Embed Size (px)
Citation preview

1021-4437/03/5004- $25.00 © 2003
MAIK “Nauka
/Interperiodica”0449
Russian Journal of Plant Physiology, Vol. 50, No. 4, 2003, pp. 449–454. Translated from Fiziologiya Rastenii, Vol. 50, No. 4, 2003, pp. 505–510.Original Russian Text Copyright © 2003 by Deryabin, Trunova, Dubinina, Burakhanova, Sabel’nikova, Krylova, Romanov.
INTRODUCTION
It was established long ago that plants accumulatesoluble sugars during adaptation to chilling and freez-ing, and such accumulation was shown to correlate withthe development of cold and frost resistance [1–3]. Theevidence demonstrating an enhanced resistance of planttissues and organs and of intact plants after hardeningon the exogenous sugar solutions is a proof of protec-tive activity of sugars [2, 4]. The role of low-molecular-weight carbohydrates in plant tolerance and adaptationunder chilling stress was substantiated by the data onsugar capacities for osmoregulation and protection ofprotein and lipid cell components [5]. Sugar ability tocombine free radicals and in this way to prevent cold-induced destructive oxidation processes, such as POLin the biological membranes, is one of the mechanismsof sugar protective activity under low temperatures [6].
Chilling results in the hydrolysis of sucrose andfructan polymers [7–9]. By breaking up glycosyl–gly-
coside bonds [10–13], such enzyme of carbohydratemetabolism as invertase (
β
-D-fructofuranoside-fructo-hydrolase, EC 3.2.1.26) plays a key role in changing thecomposition and ratio of various fractions of solublecarbohydrates. The changes in invertase activity undercold stress were reported in many plant species; thesechanges are thought to arise from the reversible transferof soluble to bound forms of the enzyme [14] and alsofrom invertase synthesis
de novo
[15]. An enhancedinvertase activity under cold stress was observed onlyin uninjured plants [16], and we therefore believe thatsuch increase in invertase activity reflects plant defen-sive and adaptive reactions, rather than the destructiveprocesses. An additional proof to this statement comesfrom the fact that the increase in invertase activity rap-idly develops in cold-treated plants [16]. The enhancedinvertase activity with the concomitant increase in thecontent of low-molecular-weight carbohydrates werereported among the responses to low temperature inboth frost-resistant (wheat) and chilling-tolerant(tomato) plant species [17, 18]. Although the interrela-tion between the content of low-molecular-weight car-bohydrates and plant cold tolerance is well established,
Chilling Tolerance of Potato Plants Transformed with a Yeast-Derived Invertase Gene under the Control
of the
B33
Patatin Promoter
A. N. Deryabin, T. I. Trunova, I. M. Dubinina, E. A. Burakhanova, E. P. Sabel’nikova, E. M. Krylova, and G. A. Romanov
Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, Botanicheskaya ul. 35, Moscow, 127276 Russia;fax: 7 (095) 977-8018; e-mail: [email protected]
Received April 24, 2002
Abstract
—Tolerance to chilling was compared under
in vitro
conditions in potato plants (
Solanum tuberosum
L., cv. Désirée) transformed with a yeast-derived invertase gene under the control of the
B33
class 1 tuber-spe-cific promoter (the
B33-inv
plants) and potato plants transformed only with a reporter gene (the control plants).The expression of the inserted yeast invertase gene was proved by following the acid and alkaline invertaseactivities and sugar contents in the leaves under the optimum temperature (22
°
C). The total activities of acidand alkaline invertases in the
B33-inv
plants exceeded those in the control plants by the factors of 2–3 and 1.3,respectively. In the
B33-inv
plants, the activity of acid invertase twice exceeded that of the alkaline invertase,whereas the difference equaled 12% in the control plants. The contents of sucrose and glucose increased in the
B33-inv
plants by 21 and 13%, respectively, as compared to the control. Chilling at +3 and –1
°
C for 1, 3, and6 h did not affect the rate of lipid peroxidation, as measured by the content of malonic dialdehyde (MDA) inthe leaves of the genotypes under study. Only the longer exposures (24 h at +3 and –1
°
C and 7 days at +5
°
C)produced a significant decline in the MDA content in the
B33-inv
plants, as compared to the control. Followingshort freezing (20 min at –9
°
C), the content of MDA increased by 50% in the leaves of the control plants, whilein the
B33-inv
plants, cold-treated and control plants did not differ in the MDA content. The authors presumethat the potato plants transformed with the yeast invertase gene acquire a higher tolerance to low temperaturesas compared to the control plants, apparently due to the changes in sugar ratio produced by the foreign invertase.
