Embed Size (px)
Citation preview

Plant Cell and Environment
(2005)
28
997ndash1011
copy 2005 Blackwell Publishing Ltd
997
No claim to original US government works
Blackwell Science LtdOxford UKPCEPlant Cell and Environment0016-8025Blackwell Science Ltd 2005 2005
289971011Original Article
Crop responses to ozoneE L Fiscus
et al
Correspondence EL Fiscus Fax
+
1 919 515 3593 e-mailedfiscus01sprynetcom
Crop responses to ozone uptake modes of action carbon assimilation and partitioning
EDWIN L FISCUS FITZGERALD L BOOKER amp KENT O BURKEY
US Department of Agriculture Agricultural Research Service Plant Science Research Unit and Department of Crop Science North Carolina State University 3908 Inwood Road Raleigh NC 27603 USA
ABSTRACT
The inhibitory effects of tropospheric O
3
on crop photosyn-thesis growth and yield have been documented in numer-ous studies over the past 35 years In large part the resultsof this research supported governmental regulationsdesigned to limit tropospheric O
3
levels to concentrationsthat affected crop production at economically acceptablelevels Recent studies have brought into question the effi-cacy of these concentration-based O
3
standards comparedwith flux-based approaches that incorporate O
3
uptakealong with environmental and biotic factors that influenceplant responses In addition recent studies provide insightinto the biochemical mechanisms of O
3
injury to plantsCurrent interpretations suggest that upon entry into theleaf intercellular space O
3
rapidly reacts with componentsof the leaf apoplast to initiate a complex set of responsesinvolving the formation of toxic metabolites and generationof plant defence responses that constitute variably effectivecountermeasures Plant species and cultivars exhibit arange of sensitivity to O
3
evident as heritable characteris-tics that must reflect identifiable biochemical and molecu-lar processes that affect sensitivity to O
3
injury althoughtheir exact makeup remains unclear Ozone clearly impairsphotosynthetic processes which might include the effectson electron transport and guard cell homeostasis as well asthe better-documented effects on carbon fixation viadecreased Rubisco activity Translocation of photosynthatecould be inhibited by O
3
exposure as well Further theinfluence of tropospheric O
3
needs to be considered whenassessing potential effects of rising concentrations of atmo-spheric CO
2
on crop production Advances in O
3
flux mod-elling and improved understanding of biochemical andmolecular effects of O
3
on photosynthetic gas exchange andplant defence processes are leading to more complete inte-grated assessments of O
3
impacts on crop physiology thatcontinue to support the rationale for maintaining orimproving current O
3
air quality standards as well as pro-viding a basis for development of more O
3
-tolerant croplines
Key-words
air pollution flux ozone partitioningphotosynthesis reactive oxygen species soybeantranslocation uptake yield
INTRODUCTION
Tropospheric O
3
is currently viewed as a widespread andgrowing problem that suppresses crop productivity on alarge scale (US Environmental Protection Agency 1996Mauzerall amp Wang 2001 Fuhrer amp Booker 2003) The prob-lem was originally perceived to be limited in nature con-fined to urban centres proximity to power plants and areasdownwind or nearby these sources However the scale ofthe problem has increased in scope in the last 25 years as aresult of increasing population densities industrializationand transportation-related activities in large parts of theworld particularly in the less developed countries In addi-tion there is increased recognition of transboundary trans-port of O
3
precursors in the troposphere In response anumber of efforts have been made to define the impact ofO
3
on crop productivity use this information to predict croplosses and to set air quality standards that should keep croplosses to an economically acceptable level (US Environ-mental Protection Agency 1996 UNECE 2000)
Current worldwide average tropospheric O
3
levels wereapproximately 50 nmol mol
-
1
in the year 2000 already 25above the threshold established for damage to sensitiveplants (Fuhrer Skarby amp Ashmore 1997) While global meanvalues have increased from an estimated pre-industriallevel of 38 nmol mol
-
1
[25ndash45 nmol mol
-
1
8-h summer sea-sonal average (US Environmental Protection Agency1996)] to about 50 nmol mol
-
1
in 2000 the most pessimisticprojections suggest a further increase to 80 nmol mol
-
1
by2100 Most of this increase would be driven by a nearlythree-fold increase in NO
X
and CH
4
emissions (Prather
et al
2001) It is critical to understand that these globalmeans are comprised of local means many of which arealready substantially above the projections and typicallyoccur sometime during the cropping season
Although the actual economic costs of O
3
-induced croplosses are difficult to assess the total benefits resulting fromvarious regulatory scenarios mostly involving reductions ofcurrent ambient levels ranged from about 01ndash25 B$ (in1980 US dollars) in the United States (Adams amp Horst2003) Additionally the US Environmental Protection
998
E L Fiscus
et al
copy 2005 Blackwell Publishing Ltd
Plant Cell and Environment
28
997ndash1011No claim to original US government works
Agency in its 1996 criteria document estimated annualnational level losses to major crops to be in excess of 1 B$(in 1990 US dollars) (US Environmental ProtectionAgency 1996) Further Mauzerall amp Wang (2001) cite sev-eral recent studies estimating benefits of 2ndash33 B$ in the USby eliminating O
3
precursors from motor vehicle emissions(Murphy
et al
1999) and 310 M
euro
in the Netherlands (Kuik
et al
2000) and 2 B$ in China (Mauzerall amp Wang 2001) byreducing O
3
to background levels (see also Ashmore 2005this issue) Also due to the non-linear shape of many crop-O
3
dosendashresponse curves we might expect a disproportion-ately larger effect for each unit increase in global averageO
3
concentrationsThe yield or productivity responses to O
3
of a wide rangeof crops clearly show a negative relationship and thereseems little to be gained by dwelling further on that issueexcept to emphasize that a wide range of responses andlevels of sensitivity exists among the many plants that havebeen tested (Heck
et al
1983 Heagle 1989 US Environ-mental Protection Agency 1996 Fuhrer
et al
1997 MorganAinsworth amp Long 2003) The past decade has seen sub-stantial progress in interpreting the effects of O
3
on plantsand the mechanisms by which those effects are mediatedHowever there is still much to be done in a practical sensebefore that information can be translated into usefulproducts
It is our purpose in this paper to review recent progressin understanding how O
3
brings about its effects and toraise occasional questions or note exceptions to the currentdogma We start by examining the rationale for flux-basedcharacterizations of O
3
exposures which may prove advan-tageous for formulation and implementation of environ-mental impact assessment and regulation Then we considerO
3
entry into leaves and the immediate reactions that occurin the leaf apoplast the protective mechanisms that may beactivated and possible signals generated that result in mod-ified plant behaviour Then since photosynthesis seemsconsistently affected by O
3
exposure especially during thereproductive phase of crop development and since photo-synthetic processes directly contribute to crop productivitywe review some recent evidence detailing the effects of O
3
exposure on photosynthetic systems in various crop speciesIn addition the distribution of the resultant photosynthateis considered in relation to O
3
stress and how perturbationsin that distribution might contribute to the overall impactPotential interactions between rising concentrations ofatmospheric CO
2
and O
3
are briefly addressed Finally wetouch upon some of the sources of experimental variabilityand the consequences thereof
O
3
EXPOSURE AND UPTAKE
First interpretations of cause and effect relationshipsbetween O
3
and plant responses are commonly based onthe O
3
concentrations in the surrounding air For examplevarious plant responses to O
3
have been correlated withaverage O
3
concentration over some time interval (egdaily 7 8 or 12 h averages) Other more biologically rele-
vant schemes illustrated in Fig 1 involve summing O
3
con-centrations that exceed some threshold value expressedeither as a step function (eg SUM06 AOT40) or a con-tinuous sigmoidal weighting function (eg W126) Still oth-ers involve a combination of hourly averages and numberof peak values (Lefohn 1992 US Environmental ProtectionAgency 1996) The advantages of concentration-basedapproaches are clear measurements of O
3
concentrationand computation of exposure indices are straightforwardand air quality standards can be based on verifiable dataThe validity of this approach is supported by numerouscontrolled environment greenhouse and field experimentsthat show a consistent negative relationship between O
3
exposure and photosynthesis growth and yield (Miller1987b Heagle 1989 Fuhrer
et al
1997)Atmospheric O
3
concentration indices are used world-wide for establishing air quality standards Experimentalresults however indicate a wide range in relative toleranceto O
3
among crops species and cultivars which is influencedby a number of environmental factors and other air pollut-ants experienced during plant growth (US Environmental
Figure 1
Illustration of the SUM06 AOT40 and the continuous sigmoidal weighting scheme imposed on a typical summertime daily average [O
3
] curve for Raleigh NC USA Each of the functions use hourly average [O
3
] and are defined as AOT40
=
S
([O
3
]
-
40) for [O
3
]
gt
40 SUM06
=
S
[O
3
] for [O
3
]
ge
60 and W126
=
S
([O
3
] W) where the sigmoidal weighting function W
=
1(1
+
4403(exp(
-
0126 [O
3
]))) (US Environmental Protection Agency (1996) All concentrations are in ppb or the SI equivalent and W126 is applied to all the hourly averages Units for all are ppbh
Time of day
0 4 8 12 16 20 24
[O3]
(p
pb
)
0
20
40
60
0
20
40
60
80
AOT40190 ppb h
W126195 ppb h
Mean 8ndash20 h [O3] = 55 ppb
SUM06375 ppb h
Crop responses to ozone
999
copy 2005 Blackwell Publishing Ltd
Plant Cell and Environment
28
997ndash1011No claim to original US government works
Protection Agency 1996 Fuhrer
et al
1997) In addition theO
3
concentration at measuring height (3ndash10 m) is usuallynot corrected for the lower concentration received at can-opy height (Tuovinen 2000) However a protocol for mak-ing this correction has been devised (UNECE 2004b)
Efforts to understand the discrepancies between O
3
con-centration in the air and variable plant responses havefocused on molecular and biochemical mechanismsinvolved in O
3
detoxification processes and certain interact-ing environmental factors affecting stomatal conductance(
g
s
) (Fuhrer
et al
1997 Massman 2004 Pleijel
et al
2004)Environmental factors influencing
g
s
are important becauseO
3
uptake mainly occurs through leaf stomata (Runeckles1992 Long amp Naidu 2002) Various modelling results haveindicated that a major factor affecting plant responses toO
3
appears to be
g
s
(Runeckles 1992 Massman 2004 Pleijel
et al
2004) This is supported by experiments with a varietyof crops exposed simultaneously to O
3
and elevated atmo-spheric CO
2
an effective antitranspirant that showedreduced O
3
injury presumably due in large part to loweredO
3
flux (McKee Farage amp Long 1995 Fiscus
et al
1997Reid amp Fiscus 1998 Reid Fiscus amp Burkey 1998 Heagle
et al
1999 Reid Fiscus amp Burkey 1999 Olszyk
et al
2000Fiscus
et al
2002 Morgan
et al
2003 Booker Fiscus ampMiller 2004 Booker
et al
2005) Reduced O
3
damage towater-stressed soybean [
Glycine max
(L) Merr] comparedwith well-watered plants was attributed to lower g
s
as well(Tingey amp Hogsett 1985 Vozzo
et al
1995)The major environmental factors controlling
g
s
includeleaf temperature water vapour pressure differencebetween the leaf and the surrounding air (VPD) photosyn-thetic photon flux density (PPFD) soil water availabilityand atmospheric CO
