Embed Size (px)
Citation preview

Comp. Biochem. Physiol. Vol. 99B, No. 2, pp. 287-294, 1991 0305-0491/91 $3.00 + 0.00 Printed in Great Britain © 1991 Pergamon Press plc
COMPARATIVE ENZYMATIC COMPOSITION OF BRAZILIAN CORAL SNAKE (MICRURUS) VENOMS
STEVEN D. AIRI)*t~ and NELSON JORGE DA SILVA, JR t *Natural Product Sciences, Inc., 420 Chipeta Way, Suite 240, Salt Lake City, UT 84108, USA (Tel: 801 583 4939; Fax: 801 583 4961); and #Centro de Estudos e Pesquisas Biol6gicas, Universidade Cat61ica de
Goi~s, Avenida Universit~iria , 1440, Setor Universitdrio, Goi~nia, Goi~is, CEP 74.000, Brasil
(Received 2 November 1990)
Abstract--1. Venoms of 11 coral snake taxa, including Micrurus albicinctus, M. corallinus, M. frontalis altirostris, M. f. brasiliensis, M. f. frontalis, M. fulvius fulvius, M. ibiboboca, M. lemniscatus ssp., M. rondonianus, M. spixii spixii, and M. surinamensis surinamensis, were examined for 13 enzymatic activities.
2. These were compared with venoms of three outgroup taxa: Naja naja kaouthia, Bungarus multicinctus, and Bothrops moojeni.
3. Enzyme activity levels in Micrurus venoms were highly variable from species to species. 4. All venoms possessed phospholipase activity. 5. Protease activity against synthetic or dyed natural substrates was generally negligible in all elapid
venoms examined. By contrast, most Micrurus venoms displayed ample L-leucine aminopeptidase activity. 6. Venom of M.s. surinamensis was significantly different from those of its congeners in most assays.
INTRODUCTION
Coral snakes (Genera Micrurus, Leptomicrurus, and Micruroides) constitute a taxonomic assemblage of more than 120 species and subspecies (Roze, 1982, 1989; Roze and Bernal-Carlo, 1987). They range from the southern United States to southern South America, achieving their greatest abundance and species diversity in lower latitudes. Micrurus is the most speciose of the three genera and the most widely distributed as well.
Human envenomations by coral snakes are relati- vely rare because of their semi-fossorial biology. From June 1986 to December 1989 the Programa Nacional de Ofidismo of the Brazilian Ministry of Health reported 70,600 snakebite accidents in Brazil. Of these, coral snakes accounted for only 333 (0.47%) (Resende et al., 1989; Anonymous, 1990). Despite their relatively low incidence, coral snake bites are frequently extremely serious. Clinical data regarding Micrurus envenomations are sketchy, and they suggest a considerable spectrum of pharmacological activities (McCullough and Gennaro, 1963; Mosely, 1966; Parrish and Khan, 1967; Rosenfeld, 1971; Kitchens and Van Mierop, 1987; Pettigrew and Glass, 1985).
Micrurus venoms are known to be neurotoxic (Vital Brazil, 1965, 1980, 1987; Jimrnez-Porras, 1973; Snyder et aL, 1973; Patrfio-Lomba et al., 1973; Dal6 et al., 1989; Vital Brazil et al., 1977; Moussatch6 and Melrndez, 1979; Vital Brazil and Fontana, 1984). They also cause myonecrosis (Gutirrrez et aL, 1980, 1983, 1986; Arroyo et al., 1987) and they exhibit hemorrhagic effects (Moussatch6 et al., 1976; G6mez- Leiva, 1978) and cardiovascular effects (Weis and Mclsaac, 1971; Ramsey et al., 1971, 1972a,b; Vital Brazil et al., 1977).
:~To whom correspondence should be addressed.
Coral snake venom chemistry has received little attention. In part, this is because the fossorial habits of coral snakes make them difficult to obtain. In addition they are difficult to maintain in captivity by virtue of their specialized diets. Moreover, venom yields are small for many taxa and extraction is often difficult. To date most comparative studies of Micru- rus venoms have employed cross-neutralization of antivenoms against crude venoms (Cohen et al., 1967, 1968, 1971; Bolafios et al., 1975, 1978). Stevan and Seligrnann (1970) examined venoms from 13 Micru- rus species and subspecies and from Micruroides using agar-gel and polyacrylamide gel electrophoresis and attempted to draw some taxonomic inferences from the electrophoretic profiles. Gutirrrez et aL (1983) also compared local effects of six Micrurus venoms in mice. Further complicating the picture, geographic variation of venoms within taxa has also been documented (Vital Brazil et al., 1977; Vital Brazil, 1987).
To the best of our knowledge sequence data have been gathered on only four Micrurus venom proteins to date. Possani et al. (1979) obtained an N-terminal sequence of a phospholipase from the venom of Micrurus fulvius microgalbineus. Mochca-Morales et al. (1990) reported N-terminal sequences on three toxic phospholipases from the venom of Micrurus nigrocinctus. These sequences show considerable simi- larity to other phospholipases from elapid venoms.
We commenced our studies of coral snake venom chemistry by examining the following taxa: Micrurus fulvius fulvius, M. albicinctus, M. corallinus, M. fron- talis altirostris, M. f . brasiliensis, M. f . frontalis, M. ibiboboca, M. lemniscatus ssp., M. rondonianus, M. spixii spixii, and M. surinamensis surinamensis. Ven- oms were compared chromatographically and in terms of their enzymatic compositions. Masses of all constituents (mol. wt ~< 30,000), purified by a series of
287

288 S'ixvEs D. AIRD and NELSON JORGE DA SILVA, JR
chromatographic procedures, were determined by mass spectrometry. Sequencing of venom proteins by tandem mass spectrometry and automated Edman degradation is underway. The present paper reports results o f our comparat ive enzymatic studies using crude venoms. Other manuscripts now in preparat ion will report chromatographic and sequencing results separately,
MATERIALS AND METHODS
Venoms
Specimens of Bothrops moojeni, Micrurus f . frontalis and M . f . brasiliensis were captured in the state of Goifis, and held at Centro de Estudos e Pesquisas Biolrgicas (CEPB) at the Universidade Cat61ica de Goifis. M. albicinc- tus, M. rondonianus, M. s. spixii, and M. lemniscatus ssp. were collected under the auspices of Operaq~_o Jamari, a faunal salvage operation conducted by Eletronorte during reservoir flooding at the Samual Hydroelectric Power Plant in Rond6nia, Brazil (Roze and Jorge da Silva, 1990). Micrurus s. surinamensis venom was obtained from the Centro de Ofidismo, Instituto de Medicina Tropical de Manaus (Manaus, Amazonas). Venoms of M. corallinus and M. ibiboboca were obtained from Instituto Vital Brazil (Niter6i, Rio de Janeiro). Micrurus frontalis altirostris venom was a gift from Pontificia Universidade Cat61ica do Rio Grande do Sul (Porto Alegre, Rio Grande do Sul). Lyophilized Micrurus f . fulvius venom was purchased from Biotoxins (St Cloud, Florida). Lyophilized Naja naja kaouthia and Bungarus multicinctus venoms were purchased from Ventoxin Laboratories, Inc. (Frederick, Maryland). Locality data for all samples are provided in Table 1.
