Embed Size (px)
Citation preview

Development 1989 Supplement, 37-51 (1989)Printed in Great Britain © T h e Company of Biologists Limited 1989
37
Cortical rotation of the Xenopus egg: consequences for the anteroposterior
pattern of embryonic dorsal development
J. GERHART, M. DANILCHIK, T. DONIACH, S. ROBERTS, B. ROWNING and R. STEWART
Department of Molecular Biology, University of California, Berkeley CA 94720 USA
Summary
We first review cortical-cytoplasmic rotation, a micro-tubule-mediated process by which the Xenopus egg, likeother amphibian eggs, transforms its polarized cylindri-cal symmetry into bilateral symmetry within the first cellcycle after fertilization. This transformation, the earliestof many steps leading to dorsal development, involvesthe displacement of the egg's cortex relative to itscytoplasmic core by 30° in an animal-vegetal direction.As rotation is progressively reduced by microtubule-depolymerizing agents, embryos develop with body axesprogressively deleted for dorsal structures at the an-terior end. With no rotation, ventralized embryos areformed. In an effort to comprehend this progressiveeffect on embryonic organization, we go on to reviewsubsequent developmental processes depending on ro-tation, and we propose, with evidence, that reducedrotation leads to a reduced number of vegetal dorsalizing
cells, which induce during the blastula stage a Spemannorganizer region of smaller than normal size. Thereduced organizer then promotes a reduced amount ofcell rearrangement (morphogenesis) at gastrulation.Reduced morphogenesis seems the proximate cause ofthe incompleteness of axial pattern, as shown further bythe fact that embryos that are normal until the gastrulastage, if exposed to inhibitors of morphogenesis, developbody axes that are progressively less complete in theiranterior dorsal organization the earlier their gastru-lation had been blocked. We discuss why axial patternmight depend systematically on morphogenesis.
Key words: cortical rotation, Xenopus laevis, mesoderminduction, microtubules, gastrulation, polarity, FGF, TGF/J,anteroposterior pattern, dorsoventral pattern, lithium,D2O, organizer.
I. Organization and reorganization of the egg:
The egg's initial symmetry arises in oogenesisThe unfertilized Xenopus egg displays polarized cylin-drical symmetry around an axis connecting the poles ofthe animal and vegetal hemispheres (these hemisphereshenceforth abbreviated as AH and VH). The twohemispheres differ as the result of various localizationmechanisms segregating materials during oogenesis,perhaps in relation to the oocyte's inherent axis com-prising the nucleus, centrosome and division bridge(Tourte et al. 1984; Wylie et al. 1985; Danilchik &Gerhart, 1987; Yisraeli et al. this volume). The bound-ary between hemispheres (the 'equator') is roughlyidentified externally as the limit of dark pigment of theanimal half, although this marking may not quitecoincide with the internal interface of materials specificto the two hemispheres. Oocyte organization corre-sponds roughly to the germ layer organization of theembryo: ectodermal tissues arise from AH materials,endodermal tissues from VH materials, and mesodermand archenteron roof endoderm from equatorial ma-terials (reviewed in Gerhart & Keller, 1986).
All pole-to-pole meridians of the egg surface are
indistinguishable before fertilization. Sperm can beapplied at any point in the animal hemisphere (Elinson,1980; K. Hara, unpublished; J. Gerhart, unpublished),and that point of entry identifies a meridian thatcoincides approximately but not perfectly with theeventual ventral midline of the embryo. The oppositemeridian falls on the dorsal midline of the embryo, ormore correctly, on the midline of the neural plate.Many experiments, some discussed later, indicate thatthe egg has an initial capacity to develop dorsally at any,several, or even all, meridians, and the same is true ofventral development. Normal embryonic developmentarises from the proportioned, patterned use of thispotential, normally in relation to the sperm entry point.
Bilateralization of the egg after fertilizationThe Xenopus egg reorganizes its cytoplasmic contentsin an interval between 45 and 90min after fertilization,a G2-like period of the first cell cycle (100min, 18°C), aperiod absent from the next 11 or 12 rapid cycles(35 min each). Within this interval, the egg behaves as ifcomposed of two rigid units, a thin cortex (2-5 /zmthick) and a large spherical core (1200/im diameter). Asdiagrammed in Fig. 1, these rotate relative to one
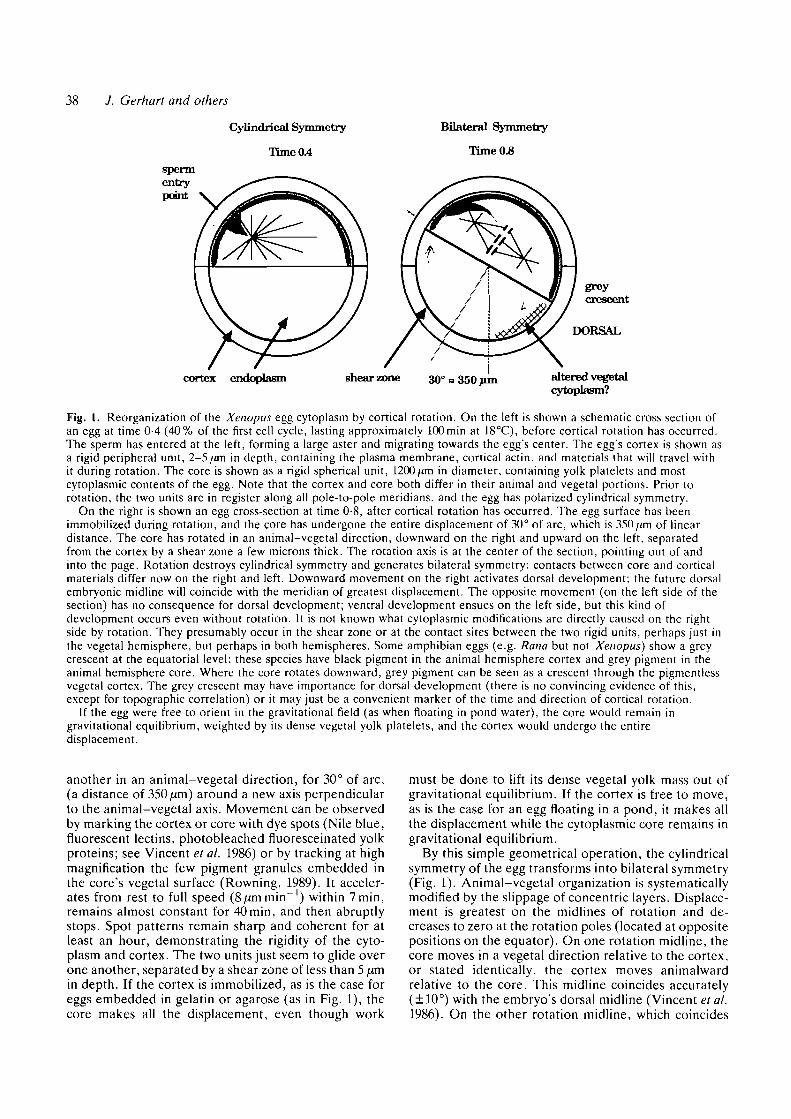
38 J. Gerhart and others
Cylindrical Symmetry
lime 0.4
Bilateral Symmetry
Time 0.8
cortex endoplasm 350jim
greycrescent
DORSAL
altered vegetalcytoplasm?
Fig. 1. Reorganization of the Xenopus egg cytoplasm by cortical rotation. On the left is shown a schematic cross section ofan egg at time 0-4 (40% of the first cell cycle, lasting approximately 100min at 18CC), before cortical rotation has occurred.The sperm has entered at the left, forming a large aster and migrating towards the egg's center. The egg's cortex is shown asa rigid peripheral unit, 2-5/an in depth, containing the plasma membrane, cortical actin, and materials that will travel withit during rotation. The core is shown as a rigid spherical unit, 1200/an in diameter, containing yolk platelets and mostcytoplasmic contents of the egg. Note that the cortex and core both differ in their animal and vegetal portions. Prior torotation, the two units are in register along all pole-to-pole meridians, and the egg has polarized cylindrical symmetry.
On the right is shown an egg cross-section at time 0-8, after cortical rotation has occurred. The egg surface has beenimmobilized during rotation, and the core has undergone the entire displacement of 30° of arc, which is 350/im of lineardistance. The core has rotated in an animal-vegetal direction, downward on the right and upward on the left, separatedfrom the cortex by a shear zone a few microns thick. The rotation axis is at the center of the section, pointing out of andinto the page. Rotation destroys cylindrical symmetry and generates bilateral symmetry: contacts between core and corticalmaterials differ now on the right and left. Downward movement on the right activates dorsal development; the future dorsalembryonic midline will coincide with the meridian of greatest displacement. The opposite movement (on the left side of thesection) has no consequence for dorsal development; ventral development ensues on the left side, but this kind ofdevelopment occurs even without rotation. It is not known what cytoplasmic modifications are directly caused on the rightside by rotation. They presumably occur in the shear zone or at the contact sites between the two rigid units, perhaps just inthe vegetal hemisphere, but perhaps in both hemispheres. Some amphibian eggs (e.g. Rana but not Xenopus) show a greycrescent at the equatorial level; these species have black pigment in the animal hemisphere cortex and grey pigment in theanimal hemisphere core. Where the core rotates downward, grey pigment can be seen as a crescent through the pigmentlessvegetal cortex. The grey crescent may have importance for dorsal development (there is no convincing evidence of this,except for topographic correlation) or it may just be a convenient marker of the time and direction of cortical rotation.
If the egg were free to orient in the gravitational field (as when floating in pond water), the core would remain ingravitational equilibrium, weighted by its dense vegetal yolk platelets, and the cortex would undergo the entiredisplacement.
another in an animal-vegetal direction, for 30° of arc,(a distance of 350 ^m) around a new axis perpendicularto the animal-vegetal axis. Movement can be observedby marking the cortex or core with dye spots (Nile blue,fluorescent lectins, photobleached fluoresceinated yolkproteins; see Vincent et al. 1986) or by tracking at highmagnification the few pigment granules embedded inthe core's vegetal surface (Rowning, 1989). It acceler-ates from rest to full speed (8/<mmin~') within 7 min,remains almost constant for 40min, and then abruptlystops. Spot patterns remain sharp and coherent for atleast an hour, demonstrating the rigidity of the cyto-plasm and cortex. The two units just seem to glide overone another, separated by a shear zone of less than 5 /«nin depth. If the cortex is immobilized, as is the case foreggs embedded in gelatin or agarose (as in Fig. 1), thecore makes all the displacement, even though work
must be done to lift its dense vegetal yolk mass out ofgravitational equilibrium. If the cortex is free to move,as is the case for an egg floating in a pond, it makes allthe displacement while the cytoplasmic core remains ingravitational equilibrium.