2
concentration (Jarvis 1976) Thusresponses to some O
3
concentration for a particular cropand cultivar undoubtedly depend upon inherent geneticdifferences and environmental conditions experienced dur-ing the growing season In order to incorporate the effectsof environmental conditions relevant to
g
s
into an exposureindex the UNECE ICP Vegetation project is investigatingthe use of VPD estimates in conjunction with O
3
concen-tration for defining the critical O
3
level that produces visibleinjury in crops (UNECE 2004b) Stomatal conductance istypically higher in warm humid environments comparedwith hot dry conditions Thus crops in warm humid envi-ronments are likely at increased risk of O
3
injury Thisapproach holds promise as a relatively simple method forincluding environmental factors that influence g
s
into O
3
exposure indices However other factors such as leaf tem-perature leaf hydration or PPFD (Massman 2004) couldindependently affect O
3
toxicity which would require addi-tional modifications of such an index
From a toxicological viewpoint it is the absorbed cumu-lative dose of O
3
that is most relevant in determining causeand effect relationships and quantifying dosendashresponsesThus O
3
flux models using a multiplicative algorithm basedon the approach of Jarvis (1976) to model
g
s
have incorpo-rated VPD air temperature PPFD and soil moisture deficitcoefficients to compute estimates of O
3
uptake (Emberson
et al
2000 Pleijel
et al
2004) A phenological componentalso has been added to account for changes in
g
s
during thegrowing season This approach indicated that phenologyand VPD were important factors in estimating O
3
fluxComparisons between flux estimates and O
3
exposure mapsof Europe suggested that areas with the highest O
3
concen-trations differed from those regions calculated as havingthe highest O
3
fluxes (Emberson
et al
2000) The highestO
3
concentrations occurred in the southern Mediterraneanregion whereas the highest O
3
fluxes for wheat occurred insouthern Scandinavia and northern Europe
However some crops and cultivars with similar gs havedifferent tolerances to the same O3 concentrations Thus ithas been suggested that O3 flux models include a coefficientfor O3 detoxification capacity (Musselman amp Massman1999 Fuhrer amp Booker 2003 Massman 2004) Musselmanamp Massman (1999) defined the lsquoeffective fluxrsquo as the bal-ance between uptake into the leaf at a given time and thedefence response at that time The defence response factorwas proportional to the effect of O3 on gross photosynthe-sis Effective flux integrated over time yields cumulativeeffective loading Using a relatively comprehensive model-ling approach for O3 deposition and uptake Massman(2004) estimated that the period of highest effective loadingoccurred between 1300 and 1600 h
The cumulative exposure indices (SUM06 W126AOT40) are based on O3 injury thresholds for effects of O3
concentration on plant growth and yield In the same man-ner cumulative O3 flux models show improved perfor-mance when a threshold O3 flux factor is included in themodel eg Pleijel et al (2004) Likewise Martin et al(2000) modified WIMOVAC (Windows Intuitive Model ofVegetation Response to Atmosphere and Climate Change)(Humphries amp Long 1995) to include O3 detoxification pro-cesses when calculating threshold flux-based rather thanconcentration-based dosendashresponse models Howeverdetermination of the threshold coefficient in the Martinet al (2000) model requires experimentally derived assess-ments of O3 effects on Vcmax The situation is a bit morecomplicated because O3 can damage plant photosynthesiswhich in turn may reduce the plantrsquos detoxification ability(Massman 2004) In effect there are interacting mecha-nisms at work that may alter O3 injury thresholds as dam-age accumulates (Massman 2004) Ozone increases therelative cost of detoxification on the one hand anddecreases the need through Ci-driven reductions in gs onthe other A flux-based approach that includes thresholdsrequires further experimentation and advances in ourunderstanding of fundamental processes involved in O3
action among different plant species Furthermore thisapproach needs to be scaled to the canopy level to begenerally applicable Micrometeorological soil-vegetation-atmosphere transfer (SVAT) models have been developedthat estimate stomatal O3 flux and cuticular depositionbased on O3 concentration atmospheric boundary layerand bulk canopy resistances to describe the verticalexchange of O3 to the plant-soil system (Gruumlnhage Haenelamp Jaumlger 2000)
1000 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
In the field canopy structure and foliage density affectthe concentration of O3 that leaves experience while envi-ronmental factors such as PPFD leaf temperature and VPDvary within the leaf canopy and can affect gs and thus O3
flux However after canopy closure for crops such as soy-bean cotton (Gossypium hirsutum L) and peanut (Ara-chis hypogaea L) leaves at the top of the canopy providethe majority of photosynthate for seed production Ozoneflux to the upper portion of the canopy is likely most criticalto impacts on yield For crops such as wheat (Triticum aes-tivum L) and rice (Oryza sativa L) flux to the entirecanopy might be important Efforts to parameterize canopyO3 flux have included leaf area index as a way to estimatecanopy gs (Massman 2004) which may be effective inaccounting for differences among crop canopy structuresHowever the ICP Vegetation protocol utilizes projectedleaf area namely leaf area of sunlit leaves at the top of thecanopy to calculate O3 flux (Pleijel et al 2004 UNECE2004b) Ozone flux to the most productive leaves is viewedas having the greatest impact on yield because even thoughthe soil and lower parts of the canopy consume largeamounts of O3 it is likely these fluxes have much less effecton yield (Pleijel et al 2004 Ashmore 2005)
Lastly the ICP Vegetation program (UNECE 2004a) iscurrently comparing economic crop loss assessments ofconcentration-based and flux-based methodologies Thisanalysis should suggest whether the flux approach providesa significantly better assessment of crop losses due to O3
pollution compared with concentration-based exposureindices
MODES OF ACTION
Apoplastic and symplastic reactions and responses
The internal air spaces within the leaf are a potential sitefor O3 reactions with volatile compounds produced by theplant Isoprene is one example of a compound emitted frommany angiosperms and conifers (Sharkey amp Yeh 2001) thatcould react with O3 However volatile compounds releasedby crop species are mainly monoterpenes which have beenlittle studied in this context although parallels betweenthese volatile organic compounds (VOCs) and O3 reactionswith isoprene are likely At present hypotheses are contra-dictory regarding the role of isoprene in plant responses toO3 Initially isoprene was thought to enhance O3 damagethrough the formation of hydroperoxides (Hewitt Kok ampRall 1990) More recently isoprene was found to exert aprotective effect either through action as a radical scaven-ger or membrane stabilizer (Loreto amp Velikova 2001) Theconcept of isoprene as an antioxidant is intriguing from theperspective of O3 scavenging reactions outside the leaf thatcould increase boundary layer resistance to O3 and effec-tively reduce the dose The potential benefits of such amechanism must be balanced against the metabolicdemand for carbon to support isoprene synthesis (Sharkeyamp Yeh 2001) and the consequences of increased plant-
derived VOCs on atmospheric chemistry leading to O3
formationOzone that has passed through leaf internal air spaces
will then dissolve in the aqueous layer surrounding leafcells The breakdown of O3 in pure water produces hydroxyland peroxyl radicals and superoxide although the reactionsproceed very slowly at neutral pH (Heath 1987) GrimesPerkins amp Boss (1983) demonstrated that the rate ofhydroxyl radical formation from O3 in aqueous solution issignificantly greater when phenolic compounds are presentsuggesting that biologically relevant compounds enhanceO3 reactions Based on in vitro and in vivo studies (Heath1987 Runeckles amp Chevone 1992 Pryor 1994 Kirichenkoet al 1996) O3 is known to react with a diverse set ofmolecules that would be encountered within the cell walland on the plasma membrane surface Initial targets for O3
include plasma membrane lipids susceptible amino acids inplasma membrane proteins or apoplastic enzymes and avariety of organic metabolites localized in the cell wallPotentially ozonolysis products from the oxidation of lipidsand metabolites containing carbon-carbon double bonds(Heath 1987 Runeckles amp Chevone 1992) and alteredplasma membrane protein function (Dominy amp Heath1985 Castillo amp Heath 1990) could serve as the initial sig-nals leading to O3 responses However the primary O3
reaction products in plants have not been identified due todifficulties in following O3 reactions within the complexbiochemical network of the leaf apoplast Knowledge ofthese early events is critical to understanding the metabolicsignals that initiate O3 responses and may provide insightinto O3 tolerance mechanisms localized in the leaf apoplastOzone incorporation into the cells and extracellular lipid-protein complexes of rat lung tissue has been studied usinga combination of 18O3 and mass spectrometry (Gunnison ampHatch 1999) an approach that could be employed in con-junction with available methods for isolation of leaf apo-plast components and plasma membranes to addressquestions regarding the initial O3 reaction products inplants
Formation of reactive oxygen species (ROS) such assuperoxide hydrogen peroxide and hydroxyl radical arethought to be associated with the initial breakdown of O3
in the leaf apoplast ESR approaches based on spin trap-ping suggest that free radicals are involved in the earlystages of O3 response (Grimes et al 1983 Mehlhorn Tab-ner amp Wellburn 1990 Runeckles amp Vaartnou 1997) but itis not clear whether the observed signals originate fromdirect O3 breakdown or the initial reactions with cellularconstituents such as phenolic compounds Localizedregions of ROS formation either hydrogen peroxide orsuperoxide depending on the plant species are associatedwith O3 response in sensitive plants (Wohlgemuth et al2002 Pendall et al 2004) The kinetics of ROS formation inthe leaf apoplast following O3 exposure seem to involve twophases an initial phase that is associated with direct effectsof O3 and a second phase associated with a plant-derivedsecondary oxidative burst (Schraudner et al 1998) The sec-ondary oxidative burst is initially localized in the leaf apo-
Crop responses to ozone 1001
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
plast and cell wall but later expands into the cytoplasm andsubcellular compartments leading to the formation of visi-ble lesions (Pendall et al 2004)
The initial signals induced by O3 in the leaf apoplastmight then be translated into responses at the tissue levelResponses vary and include unregulated cell death hyper-sensitive response leading to programmed cell death andaccelerated senescence (Pell Schlagnhaufer amp Arteca 1997Sandermann et al 1998) Evidence is accumulating thatthese responses are modulated by ethylene jasmonic andsalicylic acid levels and the interactions among their signal-ling pathways (Kangasjaumlrvi et al 1994 Sharma et al 1996Pell et al 1997 Overmyer et al 2000 Rao et al 2000 Raoamp Davis 2001 Rao Lee amp Davis 2002 Overmyer Broscheamp Kangasjaumlrvi 2003 Tamaoki et al 2003 Vahala et al 2003see also Kangasjaumlrvi et al 2005 this issue) For exampleArabidopsis thaliana mutants that overproduce ethylene(eto1) or are insensitive to jasmonic acid (jar1) are moreextensively injured by O3 than wild-type plants (Overmyeret al 2000 Rao et al 2000 Tamaoki et al 2003) Ethyleneacts as a promoting factor for O3 injury whereas jasmonatesmight have a role in minimizing injury (Overmyer et al2000 2003)
