Embed Size (px)
Citation preview

Conservation genetics of Cicindela deserticoloides, an endangeredtiger beetle endemic to southeastern Spain
A.C. Diogo,1 A.P. Vogler,1,2* A. Gimenez,3 D. Gallego3 and J. Galian4
1 The Natural History Museum, Department of Entomology, Cromwell Road, London, SW7 5BD, UK2 Department of Biology, Imperial College at Silwood Park, Ascot, Berkshire SL5 7PY, UK3 Department of Ecology and Hydrology, and 4 Department of Animal Biology,University of Murcia, 30017 Murcia, Spain
Received 12 August 1998; accepted 6 November 1998
Population surveys of the tiger beetle, Cicindela (Cephalota) deserticoloides, endemic to the few remaining salt steppes ofsoutheastern Spain revealed only four extant colonies. DNA sequencing of some 1896 base pairs of mitochondrial DNA for onespecimen each from three populations revealed only a single base pair change confined to a single of the three specimens, thusindicating an extremely low level of differentiation when compared to similar populations of Cicindela (s.l.) elsewhere. Diver-gence of C. deserticoloides from the closest relatives in the Iberian Peninsula was between 6.9 and 9.9%, attesting to theuniqueness of the species and its high conservation status. Habitat requirements appear to be phylogenetically conserved withinCephalota, but C. deserticoloides seems to be more narrowly confined to relatively drier conditions than its less endangeredrelatives. The geographic range of the relatives is wider and their local abundance higher, indicating that habitat specialization,low abundance and small geographic range in C. deserticoloides are correlated and in sum are responsible for its vulnerability toextinction.
Keywords: mitochondrial DNA, insect conservation, Cicindelidae, Iberian biodiversity, rarity, salt flats
J o u r n a l o f I n s e c t C o n s e r v a t i o n , 3 , 1 1 7 – 1 2 3 ( 1 9 9 9 )
Introduction
Many species of tiger beetles (genus Cicindela s.l.) areconfined to saline soils where they inhabit isolatedpatches of habitat, frequently near temporal or perma-nent sources of open water. There is increasing pressureto utilize these areas for agriculture, leading to altera-tion of the water table and the chemical compositionof the habitat. Because of the narrow habitat require-ments of most tiger beetles (Pearson, 1988) these changesinevitably lead to the disappearance of specialized spe-cies. Several cases of local population extinctions havebeen documented and were shown to be directly corre-lated to anthropogenic habitat disturbance (Stamatov,1972; Schultz, 1989; Knisley and Hill, 1992). In arid south-eastern Spain several species in the genus Cicindela s.l.are confined to saline habitats in coastal and inlandareas. One of these, C. (Cephalota) deserticoloides Codina(Codina, 1931), is a narrow endemic to the regionaround Murcia and Southern Alicante where it hasbeen recorded only from very few locations in dry saltsteppe habitat. Unfortunately, the unique salt steppe ofsouthern Spain has been heavily disturbed, strongly
* To whom correspondence should be addressed at: Department of Entomolo
1366–638X © 1999 Kluwer Academic Publishers
affecting C. deserticoloides. Localities for the specieshave a long history of being altered by drainage anddesalination for transformation into agricultural land(Palao, 1909) and are now being used for industrialdevelopment or rubbish dumps, often accompanied bythe construction of illegal buildings. The remainingpopulations are confined to isolated patches of suitedhabitat and have little or no contact, a situation that isaggravated by the increasing development and frag-mentation of natural areas. These factors impact on C.deserticoloides throughout its entire geographic rangeand thus put the species in danger of extinction.
For an effective conservation of C. deserticoloides,more information is needed to establish the current dis-tribution and the level of threat. We also requireimproved taxonomic information on C. deserticoloides,in particular with respect to two issues. First, C. deserti-coloides occurs in disjunct populations, which may berecognizably distinct genetically and therefore requireseparate management. Second, C. deserticoloides co-occurs with several morphologically similar species,whose level of divergence is unclear. We have usedDNA sequences to address these issues, as a basis for
gy, The Natural History Museum, Cromwell Road, London SW7 5BD, UK.

A . C . D i o g o e t a l .
evaluating conservation priorities and for designatingscientifically sound conservation strategies for thishighly endangered species and its unique habitat.
