Embed Size (px)
Citation preview

q 2003 The Paleontological Society. All rights reserved. 0094-8373/00/2904-0010/$1.00
Paleobiology, 29(4), 2003, pp. 576–604
Comparative geographic and environmental diversity dynamicsof gastropods and bivalves during the Ordovician Radiation
Philip M. Novack-Gottshall and Arnold I. Miller
Abstract.—Bivalves and gastropods, prominent members of the Modern Evolutionary Fauna, aretraditionally noted for sharing remarkably similar global diversity trajectories and environmentaldistributions throughout the Phanerozoic. By comparing their fossil occurrences at several scaleswithin a finely resolved geographic, environmental, and temporal framework, it is possible to eval-uate whether such similarities are caused primarily by intrinsic macroevolutionary factors or ex-trinsic ecological factors. Using a database of 7779 global gastropod and bivalve genus occurrences,we investigate the geographical and environmental attributes of bivalves and gastropods duringthe Ordovician Period at scales ranging from global, to a comparison among five paleocontinents,to an intracontinental comparison of four regions within Laurentia. Although both classes sharedstatistically indistinguishable global diversity trajectories and broadly similar environmental dis-tributions during the Ordovician, their environmental distributions differed in several significantfeatures. Furthermore, the diversity trajectories and environmental distributions of these classesdiffered significantly among paleocontinents and among regions within Laurentia. Bivalves wereconsistently most diverse in deeper water, siliciclastic-rich settings in higher-latitude paleoconti-nents whereas gastropods were consistently most diverse in shallower, carbonate-rich settings inmore-equatorial paleocontinents. Notably, these environmental differences were robust to chang-ing physical parameters within paleocontinents, with each class consistently tracking its preferredenvironmental setting. These results suggest that environmental factors played significant, albeitdistinct, roles in the Ordovician diversifications of gastropods and bivalves. However, their similarglobal diversity trajectories suggest that shared, intrinsic macroevolutionary attributes also mayhave played an important role in the evolution of these classes during the Ordovician Radiation.
Philip M. Novack-Gottshall. Department of Biology, Box 90338, Duke University, Durham, North Caro-lina 27708-0338. E-mail: [email protected]
Arnold I. Miller. Department of Geology, Post Office Box 210013, University of Cincinnati, Cincinnati,Ohio 45221-0013. E-mail: [email protected]
Accepted: 23 April 2003
IntroductionGiven the wide variation in diversity trajec-
tories of marine groups through the Phaner-ozoic, it is remarkable that gastropods and bi-valves consistently show similar diversity tra-jectories (Sepkoski 1981; Bambach 1985), back-ground turnover rates at several taxonomicscales (Stanley 1985; Raup and Boyajian 1988;Valentine et al. 1991), and extinction rates dur-ing mass extinctions (Erwin 1990). It is un-likely that such overwhelming similarities aremere coincidence: one may expect an intrinsicor extrinsic cause. With such similarities, how-ever, it is not surprising that the global Phan-erozoic diversity histories of these classes cor-relate when subjected to quantitative analyses(Sepkoski 1981, 1984). Combined with theirprominence in the post-Paleozoic, these simi-larities led to their characterization as domi-nant members of the Modern EvolutionaryFauna (Sepkoski 1981).
Implicit in Sepkoski’s (1984, 1991) kineticmodels of Phanerozoic global diversificationwas the assumption that shared intrinsic mac-roevolutionary dynamics were the drivingforce behind the diversity histories and sub-sequent replacement of each evolutionary fau-na, including gastropods and bivalves. Al-though this model of evolutionary successionwas originally formulated from global diver-sity data, an analysis of Paleozoic fossil com-munities from Laurentia along an environ-mental gradient illustrated that the evolution-ary faunas, to a large degree, had distinct en-vironmental, as well as temporal, patterns(Sepkoski and Sheehan 1983; Sepkoski andMiller 1985). Furthermore, the faunas dis-played a general onshore-offshore pattern inwhich succeeding evolutionary faunas origi-nated in shallow water and expanded intodeeper-water environments during their his-tory (Jablonski et al. 1983; Sepkoski and Shee-

577CLAMS AND SNAILS AT ORDOVIC SCALES
han 1983; Sepkoski and Miller 1985; Sepkoski1991; but see Westrop and Adrain 1998). Inthese onshore-offshore models, the ModernEvolutionary Fauna, and particularly gastro-pods and bivalves, were characterized pri-marily as limited to nearshore and lagoonalhabitats from their initial radiations until theDevonian, when they also became establishedin deep-water environments (e.g., Kammer etal. 1986).
Such observations were intended as gener-alizations about Paleozoic marine communityevolution, but their support in other studies(e.g., Bretsky 1968, 1969, 1970a,b; Boucot 1975;Steele-Petrovic 1979; Westrop et al. 1995) cre-ated an expectation that early Paleozoic gas-tropods and bivalves were restricted to near-shore, shallow-water environments. It was notuntil the more focused work of Frey (1987)and Miller (1988, 1989) that limitations inthese models were recognized by demonstrat-ing, for example, that diverse and abundantLate Ordovician bivalve faunas of the Cincin-nati Arch also thrived in fine-grained, silici-clastic sediments representing offshore envi-ronments. Further limitations included the ob-servation that bivalves and trilobites, mem-bers of different evolutionary faunas,commonly overlapped in their environmentaldistributions (see also Westrop and Adrain1998).
The greatest limitation, however, of theseonshore-offshore models was their reliance ondata from the single paleocontinent of Lauren-tia. For mass extinctions, a limited geographicfocus may not necessarily confound studies,as these processes are, by definition, globalevents of limited temporal duration. For ex-ample, Raup and Jablonski (1993; Jablonskiand Raup 1995) showed no differential geo-graphic overprint on marine bivalve extinc-tions during the end-Cretaceous extinction. Itis now clear, however, that evolutionary radi-ations may display significant heterogeneityamong paleocontinents. Although much ofthis work has focused on the Ordovician Ra-diation (e.g., Miller 1997a, 1998 for global fau-nas; Cope and Babin 1999 for bivalves; Wes-trop and Adrain 1998 for trilobites), other re-search on the recovery from the end-Creta-ceous mass extinction (Jablonski 1998 for
molluscs) has also suggested differential geo-graphic patterns. For the Ordovician, expla-nations for such heterogeneity have most of-ten involved local physical factors, such assedimentology and tectonism (Babin 1993,1995; Miller and Mao 1995; Miller 1997a,b) oroceanographic and sea level changes (Guens-berg and Sprinkle 1992; Patzkowsky and Hol-land 1996). Jablonski (1998) also stressed thatbiotas recovering from mass extinctions mayalso display geographically heterogeneous di-versification dynamics for reasons not directlyrelated to ecological features, a compelling ex-tension of the so-called evolutionary foundereffect of Raup (1979).
A comparison of the dynamics of individ-ual taxonomic groups has great potential forrevealing the causal processes of the Ordovi-cian Radiation and in teasing apart the oftensubtle distinctions between intrinsic macro-evolutionary dynamics and extrinsic ecologi-cal factors. However, the usefulness of such acomparison has been realized in only a fewstudies (Miller 1998). In a series of elegant pa-pers, Babin (1993, 1995; Cope and Babin 1999)proposed that the Ordovician Radiation of bi-valves was a two-step process, in which EarlyOrdovician radiations began in siliciclastic,Gondwanan shallow shelf platforms, whereaslater Ordovician radiations of bivalves in oth-er paleocontinents, including Laurentia, Bal-tica, Siberia, and Kazakhstania, occurred pri-marily in carbonate settings (but see resultsbelow). Similarly, Guensberg and Sprinkle(1992) hypothesized that the Early Ordoviciandiversification of attached echinoderms inwestern Laurentia was prompted by the in-hibition of siliciclastic influx and the concom-itant creation of carbonate hardgroundscaused by global sea level rise. Miller (1997b;Miller and Mao 1995; Miller and Connolly2001) emphasized the role of tectonically driv-en sedimentation as a cause of the OrdovicianRadiation for several orders of brachiopods,trilobites, and benthic molluscs (but see Con-nolly and Miller 2002). It is clear that explain-ing the causes of evolutionary radiations is nota simple endeavor; different causes may befound at different taxonomic levels, differentgeographic scales, and over different timeframes. A fuller understanding requires that

578 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
TABLE 1. Sample record in global database of Ordovi-cian gastropods, bivalves, and monoplacophorans. Fordiscussion of the primary data fields, see text.
GenusGenus synonymSpeciesClassPreservationFormation
GlyptarcaHemiprionodontalusitanicaBivalviaInternal mold/steinkernFonte da Horta
Lithology
Lithcode
Mudstone interbeddedwith sandstone
SiliciclasticEnvironmentEnvironmental zoneCountryLocality
Deepshelf4.5PortugalBucaco Syncline
Secondary localitiesLatitudeLongitudePaleocontinent
Dornes, Cacemes408N0088WIberia (Mediterranean
Province)Harland SubsystemHarland Series
M-OrdL/U-Llde
New Standard SubsystemNew Standard SeriesReference
M-OrdU-DarwYoung 1985
Revision referenceFile number
Cope 1996: p. 991911
these evolutionary phenomena be evaluated atall of these scales. Among the unresolvedquestions is the extent to which prominentmembers of the same evolutionary fauna ex-hibit similar diversification dynamics at allscales: are similarities in global diversity tra-jectories manifested paleoecologically at morelocal levels?
Here, we evaluate the extent to which bi-valves and gastropods exhibited similar geo-graphic and environmental patterns duringthe Ordovician at scales ranging from theglobe, to paleocontinents, to regions withinthe paleocontinent of Laurentia. Using a lit-erature-derived database, we demonstratethat the similarity of the Ordovician historiesof bivalves and gastropods depends on thescale at which the analysis is conducted. Asexpected, both gastropods and bivalves showremarkably similar Ordovician diversity tra-jectories on a global scale, thus lending sup-port to the synoptic model of global evolu-tionary faunas. However, when these samedata are evaluated at the scales of paleoconti-nents or regions within Laurentia, a morecomplex picture of gastropod and bivalve evo-lution during the Ordovician Radiation devel-ops, in which global similarities are the resultof the accumulation of different paleoecolog-ical and evolutionary histories in different re-gions and environments throughout theworld.
Structure of the Database
The database for this study consists of 8237Ordovician fossil occurrences compiled from1090 faunal lists described in 407 paleoecolog-ical, systematic, and biostratigraphic worksfrom the global literature. These occurrencesrepresent 364 genera and 1728 species of gas-tropods, bivalves, and monoplacophoransthat were revised to accord with cladistic (e.g.,Wagner 1995a, 1999) and traditional phylo-genetic relationships (e.g., Cope 1996, 1997;Wahlman 1992). Compiled data also includegeography, lithology, inferred depositionalenvironment, mode of preservation, andstratigraphic interval of occurrence (Table 1).All fossil occurrences reported within a singlelithological unit representing a single depo-sitional environment at a single geographical
coordinate of a distinct age were pooled intoa faunal list. In cases where a stratigraphic in-terval represented more than one depositionalenvironment, the fauna was divided, whenpossible, into separate lithofacies to representthe immediate surrounding strata of each fau-nal list. However, although each faunal list ismost likely a good representation of the time-averaged fossil assemblage from a particularenvironmental setting, it is possible that allmembers of a faunal list did not co-occur si-multaneously. A complementary study (No-vack-Gottshall and Miller in press) presentsthe results of a field-based analysis of mollusc-rich strata in the Cincinnati Arch, testing forlocal co-occurrence in both taxonomic rich-ness and individual abundance at the scale offormations and single bedding planes.
Although the focus of this study is on gas-tropod and bivalve co-occurrence, we includ-ed monoplacophorans in the database becauseof the convoluted historical relationships ofmany bellerophont molluscs (see review inHarper and Rollins 2000); monoplacophoranresults are not presented here. Because thesethree classes shared similar shell chemistriesduring the Ordovician and were commonly

579CLAMS AND SNAILS AT ORDOVIC SCALES
preserved in the same ways, they serve astaphonomic control groups for one another(sensu Bottjer and Jablonski 1988). Such con-trol groups are helpful when evaluating theenvironmental distributions of bivalves andgastropods to determine whether the absenceof a particular class in a sampled unit is likelycaused by non-preservation or by true ecolog-ical absence.
We used lithological information to infer ageneral onshore-offshore environmental zo-nation for approximately two-thirds of thefaunal lists. The remaining lists either lackedlithological information, represented numer-ous depositional environments, or were am-biguously interpreted. This eight-zone frame-work is adopted from that of Sepkoski (1988)and is similar to others in the literature (e.g.,Sepkoski and Sheehan 1983; Sepkoski andMiller 1985; Miller 1988; Bottjer and Jablonski1988; Brett et al. 1993; Jablonski et al. 1997).Environmental zones were based on rock typeand sedimentary structures rather than thepresence of faunal elements, which is typicallyused when designating Benthic Assemblages(sensu Boucot 1975, 1981). The shallowest set-ting, zone 1, is indicative of peritidal to shor-eface settings characterized by desiccationcracks or large-scale cross-bedding. Zone 2 ischaracterized by protected, very shallow sub-tidal settings such as lagoons, that often con-tain restricted faunas. Zone 3 is characterizedby offshore wave-agitated environments in-cluding bars and oolitic shoals with tabular ortrough cross-bedding. We included reefs andother bioherm-rich settings in this zone, incontrast with Sepkoski (1988) and others whoexcluded them from study. Open marine shelfand deltaic settings are subdivided into shal-low (zone 3.5), middle (zone 4), and deep(zone 4.5) subtidal components on the basis offrequency of storm influence and the ratio oftempestites to fine-grained shales and mud-stones (Speyer and Brett 1988), with zone 3.5dominated by tempestite sandstones, grain-stones, and/or packstones and zone 4.5 dom-inated by shales with fewer tempestite wack-estones and/or siltstones. Zone 5 is character-ized by deep-shelf, pro-deltaic, and shelf edgesettings below storm wave base and generallylacking benthic calcareous algae. The deepest
setting, zone 6, is indicative of slope and ba-sinal settings and is characterized by turbi-dites and by hemipelagic and oxygen-deplet-ed facies.
It is important to note that zones 1 through5 are all relatively shallow from an oceano-graphic perspective. The boundary betweenzone 3 and 3.5 is defined here as fair-weatherwave base; likewise, the boundary betweenzones 4 and 5 is defined here as storm wavebase, which can vary between 10 and 50 m inmodern settings (Brett et al. 1993). When in-ferring the depositional environment of faunallists in this study, we made determinations inconsultation with appropriate sequence strat-igraphical and sedimentological works (e.g.,Crevello et al. 1989; Cope et al. 1992; Witzkeet al. 1996).
The database encompasses occurrencesfrom Australasia, Avalonia, Baltica, Laurentia,the Mediterranean Province, Siberia, Ka-zakhstania, Gondwanan South America, andvarious oceanic terranes, including the Pre-cordillera. However, because of limited dataand monographic biases from several of thelatter regions, results of paleocontinentalcomparisons are illustrated in detail here onlyfor the first five of these paleocontinents. Nev-ertheless, as the available data in the addition-al, less studied regions are still concordantwith the results from the five better-exploredpaleocontinents, qualitative summaries ofthese regions will be presented in the subse-quent discussion of paleocontinental trends.
Because of this global coverage, we used theOrdovician timescale of Webby (1998), whichmore accurately reflects global correlationsthan previous efforts; 79% of the occurrenceswere resolvable stratigraphically to the sub-series level used in analyses of diversity tra-jectories and 90% were resolvable to the serieslevel used in analyses of environmental dis-tribution. Webby (1998) does not subdividethe Ordovician series, so we used the follow-ing British stages as subseries equivalents:lower Tremadocian is Cressagian, middleDarriwilian is Abereiddian, middle Carado-cian is Burrellian/Cheneyan, and middle Ash-gillian is Cautleyan/Rawtheyan; the remain-ing subseries are easily inferred.
The nine paleocontinents in the database