By this simple geometrical operation, the cylindricalsymmetry of the egg transforms into bilateral symmetry(Fig. 1). Animal-vegetal organization is systematicallymodified by the slippage of concentric layers. Displace-ment is greatest on the midlines of rotation and de-creases to zero at the rotation poles (located at oppositepositions on the equator). On one rotation midline, thecore moves in a vegetal direction relative to the cortex,or stated identically, the cortex moves animalwardrelative to the core. This midline coincides accurately(±10°) with the embryo's dorsal midline (Vincent et al.1986). On the other rotation midline, which coincides

Rotation of the Xenopus egg 39
with the embryonic ventral midline, displacement hasthe opposite sense, and, as discussed later, this sensehas no consequence for development since ventraldevelopment occurs even without rotation. The ventralmidline just seems by default the farthest position fromthe dorsal midline.
This reorganization process has received manynames: subcortical rotation, the rotation of symmetriz-ation or cortical/cytoplasmic rotation. We will call itcortical rotation for brevity. It was first inferred byBanki in 1929 for axolotl eggs and described in detail byAncel & Vintemberger (1948) for Ranafusca eggs, whorecognized it as the process forming the grey crescent, alightly pigmented subequatorial region marking theembryo's future dorsal side, including its organizerregion. Cortical rotation has also been inferred in Ranapipiens eggs from grey crescent formation and fromdisplacement of the polar body spot (Elinson, 1980).Rotation was not detected in Xenopus eggs until ourstudies with artificially marked eggs because Xenopuseggs rarely form a grey crescent; this is simply becausethe pigment granules reside entirely in the core duringrotation. Probably most amphibian eggs make use ofcortical rotation, as do the eggs of some fish (ancientgroups such as lungfish and sturgeons) which displaygrey crescents and complete cleavage.
Cortical rotation correlates well with embryonicdorsoventral organizationRotation is normally oriented toward the sperm entrypoint (SEP), as is the dorsoventral organization of theembryo. L0vtrop (1965) has proposed that the truedorsalizing process in Xenopus eggs may not be corticalrotation but a contraction of cortical materials towardthe sperm entry point. However, there is now abundantevidence that rotation is the more crucial rearrange-ment for embryonic dorsoventral development, andthat the sperm only affects embryonic organization byinfluencing (via its sperm aster) the direction of ro-tation:
(1) Under normal conditions of fertilization anddevelopment, the direction of rotation more accuratelypredicts the embryonic dorsal midline than does theSEP (Vincent et al. 1986).
(2) Eggs can be tipped out of gravitational equilib-rium, or squeezed laterally in various directions beforeor during cortical rotation, and these treatments ran-domize the relation of the SEP to the embryonic axes,whereas the direction of rotation still accurately corre-lates (Gerhart et al. 1980; Vincent et al. 1986; Black &Vincent, 1988).
(3) Dispermic and trispermic eggs initiate a singledirection of rotation which predicts the dorsal midline(Vincent & Gerhart, 1987).
(4) Artificially activated eggs, which lack an SEP,rotate in a unique direction and, if later transplantedwith a nucleus and centrosome, will develop an embryohaving its dorsal midline at the position predicted by therotation direction (Vincent & Gerhart, 1987; J. Roberts& B. Rowning, unpublished).
(5) When fertilized eggs fail to engage in cortical
D
Fig. 2. Schematic diagram of parallel microtubules in thevegetal hemisphere of the Xenopus egg during corticalrotation. The egg is cut in the bilateral plane, showing thesperm aster in the animal hemisphere (upper half), and theparallel microtubules in the periphery of the vegetalhemisphere (lower half). The tubule array exists only in theinterval from 0-4-0-9 of the first cell cycle, at the time ofcortical rotation. The sheet of tubules is only 2-4/un thick,starting approximately 2/tm from the egg surface. Tubulesare aligned in the direction of rotation, shown by thearrows; their polarity is not known. Ventral (V) and dorsal(D) development will occur on opposite meridians, asindicated. Note that tubules describe tighter half-circles asthe rotation poles ('hubs') are approached.
rotation, they develop no dorsal embryonic structures,as discussed later, despite their SEP.
Taken in its entirety, the evidence is strong thatcortical rotation is an indispensable step in normaldorsal development. The need for it can be bypassedunder certain experimental conditions (e.g. the use oflithium ion), and it is certainly not sufficient by itself fordorsal development; but it seems to be a normal,essential, early member of a series of processes leadingto dorsal development. The role of the sperm ininfluencing the direction of rotation will be discussedlater.
Microtubules serve as tracks for cortical rotationAt the start of rotation, a thin layer of parallel, slightlysinuous, microtubules (MTs) appears in the vegetalhemisphere, approximately 2-5 ^m beneath the eggsurface (Elinson & Rowning, 1988). The MTs align inthe direction of rotation, describing tighter semicirclesnearer the rotation hubs (Fig. 2). They are dynamic andperhaps associate laterally. They disappear at the end ofrotation. The array is probably anchored to the core (B.Rowning and T. Mitchison, unpublished). If true, thecortex would then have anchored to it the motormolecules (e.g. kinesin or dynein), which presumablycrawl along the tracks of parallel MTs and generate theforce to displace the rigid cortex relative to the rigidcore. The polarity of the MTs is not known, and so wecannot yet speculate on the need for kinesin as opposedto dynein. However, since movement is unaffected byinjected sodium vanadate (10-100 ^M) combined withlong wavelength UV irradiation of the vegetal eggsurface (365 nm), we currently disfavor the possibility ofa dynein-like motor molecule.
Many agents depolymerize MTs of the array, and allof these inhibit cortical rotation. These include colchi-

40 J. Gerhart and others
cine, vinblastine, nocodazole, cold shock and hydro-static pressure. Also, UV-irradiation (254nm) of thevegetal surface prevents MT polymerization (Grant &Wacaster, 1972; Malacinski et al. 1977; Elinson &Rowning, 1988), perhaps because GTP becomes co-valently bound to tubulin (S. Roberts, 1989). Since allthese agents inhibit rotation, we can assert that MTs areneeded for rotation under normal conditions, probablyas aligned tracks for movement. Cytochalasin D, incontrast, does not interfere with rotation, precluding acontinuous role for dynamic microfilaments. Finally,cycloheximide doesn't inhibit rotation, even when ap-plied before fertilization, indicating that the entireprocess is post-translational, depending only on pre-formed maternal proteins.
What aligns the microtubule array?This is an important issue because as the array gains aunique directionality (one selected from 360° of possi-bilities), rotation is committed to that direction, pro-ducing a unique bilateral symmetry retained throughoutthe rest of development. Before rotation starts at 45 minpostfertilization, there is no detectable MT array.Whatever controls the direction of MTs in the first fewminutes of their polymerization, controls the orien-tation of subsequent embryonic development, undernormal conditions.
We have had difficulty examining the earliest stagesof polymerization because the array builds up soquickly. As rotation accelerates, a few short and poorlyaligned MTs can be seen, but within a few minutes,rotation reaches full speed and the array is complete.To explain the rapid transition from a disordered toordered array, we propose a positive feedback modelwith two provisions, namely: microtubules orient ro-tation, and rotation orients microtubules (Fig. 3). Thefirst provision follows reasonably from what is knownabout MT function, for as the first few tubules polym-erize with poor alignment, a single direction of rotationof the two rigid units (cortex versus core) must nonethe-less emerge as the vector sum of forces generated alongall MTs (their lengths and directions considered). MTsof opposed directions will cancel each other, while anyslight excess of MT polarity in one direction will dictatethe direction of rotation. At first rotation will be slowbecause the vector sum is small.
The second provision is more controversial: canrotation feed back on MT polymerization so that MTstend to arise or persist in the direction of rotation? Asbackground, we note evidence that the direction of MTpolymerization in the periphery of cultured cells re-sponds to the arrangement of elongated tubes ofendoplasmic reticulum (Terasaki et al. 1986). Endoplas-mic reticulum (ER) vesicles and tubes possess kinesin-like MAPs which associate with MTs during theirtransport and elongation (Vale, 1987), and these ves-icle-associated MAPs could in principle stabilize thewalls (rather than ends) of MTs, thus disfavoring MTdepolymerization. If rotation could elongate and alignER in the shear zone between the rigid cortex and core,then the ER might in turn preferentially stabilize MTs
Fig. 3. Positive feedback model for the unidirectionalaligning of vegetal microtubules (MTs) during rotation. Inthe left panel, MTs are first polymerizing in a small regionof the egg's vegetal periphery, and rotation is just starting(approximately time 0-4). MTs are short and poorlyaligned. Tubule polarity is indicated by the solid arrowheads. We assume MTs are anchored to the rigid core.Since kinesin or dynein-like motor molecules, which areassumed anchored to the cortex, must travelunidirectionally along MTs and exert force in the directionof each MT, rotation will start in whatever single directionconstitutes the vector sum of all tubules (tubule directionand length considered). Only one rotation direction canresult (see large open arrow) since there are only two rigidunits. At first, rotation will be slow since the vector sum issmall. Then, as rotation starts, cytoplasmic materials of theshear zone become partially aligned by shear flow (aproposal, not a fact), and these materials stabilize MTs thatform in the direction of alignment, while other tubulesbreak down. The identity of these materials is unknown.The center panel shows the MT array a few minutes later:MTs are more numerous, longer on average, and morealigned in the direction of movement. More force isgenerated in the direction of rotation and now displacementis faster (large open arrow). Faster rotation means moreshear, leading to still more alignment of cytoplasmicmaterials, and still more stabilization of MTs in thatdirection. In the right panel, a few minutes later, MTs arestill longer and better aligned, supporting full-speeddisplacement in the original direction. Since MT orientationdefines the direction of rotation, and since rotation definesthe direction of MT orientation, polymerization androtation have a positive feedback relationship to oneanother. Due to positive feedback, the egg amplifies a smallinitial asymmetry (left panel) into a large one (right panel);the polymerization-rotation process tends strongly todisrupt cylindrical symmetry, as long as it starts in ananimal-vegetal direction.
in that direction. In this way, rotation might influencethe direction of MT polymerization, through the me-diation of shear-aligned ER tubes and vesicles. Withincreased MT alignment, rotation would then be faster,leading to still more alignment of the ER and morepositive feedback on tubule polymerization. Soon MTswould lie overwhelmingly in the rotation direction,directing rotation and ensuring their own persistence inthat direction. The egg would be fully committed to onedirection of rotation, out of all directions initiallypossible (Fig. 3 right panel).