Survey of existing populations
C. deserticoloides has recently been collected in fourlocalities, and records from four additional localitiesare found in the literature (Table 1). The four siteswhere beetles are currently known to occur have onlybeen discovered in recent years after thorough surveysof potential habitats (Gallego et al., 1997) Gimenez,unpublished). Given the extent of these surveys it isunlikely that more sites sustaining substantial popula-tions of the species will be found. It is also unlikely thatthe sites reported in the literature account for addi-tional populations of C. deserticoloides. Two of the his-torical sites, Elche and Santa Pola, were surveyed by usmany times, but beetles were never encountered. Therecord from Albatera, the type locality, could never beconfirmed in our surveys; the record may actually per-tain to our location at San Isidro de Albatera, some fivekilometres south of Albatera. Finally, the reportedoccurrence at Totana (Sauleda, 1985) is about 1 kmnorth of the current site at La Alcanara, but both placesare part of the large salt steppe of the area Totana-Alhama and they may therefore not represent inde-pendent records. All current and historical sites of C.deserticoloides are confined to two (ecologically) distinctregions (Fig. 1). One is along a 100 km stretch in theGuadalentin and Segura valleys including the localitiesof La Alcanara in the west, Albatera and Arneva in themiddle of the valley and Elche and Santa Pola in theeast. The second area is the Rambla de Ajauque, a tem-porary stream tributary of the Segura river in the prox-
Table 1. Historical and present localities extant localities. LA, SA, and RA are the popDNA analysis
Localities
*La Alcanara (Alhama, Murcia) ‘LA’*Arneva (Orihuela, Alicante)*San Isidro de Albatera (Alicante) ‘SA’*Rambla de Ajauque (Fortuna, Murcia) ‘RA’Albatera (Alicante)Elche (Alicante)Santa Ploa (Alicante)Totana (Murcia)
11
imity of Arneva. Historically, the Guadalentin andSegura valley was a huge area of salt steppes that isfragmented nowadays for agricultural development.The Rambla de Ajauque constitutes a compartmental-ized ensemble of small salt steppes connected by tem-porary streams in a very arid landscape.
Habitats at all localities where C. deserticoloides wasfound can be described as the characteristic salt steppesof arid or semi-arid regions of the Iberian Peninsula(Bernaldez, 1992). Steppes are flat areas with saline soilwhich experience little or no flooding (i.e. they are dryduring most of the year) but are affected by salinesubterranean waters. The vegetation typically consistsof a shrub layer (Sarcocornia fructicosa, Saueda veraand Halocnemun strobilaceum in La Alcanara) and achamaephyte layer (Frankenia corymbosa and severalLymonium species) but vegetation cover is less than50% of the surface area (Esteve et al., 1995).
At present, available habitat is reduced to very fewisolated patches in both main regions of the geographicrange. At Arneva and San Isidro de Albatera there areonly a few patches of several hundred square meterswhich are very fragmented and altered. The locality atLa Alcanara is the best preserved although it is alsofragmented within the agricultural landscape. Someparts of the salt steppes (about 7 km2) are subject tocycles of culture-abandonment-secondary succession(Caballero et al., 1994). The patches of Ajauque arethreatened by the rise of the phreatic level due to theestablishment of recent irrigation in the watershed ofthe local aquifer. This led to the replacement ofsalt steppes by halophyte communities dominated byPhragmites australis, a habitat not suitable for C.deserticoloides.