Key words: Solanum tuberosum - B33 patatin promoter - invertase - lipid peroxidation - malonic dialdehyde -chilling
Abbreviations
: AOS—antioxidase system; CaMV—cauliflowermosaic virus; GUS—
β
-glucoronidase; POL—peroxidation oflipids; MDA—malonic dialdehyde.

450
RUSSIAN JOURNAL OF PLANT PHYSIOLOGY
Vol. 50
No. 4
2003
DERYABIN
et al
.
the functional state of hydrolases is poorly studied.Transgenic plants can help to fill this gap. Transgenicplants are increasingly used as models for studyingplant responses to stresses and as sources of tolerantplant forms [19, 20].
At the Max-Planck Institute of Plant MolecularPhysiology (Golm, Germany), potato plants were trans-formed with diverse genes, including a yeast-derivedinvertase gene under the
B33
patatin promoter [21].Expression of this gene, with the apoplastic localizationof the enzyme, produced larger tubers, though in lowernumbers; in contrast, the transformed plants with inver-tase localized in the cytosol produced smaller tubers inhigher numbers per plant [22].
We suggested that the products of the yeast invertasegene expression changed the carbohydrate status ofplant vegetative organs. Hence, the present study wasfocused on the relationship between the expression ofthe yeast invertase gene in potato plants cultured
invitro
and their tolerance to chilling.
MATERIALS AND METHODS
Our study was carried out on potato plants (
Solanumtuberosum
L., cv. Désirée) transformed with the yeast
inv
invertase gene under the tuber-specific patatin class1 promoter (
B33
) carrying also the proteinase II leaderpeptide sequence providing for the apoplastic enzymelocalization (abbreviated below as the B33-
inv
plants).Plants transformed with the marker gene encoding
β
-glucuronidase (
GUS
), which does not play any physio-logical role in higher plants, under the 35S promoter ofcauliflower mosaic virus (CaMV) were also used. Thetransformed plants also harbored the kanamycin resis-tance gene.
Plants were propagated
in vitro
by cuttings andgrown in a phytotron chamber of the Timiryazev Insti-tute of Plant Physiology, RAS, at 22
°
C and a 16-h pho-toperiod (an illuminance of 5 klx) during 5 weeks in thetest-tube culture on the Murashige and Skoog mediumcontaining 0.7% agar, 2% sucrose, 0.5 mg/l thiamine,0.5 mg/l pyridoxine, and 60 mg/l
meso
-inositol.
The expression of the patatin promoter was studiedin potato plants cv. Désirée transformed with the DNAconstruction comprising the reporter gene
GUS
attached to the class 1 patatin promoter (
B33
-promoter)[23]. The
B33
-
GUS
transformed plants were obtainedin the Laboratory of Growth and Development,Timiryazev Institute of Plant Physiology, RAS; thepresence and expression of the inserted
GUS
gene weresubstantiated by the polymerase chain reaction anddirect measurement of GUS activity. The transgenicpotato plants, cv. Désirée, with the inserted
GUS
geneexpressed under the 35S CaMV promoter (the 35S-
GUS
plants) were used as a control. Plants were grownunder standard conditions on the Murashige and Skoogmedium containing 3% sucrose.
Chilling tolerance of potato genotypes under studywas assessed by the accumulation of MDA, the second-ary POL product, in leaves. To this end, leaves wereselected from the middle part of plants, and 0.3-g sam-ples of leaves devoid of petioles were put into pre-cooled weighing bottles and transferred into a coolingchamber wherein they were exposed to +3, –1, and
−
9
°
C for the periods ranging from 20 min to 24 h. Toassess the effects of prolonged cooling (for 7 days),test-tube plants were kept in the phytotrone chamberunder the light (5 klx) at +5
°
C. MDA content was mea-sured by the color thiobarbiturate reaction, and the opti-cal density was determined using an SF-46 spectropho-tometer (LOMO, Russia). MDA concentration was cal-culated from the change in molar extinction [24] andpresented as
µ
mol/g fr wt of leaves. Each replicationrepresents a sample collected from 2–4 plants.