Rotation of the Xenopus egg 41
The initial bias can be smallAccording to this model, any initial slight departurefrom a perfectly random array of MT (a vector sum ofzero) will quickly amplify into a large departure, theunidirectional array. What initial bias might allow a fewmore MTs to form in one direction than in others,thereby controlling the orientation of later embryonicorganization? To begin with, one could argue that, evenif there were no bias, the first MTs are very unlikely totake up a perfectly random orientation, and thereforethe egg will always be able to escape from cylindricalsymmetry, due to positive feedback. A near-randomcondition may be approached in carefully manipulatedeggs which are artificially activated by needle punctureor electric shock. In these, the direction of rotationcan't be predicted beforehand. Usually they succeed inrotating for the full distance at the correct time, thoughoccasionally they choose a skewed or equatorial path.A reliable bias can be introduced in such an egg simplyby tipping it out of gravitational equilibrium for5-10min, at some time before rotation, to achieve asmall displacement of the core relative to the cortex inan animal-vegetal direction (Ancel & Vintemberger,1948; Rowning, 1989). Remarkably this forced displace-ment, which occurs in the absence of MTs, can beterminated long before rotation begins (15-30 min be-fore), and nonetheless, when MTs appear, they align inthe direction of the earlier forced displacement. MT-mediated rotation then continues what tipping hadstarted. Materials of the egg periphery must have beenoriented by the forced movement; these must havepersisted and later biased the direction of MT polym-erization and/or stability. In addition to demonstratingan experimentally imposed bias, this is our best evi-dence that similar displacements might influence thedirection of MT polymerization during the normalcortical rotation period, as part of the positive feedbackmechanism, and that the influence makes use of some-thing other than MTs.
Under normal conditions, though, the sperm pro-vides the dominant bias (Manes and Barbieri, 1977).MTs of the sperm aster extend through the AH longbefore vegetal cortical MTs appear (Stewart-Savage &Grey, 1982; Ubbels et al. 1983). Perhaps MTs of theaster enter the vegetal hemisphere at an early time andsupport a small amount of cortical displacement, leav-ing materials of the shear zone slightly oriented in thesame way as can gravitationally forced displacement.Any slight orientation of materials with respect to theSEP would suffice to bias the whole MT array to buildup in that direction. Actually, the sperm's effect is notstrong; if gravity or centrifugal force is used to displacematerials in another direction in a fertilized egg, thatdisplacement easily comes to provide the dominant bias(Black & Gerhart, 1985).
In general, once the array of MT becomes alignedand rotation reaches full speed, it is difficult to changedirection (Vincent & Gerhart, 1987), although this canoccasionally happen in eggs briefly subjected to lowtemperatures to reversibly depolymerize the MT arrayduring rotation (Vincent & Gerhart, 1987) or can
Fig. 4. Dorsalized and ventralized embryos of Xenopuslaevis. The right embryo developed from an egg UV-irradiated (254 nm, 44 fAV cm"2, 2 min) at time 0-4 of thefirst cell cycle to prevent cortical rotation; that on the leftfrom an egg treated with 60% D2O for 4 min at time 0-28.The right embryo lacks dorsal axial structures, but hasdifferentiated a ciliated epidermis, red blood cells, coelomiccompartments, and a short gut. The left embryo has a largeheart, and circumferential bands of eye pigment and ofcement gland. Both embryos retain the egg's cylindricalsymmetry in their organization.
frequently happen in eggs compressed laterally duringrotation to force movement into a new path (Black &Vincent, 1988). If the MT array is really dynamic and ifpositive feedback operates continuously to define MTdirectionality, these changes would be explicable.
Dorsal development requires rotationEmbryonic organization is drastically altered whenrotation is prevented. The egg, after it has been treatedbriefly (2-4 min) with nocodazole, cold shock, hydro-static pressure, or UV irradiation (of the vegetal surfaceonly) to prevent MT polymerization and cortical ro-tation (Vincent et al. 1987; Elinson & Rowning, 1988),cleaves normally but then gastrulates abnormally, form-ing a ventral-type of blastopore lip which is late andsynchronous around the gastrula circumference (Scharf& Gerhart, 1980, 1983). Although the embryo does notneurulate or form dorsal components of the body axis,it still develops a limited set of ventral structures at allmeridians, preserving the cylindrically symmetric or-ganization of the oocyte (Fig. 4, right). Tissues of allthree germ layers are present in these ventralized 'limitforms': ciliated epidermis, coelomic mesoderm andblood islands (up to 15 times the normal amount of redblood cells; Cooke and Smith, 1987), and a short gut.The total amount of mesoderm is the same as that ofnormal embryos; but dorsal mesoderm is missing(Cooke & Smith, 1987). Dorsal development clearlydepends on rotation whereas ventral development doesnot.
With intermediate amounts of rotation, the eggdevelops certain dorsal structures, in an interestinganteroposterior deletion series that depends on theamount of rotation. The more rotation, the more

42 J. Gerhart and others
Fig. 5. Schematic diagram of the anteroposterior order ofdeletion of dorsal structures from the embryonic body axisas cortical rotation is reduced. Although the embryonicbody axis can be truncated at any level, grades (oranatomical types) of embryos are defined to establish ascoring system, for the rough quantification of embryonicdefects. This scale originated with Malacinski etal. (1977),was modified by Scharf & Gerhart (1980), and expanded toinclude excessively dorsalized embryos by Kao & Elinson(1988). Note that the scores described here are notinterchangeable with those described in the earlier papers.A grade 5 embryo has all normal axial structures, whereas agrade 0 embryo lacks all dorsal structures, as well as someventral ones such as the heart, gills, and probably someparts of the digestive system. However, grade 0 embryosstill develop red blood cells and coelomic mesoderm(organized in two chambers), and are covered with ciliatedepidermis (see Fig. 4). They digest their vegetal yolk inwhat may be a simple gut. Ventral structures of the normalbelly region (i.e. excluding gills, heart, and anus) developalong all meridians of the egg, rather than just on theventral-most. This is the "limit form" obtained withoutcortical rotation, or without vegetal dorsalizing cells (a'Nieuwkoop center'), or without a Spemann organizer.Intermediate grades have the structures of grade 0, plusmore and more of the dorsal structures, as shown by thecontour lines on the diagram. Embryonic structures arereduced and lost conceitedly in the anteroposterior anddorsoventral directions. The pattern of progressive deletioncan be more easily rationalized by examining the fate mapof the early gastrula, than by examining the final larva.
anterior is the truncation level beyond which dorsalelements are missing from the body axis. Thus, whilethe direction of rotation defines the position of thedorsal midline, the extent of rotation defines theanterior extent of dorsal development. Fig. 5 shows theprogression by which body parts are gained and lost.
While all eggs require rotation, they differ in thequantity required; some need at least 25° and othersonly 12° for development of the complete body axis(Fig. 6A). Normally eggs accomplish about 30° ofrotation, an amount sufficient for the normal develop-ment of even the most insensitive members of thepopulation. Presumably the difference reflects a vari-able effectiveness of rotation on its target, a componentinvolved in a subsequent process of the series leadingtoward dorsal development.
Rotation without microtubules still allows dorsaldevelopmentWhen eggs are treated with anti-microtubule agents sothat they have no tracks along which to effect rotation,they can nonetheless be forced to rotate artificially by
tipping them out of gravitational equilibrium, whileimmobilizing the egg surface. The vegetally weightedcore then slips relative to the cortex, driven by theartificial motive force of gravity. Tipped eggs, it turnsout, are well rescued for dorsal development (Scharfand Gerhart, 1980), and the amount of forced displace-ment correlates with the amount of rescue. The anteriorend of the body axis is rescued only in eggs experiencingthe greatest displacements (Vincent & Gerhart, 1987).During the period of forced displacement in these eggs,MTs cannot be detected in the vegetal periphery.Rotation, driven by artificial means, seems sufficient toinitiate dorsal development. Thus, we think that innormal eggs the parallel MT array functions only astracks for cortical rotation in the first cell cycle, and thatrotation itself, not the MTs, causes a dorsalizing changein the egg periphery.
What and where is the immediate target of rotation?When rotation stops at time 0-85 of the first cell cycle,what cytoplasmic alteration persists, with consequencesfor later steps of dorsal development? Unfortunatelywe don't know, but we have considered two ratherdifferent possibilities: either a change of the contactsbetween the cortex and core of the egg, or an alignmentand polarization of materials in the shear zone betweenthe cortex and core.
Regarding the first, the animal core cytoplasm, forexample, comes into contact with vegetal cortex on theprospective dorsal side during rotation. This kind ofcontact is not formed on the prospective ventral side.However, this contact may not be important sincenormal, and not excessively dorsalized, development isstill obtained from eggs in which the core has beencompletely inverted at time 0-4 in the first cell cycle,bringing animal core cytoplasm into total contact withvegetal cortex (S. Black, unpublished). These eggsgastrulate normally from the hemisphere containing thevegetal yolk mass and animal cortex. The core alonemay determine the animal-vegetal differences of devel-opment, and rotation-dependent modifications of thecore may determine dorsoventral differences.
