Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Climate-Growth Relationships for Native and Nonnative Pinaceae in NorthernMichigan's Pine BarrensAuthor(s): Jason S. Kilgore and Frank W. TelewskiSource: Tree-Ring Research, 60(1):3-13. 2004.Published By: Tree-Ring SocietyDOI: http://dx.doi.org/10.3959/1536-1098-60.1.3URL: http://www.bioone.org/doi/full/10.3959/1536-1098-60.1.3
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

Copyright q 2004 by the Tree-Ring Society 3
TREE-RING RESEARCH, Vol. 60(1), 2004, pp. 3–13
CLIMATE-GROWTH RELATIONSHIPS FOR NATIVE AND NONNATIVEPINACEAE IN NORTHERN MICHIGAN’S PINE BARRENS
JASON S. KILGORE* and FRANK W. TELEWSKI
W. J. Beal Botanical GardenDepartment of Plant Biology
Michigan State UniversityEast Lansing, MI 48824
ABSTRACT
Secondary growth responses of native and nonnative trees exposed to the same climatic conditionscan elucidate sensitivities and thus adaptability to a particular region. A long-term mixed-species plantingin the pine barrens of northern lower Michigan presented an opportunity to discriminate responses fromspecies commonly planted in this region. Mean ring-width chronologies from living native Pinus resinosaAit. and P. strobus L. and nonnative P. sylvestris L. and Picea abies (L.) Karst. at this plantation weregenerated, standardized, and analyzed by correlation analysis against mean monthly climatic variables. Thenative pine chronologies had the highest mean ring widths and signal-to-noise ratios (SNR), were highlycorrelated to each other, and exhibited positive responses to years with above-normal April temperaturesbut no significant relationships to variations in precipitation. The P. sylvestris chronology was highlycorrelated to the other two pine chronologies and responded similarly to April temperatures but exhibitednegative correlations to January and April precipitation and positive correlations to September precipitation.The P. abies chronology had the highest mean sensitivity and was correlated with the P. strobus chronologybut only responded positively to precipitation from the previous December. The low SNR (P. sylvestris, P.abies), high mean sensitivity (P. abies), and larger number of significant correlations to variations inmonthly climatic variables (P. sylvestris) suggest that these nonnative species are more sensitive to thislocal climate. These results provide insights to the adaptability, establishment, and geographic distributionof the nonnative Pinaceae.
Keywords: growth response, native, exotic, pine barrens, dendroecology, Michigan.
INTRODUCTION
The response of a species to the spatial andtemporal variation in resources, as a function ofthe physical environment, natural enemies, andcompetitors, is fundamental to its success in a giv-en community (Shea and Chasson 2002). This ap-proach to understanding niche opportunities (Sheaand Chesson 2002) provides an avenue for inves-tigating the ability of nonnative species to estab-lish and possibly even invade native communities.Response to climatic variation is traditionally con-sidered a primary determinant of biogeographicalrange for a particular species. Although the meanclimate regulates species establishment and surviv-al, the extreme climatic events (Cook et al. 1987),
*Corresponding Author: [email protected].
such as early and late-season frosts, extreme win-ter temperatures, and droughts, affect growth andsuccess in regeneration and therefore restrict thelong-term species pool (Woodward 1987). Givenspecies with similar ecological tolerances andfunction but different native biogeographical rang-es, those species that are best able to exploit theavailable resources will be most successful whengrown together. This has important implications toassessing the ability of a nonnative species to growand ultimately invade, via successful regeneration,a community dominated by native species withsimilar fundamental niches. Dendrochronologyprovides a useful tool to measure the tree’s abilityto take advantage of local resources. The limitingclimatic factors to growth (Fritts 1966, 1976; LaMarche 1978; Fritts and Swetnam 1989) for a treespecies provide information for assessing the suit-

4 KILGORE and TELEWSKI
ability of that species to its particular environmentand also provide baseline climatic response infor-mation for longer-term climatic change studies(e.g. Fritts 1991).
Following the 19th Century wildfires in thepine barrens of northern Michigan, native Pinusresinosa, P. strobus, and P. banksiana Lamb. andnonnative P. sylvestris were planted extensivelyand the nonnative Picea abies was planted inter-mittently. Many of these plantations are now beingharvested, but many stands of the nonnative Pin-aceae remain. All of these species share a similarfundamental niche (Burns and Honkala 1990). Be-cause both Picea abies and Pinus sylvestris arecapable of regenerating naturally in this ecosystem(Kilgore 2002), these nonnative species may beinvasive under particular conditions. Consequent-ly, investigating their dendrochronological re-sponse to the current climate regime will providea baseline for assessing reactions to climate changeand will lead to greater predictability of their po-tential for long-term establishment. All of the spe-cies are expected to express sensitivity in their ra-dial growth in response to the droughty, infertilesoil and cold continental climate in the region(Whitney 1986), but particular sensitivities mayindicate climatic limitations to geographic rangefor the nonnative species.
The purpose of this study is to assess the cli-matic sensitivity, or exploitation, by native andnonnative Pinaceae in northern Michigan’s pinebarrens through the influence of climate on radialgrowth. No other study has examined the climaticresponse for these species in the pine barren eco-system of the Great Lakes region, and the use ofclimatic response information has not been used toassess potential for invasiveness in the Pinaceae.Because of the short growing season and limitingsoil conditions, we predicted that radial growth forall species would be restricted by cool early andlate growing season temperatures and dry sum-mers. Although pine barrens fall within the eco-logical range for all of these species, we predictedthat the native species would be more complacentto variations in climate because their seed sourcewould more likely originate from a locally adaptedpopulation. A historically important plantation ofPinaceae located in the pine barrens of northern
lower Michigan was studied to address these pre-dictions.