To determine invertase activity, 1-g leaf sampleswere homogenized in cold (+4
°
C) with a mortar andpestle in a phosphate–citrate buffer, and, in order toremove soluble sugars, the slurry thus obtained wasdialyzed for 20 h against 10-fold diluted buffer, pH 7.0,comprising 0.1 M citrate and 0.2 M Na
2
HPO
4
· 2H
2
O.Following the dialysis, the homogenate was centri-fuged at 18000
g
for 20 min, and the supernatant frac-tion (cytosol) comprising the soluble cell fraction withadmixture of microsomes, which were devoid of inver-tase activity, was used for determination of solubleinvertase activity. The pellet (cell-wall fraction) wasrinsed three times with the diluted buffer, with centri-fugation at 200
g
for 10 min each time, and used fordetermination of precipitated invertase. The incubationmixture comprised 0.2 ml of enzyme preparation,0.3 ml of buffered sucrose solution (the final sucroseconcentration in the incubation mixture was 150 mM),and one drop of toluene. To determine acid invertaseactivity, we used 1 M acetate buffer, pH 4.7; for alkalineinvertase activity, citrate buffer, pH 7.5. In the controlsamples, invertase was preliminarily inactivated byboiling for 2 min. Mixtures were incubated for 1 h at30
°
C, and enzyme activities were assessed by accumu-lation of glucose (
µ
mol/g fr wt) determined by the glu-cooxidase method [25]. Sucrose and fructose contentsin leaf tissues were determined with the method of Roe
et al.
[25]. Tables present the means and their standarderrors from three experiments. The significance of dif-ferences between the treatments was estimated usingthe Student’s criterion.
RESULTS AND DISCUSSION
In the context of the present study, it was critical tofind out whether the tuber-specific patatin promoter
B33
is expressed in other potato organs. The activity ofthe patatin promoter was measured in the vegetativeorgans of 2.5-month-old transgenic potato plants, cv.Désirée, where this promoter controlled the expressionof the reporter
GUS
gene
.
In potato vegetative organs,such as stem, root, and leaf, we observed low and none-
RUSSIAN JOURNAL OF PLANT PHYSIOLOGY
Vol. 50
No. 4
2003
CHILLING TOLERANCE OF POTATO PLANTS 451
theless significant activity of foreign
GUS
(Table 1);this evidence indicated the limited expression of thepatatin promoter in these organs. Among the vegetativeorgans of the
B33-GUS
plants, leaves manifested thehighest activity of the patatin promoter: about 10% ofthe corresponding activity in the vegetative organs ofthe 35S-
GUS
plants (Table 1).
The efficiency of the inserted yeast invertase geneswas confirmed by comparing the activities of two formsof invertase in the leaves of two genotypes under studyat 22
°
C. The introduction of the yeast invertase geneenhanced the total activity of acid invertase two–three-fold (Table 2), whereas the total activity of alkalineinvertases increased only by the factor of 1.3, as com-pared to the control plants. The activity of acid inver-tase in the
B33-inv
plants exceeded twofold that ofalkaline invertase, and in the control plants, only by12%. Similar to plant invertases, yeast-derived inver-tase is known to exist in several isoforms, which differin their subcellular location: vacuolar, exocellular, orapoplastic, and cytosolic [26]. The optimum pH forcell-wall invertase activity is in the acidic region. In ourexperiments, we used the plants with apoplastic inver-tase location. The apoplastic medium with pH 4.5–4.7[27] provided for the highest activity of the yeast inver-tase [28]. The twofold increase in cell-wall invertaseactivity observed in the
B33-inv
plants, as compared tothe control plants, was apparently related to the adsorp-tion of secreted invertase [28]. We presumed that theincrease in the leaf invertase activity would affect thecontents of mono- and disaccharides and found thatsucrose and glucose contents in leaves of the
B33-inv
plants exceeded by 21 and 13%, respectively, the corre-
sponding indices in the control plants (Table 3). Manyauthors working with transgenic plants are of opinionthat sugar accumulation takes place when the activeapoplastic invertase hydrolyzes sucrose partitioned fortransport [29] and hexoses thus produced cannot enterthe phloem [30]. Hexoses come back to the mesophyllcells [31], are rephosphorylated by hexokinase andfructokinase and enter the pathway leading to sucrosesynthesis. Thus, the expression of apoplastic invertasein the
B33-inv
potato leaves would result in the accu-mulation of sugars, primarily glucose and fructose.
