT AGC GTG TTC ACC TTC TCT
Thr Leu Set Asn Ser Val Phe Thr Phe Set
330
AAG CAG AAA ACT GTG GAG TGG GTT GGT TCC CTT
Lys Gln Lys Thr Val Glu Trp Val Gly Set Leu
350
360
TCC AAG ACT CAG TGA TTTAAGGATGACAGAGATGTGCATGTGGGTGTTGATAGGAGATGTCATCAAGCTCCACCCTCC
Ser Lys Thr Gln End
170
TGGCTTCATACAGAAGGTGACAAGGCACAAGCCCAGGTCCTGCTGCCTCCCTTTCACACAATGCCCAACTTATTGTCTTCTTCATGTCATCAAAATCTGCACAGTCACTGGCCCAACCA
GAAGCTTTCTGTCCTAATCATACGCCAGAGGACAGACAATATGATTTGCTGTTACCCAATCTCAGTGGCTGATTCTGAATGACTGTGGTCACTTTTAACTTGAGGTTCTCTTTTGACTA
ATGGAGCTAATTGAGCTCCATTT•CCCTCTTAAATGAGAAAGCATTTCTTTTCTCTTTAAT•TC•TATTACTTCACACAGTTCAATGTAAAGAGCAATAAATGTTTTAATGCTTAACCT
-i
90
180
270
360
450
540
630
720
810
900
990
10
80
1185
1304
1423
1542
GGAAAAAAAAAAAAAAAAA
1561

106
conservative changes are found at 25 additional positions, thus leading to an overall similarity of 83%, indicating that this enzyme is well conserved in mammalian species. This conclusion is particu- larly well supported by the high degree of similar- ity observed between hydropathy profiles of
macaque (Fig. 4) and bovine and human 3f l -HSD proteins (Luu-The et al., 1989; Zhao et al., 1989).
The predicted macaque 3f l -HSD protein con- tains one potential N-glycosylat ion site (Hubbard and Watt, 1981) at the Asn residue located at posit ion 52 (Fig. 2) as found in the deduced hu-
10 Bovine rfi]AlG W S C L V T GIGIG G F L G Q R Macaque ]MITIG W S C L V T G AIG G F L G Q R
I I I I I
Human IMITIGWS C L V T G A I G G F L G Q R
20 30 40 ' l ' C ~ ' - ~ E [ ' ~ D [ ' q Q I E I R V L D KIvlTIRI-~Iv I 1 V R I L L VlEIE KIEILIKIE I R V L D K AIFIRIP ElL [ I I RILL VIKIE KIEILIK[E l R V L D KJAIFIGIP E~L
50 R E E F S K L R E E F S K L R E E F S K L
Bovine Macaque Human
60 70 80
NIKITI K L TIV L E G D I L D E P FIL KIRIA C QI D VIS VlVll H T AIC NIKITIK L T I V L E G D I L D E P ElL KIRIA C Q I D V I S V I l l i H T A I C
90 100
1 F G V T H S I M N V I F G V T H S I M N V
110 120
v T o L g g V O V V ,IH T Macaque NV-K G T Q L L L E A C V Q A S V P V F 1 Y T L E V Human N V K G T Q L L L E A C V Q A S V P V F I Y S I E V
130 140 150 A G P N S Y I R D R II
I
A G P N S Y KIE I ' QINIGIHIE z EIP L N m A G P N S Y KIE 1 1 QINLGJHIE E EIP L N T