580 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
encompass the majority of the Ordovicianglobe, excluding North and South China andportions of South Africa, Antarctica, and theNear and Middle East. Although China haddiverse marine biotas throughout the Ordo-vician (e.g., Wang et al. 1987; BGMRGP 1988;Rigby et al. 1995; Wang and Chen 1999; sum-marized in Miller 1997a; Miller and Mao1998), their reported molluscan diversity wasrelatively minor and composed of cosmopol-itan genera. Further systematic studies of themollusc fauna are encouraged, but the addi-tion of currently published data from these re-gions to our database is unlikely to change theresults presented here.
Because the compilation for Laurentia wasthe most extensive of any paleocontinent inthe database and because there was significanttemporal variation in environmental condi-tions within this paleocontinent during theOrdovician, we analyzed the radiation at finergeographical scales within four regions of thispaleocontinent: Appalachian Basin, Cincin-nati Arch, Mississippi Valley, and Great Basin.These regions comprise 87% of the occurrenc-es in the Laurentian database and thus likelyrepresent the cumulative dynamics of Lauren-tia as a whole. Additionally, during the LateOrdovician, they composed a broad transectfrom predominantly siliciclastic settings inthe eastern Appalachian Basin proximal to theTaconic Orogeny, to mixed-siliciclastic-car-bonate settings in the Cincinnati Arch andMississippi Valley showing orogenic influencemainly in the form of fine-grained siliciclas-tics, and, finally, to predominantly carbonatesin the Great Basin at the most distal portion oforogenic influence. This variation in environ-mental settings serves as a backdrop againstwhich to evaluate the paleoecological charac-teristics of these classes.
Sampling intensity for data used to compilediversity trajectories and environmental dis-tributions for the various paleocontinents andregions within Laurentia are listed in Appen-dices 1 and 2; information includes the num-ber of faunal lists; the total number of gastro-pod, bivalve, and monoplacophoran occur-rences; and the number of primary referencesfrom which faunal lists were compiled. Dif-ferences in tallied data between the two ap-
pendices are a result of the different criteriaused in compiling each data set. The four Lau-rentian regions and the paleocontinents, ex-cept for Laurentia, are all comparably sam-pled. The greater sampling intensity in Lau-rentia likely results from its greater Ordovi-cian rock volume instead of a biased samplingof the literature (Ross et al. 1982).
Despite this overall comparability, the fossilrecord during certain intervals in some paleo-continents and regions may be less well doc-umented, because of diagenetic alteration,lack of exposed strata, or lack of recent sys-tematic study. The first two biases should af-fect bivalves and gastropods equally, whereasthe last bias is lessened by the relatively stablegenus-level taxonomy of these classes and therelatively small number of Ordovician bivalveand gastropod systematists (Schneider 2001;Wagner 2001). Baltica, the MediterraneanProvince, and Laurentia contain generallywell-studied and well-documented molluscanrecords throughout the Ordovician. In con-trast, Australasia, Avalonia, and the Lauren-tian Great Basin suffer from one or more ofthese sampling limitations during at least oneinterval, the details of which are described be-low. Although in some cases faunal lists weresufficient to make statistical comparisons forthese venues, their interpretation in these lim-ited instances should be considered prelimi-nary pending corroboration from additionalfield effort. As the primary aim of this study,however, is a comparison, at several scales, ofthe diversification dynamics of gastropodsand bivalves, it is most important that thesampling effort within an interval be compa-rable for both classes in each region; this aimis satisfied given the comparable preservationof these classes.
Methods
Diversity Trajectories. Diversity trajectoriesof taxa through time were calculated on theassumption that taxa range through the inter-val demarcated by their stratigraphical firstand last occurrences. Although this assump-tion makes sense for global compilations, itmay be problematical for smaller geographi-cal scales because of immigration or emigra-tion of taxa (Patzkowsky and Holland 1996).

581CLAMS AND SNAILS AT ORDOVIC SCALES
Lacking evidence to the contrary, we assumedthat a taxon was present throughout its strat-igraphical range within a region.
To compare quantitatively gastropod andbivalve diversity trajectories while avoidingautocorrelation (McKinney 1990), we calculat-ed proportional changes in diversity overtime, or ‘‘first differences’’ (Gould and Callo-way 1980; Foote 1994; Wagner 1995a):
m 5 q • (D 2 D )a amax at at21 (1)
where ma is the proportional change in diver-sity (or slope) between adjacent time intervalst and t21, Da is the diversity in a time interval,and qamax is the reciprocal of the maximum di-versity attained by the class of interest (a) dur-ing the Ordovician Period. Once the slopeswere calculated for both bivalves and gastro-pods, the nonparametric Spearman’s rank cor-relation coefficient was calculated to test thesignificance of the correlation between the twodiversity trajectories.
Environmental Distribution. Environmentaldistributions were determined by countingthe number of unique genera in a specific en-vironmental zone during a time interval; noassumptions were made regarding first andlast occurrences. The data were plotted on atime-environment diagram with time arrayedon the vertical dimension and the environ-mental zonation arrayed along the horizontaldimension. The diversities in each time-envi-ronment cell were enclosed by shaded contourlines, with increasingly darker shades denot-ing greater diversity; unshaded, empty cellscontain no known occurrences of either gas-tropods or bivalves in the database. Tapho-nomic controls, representing the occurrence ofat least one gastropod, bivalve, or monopla-cophoran fossil, were symbolized with a filledcircle within each cell. Because all three clas-ses exhibited similar preservational potentialsduring the Ordovician (Valentine 1989;Palmqvist 1991; Foote and Sepkoski 1999), thepresence of a taphonomic marker (i.e., mono-placophorans) in an environmental cell lack-ing both gastropods and bivalves providesgreater confidence that the absence has trueecological significance. In contrast, the dualabsence of a taphonomic marker and a gastro-pod or bivalve genus in a zone could be the
result of a true absence of all three molluscanclasses or non-preservation of such fossils inthat setting at that time. Because no attemptwas made to determine whether non-mollus-can collections are available for these emptycells, no valid conclusions can be made with-out further information. A comprehensiveanalysis of the Ordovician benthos is beyondthe present study, but our results are evalu-ated, where possible, by comparison with ad-ditional studies.
Because of the limited environmental cov-erage within most paleocontinents and re-gions with Laurentia, analyses of environ-mental distributions at these finer geographicscales were conducted only at the level of Or-dovician series rather than subseries; 64% ofglobal fossil occurrences could be categorizedwithin this framework. Although severaltime-environment cells contain no known re-cord of gastropods, bivalves, or monoplaco-phorans (Appendix 2), the comparability ofsampling effort among the four Laurentian re-gions and the paleocontinents and the similarpreservation potential of these classes allowstatistically meaningful comparisons to bemade.
Because the environmental framework is adiscrete adaptation of a continuous variable,we used the Kolmogorov-Smirnov two-sam-ple test to test the similarity of bivalve andgastropod environmental distributions (Sokaland Rohlf 1995). Distributions were tested atthe level of resolvable subseries for the globalcomparison; data were tested for all geo-graphic scales at the level of Lower (Trema-docian–Latorpian), Middle (Volkhovian–Dar-riwilian), and Upper Ordovician (Carado-cian–Ashgillian) subsystems (Webby 1998).Before testing, data were percent-maximumtransformed by dividing the diversity of eachclass in each time-environmental cell by themaximum diversity attained by that class inany cell during the Ordovician. This transfor-mation ensured that the test among distribu-tions compared the shape of that distributionindependently of confounding differences inthe diversity of each class.
Although useful as a test for statisticallydistinct environmental distributions, the Kol-mogorov-Smirnov test cannot determine the

582 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
FIGURE 1. Global Ordovician diversity trajectories forgastropod and bivalve genera. In this figure, and in allfollowing diversity charts, solid lines depict the gastro-pod trajectory and dotted lines depict the bivalve tra-jectory. The timescale, with time proceeding from left toright, is based on that of Webby (1998), using abbrevi-ations for lower, middle, and upper subseries, where ap-propriate: (T) Tremadocian, (L) Latorpian, (V) Volkhov-ian, (D) Darriwilian, (C) Caradocian, and (A) Ashgilli-an.
basis of such differences. In cases where twodistributions were statistically distinct, weused the one-tailed, nonparametric Mann-Whitney U-test to test whether the environ-mental distribution of one class was skewedsignificantly toward deeper or shallowerdepths than the other (Sokal and Rohlf 1995).These tests were conducted only at the sub-system level to ensure adequate sample sizes.
Global Results and Discussion
Analyses conducted at each of the threegeographic scales are presented separately be-low, with a summary of scale-transcendentand scale-dependent patterns for all scales inthe concluding remarks.
Global Diversity Trajectories. Figure 1 de-picts the global diversity trajectories of gastro-pod and bivalve genera through the Ordovi-cian Period. Gastropods and bivalves showsignificantly correlated trajectories based onfirst differences (rs 5 0.924; p , 0.005), al-though gastropods were typically more di-verse than bivalves throughout the Ordovi-cian. Both classes shared a dramatic initial risein diversity through the Lower Ordovician,stability in the Volkhovian, diversification intothe Caradocian, and descent through the endof the period.
These trajectories are similar to those plot-ted by Sepkoski (1995: Fig. 4) for both of theseclasses and by Wagner (1995a: Fig. 5) for bothphylogenetically and taxonomically classifiedgastropods; minor differences between theirresults and those in Figure 1 are caused by thedifferent timescale used in this study and theindependent compilation of the respective da-tabases. Despite the similarities of these clas-ses, however, it is possible that the raw trajec-tories may not entirely reflect the true Ordo-vician biological history of these classes be-cause no attempt was made to standardizesampling biases (e.g., rock volume or sam-pling intensity) that can substantially alter ob-served diversity patterns (Miller and Foote1996; Alroy 2000a,b; Alroy et al. 2001). How-ever, as the focus of the present study is to testfor overall similarity in Ordovician trajecto-ries of these classes, and assuming reasonablythat the biases are not skewed in favor of anindividual class (Foote and Sepkoski 1999),the lack of such standardization does not af-fect the comparisons presented here.
The Upper Ordovician decrease is in part asampling artifact caused by an absence of datafor Silurian occurrences that eliminates taxaranging through the Ashgillian, i.e., a lack ofthe so-called Pull of the Silurian (sensu Millerand Foote 1996). It likely also reflects the ini-tial phase of the Late Ordovician mass extinc-tion that occurred at the end of the middleAshgillian and is observed in the depleted di-versity of upper Ashgillian (Hirnantian) taxa(Brenchley 1990). The low diversity of bothclasses in the Lower Ordovician is not likelyan artifact of range truncation, because onlyeight gastropod genera have been document-ed from the Upper Cambrian (Wagner 1995a)and only four of those range into the Ordo-vician. Similarly, only four unequivocal bi-valve genera are known from the Cambrian,and none of those are known in the Ordovi-cian (Cope 1997; Cope and Babin 1999).
Global Environmental Distributions. Figure2 displays the global environmental distribu-tions of gastropods and bivalves during theOrdovician Period. Gastropods were relative-ly widespread throughout most Lower Ordo-vician environments (zones 1 through 5), withthe greatest concentration in shallow offshore

583CLAMS AND SNAILS AT ORDOVIC SCALES
FIGURE 2. Contoured time-environment diagram depicting the global genus diversity of bivalves (left) and gas-tropods (right) during the Ordovician. The dark circles within some cells are taphonomic markers that representthe occurrence of at least one gastropod, bivalve, or monoplacophoran fossil (see text for details). The timescale isthe same as in Figure 1. The key at top depicts the genus diversity of each class within shaded contours.
settings (zones 3 and 3.5); in contrast, bivalveswere somewhat more restricted then to shelfsettings (zones 4 through 5), with few generain nearshore settings (zone 1). Thus, althoughboth classes inhabited a similarly wide rangeof environments, bivalves were generallymore diverse in deeper water settings and gas-tropods in shallower ones.
Given the diverse gastropod assemblages inshallow settings (zones 2 through 3.5) duringthis time, it is likely that the absence of bi-valves in these settings is ecologically signifi-cant. Five genera of gastropods are alsoknown from Laurentian lagoonal settings(zone 2) during the Tremadocian (Poulsen1937; Bass and Northrup 1953; Aitken andNorford 1967), but a lack of precise subseriesresolution prevented them from being includ-ed in Figure 1; these genera are included in theseries-level data from Laurentia below.
The apparent bottleneck exhibited by bothclasses in the Volkhovian and lower Darriwil-ian has several possible explanations. Al-though it may reflect an ecologically or envi-ronmentally meaningful restriction, a lack ofwell-studied faunas from non-shelf environ-ments could also contribute to the pattern ob-served; indeed, there are few available faunal
lists during these time intervals (Appendix 2),although the coverage in zones 3.5 and 4 iscomparable to other intervals. Whether real orpreservational, the restriction appears to af-fect more than molluscs; Sepkoski and Shee-han (1983) illustrated a similarly reduced en-vironmental coverage for this time intervalamong benthic fauna of Laurentia. Erosionduring the Sauk-Tippecanoe unconformitythat affected Laurentia, Siberia, and possiblyGondwana and other paleocontinents (Ross etal. 1982; Barnes et al. 1996; J. E. Repetski per-sonal communication 2002) also may haveplayed a role. Regardless of the cause, bothclasses were present in similar diversities inthe same environments during these intervals,and paleontological exploration in additionalfacies could be worthwhile.
In the middle and upper Darriwilian, bothgastropods and bivalves ranged throughoutmost environmental zones with few absences.Throughout the Caradocian and Ashgillian,gastropods continued to display their greatestdiversity in nearshore through midshelf set-tings (zones 1 through 4), withdrawing fromdeeper water (zones 5 and 6) in the Ashgillian;in contrast, bivalves were most diverse indeeper shelf settings (zones 4 through 5).
584 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
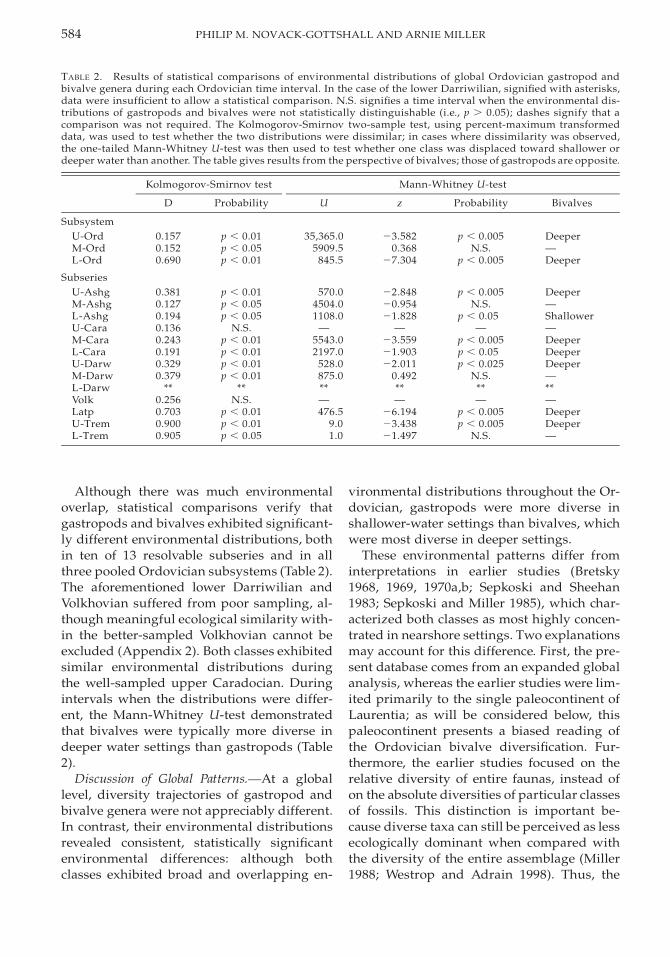
TABLE 2. Results of statistical comparisons of environmental distributions of global Ordovician gastropod andbivalve genera during each Ordovician time interval. In the case of the lower Darriwilian, signified with asterisks,data were insufficient to allow a statistical comparison. N.S. signifies a time interval when the environmental dis-tributions of gastropods and bivalves were not statistically distinguishable (i.e., p . 0.05); dashes signify that acomparison was not required. The Kolmogorov-Smirnov two-sample test, using percent-maximum transformeddata, was used to test whether the two distributions were dissimilar; in cases where dissimilarity was observed,the one-tailed Mann-Whitney U-test was then used to test whether one class was displaced toward shallower ordeeper water than another. The table gives results from the perspective of bivalves; those of gastropods are opposite.
Kolmogorov-Smirnov test
D Probability
Mann-Whitney U-test
U z Probability Bivalves
SubsystemU-OrdM-OrdL-Ord
0.1570.1520.690
p , 0.01p , 0.05p , 0.01
35,365.05909.5
845.5
23.5820.368
27.304
p , 0.005N.S.
p , 0.005
Deeper—Deeper
SubseriesU-AshgM-AshgL-AshgU-CaraM-CaraL-CaraU-DarwM-DarwL-Darw
0.3810.1270.1940.1360.2430.1910.3290.379
**
p , 0.01p , 0.05p , 0.05
N.S.p , 0.01p , 0.01p , 0.01p , 0.01
**
570.04504.01108.0
—5543.02197.0
528.0875.0**
22.84820.95421.828
—23.55921.90322.011
0.492**
p , 0.005N.S.
p , 0.05—
p , 0.005p , 0.05p , 0.025
N.S.**
Deeper—Shallower—DeeperDeeperDeeper—**
VolkLatpU-TremL-Trem
0.2560.7030.9000.905
N.S.p , 0.01p , 0.01p , 0.05
—476.5
9.01.0
—26.19423.43821.497
—p , 0.005p , 0.005
N.S.
—DeeperDeeper—
Although there was much environmentaloverlap, statistical comparisons verify thatgastropods and bivalves exhibited significant-ly different environmental distributions, bothin ten of 13 resolvable subseries and in allthree pooled Ordovician subsystems (Table 2).The aforementioned lower Darriwilian andVolkhovian suffered from poor sampling, al-though meaningful ecological similarity with-in the better-sampled Volkhovian cannot beexcluded (Appendix 2). Both classes exhibitedsimilar environmental distributions duringthe well-sampled upper Caradocian. Duringintervals when the distributions were differ-ent, the Mann-Whitney U-test demonstratedthat bivalves were typically more diverse indeeper water settings than gastropods (Table2).
Discussion of Global Patterns. At a globallevel, diversity trajectories of gastropod andbivalve genera were not appreciably different.In contrast, their environmental distributionsrevealed consistent, statistically significantenvironmental differences: although bothclasses exhibited broad and overlapping en-
vironmental distributions throughout the Or-dovician, gastropods were more diverse inshallower-water settings than bivalves, whichwere most diverse in deeper settings.
These environmental patterns differ frominterpretations in earlier studies (Bretsky1968, 1969, 1970a,b; Sepkoski and Sheehan1983; Sepkoski and Miller 1985), which char-acterized both classes as most highly concen-trated in nearshore settings. Two explanationsmay account for this difference. First, the pre-sent database comes from an expanded globalanalysis, whereas the earlier studies were lim-ited primarily to the single paleocontinent ofLaurentia; as will be considered below, thispaleocontinent presents a biased reading ofthe Ordovician bivalve diversification. Fur-thermore, the earlier studies focused on therelative diversity of entire faunas, instead ofon the absolute diversities of particular classesof fossils. This distinction is important be-cause diverse taxa can still be perceived as lessecologically dominant when compared withthe diversity of the entire assemblage (Miller1988; Westrop and Adrain 1998). Thus, the

