Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 92, pp. 2328-2332, March 1995Biochemistry
Cloning, expression, and characterization of cDNAs encodingArabidopsis thaliana squalene synthase
(plant/sterol biosynthesis/recombinant protein/dehydrosqualene)
TAKAYUKI NAKASHIMA*t, TAKAYUKI INOUE*t, ATSUHIRO OKA§, TOKUZO NISHINOT, TAKASHI OSUMI*,AND SHINGO HATA*II*Faculty of Science, Himeji Institute of Technology, Ako-gun, Hyogo 678-12, Japan; §Institute for Chemical Research, Kyoto University, Uji, Kyoto 611, Japan;and 1Department of Biochemistry and Engineering, Tohoku University, Sendai, Miyagi 980, Japan
Communicated by David B. Sprinson, St. Luke's/Roosevelt Hospital Center, New York NY October 14, 1994 (received for review August 23, 1994)
ABSTRACT We have isolated and characterized two over-lapping cDNA clones for Arabidopsis thaliana squalene syn-thase. Their nucleotide sequences contained an open readingframe for a 410-amino acid polypeptide (calculated molecularmass, 47 kDa). The deduced amino acid sequence of theArabidopsis polypeptide was significantly homologous (42-44% identical) to the sequences of known squalene synthasesof several species, from yeast to man, but it was much lesshomologous to that of tomato phytoene synthase. To expressthe Arabidopsis enzyme in Escherichia coli, the entire codingregion was subcloned into an expression vector. A cell-freeextract of E. coli transformed with the recombinant plasmid,in the presence of NADPH and Mg2+, efficiently converted['4C]farnesyl diphosphate into squalene. On the other hand,in the absence of NADPH and the presence of Mn2 , thecell-free extract formed dehydrosqualene as a secondaryproduct. Another E. coli extract expressing mouse squalenesynthase showed the same activity as the Arabidopsis enzyme.Therefore, both the structure and reaction mechanism ofsqualene synthases are markedly conserved in taxonomicallyremote eukaryotes.
Squalene synthase (farnesyl-diphosphate: farnesyl-diphos-phate farnesyltransferase, EC 2.5.1.21) is an endoplasmicreticulum membrane enzyme that converts two molecules offarnesyl diphosphate (FPP) into squalene. It is generallythought to be one of the key enzymes of sterol biosynthesis,since it catalyzes the first pathway-specific reaction on thesterol branch of the isoprenoid pathway (1). Whereas highlypurified preparations of squalene synthase have been obtainedfrom yeast (2) and rat (3), the enzyme has been only partiallypurified from plants (4, 5).
Squalene synthase is also known as a bifunctional mono-meric enzyme that catalyzes the following two steps. First, twoFPPs are condensed to form an intermediate, presqualenediphosphate (PSPP) (6, 7). Second, PSPP is reductively rear-ranged into squalene in the presence of NADPH and Mg2+.We previously found that partially purified yeast squalenesynthase converted FPP and PSPP to dehydrosqualene, in-stead of squalene, in the absence of NADPH and in thepresence of Mn2+ (8, 9). This observation might be useful forelucidating the mechanistic details.Because there are many difficulties to be overcome for
experiments with plants, studies on the regulation of isopre-noid synthesis and the biological functions of sterols in plantslag behind those in mammals and yeast. Arabidopsis thaliana isan excellent model for investigating the molecular biology andgenetics of plants (10). In the present paper, we reportmolecular cloning and characterization of Arabidopsissqualene synthase. * *
MATERIALS AND METHODScDNA Cloning and Sequencing. An oligo(dT)-primed AgtlO
library containing Arabidopsis (Columbia ecotype) cDNAinserts was a gift from M. Learned (Massachusetts Institute ofTechnology). Recombinant phages were plated with Esche-richia coli NM514 and then transferred to Hybond-N+ mem-branes (Amersham; 4 x 104 plaques per sheet). They werescreened with a 32P-labeled EcoRI-Pvu II fragment, 1.3 kb insize, which contained the entire coding sequence of the mousesqualene synthase cDNA (11). Hybridization and washing wereperformed as described (12).The cDNA inserts of two positive clones were cut out with
EcoRI and then inserted in the EcoRI site of pBluescript IIKS( -) (Stratagene). The resulting plasmids, designatedpATSS-4 and pATSS-10, were digested with exonuclease III,
and an overlapping series of deletions was prepared for eachplasmid. The nucleotide sequences of both strands of eachplasmid were determined by the chain-termination method(13) using Sequenase version 2.0 (United States Biochemical)and [a-32P]dCTP. Sequence data were analyzed using GENE-WORKS (IntelliGenetics) and GENETYX (Software Develop-ment, Tokyo) software.