METHODS
Study Site
The regional setting for this study was thepine barrens of the Midwest, which were formedupon the thick deposits of glacial outwash, largelycomposed of coarse soils, laid down 12000 yearsago at the end of the Pleistocene glaciation (Whit-ney 1986; Werlein 1998). This region is now cov-ered by at least second-growth upland pine forestsand barrens growing on sandy soils interspersedwith bogs and conifer swamps bordering formermeltwater channels. The pine forests are dominat-ed by Pinus strobus and P. resinosa, specieswhose native ranges overlap (Figure 1). The bar-rens are dominated by P. banksiana with scatteredP. resinosa (Curtis 1971; Whitney 1986, 1987;Comer 1996). In addition to these native pines, thenonnative P. sylvestris has been planted in the pinebarrens for timber and Christmas tree production(Burns and Honkala 1990), and the nonnative Pi-cea abies is planted largely for windbreaks. Thesandy soils of the barrens contain low nutrients,organic matter, and water-holding capacity (Mok-ma and Vance 1989).
The study site is the Grayling Beal Plantation(GBP) located southeast of the town of Graylingin Crawford County, Michigan (44839 9N,848429W, 346 m a.s.l.; Figure 1). In 1888–1889,seedlings, cuttings, and seed (Pinus rigida Mill.only) from 41 native and nonnative tree specieswere planted in a 5-acre (2.0 ha) sandy field todetermine which species would grow best for re-forestation and production (Beal 1888; Kilgore2002). Prior to planting, the site experienced fre-quent fires leaving scattered P. resinosa and P.banksiana, scrubby Quercus spp., and dwarf Pru-nus spp. (Kedzie 1888), but no record or evidenceof fire exists at this site since planting. The topog-raphy is level, and the homogenous soil (M.Kroell, personal communicaton) is mapped as acoarse, loose, strongly acidic, and excessivelydrained sand of the Grayling series (Typic, FrigidUdipsamment) (Mokma and Vance 1989; Werlein1998). Although the depth to the water table was

5Native and Nonnative Pinaceae in Michigan
Figure 1. Map of North America showing the native distribution (redrawn from Little 1971) of Pinus resinosa and P. strobusand the Grayling Beal Plantation study site in northern lower Michigan.
not measured in this study, Werlein (1998) notesthe water table to be . 6 ft (1.83 m) in this soiltype. Government Land Office Survey recordsshow that pre-European settlement forests on thissoil type were largely composed of Pinus banksi-ana (Whitney 1986).
Sample Collection and Data Analysis
All of the surviving planted stems from thenonnative Picea abies, native Pinus resinosa, andnative P. strobus and all of the large diameter re-generating stems from nonnative P. sylvestris weresampled to generate mean chronologies (Table 1);no other species, including P. banksiana that wasnot planted, had surviving or regenerating stemsin sufficient numbers to analyze (Kilgore 2002).Trees were cored perpendicular to the stem lean toavoid reaction wood with 5.15-mm diameter in-crement borers in 1998–2000. One full-diametercore or two opposite radial cores were obtained at
approximately 0.3 m above the ground to obtainas many growth rings as possible. Increment coreswere prepared, crossdated, and measured to 0.01mm using standard dendrochronological tech-niques (Stokes and Smiley 1996). Ring-widthmeasurements within each time series were com-puter-analyzed to detect anomalies (COFECHAVersion 6.06P, Holmes 2000). Mean raw ringwidths were compared across species using theconservative Tukey HSD Multiple Comparisonprocedure (Systat Version 9.01, SPSS Science, Inc.1998), which allows comparisons among meanswith varying sample sizes (Sokal and Rohlf 1995).
All series of raw data were converted to meanchronologies for climate response analysis usingthe same program settings to ensure consistentcomparisons among species and sites. Each serieswas detrended with a single-pass high-frequencyfilter, i.e. a negative exponential or negative linearregression was fit to the series (ARSTAN Version6.04P, Holmes 2000). This conservative technique

6 KILGORE and TELEWSKI
Table 1. Dendrochronological characteristics for the raw ring width data and ARSTAN (ARS) mean chronology for each of thePinaceae at the Grayling Beal Plantation study site. All values calculated by ARSTAN (Holmes 2000).
Characteristics Picea abies Pinus resinosa Pinus strobus Pinus sylvestris
No. treesNo. coresTotal yearsMean ring width (mm)1
SNR (detrended)2
1125
1051.788b3.697
2282
1012.177a9.818
1021
1052.167ab5.949
1020851.896ab1.131
%PC1 (detrended)3
Mean index (ARS)Mean sensitivity (ARS)Std. deviation (ARS)Skewness (ARS)Kurtosis (ARS)
43.61.00630.25690.2511
20.30410.7971
36.41.04460.13300.36462.16106.8889
49.50.98630.18100.2232
20.20520.2696
35.60.96030.13490.23890.46110.9911
Partial Autocorrelation (ARS)
1st order2nd order3rd order
Min sample size (no. trees)4
0.131820.062320.119710
0.802120.030420.076513
0.504920.012120.1053
6
0.739920.1707
0.044338
1Mean ring width values with different letters across species are significantly different (Tukey HSD, p , 0.01).2Signal-to-noise ratio (SNR) based on detrended series.3Percentage of variation accounted for by the first principal component of the correlation matrix of individual detrended series(Graumlich 1993).4Minimum sample size based on a signal strength threshold of 0.85 (Briffa and Jones 1990).