160 170 Bovine ~ S S [ - ~ ' ~ Y { S K K L A E K A V L I G ~ - ~ A [ L K N G G Macaque P AlP Y PIYIS K K L A E K A V L A I A N G W l T I L K N G G Human P A{P Y PIIIIS K K L A E K A V L AIA N G W]N{L K N G G
180 190 200
T L Y T C A L R P M Y I Y G E G G T L Y T C A L R P M Y 1 Y G E
210 220 230 240 250 Bovine Y M H / L N N N G I L I T N I t C G A ~ - - ~ ' ~ I R V N e V Y V G N V A W A H I L A L R A L R I [ ' ~ - ~ [ '~ V N I Q Macaque S I N E A I [ L N N N G 1 L S S VGIK F SITIVN P V Y V G N V A W A i t I L A L R A LIRID P K KIAIPIS V Q Human S I N E A L N N N G I L S S VGIK F S I T I V N P V Y V G N V A W A H I L A L R A LIQIDP KKIAIPIS I R
260 270
Macaque Q F Y Y I S D D T P H Q S Y D N I L N Y I I I L S K EIFIGIL Human Q F Y Y I S D D T P H Q s Y D NIL N YITIL S K EIFIGIL
280 290 300 C F ~ - - R ] M ~ ] L ~ i I S ~ Q ~ - ~ L A ~ I
A M G V CIL D S RJWISILIPIL I RIL D S RIWI~FIPIL S M I G I
310 320 330 340 350
Macaque S F L L SlPIVlYISlYIQIPIPIF N R tqT/V T L S N S V F T F S Y K K A Q R D L A[YIKIP L YISIW E E A K Human S F L L RIPI IIYITIYIRIPIPIF N R tl ] IV T L S N S V F T F S Y K K A Q R D L AIYIK[P L YIS[WE E A K
Bovine Macaque Human
360 370 Q K T I K ~ - - " ~ I ~ ' - L ' - ~ K QIH K E T L K I T ~ ] I tl Q K T VIE WIVIG S L ViD R II K Z T L K SIKIT Q Q K T VIE WIV[G S C V]D R ff K E T C K SIKIT Q
Fig. 3. Comparison of the deduced amino acid sequences of bovine, macaque and human 3fl-HSD proteins. Amino acid sequences are designated by the universal single-letter code. Amino acid residues are numbered relative to the first NH2-terminal methionine.
A 30
=~ l o
0 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1 - 30 1 , l I , I = I , I , I , I 0 5 0 100 150 2 0 0 2 5 0 3 0 0 3 5 0
B
Bovine
Macaque
Human
m
m
m
m m m
m m
160 1 5 0 2 0 0 2 5 0 3 0 0 350 AMINO ACID NUMBER
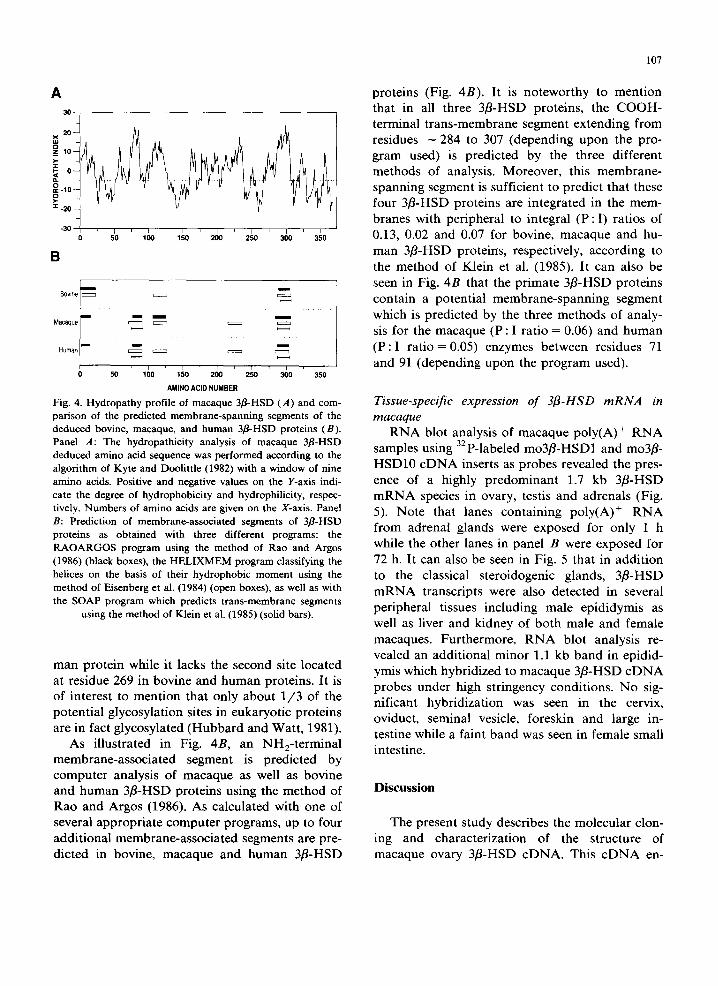