Regarding the second possibility, the zone betweenthe core and cortex must experience high shear as thetwo rigid units move past one another for 350 ^m whileseparated by only 2-5 /.im. Shear could in principle leadto the alignment and deformation of materials withinthe zone. High voltage electron microscopy shows theshear zone to contain large numbers of membrane-bounded vesicles (Rowning, 1989). Perhaps these be-come elongated and aligned during rotation. If suchvesicles are important in influencing the direction oftubule polymerization and hence the direction of ro-tation, as described earlier, shear forces would have topolarize them. Furtherrrfore, aligned vesicles wouldhave to retain different polarities on opposite sides ofthe egg if they are to promote dorsal development onjust one side. To our knowledge, this proposal ofpolarized vesicles gains as yet no support from studiesof other types of cells. Nonetheless, shear-dependent
Rotation of the Xenopus egg 43
-3a
01
33O
oQ
-OO O—O0]0-0«0«>-
OO O O O-CDOOOO O
O O OO (TOO
OO aXDQQOOOO OO
O OQQ0"OO O
t
0 ©I>-O-O-00-O-0 10 20
Rotation (degrees)30
100
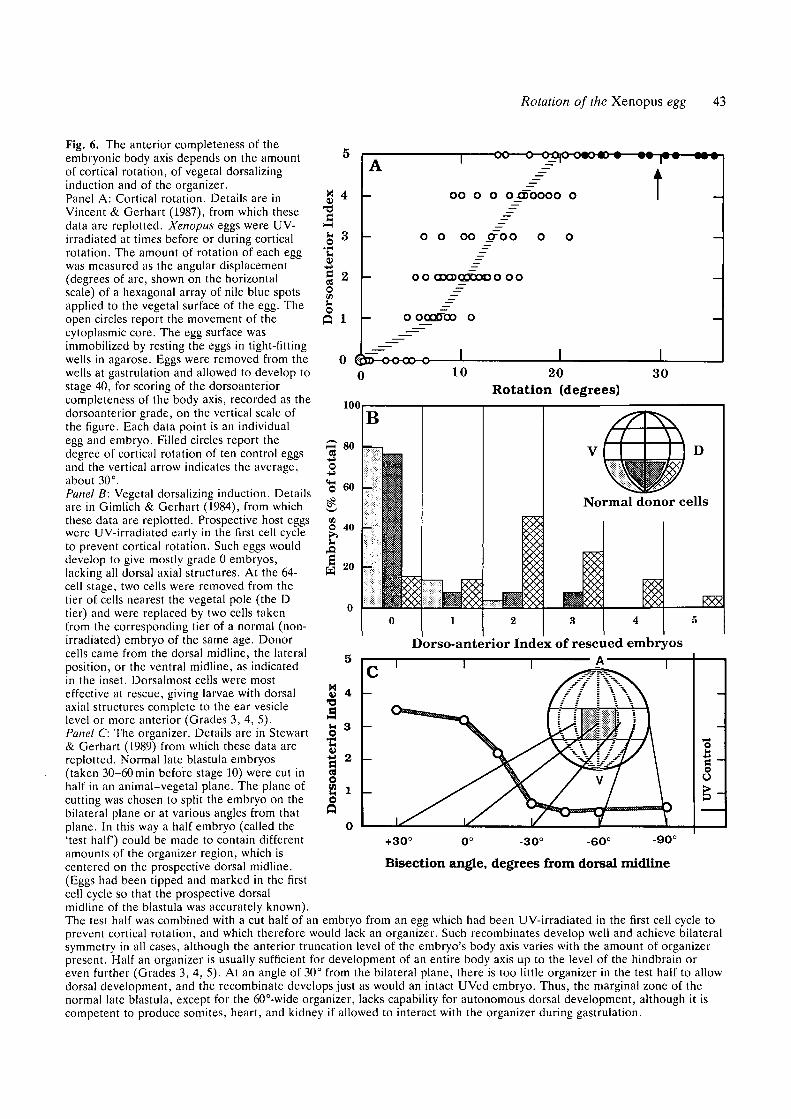
Fig. 6. The anterior completeness of theembryonic body axis depends on the amountof cortical rotation, of vegetal dorsalizinginduction and of the organizer.Panel A: Cortical rotation. Details are inVincent & Gerhart (1987), from which thesedata are replotted. Xenopus eggs were UV-irradiated at times before or during corticalrotation. The amount of rotation of each eggwas measured as the angular displacement(degrees of arc, shown on the horizontalscale) of a hexagonal array of nile blue spotsapplied to the vegetal surface of the egg. Theopen circles report the movement of thecytoplasmic core. The egg surface wasimmobilized by resting the eggs in tight-fittingwells in agarose. Eggs were removed from thewells at gastrulation and allowed to develop tostage 40, for scoring of the dorsoanteriorcompleteness of the body axis, recorded as thedorsoanterior grade, on the vertical scale ofthe figure. Each data point is an individualegg and embryo. Filled circles report thedegree of cortical rotation of ten control eggsand the vertical arrow indicates the average,about 30°.
Panel B: Vegetal dorsalizing induction. Detailsare in Gimlich & Gerhart (1984), from whichthese data are replotted. Prospective host eggswere UV-irradiated early in the first cell cycleto prevent cortical rotation. Such eggs woulddevelop to give mostly grade 0 embryos,lacking all dorsal axial structures. At the 64-cell stage, two cells were removed from thetier of cells nearest the vegetal pole (the Dtier) and were replaced by two cells takenfrom the corresponding tier of a normal (non-irradiated) embryo of the same age. Donorcells came from the dorsal midline, the lateralposition, or the ventral midline, as indicatedin the inset. Dorsalmost cells were mosteffective at rescue, giving larvae with dorsalaxial structures complete to the ear vesiclelevel or more anterior (Grades 3, 4, 5).Panel C: The organizer. Details are in Stewart& Gerhart (1989) from which these data arereplotted. Normal late blastula embryos(taken 30-60 min before stage 10) were cut inhalf in an animal-vegetal plane. The plane ofcutting was chosen to split the embryo on thebilateral plane or at various angles from thatplane. In this way a half embryo (called the'test half) could be made to contain differentamounts of the organizer region, which iscentered on the prospective dorsal midline.(Eggs had been tipped and marked in the firstcell cycle so that the prospective dorsalmidline of the blastula was accurately known).
The test half was combined with a cut half of an embryo from an egg which had been UV-irradiated in the first cell cycle toprevent cortical rotation, and which therefore would lack an organizer. Such recombinates develop well and achieve bilateralsymmetry in all cases, although the anterior truncation level of the embryo's body axis varies with the amount of organizerpresent. Half an organizer is usually sufficient for development of an entire body axis up to the level of the hindbrain oreven further (Grades 3, 4, 5). At an angle of 30° from the bilateral plane, there is too little organizer in the test half to allowdorsal development, and the recombinate develops just as would an intact UVed embryo. Thus, the marginal zone of thenormal late blastula, except for the 60°-wide organizer, lacks capability for autonomous dorsal development, although it iscompetent to produce somites, heart, and kidney if allowed to interact with the organizer during gastrulation.
1r•a 2
OQ
Dorso-anterior Index of rescued embryosAT
+30° -30° -60° -90°
Sioo
Bisection angle, degrees from dorsal midline

44 J. Gerhart and others
polarization is worth mentioning as a very differentpossibility from that of altered core-cortex contacts.
In addition to our ignorance of what is affected, wedon't know where the effect of rotation is first regis-tered, namely, in the AH or VH, or both, even thoughwe know that the effect must occur near the prospectivedorsal midline, probably in the shear zone between thecore and cortex. There are several indications that thevegetal hemisphere contains the target: Elinson &Pasceri (1989) have recently shown that UV-irradiatedoocytes give rise to eggs that form vegetal microtubulesand that rotate, but do not thereafter develop dorsalstructures; these are presumably damaged in materialsthat rotation would normally activate. The target ofirradiation is in the vegetal hemisphere of the oocyte.By the 16- or 32-cell stage, the vegetal hemisphere iscertainly specialized for promoting dorsal develop-ment; it contains a dorsalizing inductive quadrant(discussed later) centered on the midline of greatestrotational displacement.
However, Cardellini (1988) has presented evidencethat the animal hemisphere is initially specialized nearthe prospective dorsal midline, presumably by rotation,and that it passes its effect to the vegetal hemisphere atthe 8- to 16-cell stage. Kageura & Yamana (1984) havealso found cases where dorsal animal blastomeres, iftransplanted at the 8-cell stage, can initiate dorsaldevelopment independent of the state of vegetal blasto-meres. (We have unfortunately not been able to repro-duce these results; in our hands, the animal hemispherecarries no dominant dorsalizing bias at the 8-cell stage,and the vegetal hemisphere is already fixed in its controlof dorsal development.) Furthermore, London et al.(1988) report that AH cells at the 8-cell cleavage stagediffer on the prospective dorsal and ventral sides:isolated and cultured AH prospective ventral cells canautonomously express at a later time an antigen charac-teristic of normal embryonic epithelium, whereas pro-spective dorsal AH cells cannot. Non-expression ischaracteristic of the neural plate region of the intactembryo, a region that normally develops from the AHprospective dorsal cells. However, the isolated AHprospective dorsal cells do not form a neural plateautonomously. Thus, there is evidence that the animalhemisphere is regionally modified at least to someextent by rotation.
At this time, the most inclusive interpretation wouldbe that the entire dorsal meridian of the egg, in bothhemispheres, is modified by rotation, and the particularlocal developmental expression of this modification willdiffer depending on the animal, equatorial, or vegetallevel. Finally, rotation may only initiate a first transientcytoplasmic change, soon leading to other longer-lasting modifications. In this connection, it is observ-able that pigmentation differences continue to developin the animal hemisphere until at least the 4-cell stage,that deep cytoplasmic materials of the AH circulate inrelation to the direction of rotation, and that corticalmaterials ingress into the egg interior along new cleav-age furrows which differ in detail on the prospectivedorsal and ventral sides (Phillips, 1985; M. Danilchik
and J. Degnen, unpublished). It remains to be seen howmany of these changes are indispensible for dorsaldevelopment, and how many are epiphenomena ofrotation.
Excessive dorsoanterior developmentWhen eggs are centrifuged twice in opposite directions,to achieve two forced displacements, they frequentlydevelop as twins, with dorsal midlines at positionsrelated to two force vectors (Black & Gerhart, 1986). AlOmin interval must separate the two centrifugations inorder for both displacements to have effect; withshorter intervals only the second displacement initiatesdorsal development. The existence of these twins showsthat the egg has the capacity to initiate dorsal develop-ment in excess of the normal amount.
The egg can accomplish still more dorsoanteriordevelopment, at the expense of ventroposterior devel-opment, if treated with D2O early in the first cell cycle(Scharf et al. 1989). In the extreme, such embryospossess a large heart, a core of unextended notochord,and circumferential bands of eye pigment and cementgland. They lack somites and red blood cells. Theyretain the cylindrical symmetry of the unfertilized egg,with dorsal development initiated on all meridians. Inexaggerating dorsoanterior development, they definethe opposite end of the anatomical spectrum fromventralized embryos (Fig. 4, left). Partially affectedeggs develop as embryos with enlarged heads andtrunks, but lacking tails, or as enlarged heads lackingtrunks and tails (Scharf et al. 1989). As discussed later,similar embryos arise from blastulae treated with lith-ium ion at the 64- to 128-cell stage (Kao & Elinson,1988). Cooke and Smith (1988) estimate that hyperdor-salized embryos contain the same total amount ofmesoderm as normal embryos, but dorsal mesodermhas been increased at the expense of its ventral counter-part.
These hyperdorsal forms arise only if eggs are treatedwith D2O before cortical rotation. Once rotation be-gins, D2O has no effect. D2O causes a precociouspolymerization of MTs in the vegetal periphery, in adense random array that persists into the rotationperiod. The D2O-stabilized MTs are crucial since anysubsequent treatment that eliminates them also elimin-ates hyperdorsal development. The action of D2Oremains a puzzle to us. Even though unified corticalrotation is strongly inhibited, probably because therandom MT array cannot acquire unidirectionality,there may occur multiple local displacements of cortexand core materials. The egg behaves as if it had rotatedin many directions, and among the mildly affectedcases, twinning occurs frequently.