should remove exogenous variation and the age-related growth trend while retaining the climati-cally induced high-frequency variation (Fritts1976; Cook et al. 1990). Of the three types ofmean chronologies (STANDARD, RESID, andARSTAN [ARSTAN Version 6.04P, Holmes2000]) constructed from the detrended series, theARSTAN chronology expresses the greatest sen-sitivity to climatic variations (Cook and Holmes1997) and was thus used exclusively for the den-droclimatic analyses. Standard chronology statis-tics, such as signal-to-noise ratio (SNR), meansensitivity (MS), partial autocorrelation coeffi-cients, and the percentage of intraseries variationexplained by the first principal component(%PC1), were calculated by the program AR-STAN Version 6.04P (Holmes 2000). BecauseSNR is influenced by sample size, %PC1 is a bet-ter measure of variance within each series and canbe compared across chronologies (Graumlich1993). Similar temporal response patterns amongthe ARSTAN chronologies constructed for eachspecies were compared by simple correlation anal-
ysis (Pearson correlation, Bonferroni probability,Systat Version 9.01, SPSS Science, Inc. 1998).
The relative importance of monthly precipi-tation and monthly mean temperature in influenc-ing tree-ring growth for each species was deter-mined by correlation and response function anal-ysis (RESPO Version 6.06P, Holmes 2000). Cor-relation functions are the correlation coefficientsbetween the climatic variables and mean chronol-ogies, whereas the response function is a result ofa multivariate regression analysis on a set of prin-cipal components (PCs) calculated from climaticvariables (Fritts 1976). PCs enter the stepwise re-gression if the cumulative multiple of eigenvalueis greater than one, thus the number of PCs re-tained varies by chronology (RESPO Version6.06P, Holmes 2000). Although response functionanalysis has been shown to be highly effective atshowing relationships between climate and growth(Fritts and Wu 1986), the response function can bemore difficult to replicate and interpret (Blasing etal. 1984). Caution must be given regarding the in-terpretation of the direction of correlation coeffi-
7Native and Nonnative Pinaceae in Michigan
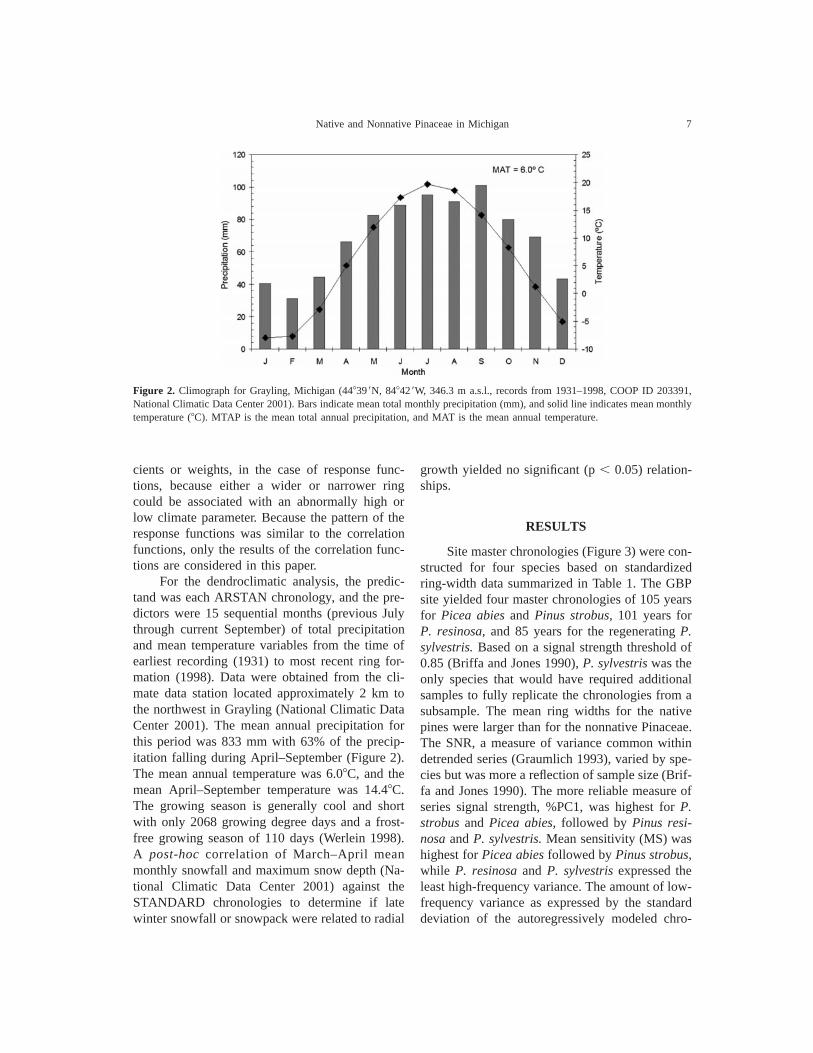
Figure 2. Climograph for Grayling, Michigan (448399N, 848429W, 346.3 m a.s.l., records from 1931–1998, COOP ID 203391,National Climatic Data Center 2001). Bars indicate mean total monthly precipitation (mm), and solid line indicates mean monthlytemperature (8C). MTAP is the mean total annual precipitation, and MAT is the mean annual temperature.
cients or weights, in the case of response func-tions, because either a wider or narrower ringcould be associated with an abnormally high orlow climate parameter. Because the pattern of theresponse functions was similar to the correlationfunctions, only the results of the correlation func-tions are considered in this paper.