Venom was extracted manually from specimens of the aforementioned taxa at CEPB using glass micropipettes. Venoms were clarified by centrifugation and the supernatant was promptly vacuum-dried.
Enzyme assays
Crude venoms were assayed for 13 enzymatic activities. Reagents were purchased from Sigma. All assays except phospholipase assays employed crude venom (40 mg/ml) dissolved in 50 mM Na2HPO4/H3PO 4 with 150 mM NaCI (pH 7.0). Blanks consisted of an equal volume of the foregoing buffer unless otherwise indicated. All samples were assayed in triplicate.
Acetylcholinesterase
This assay employed a modification of the method of Tan and Tan (1988). The reaction mixture consisted of 825/~1 phosphate-buffered saline (PBS), 100/~1 125 mM acetylthio- choline iodide, 25#1 400mM 5,5'-dithio-bis-nitrobenzoic acid in ethanol, 50 ml sodium bicarbonate (31.25 mg/ml) and 2/d diluted crude venom. The reaction was run for 10 min at room temperature and samples were then diluted
1:9 with 18 Mohm water in order to read the absorbance at 432 nm.
Alkaline phosphomonoesterase
The technique of Tan and Tan (1988) was modified as follows. The reaction mixture employed 250~1 0.5M glycine/NaOH, 250121 10raM p-nitrophenylphosphate, 150#1 10mM MgSO4 and 2/~1 diluted crude venom. The reaction proceeded at room temperature for 20 rain. Then 850/tl of 0.2 M NaOH was added and the reaction was allowed to stand for an additional 20min, before determining the absorbance at 440 nm.
Hyaluronidase
This assay modified the technique of Pukrittayakamee et al. (1988). The reaction mixture consisted of 400 #1200 mM sodium acetate (pH 6.0), 100/d hyaluronic acid (0.5 mg/ml) in 200 mM sodium acetate (pH 6.0), and 2/~1 diluted crude venom. A 0% turbidity blank consisted of 500/~1 of buffer and a 100% turbidity blank consisted of buffer and sub- strate without venom. Buffer, venom and substrate were mixed and incubated for 15 min at 37°C. Then 1 ml of 2.5% cetyltrimethylammonium bromide in 2% NaOH was added to samples and to both blanks. Absorbance was determined at 400nm against the 0% turbidity blank. The 100% turbidity blank was used as the no hyaluronidase activity standard.
L-Amino acid dehydrogenase
The reaction mixture consisted of 8901~1 100mM Tris-HC1 (pH 8.0), 100/tl 5 mM L-kynurenine SO 4 in the above buffer (153mg/100ml), and 10/tl diluted crude venom. This was a modification of the method of Weissbach et al. (I 960). Samples and blanks were incubated at 37°C for 30 min and absorbance was determined at 332nm.
5'-Nucleotidase
Samples of diluted crude venoms (2/d) were assayed in 1.0 ml of 5'-nucleotidase reagent solution in accordance with Sigma Technical Bulletin 265-UV. Absorbance was moni- tored at 340 nm.
Phosphodiesterase
The method of Koerner and Sinsheimer (1957) was modified as follows. Diluted crude venom (2 #1) was incu- bated for 1 hr at 37°C with 1.0 ml of reagent salt solution consisting of 40 ml Ca 2÷ bis p-nitrophenyl phosphate [1 mM in 10 mM ammonium acetate (pH 4.5)], 12 ml 0.3 M mag- nesium chloride, 40 m] 0.1 M Tris/acetic acid (pH 8.8), and 24ml 18 Mohm water. Activity was quantified by an in- crease in absorbance at 400 nm.
Phospholipase A 2
The phenol red method of Lrbo de Arafijo and Radvanyi (1987) was employed, using 5#1 diluted crude venom (2 #g/#l in 18 Mohm water) in 3 ml phenol red substrate solution. Phosphatidylcholine served as the substrate. Ab- sorbance was monitored at 558 nm. Reaction progress was
Table 1. Locality data for Micrurus venom samples. All samples represent pooled contributions of two or more animals from the same locality
No. of Taxon Locality specimens
M. albicinctus M. corallinus M. f altirostris M. f brasiliensis M. f frontalis M. ibiboboca M. lemniscatus ssp. M. rondonianus M. s. spixii M. s. surinamensis
UHE Samuel, Porto Velho County, Rondrnia 15 Angra dos Reis County, Rio de Janeiro 2 Cachoeira do Sul County, Rio Grande do Sul 7 Goifmia County, Goifis 2 Jatai County, Goi~is 2 Itaparica County, Bahia ? UHE Samuel, Porto Velho County, Rondbnia 10 UHE Samuel, Porto Velho County, Rondrnia 25 UHE Samuel, Porto Velho County, Rondbnia 10 Manaus County, Amazonas ?
Enzymatic composition
tracked using a Hewlett Packard 8452A diode array spectro- photometer in kinetics mode. Absorbances were taken every 15 sec for 20 min.
Six substrates were employed to assay protease and peptidase activities.
L-Leucine aminopeptidase
This assay consisted of 2/~1 diluted crude venom in I ml L-leueine p-nitroanilide (1 mg/ml) in PBS. Samples were incubated for 2 hr at 37°C and absorbance was determined at 390 nm.