The existence of hyperdorsal embryos shows that theegg has maternal resources for more extensive dorsaldevelopment than it normally uses. Presumably thispotential resides at all meridians of the egg's circumfer-ence, in a latent state, just as does the potential forventral development. Unified cortical rotation normallyactivates the use of this potential in just one region,while leaving the remainder of the circumference for

Rotation of the Xenopus egg 45
ventral development, thereby establishing the pro-portions of the normal embryonic pattern.
II. The quantitative dependence of later events oncortical rotation
Why does the quantity of cortical rotation in the first cellcycle delimit the anterior completeness of the embry-onic body axis? To consider this relationship, we willsketch three steps of later development in which thesame truncation series of embryos can be generated byexperimental interference. We propose that the amountof cortical rotation determines: (1) the number ofvegetal dorsalizing cells which in turn, by way ofinduction in the blastula period, determine (2) thenumber of the cells of the Spemann organizer regionwhich in turn, by way of inductive interactions in thegastrula period, determine (3) the extent of morpho-genesis and patterning during gastrulation.
Vegetal dorsalizing inductionAs Nieuwkoop discovered (1973), cells of the vegetalhemisphere of the midblastula embryo (4000-cell stage)induce neighboring cells of the animal hemisphere tobehave as marginal zone cells at gastrulation and todifferentiate ultimately as many types of mesoderm andas archenteron roof endoderm. The dorsalmost quad-rant of the vegetal hemisphere differs from othervegetal regions: it induces the organizer quadrant of themarginal zone, which initiates much of the morphogen-esis of gastrulation, and which usually forms the noto-chord, a central mesodermal tissue of the embryonicdorsal midline. Dorsal development beyond the blas-tula stage depends on the presence of these specialvegetal cells.
We extended these studies to earlier stages in anattempt to connect cortical rotation to the founding ofthis specialized vegetal quadrant. At the 32- to 64-cellblastula stage, two vegetal cells can be removed fromthe prospective dorsal midline, as predicted by thedirection of cortical rotation, and these cells can betransplanted into another blastula of the same age, forexample, replacing two vegetal cells of a blastuladerived from a UV-irradiated egg, one that would on itsown develop only ventral structures. From thesegrafted embryos arise well-rescued tadpoles, withnearly complete body axes containing normal dorsalstructures of the head, trunk and tail. When graft cellsare preloaded with a fluorescent lineage tracer, we seethat progeny cells of the graft do not populate therescued embryo's dorsal structures, but only the ventralyolk mass. Graft cells induce neighboring equatorialcells of the host to develop the entire body axis.Gastrulation begins near the graft, and the dorsalmidline of the rescued embryo is centered on themeridian of grafting (Gimlich & Gerhart, 1964; Gim-lich, 1986).
Rescuing graft cells can only be obtained frompositions close to the prospective dorsal midline of thedonor, within 45° or less of the midline; cells from
lateral or opposite vegetal positions do not rescue(Figure 6B). The effective donor region includes thetwo vegetal tiers of cells present at the 32- to 64-cellstage, that is, from the equator to the vegetal pole. Theinductive strength of cells of the two tiers differs fromembryo to embryo, perhaps because cleavage furrowsare variably related to the locus of specialized cyto-plasm (Ginilich, 1986). In the absence of other names,we call this the 'vegetal dorsalizing region', or the'organizer inducing region', or the 'Nieuwkoop center'.
The normal embryo requires cortical rotation for thisregion's formation, and the region arises on the merid-ian of greatest cytoplasmic displacement, on the sidewhere the core moves vegetally vis a vis the cortex.While any part of the VH is latently capable of forminga dorsalizing center, only one region actually does innormal development: that part affected by the single,unidirectional rotation. As discussed before, rotationmay directly activate cytoplasm of the vegetal dorsal-izing center, presumably bordering the shear zone or atnew contact regions between the cortex and core, or itmay activate materials of the animal hemisphere, whichsecondarily activate the vegetal center, as suggested byCardellini (1988). If the latter, the steps of transfer areprobably complete by the 8- to 16-cell stage.
The rest of the vegetal hemisphere, unaffected byrotation, also engages in an induction of the marginalzone, but this does not lead to organizer formation or todifferentiation of dorsal mesoderm. It leads to theformation of the 'indifferent' or 'ventral' portion of themarginal zone. This type of ventralizing induction doesnot depend on rotation, for it occurs in embryosprevented from rotation by MT depolymerizing agents.As discussed elsewhere in this Volume (see chapters bySlack et al.; Smith et al.; Yisraeli et al.), the VH mayexert its inductive effects by secreting growth-factor-like proteins. By these accounts, cortical rotation mightbe expected to enable cells of the vegetal dorsalizingregion to specifically release a TGF-/3 homolog,whereas all vegetal cells even without rotation wouldrelease an FGF-like material. At this time we have noidea how cortical rotation might alter the secretoryproperties of cells derived from one region. Since TGF-/S and FGF homologs are present as proteins even in theunfertilized egg (Kimelman et al. 1988; Slack & Isaacs,1989; Dale et al. 1989; Tannahill & Melton, 1989), theirrelease could involve the region-specific activation of astep of processing or externalization, rather than oftranscription or translation.
As an aside, lithium ion deserves special mentionbecause it obviates the need for cortical rotation and fora Nieuwkoop center. Eggs that have failed to rotate canbe rescued to form near-normal embryos by an injec-tion of lithium chloride into equatorial cells at the 16- to32-cell stage (tier C; Busa & Gimlich, 1989). Uniformexposure of such eggs to lithium solutions leads to thedevelopment of cylindrically symmetric hyperdorsa-lized forms resembling D2O embryos (Kao & Elinson,1988). Apparently lithium acts not by stimulating veg-etal cells to release organizer inducing (dorsalizing)factors, but by sensitizing animal hemisphere cells to

46 J. Gerhart and others
respond to the vegetal ventralizing inductors as if theywere dorsalizing inductors, i.e. to an FGF homolog as ifit were a TGF-/3 homolog (Nieuwkoop, 1970; Kao &Elinson, 1989; Slack etal. 1988; Cooke & Smith, 1988).Hence, the embryo circumvents the normal require-ments for cortical rotation and the Nieuwkoop center.Given this interesting response to lithium ion, it seemsplausible that the normal vegetal dorsalizing inductor,like lithium, just enhances the response of neighboringAH cells to the ubiquitous vegetal ventralizing induc-tor, and these responding cells become the organizer. Inthis regard, TGF-/J1 is known to synergize the FGF-dependent response by explanted AH cells to producedorsal mesoderm (Kimelman & Kirschner, 1987).
Rescue by cells of the Nieuwkoop center is oftenincompleteVegetal dorsalizing cells do not always promote com-plete rescue when grafted into UVed recipients. Whenthe body axis is incomplete, it is always truncated fromthe anterior end, and the same scoring scale can be usedas for embryos inhibited in cortical rotation (see Fig. 5and 6B). Grafts with one, two, or four cells from the Cand D tiers of the donor give increasingly frequent well-rescued embryos. Truncated embryos of the same typescan also be produced from normal blastulae by elimi-nating cells from the Nieuwkoop center of the 32-cellblastula (Gimlich, 1986). The elimination of more cellsleads to greater anterior defectiveness in the finalembryos.
These results, while not well quantified yet, implythat later dorsal development depends systematicallyon the amount of Nieuwkoop center present in theblastula stage embryo. Since experimental pertur-bations of vegetal induction produce the same finalbody patterns as do perturbations of rotation, wesuggest that the amount of rotation determines theamount of vegetal dorsalizing induction. With no ro-tation, there is no such induction. Intermediate rotationgives intermediate induction. Two opposite rotationslead to two opposite Nieuwkoop centers. And afterD2O treatment, most of the vegetal hemisphere be-comes a fully active Nieuwkoop center. Presumably theamount of rotation determines the amount of secretionof a specific inductor protein from cells of the Nieuw-koop center. Despite this quantitative correspondence,however, we can't yet discern why the anterior end ofthe body axis is the end missing in deficient embryos.
The amount of organizerBy the end of the blastula period, the marginal zone issufficiently patterned for the initiation of normal gastru-lation. In Xenopus, the marginal zone's organizerregion, which arises directly above the vegetal dorsal-izing region, is approximately 60° wide; whereas theindifferent part of the marginal zone is 300° wide,arising above the non-Nieuwkoop part of the VH(Smith & Slack, 1983; Dale & Slack, 1987; Stewart &Gerhart, 1989).
The organizer can be reduced in size by surgery at thelate blastula stage. This allows us to test the effects on
development of subnormal amounts of organizer. Asone way to do this, the late blastula (stage 9, approx.15 000 cells, 30-60min before blastopore formation) iscut vertically on the bilateral plane (i.e. in the animal-vegetal direction through the organizer region), and theleft or right half is recombined with half an embryo ofequal age, cut from an egg which had been UV-irradiated in the first cell cycle to prevent its rotationand to preclude its formation of a Nieuwkoop center.This latter half, lacking an organizer, would on its ownbe incapable of any dorsal development. Although suchrecombinates have each just half an organizer, theydevelop remarkably well and produce near-perfectbilateral embryos, in many cases with near-completebody axes containing normal dorsal structures of thehead, trunk and tail. One side of the body is contributedby the would-be ventralized half, as shown by lineagetracing. Still, some recombinates give bilateral embryoswith incomplete body axes (Fig. 6C), and when dorsalstructures are missing, the body axis is truncated fromthe anterior end, just as we have seen before, and thesame scoring system can be used.
In the same kind of operation, the organizer can beintentionally cut off center (e.g. 15° from the midline),to include a still smaller amount of organizer in therescuing normal half embryo used to make a recombi-nate with a UVed half. When this is done, the recombi-nate tends to have still fewer anterior structures(Fig. 6C), though always in a bilateral arrangement towhich the UVed half contributes one side. The relation-ship seems monotonic: the less organizer, the lessanterior development.