Fig. 4. Hydropathy profile of macaque 3fl-HSD (A) and com- parison of the predicted membrane-spanning segments of the deduced bovine, macaque, and human 3fl-HSD proteins (B). Panel A: The hydropathicity analysis of macaque 3fl-HSD deduced amino acid sequence was performed according to the algorithm of Kyte and Doolittle (1982) with a window of nine amino acids. Positive and negative values on the Y-axis indi- cate the degree of hydrophobicity and hydrophilicity, respec- tively. Numbers of amino acids are given on the X-axis. Panel B: Prediction of membrane-associated segments of 3fl-HSD proteins as obtained with three different programs: the RAOARGOS program using the method of Rao and Argos (1986) (black boxes), the HELIXMEM program classifying the helices on the basis of their hydrophobic moment using the method of Eisenberg et al. (1984) (open boxes), as well as with the SOAP program which predicts trans-membrane segments
using the method of Klein et al. (1985) (solid bars).
man protein while it lacks the second site located at residue 269 in bovine and human proteins. It is of interest to mention that only about 1 /3 of the potential glycosylation sites in eukaryotic proteins are in fact glycosylated (Hubbard and Watt, 1981).
As illustrated in Fig. 4B, an NH2-terminal membrane-associated segment is predicted by computer analysis of macaque as well as bovine and human 3fl-HSD proteins using the method of Rao and Argos (1986). As calculated with one of several appropriate computer programs, up to four additional membrane-associated segments are pre- dicted in bovine, macaque and human 3fl-HSD
107
proteins (Fig. 4B). It is noteworthy to mention that in all three 3fl-HSD proteins, the COOH- terminal trans-membrane segment extending from residues - 2 8 4 to 307 (depending upon the pro- gram used) is predicted by the three different methods of analysis. Moreover, this membrane- spanning segment is sufficient to predict that these four 3fl-HSD proteins are integrated in the mem- branes with peripheral to integral ( P : I ) ratios of 0.13, 0.02 and 0.07 for bovine, macaque and hu- man 3fl-HSD proteirrs, respectively, according to the method of Klein et al. (1985). It can also be seen in Fig. 4B that the primate 3fl-HSD proteins contain a potential membrane-spanning segment which is predicted by the three methods of analy- sis for the macaque (P : I ratio = 0.06) and human ( P : I ratio = 0.05) enzymes between residues 71 and 91 (depending upon the program used).