BAPNA
Diluted crude venom (2/~1) was incubated for 8 hr at 37°C with 1 ml N-benzoyl-oL-arginine-p-nitroanilide (0. I mg/ml). Absorbance was monitored at 394 nm.
N-Succinyl phenylalanyl peptidase
Diluted crude venom (2/~1) was incubated with 1 ml N-succinyl phenylalanyl p-nitroanilide. Samples were incu- bated for 2 hr at 37°C and absorbance was determined at 384 nm.
Azoalbumin / azocasein
Diluted crude venom (2/zl) was incubated at 37°C for 2.5hr with 1.0ml azoalbumin or azocasein (1 mg/ml) in PBS. The reaction was stopped by adding 100/zl cold 0.5 M trichloroacetic acid (TCA). The mixture was centrifuged for 5 rain in a microcentrifuge and the absorbances were deter- mined at 342 nm.
Azocoll
Assays were performed using 2/~1 diluted crude venom and 5 mg azocoll (Calbiochem, 100-250 mesh) in 1 ml PBS. Samples were incubated for 2hr at 37°C and the ab- sorbances were determined at 520 nm.
RESULTS AND DISCUSSION
Enzymatic assays
Acetylcholinesterase was absent in venom of the outgroup species Bothrops moojeni and was very active in Naja naja kaouthia and Bungarus multicinc-
3~
._~ 2.5 >
1.5
.__
@ < o.5
0
T a x o n
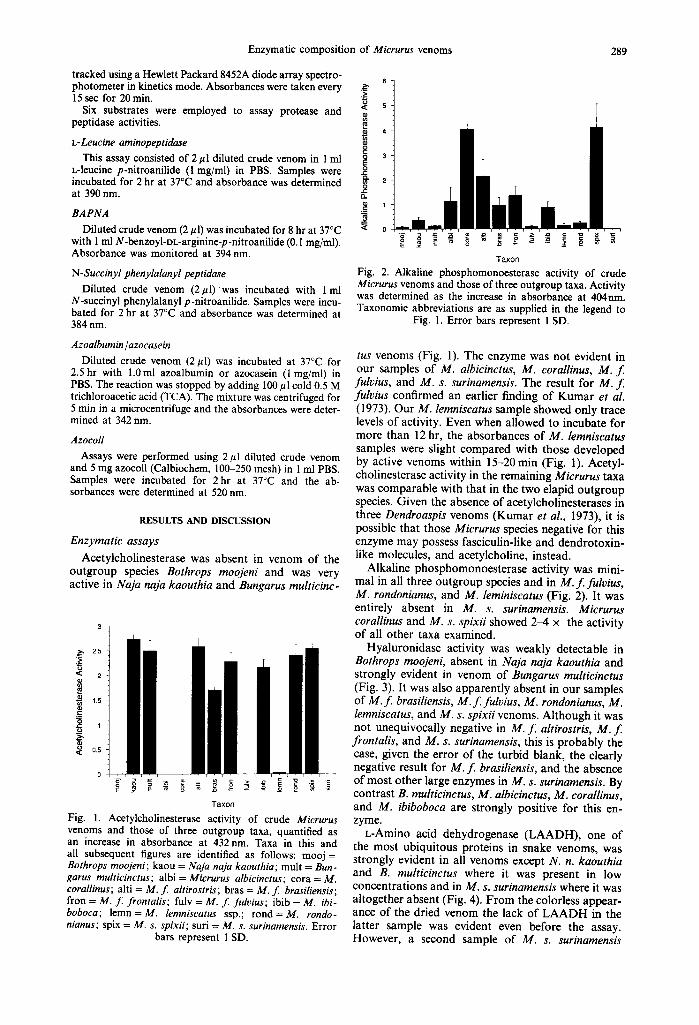
Fig. I. Acetylcholinesterase activity of crude Micrurus venoms and those of three outgroup taxa, quantified as an increase in absorbance at 432 nm. Taxa in this and all subsequent figures are identified as follows: mooj = Bothrops moojeni; kaou = Naja naja kaouthia; mult = Bun- garus multicinctus; albi = Micrurus albicinctus; cora = M. corallinus; alti = M. f. ahirostris; bras = M. f. brasiliensis; fron = M. f. frontalis; fulv = M. f. fulvius; ibib = M. ibi- boboca; lemn=M, lemniscatus ssp.; rond=M, rondo- nianus; spix = M. s. spixii; suri = M. s. surinamensis. Error
bars represent 1 SD.
of Micrurus venoms 289
6
<= 5
¢n
o
2 2
2 0 .
< 0
T a x o n
Fig. 2. Alkaline phosphomonoesterase activity of crude Micrurus venoms and those of three outgroup taxa. Activity was determined as the increase in absorbance at 404nm. Taxonomic abbreviations are as supplied in the legend to
Fig. I. Error bars represent 1 SD.
tus venoms (Fig. 1). The enzyme was not evident in our samples of M. albicinctus, M. corallinus, M. f . fulvius, and M. s. surinamensis. The result for M. f. fulvius confirmed an earlier finding of Kumar et al. (1973). Our M. lemniscatus sample showed only trace levels of activity. Even when allowed to incubate for more than 12 hr, the absorbances of M. lemniscatus samples were slight compared with those developed by active venoms within 15-20 min (Fig. 1). Acetyl- cholinesterase activity in the remaining Micrurus taxa was comparable with that in the two elapid outgroup species. Given the absence of acetylcholinesterases in three Dendroaspis venoms (Kumar et al., 1973), it is possible that those Micrurus species negative for this enzyme may possess fasciculin-like and dendrotoxin- like molecules, and acetylcholine, instead.
Alkaline phosphomonoesterase activity was mini- mal in all three outgroup species and in M. f . fulvius, M. rondonianus, and M. leminiscatus (Fig. 2). It was entirely absent in M. s. surinamensis. Micrurus corallinus and M. s. spixii showed 2-4 × the activity of all other taxa examined.