585CLAMS AND SNAILS AT ORDOVIC SCALES
generally high diversities of molluscs in Or-dovician shelf settings should not imply thatthese molluscs were the dominant taxa inthese settings; it means simply that these clas-ses were generally more diverse in shelf set-tings than in nearshore ones. The generallyheld notion of gastropod-dominated near-shore settings could as easily arise from an ex-clusion of more typically shelf-restricted bioticelements (e.g., brachiopods, bryozoans, andcrinoids) from nearshore settings as it couldfrom a propensity of these molluscs to be re-stricted to nearshore environments (Westropand Adrain 1998). Several cases are known,however, in which bivalves and gastropodswere the dominant faunal elements in Ordo-vician shelf environments. For example, sev-eral studies (Frey 1987; Miller 1989; Patz-kowsky and Holland 1999; Novack-Gottshalland Miller in press) have demonstrated Mid-dle and Late Ordovician deep subtidal set-tings where bivalves constituted more thanhalf of the genera and total number of indi-viduals within single assemblages, and shal-low settings where gastropods constitutedsimilar levels of abundance. Regardless of theexplanation, the environmental distributionsof these classes at a global scale were appre-ciably different than the nearly identical di-versity trajectories might lead one to predict.
Paleocontinental Results and Discussion
Paleocontinental Diversity Trajectories andEnvironmental Zones
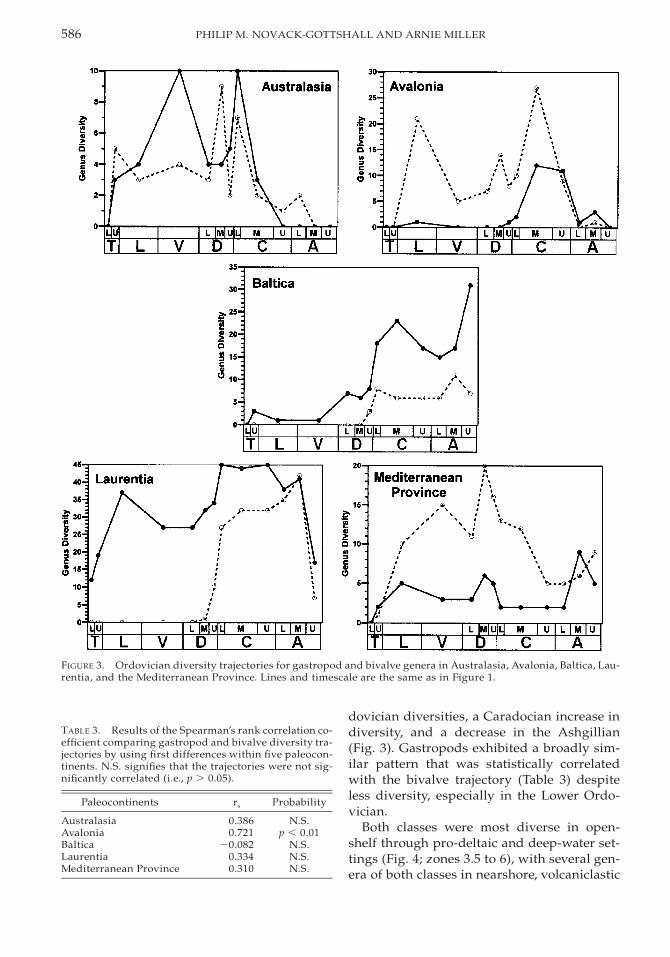
Australasia. During the Early and MiddleOrdovician, Australasia (including present-day Australia, Malaysia, and Thailand) exist-ed as a series of island arcs on the equatoralong the northeastern Gondwanan coastdominated by siliciclastic and mixed-siliciclas-tic-carbonate sedimentation (Cocks 2001). Asthe paleocontinent moved northward duringthe Late Ordovician, the paleocontinent be-came more carbonate-rich (Cocks and Fortey1988, 1997; Scotese and McKerrow 1991; Per-cival 1995). First differences indicate that gas-tropod and bivalve diversity trajectories (Fig.3) were not significantly correlated (Table 3).Gastropods were generally more diverse thanbivalves, although both classes exhibited a de-
cline in diversity through the Upper Ordovi-cian; similar diversity trajectories were re-ported for bivalves by Cope and Babin (1999).Because of poor subseries resolution, the bi-valve-rich Darriwilian Stairway Sandstonefauna (Pojeta and Gilbert-Tomlinson 1977)could not be included in these diversity tra-jectories although it is included in the series-level environmental distributions below. Oncea more precise age determination is made ofthese sediments, the diversity trajectories forAustralasian bivalves and gastropods will be-come even less comparable than presently ob-served. The Upper Ordovician, and particu-larly Ashgillian, decline is caused by both re-duced available outcrop (Cocks and Fortey1990) and a lack of recent taxonomic study(Percival 1995; Cocks and Fortey 1997).
Both classes consistently inhabited open-shelf settings (Fig. 4; zones 3.5 to 4), appearingin deeper settings (zone 4.5 to 5) in the Tre-madocian, and in shallower settings (zones 1to 3) in the Latorpian, Darriwilian, and Car-adocian. Despite such similarities, the envi-ronmental distributions were statistically dif-ferent throughout the Ordovician (Table 4).Although the data were sufficient to statisti-cally evaluate the environmental displacementof these classes during the Lower and MiddleOrdovician, such analyses are difficult to in-terpret given the patchy nature of the data.
Avalonia. During the Early Ordovician,Avalonia lay peripheral to northwesternGondwana in high southern latitudes (Torsvik1998). This island arc terrane separated fromGondwana by the Middle Ordovician andmoved northward toward Laurentia and Bal-tica, reducing the width of the Iapetus Ocean(Cocks and Fortey 1990; Torsvik 1998). Asclosing continued into the Late Ordovician,Avalonia, now in subequatorial climates, su-tured with Baltica, closing the Thor (Tornqu-ist) Ocean (Berthelsen 1998; Torsvik 1998).Sedimentation throughout the Ordovician waspredominantly siliciclastic from prolongedperiods of island arc volcanism (Traynor 1988;Scotese and McKerrow 1991). With the excep-tion of two recent collections (Cope 1996,1999) that significantly elevate the Latorpianand middle Darriwilian bivalve diversities, bi-valves showed reduced Lower and Middle Or-
586 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
FIGURE 3. Ordovician diversity trajectories for gastropod and bivalve genera in Australasia, Avalonia, Baltica, Lau-rentia, and the Mediterranean Province. Lines and timescale are the same as in Figure 1.
TABLE 3. Results of the Spearman’s rank correlation co-efficient comparing gastropod and bivalve diversity tra-jectories by using first differences within five paleocon-tinents. N.S. signifies that the trajectories were not sig-nificantly correlated (i.e., p . 0.05).
Paleocontinents rs Probability
AustralasiaAvaloniaBalticaLaurentiaMediterranean Province
0.3860.721
20.0820.3340.310
N.S.p , 0.01
N.S.N.S.N.S.
dovician diversities, a Caradocian increase indiversity, and a decrease in the Ashgillian(Fig. 3). Gastropods exhibited a broadly sim-ilar pattern that was statistically correlatedwith the bivalve trajectory (Table 3) despiteless diversity, especially in the Lower Ordo-vician.
Both classes were most diverse in open-shelf through pro-deltaic and deep-water set-tings (Fig. 4; zones 3.5 to 6), with several gen-era of both classes in nearshore, volcaniclastic

587CLAMS AND SNAILS AT ORDOVIC SCALES
settings (zone 1) in the Caradocian and a di-verse nearshore bivalve fauna (Cope 1999) inthe Darriwilian. Despite such overall similar-ities, the environmental distributions of theseclasses were statistically different in the Lowerand Middle Ordovician, although the limitedenvironmental coverage prevented recogni-tion of significant displacement with the U-test (Table 4).
Note that the diverse and important Lator-pian bivalve fauna that Cope (1996: p. 980) re-ported from a ‘‘close inshore’’ setting has beenreinterpreted here, following several lithofa-cies analyses of the strata (Traynor 1988; Be-vins et al. 1992), as representing a deep-water,fan-delta below storm wave base (zone 5). Al-though it is possible that this fauna was rela-tively close to shore given the steep slopesalong back-arc basinal deltas (Coleman andPrior 1981), the presence of sediment-gravitydebris flows, hemipelagic muds, and the au-tochthonous nature of the fauna all argue fora zone 5 designation (Traynor 1988).
Baltica. During the Early Ordovician, Bal-tica lay in temperate latitudes on the south-eastern margin of the Iapetus Ocean (Torsvik1998). As the Ordovician progressed, Balticamoved equatorward toward Laurentia and Si-beria, colliding with Laurentia in the Silurian(Torsvik et al. 1996; Cocks and Fortey 1998;Torsvik 1998). Throughout the Ordovician,condensed sedimentation dominated withcool-water carbonate production and minorsiliciclastics (Jaanusson 1973; Podhalanska1995). Gastropod diversity increased throughthe period to higher levels than bivalves (Fig.3), and the trajectories of the two classes werenot significantly correlated (Table 3). The ab-sence of gastropods and bivalves in Lower Or-dovician sediments in this region possibly re-flects ecologically significant patterns, as thesesediments bear moderately rich non-mollus-can faunas (Tjernvik 1956, 1958). However, theuncertain age determination of several gastro-pods from the Tremadocian BjorkasholmenFormation (Yochelson 1962) and the lack ofmore recent systematic study are also signifi-cant factors, although no Lower Ordovicianbivalves have been reported from this richlyfossiliferous region. Surprisingly, gastropods
seem to have been little affected by the LateOrdovician extinction in this region.
Despite limited data from the Lower Or-dovician, environmental distributions of bothclasses overlapped throughout the remainingOrdovician with greatest diversity of gastro-pods in shelf settings (Fig. 4; zones 3.5 to 4.5)and Upper Ordovician reefs (zone 3; Sheehan1979; Harland 1981). Bivalves were rare inthese reefs, moderately diverse in nearshoresettings (zone 1) in the Ashgillian, and mostdiverse in deeper shelf environments (zones 4through 6). Darriwilian bivalves were absentfrom all except the deeper midshelf (zones 4to 4.5), despite the presence of moderately di-verse gastropods in both deeper and shallow-er environments; thus, despite limited num-bers of faunal lists (Appendix 2), their absencehere is likely ecologically significant. Therewere insufficient data to test the environmen-tal distributions for the Lower Ordovician.Their distributions were significantly differ-ent during the Middle Ordovician (Table 4) al-though limited bivalve data precluded a U-test. Despite much overlap in their distribu-tions, Upper Ordovician bivalves wereskewed statistically toward deeper water thangastropods.
Laurentia. Laurentia occupied an equato-rial position throughout the Ordovician Peri-od (Scotese and McKerrow 1991). Early andMiddle Ordovician sedimentation consistedpredominantly of carbonates on passive-mar-gin ramps and relatively shallow cratonic ba-sins with occasional mixed-siliciclastic influx(James et al. 1989; Read 1989; Ross et al. 1989;Holland 1993; Ross and Ross 1995). Siliciclas-tic sedimentation increased significantly ineastern Laurentia during the Middle to LateOrdovician with initiation of the TaconicOrogeny (Holland and Patzkowsky 1996).Carbonate and mixed regimes, however, stillpredominated throughout much of the paleo-continent (Holland 1993; Ross and Ross 1995).Although Laurentia produced more faunallists than all other studied paleocontinents(Appendix 1), identifiable bivalve species arenot recorded until the Darriwilian (Fig. 3);sparse Latorpian bivalves from the SmithvilleFormation of Arkansas remain taxonomicallyindeterminate (Yochelson and Wise 1972). In