In Vitro Transcription/Translation. pATSS-4 was linear-ized by digestion with Xho I. Transcription with T7 RNApolymerase (Bethesda Research Laboratories) and translationwith a rabbit reticulocyte lysate (Amersham) and [35S]methi-onine were carried out according to the manufacturer's in-structions. The radiolabeled protein was separated by SDS/PAGE in a 10% polyacrylamide gel.RNA Blot-Hybridization Analysis. Thirty micrograms of
Arabidopsis total RNA (supplied by M. Matsuoka, NagoyaUniversity) was separated by electrophoresis in a 1.2% agarosegel containing 2.2 M formaldehyde and then blotted onto aHybond-N+ membrane. The entire cDNA fragment ofpATSS-4 was radiolabeled with [32P]dCTP and a Megaprimekit (Amersham) and used as a probe. The hybridization andwashing conditions were as described (11).
Construction of an Expression Vector of ArabidopsisSqualene Synthase. To amplify the whole coding region, aprimer, ATS03 (5'-GAATCGATGGGGAGCTTGGGGAC-GATG-3'), that is specific for the 5' end of the Arabidopsissqualene synthase coding region, was synthesized. It contained
Abbreviations: FPP, farnesyl diphosphate; PSPP, presqualene diphos-phate.tPresent address: Department of Chemistry, Faculty of Science, KyotoUniversity, Sakyo-ku, Kyoto 606-01, Japan.tPresent address: Department of Gene Sciences, Faculty of Science,Hiroshima University, Higashi-Hiroshima 724, Japan.
IlTo whom reprint requests should be sent at the present address:Laboratory of Applied Botany, Faculty of Agriculture, Kyoto Uni-versity, Sakyo-ku, Kyoto 606-01, Japan.**The nucleotide sequence reported in this paper has been deposited
in the GenBank data base (accession no. D29017).
2328
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci USA 92 (1995) 2329
a Cla I site to facilitate cloning of PCR products. The otherprimer used was a T3 primer, the sequence of which lies nextto the polylinker site in pBluescript II. The 50-,ul reactionmixture contained 20 ng of pATSS-4 plasmid, 10 mM Tris HCl(pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.01% gelatin, 0.2 mMeach of dTTP, dCTP, dGTP, and dATP, 1.5 units of TakaraTaq DNA polymerase (Takara), and 1 ,uM each of the primers.PCR was performed for 20 cycles (94°C for 1 min, 55°C for 1min, and 72°C for 2 min). The resulting products were separatedon a 1% low-gelling-temperature agarose gel; a single band ofmaterial (1.3 kb in size) was extracted and digested with Cla I andEcoRI and then subcloned between the Cla I and EcoRI sites ofpBluescript II. Partial sequencing of the resulting expressionvector, designated pCE-8, showed that the N-terminal region of/3-galactosidase (30 amino acids) and the entire Arabidopsissqualene synthase (410 amino acids) were in-frame, as expected.
Expression ofArabidopsis Squalene Synthase in E. coli. Forinduction of the recombinant protein, E. coli XL-1-Blue cellsfreshly transformed with pCE-8 were grown at 37°C in a
medium containing 1.2% Bacto tryptone, 2.4% Bacto yeastextract, 0.4% (vol/vol) glycerol, 0.23% KH2PO4, and 1.25%K2HPO4 to an OD600 of 0.3. Isopropyl 1-thio-f3-D-galactopy-ranoside (0.5 mM final concentration) was added, and thebacteria were further incubated for 3 h. Cell lysates were
prepared and passed through Sephadex G-50 (Pharmacia)columns as described (11).Enzyme Reaction. The activity of squalene formation was
assayed as follows. The reaction mixture contained, in a totalvolume of 100 ,ul, 11.4 ,uM [1,2-14C]FPP (American Radiola-beled Chemicals, St. Louis; 125 nCi; 1 Ci = 37 GBq), 3 mMNADPH, 5 mM MgCl2, 0.1 mM dithiothreitol, 10 mM KF, 50mM Tris HCl (pH 7.6), and the enzyme (85 ,ug of protein). Thereaction was carried out at 30°C for 1 h. To assay dehydro-squalene formation, the reaction was carried out as above,except that the concentration of [14C]FPP was increased to 23,uM (250 nCi), NADPH was omitted, and 2 mM MnCl2 wasused instead of MgCl2. After the reaction, 50 ng each ofsqualene and farnesol in ethanol was added to the mixture as
carriers.TLC Analysis. For normal-phase TLC, 10 pul of the reaction
mixture was applied onto a silica gel G plate (Merck), whichwas developed with ethyl acetate/benzene (1:4, vol/vol). Forreversed-phase TLC, 10 pul of the mixture was separated on a
silica gel derivative RP-18 plate (Merck) using acetone/H20(19:1, vol/vol) as the developing solvent. The radioactivity ofeach spot was quantified with a Bio-imaging analyzer BAS2000 (Fuji).