Hyaluronidase activity was weakly detectable in Bothrops moojeni, absent in Naja naja kaouthia and strongly evident in venom of Bungarus multicinctus (Fig. 3). It was also apparently absent in our samples of M. f . brasiliensis, M. f . fulvius, M. rondonianus, M. lemniscatus, and M. s. spixii venoms. Although it was not unequivocally negative in M . f . altirostris, M . f . frontalis, and M. s. surinamensis, this is probably the case, given the error of the turbid blank, the clearly negative result for M . f . brasiliensis, and the absence of most other large enzymes in M. s. surinamensis. By contrast B. multicinctus, M. albicinctus, M. corallinus, and M. ibiboboca are strongly positive for this en- zyme.
L-Amino acid dehydrogenase (LAADH), one of the most ubiquitous proteins in snake venoms, was strongly evident in all venoms except N. n. kaouthia and B. multicinctus where it was present in low concentrations and in M. s. surinamensis where it was altogether absent (Fig. 4). F rom the colorless appear- ance of the dried venom the lack of L A A D H in the latter sample was evident even before the assay. However, a second sample of M. s. surinamensis
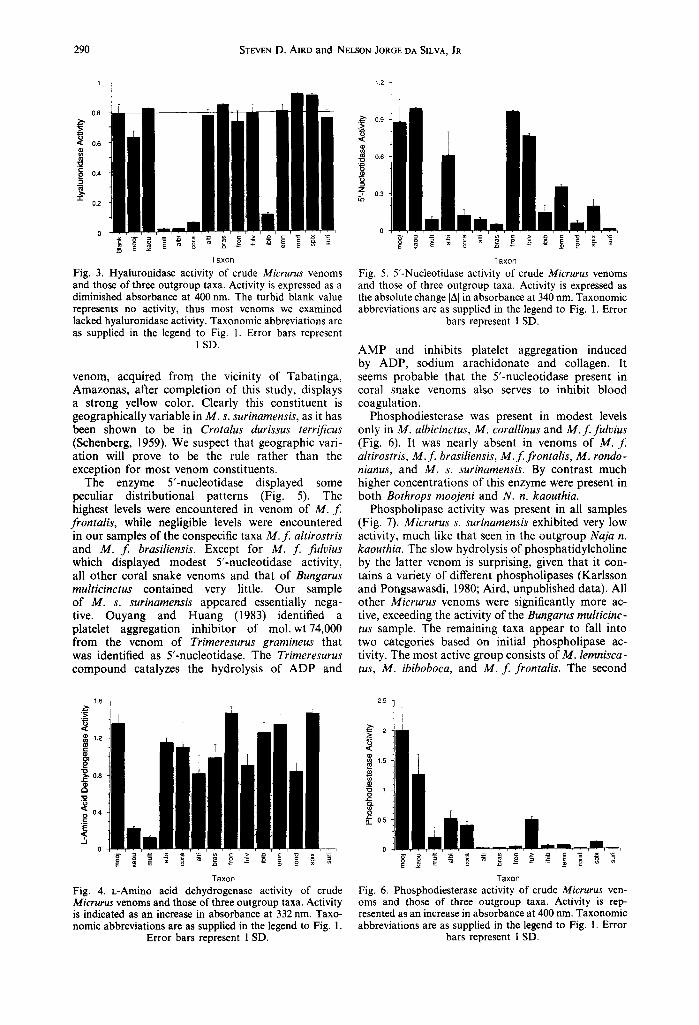
290 STEVEN D. AIRD and NELSON JORGE DA SILVA, JR
0,8
0.6
0.4
"1" 0.2
1.2 q
,.,~ 0.9
0.6
Z &~ 0.3
0
E == ~ ~ ,~ -~ = - Taxon
Fig. 3. Hyaluronidase activity of crude Micrurus venoms and those of three outgroup taxa. Activity is expressed as a diminished absorbance at 400 nm. The turbid blank value represents no activity, thus most venoms we examined lacked hyaluronidase activity. Taxonomic abbreviations are as supplied in the legend to Fig. 1. Error bars represent
1 SD.
venom, acquired from the vicinity of Tabatinga, Amazonas, after completion of this study, displays a strong yellow color. Clearly this constituent is geographically variable in M. s. surinamensis, as it has been shown to be in Crotalus durissus terrificus (Schenberg, 1959). We suspect that geographic vari- ation will prove to be the rule rather than the exception for most venom constituents.
The enzyme 5'-nucleotidase displayed some peculiar distributional patterns (Fig. 5). The highest levels were encountered in venom of M. f . frontalis, while negligible levels were encountered in our samples of the conspecific taxa M. f altirostris and M. f brasiliensis. Except for M. f fulvius which displayed modest 5'-nucleotidase activity, all other coral snake venoms and that of Bungarus multicinctus contained very little. Our sample of M. s. surinamensis appeared essentially nega- tive. Ouyang and Huang (1983) identified a platelet aggregation inhibitor of mol. wt74,000 from the venom of Trimeresurus grarnineus that was identified as 5'-nucleotidase. The Trimeresurus compound catalyzes the hydrolysis of A D P and
0
Taxon
Fig. 5. 5'-Nucleotidase activity of crude Micrurus venoms and those of three outgroup taxa. Activity is expressed as the absolute change IAI in absorbance at 340 nm. Taxonomic abbreviations are as supplied in the legend to Fig. 1. Error
bars represent 1 SD.
A M P and inhibits platelet aggregation induced by ADP, sodium arachidonate and collagen. It seems probable that the 5'-nucleotidase present in coral snake venoms also serves to inhibit blood coagulation.
Phosphodiesterase was present in modest levels only in M. albicinctus, M. corallinus and M. f . fulvius (Fig. 6). It was nearly absent in venoms of M. f. altirostris, M. f . brasiliensis, M. f . frontalis, M. rondo- nianus, and M. s. surinamensis. By contrast much higher concentrations of this enzyme were present in both Bothrops moojeni and N. n. kaouthia.
Phospholipase activity was present in all samples (Fig. 7). Micrurus s. surinamensis exhibited very low activity, much like that seen in the outgroup Naja n. kaouthia. The slow hydrolysis of phosphatidylcholine by the latter venom is surprising, given that it con- tains a variety of different phospholipases (Karlsson and Pongsawasdi, 1980; Aird, unpublished data). All other Micrurus venoms were significantly more ac- tive, exceeding the activity of the Bungarus multicinc- tus sample. The remaining taxa appear to fall into two categories based on initial phospholipase ac- tivity. The most active group consists of M. lemnisca- tus, M. ibiboboca, and M. f . frontalis. The second
1.6 q ¢
~ 1.2
c
~ 0.8
~ 0.4
E
.5 0
E , = { o -z ~ . . . . _~ .