Ozone effects at the tissue level are often characterizedas either acute or chronic responses subjective terms thatare related to the type of visible symptoms observed andwhether the O3 concentrations employed typify ambient O3
episodes For example unregulated cell death and pro-grammed cell death are usually considered acute responsesin which lesions occur within hours after exposure to rela-tively high O3 concentrations (typically gt150 nmol mol-1)In contrast chronic responses include lesions that developover days to weeks under lower O3 concentrations andaccelerated senescence where lesions might not formUnder field conditions accelerated senescence may be dif-ficult to identify in the absence of a clean air control orother reference point (eg O3-tolerant cultivar) because thecumulative effects on foliar senescence at the end of theseason are separated in time from the O3 events causingthe effects The timing of O3 episodes in relation to plantdevelopment can also have a major effect on the outcomeas in the case of soybean in which O3 exposure duringreproductive growth has a much greater impact on yieldcompared with exposure during vegetative growth (Heagleet al 1991 Miller et al 1991 Morgan et al 2003) The dis-tinction between acute and chronic responses is furthercomplicated by species and genotype differences that leadto a range of effects for a given combination of O3 concen-tration and environmental conditions
Genetic variation in O3 responses and potential O3 tolerance mechanisms
Studies of O3 effects on plants often identify genetic varia-tion as a contributing factor in the observed responseOzone-sensitive and tolerant cultivars or clones have beenreported for many plant species (Wellburn amp Wellburn1996) including crops such as soybean (Tingey Reinert amp
Carter 1972 Heagle amp Letchworth 1982 Robinson amp Britz2000) snap bean (Phaseolus vulgaris L) (Guzy amp Heath1993) tomato (Lycopersicon esculentum Miller) (Temple1990) clover (Trifolium repens L) (Heagle et al 1993)wheat (Heagle Miller amp Pursley 2000) and potato(Solanum tuberosum L) (Heagle Miller amp Pursley 2003)and natural vegetation such as black cherry (Prunus sero-tina Ehrh) (Lee et al 2002) and Plantago spp (Wolff Mor-gan-Richards amp Davison 2000) Studies have also shownthat O3 tolerance is a heritable trait (Damicone amp Manning1987 Reinert amp Eason 2000) Although much effort hasbeen made to identify the physiological and biochemicalelements of O3 tolerance no clear picture has emergedBased on the oxidative stress model formation of ROS andother oxidation products is the underlying process involvedin the generation and propagation of toxic compounds andabiotic elicitors in plants Therefore features of antioxidantmetabolism that scavenge or affect the perception of O3-derived ROS could contribute to O3 tolerance
Plants have evolved elaborate systems to combat generaloxidative stress based on enzymes that utilize ascorbic acidand glutathione (Noctor amp Foyer 1998 see also Foyer ampNoctor 2005 this issue) and these antioxidant systems havebeen linked to O3 stress tolerance (Runeckles amp Chevone1992 Smirnoff 1996 Chernikova et al 2000 Conklin ampBarth 2004) A critical role for ascorbic acid is demon-strated with the vtc1 mutant of Arabidopsis in which asignificant decrease in ascorbate content was associatedwith increased O3 sensitivity (Conklin Williams amp Last1996) A more detailed analysis of vtc1 suggested that inaddition to acting as an antioxidant ascorbic acid serves aregulatory function in the signalling networks that controlplant defence responses and leaf senescence (Conklin ampBarth 2004) Although it is clear that minimum levels ofascorbic acid and glutathione are required for normal leaffunction the concentrations of these antioxidant metabo-lites that naturally occur in leaf tissue are not always wellcorrelated with O3 tolerance (Guzy amp Heath 1993 Well-burn amp Wellburn 1996 Burkey et al 2000) suggesting amore complex mechanism that remains to be elucidated
Recent studies have found higher levels of ascorbic acidin the leaf apoplast of certain O3-tolerant snap bean geno-types compared with sensitive lines (Burkey Eason amp Fis-cus 2003) suggesting that localization of ascorbic acid inthe cell wall where initial O3 reactions occur might beimportant Leaf extracellular ascorbate content and redoxstatus are affected by O3 treatment (Castillo amp Greppin1988 Luwe amp Heber 1995 Burkey 1999) evidence thatextracellular ascorbic acid is involved in detoxification ofO3 and related ROS Ascorbic acid is initially synthesizedinside cells (Smirnoff Conklin amp Loewus 2001) followedby transport between the cytoplasm and extracellular spacevia specific carriers located in the plasma membrane (Hore-mans Foyer amp Asard 2000) A model developed by Cha-meides (1989) and expanded by others (Ploumlchl et al 2000Turcsanyi et al 2000) features ascorbic acid in the leaf apo-plast as a critical factor in the detoxification of extracellularO3 and ROS protecting plasma membranes from oxidative
1002 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
damage and preventing O3 injury Ascorbic acid potentiallycould serve as either a direct chemical scavenger of O3although this mechanism has been questioned (Jakob ampHeber 1998) or as a substrate for extracellular enzymes(eg ascorbate peroxidase) that attenuate ROS levels andthus affect the propagation of the initial O3 signal How-ever leaf apoplast ascorbic acid levels can be quite low incertain plant species [summarized in Burkey et al (2003)]suggesting that extracellular scavenging of O3 and ROS byascorbate may not be a significant factor in all plants Othercompounds with antioxidant properties also exist in the leafapoplast Identification of these compounds and theirimpact on ROS formation and propagation are subjects forfuture research on O3 tolerance mechanisms
Another approach to understanding O3 responses inplants involves investigations of the potential interactionbetween chemical signals and specific receptors in theplasma membrane Booker et al (2004) showed that theepinasty frequently observed for wild-type Arabidopsisleaves after O3 treatment did not occur in gpa1 mutantplants lacking the alpha subunit of the heterotrimeric Gprotein complex suggesting that G-proteins may beinvolved in the transduction of O3-derived signals Thebinding of O3 or more likely one of the postulated O3 reac-tion products to a G-protein receptor could activate theprotein and initiate signal cascade processes that activatetarget proteins in this signal transduction system Howeverfurther studies will be required to define the possible roleG-proteins have in O3-induced signal transduction andpropagation It is also possible that G-proteins modulate O3
responses via hormone signal transduction processes Ineither case elements of G-protein pathways involved in O3
response represent potential molecular targets that mightbe manipulated to enhance O3 tolerance although this isspeculative at present The challenge will be to identifytargets unique to O3 so that other important stressresponses remain intact (eg plant defence responsesagainst pathogen attack) Finally it is likely that any pro-tective mechanism will come with some metabolic cost thatwill require evaluation to determine its net benefits
IMPACTS ON PHOTOSYNTHESIS
Net assimilation
Other than crop yield and visible injury photosynthesis hasbeen the most broadly studied aspect of plant responses toO3 Generally O3 exposure results in decreased photosyn-thetic carbon assimilation (Miller 1987b Runeckles ampChevone 1992 Pell et al 1997 Long amp Naidu 2002 Morganet al 2003) In many cases loss of assimilation capacity wasshown to be due primarily to reduced carboxylation effi-ciency directly related to loss of Rubisco activity Theselosses of activity are thought to be due to decreases inRubisco concentration in the leaves rather than a decreasein activation state Messenger RNA transcripts for theRubisco small subunit (rbcS) have been noted to declinewith O3 treatment in potato leaves (Glick et al 1995) sug-
gesting that protein synthesis might be inhibited a notionconsistent with observations that Rubisco content in poplar(Populus spp) leaves treated with O3 does not rise as highas controls during leaf expansion (Pell et al 1997) How-ever Eckardt amp Pell (1994) under conditions where addi-tional synthesis should be minimal showed a substantialdecline in Rubisco content suggesting that the primarycause of the decline in Rubisco due to O3 exposure wasenhanced degradation rather than reduced production
Because photosynthetic measurements are typicallymade on the most recently fully expanded upper canopyleaves in determinate plants including many soybean linesit is often difficult to demonstrate differences in net photo-synthesis (An) due to O3 until near or after the time whenproduction of new upper canopy leaves has ceased Thenduring most of the reproductive stages An decreased earlierthan in control plants (accelerated senescence) This sug-gests that during vegetative growth either Rubisco synthe-sis in newly formed upper canopy leaves is not substantiallyaffected or that these leaves have not accumulated suffi-cient damage to elicit a response Vozzo et al (1995) dem-onstrated significant reductions in An on somewhat oldersoybean leaves that were relatively unaffected by shadingbut had received sufficient exposure to exhibit visible symp-toms thus lending support to this idea In contrast Morganet al (2004) using an indeterminate line were unable todemonstrate any differences due to dose accumulation in acohort of soybean leaves that developed during vegetativegrowth However they did observe decreased An in anothercohort that developed during reproductive growth andremained near the top of the canopy These results couldwell have been due to differences in exposure methodolo-gies particularly the greater atmospheric coupling to thelower canopy in OTCs as compared to FACE systems Infurther contrast to the above observations additionalreview of the data in two earlier soybean studies (Reid ampFiscus 1998 Reid et al 1998) illustrated in Fig 2 showsthat Rubisco content and activity chlorophyll content andAn were reduced by O3 early in the vegetative stage andthat all but An had reached or exceeded clean air levels byearly flowering Afterwards while chlorophyll content Anand Rubisco activity remained level or declined slightlyRubisco content continued to rise in the O3 treatmentFinally in early seed development all these parametersbegan to decline more precipitously Throughout this wholeexperiment chlorophyll content and Rubisco activitytracked each other very well This suggests that from earlyflowering through most of seed development O3 stimulatedRubisco content on a leaf area basis In addition the ques-tion of the role of accelerated senescence especially duringreproductive growth is one deserving more attention Forexample in Fig 2 the O3-induced decline in An duringreproduction may be due to the normal process of senes-cence that has been accelerated by O3 exposure or it maybe due simply to the loss of photosynthetically active leafarea due to accumulated O3 damage Thus especially inareas associated with visible lesions one would measure amixture of live and dead or dying cells and it seems rele-
Crop responses to ozone 1003
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
vant to know whether the remaining live cells are function-ing at or near normal levels for their age This possibility issuggested by the fact that from beginning of pod fill throughseed maturation when An per unit area is declining in thisstudy An per unit Rubisco remained high at 91 of the CFcontrol (difference not significant and data not shown)Further information is necessary to clarify this Anotherpoint deserving attention and bearing on the exposureaccumulation comments earlier in the paragraph is the sub-stantially reduced An early in the vegetative period Thismay have been caused by some inherent sensitivity of theseleaves or be the result of enhanced atmospheric couplingdue to the more open nature of the canopy at this growthstage