RESULTS AND DISCUSSION
cDNA Cloning and Sequencing of Arabidopsis SqualeneSynthase. Squalene synthases have been cloned from mam-
mals (14-17) and yeasts (15, 18, 19). Several approaches couldhave been taken for the cDNA cloning of plant squalenesynthase. However, it turned out that the solubilization andpurification of plant squalene synthase is extremely difficult (4,5). Moreover, functional complementation of yeast mutantsdefective in sterol synthesis often failed to work for some
unknown reason (15, 20). We adopted cross-hybridization ofdistantly related genes under conditions of low stringency. Thisstrategy was successful for isolating theArabidopsis 3-hydroxy-3-methylglutaryl-CoA reductase gene (21, 22). The mouse
squalene synthase cDNA (11) was used as a probe, because wehad found that plant cell-cycle-related genes were more similarto mammalian homologs than to yeast homologs (23-26).Two positive clones were detected and isolated from 7 x 105
clones. Both cDNA inserts were subcloned into pBluescript II.
The resulting plasmids were designated pATSS-4 and pATSS-10. Determination of their nucleotide sequences showed thattheir inserts were overlapping and that the sequence of the
overlapping region is completely identical (data not shown).The composite sequence contained a long open reading framecoding for a 410-amino acid polypeptide. pATSS-4 containeda 5'-untranslated region and the entire coding region, whereaspATSS-10 lacked the 5' end of the coding region but containeda 3'-untranslated region. The predicted molecular mass is47,141 Da.To check the size of the protein product, the full-length
clone, pATSS-4, was linearized, transcribed in vitro, and thentranslated in vitro. The apparent molecular mass of the ex-pressed protein was 47 kDa (data not shown). This valueexactly matches the calculated molecular mass.On RNA blot-hybridization analysis, a major band corre-
sponding to 1.6 kb in size was detected (data not shown).Fainter bands corresponding to 2.2 kb and 0.6 kb were alsoobserved. At present, however, it is unclear whether or not thefainter bands represent actual transcripts of the squalenesynthase gene.
Structure Comparisons. Fig. 1 shows the alignment of theamino acid sequences of squalene synthases from Arabidopsisand other species. The amino acid sequence of the Arabidopsisenzyme was 42.5% (over 407 amino acids), 42.2% (over 408amino acids), 42.2% (over 405 amino acids), 41.6% (over 380amino acids), and 44.3% (over 325 amino acids) identical tothose of the mouse (11), rat (14), human (15-17), buddingyeast (18, 19), and fission yeast (15) enzymes, respectively. Theconserved segments, A, B, and C (14), or III, IV, and V (15),of known squalene synthases are overlined in Fig. 1. The aminoacid residues in the segments were conserved in the Arabidop-sis polypeptide. Three additional sections (amino acids 48-56,69-82, and 313-318 of the Arabidopsis enzyme) were alsoconserved (Fig. 1). These results help to narrow down theessential amino acids involved in the catalytic site.
Fig. 2 shows a phylogenetic tree of squalene synthases fromvarious species. The three mammalian enzymes are closelyrelated to each other; they form a subgroup. On the otherhand, the Arabidopsis enzyme likely belongs to another sub-group. Yeast enzymes are distantly related to mammalian orplant enzymes. These results agree with our initial speculationand indicate that plant squalene synthases are more closelyrelated to mammalian than to yeast enzymes.The hydrophobicity plot of Arabidopsis enzyme (data not
shown) constructed using the Kyte-Doolittle algorithm (28)was remarkably similar to those of the reported squalenesynthases from yeast to man (14-19). The C-terminal hydro-phobic domain is especially characteristic, because it likelyfunctions as an anchor in the endoplasmic reticulum mem-brane (15, 29). It is noteworthy that the amino acid sequencein this domain is not conserved (see Fig. 1), but the hydro-phobicity is strictly retained.
Unlike mammals or yeasts, plants have phytoene synthase.This enzyme is similar to squalene synthase in that it isbifunctional, converting two molecules of geranylgeranyldiphosphate to phytoene through prephytoene diphosphate,but different in that it is located in plastids and never utilizesNADPH (30). The amino acid sequences are only 20% iden-tical (over 165 amino acids in the middle region) in theArabidopsis enzyme and tomato phytoene synthase (31). Theirhydrophobicity plots were also quite different from each other(data not shown). Moreover, as described below, the Arabi-dopsis enzyme efficiently converts FPP into squalene depend-ing on the presence of NADPH. Thus, theArabidopsis enzymeencoded by pATSS-4 and pATSS-10 is squalene synthase, notphytoene synthase.