T a x o n
Fig. 4. L-Amino acid dehydrogenase activity of crude Micrurus venoms and those of three outgroup taxa. Activity is indicated as an increase in absorbance at 332 nm, Taxo- nomic abbreviations are as supplied in the legend to Fig. 1.
Error bars represent 1 SD,
25 ~ _
" I ~2 <
1.5
o~
.c:
o ~. 0.5
0
Taxon
Fig. 6. Phosphodiesterase activity of crude Micrurus ven- oms and those of three outgroup taxa. Activity is rep- resented as an increase in absorbance at 400 nm. Taxonomic abbreviations are as supplied in the legend to Fig. 1. Error
bars represent I SD.
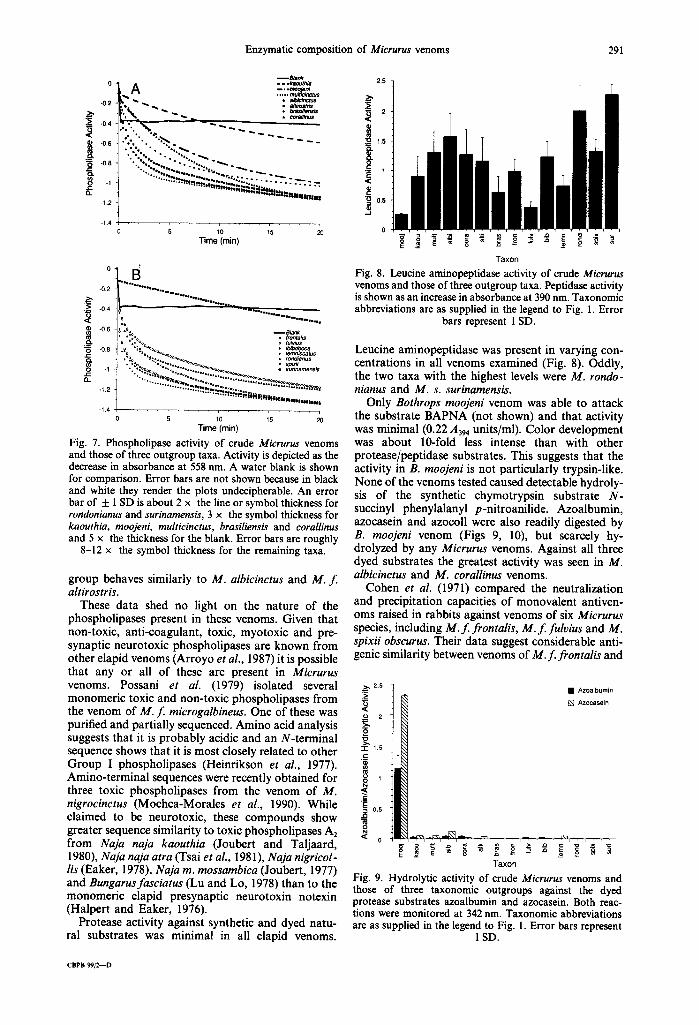
Enzymatic composition of Micrurus venoms 291
-0.2
"~; -0.4
[ -0.6
~ -0.8 }-, Q..
-1.2
-1.4
- - B m n k -..kaouthia -- ,° ni A ..... ~ . ~
"~s,~ • atbiclnctu$ • a/L~ostds
÷. % "% • . o %, , , " ~ % %%%,% o . ° o',.~, - . ~ .
~"~',~,,,,,r~.~p.~,~ ,. • . . . . - - ~ | H I l i l I I U l I I
5 10 15 20 Time (rnin)
- ' : ' ' " : ' : ............. ............................ -0.2 °°°°'°'°°°'O,-o,o~
".--~ -0.4
.~.
g o
-1,2 . . . . . . . . . "*"* ~*'~|]}ll~llllliJn~luoe,,j,**,,,¢
-I .4 5 10 15 20
Time (rain)
Fig. 7. Phospholipase activity of crude Micrurus venoms and those of three outgroup taxa. Activity is depicted as the decrease in absorbance at 558 nm. A water blank is shown for comparison. Error bars are not shown because in black and white they render the plots undecipherable. An error bar of _+ 1 SD is about 2 x the line or symbol thickness for rondonianus and surinamensis, 3 x the symbol thickness for kaouthia, moojeni, multicinctus, brasiliensis and corallinus and 5 x the thickness for the blank. Error bars are roughly
8-12 x the symbol thickness for the remaining taxa.
group behaves similarly to M. albicinctus and M. f. altirostris.
These data shed no light on the nature of the phospholipases present in these venoms. Given that non-toxic, anti-coagulant, toxic, myotoxic and pre- synaptic neurotoxic phospholipases are known from other elapid venoms (Arroyo et al., 1987) it is possible that any or all of these are present in Micrurus venoms. Possani et al. (1979) isolated several monomeric toxic and non-toxic phospholipases from the venom of M. f microgalbineus. One of these was purified and partially sequenced. Amino acid analysis suggests that it is probably acidic and an N-terminal sequence shows that it is most closely related to other Group I phospholipases (Heinrikson et al., 1977). Amino-terminal sequences were recently obtained for three toxic phospholipases from the venom of M. nigrocinctus (Mochca-Morales et aL, 1990). While claimed to be neurotoxic, these compounds show greater sequence similarity to toxic phospholipases A2 from Naja naja kaouthia (Joubert and Taljaard, 1980), Naja naja atra (Tsai et aL, 1981), Naja nigrieol- lis (Eaker, 1978), Naja m. mossambica (Joubert, 1977) and Bungarusfasciatus (Lu and Lo, 1978) than to the monomeric elapid presynaptic neurotoxin notexin (Halpert and Eaker, 1976).
Protease activity against synthetic and dyed natu- ral substrates was minimal in all elapid venoms.