For the dendroclimatic analysis, the predic-tand was each ARSTAN chronology, and the pre-dictors were 15 sequential months (previous Julythrough current September) of total precipitationand mean temperature variables from the time ofearliest recording (1931) to most recent ring for-mation (1998). Data were obtained from the cli-mate data station located approximately 2 km tothe northwest in Grayling (National Climatic DataCenter 2001). The mean annual precipitation forthis period was 833 mm with 63% of the precip-itation falling during April–September (Figure 2).The mean annual temperature was 6.08C, and themean April–September temperature was 14.48C.The growing season is generally cool and shortwith only 2068 growing degree days and a frost-free growing season of 110 days (Werlein 1998).A post-hoc correlation of March–April meanmonthly snowfall and maximum snow depth (Na-tional Climatic Data Center 2001) against theSTANDARD chronologies to determine if latewinter snowfall or snowpack were related to radial
growth yielded no significant (p , 0.05) relation-ships.
RESULTS
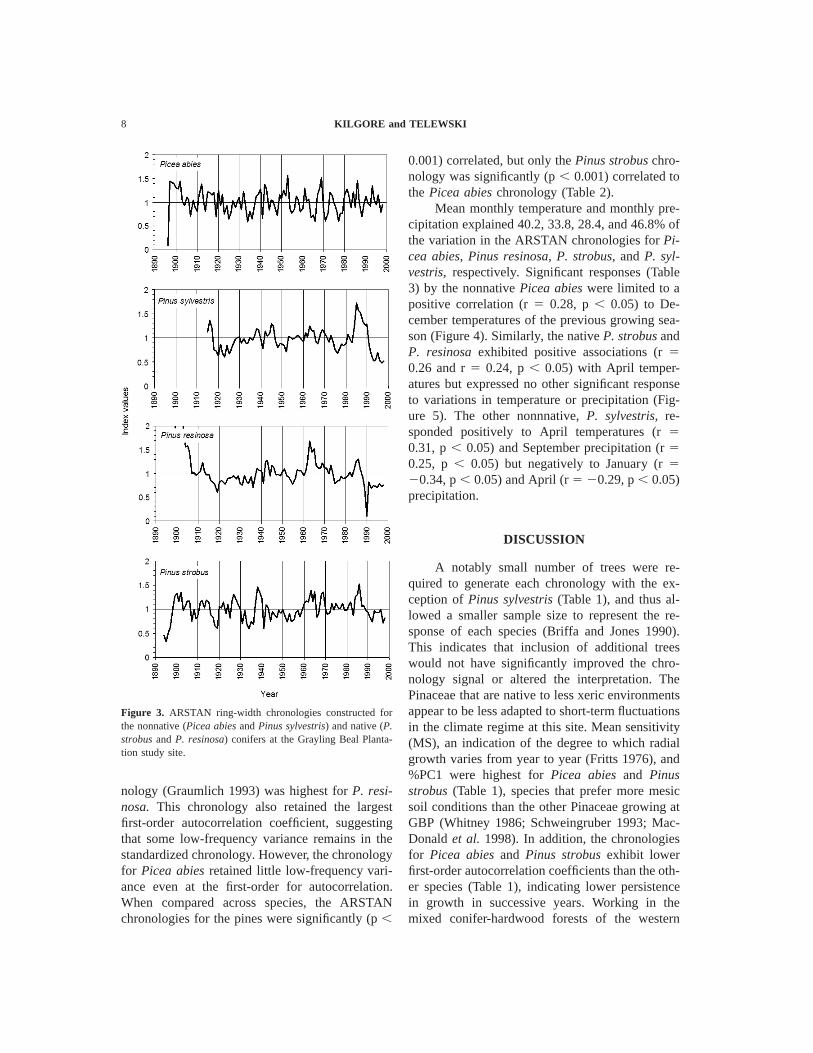
Site master chronologies (Figure 3) were con-structed for four species based on standardizedring-width data summarized in Table 1. The GBPsite yielded four master chronologies of 105 yearsfor Picea abies and Pinus strobus, 101 years forP. resinosa, and 85 years for the regenerating P.sylvestris. Based on a signal strength threshold of0.85 (Briffa and Jones 1990), P. sylvestris was theonly species that would have required additionalsamples to fully replicate the chronologies from asubsample. The mean ring widths for the nativepines were larger than for the nonnative Pinaceae.The SNR, a measure of variance common withindetrended series (Graumlich 1993), varied by spe-cies but was more a reflection of sample size (Brif-fa and Jones 1990). The more reliable measure ofseries signal strength, %PC1, was highest for P.strobus and Picea abies, followed by Pinus resi-nosa and P. sylvestris. Mean sensitivity (MS) washighest for Picea abies followed by Pinus strobus,while P. resinosa and P. sylvestris expressed theleast high-frequency variance. The amount of low-frequency variance as expressed by the standarddeviation of the autoregressively modeled chro-
8 KILGORE and TELEWSKI
Figure 3. ARSTAN ring-width chronologies constructed forthe nonnative (Picea abies and Pinus sylvestris) and native (P.strobus and P. resinosa) conifers at the Grayling Beal Planta-tion study site.
nology (Graumlich 1993) was highest for P. resi-nosa. This chronology also retained the largestfirst-order autocorrelation coefficient, suggestingthat some low-frequency variance remains in thestandardized chronology. However, the chronologyfor Picea abies retained little low-frequency vari-ance even at the first-order for autocorrelation.When compared across species, the ARSTANchronologies for the pines were significantly (p ,
0.001) correlated, but only the Pinus strobus chro-nology was significantly (p , 0.001) correlated tothe Picea abies chronology (Table 2).