In contrast to wild
Solanum
species, potato cultivarsare resistant to chilling, rather than freezing [32] andcan tolerate cooling down to –4
°
C. At –5
°
C and belowthis temperature, plants perish [33]. Hence, our goalwas to select cooling conditions producing such initialcell injuries that would differ depending on the chillingtolerance of the genotypes under study.
It is known that, in addition to many biochemicalprocesses, plant adaptation to hypothermia is accompa-nied with an increase in the content of unsaturated fattyacids [3], and POL affects their level in cell mem-branes. POL is a natural metabolic reaction, which isnormally regulated by an antioxidant system (AOS) ofsufficiently high buffer capacity. However, as thiscapacity is exhausted in plant cells exposed to strongand prolonged stress, the balance is shifted towardsPOL activation [34]. This shift is related to MDA accu-mulation due to the activation of free-radical reactionsinvolving the reactive oxygen species that participate inthe peroxidation of unsaturated fatty acids in mem-brane lipids; as a result, the amount of membrane mate-rial decreases [35–37]. Therefore, to evaluate plant tol-erance to hypothermia, we employed the method ofassessing POL by the content of MDA, one of its termi-nal products.
Table 1.
Activity of the patatin promoter in the vegetativeorgans of potato plants
Transformedplant Organ Specific GUS activity,
units/mg protein
B33
-GUS
, line L1 leaf 49.3
±
10.6
stem 11.9
±
1.6
root 4.1
±
0.2
35S-
GUS
stem 660
±
183
root 541
±
22
Table 2.
Activities of acid and alkaline invertases in potatoleaves,
µ
mol glucose produced per g fr wt
FractionControl
B33-inv
plants
acid alkaline acid alkaline
Sediment 34.3
±
2.0 25.8
±
1.8 66.2
±
4.6 34.7
±
2.4
Cytosol 36.0
±
2.7 31.6
±
2.5 91.1
±
8.2 42.9
±
3.4
Table 3.
Sugar contents in potato leaves,
µ
g/g fr wt
Sugars Control
B33-inv
plants % of the control
Sucrose 5075
±
405.0 6153
±
430.7 121
Fructose 681
±
47.6 351
±
28.1 51
Glucose 4819
±
337.3 5444
±
381.1 113
Total monosaccharides 5500
±
494.9 5795
±
521.6 105
Total sugars 10575
±
951.8 11948
±
955.8 113
452
RUSSIAN JOURNAL OF PLANT PHYSIOLOGY Vol. 50 No. 4 2003
DERYABIN et al.