for C. deserticoloides. Asterisks indicate theulation codes for the three specimens of our
References
Gallego et al. (1997)Gimenez (unpublished results)Ortiz et al. (1987); present paperGallego et al. (1997)Codina (1931); Ortiz et al. (1987)Vives and Vives (1978)Vives and Vives (1978); Sauleda (1985)Sauleda (1985)
8
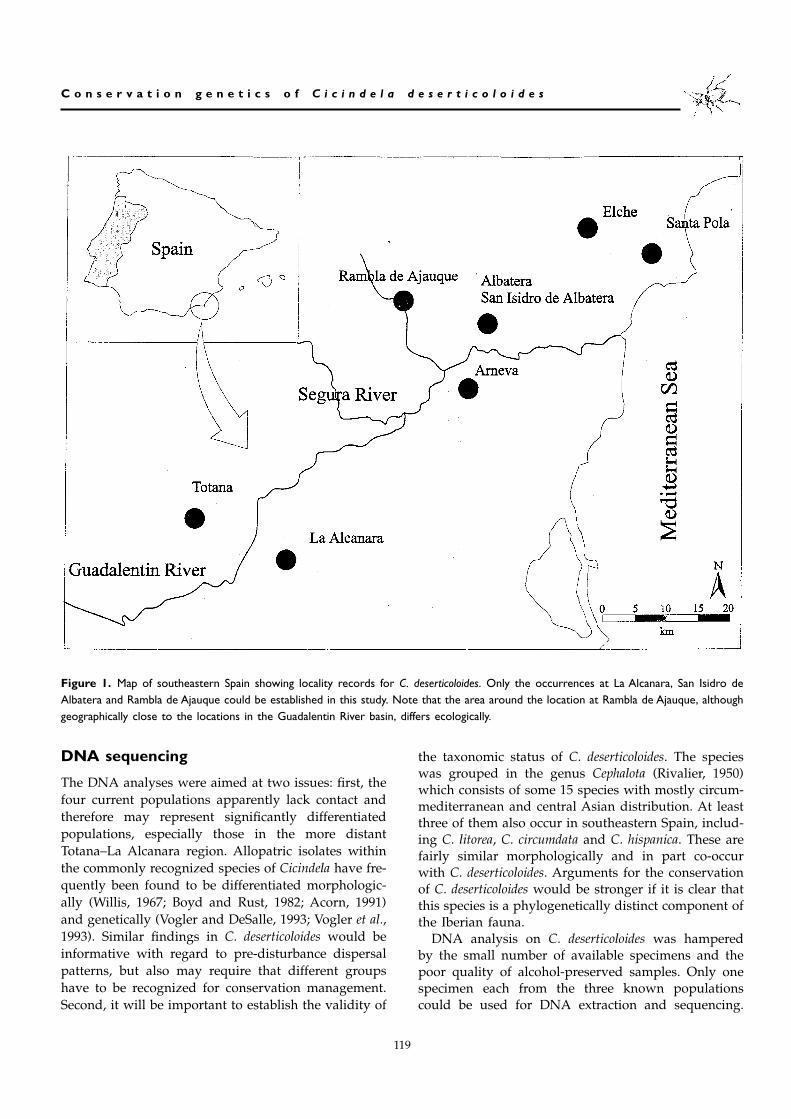
Figure 1. Map of southeastern Spain showing locality records for C. deserticoloides. Only the occurrences at La Alcanara, San Isidro deAlbatera and Rambla de Ajauque could be established in this study. Note that the area around the location at Rambla de Ajauque, althoughgeographically close to the locations in the Guadalentin River basin, differs ecologically.
C o n s e r v a t i o n g e n e t i c s o f C i c i n d e l a d e s e r t i c o l o i d e s
DNA sequencing
The DNA analyses were aimed at two issues: first, thefour current populations apparently lack contact andtherefore may represent significantly differentiatedpopulations, especially those in the more distantTotana–La Alcanara region. Allopatric isolates withinthe commonly recognized species of Cicindela have fre-quently been found to be differentiated morphologic-ally (Willis, 1967; Boyd and Rust, 1982; Acorn, 1991)and genetically (Vogler and DeSalle, 1993; Vogler et al.,1993). Similar findings in C. deserticoloides would beinformative with regard to pre-disturbance dispersalpatterns, but also may require that different groupshave to be recognized for conservation management.Second, it will be important to establish the validity of
11
the taxonomic status of C. deserticoloides. The specieswas grouped in the genus Cephalota (Rivalier, 1950)which consists of some 15 species with mostly circum-mediterranean and central Asian distribution. At leastthree of them also occur in southeastern Spain, includ-ing C. litorea, C. circumdata and C. hispanica. These arefairly similar morphologically and in part co-occurwith C. deserticoloides. Arguments for the conservationof C. deserticoloides would be stronger if it is clear thatthis species is a phylogenetically distinct component ofthe Iberian fauna.