2.5
¢U
~ 1.5
8 <C
"~ 0.5
0
Taxon Fig. 8. Leucine aminopeptidase activity of crude Micrurus venoms and those of three outgroup taxa. Peptidase activity is shown as an increase in absorbance at 390 nm. Taxonomic abbreviations are as supplied in the legend to Fig. 1. Error
bars represent 1 SD.
Leucine aminopeptidase was present in varying con- centrations in all venoms examined (Fig. 8). Oddly, the two taxa with the highest levels were M. rondo- nianus and M. s. surinamensis.
Only Bothrops moojeni venom was able to attack the substrate BAPNA (not shown) and that activity was minimal (0.22 A394 units/ml). Color development was about 10-fold less intense than with other protease/peptidase substrates. This suggests that the activity in B. moojeni is not particularly trypsin-like. None of the venoms tested caused detectable hydroly- sis of the synthetic chymotrypsin substrate N- succinyl phenylalanyl p-nitroanilide. Azoalbumin, azocasein and azocoll were also readily digested by B. moojeni venom (Figs 9, 10), but scarcely hy- drolyzed by any Micrurus venoms. Against all three dyed substrates the greatest activity was seen in M. albicinctus and M. corallinus venoms.
Cohen et al. (1971) compared the neutralization and precipitation capacities of monovalent antiven- oms raised in rabbits against venoms of six Micrurus species, including M. f. frontalis, M. f. fulvius and M. spixii obscurus. Their data suggest considerable anti- genic similarity between venoms of M. f. frontalis and
.•2.5 • Azoalbumin
~; [ ] Azocasein <E ._o 2
i 1.5 .~_
.c
~ 0.5
Taxon Fig. 9. Hydrolytic activity of crude Micrurus venoms and those of three taxonomic outgroups against the dyed protease substrates azoalbumin and azocasein. Both reac- tions were monitored at 342 nm. Taxonomic abbreviations are as supplied in the legend to Fig. 1. Error bars represent
1 SD.
CBPB 99/2--D

25
STEVEN D. AXRD a n d NELSON JORGE DA SILVA, JR
"5
-r 1
0,5
0
292
Taxon
Fig. 10. Hydrolytic activity of crude Micrurus venoms and those of three outgroup taxa against the dyed protease substrate azocoll. Azocoll hydrolysis was followed at 502 nm. Taxonomic abbreviations are as supplied in the
legend to Fig. 1. Error bars represent l SD.
M. s. obscurus. By contrast, M. f . fulvius venom appeared only distantly related to those of the other two species. Some of the enzymatic data suggest the same affinities. Micrurus f . fulvius venom lacks acetylcholinesterase activity, whereas that activity is pronounced in the other two venoms. Alkaline phosphomonoesterase activity is minimal in M. f fulvius, but significant in M. f frontalis and especially in M. s. spixii. Phosphodiesterase activity is ample in M. f fulvius and negligible in the other two taxa.
Micrurus corallinus and M. albicinctus also yielded similar results in 11 of 13 assays, suggesting a possible close phylogenetic relationship. Except for the 5'- nucleotidase assay, all three M. frontalis sub- species yielded similar results.
Most significantly, M. s. surinamensis showed little or no activity in 12 of 13 assays. Oddly, it possessed greater L-leucine aminopeptidase activity than any other venom examined. The absence of most large enzymes in M. surinamensis venom and the diver- gence of M. surinamensis from other Micrurus species with regard to more ubiquitous enzymes suggest that M. surinamensis may have a different evolutionary history from other species in the genus. This is also suggested by the venom-ant ivenom cross-neutraliz- ation data of Bolafios et al. (1978) and possibly by the immunological distance data of Cadle and Sarich (1981). Certainly the biology of this species is radi- cally different from that of all other congeners. Micrurus surinamensis is the only aquatic coral snake, feeding primarily upon fish, particularly Gymnotus carapo and Callichthys callichthys and Symbranchus marmoratus (Schmidt, 1952; Greene, 1973; Dixon and Soini, 1977; Cunha and Nascimento, 1978; Roze, 1982).
Affinities at higher taxonomic levels cannot be ascertained from the data presented here. In three of the five assays where Naja n. kaouthia and Bungarus multicinctus samples yielded significantly different results, most of the Micrurus samples paralleled the Bungarus result. However, this may merely reflect dietary similarities rather than phylogenetic relationships.
Acknowledgements--We wish to thank Eletronorte for granting permission to participate in Operaq~o Jamari and for providing logistical support for collection efforts. We thank Paulo Friedrich Bfihrnheim, Anibal Rafael Melgarejo, Juan Jose Silva Haad and Thales de Lema for providing important venom samples. We are grateful to Marta Regina Magalh~es and Fernanda E. Costa Paulino for animal maintenance, collection of venoms and laboratory work. We also thank Cyro Coimbra de Resende of the Brazilian Ministry of Health for supplying data on ophidism in Brazil. Lastly we appreciate the efforts of Janis Roze in reviewing the manuscript.
REFERENCES
Anonymous (1990) Acidentes ofidicos: contribuiqfio ao estudo da morbidade. Nflmero 41, Ano IV., Secretaria Nacional de Aq6es Bfisicas de Safide, Minst6rio da Sat]de. Brasilia, D. F.
Arroyo O., Rosso J. P., Vargas O., Guti6rrez J. M. and Cerdas L. (1987) Skeletal muscle necrosis induced by a phospholipase A 2 isolated from the venom of the coral snake Micrurus nigrocinctus nigrocinctus. Comp. Biochem. Physiol. 87B, 949-952.
Bolafios R., Cerdas L. and Abalos J. W. (1978) Venoms of coral snakes (Micrurus spp.): report on a multivalent antivenin for the Americas. Bull. Pan. Am. Hlth Org. 12, 23~7.
Bolafios R., Cerdas L. and Taylor R. (1975) The production and characteristics of a coral snake (Micrurus mipartitus hertwigi) antivenin. Toxicon 13, 139-142.
Cadle J. E. and Sarich V. M. A. (1981) An immunological assessment of the phylogenetic position of New World coral snakes. J. Zool. (Lond.) 195, 157-167.
Cohen P., Berkeley W. H. and Seligmann E. B., Jr (1971) Coral snake venoms. In vitro relation of neutralizing and precipitating antibodies. Am. J. trop. med. Hyg. 20, 646-649.