Confirmation of the Squalene Synthase Activity. E. coliXL-1-Blue cells, which do not synthesize squalene, weretransformed with the recombinant plasmid pCE-8. The har-vested cells were disrupted, the resulting homogenate waspassed through Sephadex G-50, and then the enzyme prepa-ration was assayed for squalene synthase activity in the pres-
Biochemistry: Nakashima et aL

2330 Biochemistry: Nakashima et al. Proc. NatL Acad Sci. USA 92 (1995)
M ---SLGTM LRYPDDIYPL LKMKRAIEKASGKLLQL-AL HPVEMKAALK LKFCRTPLFSMtS--LAN-RI EEIRCLCQYK L-WNDLPSYGMEFVKCLGHP EEFYNLLRFR MGGRRNFIPK)EFVKCLGHP EEFYNLLRFR MGGRRNFIPKMEFVKCLGHP EEFYNLVRFR IGGKRKVMPK
EKQIPPEPHW GFCYSMLHKV SRSFSLVIQQ LNTELRNAVCIYDQSTSPYL LHCFELLNLT SRSFAAVIRE LHPELRNCVTE-DENVPQNI RRCYQLLDMT SRSFAVVIKQ LPNGIRQAVMMDQDSLSSSL KTCYKYLNQT SRSFAAVIQA LDGDIRHAICMDRNSLSNSL KTCYKYLDQT SRSFAAVIQA LDGDIRHAVCMDQDSLSSSL KTCYKYLNQT SRSFAAVIQA LDGEMRNAVC
VYYLVLRALD TVZDDTSIPT DEKVPILIAPLFYLILRALD TIBDDMSIEH DLlIDLLRHFIFYLVLRGLD TVBDDMTLPL DKKLPILRDFVFYLVLRALD TVBDDMSISV EKKIPLLCNFVFYLILRAMD TVBDDMAISV EKRIPLLRNFIFYLVLRALD TLBDDMTISV EKKVPLLHNF
Consensus . LG.. EE ...LR. S.S .L ..CY. .L. .T SRSFAAVIQ. L.... R.AVC .FYLVLRALD TVBDDM.I.. KK.PLL ..F 100
YSCG--TKEY KILMDQFHHV SAAFLELEKG YQEAEEEITRFDGNAPDVKD RAVLTDFESI LIEFHKLKPE YQEVIKEITEFNESGPNEKD RQLLVEFDVV IKEYLNLSEG YRNVISNITKFTES--KEKD RQVLEDFPTI SLEFRNLAEK YQTVIDDICHFTES--KEKH RVVLEDFPTI SLEFRNLAEK YQTVIADICHFMES--KEKD RQVLEDFPTI SLEFRNLAEK YQTVIADICR
F.ES--.EKD R.VL.DF..I S.EF.NL.E. YQ.VI..I..
BAISNSMGLFL QITNIIRDYL BDINEIPKSR MFWPRBIWGKQLYESMGLF QKTNIIRDYN BDLVDGRS-- -FWPKEIWSQAISNSLGLFL QXVNIIRDYR EDFDDNRH-- -FWPRRIWSKECANSMGLFL QXTNITRDYL BDQQEGRK-- -FWPQEVWGRECANSMGLFL QITNIIRDYL BDQQEGRQ-- -FWPQEVWGKERANSMGLFL QxTNIIRDYL BDQQGGRE-- -FWPQEVWSR...NSMGLFL QXTNIIRDYL BD...GR.-- -.W.B.W..
VFRGVVXLRR CLTAKVIDRTVLHGNVKIRK GTTCYLILKSVFQTNVKIRK GQAVQIILHSVFKGVVKIRK CQAVTLMMDAVFKGVVIIRK QQAVTLMMDAVFKGAVXIRK CQAVTLMMDAVF.G.VKIRK GQAV.L ....
KTMADVYGAFRTLRGCVEIFVNLKNVCDLFTNMPAVKAI ITNMPAVKAI ITNMPAVKAI I
.NM..V. .I.
CCAXPQIMAIG TLLCYNNEQCAIPQVMAIA TLALVFNNRECAIPQVMLIA TLAAVFRNPDCAIPQVMAIA TLAACYNNQQCAIPQVMAIA TIJAACYNNHQCAIPQVNAIA TLAACYNNQQCAIPQVM&IA TLAACYNN .Q
RMGAGMAKF-KNGNMGADY-EMGDGMAYYAQMGCGMAEF-RNQGCGKEF-RMGIGMAEF-.MG.GMA.F-
YADKLEDLKYYAPQLKDFMKYTSSFGDLCLYIKKLEDFAKYVKKLEDFVKYVKKLGDFAK
Y. .KL.DF.K
YDFSCMLKTKDYYLRDIKSKLRYTRDIHYKYQYIEEIYHRYQYIEEIYHRYQYMEEIYHRY.Y.. .I...