DNA analysis on C. deserticoloides was hamperedby the small number of available specimens and thepoor quality of alcohol-preserved samples. Only onespecimen each from the three known populationscould be used for DNA extraction and sequencing.
9
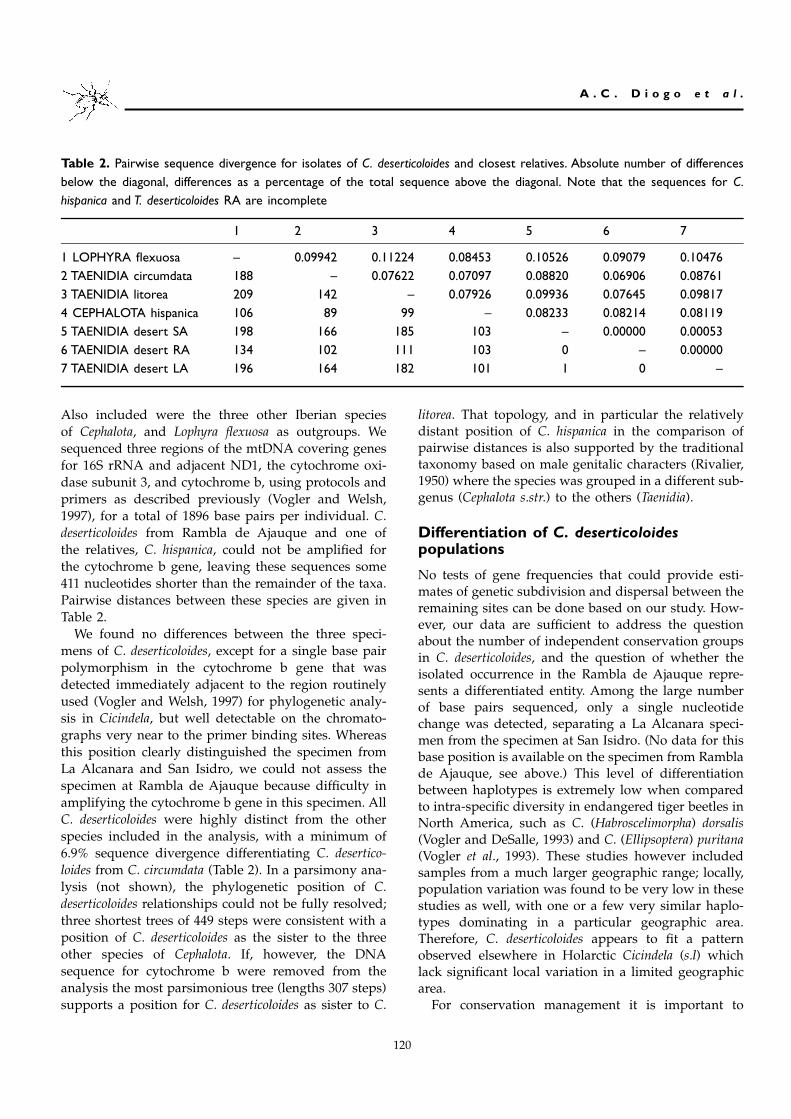
Table 2. Pairwise sequence divergence for isolates of C. deserticoloides and closest relatives. Absolute number of differencesbelow the diagonal, differences as a percentage of the total sequence above the diagonal. Note that the sequences for C.hispanica and T. deserticoloides RA are incomplete
1 2 3 4 5 6 7
1 LOPHYRA flexuosa – 0.09942 0.11224 0.08453 0.10526 0.09079 0.104762 TAENIDIA circumdata 188 – 0.07622 0.07097 0.08820 0.06906 0.087613 TAENIDIA litorea 209 142 – 0.07926 0.09936 0.07645 0.098174 CEPHALOTA hispanica 106 89 99 – 0.08233 0.08214 0.081195 TAENIDIA desert SA 198 166 185 103 – 0.00000 0.000536 TAENIDIA desert RA 134 102 111 103 0 – 0.000007 TAENIDIA desert LA 196 164 182 101 1 0 –
A . C . D i o g o e t a l .