Cohen P., Dawson J. H. and Seligmann E. B., Jr (1968) Cross-neutralization of Micrurus fulvius fulvius (coral snake) venom by anti-Micrurus carinacauda dumerilii serum. Am. J. trop. med. Hyg. 17, 308-310.
Cohen P., Seligmann E. B., Jr and Berkeley W. H. (1967) Coral snake venom: antibody response in rabbits. Nature 213, 820-822.
Cunha O. R. and Nascimento F. P. (1978) Ofidios da Amaz6nia. X. As cobras da regi~.o leste do Parfi. Publica~:6es Avulsas do Museu Emilio Goeldi No. 31, pp. 1-218.
Dal6 N., Perales J., Mufioz R., Martinez B. and Moussatch6 H. (•989) Neuromuscular blocking activity of a fraction isolated from the coral snake venom, Micrurus nigrocinc- tus. Toxicon 27, 40.
Dixon J. R. and Soini P. (1977) The reptiles of the upper Amazon Basin, Iquitos region, Peru. II. Crocodilians, turtles and snakes. Contr. BioL Geol. Milwaukee PubL Mus. 12, 1-91.
Eaker D. (1978) Studies on presynaptically neurotoxic and myotoxic phospholipases A2. In Versatih'ty of Proteins (Edited by Li C. H.), Proceedings of the International Symposium on Proteins, Taipei, Taiwan, pp. 413-431. Academic Press, New York.
Gomez-Leiva M. A. (1978) Comparative study of the hemolytic activity of venoms of Micrurus nigrocinctus, Bungarus multicinctus and Naja naja. In Toxins--Animal, Plant and Microbial (Edited by Rosenberg P.), p. 433. Pergamon Press, Oxford.
Greene H. W. (1973) The food habits and feeding behavior of New World coral snakes. M. A. thesis, University Texas at Arlington, 66 pp.
Guti6rrez J. M., Arroyo O., Chaves F., Lomonte B. and Cerdas L. (1986) Pathogenesis of myonecrosis induced by coral snake (Micrurus nigrocinctus) venom in mice. Br. J. exp. Path. 67, 1-12.

Enzymatic composition of Micrurus venoms 293
Guti6rrez J. M., Chaves F., Rojas R. and Bolafios R. (1980) Efectos locales inducidos por el veneno de la serpiente coral Micrurus nigrocinctus en Rat6n Blanco. Toxicon 18, 633-639.
Guti6rrez J. M., Lomonte B., Portilla E,, Cerdas L. and Rojas E. (1983) Local effects induced by coral snake venoms: evidence of myonecrosis after experimental in- oculations of venoms from five species. Toxicon 21, 777-783.
Halpert J. and Eaker D. (1976) Isolation and amino acid sequence of a neurotoxic phospholipase A from the venom of the Australian tiger snake Notechis scutatus scutatus. J. biol. Chem. 251, 7343-7347.
Heinrikson R. L., Krueger E. T. and Keim P. S. (1977) Amino acid sequence of phospholipase A2-~ from the venom of Crotalus adamanteus. J. biol. Chem. 252, 4913-4921.
Jim6nez-Porras J. M. (1973) Reptile toxins, In Biology Data Book, Vol. II, p. 697. FASEB, USA.
Joubert H. J. (1977) Naja mossambica mossambica venom. Purification, some properties, and the amino acid se- quences of three phospholipases A (CM-I, CM-II and CM-III). Biochim. biophys. Acta 493, 216-227,
Joubert H. J. and Taljaard N. (1980) Purification, some properties and amino acid sequences of two phospho- lipases A (CM-II and CM-III) from Naja naja kaouthia venom. Eur. J. Biochem. 112, 493-499.
Karlsson E. and Pongsawasdi P. (1980) Purification of two phospholipase A isoenzymes with anticoagulant activity from the venom of the cobra Naja naja siamensis. Toxicon 18, 409-419.
Kitchens C. S. and Van Mierop L. H. S. (1987) Envenoma- tion by the eastern coral snake (Micrurusfulviusfulvius). JAMA 258, 1615-1618.
Koerner J. F. and Sinsheimer R. L. (1957) A deoxyribo- nuclease from calf spleen. II. Mode of action. J. biol. Chem. 228, 1049-1062
Kumar V., Rejent T. A. and Elliott W. B. (1973) Anti- cholinesterase activity of elapid venoms. Toxicon 11, 131-138.
L6bo de Arafijo A. and Radvanyi F. (1987) Determination of phospholipase A 2 activity by a colorimetric assay using a pH indicator. Toxicon 25, 1181-1188.
Lu H.-S. and Lo T. B. (1978) Complete amino acid sequence of a new type of cardiotoxin of Bungarusfasciatus venom. Int. J. Pept. Protein Res. 12, 181-183.
McCollough N. C. and Gennaro J. F., Jr (1963) Coral snake bites in the United States. J. Florida reed. Ass. 49, 968-972.
Mochca-Morales J., Martin B. M., Zamudio F. Z. and Possani L. D. (1990) Isolation and characterization of three toxic phospholipases from the venom of the coral snake Micrurus nigrocinctus, p. 95, Abstracts, Third Sym- posium on Animal, Plant, and Microbial Toxins. Inter- national Society on Toxinology, Pan American Sect.
Moseley T. (1966) Coral snake bite: recovery following symptoms of respiratory paralysis. Ann. Surgery 163, 943-948.
Moussatch6 H. and Mel6ndez T. (1979) Some pharmaco- logical observations with Elapidae and Crotalidae snake venoms in the guinea pig denervated diaphragm on the specificity of the cholinergic blockade by their venoms. Rev. Bras. Biol. 39, 605~I0.
Moussatch6 H., Yates A., Mel6ndez T. and Mendoza J. (1976) Preliminary report on the fractionation of the venom of Micrurus nigrocinctus and the pharmacological properties of the fractions. Abstracts, 5th International Symposium on Animal, Plant and Microbial Toxins, San Jos& Costa Rica, p. 60. Universidad de Costa Rica, San Jos6.
Ouyang C. and Huang T. F. (1983) Inhibition of platelet aggregation by 5'-nucleotidase purified from Trimeresurus gramineus snake venom. Toxicon 21, 491-501.