588 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER

589CLAMS AND SNAILS AT ORDOVIC SCALES
←
FIGURE 4. Contoured time-environment diagrams depicting the genus diversity of gastropods (above) and bivalves(below) in Australasia, Avalonia, Baltica, Laurentia, and the Mediterranean Province during the Ordovician. Thetimescale is the same as in Figure 1. The key at top depicts the genus diversity of each class within shaded contours.
contrast to the bivalve record, the gastropodfaunas of Laurentia were the most diverse ofany Ordovician paleocontinent (Wagner1999). Consequently, gastropod and bivalvediversity trajectories were not statistically cor-related (Table 3), even when only the UpperOrdovician trajectories were compared (rs 50.700; p . 0.05). Both classes underwent dra-matic radiations in the upper Darriwilian thatcontinued into the Caradocian, coincidentwith the onset of the Taconic Orogeny. Re-gional dynamics of these classes within Lau-rentia are described below, as are the impli-cations of the dramatic change in physical en-vironment brought about by this orogenicevent.
Laurentian gastropods inhabited a range ofLower and Middle Ordovician environments(Fig. 4; zones 1 through 5), although they weremost diverse in nearshore, biohermal, andshallow shelf settings (zones 1 and 3 to 3.5);because of extensive coverage of faunal lists(Appendix 2), the near absence of bivalves inthese environments is significant. When bi-valves diversified in the Middle to Upper Or-dovician, both classes occupied a full range ofenvironments, from nearshore (zone 1) to ba-sinal (zone 6). However, Upper Ordoviciangastropods were most diverse in shallow-wa-ter and shelf settings (zones 1 through 4.5),with an absence from deep water during theAshgillian. Upper Ordovician bivalves wereconsistently most diverse in shelf settings(zones 3.5 through 5), although they were alsodiverse in nearshore and lagoonal settings(zones 1 through 2) in the Ashgillian. Thesepatterns were corroborated statistically, withMiddle Ordovician bivalves skewed towardshallower settings than gastropods, and withthe converse pattern in the Upper Ordovician(Table 4).
Mediterranean Province. The MediterraneanProvince denotes the faunas situated on smallcontinental blocks along the western Gond-wanan margin found in present-day Portugal,Spain, France, the Czech Republic, and the
Moroccan Anti-Atlas (Williams et al. 1995;Cocks and Fortey 1988). During the Ordovi-cian, these blocks bordered the Iapetus Oceanin polar and subpolar latitudes of the South-ern Hemisphere (Scotese and McKerrow1991), dominated by siliciclastic sedimenta-tion in shelf settings (Babin 1993, 1995). Thediversity trajectories of gastropods and bi-valves throughout the period (Fig. 3) were notsignificantly correlated (Table 3), with gastro-pods generally less diverse than bivalves. Al-though this paleocontinent was the second-most intensively sampled, gastropod diversi-ty was lower than any other paleocontinent orregion in Laurentia. Furthermore, gastropoddiversity here was relatively stable throughoutthe Ordovician, with only a minor increase inthe middle Ashgillian. Bivalve diversity, incontrast, increased to a Darriwilian peak anddecreased to the end of the Ordovician, witha slight increase in the upper Ashgillian.
Throughout the Ordovician, both classeswere most diverse in shelf and deep-water set-tings (Fig. 4; zones 3.5 through 6). Minor oc-currences include gastropods in nearshoresettings (zone 1) in the Tremadocian and Up-per Ordovician to the exclusion of bivalves.Additionally, gastropods also occupied shoalsettings (zone 3) throughout the Upper Or-dovician, sharing this setting with bivalves inthe Ashgillian. Despite the limited number offaunal lists from shallower environments (Ap-pendix 2), the absence of bivalves from mod-erately gastropod-rich settings in six instancesthroughout the Ordovician is likely ecologi-cally meaningful. Accordingly, these classeshad distinct environmental distributionsthroughout the Ordovician, although environ-mental displacement was not distinguishablewith the U-test (Table 4).
Discussion of Paleocontinental Patterns
Of the five Ordovician paleocontinents dis-cussed above, only Avalonia showed signifi-cantly correlated diversity trajectories amonggastropod and bivalve genera. Thus, among
590 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
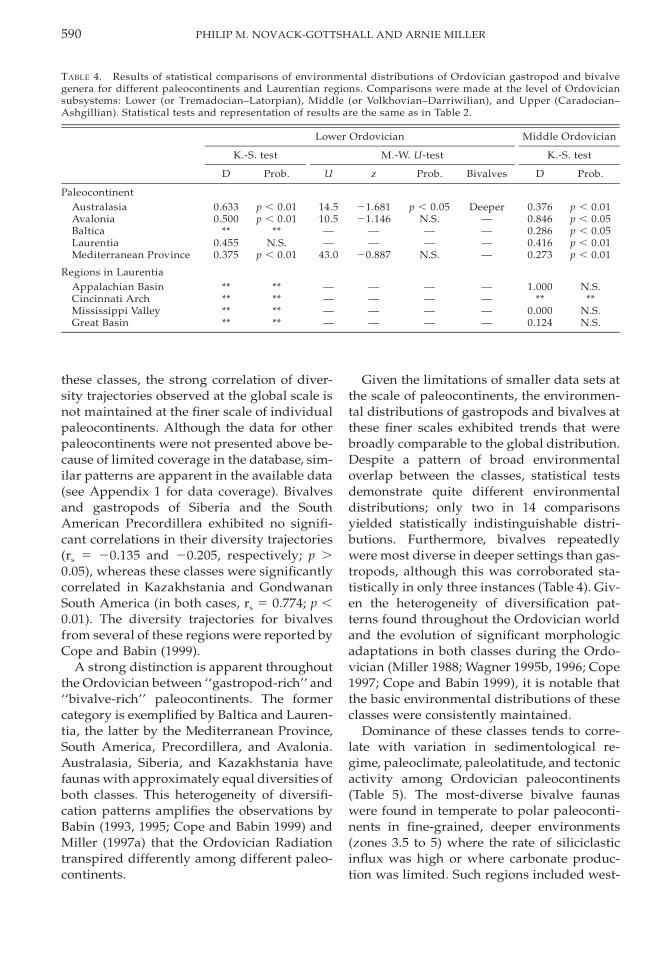
TABLE 4. Results of statistical comparisons of environmental distributions of Ordovician gastropod and bivalvegenera for different paleocontinents and Laurentian regions. Comparisons were made at the level of Ordoviciansubsystems: Lower (or Tremadocian–Latorpian), Middle (or Volkhovian–Darriwilian), and Upper (Caradocian–Ashgillian). Statistical tests and representation of results are the same as in Table 2.
Lower Ordovician
K.-S. test
D Prob.
M.-W. U-test
U z Prob. Bivalves
Middle Ordovician
K.-S. test
D Prob.
PaleocontinentAustralasiaAvaloniaBalticaLaurentiaMediterranean Province
0.6330.500
**0.4550.375
p , 0.01p , 0.01
**N.S.
p , 0.01
14.510.5——
43.0
21.68121.146
——
20.887
p , 0.05N.S.——
N.S.
Deeper————
0.3760.8460.2860.4160.273
p , 0.01p , 0.05p , 0.05p , 0.01p , 0.01
Regions in LaurentiaAppalachian BasinCincinnati ArchMississippi ValleyGreat Basin
********
********
————
————
————
————
1.000**
0.0000.124
N.S.**
N.S.N.S.
these classes, the strong correlation of diver-sity trajectories observed at the global scale isnot maintained at the finer scale of individualpaleocontinents. Although the data for otherpaleocontinents were not presented above be-cause of limited coverage in the database, sim-ilar patterns are apparent in the available data(see Appendix 1 for data coverage). Bivalvesand gastropods of Siberia and the SouthAmerican Precordillera exhibited no signifi-cant correlations in their diversity trajectories(rs 5 20.135 and 20.205, respectively; p .0.05), whereas these classes were significantlycorrelated in Kazakhstania and GondwananSouth America (in both cases, rs 5 0.774; p ,0.01). The diversity trajectories for bivalvesfrom several of these regions were reported byCope and Babin (1999).
A strong distinction is apparent throughoutthe Ordovician between ‘‘gastropod-rich’’ and‘‘bivalve-rich’’ paleocontinents. The formercategory is exemplified by Baltica and Lauren-tia, the latter by the Mediterranean Province,South America, Precordillera, and Avalonia.Australasia, Siberia, and Kazakhstania havefaunas with approximately equal diversities ofboth classes. This heterogeneity of diversifi-cation patterns amplifies the observations byBabin (1993, 1995; Cope and Babin 1999) andMiller (1997a) that the Ordovician Radiationtranspired differently among different paleo-continents.
Given the limitations of smaller data sets atthe scale of paleocontinents, the environmen-tal distributions of gastropods and bivalves atthese finer scales exhibited trends that werebroadly comparable to the global distribution.Despite a pattern of broad environmentaloverlap between the classes, statistical testsdemonstrate quite different environmentaldistributions; only two in 14 comparisonsyielded statistically indistinguishable distri-butions. Furthermore, bivalves repeatedlywere most diverse in deeper settings than gas-tropods, although this was corroborated sta-tistically in only three instances (Table 4). Giv-en the heterogeneity of diversification pat-terns found throughout the Ordovician worldand the evolution of significant morphologicadaptations in both classes during the Ordo-vician (Miller 1988; Wagner 1995b, 1996; Cope1997; Cope and Babin 1999), it is notable thatthe basic environmental distributions of theseclasses were consistently maintained.
Dominance of these classes tends to corre-late with variation in sedimentological re-gime, paleoclimate, paleolatitude, and tectonicactivity among Ordovician paleocontinents(Table 5). The most-diverse bivalve faunaswere found in temperate to polar paleoconti-nents in fine-grained, deeper environments(zones 3.5 to 5) where the rate of siliciclasticinflux was high or where carbonate produc-tion was limited. Such regions included west-
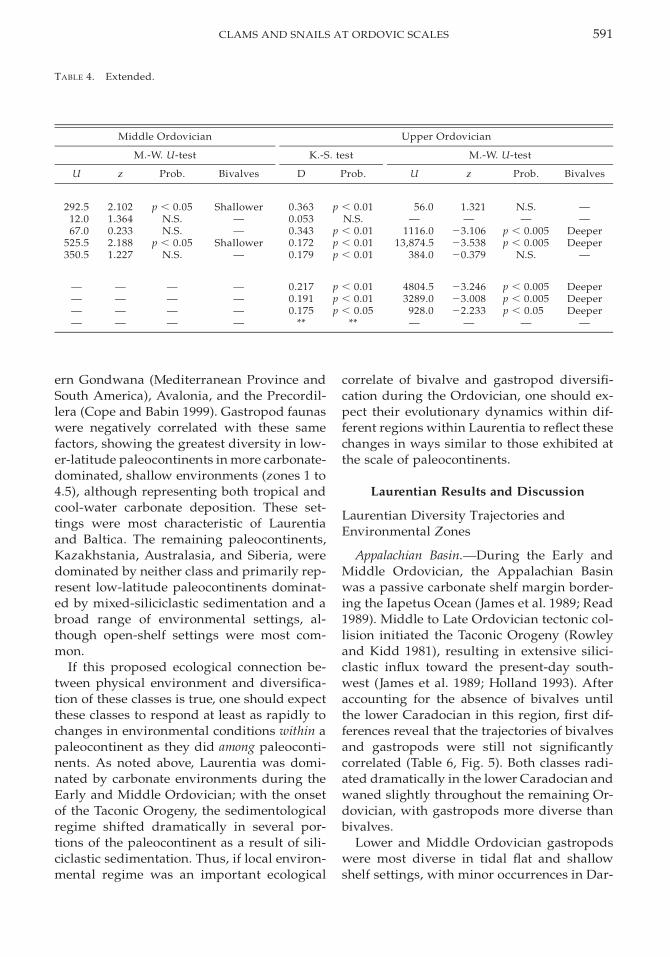
591CLAMS AND SNAILS AT ORDOVIC SCALES
TABLE 4. Extended.
Middle Ordovician
M.-W. U-test
U z Prob. Bivalves
Upper Ordovician
K.-S. test
D Prob.
M.-W. U-test
U z Prob. Bivalves
292.512.067.0
525.5350.5
2.1021.3640.2332.1881.227
p , 0.05N.S.N.S.
p , 0.05N.S.
Shallower——
Shallower—
0.3630.0530.3430.1720.179
p , 0.01N.S.
p , 0.01p , 0.01p , 0.01
56.0—
1116.013,874.5
384.0
1.321—
23.10623.53820.379
N.S.—
p , 0.005p , 0.005
N.S.
——
DeeperDeeper
—
————
————
————
————
0.2170.1910.175
**
p , 0.01p , 0.01p , 0.05
**
4804.53289.0
928.0—
23.24623.00822.233
—
p , 0.005p , 0.005p , 0.05
—
DeeperDeeperDeeper
—
ern Gondwana (Mediterranean Province andSouth America), Avalonia, and the Precordil-lera (Cope and Babin 1999). Gastropod faunaswere negatively correlated with these samefactors, showing the greatest diversity in low-er-latitude paleocontinents in more carbonate-dominated, shallow environments (zones 1 to4.5), although representing both tropical andcool-water carbonate deposition. These set-tings were most characteristic of Laurentiaand Baltica. The remaining paleocontinents,Kazakhstania, Australasia, and Siberia, weredominated by neither class and primarily rep-resent low-latitude paleocontinents dominat-ed by mixed-siliciclastic sedimentation and abroad range of environmental settings, al-though open-shelf settings were most com-mon.
If this proposed ecological connection be-tween physical environment and diversifica-tion of these classes is true, one should expectthese classes to respond at least as rapidly tochanges in environmental conditions within apaleocontinent as they did among paleoconti-nents. As noted above, Laurentia was domi-nated by carbonate environments during theEarly and Middle Ordovician; with the onsetof the Taconic Orogeny, the sedimentologicalregime shifted dramatically in several por-tions of the paleocontinent as a result of sili-ciclastic sedimentation. Thus, if local environ-mental regime was an important ecological
correlate of bivalve and gastropod diversifi-cation during the Ordovician, one should ex-pect their evolutionary dynamics within dif-ferent regions within Laurentia to reflect thesechanges in ways similar to those exhibited atthe scale of paleocontinents.
Laurentian Results and Discussion
Laurentian Diversity Trajectories andEnvironmental Zones
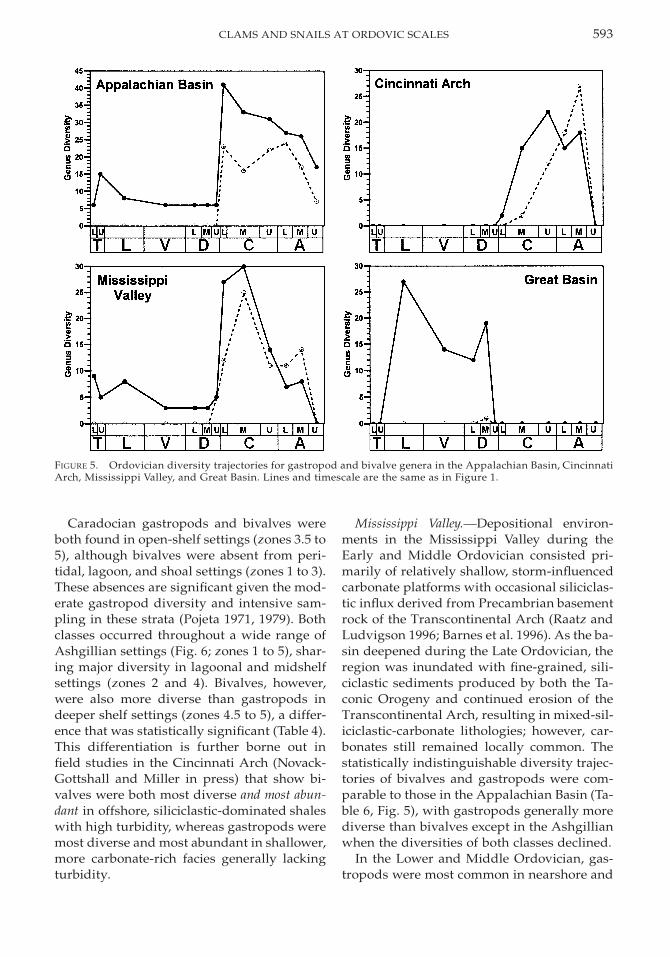
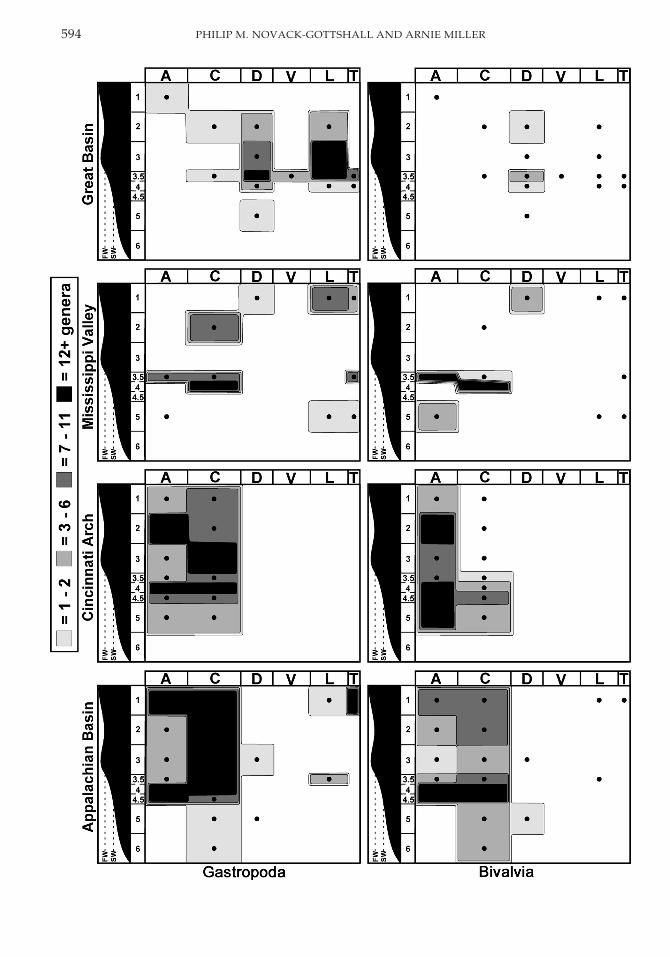
Appalachian Basin. During the Early andMiddle Ordovician, the Appalachian Basinwas a passive carbonate shelf margin border-ing the Iapetus Ocean (James et al. 1989; Read1989). Middle to Late Ordovician tectonic col-lision initiated the Taconic Orogeny (Rowleyand Kidd 1981), resulting in extensive silici-clastic influx toward the present-day south-west (James et al. 1989; Holland 1993). Afteraccounting for the absence of bivalves untilthe lower Caradocian in this region, first dif-ferences reveal that the trajectories of bivalvesand gastropods were still not significantlycorrelated (Table 6, Fig. 5). Both classes radi-ated dramatically in the lower Caradocian andwaned slightly throughout the remaining Or-dovician, with gastropods more diverse thanbivalves.
Lower and Middle Ordovician gastropodswere most diverse in tidal flat and shallowshelf settings, with minor occurrences in Dar-
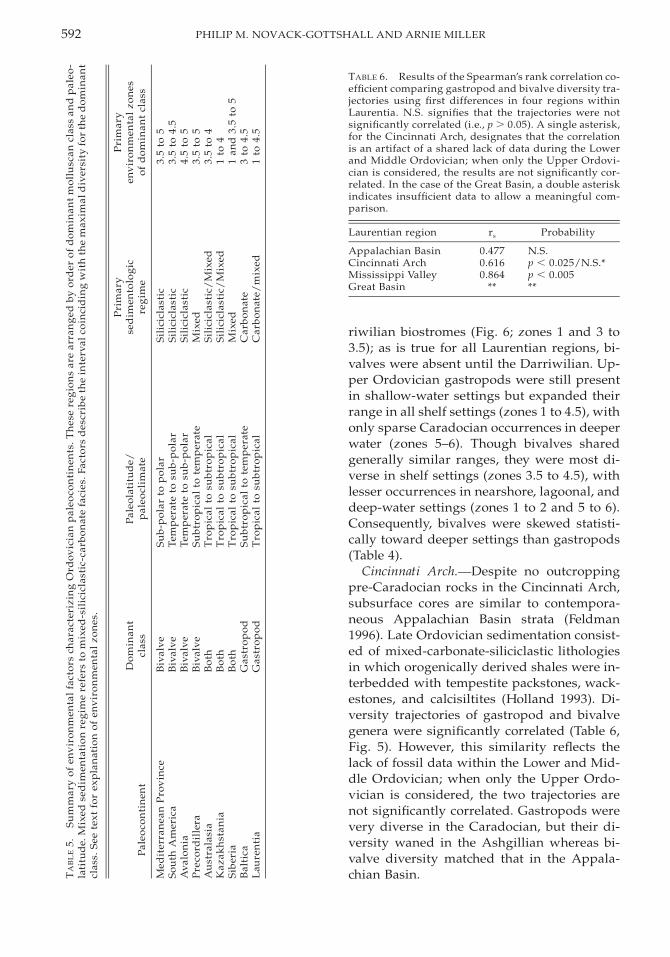
592 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
TA
BL
E5.
Sum
mar
yof
env
iron
men
tal
fact
ors
char
acte
rizi
ng
Ord
ovic
ian
pal
eoco
nti
nen
ts.T
hes
ere
gio
ns
are
arra
ng
edby
ord
erof
dom
inan
tm
ollu
scan
clas
san
dp
aleo
-la
titu
de.
Mix
edse
dim
enta
tion
reg
ime
refe
rsto
mix
ed-s
ilic
icla
stic
-car
bon
ate
faci
es.F
acto
rsd
escr
ibe
the
inte
rval
coin
cid
ing
wit
hth
em
axim
ald
iver
sity
for
the
dom
inan
tcl
ass.
See
text
for
exp
lan
atio
nof
env
iron
men
tal
zon
es.
Pal
eoco
nti
nen
tD
omin
ant
clas
sP
aleo
lati
tud
e/p
aleo
clim
ate
Pri
mar
yse
dim
ento
log
icre
gim
e
Pri
mar
yen
vir
onm
enta
lzo
nes
ofd
omin
ant
clas
s
Med
iter
ran
ean
Pro
vin
ceSo
uth
Am
eric
aA
valo
nia
Pre
cord
ille
raA
ust
rala
sia
Biv
alve
Biv
alve
Biv
alve
Biv
alve
Bot
h
Sub
-pol
arto
pol
arTe
mp
erat
eto
sub
-pol
arTe
mp
erat
eto
sub
-pol
arSu
btr
opic
alto
tem
per
ate
Tro
pic
alto
sub
trop
ical
Sili
cicl
asti
cSi
lici
clas
tic
Sili
cicl
asti
cM
ixed
Sili
cicl
asti
c/M
ixed
3.5
to5
3.5
to4.
54.
5to
53.
5to
53.
5to
4K
azak
hst
ania
Sib
eria
Bal
tica
Lau
ren
tia
Bot
hB
oth
Gas
trop
odG
astr
opod
Tro
pic
alto
sub
trop
ical
Tro
pic
alto
sub
trop
ical
Sub
trop
ical
tote
mp
erat
eT
rop
ical
tosu
btr
opic
al
Sili
cicl
asti
c/M
ixed
Mix
edC
arb
onat
eC
arb
onat
e/m
ixed
1to
41
and
3.5
to5
3to
4.5
1to
4.5
TABLE 6. Results of the Spearman’s rank correlation co-efficient comparing gastropod and bivalve diversity tra-jectories using first differences in four regions withinLaurentia. N.S. signifies that the trajectories were notsignificantly correlated (i.e., p . 0.05). A single asterisk,for the Cincinnati Arch, designates that the correlationis an artifact of a shared lack of data during the Lowerand Middle Ordovician; when only the Upper Ordovi-cian is considered, the results are not significantly cor-related. In the case of the Great Basin, a double asteriskindicates insufficient data to allow a meaningful com-parison.
Laurentian region rs Probability
Appalachian BasinCincinnati ArchMississippi ValleyGreat Basin
0.4770.6160.864
**
N.S.p , 0.025/N.S.*p , 0.005**
riwilian biostromes (Fig. 6; zones 1 and 3 to3.5); as is true for all Laurentian regions, bi-valves were absent until the Darriwilian. Up-per Ordovician gastropods were still presentin shallow-water settings but expanded theirrange in all shelf settings (zones 1 to 4.5), withonly sparse Caradocian occurrences in deeperwater (zones 5–6). Though bivalves sharedgenerally similar ranges, they were most di-verse in shelf settings (zones 3.5 to 4.5), withlesser occurrences in nearshore, lagoonal, anddeep-water settings (zones 1 to 2 and 5 to 6).Consequently, bivalves were skewed statisti-cally toward deeper settings than gastropods(Table 4).
Cincinnati Arch. Despite no outcroppingpre-Caradocian rocks in the Cincinnati Arch,subsurface cores are similar to contempora-neous Appalachian Basin strata (Feldman1996). Late Ordovician sedimentation consist-ed of mixed-carbonate-siliciclastic lithologiesin which orogenically derived shales were in-terbedded with tempestite packstones, wack-estones, and calcisiltites (Holland 1993). Di-versity trajectories of gastropod and bivalvegenera were significantly correlated (Table 6,Fig. 5). However, this similarity reflects thelack of fossil data within the Lower and Mid-dle Ordovician; when only the Upper Ordo-vician is considered, the two trajectories arenot significantly correlated. Gastropods werevery diverse in the Caradocian, but their di-versity waned in the Ashgillian whereas bi-valve diversity matched that in the Appala-chian Basin.
593CLAMS AND SNAILS AT ORDOVIC SCALES
FIGURE 5. Ordovician diversity trajectories for gastropod and bivalve genera in the Appalachian Basin, CincinnatiArch, Mississippi Valley, and Great Basin. Lines and timescale are the same as in Figure 1.
Caradocian gastropods and bivalves wereboth found in open-shelf settings (zones 3.5 to5), although bivalves were absent from peri-tidal, lagoon, and shoal settings (zones 1 to 3).These absences are significant given the mod-erate gastropod diversity and intensive sam-pling in these strata (Pojeta 1971, 1979). Bothclasses occurred throughout a wide range ofAshgillian settings (Fig. 6; zones 1 to 5), shar-ing major diversity in lagoonal and midshelfsettings (zones 2 and 4). Bivalves, however,were also more diverse than gastropods indeeper shelf settings (zones 4.5 to 5), a differ-ence that was statistically significant (Table 4).This differentiation is further borne out infield studies in the Cincinnati Arch (Novack-Gottshall and Miller in press) that show bi-valves were both most diverse and most abun-dant in offshore, siliciclastic-dominated shaleswith high turbidity, whereas gastropods weremost diverse and most abundant in shallower,more carbonate-rich facies generally lackingturbidity.
Mississippi Valley. Depositional environ-ments in the Mississippi Valley during theEarly and Middle Ordovician consisted pri-marily of relatively shallow, storm-influencedcarbonate platforms with occasional siliciclas-tic influx derived from Precambrian basementrock of the Transcontinental Arch (Raatz andLudvigson 1996; Barnes et al. 1996). As the ba-sin deepened during the Late Ordovician, theregion was inundated with fine-grained, sili-ciclastic sediments produced by both the Ta-conic Orogeny and continued erosion of theTranscontinental Arch, resulting in mixed-sil-iciclastic-carbonate lithologies; however, car-bonates still remained locally common. Thestatistically indistinguishable diversity trajec-tories of bivalves and gastropods were com-parable to those in the Appalachian Basin (Ta-ble 6, Fig. 5), with gastropods generally morediverse than bivalves except in the Ashgillianwhen the diversities of both classes declined.
In the Lower and Middle Ordovician, gas-tropods were most common in nearshore and
594 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER

595CLAMS AND SNAILS AT ORDOVIC SCALES
←
FIGURE 6. Contoured time-environment diagrams depicting the genus diversity of gastropods (above) and bivalves(below) in the Appalachian Basin, Cincinnati Arch, Mississippi Valley, and Great Basin during the Ordovician. Thetimescale is the same as in Figure 1. The key at top depicts the genus diversity of each class within shaded contours.
shallow-shelf settings (Fig. 6; zones 1 and 3.5)with sparse occurrences in deep-shelf settings(zone 5); as noted above, bivalves were absentuntil the Darriwilian when they occurred innearshore settings (zone 1) with gastropods.In the Upper Ordovician, both classes co-oc-curred in shallow and midshelf settings(zones 3.5 through 4); however, gastropodsalso occurred in lagoons (zone 2), whereas bi-valves were diverse in deep-shelf settings(zone 5). Given these differences, their envi-ronmental distributions were significantlydistinct in the Upper Ordovician, with bi-valves extending to deeper environments thangastropods (Table 4).
Great Basin. Located almost directly on theequator for much of the Ordovician, the GreatBasin was a vast carbonate platform with a va-riety of depositional environments (Ross et al.1989). Sufficient aeolian sands and silts en-tered the basin to produce mixed-carbonate-siliciclastic lithologies, and cratonic erosionduring the latest Middle Ordovician Sauk-Tippecanoe unconformity was sufficient tonearly drown the shelf with coarse-grainedsiliciclastics (Ross et al. 1989; Keller and Coo-per 1995). However, nearly pure carbonate-rich conditions resumed throughout the LateOrdovician (Droser and Sheehan 1995). Gas-tropods were quite diverse in this region inthe Lower and Middle Ordovician (Fig. 5),with Upper Ordovician gastropods either un-studied, poorly preserved, or with poor strat-igraphical resolution (Webb 1958; Droser andSheehan 1995). Because of the rarity of bi-valves from this region, statistical compari-sons of the diversity trajectories were not pos-sible (Table 4).
Gastropods were distributed throughout arange of environments in the Lower and Mid-dle Ordovician (Fig. 6; zones 2 to 5), attaininggreatest diversity in shoal and shallow-shelfsettings (zones 3 to 3.5). Upper Ordoviciangastropods were locally common in nearshoresettings (zone 1; Droser and Sheehan 1995);additional occurrences from a variety of deep-
er-water settings are also known but unpub-lished (Sheehan personal communication2003). Indeterminate bivalves have been notedonly rarely and no taxonomic descriptionshave been published from this region sinceWalcott’s classic study of the Eureka District(1884). Given the numerous occurrences ofgastropods, the rarity of bivalves in the GreatBasin is likely ecologically meaningful. Re-cently, several Middle Ordovician bivalveshave been discovered in the Great Basin (Mof-fat et al. 2001), but these are not sufficientlydiverse to change significantly the results pre-sented here. Regardless, statistical compari-sons of environmental distributions were notpossible (Table 6).
Discussion of Laurentian Patterns
The diversity trajectories of gastropods andbivalves within these four regions were simi-lar to those of Laurentia as a whole, with bi-valves absent until the latest Middle Ordovi-cian when both classes diversified dramati-cally. This diversification was not apparent inthe Great Basin, but the lack of well-preservedand recently studied Upper Ordovician mol-luscs limits further interpretation; the rarity ofpre-Caradocian bivalves in this region, how-ever, is not likely to change. Of the four re-gions studied, only the Mississippi Valley ex-hibits significant correlations between gastro-pod and bivalve diversity trajectories. Despitemoderately overlapping environmental distri-butions for both classes in all four regions,these classes had statistically different distri-butions; gastropods were most diverse inshallower-water settings than bivalves, whichwere most diverse in deep-shelf, fine-grained,siliciclastic settings.
Although gastropods were quite diverse ineach region, bivalves outnumbered them inthe Cincinnati Arch and the Mississippi Val-ley; both of these regions were dominated bystorm-influenced sedimentation and mixed-siliciclastic-carbonate lithologies. Notably, bi-valve diversification in all four regions coin-