Also included were the three other Iberian speciesof Cephalota, and Lophyra flexuosa as outgroups. Wesequenced three regions of the mtDNA covering genesfor 16S rRNA and adjacent ND1, the cytochrome oxi-dase subunit 3, and cytochrome b, using protocols andprimers as described previously (Vogler and Welsh,1997), for a total of 1896 base pairs per individual. C.deserticoloides from Rambla de Ajauque and one ofthe relatives, C. hispanica, could not be amplified forthe cytochrome b gene, leaving these sequences some411 nucleotides shorter than the remainder of the taxa.Pairwise distances between these species are given inTable 2.
We found no differences between the three speci-mens of C. deserticoloides, except for a single base pairpolymorphism in the cytochrome b gene that wasdetected immediately adjacent to the region routinelyused (Vogler and Welsh, 1997) for phylogenetic analy-sis in Cicindela, but well detectable on the chromato-graphs very near to the primer binding sites. Whereasthis position clearly distinguished the specimen fromLa Alcanara and San Isidro, we could not assess thespecimen at Rambla de Ajauque because difficulty inamplifying the cytochrome b gene in this specimen. AllC. deserticoloides were highly distinct from the otherspecies included in the analysis, with a minimum of6.9% sequence divergence differentiating C. desertico-loides from C. circumdata (Table 2). In a parsimony ana-lysis (not shown), the phylogenetic position of C.deserticoloides relationships could not be fully resolved;three shortest trees of 449 steps were consistent with aposition of C. deserticoloides as the sister to the threeother species of Cephalota. If, however, the DNAsequence for cytochrome b were removed from theanalysis the most parsimonious tree (lengths 307 steps)supports a position for C. deserticoloides as sister to C.
12
litorea. That topology, and in particular the relativelydistant position of C. hispanica in the comparison ofpairwise distances is also supported by the traditionaltaxonomy based on male genitalic characters (Rivalier,1950) where the species was grouped in a different sub-genus (Cephalota s.str.) to the others (Taenidia).
Differentiation of C. deserticoloidespopulations
No tests of gene frequencies that could provide esti-mates of genetic subdivision and dispersal between theremaining sites can be done based on our study. How-ever, our data are sufficient to address the questionabout the number of independent conservation groupsin C. deserticoloides, and the question of whether theisolated occurrence in the Rambla de Ajauque repre-sents a differentiated entity. Among the large numberof base pairs sequenced, only a single nucleotidechange was detected, separating a La Alcanara speci-men from the specimen at San Isidro. (No data for thisbase position is available on the specimen from Ramblade Ajauque, see above.) This level of differentiationbetween haplotypes is extremely low when comparedto intra-specific diversity in endangered tiger beetles inNorth America, such as C. (Habroscelimorpha) dorsalis(Vogler and DeSalle, 1993) and C. (Ellipsoptera) puritana(Vogler et al., 1993). These studies however includedsamples from a much larger geographic range; locally,population variation was found to be very low in thesestudies as well, with one or a few very similar haplo-types dominating in a particular geographic area.Therefore, C. deserticoloides appears to fit a patternobserved elsewhere in Holarctic Cicindela (s.l) whichlack significant local variation in a limited geographicarea.
For conservation management it is important to
0

C o n s e r v a t i o n g e n e t i c s o f C i c i n d e l a d e s e r t i c o l o i d e s
recognize those groups of individuals or populationsthat constitute independent gene pools, because suchgenetically independent groups have a key role in theevolutionary process which conservation programmesattempt to conserve (Vogler and DeSalle, 1994; Cracraft,1997). Principally a single nucleotide polymorphism,such as the one separating the specimens from LaAlcanara and San Isidro, can be diagnostic (sensuCracraft, 1983) distinguishing all individuals in onepopulation from those in another and indicating com-plete separation of two such groups. For example, thefinding of a single base consistently differentiating allindividuals of an isolated population of C. dorsalis fromother populations was sufficient to indicate high con-servation status as a last remnant of a larger assem-blage now largely extinct (Vogler, 1998). Because only asingle specimen from each population of C. desertico-loides was included in our analysis we cannot judgewhether or not any of the three surviving populationsshould constitute such a separate entity, and it wouldbe necessary to include further specimens for thisdetermination. However, it is reasonable to assume thatthese two distinguishable haplotypes are not confinedgeographically, and if they were, the separation is shal-low and certainly does not indicate deep evolutionarypartitions. For management purposes, it is also import-ant to keep in mind that tiger beetle species are fre-quently persisting in a network of habitat patchessubject to succession and (natural) disturbance fromwhich populations disappear. Therefore a minimumnumber of populations is required for the long-termpersistence of species; with only very few populationsof C. deserticoloides left, and in the absence of posi-tive evidence for genetic subdivision, all populationsshould therefore be considered a single entity for con-servation purposes. The analysis of additional speci-mens is unlikely to add anything to this conclusion.