Photosynthetic electron transport
Detrimental effects of O3 on photosynthetic electron trans-port has received increased attention in recent years andwhile earlier it had been difficult to demonstrate significantdeclines in function evidence is accumulating that in someplants there are demonstrable effects both on PSII func-tion as well as the xanthophyll cycle components Forinstance significant and substantial declines in FvFm whichrepresents the efficiency of excitation energy capture byPSII in dark-adapted leaves have been reported for pump-kin (Cucurbita pepo L) (Ciompi et al 1997 Castagna et al2001) wheat (T durum and T aestivum) (Reichenauer et al1998) bean (P vulgaris L) (Guidi Di Cagno amp Soladatini2000a Guidi Tonini amp Soladatini 2000b Leipner Oxbor-ough amp Baker 2001 Guidi DeglrsquoInnocenti amp Soldatini2002) tomato (Calatayud amp Barreno 2001) and Festucapratensis and turnip (Brassica napus L) (Plazek Rapacz ampSkoczowski 2000) Of these particular studies three dem-
onstrated an increase in Fo which suggested damaged ordeactivated PSII centers (Guidi et al 2000a Leipner et al2001 Guidi et al 2002) all occurring in bean In additionlight saturated photosynthesis (ASAT) decreased in bean(Guidi et al 2000a Guidi et al 2002) pumpkin (Ciompiet al 1997 Castagna et al 2001) and wheat (Reichenaueret al 1998) although in the latter the effect was confined toolder leaves in two of the three lines and was partiallyreversible in some lines and fully reversible overnight inone It was unknown whether reversibility was due to anight-time epoxidation of the xanthophyll cycle decreasingsubsequent thermal quenching of PSII or to overnightrepair of damaged PSII centres This latter point becomesrelevant since a decrease in FvFm may be caused by both arelatively fast reversible down-regulation or a reversibledestruction (Osmond 1994) However the two processesmight be distinguished by monitoring effects on Fo in thatthermal dissipation by the xanthophyll cycle would lowerFo whereas damage to PSII would increase Fo as was foundin bean (Guidi et al 2000a 2002 Leipner et al 2001) EPRmeasurements also showed increases in free radicals whichReichenauer et al (1998) suggested might contribute todamage of chloroplast membranes in the sensitive wheatlines Runeckles amp Vaartnou (1997) also observed light-dependent EPR signals in bluegrass (Poa pratensis L)ryegrass (Lolium perenne L) and radish (Raphanus sativaL) consistent with superoxide anion characteristics onexposure to O3 The light dependence suggested that thesignal was localized in the chloroplast and significantly itsappearance was reduced in leaves with increased levels ofapoplastic ascorbic acid
High light is another factor that might interact with O3
in such a way as to increase or aggravate the effects of O3
alone through the process of photoinhibition with a result-
Figure 2 Rubisco quantity and activity An and chlorophyll content ratios for soybean (cv Essex) leaves treated with charcoal-filtered air (CF) 25 nmol O3 mol-1 and 15 yen ambient O3 (75 nmol mol-1 12 h average) in open-top field chambers in 1994 The ratios represent the response to O3 and are calculated as the quantity present in the O3 treatment compared with the control (O3CF) Developmental stages are given in the bar above the graph Data are from Reid et al (1998) and Reid amp Fiscus (1998)
1004 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
ant increase in free radicals in the chloroplast (Massman2004) Guidi et al (2002) demonstrated that both high lightand O3 decreased FvFm and increased Fo Furthermore thecombined effects were additive Farage et al (1991) uponexposing wheat leaves to high [O3] (200 and 400 nmolmol-1) could detect no difference in atrazine binding inisolated thylakoids nor any difference in D1 protein asindicated by western blots using D1-specific antibodiesHowever Godde amp Buchhold (1992) found that the turn-over rate for D1 was increased by exposure of spruce nee-dles to O3 suggesting both increased degradation andsynthesis In addition Ciompi et al (1997) found that inyoung leaves of pumpkin engagement of the xanthophyllcycle appeared to counteract the effects of O3 on the PSIIreaction center preventing a decline in D1 content How-ever in mature leaves even though there was activation ofthe xanthophyll cycle the effect appeared inadequate toprevent photoinhibition and loss of D1 protein The authorssuggested that this effect in mature leaves may have beendue to the normal decline in antioxidant activity in olderleaves Clearly high light has the potential to induce photo-inhibition which could be already sensitized by O3 exposurein field crops or inversely and is an area that requiresfurther investigation especially under more realistic fieldconditions
Direct effects on stomata
Another possible point of attack by O3 that cannot beignored is the guard cells Although much of the evidenceto date indicates that reduced gs is the result and not thecause of reduced assimilation (McKee et al 1995 Fiscuset al 1997 Long amp Naidu 2002) as suggested anddeduced on the basis of water use efficiency studies(Reich Schoettle amp Amundsen 1985) there is someintriguing work being done on possible direct effects ofO3 on stomatal response which links oxidative stress inthe form of H2O2 Ca2+ homeostasis of guard cells andABA response This work was recently reviewed byMcAinsh et al (2002) while the review by Schroeder et al(2001) provides a more detailed context for these studiesThe studies showed that H2O2 stimulated increased wholeplant cytosolic free Ca ions [Ca2+]cyt The stimulation washighly dose dependent with biphasic kinetics similar tothe Ca signature induced by exposure to O3 which wasalso highly dose dependent Using various techniquesincreased [Ca2+]cyt was also demonstrated in guard cells inresponse to both O3 and H2O2 In addition Pei et al(2000) demonstrated H2O2-induced stomatal closure andthat ABA induces H2O2 production in guard cells andactivates plasma membrane Ca-permeable channels Allof these responses were disrupted in the ABA-insensitiveArabidopsis mutant gca2 Taken together these and otherstudies reviewed (McAinsh et al 2002) raise the possibil-ity that O3 might directly affect stomata through theaction of H2O2 formed when O3 enters solution on theCa-permeable channels in the guard cell plasma mem-branes
O3ndashCO2 interactions on biomass and yield
Along with O3 atmospheric CO2 concentrations are alsorising and numerous studies indicate that elevated CO2
might enhance the productivity of current cropping sys-tems Long et al (2004) have shown that the magnitude ofthe response to elevated CO2 is highly variable not onlyacross but within species and cultivars Although thereappears to be a genetic component to this variability someof it may be due to environmental stresses that occurredduring plant growth Ozone is one such possible stress andit would not be surprising that there are significant physio-logical interactions between elevated CO2 and O3 The mostrelevant interaction for present purposes is the reductionin gs at elevated levels of CO2 that has the effect of prevent-ing reductions of An growth and yield in many crops grownin the presence of toxic levels of O3 (Fiscus et al 1997Olszyk et al 2000 Fiscus et al 2002 Morgan et al 2003Booker et al 2005) Figure 3 illustrates this interaction insoybean grown in both elevated CO2 and O3 (compare withAn ratio in Fig 2) and although it is expected that theassimilation ratios in elevated CO2 are substantially higherthan the control it is interesting to note that elevated CO2
did not seem to prevent the accelerated senescence charac-teristic of an O3 fumigation at current levels of CO2 Anal-ysis of compiled shoot biomass and yield data (Fiscus et al2002) show that increased An does get translated into anaverage 30 increase in biomass but often not into yieldin clean (charcoal-filtered) air (Fig 4) (Fiscus et al 2001)The variability in both biomass and yield is large with yieldsin doubled CO2 ranging from -37 to +41 in CF air witha mean of +6 across the crops studied However inincreasing O3 concentrations both the apparent biomassand yield due to elevated CO2 (CO2 fertilization effect)increased dramatically but the variability of the responsesremained large Of course in elevated CO2 gs will bereduced typically by 30 in these studies so O3 flux intothe leaves and the effective exposure is much reduced Thusthe apparent dramatic yield increases in some non-filteredair experiments might be caused in part by the fact thatyields are already suppressed by 5ndash15 by O3 in ambientair control plants and the best that elevated CO2 seemedto do in these experiments was to restore yields to the cleanair levels The wide range of variability in these experimentssuggests that in some CO2 was not the limiting resourceduring reproduction In fact it seems possible given thelarge increases in vegetative biomass that early resourcedepletion under elevated CO2 actually may have reducedyield One would expect that any environmental stress thataffects gs (eg drought) would also influence the directeffects of O3 and lead to large apparent CO2-stimulatedincreases in growth and yield The cause of the distinctvariability shown in these experiments is an area thatdeserves further research
Impacts on translocation and partitioning
Early in the season chronic exposure to O3 can inhibit An
(Fig 2) and plant growth (Fig 5) but often the decline in
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005
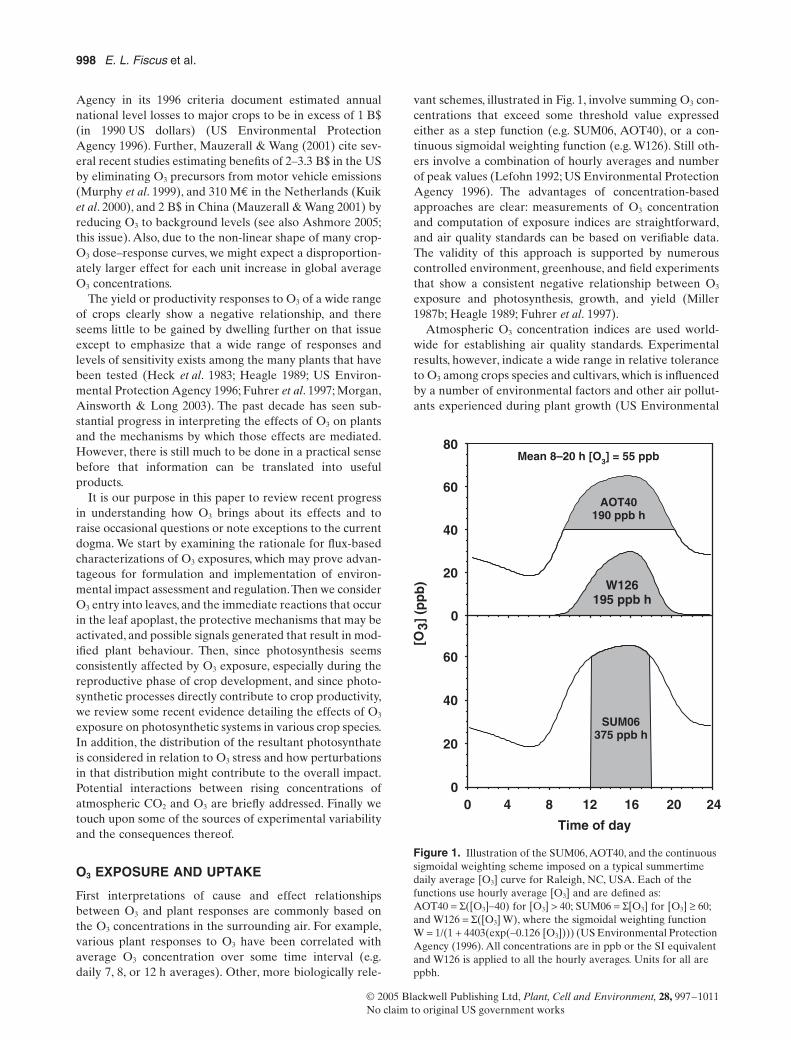
998
E L Fiscus
et al
copy 2005 Blackwell Publishing Ltd
Plant Cell and Environment
28
997ndash1011No claim to original US government works
Agency in its 1996 criteria document estimated annualnational level losses to major crops to be in excess of 1 B$(in 1990 US dollars) (US Environmental ProtectionAgency 1996) Further Mauzerall amp Wang (2001) cite sev-eral recent studies estimating benefits of 2ndash33 B$ in the USby eliminating O
3
precursors from motor vehicle emissions(Murphy
et al
1999) and 310 M
euro
in the Netherlands (Kuik
et al
2000) and 2 B$ in China (Mauzerall amp Wang 2001) byreducing O
3
to background levels (see also Ashmore 2005this issue) Also due to the non-linear shape of many crop-O
3
dosendashresponse curves we might expect a disproportion-ately larger effect for each unit increase in global averageO
3
concentrationsThe yield or productivity responses to O
3
of a wide rangeof crops clearly show a negative relationship and thereseems little to be gained by dwelling further on that issueexcept to emphasize that a wide range of responses andlevels of sensitivity exists among the many plants that havebeen tested (Heck
et al
1983 Heagle 1989 US Environ-mental Protection Agency 1996 Fuhrer
et al
1997 MorganAinsworth amp Long 2003) The past decade has seen sub-stantial progress in interpreting the effects of O
3
on plantsand the mechanisms by which those effects are mediatedHowever there is still much to be done in a practical sensebefore that information can be translated into usefulproducts
It is our purpose in this paper to review recent progressin understanding how O
3
brings about its effects and toraise occasional questions or note exceptions to the currentdogma We start by examining the rationale for flux-basedcharacterizations of O
3
exposures which may prove advan-tageous for formulation and implementation of environ-mental impact assessment and regulation Then we considerO
3
entry into leaves and the immediate reactions that occurin the leaf apoplast the protective mechanisms that may beactivated and possible signals generated that result in mod-ified plant behaviour Then since photosynthesis seemsconsistently affected by O
3
exposure especially during thereproductive phase of crop development and since photo-synthetic processes directly contribute to crop productivitywe review some recent evidence detailing the effects of O
3
exposure on photosynthetic systems in various crop speciesIn addition the distribution of the resultant photosynthateis considered in relation to O