Parrish H. N. and Kahn M. S. (1967) Bites by coral snakes: report of 11 representative cases. Am. J. med. Sci. 253, 561-568.
Patr,~o-Lomba M., Fontana M. D. and Vital Brazil O. (1973) Sobre o mecanismo da aq~o neuromuscular de pe~onha de Micrurus frontalis. Acta phys. Latinoameri- cana 23, 674.
Pettigrew L. C. and Glass J. P. (1985) Neurologic compli- cations of a coral snake bite. Neurology 35, 589-592.
Possani L. D., Alag6n A. C., Fletcher P. L., Jr, Varela M. J. and Julifi J, Z. (1979) Purification and characteriz- ation of a phospholipase A 2 from the venom of the coral snake, Micrurus fulvius microgalbineus. Biochem. J. 179, 603-606.
Pukrittayakamee S., Warrell D. A., Desakorn V., McMichael A. J., White N. J. and Bunnag D. (1988) The hyaluronidase activities of some southeast Asian snake venoms. Toxicon 26, 629~537.
Ramsey H. W., Snyder G. K., Kitchen H. and Taylor W. J. (1972a) Fractionation of coral snake venom. Preliminary studies on the separation and characterization of the protein fractions. Toxicon 10, 67-72.
Ramsey H. W., Snyder G. K. and Taylor W. J. (1971) The effect of Micrurusf. fulvius (Coral) venom on myocardial contractility of the isolated perfused rabbit heart. Clin. Res. 19, 66.
Ramsey H. W., Taylor W. J., Boruchow I. B. and Snyder G. K. (1972b) Mechanism of shock produced by an elapid snake (Micrurus f. fulvius) venom in dogs. Am. J. Phys. 222, 782-786.
Resende C. C., Arafijo F. A. A. and Sallenave R. N. V. R. (1989) Amilise epidemiol6gica dos acidentes ofidicos: Brasil--junho de 1986 a dezembro de 1987. Secretaria Nacional de Aq6es Bfisicas de Safide, Minist6rio da Safide Brasilia D. F.
Rosenfeld G. (1971) Symptomatology, pathology, and treat- ment of snake bites in South America. In Venomous Animals and their Venoms. Vok II, Venomous Vertebrates (Edited by Biicherl W. and Buckley E.), pp. 345-384. Academic Press, New York.
Roze J. A. (1982) New world coral snakes (Elapidae): a taxonomic and biological summary. Mere. Inst. Butantan 46, 305-338.
Roze J. A. (1989) New species and subspecies of coral snakes, Genus Micrurus (Elapidae), with notes on type specimens of several species. Am. Mus. Novitates 2932, 1-15.
Roze J. A. and Bernal-Carlo A. (1987) Las serpientes corales venenosas del genero Leptomicrurus (Serpentes, Elapidae) de Suram6rica con descripci6n de una nueva subespecie. Boll. Mus. Reg. Sci. Nat. Torino 5, 573~08.
Roze J. A. and Jorge da Silva N. (1990) Coral snakes (Serpentes, Elapidae) from hydroelectric power plant of Samuel, Rond6nia, Brazil, with description of a new species. Bull. Aid. Herp. Soc. (in press).
Schenberg S. (1959) Geographical pattern of crotamine distribution in the same rattlesnake subspecies. Science 129, 1361-1363.
Schmidt K. P. (1952) The Surinam coral snake, Micrurus surinamensis. Fieldiana Zool. 34, 25-34.
Snyder G. K., Ramsey H. W., Taylor W. J. and Chiou C. Y. (1973) Neuromuscular blockade of chick biventer cervicis nerve-muscle preparations by a fraction from coral snake venom. Toxicon 11, 505-508.
Stevan L. J. and Seligmann E. B., Jr (1970) Agar-gel and acrylamide-disc electrophoresis of coral snake venoms. Toxicon 8, 11-14.
Tan N.-H. and Tan C.-S. (1988) A comparative study of cobra (Naja) venom enzymes. Comp. Biochem. Physiol. 90B, 745-750.
Tsai I.-H., Wu S.-H. and Lo T.-B. (1981) Complete amino acid sequence of a phospholipase A2 from the venom of Naja naja atra (Taiwan cobra). Toxicon 19, 141-152.

294 STEVEN D. Aim) and NELSON JORGE DA SILVA, JR
Vital Brazil O. (1965) A ~ o neuromuscular da peqonha de Micrurus. 0 Hospital 68, 183-224.
Vital Brazil O. (1980) Venenos ofidicos neurot6xicos. Rev. Ass. Med. Brasil. 26, 212-218.
Vital Brazil O. (1987) Coral snake venoms: mode of action and pathophysiology of experimental en- venomation. Rev. Inst. reed. Trop. Sao Paulo 29, 119-126.
Vital Brazil O. and Fontana M. D. (1984) A~6es pr6- juncionais e p6sjuncionais da peqonha da cobra coral Micrurus corallinus na jung:~o neuromuscular. Mere. Inst. Butantan 47/48, 13-26.
Vital Brazil O., Fontana M. D. and Pellegrini Filho A. (1977) Physiopathologie et th6rapeutique de l'enveno- mation exp6rimentale caus6e par le venin de Micrurus frontalis. Mem. Inst. Butantan 40/41, 221-240.
Weis R. and McIsaac R. J. (1971) Cardiovascular and muscular effects of venom from coral snake, Micrurus fulvius. Toxicon 9, 219-228.
Weissbach H., Robertson A. V., Witkop B. and Udenfriend S. (1960) Rapid spectrophotometric assays for snake venom L-amino acid oxidase based on the oxidation of L-kynurenine or 3,4-dehydroproline. Analyt. Biochem. 1, 286-290.













![Peptidomic Analysis of Animal Venoms - IntechOpen · Peptidomic Analysis of Animal Venoms ... Venomous and poisonous invertebrates include cnidarians [1, 2] (sea anemones, jellyfish](https://img.pdfslide.net/doc/110x75/5f0a94497e708231d42c5393/peptidomic-analysis-of-animal-venoms-intechopen-peptidomic-analysis-of-animal.jpg)