Sequence divergence in closely relatedspecies of Cephalota
While the different isolates within C. deserticoloidesare little divergent, they exhibit substantial DNAdivergence to their closest relatives in the genusCephalota. Uncorrected levels of sequence divergencewere between 6.9% and 9.9% (Table 2). Compared toother species-level phylogenies for similar subgroupswithin the genus Cicindela (s.l.) from North America,divergence between species ranged between 0.7% and8.9% in the C. maritima group and 0.8% and 9.5% in thegenus Ellipsoptera. Thus, taxa within Cephalota are gen-erally at the upper level of those found between species
12
in the other species groups. It has been proposed that aspecies is more valuable to conserving biotic diversitythe more divergent it is from its closest relatives. Thisreasoning led authors to set conservation prioritiesbased on the total amount of diversity that is unique toa particular species (feature diversity of Faith, 1992).MtDNA with its generally clock-like rate of mutationprovides a good approximation for the amount ofdiversity contained in a lineage and its unique features.Accepting the rationale of conservation priorities basedon phylogenetic distances, data on branch length in C.deserticoloides indeed strengthen the case for protectionof this species.
This conclusion, however, has to be considered in thelight of incomplete sampling of the genus Cephalota. Assuggested by its name, C. deserticoloides is similar inmorphological features to C. deserticola, a species withwide geographic distribution in central Asia. The spe-cies is rare in collections (Werner, 1992), and very littleis known about its current distribution and abundance.Inclusion of this species in the DNA analysis maychange the conclusions about the level of divergencefrom the closest relative of C. deserticoloides and theuniqueness of its features. However, in the absence ofany reliable data on C. deserticola, and given the factthat the nearest locations for that species are inArmenia and northern Iran, it is probably justified toconsider the status of C. deserticoloides irrespectiveof the situation in the related species. Therefore C.deserticoloides should be considered a unique element ofthe Iberian fauna deserving high priority for conser-vation.
The causes of endangerment
Why is C. deserticoloides more endangered than its closerelatives? Habitat requirements in Cicindela are evolu-tionarily conservative, and in many cases small groupsof closely related species occur in very similar habitat(Vogler and Goldstein, 1997). This is also true for thespecies of Cephalota which occur associated with salineconditions, near water bodies and small lagoons (Table3). Related species should therefore be similarly en-dangered which does not seem to be the case, butremains to be investigated in more detail. However, C.deserticoloides appears to be more limited in its habitatrequirements than its congeners. Animal and plantcommunities associated with salt steppes are organizedaccording to the ecological gradients determined by thewater availability and salinity of the soil. Tiger beetlecommunities are organized according to the same gra-dient (Ganeshaiah and Belavadi, 1986; Hidalgo et al.,
1

Table 3. Iberian species in the genus Cephalota, their taxonomic affinities and their distributional range. Data on distribution andhabitat from Werner (1992)
Species Subgenus Distribution Range Habitat
C. hispanica Cephalota southern Iberia narrow brackish water
C. litorea Taenidia circum-Mediterranean,Red Sea, Sudan, Somalia
widespread halophilous, sea andlakes
C. circumdata Taenidia circum-Mediterranean widespread saline riparian
C. deserticoloides Taenidia southern Spain narrow dry saline areas
A . C . D i o g o e t a l .
1995). Although there are no detailed studies abouthabitat selection in C. deserticoloides its requirements seemto be very stringent. While co-occurring tiger beetlesspecies such as C. litorea and Megacephala euphratica alsoselect habitats where the soil is wet or even inundatedfor longer periods of time, the preferred habitat of C.deserticoloides is narrowly limited to the dry parts ofthe area. This fact could account for differences in thedegree of endangerment between close relatives pos-sibly because dryer locations are less common or maybe more affected by current development.