3
stress and how perturbationsin that distribution might contribute to the overall impactPotential interactions between rising concentrations ofatmospheric CO
2
and O
3
are briefly addressed Finally wetouch upon some of the sources of experimental variabilityand the consequences thereof
O
3
EXPOSURE AND UPTAKE
First interpretations of cause and effect relationshipsbetween O
3
and plant responses are commonly based onthe O
3
concentrations in the surrounding air For examplevarious plant responses to O
3
have been correlated withaverage O
3
concentration over some time interval (egdaily 7 8 or 12 h averages) Other more biologically rele-
vant schemes illustrated in Fig 1 involve summing O
3
con-centrations that exceed some threshold value expressedeither as a step function (eg SUM06 AOT40) or a con-tinuous sigmoidal weighting function (eg W126) Still oth-ers involve a combination of hourly averages and numberof peak values (Lefohn 1992 US Environmental ProtectionAgency 1996) The advantages of concentration-basedapproaches are clear measurements of O
3
concentrationand computation of exposure indices are straightforwardand air quality standards can be based on verifiable dataThe validity of this approach is supported by numerouscontrolled environment greenhouse and field experimentsthat show a consistent negative relationship between O
3
exposure and photosynthesis growth and yield (Miller1987b Heagle 1989 Fuhrer
et al
1997)Atmospheric O
3
concentration indices are used world-wide for establishing air quality standards Experimentalresults however indicate a wide range in relative toleranceto O
3
among crops species and cultivars which is influencedby a number of environmental factors and other air pollut-ants experienced during plant growth (US Environmental
Figure 1
Illustration of the SUM06 AOT40 and the continuous sigmoidal weighting scheme imposed on a typical summertime daily average [O
3
] curve for Raleigh NC USA Each of the functions use hourly average [O
3
] and are defined as AOT40
=
S
([O
3
]
-
40) for [O
3
]
gt
40 SUM06
=
S
[O
3
] for [O
3
]
ge
60 and W126
=
S
([O
3
] W) where the sigmoidal weighting function W
=
1(1
+
4403(exp(
-
0126 [O
3
]))) (US Environmental Protection Agency (1996) All concentrations are in ppb or the SI equivalent and W126 is applied to all the hourly averages Units for all are ppbh
Time of day
0 4 8 12 16 20 24
[O3]
(p
pb
)
0
20
40
60
0
20
40
60
80
AOT40190 ppb h
W126195 ppb h
Mean 8ndash20 h [O3] = 55 ppb
SUM06375 ppb h
Crop responses to ozone
999
copy 2005 Blackwell Publishing Ltd
Plant Cell and Environment
28
997ndash1011No claim to original US government works
Protection Agency 1996 Fuhrer
et al
1997) In addition theO
3
concentration at measuring height (3ndash10 m) is usuallynot corrected for the lower concentration received at can-opy height (Tuovinen 2000) However a protocol for mak-ing this correction has been devised (UNECE 2004b)
Efforts to understand the discrepancies between O
3
con-centration in the air and variable plant responses havefocused on molecular and biochemical mechanismsinvolved in O
3
detoxification processes and certain interact-ing environmental factors affecting stomatal conductance(
g
s
) (Fuhrer
et al
1997 Massman 2004 Pleijel
et al
2004)Environmental factors influencing
g
s
are important becauseO
3
uptake mainly occurs through leaf stomata (Runeckles1992 Long amp Naidu 2002) Various modelling results haveindicated that a major factor affecting plant responses toO
3
appears to be
g
s
(Runeckles 1992 Massman 2004 Pleijel
et al
2004) This is supported by experiments with a varietyof crops exposed simultaneously to O
3
and elevated atmo-spheric CO
2
an effective antitranspirant that showedreduced O
3
injury presumably due in large part to loweredO
3
flux (McKee Farage amp Long 1995 Fiscus
et al
1997Reid amp Fiscus 1998 Reid Fiscus amp Burkey 1998 Heagle
et al
1999 Reid Fiscus amp Burkey 1999 Olszyk
et al
2000Fiscus
et al
2002 Morgan
et al
2003 Booker Fiscus ampMiller 2004 Booker
et al
2005) Reduced O
3
damage towater-stressed soybean [
Glycine max
(L) Merr] comparedwith well-watered plants was attributed to lower g
s
as well(Tingey amp Hogsett 1985 Vozzo
et al
1995)The major environmental factors controlling
g
s
includeleaf temperature water vapour pressure differencebetween the leaf and the surrounding air (VPD) photosyn-thetic photon flux density (PPFD) soil water availabilityand atmospheric CO
2
concentration (Jarvis 1976) Thusresponses to some O
3
concentration for a particular cropand cultivar undoubtedly depend upon inherent geneticdifferences and environmental conditions experienced dur-ing the growing season In order to incorporate the effectsof environmental conditions relevant to
g
s
into an exposureindex the UNECE ICP Vegetation project is investigatingthe use of VPD estimates in conjunction with O
3
concen-tration for defining the critical O
3
level that produces visibleinjury in crops (UNECE 2004b) Stomatal conductance istypically higher in warm humid environments comparedwith hot dry conditions Thus crops in warm humid envi-ronments are likely at increased risk of O
3
injury Thisapproach holds promise as a relatively simple method forincluding environmental factors that influence g
s
into O
3
exposure indices However other factors such as leaf tem-perature leaf hydration or PPFD (Massman 2004) couldindependently affect O
3
toxicity which would require addi-tional modifications of such an index
From a toxicological viewpoint it is the absorbed cumu-lative dose of O
3
that is most relevant in determining causeand effect relationships and quantifying dosendashresponsesThus O
3
flux models using a multiplicative algorithm basedon the approach of Jarvis (1976) to model
g
s
have incorpo-rated VPD air temperature PPFD and soil moisture deficitcoefficients to compute estimates of O
3
uptake (Emberson
et al
2000 Pleijel
et al
2004) A phenological componentalso has been added to account for changes in
g
s
during thegrowing season This approach indicated that phenologyand VPD were important factors in estimating O
3
fluxComparisons between flux estimates and O
3
exposure mapsof Europe suggested that areas with the highest O
3
concen-trations differed from those regions calculated as havingthe highest O
3
fluxes (Emberson
et al
2000) The highestO
3
concentrations occurred in the southern Mediterraneanregion whereas the highest O
3
fluxes for wheat occurred insouthern Scandinavia and northern Europe
However some crops and cultivars with similar gs havedifferent tolerances to the same O3 concentrations Thus ithas been suggested that O3 flux models include a coefficientfor O3 detoxification capacity (Musselman amp Massman1999 Fuhrer amp Booker 2003 Massman 2004) Musselmanamp Massman (1999) defined the lsquoeffective fluxrsquo as the bal-ance between uptake into the leaf at a given time and thedefence response at that time The defence response factorwas proportional to the effect of O3 on gross photosynthe-sis Effective flux integrated over time yields cumulativeeffective loading Using a relatively comprehensive model-ling approach for O3 deposition and uptake Massman(2004) estimated that the period of highest effective loadingoccurred between 1300 and 1600 h
The cumulative exposure indices (SUM06 W126AOT40) are based on O3 injury thresholds for effects of O3
concentration on plant growth and yield In the same man-ner cumulative O3 flux models show improved perfor-mance when a threshold O3 flux factor is included in themodel eg Pleijel et al (2004) Likewise Martin et al(2000) modified WIMOVAC (Windows Intuitive Model ofVegetation Response to Atmosphere and Climate Change)(Humphries amp Long 1995) to include O3 detoxification pro-cesses when calculating threshold flux-based rather thanconcentration-based dosendashresponse models Howeverdetermination of the threshold coefficient in the Martinet al (2000) model requires experimentally derived assess-ments of O3 effects on Vcmax The situation is a bit morecomplicated because O3 can damage plant photosynthesiswhich in turn may reduce the plantrsquos detoxification ability(Massman 2004) In effect there are interacting mecha-nisms at work that may alter O3 injury thresholds as dam-age accumulates (Massman 2004) Ozone increases therelative cost of detoxification on the one hand anddecreases the need through Ci-driven reductions in gs onthe other A flux-based approach that includes thresholdsrequires further experimentation and advances in ourunderstanding of fundamental processes involved in O3
action among different plant species Furthermore thisapproach needs to be scaled to the canopy level to begenerally applicable Micrometeorological soil-vegetation-atmosphere transfer (SVAT) models have been developedthat estimate stomatal O3 flux and cuticular depositionbased on O3 concentration atmospheric boundary layerand bulk canopy resistances to describe the verticalexchange of O3 to the plant-soil system (Gruumlnhage Haenelamp Jaumlger 2000)
1000 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
In the field canopy structure and foliage density affectthe concentration of O3 that leaves experience while envi-ronmental factors such as PPFD leaf temperature and VPDvary within the leaf canopy and can affect gs and thus O3
flux However after canopy closure for crops such as soy-bean cotton (Gossypium hirsutum L) and peanut (Ara-chis hypogaea L) leaves at the top of the canopy providethe majority of photosynthate for seed production Ozoneflux to the upper portion of the canopy is likely most criticalto impacts on yield For crops such as wheat (Triticum aes-tivum L) and rice (Oryza sativa L) flux to the entirecanopy might be important Efforts to parameterize canopyO3 flux have included leaf area index as a way to estimatecanopy gs (Massman 2004) which may be effective inaccounting for differences among crop canopy structuresHowever the ICP Vegetation protocol utilizes projectedleaf area namely leaf area of sunlit leaves at the top of thecanopy to calculate O3 flux (Pleijel et al 2004 UNECE2004b) Ozone flux to the most productive leaves is viewedas having the greatest impact on yield because even thoughthe soil and lower parts of the canopy consume largeamounts of O3 it is likely these fluxes have much less effecton yield (Pleijel et al 2004 Ashmore 2005)
Lastly the ICP Vegetation program (UNECE 2004a) iscurrently comparing economic crop loss assessments ofconcentration-based and flux-based methodologies Thisanalysis should suggest whether the flux approach providesa significantly better assessment of crop losses due to O3
pollution compared with concentration-based exposureindices
MODES OF ACTION
Apoplastic and symplastic reactions and responses
The internal air spaces within the leaf are a potential sitefor O3 reactions with volatile compounds produced by theplant Isoprene is one example of a compound emitted frommany angiosperms and conifers (Sharkey amp Yeh 2001) thatcould react with O3 However volatile compounds releasedby crop species are mainly monoterpenes which have beenlittle studied in this context although parallels betweenthese volatile organic compounds (VOCs) and O3 reactionswith isoprene are likely At present hypotheses are contra-dictory regarding the role of isoprene in plant responses toO3 Initially isoprene was thought to enhance O3 damagethrough the formation of hydroperoxides (Hewitt Kok ampRall 1990) More recently isoprene was found to exert aprotective effect either through action as a radical scaven-ger or membrane stabilizer (Loreto amp Velikova 2001) Theconcept of isoprene as an antioxidant is intriguing from theperspective of O3 scavenging reactions outside the leaf thatcould increase boundary layer resistance to O3 and effec-tively reduce the dose The potential benefits of such amechanism must be balanced against the metabolicdemand for carbon to support isoprene synthesis (Sharkeyamp Yeh 2001) and the consequences of increased plant-
derived VOCs on atmospheric chemistry leading to O3