Tissue-specific expression of 3fl-HSD mRNA in macaque
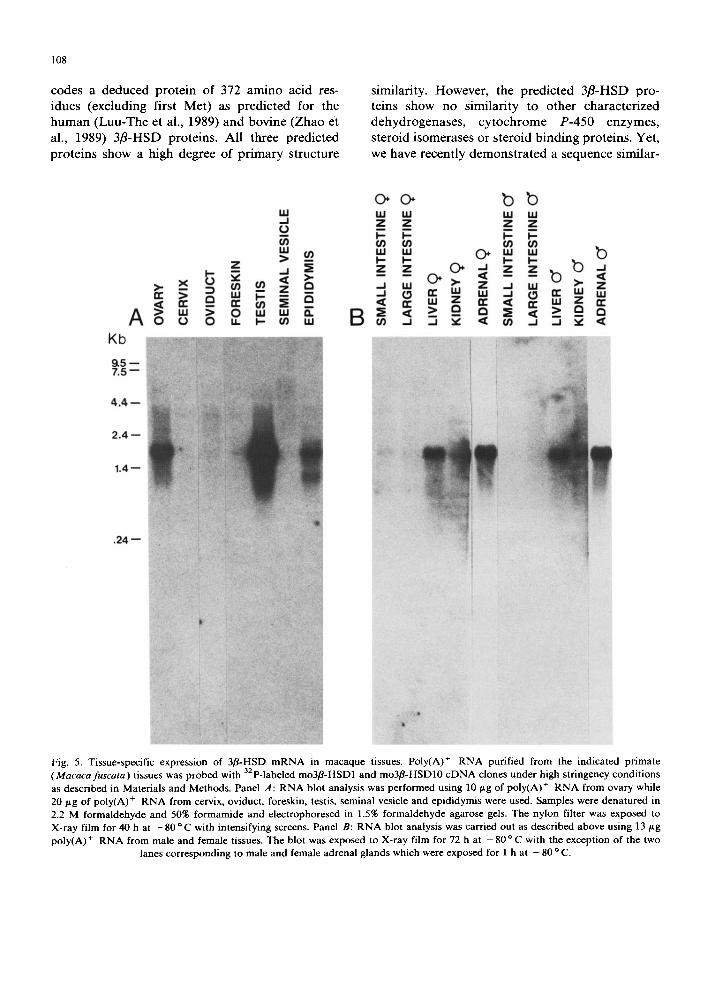
RNA blot analysis of macaque poly(A) + RNA samples using 3zp-labeled mo3fl-HSD1 and mo3fl- HSD10 cDNA inserts as probes revealed the pres- ence of a highly predominant 1.7 kb 3fl-HSD m R N A species in ovary, testis and adrenals (Fig. 5). Note that lanes containing poly(A) ÷ R N A from adrenal glands were exposed for only 1 h while the other lanes in panel B were exposed for 72 h. It can also be seen in Fig. 5 that in addition to the classical steroidogenic glands, 3fl-HSD m R N A transcripts were also detected in several peripheral tissues including male epididymis as well as liver and kidney of both male and female macaques. Furthermore, RNA blot analysis re- vealed an additional minor 1.1 kb band in epidid- ymis which hybridized to macaque 3fl-HSD c D N A probes under high stringency conditions. No sig- nificant hybridization was seen in the cervix, oviduct, seminal vesicle, foreskin and large in- testine while a faint band was seen in female small intestine.
Discussion
The present study describes the molecular clon- ing and characterization of the structure of macaque ovary 3fl-HSD cDNA. This cDNA en-
108
c o d e s a d e d u c e d p r o t e i n o f 372 a m i n o ac id res-
i d u e s ( e x c l u d i n g f i r s t M e t ) as p r e d i c t e d fo r t h e
h u m a n ( L u u - T h e e t al., 1989) a n d b o v i n e ( Z h a o e t
al., 1989) 3 f l - H S D p r o t e i n s . A l l t h r e e p r e d i c t e d
p r o t e i n s s h o w a h i g h d e g r e e o f p r i m a r y s t r u c t u r e
s imi l a r i t y . H o w e v e r , t he p r e d i c t e d 3 / 3 - H S D p r o -
t e i n s s h o w n o s i m i l a r i t y to o t h e r c h a r a c t e r i z e d
d e h y d r o g e n a s e s , c y t o c h r o m e P - 4 5 0 e n z y m e s ,
s t e r o i d i s o m e r a s e s o r s t e r o i d b i n d i n g p r o t e i n s . Ye t ,
we h a v e r e c e n t l y d e m o n s t r a t e d a s e q u e n c e s i m i l a r -
bb W I.u LU LLI UJ d Z Z Z Z
r-- F- r-- F-- or) (/) ¢J') (/) " ' " ' " ' " ' b
a ...I w Z ~ ,J u.I
K b
9 . 5 - - 7 . 5 - -
4 . 4 - -
2 . 4 - -
1 . 4 - -
b
l
. 2 4 - -
Fig. 5. Tissue-specific expression of 3fl-HSD mRNA in macaque tissues. Poly(A) + RNA purified from the indicated primate (Macaca fuscata) tissues was probed with 32p-labeled mo3fl-HSD1 and mo3fl-HSD10 eDNA clones under high stringency conditions as described in Materials and Methods. Panel A: RNA blot analysis was performed using 10 p.g of poly(A) + RNA from ovary while 20/tg of poly(A) + RNA from cervix, oviduct, foreskin, testis, seminal vesicle and epididymis were used. Samples were denatured in 2.2 M formaldehyde and 50% formamide and electrophoresed in 1.5% formaldehyde agarose gels. The nylon filter was exposed to X-ray film for 40 h at - 8 0 °C with intensifying screens. Panel B: RNA blot analysis was carried out as described above using 13 ttg poly(A) + RNA from male and female tissues. The blot was exposed to X-ray film for 72 h at - 8 0 °C with the exception of the two
lanes corresponding to male and female adrenal glands which were exposed for 1 h at - 80 ° C.