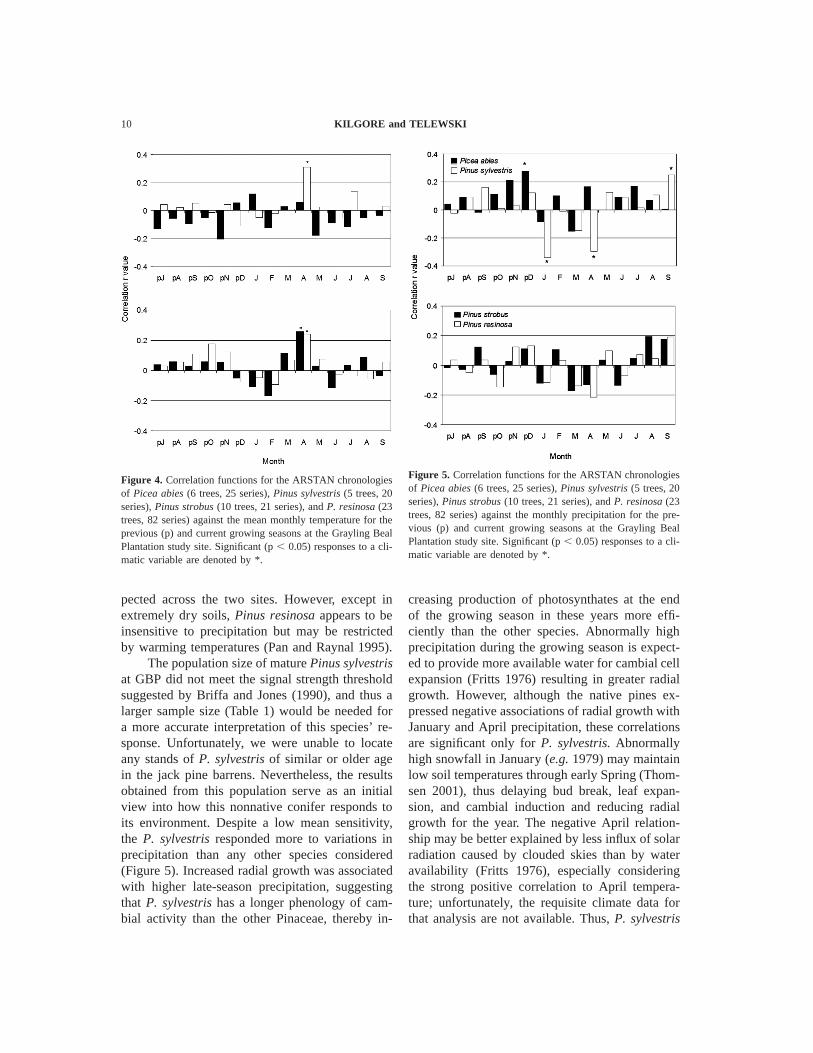
Mean monthly temperature and monthly pre-cipitation explained 40.2, 33.8, 28.4, and 46.8% ofthe variation in the ARSTAN chronologies for Pi-cea abies, Pinus resinosa, P. strobus, and P. syl-vestris, respectively. Significant responses (Table3) by the nonnative Picea abies were limited to apositive correlation (r 5 0.28, p , 0.05) to De-cember temperatures of the previous growing sea-son (Figure 4). Similarly, the native P. strobus andP. resinosa exhibited positive associations (r 50.26 and r 5 0.24, p , 0.05) with April temper-atures but expressed no other significant responseto variations in temperature or precipitation (Fig-ure 5). The other nonnnative, P. sylvestris, re-sponded positively to April temperatures (r 50.31, p , 0.05) and September precipitation (r 50.25, p , 0.05) but negatively to January (r 520.34, p , 0.05) and April (r 5 20.29, p , 0.05)precipitation.
DISCUSSION
A notably small number of trees were re-quired to generate each chronology with the ex-ception of Pinus sylvestris (Table 1), and thus al-lowed a smaller sample size to represent the re-sponse of each species (Briffa and Jones 1990).This indicates that inclusion of additional treeswould not have significantly improved the chro-nology signal or altered the interpretation. ThePinaceae that are native to less xeric environmentsappear to be less adapted to short-term fluctuationsin the climate regime at this site. Mean sensitivity(MS), an indication of the degree to which radialgrowth varies from year to year (Fritts 1976), and%PC1 were highest for Picea abies and Pinusstrobus (Table 1), species that prefer more mesicsoil conditions than the other Pinaceae growing atGBP (Whitney 1986; Schweingruber 1993; Mac-Donald et al. 1998). In addition, the chronologiesfor Picea abies and Pinus strobus exhibit lowerfirst-order autocorrelation coefficients than the oth-er species (Table 1), indicating lower persistencein growth in successive years. Working in themixed conifer-hardwood forests of the western

9Native and Nonnative Pinaceae in Michigan
Table 2. Pearson correlation coefficients (r) and significance (Bonferroni probability) between the ARSTAN master chronologiesdeveloped for each species at the Grayling Beal Plantation study site. All values calculated by Systat Version 9.01 (SPSS Science,Inc. 1998).
Picea abies Pinus resinosa Pinus strobus Pinus sylvestris
Picea abiesPinus resinosaPinus strobusPinus sylvestris
1.0000.2340.408***0.313
1.0000.594****0.583****
1.0000.429*** 1.000
*p , 0.05, **p , 0.01, ***p , 0.001, ****p , 0.0001.