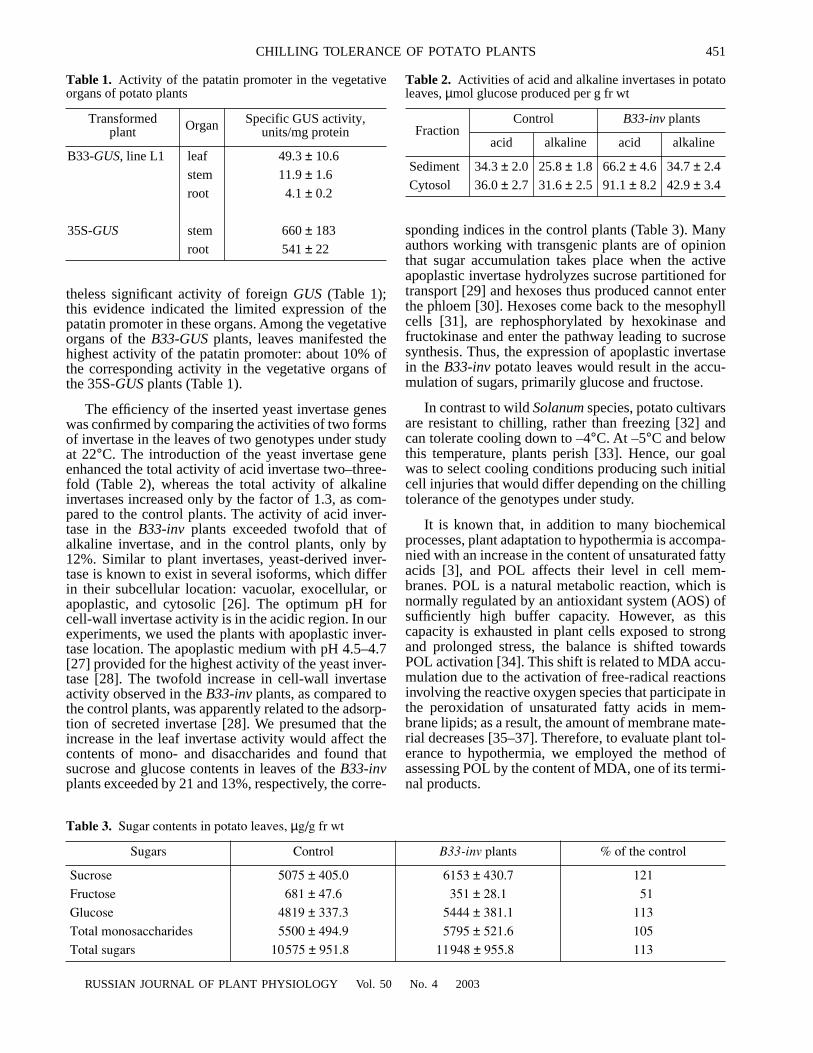
We observed that, in each experiment carried out at22°C, the MDA content in the plants transformed withthe yeast invertase exceeded the control value by 20–40%. Following a one-hour-long cooling at +3°C, theMDA content in the leaves somewhat declined in bothgenotypes (Fig. 1a). With extending the period of coldexposure to 3 and 6 h, the content of MDA in the con-trol plants increased, while in the B33-inv leaves, POLactivity even slightly diminished, though the differencebetween two treatments was not significant. Only 24-h-
long chilling produced an increase in MDA content: 5%in the B33-inv plants and 17% in the control. We relatethe difference to the higher AOS activity mobilized inthe former case.
Exposing plants to –1°C for 1, 3, and 6 h also did notdemonstrate any difference in the POL rates: in bothgenotypes, the content of MDA slightly decreased ascompared to unexposed plants. Extending cold expo-sure to 24 h produced a dramatic POL activation: theMDA content increased by 62 and 36% in leaves of thecontrol and B33-inv plants, respectively (Fig. 1b).Apparently, high cold tolerance of potato plants pre-vented us from finding significant differences in MDAcontents between the genotypes under study, when weexposed plants to 3°C and –1°C for 1, 3, and 6 h. Onlyfollowing the prolonged chilling exposure, such differ-ences became evident, with the MDA content in theB33-inv plants significantly lower than in the control.
Similar data were obtained when potato plants wereexposed to 5°C for 7 days in the light (Table 4). Weobserved a twofold increase of the MDA content in thecontrol leaves, whereas in the B33-inv plants, this indexincreased only by the factor of 1.5. We believe that sucha long exposure to chilling exhausted the reserves ofplant AOS capacity and produced a burst of POL activ-ity, although such burst in the B33-inv plants laggedbehind the control.
To discern between the cold-tolerance indices of thecontrol and B33-inv plants and to demonstrate the pri-mary POL activation, it is necessary to select a combi-nation of temperatures and exposure periods that wouldpreclude ice crystallization in plant tissues and celldeath and would maintain plant tissues in supercooledstate [34]. POL belongs to free-radical chain reactions,and the POL products can be found as soon as 0.5–1 hafter the beginning of plant exposure to cold stress [38];hence we subjected potato plants to short (20 min) andstringent (–9°C) freezing. Our data (Table 4) demon-strated that the MDA content in the control leavesincreased by 50%; however, in the B33-inv plants, theMDA content stayed at the level of plants untreatedwith cold presuming that the B33-inv plants developedhigher cold tolerance.