ity between bovine and human 3fl-HSD and plant dihydroflavonol reductase (Baker et al., 1990).
Subcellular distribution studies have shown that the 3fl-HSD enzyme is a membrane-bound protein located in microsomal as well as mitochondrial membranes (Ishii-Ohba et al., 1986, 1987; Luu-The et al., 1989, 1990a; Inano et al., 1990). Such find- ings are in close agreement with the present com- puter analysis of bovine, macaque and human 3fl-HSD proteins indicating the presence of several potential membrane-associated segments (Fig. 4B).
The present study clearly demonstrates that the 32p-labeled macaque 3fl-HSD cDNA probe specifically hybridizes with a 1.7 kb mRNA tran- script(s) in ovary, testis and adrenal glands in agreement with the presence of a 1.7 kb 3fl-HSD mRNA species in the bovine ovary (Zhao et al., 1989) and in human placenta (Luu-The et al., 1989). It is of special interest that 3fl-HSD mRNA transcripts were also detected in macaque periph- eral tissues as diverse as the epididymis, liver and kidney while a faint 1.7 kb band was also revealed after longer exposure time in female small in- testine.
The present study thus demonstrates for the first time 3/3-HSD gene expression in these periph- eral tissues. This finding in the primate is of special interest following the observation of 3/3- HSD activity in several normal and cancerous peripheral tissues including the prostate (Abalain et al., 1989; Bartsch et al., 1990; Lacoste et al., 1990), breast (Abul-Hajj, 1975), endometrium (Bonney et al., 1985), skin (Cameron et al., 1966), and lung (Milewich et al., 1977). The importance of sex steroid formation in peripheral tissues is also indicated by the widespread distribution of expression of other steroidogenic enzymes, espe- cially 17/3-hydroxysteroid dehydrogenase, aroma- tase and 5a-reductase (Labrie et al., 1985, 1989; Milewich et al., 1989; Bartsch et al., 1990; Castagnetta et al., 1990; Killinger et al., 1990; Luu-The et al., 1990b; and references therein). These findings led to the new concept of in- tracrine activity whereby locally produced steroids can exert their activity inside the same cells where biosynthesis takes place (Labrie et al., 1988).
The interest of 3/3-HSD expression in periph- eral tissues also pertains to the observation of
109
steroid 21-hydroxylation in extraadrenal tissues in the rat and human (Casey and MacDonald, 1982; Casey et al., 1983). Furthermore, it has been pos- tulated that in guinea-pig kidney, 3B-HSD can be involved in ll-deoxycorticosterone formation in situ (Devine et al., 1986).
The availability of the macaque 3fl-HSD cDNA should permit detailed investigation of the hormonal control of 3/3-HSD gene expression in classical steroidogenic glands as well as in periph- eral tissues using highly sensitive and specific methods such as S1 nuclease mapping and ribonuclease A protection analysis. Since the monkey adrenal is unique among all experimental animals to secrete large amounts of precursor sex steroids (Cutler et al., 1978) which can be con- verted into active androgens and estrogens in pe- ripheral tissues, the present model offers a unique opportunity to study the regulation of 3fl-HSD and other sex steroid forming enzymes in periph- eral tissues and thus gain knowledge about the human situation when peripheral in situ formation plays an important role (Labrie et al., 1985, 1988).