formationOzone that has passed through leaf internal air spaces
will then dissolve in the aqueous layer surrounding leafcells The breakdown of O3 in pure water produces hydroxyland peroxyl radicals and superoxide although the reactionsproceed very slowly at neutral pH (Heath 1987) GrimesPerkins amp Boss (1983) demonstrated that the rate ofhydroxyl radical formation from O3 in aqueous solution issignificantly greater when phenolic compounds are presentsuggesting that biologically relevant compounds enhanceO3 reactions Based on in vitro and in vivo studies (Heath1987 Runeckles amp Chevone 1992 Pryor 1994 Kirichenkoet al 1996) O3 is known to react with a diverse set ofmolecules that would be encountered within the cell walland on the plasma membrane surface Initial targets for O3
include plasma membrane lipids susceptible amino acids inplasma membrane proteins or apoplastic enzymes and avariety of organic metabolites localized in the cell wallPotentially ozonolysis products from the oxidation of lipidsand metabolites containing carbon-carbon double bonds(Heath 1987 Runeckles amp Chevone 1992) and alteredplasma membrane protein function (Dominy amp Heath1985 Castillo amp Heath 1990) could serve as the initial sig-nals leading to O3 responses However the primary O3
reaction products in plants have not been identified due todifficulties in following O3 reactions within the complexbiochemical network of the leaf apoplast Knowledge ofthese early events is critical to understanding the metabolicsignals that initiate O3 responses and may provide insightinto O3 tolerance mechanisms localized in the leaf apoplastOzone incorporation into the cells and extracellular lipid-protein complexes of rat lung tissue has been studied usinga combination of 18O3 and mass spectrometry (Gunnison ampHatch 1999) an approach that could be employed in con-junction with available methods for isolation of leaf apo-plast components and plasma membranes to addressquestions regarding the initial O3 reaction products inplants
Formation of reactive oxygen species (ROS) such assuperoxide hydrogen peroxide and hydroxyl radical arethought to be associated with the initial breakdown of O3
in the leaf apoplast ESR approaches based on spin trap-ping suggest that free radicals are involved in the earlystages of O3 response (Grimes et al 1983 Mehlhorn Tab-ner amp Wellburn 1990 Runeckles amp Vaartnou 1997) but itis not clear whether the observed signals originate fromdirect O3 breakdown or the initial reactions with cellularconstituents such as phenolic compounds Localizedregions of ROS formation either hydrogen peroxide orsuperoxide depending on the plant species are associatedwith O3 response in sensitive plants (Wohlgemuth et al2002 Pendall et al 2004) The kinetics of ROS formation inthe leaf apoplast following O3 exposure seem to involve twophases an initial phase that is associated with direct effectsof O3 and a second phase associated with a plant-derivedsecondary oxidative burst (Schraudner et al 1998) The sec-ondary oxidative burst is initially localized in the leaf apo-
Crop responses to ozone 1001
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
plast and cell wall but later expands into the cytoplasm andsubcellular compartments leading to the formation of visi-ble lesions (Pendall et al 2004)
The initial signals induced by O3 in the leaf apoplastmight then be translated into responses at the tissue levelResponses vary and include unregulated cell death hyper-sensitive response leading to programmed cell death andaccelerated senescence (Pell Schlagnhaufer amp Arteca 1997Sandermann et al 1998) Evidence is accumulating thatthese responses are modulated by ethylene jasmonic andsalicylic acid levels and the interactions among their signal-ling pathways (Kangasjaumlrvi et al 1994 Sharma et al 1996Pell et al 1997 Overmyer et al 2000 Rao et al 2000 Raoamp Davis 2001 Rao Lee amp Davis 2002 Overmyer Broscheamp Kangasjaumlrvi 2003 Tamaoki et al 2003 Vahala et al 2003see also Kangasjaumlrvi et al 2005 this issue) For exampleArabidopsis thaliana mutants that overproduce ethylene(eto1) or are insensitive to jasmonic acid (jar1) are moreextensively injured by O3 than wild-type plants (Overmyeret al 2000 Rao et al 2000 Tamaoki et al 2003) Ethyleneacts as a promoting factor for O3 injury whereas jasmonatesmight have a role in minimizing injury (Overmyer et al2000 2003)
Ozone effects at the tissue level are often characterizedas either acute or chronic responses subjective terms thatare related to the type of visible symptoms observed andwhether the O3 concentrations employed typify ambient O3
episodes For example unregulated cell death and pro-grammed cell death are usually considered acute responsesin which lesions occur within hours after exposure to rela-tively high O3 concentrations (typically gt150 nmol mol-1)In contrast chronic responses include lesions that developover days to weeks under lower O3 concentrations andaccelerated senescence where lesions might not formUnder field conditions accelerated senescence may be dif-ficult to identify in the absence of a clean air control orother reference point (eg O3-tolerant cultivar) because thecumulative effects on foliar senescence at the end of theseason are separated in time from the O3 events causingthe effects The timing of O3 episodes in relation to plantdevelopment can also have a major effect on the outcomeas in the case of soybean in which O3 exposure duringreproductive growth has a much greater impact on yieldcompared with exposure during vegetative growth (Heagleet al 1991 Miller et al 1991 Morgan et al 2003) The dis-tinction between acute and chronic responses is furthercomplicated by species and genotype differences that leadto a range of effects for a given combination of O3 concen-tration and environmental conditions
Genetic variation in O3 responses and potential O3 tolerance mechanisms
Studies of O3 effects on plants often identify genetic varia-tion as a contributing factor in the observed responseOzone-sensitive and tolerant cultivars or clones have beenreported for many plant species (Wellburn amp Wellburn1996) including crops such as soybean (Tingey Reinert amp
Carter 1972 Heagle amp Letchworth 1982 Robinson amp Britz2000) snap bean (Phaseolus vulgaris L) (Guzy amp Heath1993) tomato (Lycopersicon esculentum Miller) (Temple1990) clover (Trifolium repens L) (Heagle et al 1993)wheat (Heagle Miller amp Pursley 2000) and potato(Solanum tuberosum L) (Heagle Miller amp Pursley 2003)and natural vegetation such as black cherry (Prunus sero-tina Ehrh) (Lee et al 2002) and Plantago spp (Wolff Mor-gan-Richards amp Davison 2000) Studies have also shownthat O3 tolerance is a heritable trait (Damicone amp Manning1987 Reinert amp Eason 2000) Although much effort hasbeen made to identify the physiological and biochemicalelements of O3 tolerance no clear picture has emergedBased on the oxidative stress model formation of ROS andother oxidation products is the underlying process involvedin the generation and propagation of toxic compounds andabiotic elicitors in plants Therefore features of antioxidantmetabolism that scavenge or affect the perception of O3-derived ROS could contribute to O3 tolerance
Plants have evolved elaborate systems to combat generaloxidative stress based on enzymes that utilize ascorbic acidand glutathione (Noctor amp Foyer 1998 see also Foyer ampNoctor 2005 this issue) and these antioxidant systems havebeen linked to O3 stress tolerance (Runeckles amp Chevone1992 Smirnoff 1996 Chernikova et al 2000 Conklin ampBarth 2004) A critical role for ascorbic acid is demon-strated with the vtc1 mutant of Arabidopsis in which asignificant decrease in ascorbate content was associatedwith increased O3 sensitivity (Conklin Williams amp Last1996) A more detailed analysis of vtc1 suggested that inaddition to acting as an antioxidant ascorbic acid serves aregulatory function in the signalling networks that controlplant defence responses and leaf senescence (Conklin ampBarth 2004) Although it is clear that minimum levels ofascorbic acid and glutathione are required for normal leaffunction the concentrations of these antioxidant metabo-lites that naturally occur in leaf tissue are not always wellcorrelated with O3 tolerance (Guzy amp Heath 1993 Well-burn amp Wellburn 1996 Burkey et al 2000) suggesting amore complex mechanism that remains to be elucidated
Recent studies have found higher levels of ascorbic acidin the leaf apoplast of certain O3-tolerant snap bean geno-types compared with sensitive lines (Burkey Eason amp Fis-cus 2003) suggesting that localization of ascorbic acid inthe cell wall where initial O3 reactions occur might beimportant Leaf extracellular ascorbate content and redoxstatus are affected by O3 treatment (Castillo amp Greppin1988 Luwe amp Heber 1995 Burkey 1999) evidence thatextracellular ascorbic acid is involved in detoxification ofO3 and related ROS Ascorbic acid is initially synthesizedinside cells (Smirnoff Conklin amp Loewus 2001) followedby transport between the cytoplasm and extracellular spacevia specific carriers located in the plasma membrane (Hore-mans Foyer amp Asard 2000) A model developed by Cha-meides (1989) and expanded by others (Ploumlchl et al 2000Turcsanyi et al 2000) features ascorbic acid in the leaf apo-plast as a critical factor in the detoxification of extracellularO3 and ROS protecting plasma membranes from oxidative
1002 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
damage and preventing O3 injury Ascorbic acid potentiallycould serve as either a direct chemical scavenger of O3although this mechanism has been questioned (Jakob ampHeber 1998) or as a substrate for extracellular enzymes(eg ascorbate peroxidase) that attenuate ROS levels andthus affect the propagation of the initial O3 signal How-ever leaf apoplast ascorbic acid levels can be quite low incertain plant species [summarized in Burkey et al (2003)]suggesting that extracellular scavenging of O3 and ROS byascorbate may not be a significant factor in all plants Othercompounds with antioxidant properties also exist in the leafapoplast Identification of these compounds and theirimpact on ROS formation and propagation are subjects forfuture research on O3 tolerance mechanisms
Another approach to understanding O3 responses inplants involves investigations of the potential interactionbetween chemical signals and specific receptors in theplasma membrane Booker et al (2004) showed that theepinasty frequently observed for wild-type Arabidopsisleaves after O3 treatment did not occur in gpa1 mutantplants lacking the alpha subunit of the heterotrimeric Gprotein complex suggesting that G-proteins may beinvolved in the transduction of O3-derived signals Thebinding of O3 or more likely one of the postulated O3 reac-tion products to a G-protein receptor could activate theprotein and initiate signal cascade processes that activatetarget proteins in this signal transduction system Howeverfurther studies will be required to define the possible roleG-proteins have in O3-induced signal transduction andpropagation It is also possible that G-proteins modulate O3
responses via hormone signal transduction processes Ineither case elements of G-protein pathways involved in O3
response represent potential molecular targets that mightbe manipulated to enhance O3 tolerance although this isspeculative at present The challenge will be to identifytargets unique to O3 so that other important stressresponses remain intact (eg plant defence responsesagainst pathogen attack) Finally it is likely that any pro-tective mechanism will come with some metabolic cost thatwill require evaluation to determine its net benefits
IMPACTS ON PHOTOSYNTHESIS
Net assimilation