A second factor determining the degree of endanger-ment is the level of rarity, both with regard to localabundance and the extent of the total geographicrange. Although we have not made an accurate estima-tion of population size, C. deserticoloides populationsseem to be small compared to related species such as C.litorea which is more widely distributed and was con-sistently found in higher numbers. The spatial matrixin which populations exist depends on the availabilityof suitable saline flats in the area, which also stronglyaffect the overall abundance and metapopulation struc-ture. Even before widespread development in theregion, C. deserticoloides has been limited to a small setof locations, indicating either narrow habitat require-ments not permitting the use of additional sites, orpoor dispersal capabilities. Both would affect the over-all abundance of this species and could result in shortpersistence times at a given location and low chances ofrecolonization. This could also affect the extent of thetotal distributional range. In contrast to C. litorea and C.circumdata with their wide distribution throughout theMediterranean and Asia (Table 3), C. deserticoloides is anarrow endemic of southeastern Spain. It is possiblethat factors causing low local abundance are reflectedalso in the small global range of the species, as thetendency to extinction on a local level could translateinto more limited global distributions.
122
Implications for regional conservationmanagement
Because of their narrow ecological requirements cicin-delids are excellent taxa to monitor environmentalchanges and degradation affecting their habitats (Pear-son and Cassola, 1992). The current decline of the cicin-delid fauna in the salt flats of southeastern Spaincertainly reflects vulnerability of other components ofthis unique ecosystem, including plants, birds andother insects. The presence of the endemic C. desertico-loides might indicate the existence of other endemicforms in this region, such as the tenebrionid Alphasidalorcana. C. deserticoloides is therefore a useful focal taxonfor conservation, indicative of a relict assemblage froma formerly much wider distribution that was appar-ently only able to persist in this particular region. Pro-tection of the remaining locations of C. deserticoloides istherefore of high priority and will also be useful in pro-tecting a larger assemblage of genetically unique com-ponents. This has been recognized by the authorities,and the salt steppes of La Alcanara and Ajauque aredesignated protected areas by a Murcian regional law(4/1992; BORM). Unfortunately, their protection is notcurrently enforced.
Acknowledgements
We thank J. Serrano, M.A. Esteve and J.L. Lencina forcomments and locality information. Grant support byPortuguese JNICT (to ACD), British NERC (grant GR3/10632, to APV) and Spanish DGICYT (PB95–1005, toJG) is gratefully acknowledged.
References
Acorn, J.H. (1991) Habitat associations, adult life histories,and species interactions among sand dune tiger beetles inthe Southern Canadian prairies (Coleoptera: Cicindelidae).Cicindela, 23, 17–47.

C o n s e r v a t i o n g e n e t i c s o f C i c i n d e l a d e s e r t i c o l o i d e s
Bernaldez, F.G. (1992) Los paisajes del agua: Terminologıapopular de los humedales. J.M.Reyero, Madrid, Pages.
Boyd, H.P. and Rust, R.W. (1982) Intraspecific and geo-graphical variations in Cicindela dorsalis (Coleoptera:Cicindelidae). Coleopts Bull., 36, 221–39.
Caballero, J.M., Esteve, M.A., Calvo, J.F. and Pujol, J.A.(1994) Estructura de la vegetacion de los saladares delGuadalentın (Region de Murcia). Estudia Oecologica,171–83.
Codina, A. (1931) Una Cicindela (Col.) nueva de Espana. Bol.Soc. Ent. Esp., 14, 161–64.
Cracraft, J. (1997) Species concepts in systematics and con-servation biology – an ornithological viewpoint. In: M.F.Claridge, H.A. Dawah and M.R. Wilson (ed.) Species. Theunits of biodiversity , pp. 325–40. London: Chapman andHall.
Esteve, M.A., Caballero, J.M., Gimenez, A., Aledo, E., Baraza,F., Guirao, J., Robledano, F. and Torres, A. (1995) Los pai-sajes del agua en la Region de Murcia. Caracterizacionambiental y perspectivas de gestion de humedales. In: M.Senent and F. Cabezas (eds) Agua y futuro en la Region deMurcia, pp. 570. Murcia: Asamblea Regional.