References
Abalain, J.H., Quemener, E., Carre, J.L., Simon, B., Amet, Y., Mangin, P. and Floch, H.H. (1989) J. Steroid Biochem. 34, 467-471.
Abui-Hajj, Y.J. (1975) Steroids 26, 488-500. Aviv, H. and Leder, P. (1972) Proc. Natl. Acad. Sci. U.S.A. 69,
1408-1412. Baker, M.E., Luu-The, V., Simard, J. and Labile, F. (1990)
Biochem. J. 269, 558-559. Bartsch, W., Klein, H., Schiemann, U., Bauer, H.W. and Voigt,
K.D. (1990) Ann. N.Y. Acad. Sci. 595, 53-66. Bauer, H.C. and Bauer, H. (1989) J. Steroid. Biochem. 33,
643-646. Brlanger, B., Brlanger, A., Labile, F., Dupont, A., Cusan, L.
and Monfette, G. (1989) J. Steroid Biochem. 32, 695-698. Brrubr, D., Luu-The, V., Lachance, Y., Gagnr, R. and Labile,
F. (1989) Cytogenet. Cell Genet. 52, 199-200. Bonney, R.C., Reed, M.J., Beranek, P.A. and James, V.HT.
(1985) J. Steroid Biochem. 23, 347-352. Cameron, E.M., Baillie, A.M., Grant, J.K., Milne, J.A. and
Thompson, J. (1966) J. Endocrinol. 35, 19-20. Casey, M.L. and McDonald, P.C. (1982) Endocr. Rev. 3,
396-403. Casey, M.L., Winkel, C.A. and MacDonald, P.C. (1983) J.
Steroid Biochem. 18, 449-452. Castagnetta, L., d'Aquino, S., Labile, F. and Bradlow, H.L.
(1990) Ann. N.Y. Acad. Sci. 595, 489p. Chirgwin, J.M., Przybyla, A.E., MacDonald, R.J. and Rutter,
W.J. (1979) Biochemistry 18, 5294-5299.

110
Cutler, Jr., G.B., Glenn, M., Bush, M., Hodgen, G.D., Graham, C.E. and Loriaux, D.L. (1978) Endocrinology 103, 2112- 2118.
Devereux, J., Haeberli, P. and Smithies, O. (1984) Nucleic Acids Res. 12, 387-395.
Devine, P.L., Kelly, N. and Adams, J.B. (1986) J. Steroid Biochem. 25, 265-271.
Dupont, E., Luu-The, V., Labile, F. and Pelletier, G. (1990a) Endocrinology 126, 2906-2909.
Dupont, E., Zhao, H.F., Rh6aume, E., Simard, J., Luu-The, V., Labile, F. and Pelletier, G. (1990b) Endocrinology 127, 1394-1403.
Eisenberg, D.S., Schwartz, E., Komaromy, M. and Wall, R. (1984) J. Mol. Biol. 179, 125-142.
Feinberg, A.P. and Vogelstein, B. (1983) Anal. Biochem. 132, 6-13.
Hubbard, S.C. and Watt, R.J. (1981) Annu. Rev. Biochem. 50, 555-583.
Inano, H., Ishii-Ohba, H., Sugimoto, Y., Ohta, Y., Morikawa, T., Yoshida, M. and Tamaoki, B. (1990) Ann. N.Y. Acad. Sci. 595, 17-25.
Ishii-Ohba, H., Saiki, N., Inano, H. and Tamaoki, B.I. (1986) J. Steroid Biochem. 24, 753-760.
Ishii-Ohba, H., lnano, H. and Tamaoki, B.I. (1987) J. Steroid Biochem. 27, 775-779.
Jung-Testas, I., Hu, Z.Y., Beaulieu, E.E. and Robel, P. (1989) Endocrinology 125, 2083-2091.