-ICQE -----ILDENYNLNGSLAEKNDGFS-VDKD------LNKD------LDKH-----
EENTNKSVQCPENEQLGLDCPDNSEKALECPENVDVAVQCPENVDVAVKCPENIDKAVQCPEN ... .AV.C
AVETVDDYDEY CRYAGLVGL GLSKLFLAAGLQTVHDYDVY CHYVAGLVGD GLTRLIVIAKVETIEDFNKY CRYVAGLVGI GLSRLFAQSKVTSKQDWDKY CK!VAGLVGI GLSRLFSASEVTSKQDWDKY CNYVAGLVGI GLSRLFSASEVTSEQEWDKY CEYVAGLVGI GLSRLFSASE
V....D.DKY CNYVAGLVGI GLSRLF.AS.
LNEMVTNALM IIEDCLKYMV SLRDPSIFRFINHLVLNALS 8VIDVLTYLA SIHEQSTFQFLSDMTANALT HATDALVYLS QLKTQEYFNFLNELITNTLQ HIPDVLTYLS RLRNQSVFNFLNELITNALQ EIPDVITYLS RLRNQSVFNFLNELITNALH IIPDVITYLS RLRNQSVFNFLNEL.TNAL. N3I.DVLTYLS .LR.QS.FNF
VDKNDPNASK TLNRLEAV-- --QKL--CRD AG--VLQNRKLAVQDPNFLK LNIQISKIEQ FMEEMYQDKL PPNVKPNETPNTPKDPNFLK ISIECGKIEQ VSESLFPRRF REMYEKAYVSIPNSDPSSSX TKQVISKIR- -TQNLPNCQL IS--RSHYSPVPNSDPSASK AKQLISNIR- -TQSLPNCQL IS--RSHYSPIPDSDPSSSX TRQIISTIR- -TQNLPNCQL IS--RSHYSP
....DP ..SX .... IS.1I.- -.Q.L..C.L ...--... Y.P
188198195191191191200
287295292288288288300
379395392384384384400
FIIMVVIL-L AIVFA-----EEEYKFNMVL SIILSVLLGFKELYRKDL-Q KLGISILFVFWQ-YLSTL-S QVTED-----WQ-YLSTL-S QVTED-----WQ-YLTTL-S QVTED-----
.Y.. .L-.
-YLR------YYIYT-----FIILVCLAVI-YVQR------YVQR------YVQT-----_y. _ _
--------AN--------LHRFYVFNIRIHW-------EH--------EH-------GEH-
A--------SDFKELNLF
-------.H- ---------
FIG. 1. Amino acid sequence alignment of squalene synthase polypeptides from Arabidopsis, budding yeast (18, 19), fission yeast (15), mouse
(11), rat (14), and man (15-17). Amino acid residues common to more than four polypeptides are denoted as consensus ones. Amino acids identicalthroughout are shown in boldface. Dashes indicate gaps that were introduced to maximize the alignment. The conserved segments A, B, and C(14) are overlined.
ence of NADPH and Mg2+. As shown in Fig. 3, the efficientconversion of FPP into squalene was confirmed by normal-phase TLC (lane 1). An apparent specific activity of 102 pmolper mg of protein per min was observed. This activity was
comparable to that of mouse squalene synthase expressed inthe same way (lane 3; ref. 11). On the other hand, the enzymepreparation of E. coli transformed with pBluescript II withoutan insert showed no activity (lane 2). Farnesol seems to begenerated by endogeneous phosphatases in E. coli. To furtherconfirm that the reaction product is squalene, reversed-phaseTLC was carried out. Again, the product comigrated with
authentic squalene (Fig. 4, lane 1). Expression of the recom-
binant protein in E. coli may facilitate purification of the plantenzyme and may lead to detailed analysis of the structure ofthe enzyme at the atomic level.Formation of Dehydrosqualene. When the enzyme prepa-
rations of Arabidopsis and mouse were incubated with[14C]FPP in the absence of NADPH and in the presence ofMn2+, the TLC pattern of the reaction products drasticallychanged (Fig. 3, lanes 4 and 6). This pattern was identical tothat observed by Takatsuji et al. (9). Unidentified compoundsA and B proved to be generated by squalene synthases, since
3Mouse
0.02f'0.006)N34O6 Rat
(*o.01 o)0.6
uman
Arabidopsi s
S. cerevisiae
S. polbe
FIG. 2. Phylogenetic tree for the six squalene synthase polypeptides. The unweighted pair group method with arithmetic mean tree was
constructed by the method of Nei (27) using GENEWORKS software. The horizontal distances to the branching points correspond to the relativedegrees of identity among the polypeptides. The values on the branches give the branch lengths in the number of amino acid substitutions per site.The gray lines and values in parentheses at branch points in the tree are error bars and standard errors, respectively.