596 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
cided with influx of fine-grained sedimentsfrom either orogenic activity or erosion of cra-tonic highlands. Gastropods exhibited similarbut reduced patterns in the Upper Ordovician,and they maintained diversity throughoutshallow carbonate strata throughout the Or-dovician. These environmental patterns par-allel observations made at the scales of paleo-continents and the world as a whole, in whichgastropods were generally most diverse inshallower water than bivalves. Furthermore,they support the earlier observation from thecomparisons among paleocontinents that thenature of the physical environment was inti-mately related to the diversification of theseclasses.
General Discussion and Conclusions
The Ordovician Radiation of gastropodsand bivalves was a complex but especially im-portant period in the history of these classes,responsible for dramatic increases in the num-ber of genera and the origins of many highertaxonomic groups. Although gastropods andbivalves displayed statistically indistinguish-able global diversity dynamics throughout theOrdovician, this study documents more dis-parate dynamics at the finer scales of five pa-leocontinents and four regions within Lauren-tia. Furthermore, there was significant hetero-geneity in diversity patterns among paleocon-tinents, with bivalves diversifying first in theGondwanan paleocontinents before occurringlater in Laurentia and Baltica (see also Babin1993, 1995 and Cope and Babin 1999), wheregastropods were already well established.Among these paleocontinents, only Avaloniaexhibited significantly correlated bivalve andgastropod diversity trajectories. These statis-tical differences were further observed at thefinest scale of regions within Laurentia.
This scale-dependence of diversity trajec-tories contrasts with the scale-independentenvironmental distributions of these classes.Regardless of whether Ordovician gastropodand bivalve genus occurrences are consideredin global aggregate or at the finer scales of pa-leocontinents or regions within Laurentia, asimilar environmental picture emerges in cas-es where there are sufficient data to permit ananalysis. Gastropods were most diverse in
shallow water, carbonate-rich platforms inlow latitudes; in contrast, bivalves were mostdiverse in deeper-water, higher-latitude set-tings in which fine-grained siliciclastics pre-dominated, whether resulting from tectonicactivity or a lack of carbonate production.
Although our study supports the conclu-sions of Babin (1993, 1995; Cope and Babin1999) regarding the paleocontinental diversi-fication of bivalves, our results concerning en-vironmental trends may appear at first glanceto be at odds with those Cope and Babin(1999) in two significant details. First, they ar-gued that Ordovician, and particularly LowerOrdovician, bivalves were generally restrictedto nearshore clastic settings. Although it istrue that bivalves did inhabit shallow-watersettings throughout the Ordovician, our re-sults, and those of several other studies (e.g.,Babin et al. 1982; Frey 1987; Miller 1988, 1989)clearly demonstrate that bivalves were neithermost diverse nor restricted to these settings atany point during the Ordovician. In fact, withthree low-diversity, nearshore exceptions(Harrington 1938; Nikitin et al. 1986; Shergoldet al. 1991), most Lower Ordovician bivalvesoccurred in deeper-water settings, includingthe diverse Latorpian fauna (Cope 1996) re-interpreted here as representing deep-water,fan-deltaic facies (zone 5). Reconciliation maybe found by differences in our definitions ofshallow water, as even our zone 5 shelf may beconsidered ‘‘relatively shallow’’ or ‘‘onshore’’compared with slope and basinal deposits(see discussion in Structure of the Databaseabove).
A second, more significant point of differ-ence is their conclusion that Late Ordovicianbivalve diversification in Laurentia wasspurred on by the presence of carbonate plat-forms. Although Laurentia was indeed the siteof extensive Late Ordovician bivalve diversi-fication (Fig. 3), this radiation was limited pri-marily to settings influenced by siliciclasticsedimentation in a significant part of the cen-tral and eastern United States and Canada(Fig. 5). It is possible that Cope and Babin(1999) implied a role for carbonates because ofthe tropical location of Laurentia during theOrdovician. However, direct assessment ofMiddle and Upper Ordovician strata makes
597CLAMS AND SNAILS AT ORDOVIC SCALES
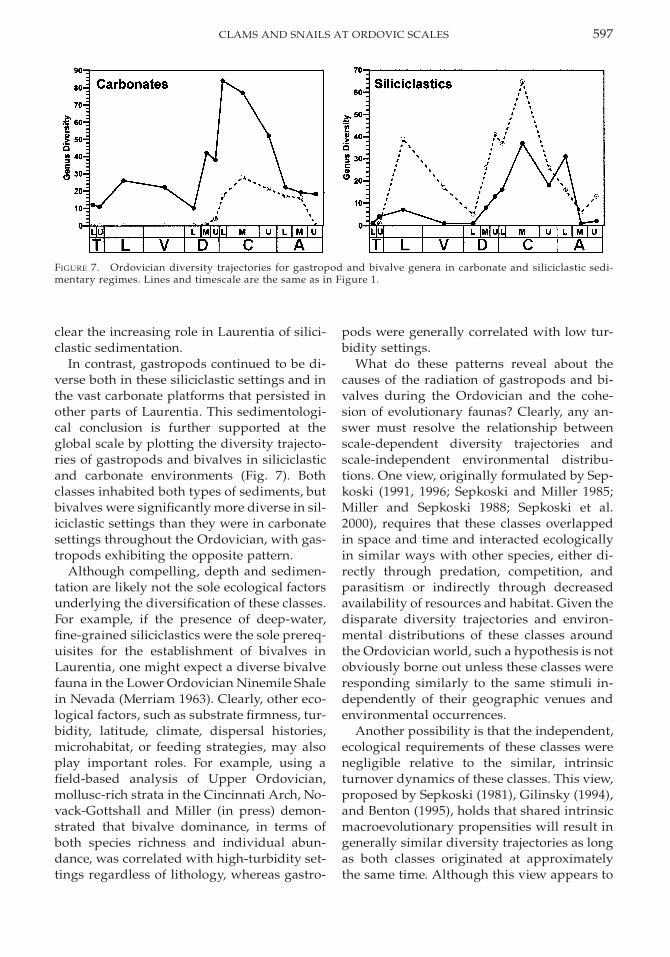
FIGURE 7. Ordovician diversity trajectories for gastropod and bivalve genera in carbonate and siliciclastic sedi-mentary regimes. Lines and timescale are the same as in Figure 1.
clear the increasing role in Laurentia of silici-clastic sedimentation.
In contrast, gastropods continued to be di-verse both in these siliciclastic settings and inthe vast carbonate platforms that persisted inother parts of Laurentia. This sedimentologi-cal conclusion is further supported at theglobal scale by plotting the diversity trajecto-ries of gastropods and bivalves in siliciclasticand carbonate environments (Fig. 7). Bothclasses inhabited both types of sediments, butbivalves were significantly more diverse in sil-iciclastic settings than they were in carbonatesettings throughout the Ordovician, with gas-tropods exhibiting the opposite pattern.
Although compelling, depth and sedimen-tation are likely not the sole ecological factorsunderlying the diversification of these classes.For example, if the presence of deep-water,fine-grained siliciclastics were the sole prereq-uisites for the establishment of bivalves inLaurentia, one might expect a diverse bivalvefauna in the Lower Ordovician Ninemile Shalein Nevada (Merriam 1963). Clearly, other eco-logical factors, such as substrate firmness, tur-bidity, latitude, climate, dispersal histories,microhabitat, or feeding strategies, may alsoplay important roles. For example, using afield-based analysis of Upper Ordovician,mollusc-rich strata in the Cincinnati Arch, No-vack-Gottshall and Miller (in press) demon-strated that bivalve dominance, in terms ofboth species richness and individual abun-dance, was correlated with high-turbidity set-tings regardless of lithology, whereas gastro-
pods were generally correlated with low tur-bidity settings.
What do these patterns reveal about thecauses of the radiation of gastropods and bi-valves during the Ordovician and the cohe-sion of evolutionary faunas? Clearly, any an-swer must resolve the relationship betweenscale-dependent diversity trajectories andscale-independent environmental distribu-tions. One view, originally formulated by Sep-koski (1991, 1996; Sepkoski and Miller 1985;Miller and Sepkoski 1988; Sepkoski et al.2000), requires that these classes overlappedin space and time and interacted ecologicallyin similar ways with other species, either di-rectly through predation, competition, andparasitism or indirectly through decreasedavailability of resources and habitat. Given thedisparate diversity trajectories and environ-mental distributions of these classes aroundthe Ordovician world, such a hypothesis is notobviously borne out unless these classes wereresponding similarly to the same stimuli in-dependently of their geographic venues andenvironmental occurrences.
Another possibility is that the independent,ecological requirements of these classes werenegligible relative to the similar, intrinsicturnover dynamics of these classes. This view,proposed by Sepkoski (1981), Gilinsky (1994),and Benton (1995), holds that shared intrinsicmacroevolutionary propensities will result ingenerally similar diversity trajectories as longas both classes originated at approximatelythe same time. Although this view appears to

598 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
be borne out at the global scale, it is insuffi-cient, by itself, to explain the rather differentdynamics of these classes in different geo-graphic and environmental settings. Further-more, such a macroevolutionary explanationrequires that the strong correlation betweenthe diversification of these classes and thephysical setting within these paleocontinentsis coincidental, which is unlikely given thegeneral observation that wherever shallowcarbonate settings flourished throughout theregions covered in this database, gastropodswere diverse. In addition, wherever there wassignificant influx of or sufficient sedimento-logical conditions for fine-grained siliciclasticsediments into deep subtidal settings, bi-valves were more likely to flourish. These re-lationships were best exemplified in Lauren-tia, where the Taconic Orogeny dramaticallyaltered the physical setting of the paleoconti-nent by introducing large amounts of silici-clastic sedimentation. Only after the advent ofwidespread siliciclastic sedimentation did bi-valves become established there.
Thus, the Ordovician Radiation of gastro-pods and bivalves appears to be best charac-terized by a dynamic interaction between theconsistent environmental propensities of eachclass in a world experiencing large-scale phys-ical transitions, including tectonic, climatolog-ical, and oceanographic changes (Miller1997b, 1998). However, given that these clas-ses responded differently to these environ-mental transitions, and that the compositeglobal diversity trajectories of these classeswere statistically indistinguishable, this con-clusion does not reject the claim that intrinsicmacroevolutionary properties of these classeswere also important contributors to theirglobal diversification (see Connolly and Miller2002). For example, intrinsic factors may haveplayed significant roles in determining theturnover rates, carrying capacities, or othermacroevolutionary factors underlying the dy-namics of their radiations. However, our re-sults suggest that the initial capacity of theseclasses to diversify during the Ordovician wasaffected greatly by their environmental pro-pensities in a physically dynamic Ordovicianworld.
Acknowledgments
We thank the following for discussion: C. E.Brett, C. Ciampaglio, S. M. Holland, D. W.McShea, D. L. Meyer, S. E. Novack-Gottshall,E. Venit, and P. J. Wagner. Comments by M. E.Patzkowsky and P. M. Sheehan greatly im-proved the manuscript. A bibliography of ref-erences and complete data used in compilingthis database are available from Novack-Gottshall. This study is based on a chapter ofthe M.S. thesis by Novack-Gottshall at theUniversity of Cincinnati. It was supported bygrants from the University of Cincinnati, thePaleontological Society, the Geological Societyof America, and Sigma Xi to Novack-Gottsh-all; and from the National Aeronautic andSpace Administration’s Program in Exobiolo-gy to Miller (Grants NAGW-3307, NAG5-6946,and NAG5-9418).
Literature Cited
Aitken, J. D., and B. S. Norford. 1967. Lower Ordovician SurveyPeak and Outran Formations, southern Rocky Mountains ofAlberta. Bulletin of Canadian Petroleum Geology 15:150–207.
Alroy, J. 2000a. New methods for quantifying macroevolution-ary patterns and processes. Paleobiology 26:707–733.
———. 2000b. Successive approximations of diversity curves:ten more years in the library. Geology 28:1023–1026.
Alroy, J., C. R. Marshall, R. K. Bambach, K. Bezusko, M. Foote,F. T. Fursich, T. A. Hansen, S. M. Holland, L. C. Ivany, D. Ja-blonski, D. K. Jacobs, D. C. Jones, M. A. Kosnik, S. Lidgard, S.Low, A. I. Miller, P. M. Novack-Gottshall, T. D. Olszewski, M.E. Patzkowsky, D. M. Raup, K. Roy, J. J. Sepkoski, Jr., M. G.Sommers, P. J. Wagner, and A. Webber. 2001. Effects of sam-pling standardization on estimates of Phanerozoic marine di-versification. Proceedings of the National Academy of Scienc-es USA 98:6261–6266.
Babin, C., R. Courtessole, M. Melou, J. Pillet, D. Vizcaino, and E.L. Yochelson. 1982. Brachiopodes (Articules) et mollusques(bivalves, rostroconches, monoplacophores, gasteropodes) del’Ordovicien inferieur (Tremadocien-Arenigien) de la Mon-tagne Noire (France meridionale). Memoires de la Societed’Etudes Scientifiques de l’Aude, Carcassonne.
Babin, C. 1993. Role des plates-formes gondwaniennes dans lesdiversifications des mollusques bivalves durant l’Ordovicien.Bulletin de la Societe Geologique de France 164:141–153.
———. 1995. The initial Ordovician bivalve mollusc radiationson the western Gondwanan shelves. Pp. 491–498 in Cooper etal. 1995.
Bambach, R. K. 1985. Classes and adaptive variety: the ecologyof diversification in marine faunas through the Phanerozoic.Pp. 191–253 in J. W. Valentine, ed. Phanerozoic diversity pat-terns: profiles in macroevolution. Princeton University Press,Princeton, N.J.
Barnes, D. A., W. B. Harrison III, and T. H. Shaw. 1996. Lower-Middle Ordovician lithofacies and interregional correlation,Michigan basin, U.S.A. Pp. 35–54 in Witzke et al. 1996.
Bass, N.W., and S.A. Northrop. 1953. Dotsero and Manitou For-mations, White River Plateau, Colorado, with special refer-ence to Clinetop Algal Limestone Member of Dotsero For-

599CLAMS AND SNAILS AT ORDOVIC SCALES
mation. Bulletin of the American Association of PetroleumGeologists 37:889–912.
Benton, M. J. 1995. Diversification and extinction in the historyof life. Science 268:52–58.
Berthelsen, A. 1998. The Tornquist Zone northwest of the Car-pathians: an intraplate pseudosuture. Geologiska Forenin-gens Forhandlingen 120:223–230.
Bevins, R. E., B. J. Bluck, P. J. Brenchley, R. A. Fortey, C. P.Hughes, J. K. Ingham, and A. W. A. Rushton. 1992. Ordovi-cian. In J. C. W. Cope, J. K. Ingham, and P. F. Rawson, eds.Atlas of palaeogeography and lithofacies. Geological Societyof London Memoir 13:19–36.
Bottjer, D. J., and D. Jablonski. 1988. Paleoenvironmental pat-terns in the evolution of post-Paleozoic benthic marine inver-tebrates. Palaios 3:540–560.
Boucot, A. J. 1975. Evolution and extinction rate controls. Elsev-ier, Amsterdam.
———. 1981. Principles of benthic marine paleoecology. Aca-demic Press, New York.
Brenchley, P. J. 1990. End Ordovician. Pp. 181–184 in D. E. G.Briggs and P. R. Crowther, eds. Palaeobiology: a synthesis.Blackwell Scientific, London.
Bretsky, P. W. 1968. Evolution of Paleozoic marine invertebratecommunities. Science 159:1231–1233.
———. 1969. Central Appalachian Late Ordovician communi-ties. Geological Society of America Bulletin 80:193–202.
———. 1970a. Upper Ordovician ecology of the central Appa-lachians. Peabody Museum of Natural History Bulletin 34.
———. 1970b. Late Ordovician benthic communities in north-central New York. New York State Museum and Science Ser-vice Bulletin 414.
Brett, C. E., A. J. Boucot, and B. Jones. 1993. Absolute depths ofSilurian benthic assemblages. Lethaia 26:25–40.
Bureau of Geology and Mineral Resources of Guangdong Prov-ince (BGMRGP). 1988. Regional Geology of Guangdong Prov-ince. Geological Publishing House, Beijing.
Cocks, L. R. M. 2001. Ordovician and Silurian global geography.Journal of the Geological Society, London 158:197–210.
Cocks, L. R. M., and R. A. Fortey. 1988. Lower Palaeozoic faciesand faunas around Gondwana. In M. G. Audley-Charles andA. Hallam, eds. Gondwana and Tethys. Geological SocietySpecial Publication 37:183–200.Oxford University Press, NewYork.
———. 1990. Biogeography of Ordovician and Silurian faunas.In W. S. McKerrow and C. R. Scotese, eds. Paleozoic palaeo-geography and biogeography. Geological Society of AmericaMemoir 12:97–104.
———. 1997. A new Hirnantian fauna from Thailand and the bio-geography of the latest Ordovician of south-east Asia. Geo-bios Memoir 20:117–126.
———. 1998. The Lower Palaeozoic margins of Baltica. Geolo-giska Foreningens Forhandlingen 120:173–179.
Coleman, J. M., and D. B. Prior. 1981. Deltaic environments ofdeposition. In P. A. Scholle and D. Spearing, eds. Sandstonedepositional environments. AAPG Memoir 31:139–178.American Association of Petroleum Geologists,Tulsa, Okla.
Connolly, S. R., and A. I. Miller. 2002. Global Ordovician faunaltransitions in the marine benthos: ultimate causes. Paleobi-ology 28:26–40.
Cooper, J. D. M. L. Droser, and S. C. Finney, eds. 1995. Ordo-vician odyssey: short papers for the seventh internationalsymposium on the Ordovician System. Society for Sedimen-tary Geology (SEPM), Fullerton, Calif.
Cope, J. C. W. 1996. Early Ordovician (Arenig) bivalves from theLlangyod Inlier, South Wales. Palaeontology 39:979–1025.
———. 1997. The early phylogeny of the class Bivalvia. Palaeon-tology 40:713–746.
———. 1999. Middle Ordovician bivalves from mid-Wales andthe Welsh Borderland. Palaeontology 42:467–499.
Cope, J. C. W. and C. Babin. 1999. Diversification of bivalves inthe Ordovician. Geobios 32:175–185.
Cope, J. C. W., J. K. Ingham, and P. F. Rawson, eds. 1992. Atlasof palaeogeography and lithofacies. Geological Society ofLondon Memoir 13.
Crevello, P. D., J. L. Wilson, J. F. Sarg, and J. F. Read. 1989. Con-trols on carbonate platform and basin development. SEPMSpecial Publication 44.
Droser, M. L., and P. M. Sheehan. 1995. Paleoecology of the Or-dovician Radiation and the Late Ordovician extinction event.Pp. 64–106 in J. Cooper, ed. Ordovician of the Great Basin:fieldtrip guidebook and volume for the seventh internationalsymposium on the Ordovician System. Society for Sedimen-tary Geology (SEPM), Fullerton, Calif.
Erwin, D. H. 1990. Carboniferous-Triassic gastropod diversitypatterns and the Permo-Triassic mass extinction. Paleobiolo-gy 16:187–203.
Feldmann, R. M., ed. 1996. Fossils of Ohio. Ohio Division ofGeological Survey Bulletin 70.
Foote, M. 1994. Temporal variation in extinction risk and tem-poral scaling of extinction metrics. Paleobiology 20:424–444.
Foote, M., and J. J. Sepkoski Jr. 1999. Absolute measures of thecompleteness of the fossil record. Nature 398:415–417.
Frey, R. C. 1987. The occurrence of Pelecypods in Early Paleo-zoic epeiric-sea environments, Late Ordovician of the Cincin-nati, Ohio area. Palaios 2:3–23.
Gilinsky, N. L. 1994. Volatility and the Phanerozoic decline ofbackground extinction intensity. Paleobiology 20:445–458.
Gould, S. J., and C. B. Calloway. 1980. Clams and brachiopods:ships that pass in the night. Paleobiology 6:383–396.
Guensberg, T. E., and J. S. Sprinkle. 1992. Rise of echinodermsin the Paleozoic evolutionary fauna: significance of paleoen-vironmental controls. Geology 20:407–410.
Harland, T. L. 1981. Middle Ordovician reefs of Norway. Lethaia14:169–188.
Harper, J. A., and H. B. Rollins. 2000. The Bellerophont contro-versy revisited. American Malacological Bulletin 15:147–156.
Harrington, H. J. 1938. Sobre las faunas del Ordovıcico inferiordel norte Argentino. Revista Museo de La Plata, new series 1:109–289.
Holland, S. M. 1993. Sequence stratigraphy of a carbonate-clas-tic ramp: the Cincinnatian Series (Upper Ordovician) in itstype area. Geological Society of America Bulletin 105:306–322.
Holland, S. M., and M. E. Patzkowsky. 1996. Sequence stratig-raphy and long-term lithologic change in the Middle and Up-per Ordovician of the eastern United States. Pp. 117–130 inWitzke et al. 1996.
Jaanusson, V. 1973. Aspects of carbonate sedimentation in theOrdovician of Baltoscandia. Lethaia 6:11–34.
Jablonski, D. 1998. Geographic variation in the molluscan re-covery from the End-Cretaceous extinction. Science 279:1327–1330.
Jablonski, D., and D. M. Raup. 1995. Selectivity of End-Creta-ceous marine bivalve extinctions. Science 268:389–391.
Jablonski, D., J. J. Sepkoski Jr., D. J. Bottjer, and P. M. Sheehan.1983. Onshore-offshore patterns in the evolution of Phaner-ozoic shelf communities. Science 222:1123–1125.
Jablonski, D., S. Lidgard, and P. D. Taylor. 1997. Comparativeecology of bryozoan radiations: origin of novelties in cyclo-stomes and cheilostomes. Palaios 12:505–523.
James, N. P., R. K. Stevens, C. R. Barnes, and I. Knight. 1989.Evolution of a Lower Paleozoic continental-margin carbonateplatform, northern Canadian Appalachians. Pp. 123–146 inCrevello et al. 1989.
Kammer, T. W., C. E. Brett, D. R. Boardman, and R. H. Mapes.