Table 3. Significant (p , 0.05) correlations between ARSTAN master chronologies and monthly climate variables at the GraylingBeal Plantation study site. All values calculated by RESPO Version 6.06P (Holmes 2000).
Species Correlation Variable
Picea abiesPinus resinosaPinus strobusPinus sylvestris
0.27770.23920.25700.3113
Total prior December precipitationMean April temperatureMean April temperatureMean April temperature
20.340320.2970
0.2519
Total January precipitationTotal April precipitationTotal September precipitation
Great Lakes, Graumlich (1993) found similar re-sults for P. strobus (MS 5 0.18, %PC1 5 46.3)and P. resinosa (MS 5 0.16, %PC1 5 33.9). Datacollected by Fritts (DeWitt and Ames 1978) for P.resinosa in northern Minnesota indicated similarmean sensitivities (0.18, 0.11, and 0.21). A studyby Koop (1985) of old-growth P. resinosa grow-ing at a dry mesic site (Hartwick Pines State Park,HRP) approximately 10 km north of GBP found amean sensitivity (ARSTAN chronology) of 0.15.Because radial growth is affected by climate (Fritts1976), higher mean sensitivity gives an indicationof the strength of dependence on a limiting fac-tor(s) having high-frequency variance, presumablyclimate. Consequently, allocation to secondary ra-dial growth in Picea abies appears to be more sen-sitive to available resources and subsequent pho-tosynthate supply in the short-term at this locationthan for the other species.
The chronologies for the native Pinus strobusand P. resinosa were significantly correlated tovariations in April temperatures but showed nosignificant relationships to any other climatic var-iable examined in this study. Because of their ev-ergreen foliage and large earlywood xylem, coni-
fers are able to take advantage of warm tempera-tures early in the growing season, thus a largergrowth ring is formed (Fritts 1976). Koop (1985)examined only the current year’s climatic param-eters with a P. resinosa chronology (RESID):April (r 5 0.22, p , 0.05), June (r 5 20.34, p, 0.005), and July (r 5 20.29, p , 0.05) tem-peratures and June precipitation (r 5 0.218, p ,0.05) were significant. The difference in Summerprecipitation responses could be attributable to theuse of a different number of climatic parametersin the analysis. Furthermore, although GBP is arelatively even-aged closed canopy forest on levelground, the crowns of the uneven-aged trees atHRP are more exposed to solar radiation and wind,and the roots are presumably further from the soilwater because they grow along a ridge (Koop1985). Consequently, these trees may suffer great-er water stress through evaporative water loss andhave less access to soil water than those trees atGBP. In addition, the pines at HRP are larger andolder, and thus have a greater overall respiratorydemand and are less physiologically robust (Koz-lowski 1971). A differential response to variationin temperature and precipitation would thus be ex-
10 KILGORE and TELEWSKI
Figure 4. Correlation functions for the ARSTAN chronologiesof Picea abies (6 trees, 25 series), Pinus sylvestris (5 trees, 20series), Pinus strobus (10 trees, 21 series), and P. resinosa (23trees, 82 series) against the mean monthly temperature for theprevious (p) and current growing seasons at the Grayling BealPlantation study site. Significant (p , 0.05) responses to a cli-matic variable are denoted by *.
Figure 5. Correlation functions for the ARSTAN chronologiesof Picea abies (6 trees, 25 series), Pinus sylvestris (5 trees, 20series), Pinus strobus (10 trees, 21 series), and P. resinosa (23trees, 82 series) against the monthly precipitation for the pre-vious (p) and current growing seasons at the Grayling BealPlantation study site. Significant (p , 0.05) responses to a cli-matic variable are denoted by *.
pected across the two sites. However, except inextremely dry soils, Pinus resinosa appears to beinsensitive to precipitation but may be restrictedby warming temperatures (Pan and Raynal 1995).
The population size of mature Pinus sylvestrisat GBP did not meet the signal strength thresholdsuggested by Briffa and Jones (1990), and thus alarger sample size (Table 1) would be needed fora more accurate interpretation of this species’ re-sponse. Unfortunately, we were unable to locateany stands of P. sylvestris of similar or older agein the jack pine barrens. Nevertheless, the resultsobtained from this population serve as an initialview into how this nonnative conifer responds toits environment. Despite a low mean sensitivity,the P. sylvestris responded more to variations inprecipitation than any other species considered(Figure 5). Increased radial growth was associatedwith higher late-season precipitation, suggestingthat P. sylvestris has a longer phenology of cam-bial activity than the other Pinaceae, thereby in-
creasing production of photosynthates at the endof the growing season in these years more effi-ciently than the other species. Abnormally highprecipitation during the growing season is expect-ed to provide more available water for cambial cellexpansion (Fritts 1976) resulting in greater radialgrowth. However, although the native pines ex-pressed negative associations of radial growth withJanuary and April precipitation, these correlationsare significant only for P. sylvestris. Abnormallyhigh snowfall in January (e.g. 1979) may maintainlow soil temperatures through early Spring (Thom-sen 2001), thus delaying bud break, leaf expan-sion, and cambial induction and reducing radialgrowth for the year. The negative April relation-ship may be better explained by less influx of solarradiation caused by clouded skies than by wateravailability (Fritts 1976), especially consideringthe strong positive correlation to April tempera-ture; unfortunately, the requisite climate data forthat analysis are not available. Thus, P. sylvestris

11Native and Nonnative Pinaceae in Michigan
could be taking advantage of the local resources,including climate, by extending its phenology, re-sulting in greater biomass accumulation, or is clos-er to the center of its ecological range than the twonative pines (Figure 1). If P. sylvestris is moreadapted than the native pines to the local environ-ment, as has been observed by others in NorthAmerica (Pan and Raynal 1995), then the appli-cation of dendrochronological techniques may beuseful in identifying the beneficial environmentalfactor that may be supporting the establishmentand success of the species. However, the smallsample size, low SNR, and low %PC1 for this spe-cies necessitates concern over the interpretation ofthese results. In any case, the broad ecological dis-tribution (Burns and Honkala 1990) of P. sylves-tris and its potential ability to take advantage oflocal climate conditions supports the notion that P.sylvestris is at least a successful pioneer species(Gutierrez 1989).