ArabidopsisS.cerevisiaeS.pombeMouseRatHuman
HRHIYDTDWHHEKLLLTKWSYKTIEVEGWTHTFLYDPEWRHTFLYEPEWRHSFLYQPDWRH..LY...W.
SE-VLTPDWEFANESLYSNELEDPDLAHSQFEDPIVGEDIFEDPIVGEDTFEDPLVGEDTFEDP ......
979995
100100100
ArabidopsisS.cerevisiaeS.pombeMouseRatHuman
Consensus
ArabidopsisS.cerevisiaeS.pombeMouseRatHumanConsensus
ArabidopsisS.cerevisiaeS.pombeMouseRatHumanConsensus
Arabidops isS .cerevisiaeS .pombeMouseRatHuman
Consensus
SYV------NIFLKVKERSRKLSEQKKGNGIYL------SIYL------SIYL------S
IYL-------
--DKGQPNSVYDDELVPTQQTQKAILNDEQF-IMLLAALSF-IMLLAALSF-IMLLAALS. -..LL ....
410444460416416417469
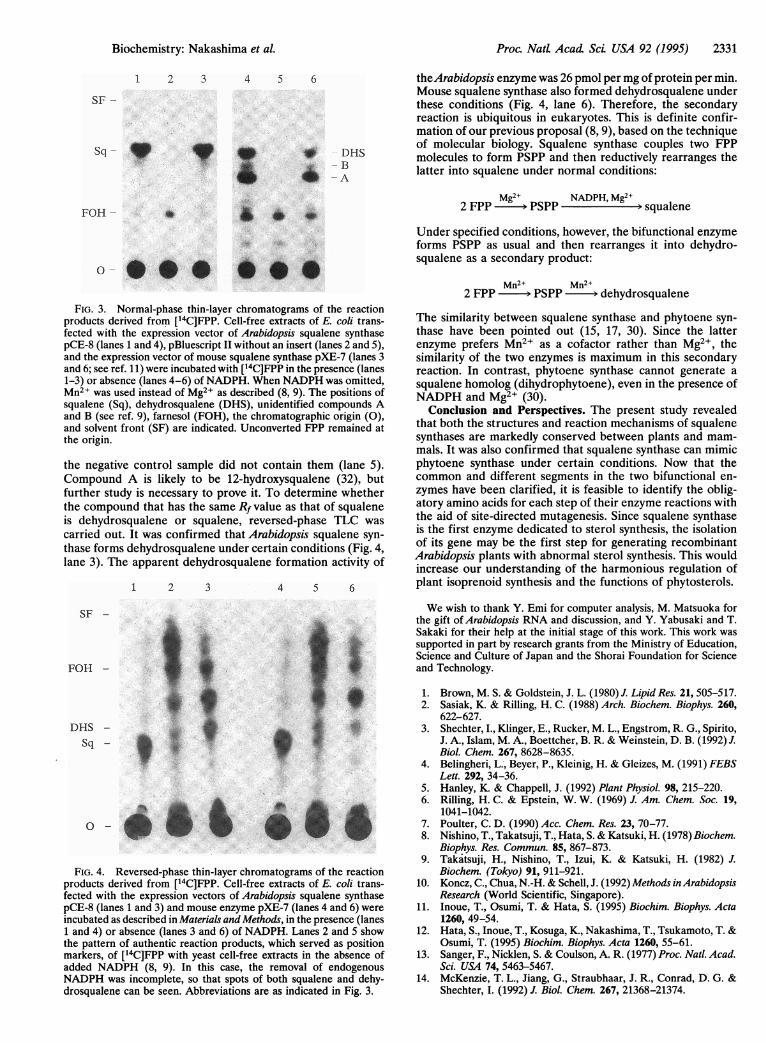
Proc. NatL Acad Sci USA 92 (1995) 2331
1 2 3 4 5 6
SF --
Sq- qp * DHS4 - Bw --A
FOH
O**. **.