Summing up our evidence, we presume that the cellmembrane complex in the potato plants transformedwith the yeast-derived invertase gene manifests higherresistance to low temperatures than that of the controlplants. This difference is especially vivid under pro-longed (24 h) exposure to +3 and –1°C or under short(20 min) and stringent (–9°C) hypothermia. Appar-ently, an enhanced cold resistance of the B33-inv plantsresulted from the changes in sugar composition pro-duced by the foreign invertase.
ACKNOWLEDGMENTS
The study was supported by the Russian Foundationfor Basic Research, project no. 01-04-49085.
~~
~~
1
120
110
100
90
0
160
140
120
100
80
60
01 3 6 24
Cold exposure, h
MD
A c
onte
nts,
% o
f th
e co
ntro
l(a)
2
1
2
(b)
Fig. 1. The rates of lipid peroxidation in potato leaves fol-lowing plant exposure to (a) +3°C and (b) –1°C.(1) Control; (2) B33-inv plants.
Table 4. The contents of MDA in potato leaves followingchilling exposure, µmol/g fr wt
Treatment Control B33-inv plants
Without chilling 1.55 ± 0.08 1.85 ± 0.10
–9°C, 20 min 2.35 ± 0.12 2.00 ± 0.14
% of the nonchilled control
151.6 108.1
Without chilling 2.53 ± 0.13 3.22 ± 0.17
+5°C, 7 days 5.18 ± 0.21 5.05 ± 0.27
% of the nonchilled control
204.7 156.8

RUSSIAN JOURNAL OF PLANT PHYSIOLOGY Vol. 50 No. 4 2003
CHILLING TOLERANCE OF POTATO PLANTS 453
REFERENCES1. Maksimov, N.A., Izbrannye raboty po zasukhoustoichi-
vosti i zimostoikosti rastenii. T. 2. Zimostoikost’ rastenii(Selected Works on Plant Drought and Winter Tolerance,vol. 2, Plant Winter Tolerance), Moscow: Akad. NaukSSSR, 1952.
2. Tumanov, I.I., Fiziologiya zakalivaniya i morozostoiko-sti rastenii (Physiology of Hardening and Frost-Resis-tance of Plants), Moscow: Nauka, 1979.
3. Levitt, J., Responses of Plants to EnvironmentalStresses, vol. 1, Chilling, Freezing and High Tempera-ture Stresses, New York: Academic, 1980.
4. Trunova, T.I., Various Sugar Form Significance for anIncrease in the Frost Resistance of Winter CerealColeoptiles, Fiziol. Rast. (Moscow), 1963, vol. 10,pp. 588–594 (Sov. Plant Physiol., Engl. Transl.).
5. Yoshida, M., Abe, J., Moriyama, M., Shimokawa, S., andNakamura, Y., Seasonal Changes in the Physical State ofCrown Water Associated with Freezing Tolerance in Win-ter Wheat, Physiol. Plant., 1997, vol. 99, pp. 363–370.
6. Aver’yanov, A.A. and Lapikova, V.P., Interaction of Sug-ars and Hydroxyl Radicals as Related to Antifungal Tox-icity of Leaf Secretions, Biokhimiya, 1989, vol. 54,pp. 1646–1651.
7. Jeong, B.R. and Housley, T.L., Fructan Metabolism inWheat in Alternating Warm and Cold Temperatures,Plant Physiol., 1990, vol. 93, pp. 902–906.
8. Pollock, C.J. and Cairns, A.J., Fructan Metabolism inGrasses and Cereals, Annu. Rev. Plant Physiol. PlantMol. Biol., 1991, vol. 42, pp. 77–101.
9. Yukawa, T., Kobayashi, Y., Watanabe, Y., and Yamamo-to, S., Studies on Fructan Accumulation in Wheat (Triti-cum aestivum L.): 4. Fructan Accumulation under ColdTreatments and Its Varietal Difference in Relation to theActivities of Sucrose: Sucrosefructosyltransferase andFructan Exohydrolase, Jpn. J. Crop Sci., 1995, vol. 64,pp. 801–806.
10. Crespi, M.D., Zabaleta, E.J., Pontis, H.G., and Saler-no, G.L., Sucrose Synthase Expression during ColdAcclimation in Wheat, Plant Physiol., 1991, vol. 96,pp. 887–891.