The chronology for Picea abies expressed norelationship to variations in temperature, unlike theother Pinaceae, but a significant positive responseto precipitation in the December preceding thegrowing season. Sensitivity in Picea abies to con-ditions during the previous growing season hasbeen confirmed when growing within its nativeEuropean range (Makinen et al. 2001) and towarmer winters in an Adirondack Mountain plan-tation (Pan and Raynal 1995). Although speciesliving at the edge of their range express the great-est sensitivity to limiting factors like climate (Fritts1976), the geographic range for P. abies includesmuch colder and more mesic environments(Schweingruber 1993) than Grayling; the physicalfeatures (e.g. short needles and fastigiate shape) ofthe P. abies at GBP resemble the more northernraces of the species (Telewski, personal observa-tion). As a result of this species’ strong commonvariance (%PC1), relatively high MS, but low cor-relation to climatic variables during the growingseason, the P. abies trees are neither limited bynor exploiting the climatic resources at this site.
SUMMARY AND CONCLUSIONS
The use of dendrochronological techniques tocompare the high frequency radial growth re-
sponse of native and nonnative species to climaticvariables may elucidate ecological tolerances, orrange limits, and consequently adaptability, in par-ticular environments. Radial growth is more thana simple response to climate; the particular envi-ronmental conditions, including temperature, soilnutrients and available water, disturbances, patho-gens, and insects, affect the ultimate product ofannual xylem formation. When several species areplanted and grown in a homogeneous environ-ment, such as at the GBP, the exposure to restric-tive growing conditions is approximately uniform.Furthermore, pines have been shown to havegreater species-specific responses across differentgrowing conditions than interspecific variationwithin a climate regime (Graumlich 1993). Con-sequently, comparisons of growth responses acrossspecies at one location are legitimate. Unfortu-nately, few have investigated the dendroclimaticresponses for these species in North America, es-pecially in the Great Lakes region. Comparisonsof the results from this study to other studies areaccordingly limited.
In this study, the nonnative Pinus sylvestrisand Picea abies demonstrated more differential re-sponse to climatic variations than did the nativePinus resinosa and P. strobus. By extending itsphenology in years of exceptional climatic re-sources, P. sylvestris may be gaining advantage inits growth in the pine barrens ecosystem. Piceaabies does not appear to be affected by any vari-ations in growing season climate yet is highly sen-sitive to current year’s photosynthate production.Examining these climatic relationships for thesespecies at other pine barren sites should supportthe application of dendrochronology in the field ofinvasion ecology.
ACKNOWLEDGMENTS
The authors appreciate the field assistancefrom Pam Kilgore, Robyn Ast, Steve Nimcheski,Steve Lettero, Denise Kemp, and Alex Zornig.Chris Baisan, Rex Adams, Tom Harlan, and Har-old Fritts kindly spent time discussing this projectwith the authors, and Marty Kroell provided soilinformation. We also thank Steven Leavitt, YvesBegin, and two anonymous reviewers for com-

12 KILGORE and TELEWSKI
ments that greatly improved earlier versions of thismanuscript. This research was supported by agrant from Sigma Xi (The Scientific Research So-ciety), with logistical support provided by the Di-vision of Campus Park and Planning at MichiganState University.
REFERENCES CITED
Beal, W. J.1888 Report of the Botanist of the Experiment Station. In
Twenty-Seventh Annual Report of the Secretary of theState Board of Agriculture of the State of Michigan,1888, Thorp and Godfrey, State Printers and Binders,Lansing; pp. 171–193.
Blasing, T. J., A. M. Solomon, and D. N. Duvick1984 Response functions revisited. Tree-Ring Bulletin 44:
1–15.Briffa, K., and P. D. Jones
1990 Basic chronology statistics and assessment. In Meth-ods of Dendrochronology: Applications in the Envi-ronmental Sciences, edited by E. R. Cook and L. A.Kairiukstis, Kluwer Academic Publishers, Dordrecht;pp. 137–152.
Burns, R. M., and B. H. Honkala1990 Silvics of North America: 1. Conifers. Agricultural
Handbook 654. US Department of Agriculture, For-est Service, Washington, D.C.
Comer, P. J.1996 Natural Community Abstract for Pine Barrens. Mich-
igan Natural Features Inventory, Lansing.Cook, E. R., A. H. Johnson, and T. J. Blasing
1987 Forest decline: modeling the effect of climate in treerings. Tree Physiology 3:27–40.
Cook, E. R., K. R. Briffa, S. G. Shiyatov, and V. Mazepa1990 Tree-ring standardization and growth-trend estima-
tion. In Methods of Dendrochronology: Applicationsin the Environmental Sciences, edited by E. R. Cookand L. A. Kairiukstis, Kluwer Academic Publishers,Dordrecht; pp. 104–123.
Cook, E. R., and R. L. Holmes1997 ARSTAN: chronology development. In Documenta-
tion to the International Tree-Ring Data Bank Pro-gram Library Version 2.1, edited by H. D. Grissino-Mayer, R. L. Holmes, and H. C. Fritts, URL: http://tree.ltrr.arizona.edu/software.html; pp. 75–87.