FIG. 3. Normal-phase thin-layer chromatograms of the reactionproducts derived from [14C]FPP. Cell-free extracts of E. coli trans-fected with the expression vector of Arabidopsis squalene synthasepCE-8 (lanes 1 and 4), pBluescript II without an insert (lanes 2 and 5),and the expression vector of mouse squalene synthase pXE-7 (lanes 3and 6; see ref. 11) were incubated with [14C]FPP in the presence (lanes1-3) or absence (lanes 4-6) of NADPH. When NADPH was omitted,Mn2+ was used instead of Mg2+ as described (8, 9). The positions ofsqualene (Sq), dehydrosqualene (DHS), unidentified compounds Aand B (see ref. 9), farnesol (FOH), the chromatographic origin (0),and solvent front (SF) are indicated. Unconverted FPP remained atthe origin.
the negative control sample did not contain them (lane 5).Compound A is likely to be 12-hydroxysqualene (32), butfurther study is necessary to prove it. To determine whetherthe compound that has the same Rf value as that of squaleneis dehydrosqualene or squalene, reversed-phase TLC was
carried out. It was confirmed that Arabidopsis squalene syn-thase forms dehydrosqualene under certain conditions (Fig. 4,lane 3). The apparent dehydrosqualene formation activity of
1 2 3 4 5 6
theArabidopsis enzyme was 26 pmol per mg of protein per min.Mouse squalene synthase also formed dehydrosqualene underthese conditions (Fig. 4, lane 6). Therefore, the secondaryreaction is ubiquitous in eukaryotes. This is definite confir-mation of our previous proposal (8, 9), based on the techniqueof molecular biology. Squalene synthase couples two FPPmolecules to form PSPP and then reductively rearranges thelatter into squalene under normal conditions:
Mg2+ NADPH, Mg2+2 FPP - PSPP >squalene
Under specified conditions, however, the bifunctional enzymeforms PSPP as usual and then rearranges it into dehydro-squalene as a secondary product:
Mn2+ Mn2+2 FPP -> PSPP - > dehydrosqualene
The similarity between squalene synthase and phytoene syn-thase have been pointed out (15, 17, 30). Since the latterenzyme prefers Mn2+ as a cofactor rather than Mg2+, thesimilarity of the two enzymes is maximum in this secondaryreaction. In contrast, phytoene synthase cannot generate asqualene homolog (dihydrophytoene), even in the presence ofNADPH and Mg2+ (30).
Conclusion and Perspectives. The present study revealedthat both the structures and reaction mechanisms of squalenesynthases are markedly conserved between plants and mam-mals. It was also confirmed that squalene synthase can mimicphytoene synthase under certain conditions. Now that thecommon and different segments in the two bifunctional en-zymes have been clarified, it is feasible to identify the oblig-atory amino acids for each step of their enzyme reactions withthe aid of site-directed mutagenesis. Since squalene synthaseis the first enzyme dedicated to sterol synthesis, the isolationof its gene may be the first step for generating recombinantArabidopsis plants with abnormal sterol synthesis. This wouldincrease our understanding of the harmonious regulation ofplant isoprenoid synthesis and the functions of phytosterols.
SF -
FOH -
DHSMq..9
Sq~~~ .
;.....
0o-6S
FIG. 4. Reversed-phase thin-layer chromatograms of the reactionproducts derived from [14C]FPP. Cell-free extracts of E. coli trans-fected with the expression vectors of Arabidopsis squalene synthasepCE-8 (lanes 1 and 3) and mouse enzyme pXE-7 (lanes 4 and 6) wereincubated as described in Materials and Methods, in the presence (lanes1 and 4) or absence (lanes 3 and 6) of NADPH. Lanes 2 and 5 showthe pattern of authentic reaction products, which served as positionmarkers, of [14C]FPP with yeast cell-free extracts in the absence ofadded NADPH (8, 9). In this case, the removal of endogenousNADPH was incomplete, so that spots of both squalene and dehy-drosqualene can be seen. Abbreviations are as indicated in Fig. 3.
We wish to thank Y. Emi for computer analysis, M. Matsuoka forthe gift of Arabidopsis RNA and discussion, and Y. Yabusaki and T.Sakaki for their help at the initial stage of this work. This work wassupported in part by research grants from the Ministry of Education,Science and Culture of Japan and the Shorai Foundation for Scienceand Technology.
1. Brown, M. S. & Goldstein, J. L. (1980) J. Lipid Res. 21, 505-517.2. Sasiak, K. & Rilling, H. C. (1988) Arch. Biochem. Biophys. 260,
622-627.3. Shechter, I., Klinger, E., Rucker, M. L., Engstrom, R. G., Spirito,
J. A., Islam, M. A., Boettcher, B. R. & Weinstein, D. B. (1992) J.Biol. Chem. 267, 8628-8635.
4. Belingheri, L., Beyer, P., Kleinig, H. & Gleizes, M. (1991) FEBSLett. 292, 34-36.
5. Hanley, K. & Chappell, J. (1992) Plant Physiol. 98, 215-220.6. Rilling, H. C. & Epstein, W. W. (1969) J. Am. Chem. Soc. 19,
1041-1042.7. Poulter, C. D. (1990) Acc. Chem. Res. 23, 70-77.8. Nishino, T., Takatsuji, T., Hata, S. & Katsuki, H. (1978) Biochem.