11. Guy, C.L., Huber, J.L.A., and Huber, S.C., SucrosePhosphate Synthase and Sucrose Accumulation at LowTemperature, Plant Physiol., 1992, vol. 100, pp. 502–508.
12. Tymowska-Lalanne, Z. and Kreis, M., The Plant Inver-tases: Physiology, Biochemistry and Molecular Biology,Adv. Bot. Res., 1998, vol. 28, pp. 71–117.
13. Winter, H. and Huber, S.C., Regulation of SucroseMetabolism in Higher Plants: Localization and Regula-tion of Activity of Key Enzymes, Crit. Rev. Biochem.Mol. Biol., 2000, vol. 35, pp. 253–289.
14. Wyse, R., Effect of Harvest Injury on Respiration andSucrose Loss in Sugar Beet Roots during Storage, J. Am.Soc. Sugar Beet Technol., 1978, vol. 20, pp. 193–197.
15. Heineke, D., Wildenberger, K., Sonnewald, U., Will-mitzer, L., and Heldt, H.W., Accumulation of Hexoses inLeaf Vacuoles: Studies with Transgenic Tobacco PlantsExpressing Yeast-Derived Invertase in the Cytosol, Vac-uole or Apoplast, Planta, 1994, vol. 194, pp. 29–33.
16. Kolupaev, Yu.E. and Trunova, T.I., Hypothermia and SaltStress Influence on Invertase Activity and Carbohydrate
Content in Wheat Coleoptiles, Fiziol. Rast. (Moscow),1994, vol. 41, pp. 552–557 (Russ. J. Plant Physiol.,Engl. Transl.).
17. Kolupaev, Yu.E., Trunova, T.I., and Ryabchoun, N.I.,Control of Invertase Activity of Winter Wheat underStress Condition, Hortic. Veget. Growing, 1998, vol. 17,pp. 379–386.
18. Klimov, S.V., Popov, V.N., Dubinina, I.I., Burakhano-va, E.A., and Trunova, T.I., The Decreased Cold-Resis-tance of Chilling Sensitive Plants Is Related to Sup-pressed CO2 Assimilation in Leaves and Sugar Accumu-lation in Roots, Fiziol. Rast. (Moscow), 2002, vol. 49,pp. 871–877 (Russ. J. Plant Physiol., Engl. Transl.).
19. Piruzyan, E.S., Kobets, N.S., Mett, V.L., Serebriiskaya, T.S.,Neumyvakin, L.V., Alizade, Kh., Lenets, A.A., Simo-nova, M.L., Shevelukha, V.S., and Goldenkova, I.V.,Transgenic Plants Expressing Foreign Genes as a Modelfor Studying Plant Stress Responses and a Source forResistant Plant Forms, Fiziol. Rast. (Moscow), 2000,vol. 47, pp. 370–381 (Russ. J. Plant Physiol., Engl.Transl.).
20. Küzniak, E., Transgenic Plants: An Insight into Oxida-tive Stress Tolerance Mechanisms, Acta Physiol. Plant.,2002, vol. 24, pp. 97–113.
21. Bussis, D., Heineke, D., Sonnewald, U., Willmitzer, L.,Raschke, K., and Heldt, H.W., Solute Accumulation andDecreased Photosynthesis in Leaves of Potato PlantsExpressing Yeast-Derived Invertase either in the Apo-plast, Vacuole or Cytosol, Planta, 1997, vol. 202,pp. 126–136.
22. Sonnewald, U., Hajlrezaei, M.-R., Kossmann, J.,Heyer, A., Thethewey, R.N., and Willmitzer, L.,Increased Potato Tuber Size Resulting from ApoplasticExpression of a Yeast Invertase, Nature Biotech., 1997,vol. 15, pp. 794–797.
23. Rocha-Sosa, M., Sonnewald, U., Frommer, W., Strat-mann, M., Schell, J., and Willmeitzer, L., Both Develop-mental and Metabolic Signals Activate the Promoter of aClass I Patatin Gene, EMBO J., 1989, vol. 8, pp. 23–29.
24. Lukatkin, A.S. and Golovanova, V.S., Activity of LipidPeroxidation in the Chilled Leaves of Chilling-SensitivePlants, Fiziol. Rast. (Moscow), 1988, vol. 35, pp. 773–780(Sov. Plant Physiol., Engl. Transl.).