600 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
1986. Ecologic stability of the dysaerobic biofacies during thelate Paleozoic. Lethaia 19:109–121.
Keller, M., and J. Cooper. 1995. Paleokarst in the lower MiddleOrdovician of southeastern California and adjacent Nevadaand its bearing on the Sauk-Tippecanoe boundary problem.Pp. 323–327 in Cooper et al. 1995.
McKinney, F. K. 1990. Classifying and analyzing evolutionarytrends. Pp. 28–58 in K. J. MacNamara, ed. Evolutionarytrends. University of Arizona Press, Tucson.
Merriam, C. W. 1963. Paleozoic rocks of Antelope Valley, Eurekaand Nye Counties, Nevada: U.S. Geological Survey Profes-sional Paper 423.
Miller, A. I. 1988. Spatio-temporal transitions in Paleozoic Bi-valvia: an analysis of North American fossil assemblages.Historical Biology 1:251–273.
———. 1989. Spatio-temporal transitions in Paleozoic Bivalvia:a field comparison of Upper Ordovician and Upper Paleozoicbivalve-dominated fossil assemblages. Historical Biology 2:227–260.
———. 1997a. Comparative diversification dynamics among pa-leocontinents during the Ordovician Radiation. GeobiosMemoir 20:397–406.
———. 1997b. Dissecting global diversity patterns: examplesfrom the Ordovician Radiation. Annual Review of Ecologyand Systematics 28:85–104.
———. 1998. Biotic transitions in global marine diversity. Sci-ence 281:1157–1160.
Miller, A. I., and S. R. Connolly. 2001. Substrate affinities ofhigher taxa and the Ordovician Radiation. Paleobiology 27:768–778.
Miller, A. I., and M. Foote. 1996. Calibrating the Ordovician Ra-diation of marine life: implications for Phanerozoic diversitytrends. Paleobiology 22:304–309.
Miller, A. I., and S. Mao. 1995. Association of orogenic activitywith the Ordovician Radiation of marine life. Geology 23:305–308.
———. 1998. Scales of diversification and the Ordovician Ra-diation. Pp. 288–310 in M. L. McKinney, ed. Biodiversity dy-namics: turnover of populations, taxa, and communities. Co-lumbia University Press, New York.
Miller, A. I., and J. J. Sepkoski Jr. 1988. Modeling bivalve diver-sification: the effect of interaction on a macroevolutionarysystem. Paleobiology 14:364–369.
Moffat, H., M. L. Droser, and D. Thiel. 2001. Middle Ordovicianbivalves of the Basin and Range. Geological Society of Amer-ica Abstracts with Programs 33:A-73.
Nikitin, I. F., M. K. Apollonov, D. T. Tzaj, V. G. Koroljov, A. I.Kim, M. V. Erina, N. M. Larin, and A. N. Golicov. 1986. TheOrdovician System in Kazakhstan and Middle Asia: correla-tion chart and explanatory notes. International Union of Geo-logical Sciences, Publication 21.
Novack-Gottshall, P. M., and A. I. Miller. In press. Comparativetaxonomic richness and abundance of Late Ordovician gas-tropods and bivalves in mollusc-rich strata of the CincinnatiArch.. Palaios.
Palmqvist, P. 1991. Differences in the fossilization potential ofbivalve and gastropod species related to their life sites andtrophic resources. Lethaia 24:287–288.
Patzkowsky, M. E., and S. M. Holland. 1996. Extinction, inva-sion, and sequence stratigraphy: patterns of faunal change inthe Middle and Upper Ordovician of the eastern UnitedStates. Pp. 131–142 in Witzke et al. 1996.
———. 1999. Biofacies replacement in a sequence stratigraphicframework: Middle and Upper Ordovician of the NashvilleDome, Tennessee, USA. Palaios 14:301–323.
Percival, I. G. 1995. Ordovician gastropods of New South Wales:ecologic and biogeographic relationships. Pp. 453–456 in Coo-per et al. 1995.
Podhalanska, T. 1995. Early/Middle Ordovician condensationin the Polish part of the Baltic basin. Pp. 381–284 in Cooper etal. 1995.
Pojeta, J., Jr. 1971. Review of Ordovician Pelecypods. U.S. Geo-logical Survey Professional Paper 695.
———. 1979. The Ordovician paleontology of Kentucky andnearby states: introduction. U.S. Geological Survey Profes-sional Paper 1066-A.
Pojeta, J., Jr., and J. Gilbert-Tomlinson. 1977. Australian Ordo-vician pelecypod molluscs. Bureau of Mineral Resources, Ge-ology and Geophysics, Australia, Bulletin 174.
Poulsen, C. 1937. On the Lower Ordovician faunas of eastGreenland. Meddelelser om Grønland 119(3).
Raatz, W. D., and G. A. Ludvigson. 1996. Depositional environ-ments and sequence stratigraphy of Upper Ordovician epi-continental deep water deposits, eastern Iowa and southernMinnesota. Pp. 143–160 in Witzke et al. 1996.
Raup, D. M. 1979. Size of the Permo-Triassic bottleneck and itsevolutionary implications. Science 206:217–218.
Raup, D. M., and G. E. Boyajian. 1988. Patterns of generic ex-tinction in the fossil record. Paleobiology 14:109–125.
Raup, D. M., and D. Jablonski. 1993. Geography of End-Creta-ceous marine bivalve extinctions. Science 260:971–973.
Read, J. F. 1989. Controls on evolution of Cambrian-Ordovicianpassive margin, U.S. Appalachians. Pp. 147–166 in Crevello etal. 1989.
Rigby, J. K., M. H. Nitecki, Z. Zhongde, L. Bingli, and J. Yang-wen. 1995. Lower Ordovician reefs of Hubei, China, and thewestern United States. Pp. 423–426 in Cooper et al. 1995.
Ross, C. A., and J. R. P. Ross. 1995. North American Ordoviciandepositional sequences and correlations. Pp. 309–313 in Coo-per et al. 1995.
Ross, R. J., F. J. Adler, T. W. Amsden, D. Bergstrom, S. M. Berg-strom, C. Carter, M. Churkin, E. A. Cressman, J. R. Derby, J.T. Dutro, R. L. Ethington, S. C. Finney, D. W. Fisher, J. H. Fish-er, A. G. Harris, L. F. Hintze, K. B. Ketner, D. L. Kolata, E.Landing, R. B. Neuman, W. C. Sweet, J. Pojeta Jr., A. W. Potter,E. K. Rader, J. E. Repetski, R. H. Shaver, T. L. Thompson, andG. F. Webers. 1982. The Ordovician System in the UnitedStates of America: correlation chart and explanatory notes. In-ternational Union of Geological Sciences Publication 12.
Ross, R. J. R., Jr., N. P. James, L. F. Hintze, and F. G. Poole. 1989.Architecture and evolution of a Whiterockian (early MiddleOrdovician) carbonate platform, Basin Ranges of westernU.S.A. Pp. 167–186 in Crevello et al. 1989.
Rowley, D. B., and W. S. F. Kidd. 1981. Stratigraphic relation-ships and detrital composition of the Medial Ordovicianflysch of western New England: implications for the tectonicevolution of the Taconic Orogeny. Journal of Geology 89:199–218.
Schneider, J. A. 2001. Bivalve systematics during the 20th cen-tury. Journal of Paleontology 75:1119–1127.
Scotese, C. R., and W. S. McKerrow. 1991. Ordovician plate tec-tonic reconstructions. In C. R. Barnes and S. H. Williams, eds.Advances in Ordovician geology. Geological Survey of Can-ada Paper 90-9:271–282.
Sepkoski, J. J., Jr. 1981. A factor analytic description of the ma-rine fossil record. Paleobiology 7:36–53.
———. 1984. A kinetic model of Phanerozoic taxonomic diver-sity. III. Post-Paleozoic marine families and mass extinctions.Paleobiology 10:246–257.
———. 1988. Alpha, beta, or gamma: where does all the diver-sity go? Paleobiology 14:221–234.
———. 1991. A model of onshore-offshore change in faunal di-versity. Paleobiology 17:58–77.
———. 1995. The Ordovician radiations: diversification and ex-tinction shown by global genus level taxonomic data. Pp. 393–396 in Cooper et al. 1995.