Finally, when the cut departs 30° or more from thebilateral plane, the piece lacking the dorsal meridianhas no capacity for rescue; the recombinate develops asa cylindrically symmetric ventralized embryo. The or-ganizer, at least in its capacity to initiate dorsal axialdevelopment, apparently extends just 30° to either sideof the dorsal meridian, i.e. it is 60° wide. The remainingmarginal zone material of a normal late blastula re-sembles the marginal zone of a ventralized embryo,with no autonomy for dorsal development, even thoughthis part is destined in normal development to producethe bulk of the mesodermal tissues of the embryo,namely, the somites, kidney and heart, in fact, almostall mesoderm except the notochord and some headmesenchyme. This indifferent region must interact withorganizer cells during gastrulation to generate its sub-stantial part of the normal embryo's axial organization.The major share of definitive embryonic organization,along both the dorsoventral and anteroposterior axes,must be established during and/or after gastrulation, asshown by the performance of UVed halves brought intocontact with an organizer, and by the performance ofnormal indifferent marginal zone regions deprived of anorganizer.
We can now extend our causal chain of quantitativerelationships by one more step since progressive re-ductions in the amount of organizer material in the lateblastula result in embryonic body axes progressivelytruncated from the anterior end. We suggest that the

Rotation of the Xenopus egg 47
amount of rotation defines the amount of Nieuwkoopcenter, which defines the amount of organizer. With norotation, there is no vegetal dorsalizing induction andno organizer. Partial rotation leads to partial inductionand a reduced organizer. However, despite our causalchain, we haven't answered why, when the organizer isreduced in size, the anterior end of the embryonic axisis missing.
Definitive axis formationGastrulation transforms egg organization into embry-onic organization. In this process, cells of the marginalzone generate the dorsoventral and anteroposteriordimensions of the body axis as the result of theirmigration, repacking and manifold interactions (Keller& Danilchik, 1988; Wilson & Keller, 1989). In undis-turbed development, the first cells to gastrulate arethose of the organizer region, which comprises the'dorsal lip' of the blastopore. Cells of the organizerregion are special in their ability, as a population, toconverge toward the prospective dorsal midline and toextend in the anteroposterior direction. They induceneighboring cells of the indifferent marginal zone toundertake similar movements and they induce ectoder-mal cells to undertake neurulation. Eventually theycontribute to dorsoanterior elements of the body axis,such as pharyngeal endoderm, head mesenchyme,archenteron roof, notoplate and notochord (see Ger-hart & Keller, 1986, for a review).
Because organizer cells are so special, we initiallyassumed that they are fully determined at the start ofgastrulation, not only for their gastrulation movementsand induction abilities, but also for their ultimate celltype differentiations. Fate maps show that the firstinvaginated cells of the organizer tend to populate themost anterior parts of the body, and later ones contrib-ute more posteriorally. The organizer might havewithin it a cryptic anteroposterior and a dorsoventralpattern of cellular differences, a pattern that wouldpropagate to the rest of the embryo during morphogen-esis. By this view, the organizer would itself be alreadyfully organized at the start of gastrulation, a miniaturemosaic of the future embryo's axial organization. Inmany respects, this interpretation could explain theseries of systematically truncated embryos produced byour experimental interventions just described. Forexample, reduced rotation and reduced vegetal induc-tion of the organizer might just lead to the formation ofan organizer lacking the most anterior-dorsal parts ofits cryptic pattern, if we assume that qualitative differ-ences within the organizer originate from quantitativedifferences set up by vegetal induction. In such manipu-lated embryos, gastrulation does in fact start later, andcells accomplish less convergence and extension. Thus,we thought it plausible that the diminished organizerjust lacked cells for organizing the most anterior dorsalparts of the body axis. Of course, it must be admittedthat the results on reducing the size of the organizer bysurgery at the start of gastrulation would not readily fitthis interpretation.
The situation is considerably more subtle, though, as
shown by the results of experiments in which embryosare allowed to develop normally up to and into thegastrula stage (so as to have a well-organized organ-izer), and then at various times are prevented fromcontinuing gastrulation. Inhibitors of Xenopus gastru-lation are not easy to find: there is no significant effectfrom RGD peptides, p-nitrophenyl-xylosides, or/3-aminopropionitrile, agents that arrest gastrulation insea urchins (Wessel & McClay, 1987; Lane & Solursh,1988) and Pleurodeles (Boucaut et al. 1984). However,polysulfonated compounds such trypan blue and sura-min (germanin) are very effective (Waddington &Perry, 1956). These are powerful teratogens in mam-mals, acting at the primitive streak stage; they anima-lize sea urchins by arresting gastrulation (Beck &Lloyd, 1966). Suramin disrupts the binding of proteinligands to cell surface receptors, the examples includinggrowth factors (FGF and TGF-/3, Coffey et al. 1987),vitellogenin, and LDL (Peacock etal. 1988). High dosesof trypan blue or suramin (20 HM in the blastocoel) stopconvergent extension movements in vivo, though allow-ing the blastopore to close. The agents effectively arrestthese same movements of cells in explanted prep-arations of the organizer (Danilchik et al. 1989).
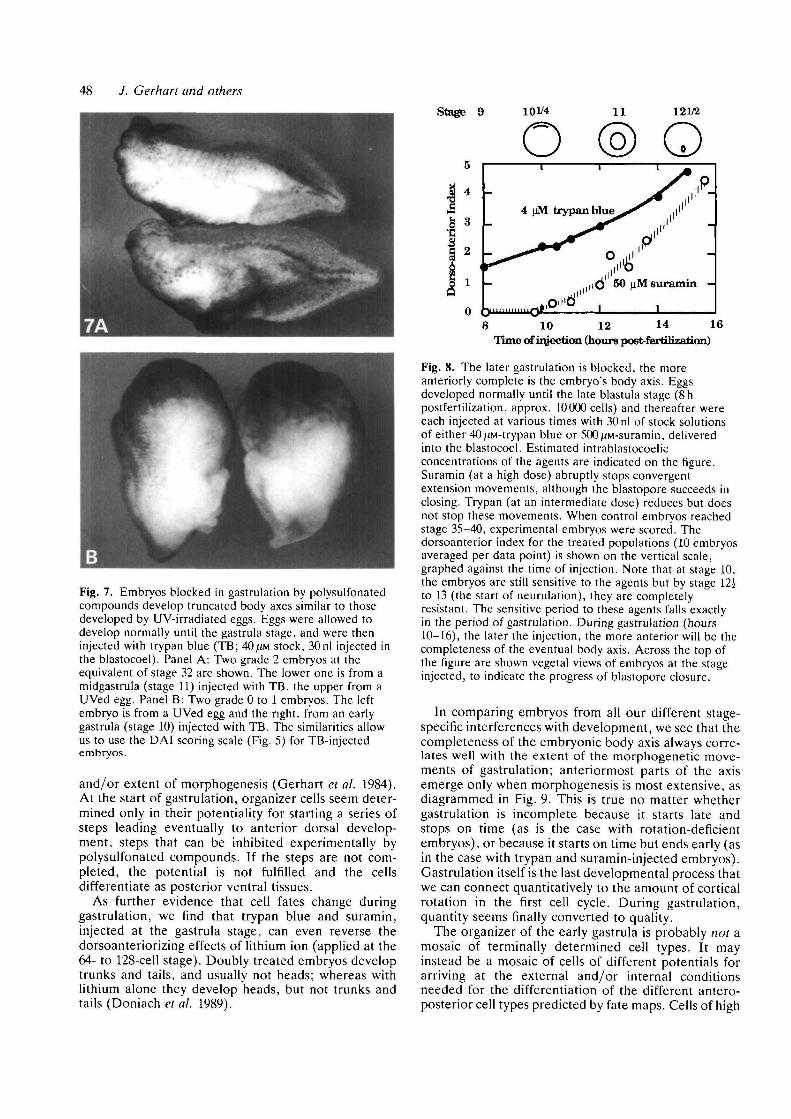
The clearest demonstration of their systematic effecton axis formation comes when these agents are injectedinto the blastocoel at different times during the 6hperiod of gastrulation (stages 10 to 13): embryos there-after develop body axes truncated at different antero-posterior levels depending on the time of injection(Figure 7 and 8). If injected when the dorsal lip is firstinvaginating, the resulting embryo develops no head ortrunk, and sometimes no tail as well (Fig. 8B). Somitesand notochord are absent; the embryo resembles thecylindrically symmetric ventralized forms seen whenrotation fails or when the Nieuwkoop center or organ-izer is ablated. If the agents are injected midway ingastrulation, embryos develop without heads, but withtrunks and tails (Fig. 8A). Finally, if injected at the endof gastrulation, the embryo is completely unaffected,even though the agents are present throughout theperiods of neurulation and cell differentiation. Thus,the polysulfonated agents are not indiscriminatelytoxic, but act in a stage-specific way during gastrulation,inhibiting some process perhaps unique to gastrulation,or most exaggerated or critical at that time.
Gastrulation is the latest stage at which we have beenable to produce the familiar series of truncated anat-omies for which we can use the same dorsoanteriorindex scoring system (Fig. 5). In preliminary exper-iments with injected fluorescent lineage tracers, we findthat the leading cells of the dorsal marginal zone (theorganizer region) differentiate into whatever parts ofthe body axis are most anterior in the truncatedembryo. Also, in preliminary experiments, the extentof gastrulation (the length of the archenteron) corre-lates with the completeness of the body axis. In contra-diction to the expectations of the miniature mosaicmodel, organizer cells do not seem to have a fixed fateat the start of gastrulation but seem to undergo aprogressive dorsoanteriorization in relation to the time
48 J. Gerhart and others
Stage 9 101/4
Fig. 7. Embryos blocked in gastrulation by polysulfonatedcompounds develop truncated body axes similar to thosedeveloped by UV-irradiated eggs. Eggs were allowed todevelop normally until the gastrula stage, and were theninjected with trypan blue (TB; 40/.m stock, 30nl injected inthe blastocoel). Panel A: Two grade 2 embryos at theequivalent of stage 32 are shown. The lower one is from amidgastrula (stage 11) injected with TB, the upper from aUVed egg. Panel B: Two grade 0 to 1 embryos. The leftembryo is from a UVed egg and the right, from an earlygastrula (stage 10) injected with TB. The similarities allowus to use the DAI scoring scale (Fig. 5) for TB-injectedembryos.
and/or extent of morphogenesis (Gerhart et al. 1984).At the start of gastrulation, organizer cells seem deter-mined only in their potentiality for starting a series ofsteps leading eventually to anterior dorsal develop-ment, steps that can be inhibited experimentally bypolysulfonated compounds. If the steps are not com-pleted, the potential is not fulfilled and the cellsdifferentiate as posterior ventral tissues.
As further evidence that cell fates change duringgastrulation, we find that trypan blue and suramin,injected at the gastrula stage, can even reverse thedorsoanteriorizing effects of lithium ion (applied at the64- to 128-cell stage). Doubly treated embryos developtrunks and tails, and usually not heads; whereas withlithium alone they develop heads, but not trunks andtails (Doniach et al. 1989).