Other than crop yield and visible injury photosynthesis hasbeen the most broadly studied aspect of plant responses toO3 Generally O3 exposure results in decreased photosyn-thetic carbon assimilation (Miller 1987b Runeckles ampChevone 1992 Pell et al 1997 Long amp Naidu 2002 Morganet al 2003) In many cases loss of assimilation capacity wasshown to be due primarily to reduced carboxylation effi-ciency directly related to loss of Rubisco activity Theselosses of activity are thought to be due to decreases inRubisco concentration in the leaves rather than a decreasein activation state Messenger RNA transcripts for theRubisco small subunit (rbcS) have been noted to declinewith O3 treatment in potato leaves (Glick et al 1995) sug-
gesting that protein synthesis might be inhibited a notionconsistent with observations that Rubisco content in poplar(Populus spp) leaves treated with O3 does not rise as highas controls during leaf expansion (Pell et al 1997) How-ever Eckardt amp Pell (1994) under conditions where addi-tional synthesis should be minimal showed a substantialdecline in Rubisco content suggesting that the primarycause of the decline in Rubisco due to O3 exposure wasenhanced degradation rather than reduced production
Because photosynthetic measurements are typicallymade on the most recently fully expanded upper canopyleaves in determinate plants including many soybean linesit is often difficult to demonstrate differences in net photo-synthesis (An) due to O3 until near or after the time whenproduction of new upper canopy leaves has ceased Thenduring most of the reproductive stages An decreased earlierthan in control plants (accelerated senescence) This sug-gests that during vegetative growth either Rubisco synthe-sis in newly formed upper canopy leaves is not substantiallyaffected or that these leaves have not accumulated suffi-cient damage to elicit a response Vozzo et al (1995) dem-onstrated significant reductions in An on somewhat oldersoybean leaves that were relatively unaffected by shadingbut had received sufficient exposure to exhibit visible symp-toms thus lending support to this idea In contrast Morganet al (2004) using an indeterminate line were unable todemonstrate any differences due to dose accumulation in acohort of soybean leaves that developed during vegetativegrowth However they did observe decreased An in anothercohort that developed during reproductive growth andremained near the top of the canopy These results couldwell have been due to differences in exposure methodolo-gies particularly the greater atmospheric coupling to thelower canopy in OTCs as compared to FACE systems Infurther contrast to the above observations additionalreview of the data in two earlier soybean studies (Reid ampFiscus 1998 Reid et al 1998) illustrated in Fig 2 showsthat Rubisco content and activity chlorophyll content andAn were reduced by O3 early in the vegetative stage andthat all but An had reached or exceeded clean air levels byearly flowering Afterwards while chlorophyll content Anand Rubisco activity remained level or declined slightlyRubisco content continued to rise in the O3 treatmentFinally in early seed development all these parametersbegan to decline more precipitously Throughout this wholeexperiment chlorophyll content and Rubisco activitytracked each other very well This suggests that from earlyflowering through most of seed development O3 stimulatedRubisco content on a leaf area basis In addition the ques-tion of the role of accelerated senescence especially duringreproductive growth is one deserving more attention Forexample in Fig 2 the O3-induced decline in An duringreproduction may be due to the normal process of senes-cence that has been accelerated by O3 exposure or it maybe due simply to the loss of photosynthetically active leafarea due to accumulated O3 damage Thus especially inareas associated with visible lesions one would measure amixture of live and dead or dying cells and it seems rele-
Crop responses to ozone 1003
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
vant to know whether the remaining live cells are function-ing at or near normal levels for their age This possibility issuggested by the fact that from beginning of pod fill throughseed maturation when An per unit area is declining in thisstudy An per unit Rubisco remained high at 91 of the CFcontrol (difference not significant and data not shown)Further information is necessary to clarify this Anotherpoint deserving attention and bearing on the exposureaccumulation comments earlier in the paragraph is the sub-stantially reduced An early in the vegetative period Thismay have been caused by some inherent sensitivity of theseleaves or be the result of enhanced atmospheric couplingdue to the more open nature of the canopy at this growthstage
Photosynthetic electron transport
Detrimental effects of O3 on photosynthetic electron trans-port has received increased attention in recent years andwhile earlier it had been difficult to demonstrate significantdeclines in function evidence is accumulating that in someplants there are demonstrable effects both on PSII func-tion as well as the xanthophyll cycle components Forinstance significant and substantial declines in FvFm whichrepresents the efficiency of excitation energy capture byPSII in dark-adapted leaves have been reported for pump-kin (Cucurbita pepo L) (Ciompi et al 1997 Castagna et al2001) wheat (T durum and T aestivum) (Reichenauer et al1998) bean (P vulgaris L) (Guidi Di Cagno amp Soladatini2000a Guidi Tonini amp Soladatini 2000b Leipner Oxbor-ough amp Baker 2001 Guidi DeglrsquoInnocenti amp Soldatini2002) tomato (Calatayud amp Barreno 2001) and Festucapratensis and turnip (Brassica napus L) (Plazek Rapacz ampSkoczowski 2000) Of these particular studies three dem-
onstrated an increase in Fo which suggested damaged ordeactivated PSII centers (Guidi et al 2000a Leipner et al2001 Guidi et al 2002) all occurring in bean In additionlight saturated photosynthesis (ASAT) decreased in bean(Guidi et al 2000a Guidi et al 2002) pumpkin (Ciompiet al 1997 Castagna et al 2001) and wheat (Reichenaueret al 1998) although in the latter the effect was confined toolder leaves in two of the three lines and was partiallyreversible in some lines and fully reversible overnight inone It was unknown whether reversibility was due to anight-time epoxidation of the xanthophyll cycle decreasingsubsequent thermal quenching of PSII or to overnightrepair of damaged PSII centres This latter point becomesrelevant since a decrease in FvFm may be caused by both arelatively fast reversible down-regulation or a reversibledestruction (Osmond 1994) However the two processesmight be distinguished by monitoring effects on Fo in thatthermal dissipation by the xanthophyll cycle would lowerFo whereas damage to PSII would increase Fo as was foundin bean (Guidi et al 2000a 2002 Leipner et al 2001) EPRmeasurements also showed increases in free radicals whichReichenauer et al (1998) suggested might contribute todamage of chloroplast membranes in the sensitive wheatlines Runeckles amp Vaartnou (1997) also observed light-dependent EPR signals in bluegrass (Poa pratensis L)ryegrass (Lolium perenne L) and radish (Raphanus sativaL) consistent with superoxide anion characteristics onexposure to O3 The light dependence suggested that thesignal was localized in the chloroplast and significantly itsappearance was reduced in leaves with increased levels ofapoplastic ascorbic acid
High light is another factor that might interact with O3
in such a way as to increase or aggravate the effects of O3
alone through the process of photoinhibition with a result-
Figure 2 Rubisco quantity and activity An and chlorophyll content ratios for soybean (cv Essex) leaves treated with charcoal-filtered air (CF) 25 nmol O3 mol-1 and 15 yen ambient O3 (75 nmol mol-1 12 h average) in open-top field chambers in 1994 The ratios represent the response to O3 and are calculated as the quantity present in the O3 treatment compared with the control (O3CF) Developmental stages are given in the bar above the graph Data are from Reid et al (1998) and Reid amp Fiscus (1998)
1004 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
ant increase in free radicals in the chloroplast (Massman2004) Guidi et al (2002) demonstrated that both high lightand O3 decreased FvFm and increased Fo Furthermore thecombined effects were additive Farage et al (1991) uponexposing wheat leaves to high [O3] (200 and 400 nmolmol-1) could detect no difference in atrazine binding inisolated thylakoids nor any difference in D1 protein asindicated by western blots using D1-specific antibodiesHowever Godde amp Buchhold (1992) found that the turn-over rate for D1 was increased by exposure of spruce nee-dles to O3 suggesting both increased degradation andsynthesis In addition Ciompi et al (1997) found that inyoung leaves of pumpkin engagement of the xanthophyllcycle appeared to counteract the effects of O3 on the PSIIreaction center preventing a decline in D1 content How-ever in mature leaves even though there was activation ofthe xanthophyll cycle the effect appeared inadequate toprevent photoinhibition and loss of D1 protein The authorssuggested that this effect in mature leaves may have beendue to the normal decline in antioxidant activity in olderleaves Clearly high light has the potential to induce photo-inhibition which could be already sensitized by O3 exposurein field crops or inversely and is an area that requiresfurther investigation especially under more realistic fieldconditions
Direct effects on stomata
Another possible point of attack by O3 that cannot beignored is the guard cells Although much of the evidenceto date indicates that reduced gs is the result and not thecause of reduced assimilation (McKee et al 1995 Fiscuset al 1997 Long amp Naidu 2002) as suggested anddeduced on the basis of water use efficiency studies(Reich Schoettle amp Amundsen 1985) there is someintriguing work being done on possible direct effects ofO3 on stomatal response which links oxidative stress inthe form of H2O2 Ca2+ homeostasis of guard cells andABA response This work was recently reviewed byMcAinsh et al (2002) while the review by Schroeder et al(2001) provides a more detailed context for these studiesThe studies showed that H2O2 stimulated increased wholeplant cytosolic free Ca ions [Ca2+]cyt The stimulation washighly dose dependent with biphasic kinetics similar tothe Ca signature induced by exposure to O3 which wasalso highly dose dependent Using various techniquesincreased [Ca2+]cyt was also demonstrated in guard cells inresponse to both O3 and H2O2 In addition Pei et al(2000) demonstrated H2O2-induced stomatal closure andthat ABA induces H2O2 production in guard cells andactivates plasma membrane Ca-permeable channels Allof these responses were disrupted in the ABA-insensitiveArabidopsis mutant gca2 Taken together these and otherstudies reviewed (McAinsh et al 2002) raise the possibil-ity that O3 might directly affect stomata through theaction of H2O2 formed when O3 enters solution on theCa-permeable channels in the guard cell plasma mem-branes
O3ndashCO2 interactions on biomass and yield
Along with O3 atmospheric CO2 concentrations are alsorising and numerous studies indicate that elevated CO2
might enhance the productivity of current cropping sys-tems Long et al (2004) have shown that the magnitude ofthe response to elevated CO2 is highly variable not onlyacross but within species and cultivars Although thereappears to be a genetic component to this variability someof it may be due to environmental stresses that occurredduring plant growth Ozone is one such possible stress andit would not be surprising that there are significant physio-logical interactions between elevated CO2 and O3 The mostrelevant interaction for present purposes is the reductionin gs at elevated levels of CO2 that has the effect of prevent-ing reductions of An growth and yield in many crops grownin the presence of toxic levels of O3 (Fiscus et al 1997Olszyk et al 2000 Fiscus et al 2002 Morgan et al 2003Booker et al 2005) Figure 3 illustrates this interaction insoybean grown in both elevated CO2 and O3 (compare withAn ratio in Fig 2) and although it is expected that theassimilation ratios in elevated CO2 are substantially higherthan the control it is interesting to note that elevated CO2