25. Turkina, N.V. and Sokolova, S.V., Methods forMonosaccharide and Oligosaccharide Determination,Biokhimicheskie metody v fiziologii rastenii (Biochemi-cal Methods in Plant Physiology), Pavlinova, O.A., Ed.,Moscow: Nauka, 1971, pp. 7–34.
26. Sturm, A., Invertases. Primary Structures, Functions,and Roles in Plant Development and Sucrose Partition-ing, Plant Physiol., 1999, vol. 121, pp. 1–8.
27. Kosegarten, H.U., Hoffmann, B., and Mengel, K., Apo-plastic pH and Fe3+ Reduction in Intact SunflowerLeaves, Plant Physiol., 1999, vol. 121, pp. 1069–1080.
28. Immobilised Cells and Enzymes. A Practical Approach,Woodwords, J., Ed., Oxford, 1985. Translated under thetitle Immobilizirovannye kletki i fermenty. Metody, Mos-cow: Mir, 1988.
29. Von Schaewen, A., Stitt, M., Schmidt, R., Sonnewald, U.,and Willmitzer, L., Expression of a Yeast-Derived Inver-tase in the Cell Wall of Tobacco and Arabidopsis PlantsLeads to Accumulation of Carbohydrate and Inhibition

454
RUSSIAN JOURNAL OF PLANT PHYSIOLOGY Vol. 50 No. 4 2003
DERYABIN et al.
of Photosynthesis and Strongly Influences Growth andPhenotype of Transgenic Tobacco Plants, EMBO J.,1990, vol. 9, pp. 3033–3044.
30. Kallarakal, J. and Komor, E., Transport of Hexoses bythe Phloem of Ricinus communis L. Seedling, Planta,1989, vol. 177, pp. 336–341.
31. Maynard, J.W. and Lucas, W.J., Sucrose and GlucoseUptake into Beta vulgaris Leaf Tissues. A Case for Gen-eral (Apoplastic) Retrieval Systems, Plant Physiol.,1982, vol. 70, pp. 1436–1443.
32. Seppanen, M.M., Majaharju, M., Somersalo, S., andPehu, E., Freezing Tolerance, Cold-Acclimation andOxidative Stress in Potato: Paraquat Tolerance Is Relatedto Acclimation but Is a Poor Indicator of Freezing Toler-ance, Physiol. Plant., 1998, vol. 102, pp. 454–460.
33. Estrada, R.N., Utilization of Wild and Cultivated DiploidPotato Species to Transfer Frost Resistance into the Tet-raploid Common Potato, Solanum tuberosum L., PlantCold Hardiness, Li, P.H. and Sakai, A., Eds., New York:Academic, 1987, vol. 5, pp. 339–353.
34. Baraboi, V.A., Mechanisms of Stress and Lipid Peroxi-dation, Usp. Sovrem. Biol., 1991, vol. 111, pp. 923–931.
35. Vladimirov, Yu.A. and Archakov, A.I., Perekisnoeokislenie lipidov v biologicheskikh membranakh (LipidPeroxidation in Biological Membranes), Moscow:Nauka, 1972.
36. Zhirov, V.K., Merzlyak, M.N., and Kuznetsov, L.V.,Membrane Lipid Peroxidation in Cold-Resistant Plantsunder Belowzero Temperatures, Fiziol. Rast. (Moscow),1982, vol. 29, pp. 1045–1052 (Sov. Plant Physiol., Engl.Transl.).
37. Merzlyak, M.N., Reactive Oxygen and Oxidative Pro-cesses in the Membranes of Plant Cells, Itogi NaukiTekh., Ser. Fiziol. Rast., 1989, vol. 6.
38. Lukatkin, A.S., Sharkaeva, E.Sh., and Zauralov, O.A.,Lipid Peroxidation in the Leaves of Heat-Loving Plantsas Dependent on the Duration of Cold Stress, Fiziol.Rast. (Moscow), 1995, vol. 42, pp. 607–611 (Russ. J.Plant Physiol., Engl. Transl.).





