8 10 12 14 16Time of injection (hours poetfertQization)
Fig. 8. The later gastrulation is blocked, the moreanteriorly complete is the embryo's body axis. Eggsdeveloped normally until the late blastula stage (8hpostfertilization, approx. 10000 cells) and thereafter wereeach injected at various times with 30 nl of stock solutionsof either 40;iM-trypan blue or 500/iM-suramin, deliveredinto the blastocoel. Estimated intrablastocoelicconcentrations of the agents are indicated on the figure.Suramin (at a high dose) abruptly stops convergentextension movements, although the blastopore succeeds inclosing. Trypan (at an intermediate dose) reduces but doesnot stop these movements. When control embryos reachedstage 35-40, experimental embryos were scored. Thedorsoanterior index for the treated populations (10 embryosaveraged per data point) is shown on the vertical scale,gTaphed against the time of injection. Note that at stage 10,the embryos are still sensitive to the agents but by stage 12ito 13 (the start of neurulation), they are completelyresistant. The sensitive period to these agents falls exactlyin the period of gastrulation. During gastrulation (hours10-16), the later the injection, the more anterior will be thecompleteness of the eventual body axis. Across the top ofthe figure are shown vegetal views of embryos at the stageinjected, to indicate the progress of blastopore closure.
In comparing embryos from all our different stage-specific interferences with development, we see that thecompleteness of the embryonic body axis always corre-lates well with the extent of the morphogenetic move-ments of gastrulation; anteriormost parts of the axisemerge only when morphogenesis is most extensive, asdiagrammed in Fig. 9. This is true no matter whethergastrulation is incomplete because it starts late andstops on time (as is the case with rotation-deficientembryos), or because it starts on time but ends early (asin the case with trypan and suramin-injected embryos).Gastrulation itself is the last developmental process thatwe can connect quantitatively to the amount of corticalrotation in the first cell cycle. During gastrulation,quantity seems finally converted to quality.
The organizer of the early gastrula is probably not amosaic of terminally determined cell types. It mayinstead be a mosaic of cells of different potentials forarriving at the external and/or internal conditionsneeded for the differentiation of the different antero-posterior cell types predicted by fate maps. Cells of high

Rotation of the Xenopus egg 49
Stage 1X5
Fig. 9. Progressive anteriorization of the embryonic bodyaxis during Xenopus gastrulation. In all our experimentallyperturbed embryos (excluding D2O effects), the truncationlevel of the final body is related to the extent of gastrulationmovements, regardless of whether gastrulation startsnormally at stage 10 but is then ended at different times (bytrypan blue or suramin), or whether gastrulation starts later(due to reduced amounts of organizer in the marginal zone,due to inhibited rotation, or reduced numbers of vegetaldorsalizing cells, or surgical removal of parts of theorganizer), while ending at the same time (probably stage13).
The six panels are schematic cross-sections of embryos,showing different stages of the 6h period of gastrulation,from stage 10 (upper left) and to stage 12-5—13 (lowerright). The animal pole (A) is at the top of all figures. Theblastocoel is the space in the animal hemisphere. At thestart of gastrulation, some cells of the organizer part of themarginal zone (right subequatorial level) begin invaginatingfirst, followed in sequence by cells more distant laterallyand vertically. The '0' in the upper left section indicatesthat cells of the organizer, if their morphogenesis werestopped at that point, would differentiate ventral structures,like the parts of a grade 0 embryo (see Fig. 5). In the topcenter panel, leading cells of the organizer have movedfarther inward, followed by dorsal marginal zone cellswhich were initially farther from the blastopore. Theleading cells are now in a condition to differentiate asposterior dorsal structures, which they would do ifgastrulation were stopped at this point, while cells behindthem have just reached the '0' condition. These conditions,which may reflect internal or external (or both) states of thecells, change progressively with the time and/or distance ofmorphogenesis (see subsequent panels). Leading cells areunique in that they have the potential for progressingthrough the full range of conditions, finally reachingcondition '5' (lower right) sufficient for anterior dorsaldifferentiation. Cells farther back in the migration seriesmake less progress, because they start later and are lessactive. While cells of the marginal zone would be initiallygraded in their potential for progress toward anterior dorsaldevelopment, all would need to accomplish extensivemorphogenesis to fulfill this potential. Thus, inhibitors ofmorphogenesis always have a posteroventralizing effect onthe final pattern.
potential would initiate morphogenesis earlier andmore vigorously than those of low potential, the latternot having the option to progress very far in the seriesof conditions for dorsoanterior differentiation; theywould be limited to producing posterior parts of the
pattern, although even in this they would fail if morpho-genesis were inhibited.
Why is the extent of gastrulation related to the anteriorcompleteness of the axial pattern?This inquiry dates back to Spemann (1938) and Vogtwho considered the connection between dynamic deter-mination (a cell's morphogenetic behavior at gastru-lation) and its material determination (its final cell typedifferentiation). We don't know the answer, but haveconsidered two possibilities, for each of which we willgive an example:
(1) Morphogenesis does not itself accomplish pat-terning but creates conditions upon which a separatepatterning process operates. This possibility is inherentin the double gradient idea of Saxen and Toivonen(1961). All cells would have an intrinsic capacity foranterior differentiation, but many are suppressed by aposteriorizing (or caudalizing or tail-inducing) centerlocated in those regions of the marginal zone gastrulat-ing last. Suppression, and therefore fates, would begraded with distance from the center: the closest cellsbecoming tail, and more distant ones, trunk. Only theearliest, farthest advancing marginal zone cells and themost distant neurectoderm would escape the effects ofthe center and would become anterior. According tothis view, if we inhibit gastrulation movements by anymeans, we should expect to prevent cells from escapingthe posteriorizing center, i.e. from acquiring the exter-nal conditions needed for their anterior differentiation.The head end of the body axis would be deleted and thecompleteness of the axial pattern would be quantitat-ively related to morphogenesis, just as we find.
(2) Morphogenesis and axial patterning may dependjointly on yet another agency such as intercellularsignalling. This possibility resembles ones for cell aggre-gation and slug migration in Dictyostelium (see chapterby Johnson et al. this Volume). At the start of gastru-lation, organizer cells might be unique in their highintensity of intercellular signalling, due to prior stimu-lation from the Nieuwkoop center. Such a cell mighthave three responses to signals: to move, to emit moresignal, and to keep a running sum of the amount ofreceived signal, a sum that would determine the cell'santeroposterior-dorsoventral fate. If the response dur-ation were short, repeated signal reception would beneeded by cells to keep signalling and responding. Cellfate would change during gastrulation in concert withmovement, both depending on continued intercellularsignalling. The earliest invaginating cells would havethe greatest potential for movement and anterior fatessince they could maintain the longest period of signal-ling. Non-organizer cells of the marginal zone mightreceive enough signals from organizer cells to beginsignalling, moving, and changing fates. Signalling couldpropagate through the marginal zone and neurecto-derm. By this view, polysulfonated inhibitors of gastru-lation would be inhibitors of signalling; they wouldterminate both morphogenesis and progress towardsanterior fates since these jointly depend on continuedsignalling. Parenthetically, another agent that affects

50 J. Gerhart and others
signal transduction, namely lithium ion, also reducesmorphogenetic movements and gives headless em-bryos, if applied in the gastrula stage (Hall, 1942; J.Slack, personal communication; an effect not to beconfused with hyperdorsalization by treatment of theblastula). By this view, reduced rotation and vegetalinduction would lead to a diminished organizer, onewith a lowered level of signalling, and morphogenesisand anterior patterning would consequently diminish inconcert.
These rather different interpretations have equalplausibility at present. Both confer to gastrulation, as aprocess, an inherent dynamic organization by whichanterior dorsal fates arise in relation to the increasingquantity of morphogenesis, and both give to the earlysteps of development, such as cortical rotation andvegetal induction, the significance of setting the initialconditions of gastrulation, conditions that may limit itsoperation.
This work was supported by USPHS grant GM 19363 andNSF grant DCB-8517548.
References
ANCEL, P. & VINTEMBERGER, P. (1948). Recherches sur ledeterminisme de la symetrie bilaterale dans l'oeuf desAmphibiens. Bull Biol. FT. Belg. 31 (Suppl.), 1-182.
BECK, F. & LLOYD. J. B. (1966). The teratogenic effect of azo dyes.Adv. Teratol. 1, 133-191.
BLACK. S. D. & GERHART, J. C. (1985). Experimental control ofthe site of embryonic axis formation in Xenopus laevis eggscentrifuged before first cleavage Devi Biol. 108, 310-324.
BLACK, S. D. & GERHART, J. C. (1986). High frequency twinning ofXenopus laevis embryos from eggs centrifuged bidirectionallybefore first cleavage. Devi Biol. 116, 228-240.
BLACK, S. D. & VINCENT, J.-P. (1988). The first cleavage plane andthe embryonic axis are determined by separate mechanisms inXenopus laevis. Devi Biol. 128, 65-71.
BOUCAUT, J. C , DARRIBERE, T., POOLE, T. J., AOYAMA, H.,
YAMADA, K. M & THIERY. J.-P. (1984). Biologically activesynthetic peptides as probes of embryonic development. J. CellBiol. 99, 1822-1830.
BUSA, W. B. & GIMLICH, R. L. (1989). Lithium-inducedteratogenesis in frog embryos prevented by apolyphosphoinositide cycle intermediate or a diacylglycerolanalog. Devi Biol. 132, 315-324.
CARDELLINI, P. (1988). Reversal of dorsoventral polarity inXenopus laevis embryos by 180° rotation of the animalmicromeres at the 8 cell stage. Devi Biol. 128, 428-434.
COFFEY, R. J.. LEOF. E. B., SHIPLEY, G. D. & MOSES. H. L.
(1987). Suramin inhibition of growth factor receptor binding andmitogenecity in AKR-2B cells J cell. Physiol. 132, 143-148.
COOKE. J. & SMITH. E. J. (1988). The restrictive effect of earlyexposure to lithium upon body pattern in Xenopus development.studies by quantitative anatomy and immunofluorescence.Development 102, 85-100.
COOKE, J. & SMITH, J. C. (1987). The midblastula cell cycletransition and the character of mesoderm in UV-inducednonaxial Xenopus development. Development 99, 197-210.
DALE, L., MATTHEWS, G., TABE, L. & COLMAN, A. (1989).
Developmental expression of the protein product of Vgl, alocalized maternal mRNA in the frog Xenopus laevis. EMBO J.8, 1057-1065.
DALE, L. & SLACK, J. M. W. (1987). Regional specification withinthe mesoderm of early embryos of Xenopus laevis. Development100, 279-295.