Curtis, J. T.1971 The Vegetation of Wisconsin: An Ordination of Plant
Communities. University of Wisconsin Press, Madi-son.
DeWitt, E., and M. Ames, Editors1978 Tree-Ring Chronologies of Eastern North America.
The University of Arizona, Tucson.Fritts, H. C.
1966 Growth rings of trees: Their correlation with climate.Science 154:973–979.
Fritts, H. C.1976 Tree Rings and Climate. Academic Press, New York.
Fritts, H.C.1991 Reconstructing Large-Scale Climatic Patterns from
Tree-Ring Data. The University of Arizona Press,Tucson.
Fritts, H. C., and T. W. Swetnam1989 Dendroecology: A tool for evaluating variations in
past and present forest environments. Advances inEcological Research 19:111–188.
Fritts, H. C., and X. Wu1986 A comparison between response-function analysis
and other techniques. Tree-Ring Bulletin 46:31–46.Graumlich, L. J.
1993 Response of tree growth to climatic variation in themixed conifer and deciduous forest of the UpperGreat Lakes region. Canadian Journal of Forest Re-search 23:133–143.
Gutierrez, E.1989 Dendroclimatological study of Pinus sylvestris L. in
southern Catalonia (Spain). Tree-Ring Bulletin 46:1–9.
Holmes, R.2000 Dendrochronology Program Library. URL: ftp://
ftp.cricyt.edu.ar/users/dendro/.Kedzie, R. C.
1888 The jack pine plains, Bulletin No. 37—Chemical De-partment. In Twenty-Seventh Annual Report of theSecretary of the State Board of Agriculture of theState of Michigan, 1888, Thorp and Godfrey, StatePrinters and Binders, Lansing; pp. 207–211.
Kilgore, J. S.2002 An Historical and Dendroecological Analysis of a
Long-term Reforestation Experiment in Grayling,Michigan. M.S. thesis, Michigan State University,East Lansing.
Koop, D. L.1985 Reconstruction of Past Climate in Northern Lower
Michigan Through the Analysis of Red Pine (Pinusresinosa Ait.) Tree Rings. M.S. thesis, University ofGeorgia, Athens.
Kozlowski, T. T.1971 Growth and Development of Trees, Volume I: Seed
Germination, Ontogeny, and Shoot Growth. Academ-ic Press, New York.
La Marche, V. C, Jr.1978 Tree-ring evidence of past climatic variability. Nature
276(5686):334–338.Little, E. L., Jr.
1971 Atlas of United States Trees: Volume 1. Conifers andImportant Hardwoods. Miscellaneous Publication1146. US Department of Agriculture, Forest Service,Washington, D.C.
MacDonald, G. M., L. C. Cwynar, and C. Whitlock1998 The late Quaternary dynamics of pines in northern
North America. In The Ecology and Biogeography ofPinus, edited by D. M. Richardson, Cambridge Uni-versity Press, Cambridge; pp. 122–136.

13Native and Nonnative Pinaceae in Michigan
Makinen, H., P. Nojd, and K. Mielikainen2001 Climatic signal in annual growth variation in dam-
aged and healthy stands of Norway spruce [Piceaabies (L.) Karst.] in southern Finland. Trees 15:177–185.
Mokma, D. L., and G. F. Vance1989 Forest vegetation and origin of some spodic horizons,
Michigan. Geoderma 43:311–324.National Climatic Data Center
2001 Monthly climate data for Grayling (COOP ID203391). U.S. Department of Commerce, NationalOceanic and Atmospheric Administration, and Na-tional Environmental Satellite, Data and InformationService. URL: http://lwf.ncdc.noaa.gov/oa/ncdc.html.
Pan, Y., and D. J. Raynal1995 Predicting growth of plantation conifers in the Adi-
rondack Mountains in response to climate change.Canadian Journal of Forest Research 25:48–56.
Schweingruber, F. H.1993 Trees and Wood in Dendrochronology: Morpholog-
ical, Anatomical, and Tree-Ring Analytical Charac-teristics of Trees Frequently Used in Dendrochro-nology. Springer-Verlag, Berlin.
Shea, K., and P. Chesson2002 Community ecology theory as a framework for bio-
logical invasions. TRENDS in Ecology and Evolution17(4):170–176.
Sokal, R. R., and F. J. Rohlf1995 Biometry, 3rd ed. W.H. Freeman and Company, New
York.SPSS Science, Inc.
1998 Systat Version 9.01. SPSS Science, Inc., Chicago.Stokes, M., and T. Smiley
1996 An Introduction to Tree-Ring Dating. The Universityof Arizona Press, Tucson.
Thomsen, G.2001 Response to winter precipitation in ring width chro-
nologies of Pinus sylvestris L. from the northwesternSiberian Plain, Russia. Tree-Ring Research 57(1):15–29.
Werlein, J. O.1998 Soil Survey of Crawford County, Michigan. U.S. De-
partment of Agriculture, Natural Resources Conser-vation Service, Washington, D.C.
Whitney, G. G.1986 Relation of Michigan’s presettlement pine forests to
substrate and disturbance history. Ecology 67(6):1548–1559.
Whitney, G. G.1987 An ecological history of the Great Lakes forests of
Michigan. Journal of Ecology 75:667–684.Woodward, F. I.
1987 Climate and Plant Distribution. Cambridge Univer-sity Press, Cambridge.
Received 23 September 2002; accepted 27 September 2003.




