601CLAMS AND SNAILS AT ORDOVIC SCALES
———. 1996. Competition in macroevolution: the double wedgerevisited. Pp. 211–255 in D. Jablonski, D. H. Erwin, and J. H.Lipps, eds. Evolutionary paleobiology. University of ChicagoPress, Chicago.
Sepkoski, J. J., Jr., and A. I. Miller. 1985. Evolutionary faunas andthe distribution of Paleozoic benthic communities in spaceand time. Pp, 153–190 in J. W. Valentine, ed. Phanerozoic di-versity patterns: profiles in macroevolution. Princeton Uni-versity Press, Princeton, N.J.
Sepkoski, J. J., Jr., and P. M. Sheehan. 1983. Diversification, fau-nal change, and community replacement during the Ordovi-cian radiations. Pp. 673–717 in M. J. S. Tevesz and P. L. McCall,eds. Biotic interactions in recent and fossil benthic commu-nities. Plenum, New York.
Sepkoski, J. J., Jr., F. K. McKinney, and S. Lidgard. 2000. Com-petitive displacement among post-Paleozoic cyclostome andcheilostome bryozoans. Paleobiology 26:7–18.
Sheehan, P. M. 1979. Swedish Late Ordovician marine benthicassemblages and their bearing on brachiopod zoogeography.Pp. 61–73 in J. Gray and A. J. Boucot, eds. Historical bioge-ography, plate tectonics, and the changing environment. Pro-ceedings of the 37th Annual Biology Colloquium, and SelectedPapers. Oregon State University Press, Corvallis, Oregon.
Shergold, J. H., J. D. Gorter, R. S. Nicoll, and P. W. Haines. 1991.Stratigraphy and the Pacoota Sandstone (Cambrian-Ordovi-cian) Amadeus Basin, N.T. Bureau of Mineral Resources, Aus-tralia, Bulletin 237.
Sokal, R. R., and F. J. Rohlf. 1995. Biometry: the principles andpractice of statistics in biological research, 3d ed. W. H. Free-man, San Francisco.
Speyer, S. E., and C. E. Brett. 1988. Taphofacies models for epeir-ic sea environments: middle Paleozoic examples. Palaeogeog-raphy, Palaeoclimatology, Palaeoecology 63:225–262.
Stanley, S. M. 1985. Rates of evolution. Paleobiology 11:13–26.Steele-Petrovic, H. M. 1979. The physiological differences be-
tween articulate brachiopods and filter-feeding bivalves as afactor in the evolution of marine level-bottom communities.Palaeontology 22:101–134.
Tjernvik, T. E. 1956. On the Early Ordovician of Sweden: stra-tigraphy and fauna. Bulletin of the Geological Institutions ofthe University of Uppsala 36:109–284.
———. 1958. The Tremadocian Beds at Flagabro in south-east-ern Scania (Sweden). Geologiska Foreningens i StockholmForhandlingar 80:259–276.
Torsvik, T. H. 1998. Palaeozoic palaeogeography: a North At-lantic viewpoint. Geologiska Foreningens Forhandlingen120:109–118.
Torsvik, T. H., M. A. Smethurst, J. G. Meert, R. Van der Voo, W.S. McKerrow, M. D. Brasier, B. A. Sturt, and H. J. Walderhung.1996. Continental break-up and collision in the Neoprotero-zoic and Palaeozoic: a tale of Baltica and Laurentia. Earth Sci-ence Reviews 40:229–258.
Traynor, J.-J. 1988. The Arenig in South Wales: sedimentary andvolcanic processes during the initiation of a marginal basin.Geological Journal 23:275–292.
Valentine, J. W. 1989. How good was the fossil record? Cluesfrom the Californian Pleistocene. Paleobiology 15:83–94.
Valentine, J. W., B. H. Tiffney, and J. J. Sepkoski Jr. 1991. Evolu-
tionary dynamics of plants and animals: a comparative ap-proach. Palaios 6:81–88.
Wagner, P. J. 1995a. Diversity patterns among early gastropods:contrasting taxonomic and phylogenetic descriptions. Paleo-biology 21:410–439.
———. 1995b. Testing evolutionary constraint hypotheses withearly Paleozoic gastropods. Paleobiology 21:248–272.
———. 1996. Patterns of morphologic diversification during theinitial radiation of the ‘‘Archaeogastropoda.’’ Pp. 161–169 inJ. Taylor, ed. Origin and evolutionary radiation of the Mollus-ca. Oxford University Press, Oxford.
———. 1999. Phylogenetic relationships of the earliest anisos-trophically coiled gastropods. Smithsonian Contributions toPaleobiology No. 88.
———. 2001. Gastropod phylogenetics: progress, problems, andimplications. Journal of Paleontology 75:1128–1140.
Wahlman, G. P. 1992. Middle and Upper Ordovician symmet-rical univalved molluscs (Monoplacophora and Bellerophon-tina) of the Cincinnati Arch region. United States GeologicalSurvey Professional Paper 1066-O.
Walcott, C. D. 1884. Paleontology of the Eureka District, Neva-da. United States Geological Survey Monograph 8.
Wang, X., and X. Chen. 1999. Palaeobiogeography and palaeo-climatology of Ordovician in China. Diceng Gushengwu Lun-wen Ji 27:1–27.
Wang, X., L. Xian, S. Ni, Q. Zheng, G. Xu, T. Zhou, C. Lai, andZ. Li. 1987. Biostratigraphy of the Yangtze Gorge Area, Part2. Early Paleozoic Era. Geological Publishing House, Beijing.
Webb, G. W. 1958. Middle Ordovician stratigraphy in easternNevada and western Utah. Bulletin of the American Associ-ation of Petroleum Geologists 42:2335–2377.
Webby, B. D. 1998. Steps toward a global standard for Ordovi-cian stratigraphy. Newsletters in Stratigraphy 36:1–33.
Westrop, S. R., and J. M. Adrain. 1998. Trilobite alpha diversityand the reorganization of Ordovician benthic marine com-munities. Paleobiology 24:1–16.
Westrop, S. R., J. V. Tremblay, and E. Landing. 1995. Decliningimportance of trilobites in Ordovician nearshore paleocom-munities: dilution or displacement? Palaios 10:75–79.
Williams, S. H., D. A. T. Harper, R. B. Neuman, W. D. Boyce, andC. MacNiocaill. 1995. Lower Paleozoic fossils from New-foundland and their importance in understanding the historyof the Iapetus Ocean. In J. P. Hibbard, C. R. van Staal, and P.A. Cawood, eds. Current perspectives in the Appalachian-Ca-ledonian Orogen. Geological Association of Canada SpecialPaper 41:115–126. Geological Association of Canada,St.John’s, Newfoundland.
Witzke, B. J., G. A. Ludvigson, and J. E. Day, eds. 1996. Paleozoicsequence stratigraphy: views from the North American Cra-ton. Geological Society of America Special Paper 306.
Yochelson, E. L. 1962. Early Ordovician gastropods from theOslo region, Norway. Norsk Geologisk Tidsskrift 42:239–250.
Yochelson, E. L., and A. O. Wise Jr. 1972. A life association ofshell and operculum in the Early Ordovician gastropod Cer-atopea unguis. Journal of Paleontology 46:681–684.
Young, T. P. 1985. The stratigraphy of the Upper Orovician ofcentral Portugal. Ph.D. thesis. University of Sheffield, Shef-field, England.
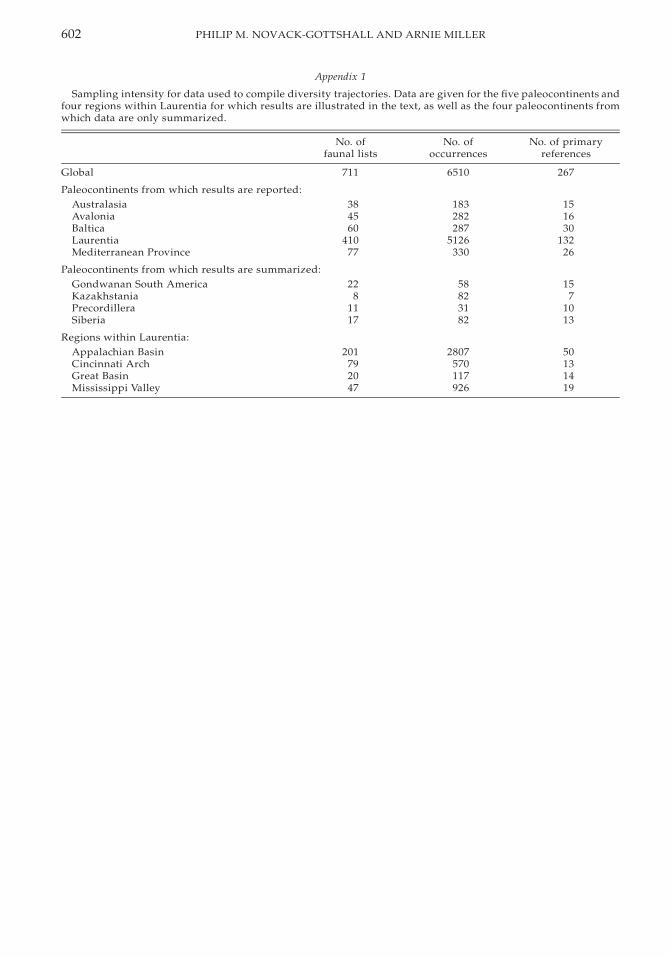
602 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
Appendix 1
Sampling intensity for data used to compile diversity trajectories. Data are given for the five paleocontinents andfour regions within Laurentia for which results are illustrated in the text, as well as the four paleocontinents fromwhich data are only summarized.
No. offaunal lists
No. ofoccurrences
No. of primaryreferences
Global 711 6510 267
Paleocontinents from which results are reported:AustralasiaAvaloniaBalticaLaurentiaMediterranean Province
384560
41077
183282287
5126330
151630
13226
Paleocontinents from which results are summarized:Gondwanan South AmericaKazakhstaniaPrecordilleraSiberia
228
1117
58823182
157
1013
Regions within Laurentia:Appalachian BasinCincinnati ArchGreat BasinMississippi Valley
201792047
2807570117926
50131419
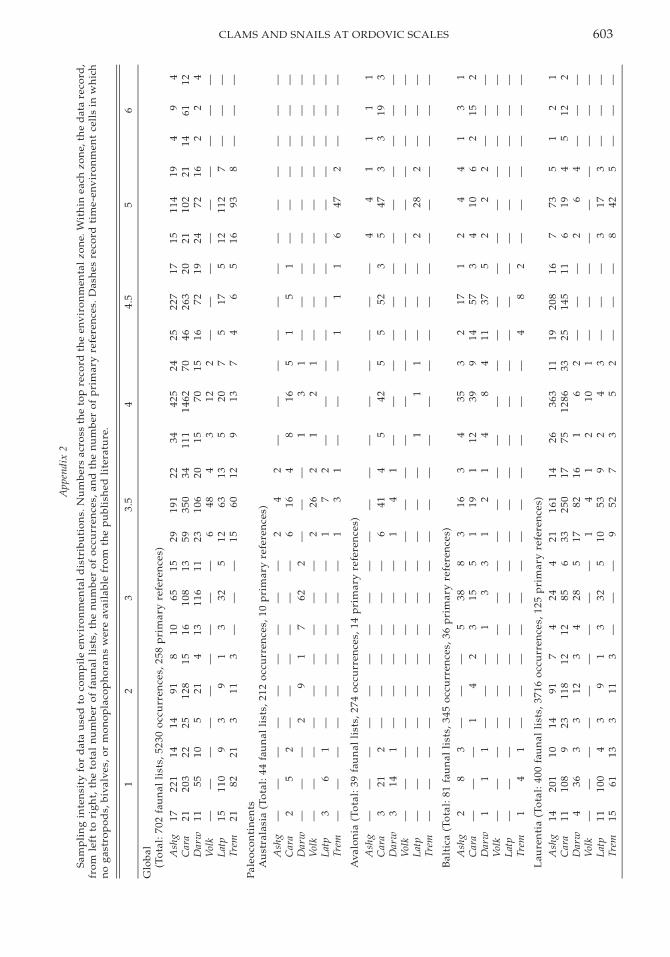
603CLAMS AND SNAILS AT ORDOVIC SCALES
App
endi
x2
Sam
pli
ng
inte
nsi
tyfo
rd
ata
use
dto
com
pil
een
vir
onm
enta
ld
istr
ibu
tion
s.N
um
ber
sac
ross
the
top
reco
rdth
een
vir
onm
enta
lzo
ne.
Wit
hin
each
zon
e,th
ed
ata
reco
rd,
from
left
tori
ght,
the
tota
ln
um
ber
offa
un
alli
sts,
the
nu
mb
erof
occu
rren
ces,
and
the
nu
mb
erof
pri
mar
yre
fere
nce
s.D
ash
esre
cord
tim
e-en
vir
onm
ent
cell
sin
wh
ich
no
gas
trop
ods,
biva
lves
,or
mon
opla
cop
hor
ans
wer
eav
aila
ble
from
the
pu
blis
hed
lite
ratu
re.
12
33.
54
4.5
56
Glo
bal
(Tot
al:7
02fa
un
alli
sts,
5230
occu
rren
ces,
258
pri
mar
yre
fere
nce
s)A
shg
Car
aD
arw
Vol
kL
atp
Trem
17 21 11 — 15 21
221
203 55 — 110 82
14 22 10 — 9 21
14 25 5 — 3 3
91 128 21 —9 11
8 15 4 — 1 3
10 16 13 — 3 —
65 108
116
— 32 —
15 13 11 — 5 —
29 59 23 6 12 15
191
350
106 48 63 60
22 34 20 4 13 12
34 111 15 3 5 9
425
1462 70 12 20 13
24 70 15 2 7 7
25 46 16 — 5 4
227
263 72 — 17 6
17 20 19 — 5 5
15 21 24 — 12 16
114
102 72 — 112 93
19 21 16 — 7 8
4 14 2 — — —
9 61 2 — — —
4 12 4 — — —
Pal
eoco
nti
nen
tsA
ust
rala
sia
(Tot
al:4
4fa
un
alli
sts,
212
occu
rren
ces,
10p
rim
ary
refe
ren
ces)
Ash
gC
ara
Dar
wV
olk
Lat
pTr
em
— 2 — — 3 —
—5
— —6
—
— 2 — — 1 —
— — 2 — — —
— —9
— — —
— — 1 — — —
— — 7 — — —
— — 62 — — —
— — 2 — — —
2 6 — 2 1 1
4 16 — 26 7 3
2 4 — 2 2 1
—8 1 1
— —
— 16 3 2— —
— 5 1 1 — —
— 1 — — — 1
—5
— — —1
— 1 — — — 1
— — — — — 6
— — — — — 47
— — — — — 2
— — — — — —
— — — — — —
— — — — — —
Ava
lon
ia(T
otal
:39
fau
nal
list
s,27
4oc
curr
ence
s,14
pri
mar
yre
fere
nce
s)A
shg
Car
aD
arw
Vol
kL
atp
Trem
— 3 3 — — —
— 21 14 — — —
— 2 1 — — —
— — — — — —
— — — — — —
— — — — — —
— — — — — —
— — — — — —
— — — — — —
— 6 1 — — —
— 41 4— — —
— 4 1 — — —
—5
— —1
—
— 42 — —1
—
— 5 — — 1 —
— 5 — — — —
— 52 — — — —
— 3 — — — —
4 5 — — 2 —
4 47 — — 28 —
1 3 — — 2 —
1 3 — — — —
1 19 — — — —
1 3 — — — —
Bal
tica
(Tot
al:8
1fa
un
alli
sts,
345
occu
rren
ces,
36p
rim
ary
refe
ren
ces)
Ash
gC
ara
Dar
wV
olk
Lat
pTr
em
2 — 1 — — 1
8—
1— —
4
3 — 1 — — 1
— 1 — — — —
—4
— — — —
— 2 — — — —
5 3 1 — — —
38 15 3— — —
8 5 3 — — —
3 1 1 — — —
16 19 2— — —
3 1 1 — — —
4 12 4— — —
35 39 8— — —
3 9 4 — — —
2 14 11 — — 4
17 57 37 — —8
1 3 5 — — 2
2 4 2 — — —
4 10 2— — —
4 6 2 — — —
1 2 — — — —
3 15 — — — —
1 2 — — — —
Lau
ren
tia
(Tot
al:4
00fa
un
alli
sts,
3716
occu
rren
ces,
125
pri
mar
yre
fere
nce
s)A
shg
Car
aD
arw
Vol
kL
atp
Trem
14 11 4 — 11 15
201
108 36 — 100 61
10 9 3 — 4 13
14 23 3 — 3 3
91 118 12 —9 11
7 12 3 — 1 3
4 12 4 — 3 —
24 85 28 — 32 —
4 6 5 — 5 —
21 33 17 1 10 9
161
250 82 4 53 52
14 17 16 1 9 7
26 75 1 2 2 3
363
1286 6 10 4 5
11 33 2 1 3 2
19 25 — — — —
208
145
— — — —
16 11 — — — —
7 6 2 — 3 8
73 19 6— 17 42
5 4 4 — 3 5
1 5 — — — —
2 12 — — — —
1 2 — — — —

604 PHILIP M. NOVACK-GOTTSHALL AND ARNIE MILLER
App
endi
x2.
Con
tin
ued
.
12
33.
54
4.5
56
Med
iter
ran
ean
Pro
vin
ce(T
otal
:69
fau
nal
list
s,33
2oc
curr
ence
s,30
pri
mar
yre
fere
nce
s)A
shg
Car
aD
arw
Vol
kL
atp
Trem
— 1 — — — 1
—5
— — —2
— 2 — — — 2
— — — — — —
— — — — — —
— — — — — —
1 1 — — — —
3 1— — — —
3 1 — — — —
— 10 — 3 1 1
— 16 — 18 3 1
— 4 — 1 1 1
1 1 3—
4 1
13 9 21 —8 1
5 3 2 — 1 1
— 1 4 — — —
—3 34 — — —
— 1 11 — — —
2 5 18 — 7 1
29 24 59 — 67 3
7 9 7 — 3 1
— 2 — — — —
— 12 — — — —
— 2 — — — —
Lau
ren
tian
Reg
ion
sA
pp
alac
hia
nB
asin
(Tot
al:1
61fa
un
alli
sts,
1251
occu
rren
ces,
44p
rim
ary
refe
ren
ces)
Ash
gC
ara
Dar
wV
olk
Lat
pTr
em
6 8 — — 3 8
138 79 — —5 43
3 5 — — 4 5
1 20 — — — —
16 85 — — — —
1 10 — — — —
1 7 1 — — —
8 57 1— — —
1 7 1 — — —
2 15 — — 1 —
12 89 — —4
—
2 10 — — 2 —
12 40 — — — —
252
276
— — — —
5 19 — — — —
8 16 — — — —
88 73 — — — —
8 4 — — — —
— 5 1 — — —
— 10 1— — —
— 3 1 — — —
1 5 — — — —
2 12 — — — —
1 3 — — — —
Cin
cin
nat
iA
rch
(Tot
al:8
0fa
un
alli
sts,
598
occu
rren
ces,
14p
rim
ary
refe
ren
ces)
Ash
gC
ara
Dar
wV
olk
Lat
pTr
em
2 1 — — — —
13 12 — — — —
2 2 — — — —
11 1 — — — —
72 10 — — — —
5 1 — — — —
2 5 — — — —
14 28 — — — —
2 1 — — — —
9 3 — — — —
31 34 — — — —
5 1 — — — —
13 7— — — —
104 44 — — — —
6 6 — — — —
13 7 — — — —
120 50 — — — —
8 4 — — — —
5 1 — — — —
57 9— — — —
4 1 — — — —
— — — — — —
— — — — — —
— — — — — —
Mis
siss
ipp
iV
alle
y(T
otal
:41
fau
nal
list
s,93
0oc
curr
ence
s,16
pri
mar
yre
fere
nce
s)A
shg
Car
aD
arw
Vol
kL
atp
Trem
— — 2 — 6 4
— — 20 — 70 12
— — 1 — 1 3
— 1 — — — —
— 22 — — — —
— 1 — — — —
— — — — — —
— — — — — —
— — — — — —
2 4 — — — 2
70 38 — — — 15
4 2 — — — 1
— 16 — — — —
— 664
— — — —
— 6 — — — —
— — — — — —
— — — — — —
— — — — — —
2 — — — 1 1
16 — — —2 1
1 — — — 1 1
— — — — — —
— — — — — —
— — — — — —
Gre
atB
asin
(Tot
al:3
5fa
un
alli
sts,
180
occu
rren
ces,
17p
rim
ary
refe
ren
ces)
Ash
gC
ara
Dar
wV
olk
Lat
pTr
em
4 — — — — —
4— — — — —
3 — — — — —
— 1 1 — 3 —
—1 5
—9
—
— 1 1 — 1 —
— — 2 — 1 —
— — 26 — 19 —
— — 3 — 3 —
— 1 9 1 4 2
—2 58 4 22 13
— 1 8 1 6 2
— —1
—2 2
— —6
—4 2
— — 2 — 3 1
— — — — — —
— — — — — —
— — — — — —
— — 1 — — —
— —5
— — —
— — 3 — — —
— — — — — —
— — — — — —
— — — — — —





