Faith, D.P. (1992) Conservation evaluation and phylogeneticdiversity. Biol. Conserv., 61, 1–10.
Gallego, D., Gimenez, A., Esteve, M.A. and Serrano, J. (1997)Seleccion de habitat y patron de actividad temporal deTaenidia deserticoloides Codina (Col., Carabidae). Abstractof the 5th Meeting of the Asociacion Espanola de EcologıaTerrestre (AEET). Cordoba, Spain.
Ganeshaiah, K.N. and Belavadi, V.V. (1986) Habitat segrega-tion in four species of adult tiger beetles (Coleoptera:Cicindelidae). Ecol. Entomol., 11, 147–54.
Hidalgo, J., Ballesta, M., Ruano, F. and Tinaut, A. (1995) Dis-tribucion de los Cicindelidos en un ambiente dunar.Punta Entinas-El Sabinar (Almerıa, Espana). (Coleoptera:Cicindelidae). Ecologia, 9, 469–74.
Knisley, C.B. and Hill, J.M. (1992) Effects of habitat changefrom ecological succession and human impact on tigerbeetles. Virginia Journal of Science, 43, 133–42.
Palao, F.M.M. (1909) Saneamiento de tierras humedas y salo-brenas. In: J.A. Gimenez (ed.), pp. 89. Murcia.
Pearson, D.L. (1988) Biology of tiger beetles. Annu. Rev.Entomol., 33, 123–47.
Pearson, D.L. and Cassola, F. (1992) World-wide species rich-
12
ness patterns of tiger beetles (Coleoptera: Cicindelidae):indicator taxon for biodiversity and conservation studies.Conserv. Biol., 6, 376–91.
Rivalier, E. (1950) Demembrement du genre Cicindela Linne.Rev. Franc. Entomol., 17, 217–44.
Sauleda, N. (1985) Caraboidea ammofilos y halofilos de laprovincia de Alicante. Ann. Univ. Alicant, 241–64.
Schultz, T.D. (1989) Habitat preference and seasonal abun-dances of eight sympatric species of tiger beetle, genusCicindela (Coleoptera: Cicindelidae) in Bastrop StatePark, Texas. Southw. Nat., 34, 468–77.
Stamatov, J. (1972) Cicindela dorsalis Say endangered on theNorthern Atlantic Coast. Cicindela, 4, 78.
Vogler, A.P. (1998) Extinction and the evolutionary process:what to conserve? In: R. DeSalle and B. Schierwate (eds)Molecular approaches in ecology and evolution, pp. 191–210.Basel: Birkhauser.
Vogler, A.P. and DeSalle, R. (1993) Phylogeographic patternsin coastal North American Tiger Beetles, Cicindela dorsalisinferred from mitochondrial DNA sequences. Evolution,47, 1192–1202.
Vogler, A.P. and DeSalle, R. (1994) Diagnosing units of con-servation management. Conserv. Biol., 8, 354–63.
Vogler, A.P. and Goldstein, P.Z. (1997) Adaptation, clado-genesis, and the evolution of habitat association in NorthAmerican tiger beetles: A phylogenetic perspective. In:T. Givnish and K. Systma (eds) Molecular evolution andadaptive radiation, pp. 353–73. Cambridge: CambridgeUniversity Press.
Vogler, A.P., Knisley, C.B., Glueck, S.B., Hill, J.M. and DeSalle,R. (1993) Using molecular and ecological data to diag-nose endangered populations of the Puritan tiger beetle,Cicindela puritana. Mol. Ecol., 2, 375–83.
Vogler, A.P. and Welsh, A. (1997) Phylogeny of North Amer-ican Cicindela tiger beetles inferred from multiple mito-chondrial DNA sequences. Mol. Phylogenet. Evol., 8,225–35.
Werner, K. (1992) The beetles of the world. Cicindelidae 2.Sciences Nat., Venette, France, Pages.
Willis, H.L. (1967) Bionomics and zoogeography of tigerbeetles of saline habitats in the central United States(Coleoptera: Cicindelidae). Univ. Kans. Sci. Bull., 47,145–313.
3