Biophys. Res. Commun. 85, 867-873.9. Takatsuji, H., Nishino, T., Izui, K. & Katsuki, H. (1982) J.
Biochem. (Tokyo) 91, 911-921.10. Koncz, C., Chua, N.-H. & Schell, J. (1992) Methods inArabidopsis
Research (World Scientific, Singapore).11. Inoue, T., Osumi, T. & Hata, S. (1995) Biochim. Biophys. Acta
1260, 49-54.12. Hata, S., Inoue, T., Kosuga, K., Nakashima, T., Tsukamoto, T. &
Osumi, T. (1995) Biochim. Biophys. Acta 1260, 55-61.13. Sanger, F., Nicklen, S. & Coulson, A. R. (1977) Proc. Natl. Acad.
Sci. USA 74, 5463-5467.14. McKenzie, T. L., Jiang, G., Straubhaar, J. R., Conrad, D. G. &
Shechter, I. (1992) J. Biol. Chem. 267, 21368-21374.
Biochemistry: Nakashima et aL
.fI IW
4

2332 Biochemistry: Nakashima et al.
15. Robinson, G. W., Tsay, Y. H., Kienzle, B. K., Smith-Monroy,C. A. & Bishop, R. W. (1993) Mo. Cell. Bio. 13, 2706-2717.
16. Jiang, G., McKenzie, T. L., Conrad, D. G. & Shechter, I. (1993)J. Bio. Chem. 268, 12818-12824.
17. Summers, C., Karst, F. & Charles, A. D. (1993) Gene 136,185-192.
18. Jennings, S., Tsay, Y. H., Fisch, T. M. & Robinson, G. W. (1991)Proc. Natl. Acad. Sci. USA 88, 6038-6042.
19. Fegueur, M., Richard, L., Charles, A. D. & Karst, F. (1991) Curr.Genet. 20, 365-372.
20. Corey, E. J., Matsuda, S. P. T. & Bartel, B. (1994) Proc. Natl.Acad. Sci. USA 91, 2211-2215.
21. Learned, R. M. & Fink, G. R. (1989) Proc. Natl. Acad. Sci. USA86, 2779-2783.
22. Caelles, C., Ferrer, A., Balcells, L., Hegardt, F. G. & Boronat, A.(1989) Plant Moi. Bio. 13, 627-638.
23. Hata, S., Kouchi, H., Suzuka, I. & Ishii, T. (1991) EMBO J. 9,2681-2688.
Proc. Nati Acad ScL USA 92 (1995)
24. Hirayama, T., Imajuku, Y., Anai, T., Matsui, M. & Oka, A. (1991)Gene 105, 159-165.
25. Hashimoto, J., Hirabayashi, T., Hayano, Y., Hata, S., Ohashi, Y.,Suzuka, I., Utsugi, T., Toh-E, A. & Kikuchi, Y. (1992) Mo. Gen.Genet. 233, 10-16.
26. Suzuka, I., Hata, S., Matsuoka, M., Kosugi, S. & Hashimoto, J.(1991) Eur. J. Biochem. 195, 571-575.
27. Nei, M. (1987) Molecular Evolutionary Genetics (Columbia Univ.Press, New York), pp. 293-298.
28. Kyte, J. & Doolittle, R. F. (1982) J. Mo. Bio. 157, 105-132.29. Zhang, D., Jennings, S. M., Robinson, G. W. & Poulter, C. D.
(1993) Arch. Biochem. Biophys. 304, 133-143.30. Dogbo, O., Laferriere, A., d'Harlingue, A. & Camara, B. (1988)
Proc. Natl. Acad. Sci. USA 85, 7054-7058.31. Bartley, G. E., Viitanen, P. V., Bacot, K. 0. & Scolnik, P. A.
(1992) J. Bio. Chem. 267, 5036-5039.32. Shigeri, Y., Nishino, T., Yumoto, N. & Tokushige, M. (1991)
Agric. Bio. Chem. 55, 589-591.
















![Home | Cancer Research - Molecular Cloning of a ......[CANCER RESEARCH 53,227-230, January 15, 1993] Advances in Brief Molecular Cloning of a Complementary DNA Encoding a Prostate-specific](https://img.pdfslide.net/doc/110x75/5f0256ef7e708231d403c8ca/home-cancer-research-molecular-cloning-of-a-cancer-research-53227-230.jpg)