Killinger, D.W., Perel, E., Daniilescu, D., Kharlys, L. and Lindsay, W.R.N. (1990) Ann. N.Y. Acad. Sci. 595, 199-211.
Klein, P., Kanehisa, M. and De Lisi, C. (1985) Biochim. Biophys. Acta 815, 468-476.
Kozak, M. (1989) J. Cell Biol. 108, 229-241. Kreil, G. (1981) Annu. Rev. Biochem. 50, 317-348. Kyte, J. and Doolittle, R.F. (1982) J. Mol. Biol. 157, 105-132. Labile, F., Dupont, A. and B61anger, A. (1985) in Important
Advances in Oncology (De Vita, V.T., Hellman, S. and Rosenberg, S.A., eds.), pp. 193-200, J.B. Lippincott, Philadelphia, PA.
Labile, C., B61anger, A. and Labile, F. (1988a) Endocrinology 123, 1412-1417.
Labile, F., B61anger, A., Veilleux, R., Lacoste, D., Labile, C., Marchetti, B., Poulin, R., Dupont, A., Cusan, L. and Luthy, I. (1988b) Bailli6re's Clin. Oncol. 2, 597-619.
Lachance, Y., Luu-The, V., Labile, C., Simard, J., Dumont, M., de Launoit, Y., Gu6rin, S., Leblanc, G. and Labile, F. (1990) J. Biol. Chem. 265 (in press).
Lacoste, D., B61anger, A. and Labile, F. (1990) Ann. N.Y. Acad. Sci. 595, 389-391.
Lax, E.R. and Schriefers, H. (1981) Acta Endocilnol. 98, 261-266.
Lorence, M.C., Muny, B.A., Trant, J.M. and Mason, J.I. (1990) Endocrinology 126, 2493-2498.
Low, K.G., Nielsen, C.P., West, N.B., Douglass, J., Brenner, R.M., Maslar, I.A. and Melner, M.H. (1989) Mol. Endo- crinol. 3, 852-857.
Luu-The, V., Lachance, Y., Labile, C., Leblanc, G., Thomas, J.L., Stilckler, R.C. and Labile, F. (1989) Mol. Endocrinol. 3, 1310-1312.
Luu-The, V., Takahashi, M. and Labile, F. (1990a) Ann. N.Y. Acad. Sci. 595, 386-388.
Luu-The, V., Labile, C., Simard, J., Lachance, Y., Zhao, H.F., Cou~t, J., Leblanc, G. and Labile, F. (1990b) Mol. Endo- crinol. 4, 268-275.
Melner, M.H., Young, S.L., Czerwiec, F.S., Lyn, D., Puett, D., Roberts, J.L. and Koos, R.D. (1986) Endocrinology 119, 2082-2088.
Milewich, L., Winters, A.J., Stephens, P. and MacDonald, P.C. (1977) J. Steroid Biochem. 8, 277-284.
Milewich, L., MacDonald, P.C. and Cart, B.R. (1989) J. Endo- crinol. 123, 509-518.
Pepe, G.J. and Albrecht, E.D. (1990) Endocr. Rev. 11,151-171. Proudfoot, N.J. and Brownlee, G.G. (1976) Nature 263, 211-
214. Rao, M.J.K. and Argos, P. (1986) Biochim. Biophys. Acta 869,
197-214. Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular
Cloning: A Laboratory Manual, 2nd edn., Cold Spring Harbor Press, Cold Spring Harbor, NY.
Sanger, E., Milken, S. and Coulson, A.R. (1977) Proc. Natl. Acad. Sci. U.S.A. 74, 5453-5467.
Tabor, S. and Richardson, C.C. (1987) Proc. Natl. Acad. Sci. U.S.A. 84, 4767-4771.
Zhao, H.F., Simard, J., Labile, C., Breton, N., Rh6aume, E., Luu-The, V. and Labile, F. (1989) FEBS Lett. 259, 153-157.





