DANILCHIK. M., DONAICH. T. & GERHART, J. C. (1989). Patterning
of the embryonic body axis during Xenopus gastrulation:experimentally reduced morphogenesis leads to anteriorallytruncated embryos. In preparation.
DANILCHIK, M. V. & GERHART, J. C. (1987). Differentiation of theanimal-vegetal axis in Xenopus laevis oocytes. I. Polarizedtranslocation of platelets establishes the yolk gradient. Devi Biol.122, 101-112.
DONIACH, T., DANILCHIK, M. & GERHART, J. C. (1989). Patterning
of the embryonic body axis during Xenopus gastrulation. theprogressive anteriorization of cell fates. In preparation.
ELINSON, R. P. (1980). The amphibian egg cortex in fertilizationand development. Symp. Soc. Devi Biol. 38, 217-234.
EUNSON, R. P. & PASCERI, P. (1989). Two UV-sensitive targets indorsoanterior specification of frog embryos. Development 106,00-00.
ELINSON, R. P. & ROWNING, B. (1988). A transient array ofparallel microtubules in frog eggs: Potential tracks for acytoplasmic rotation that specifies the dorso-ventral axis. DeviBiol. 128, 185-197.
GERHART. J. & KELLER, R. (1986). Region-specific cell activities inamphibian gastrulation. A. Rev. Cell Biol. 1, 201-229.
GERHART, J , UBBELS, G., BLACK, S., HARA, K. & KIRSCHNF.R, M.
(1980). A reinvesrigation of the role of the grey crescent in axisformation in Xenopus laevis. Nature, Land. 292, 5H-516.
GERHART, J. C , VINCENT, J.-P., SCHARF, S. R., BLACK, S. D.,
GIMLICH, R. L. & DANILCHIK, M. (1984). Localization andinduction in early development of Xenopus Phil. Trans. R. Soc.Lond. B 307, 319-330.
GIMLICH, R. L. (1986). Acquisition of developmental autonomy inthe equatorial region of the Xenopus embryo. Devi Biol 115,340-352.
GIMLICH, R. L. & GERHART, J. C. (1964). Early cellular interactionspromote embryonic axis formation in Xenopus laevis. Devi Biol.104, 117-130.
GRANT, P. & WACASTER, J. F. (1972). The amphibian grey crescent-a site of developmental information? Devi Biol. 28, 454-471.
HALL, T. S. (1942). The mode of action of lithium salts inamphibian development. J. exp. Zool. 89, 1-30.
KAGEURA, H. & YAMANA, K. (1984). Pattern regulation in defectembryos of Xenopus laevis. Devi Biol. 101,410-415.
KAO, K. R. & ELINSON, R. P. (1988). The entire mesodermalmantle behaves as a Spemann's Organizer in dorsoanteriorenhanced Xenopus laevis embryos. Devi Biol. 127, 64-77.
KAO, K. R & ELINSON, R. P. (1989). Dorsalization of mesoderminduction by lithium. Devi Biol. (in press).
KELLER, R. E. & DANILCHIK, M. (1988). Regional expression,pattern and timing of convergence and extension duringgastrulation of Xenopus laevis. Development 103, 193-210.
KIMELMAN, D. & KIRSCHNER, M. (1987). Synergistic induction ofmesoderm by FGF and TGF-/3 and the identification of anmRNA coding for FGF in the early Xenopus embryo. Cell 51,869-877.
KIMELMAN, D., ABRAHAM, J. A., HAAPARANTA, T., PALISI, T. M &
KIRSCHNER. M. W. (1988) The presence of fibroblast growthfaster in the frog egg: its role as a natural mesoderm inducer.Science 242, 1053-1056.
LANE, M. C & SOLURSH, M. (1988). Dependence of sea urchinprimary mesenchyme cell migration on xyloside and sulfatesensitive cell surface associated components. Devi Biol. 127,78-87.
LONDON, C , AKERS, R. & PHILLIPS, C. R. (1988). Expression of
epi 1, an epidermal specific marker, in Xenopus laevis embryos isspecified prior to gastrulation. Devi Biol. 129, 380-389.
L0VTRUP, S. (1965). Morphogenesis in the amphibian embryo:Fertilization and blastula formation Wilhelm Rou.x' Arch.EntwMech. Org. 156, 204-248.
MALACINSKI, G. M., BROTHERS, A. J & CHUNG, H.-M. (1977).
Destruction of components of the neural induction system of theamphibian egg with ultraviolet irradiation. Devi Biol. 56, 24-39.
MANES, M. E. & BARBIERI, F. D. (1977). On the possibility ofsperm aster involvement in dorso-ventral polarization andpronuclear migration in the amphibian egg. J Embryo!, exp.Morph 40, 187-197.

Rotation of the Xenopus egg 51
NIEUWKOOP, P. D. (1970). The formation of mesoderm in urodelanamphibians. III. The vegetalizing action of the Li ion. WilhelmRoux1 Arch. EntwMech. Org. 166, 105-123.
NIEUWKOOP, P. D. (1973). The "organization center" of theamphibian embryo: its spatial organization and morphogeneticaction. Adv. Morphogen 10, 1-39.
PEACOCK, S. L., BATES, M. P., RUSSELL, D. W., BROWN, M. S. &
GOLDSTEIN, J. L. (1988). Human low density lipoprotein receptorexpressed in Xenopus oocytes. J. biol. Chem, 263, 7838-7845.
PHILLIPS, C. R. (1985). Spatial changes in polyA concentrationsduring early embryogenesis in Xenopus laevis: analysis by in situhybridization. Devi Biol. 109, 299-310.
ROBERTS, S. (1989). Crosslinking of GTP to tubulin in Xenopuseggs: the possible basis of UV inactivation of dorsaldevelopment. In preparation.
ROWNINC, B. (1989). Microtubule-mediated cortical rotation in theXenopus egg. Thesis, University of California, Berkeley CA.,156 pp.
SAXEN, L. & TOIVONEN, S. (1961). The two gradient hypothesis inprimary induction. The combined effect of two types of inductorsin different ratios. J. Embryol. exp. Morph. 9, 514-528.
SCHARF, S. R. & GERHART, J. C. (1980). Determination of thedorso-ventral axis in eggs of Xenopus laevis: Complete rescue ofUV-impaired eggs by oblique orientation before first cleavage.Devi Biol. 79, 181-198.
SCHARF, S. R. & GERHART, J. C. (1983). Axis determination in eggsof Xenopus laevis: A critical period before first cleavage,identified by the common effects of cold, pressure, andultraviolet irradiation. Devi Biol. 99, 75-87.
SCHARF, S. R., ROWNINO, B., W U , M. & GERHART, J. C. (1989).
Hyperdorsoanterior embryos from Xenopus eggs treated withD2O. Devi Biol. (in press).
SLACK, J. M. W. & ISAACS, H. V. (1989). Presence of basicfibroblast growth factor in the early Xenopus embryo.Development 105, 147-154.
SLACK, J. M. W., ISAACS, H. V. & DARLINGTON, B. G. (1988).
Inductive effects of fibroblast growth factor and lithium ion onXenopus blastula ectoderm. Development 103, 581-590.
SMITH, J. C. & SLACK, J. M. W. (1983). Dorsalization and neuralinduction: Properties of the organizer in Xenopus laevis.J. Embryol. exp. Morph. 78, 299-317.
SPEMANN, H. (1938). Embryonic Development and Induction, YaleUniversity Press, New Haven, 401pp.
STEWART, R. & GERHART, J. C. (1989). The organization of the
marginal zone of the late blastula stage of Xenopus laevis. DeviBiol. (submitted).
STEWART-SAVAGE, J. & GREY, R. (1982). The temporal and spatialrelationship between cortical contraction, sperm trail formation,and pronuclear migration in fertilized Xenopus eggs. WilhelmRoux1 Arch, devl Biol. 191, 241-245.
TANNAHILL, D. & MELTON, D. A. (1989). Localized synthesis ofVgl protein during early Xenopus development. Development, inpress.
TERASAKI, M., CHEN, L. B. & FUJIWARA, K. (1986). Microtubules
and the endoplasmic reticulum are highly independent structures.J. Cell Biol. 103, 1557-1568.
TOURTE, M., MIGNOTTE, F. & MOUNOLOU, J.-C. (1984).
Heterogeneous distribution and replication activity ofmitochondria in Xenopus laevis oocytes. Eur. J. Cell Biol. 34,171-178.
UBBELS, G. A., HARA, K., KOSTER, C. H. & KIRSCHNER, M. W.(1983). Evidence for a functional role of the cytoskeleton indetermination of the dorsoventral axis in Xenopus laevis eggs.J. Embryol. exp. Morph. Tl, 15-37.
VALE, R. D. (1987). Intracellular transport using microtubule basedmotors. A. Rev Cell Biol. 3, 347-378.
VINCENT, J.-P. & GERHART, J. C. (1987). Subcortical rotation inXenopus eggs: an early step in embryonic axis specification. DeviBiol. 123, 526-539.
VINCENT, J.-P., OSTER, G. F. & GERHART, J. C. (1986). Kinematicsof grey crescent formation in Xenopus eggs: The displacement ofsubcortical cytoplasm relative to the egg surface. Devi Biol. 113,484-500.
VINCENT, J.-P., SCHARF, S. R. & GERHART, J. C. (1987). Subcorticalrotation in Xenopus eggs: a preliminary study of itsmechanochemical basis. Cell Mot. & Cytoskel. 8, 143-154.
WADDINGTON, C. H. & PERRY, M. M. (1956). Teratogenic effects oftrypan blue on amphibian embryos. J Embryol. exp. Morph. 4,110-119.
WESSEL, G. & MCCLAY, D. R. (1987). Gastrulation in the seaurchin requires deposition of cross linked collagen within theextracellular matrix. Devi Biol. 103, 235-245.
WILSON, P. A., OSTER, G. & KELLER, R. (1989). Cellrearrangement and segmentation in Xenopus: direct observationof cultured explants. Development 105, 155-166.
WYLIE, C. C , BROWN, D., GODSAVE, S. F., QUARMBY, J. &
HEASMAN, J. (1985). The cytoskeleton of Xenopus oocytes and itsrole in development. J. Embrvol. exp. Morph. 89 Supplement,1-15.



























![[ 149 ] the growth of the hindlimb bud of xenopus laevis and its](https://img.pdfslide.net/doc/110x75/586789b31a28ab44568b868b/-149-the-growth-of-the-hindlimb-bud-of-xenopus-laevis-and-its-.jpg)