did not seem to prevent the accelerated senescence charac-teristic of an O3 fumigation at current levels of CO2 Anal-ysis of compiled shoot biomass and yield data (Fiscus et al2002) show that increased An does get translated into anaverage 30 increase in biomass but often not into yieldin clean (charcoal-filtered) air (Fig 4) (Fiscus et al 2001)The variability in both biomass and yield is large with yieldsin doubled CO2 ranging from -37 to +41 in CF air witha mean of +6 across the crops studied However inincreasing O3 concentrations both the apparent biomassand yield due to elevated CO2 (CO2 fertilization effect)increased dramatically but the variability of the responsesremained large Of course in elevated CO2 gs will bereduced typically by 30 in these studies so O3 flux intothe leaves and the effective exposure is much reduced Thusthe apparent dramatic yield increases in some non-filteredair experiments might be caused in part by the fact thatyields are already suppressed by 5ndash15 by O3 in ambientair control plants and the best that elevated CO2 seemedto do in these experiments was to restore yields to the cleanair levels The wide range of variability in these experimentssuggests that in some CO2 was not the limiting resourceduring reproduction In fact it seems possible given thelarge increases in vegetative biomass that early resourcedepletion under elevated CO2 actually may have reducedyield One would expect that any environmental stress thataffects gs (eg drought) would also influence the directeffects of O3 and lead to large apparent CO2-stimulatedincreases in growth and yield The cause of the distinctvariability shown in these experiments is an area thatdeserves further research
Impacts on translocation and partitioning
Early in the season chronic exposure to O3 can inhibit An
(Fig 2) and plant growth (Fig 5) but often the decline in
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

Crop responses to ozone
999
copy 2005 Blackwell Publishing Ltd
Plant Cell and Environment
28
997ndash1011No claim to original US government works
Protection Agency 1996 Fuhrer
et al
1997) In addition theO
3
concentration at measuring height (3ndash10 m) is usuallynot corrected for the lower concentration received at can-opy height (Tuovinen 2000) However a protocol for mak-ing this correction has been devised (UNECE 2004b)
Efforts to understand the discrepancies between O
3
con-centration in the air and variable plant responses havefocused on molecular and biochemical mechanismsinvolved in O
3
detoxification processes and certain interact-ing environmental factors affecting stomatal conductance(
g
s
) (Fuhrer
et al
1997 Massman 2004 Pleijel
et al
2004)Environmental factors influencing
g
s
are important becauseO
3
uptake mainly occurs through leaf stomata (Runeckles1992 Long amp Naidu 2002) Various modelling results haveindicated that a major factor affecting plant responses toO
3
appears to be
g
s
(Runeckles 1992 Massman 2004 Pleijel
et al
2004) This is supported by experiments with a varietyof crops exposed simultaneously to O
3
and elevated atmo-spheric CO
2
an effective antitranspirant that showedreduced O
3
injury presumably due in large part to loweredO
3
flux (McKee Farage amp Long 1995 Fiscus
et al
1997Reid amp Fiscus 1998 Reid Fiscus amp Burkey 1998 Heagle
et al
1999 Reid Fiscus amp Burkey 1999 Olszyk
et al
2000Fiscus
et al
2002 Morgan
et al
2003 Booker Fiscus ampMiller 2004 Booker
et al
2005) Reduced O
3
damage towater-stressed soybean [
Glycine max
(L) Merr] comparedwith well-watered plants was attributed to lower g
s
as well(Tingey amp Hogsett 1985 Vozzo
et al
1995)The major environmental factors controlling
g
s
includeleaf temperature water vapour pressure differencebetween the leaf and the surrounding air (VPD) photosyn-thetic photon flux density (PPFD) soil water availabilityand atmospheric CO
2
concentration (Jarvis 1976) Thusresponses to some O
3
concentration for a particular cropand cultivar undoubtedly depend upon inherent geneticdifferences and environmental conditions experienced dur-ing the growing season In order to incorporate the effectsof environmental conditions relevant to
g
s
into an exposureindex the UNECE ICP Vegetation project is investigatingthe use of VPD estimates in conjunction with O
3
concen-tration for defining the critical O
3
level that produces visibleinjury in crops (UNECE 2004b) Stomatal conductance istypically higher in warm humid environments comparedwith hot dry conditions Thus crops in warm humid envi-ronments are likely at increased risk of O
3
injury Thisapproach holds promise as a relatively simple method forincluding environmental factors that influence g
s
into O
3
exposure indices However other factors such as leaf tem-perature leaf hydration or PPFD (Massman 2004) couldindependently affect O
3
toxicity which would require addi-tional modifications of such an index
From a toxicological viewpoint it is the absorbed cumu-lative dose of O
3
that is most relevant in determining causeand effect relationships and quantifying dosendashresponsesThus O
3
flux models using a multiplicative algorithm basedon the approach of Jarvis (1976) to model
g
s
have incorpo-rated VPD air temperature PPFD and soil moisture deficitcoefficients to compute estimates of O
3
uptake (Emberson
et al
2000 Pleijel
et al
2004) A phenological componentalso has been added to account for changes in
g
s
during thegrowing season This approach indicated that phenologyand VPD were important factors in estimating O
3
fluxComparisons between flux estimates and O
3
exposure mapsof Europe suggested that areas with the highest O
3
concen-trations differed from those regions calculated as havingthe highest O
3
fluxes (Emberson
et al
2000) The highestO
3
concentrations occurred in the southern Mediterraneanregion whereas the highest O
3
fluxes for wheat occurred insouthern Scandinavia and northern Europe
However some crops and cultivars with similar gs havedifferent tolerances to the same O3 concentrations Thus ithas been suggested that O3 flux models include a coefficientfor O3 detoxification capacity (Musselman amp Massman1999 Fuhrer amp Booker 2003 Massman 2004) Musselmanamp Massman (1999) defined the lsquoeffective fluxrsquo as the bal-ance between uptake into the leaf at a given time and thedefence response at that time The defence response factorwas proportional to the effect of O3 on gross photosynthe-sis Effective flux integrated over time yields cumulativeeffective loading Using a relatively comprehensive model-ling approach for O3 deposition and uptake Massman(2004) estimated that the period of highest effective loadingoccurred between 1300 and 1600 h
The cumulative exposure indices (SUM06 W126AOT40) are based on O3 injury thresholds for effects of O3
concentration on plant growth and yield In the same man-ner cumulative O3 flux models show improved perfor-mance when a threshold O3 flux factor is included in themodel eg Pleijel et al (2004) Likewise Martin et al(2000) modified WIMOVAC (Windows Intuitive Model ofVegetation Response to Atmosphere and Climate Change)(Humphries amp Long 1995) to include O3 detoxification pro-cesses when calculating threshold flux-based rather thanconcentration-based dosendashresponse models Howeverdetermination of the threshold coefficient in the Martinet al (2000) model requires experimentally derived assess-ments of O3 effects on Vcmax The situation is a bit morecomplicated because O3 can damage plant photosynthesiswhich in turn may reduce the plantrsquos detoxification ability(Massman 2004) In effect there are interacting mecha-nisms at work that may alter O3 injury thresholds as dam-age accumulates (Massman 2004) Ozone increases therelative cost of detoxification on the one hand anddecreases the need through Ci-driven reductions in gs onthe other A flux-based approach that includes thresholdsrequires further experimentation and advances in ourunderstanding of fundamental processes involved in O3
action among different plant species Furthermore thisapproach needs to be scaled to the canopy level to begenerally applicable Micrometeorological soil-vegetation-atmosphere transfer (SVAT) models have been developedthat estimate stomatal O3 flux and cuticular depositionbased on O3 concentration atmospheric boundary layerand bulk canopy resistances to describe the verticalexchange of O3 to the plant-soil system (Gruumlnhage Haenelamp Jaumlger 2000)
1000 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
In the field canopy structure and foliage density affectthe concentration of O3 that leaves experience while envi-ronmental factors such as PPFD leaf temperature and VPDvary within the leaf canopy and can affect gs and thus O3
flux However after canopy closure for crops such as soy-bean cotton (Gossypium hirsutum L) and peanut (Ara-chis hypogaea L) leaves at the top of the canopy providethe majority of photosynthate for seed production Ozoneflux to the upper portion of the canopy is likely most criticalto impacts on yield For crops such as wheat (Triticum aes-tivum L) and rice (Oryza sativa L) flux to the entirecanopy might be important Efforts to parameterize canopyO3 flux have included leaf area index as a way to estimatecanopy gs (Massman 2004) which may be effective inaccounting for differences among crop canopy structuresHowever the ICP Vegetation protocol utilizes projectedleaf area namely leaf area of sunlit leaves at the top of thecanopy to calculate O3 flux (Pleijel et al 2004 UNECE2004b) Ozone flux to the most productive leaves is viewedas having the greatest impact on yield because even thoughthe soil and lower parts of the canopy consume largeamounts of O3 it is likely these fluxes have much less effecton yield (Pleijel et al 2004 Ashmore 2005)
Lastly the ICP Vegetation program (UNECE 2004a) iscurrently comparing economic crop loss assessments ofconcentration-based and flux-based methodologies Thisanalysis should suggest whether the flux approach providesa significantly better assessment of crop losses due to O3
pollution compared with concentration-based exposureindices
MODES OF ACTION
Apoplastic and symplastic reactions and responses
The internal air spaces within the leaf are a potential sitefor O3 reactions with volatile compounds produced by theplant Isoprene is one example of a compound emitted frommany angiosperms and conifers (Sharkey amp Yeh 2001) thatcould react with O3 However volatile compounds releasedby crop species are mainly monoterpenes which have beenlittle studied in this context although parallels betweenthese volatile organic compounds (VOCs) and O3 reactionswith isoprene are likely At present hypotheses are contra-dictory regarding the role of isoprene in plant responses toO3 Initially isoprene was thought to enhance O3 damagethrough the formation of hydroperoxides (Hewitt Kok ampRall 1990) More recently isoprene was found to exert aprotective effect either through action as a radical scaven-ger or membrane stabilizer (Loreto amp Velikova 2001) Theconcept of isoprene as an antioxidant is intriguing from theperspective of O3 scavenging reactions outside the leaf thatcould increase boundary layer resistance to O3 and effec-tively reduce the dose The potential benefits of such amechanism must be balanced against the metabolicdemand for carbon to support isoprene synthesis (Sharkeyamp Yeh 2001) and the consequences of increased plant-
derived VOCs on atmospheric chemistry leading to O3
formationOzone that has passed through leaf internal air spaces
will then dissolve in the aqueous layer surrounding leafcells The breakdown of O3 in pure water produces hydroxyland peroxyl radicals and superoxide although the reactionsproceed very slowly at neutral pH (Heath 1987) GrimesPerkins amp Boss (1983) demonstrated that the rate ofhydroxyl radical formation from O3 in aqueous solution issignificantly greater when phenolic compounds are presentsuggesting that biologically relevant compounds enhanceO3 reactions Based on in vitro and in vivo studies (Heath1987 Runeckles amp Chevone 1992 Pryor 1994 Kirichenkoet al 1996) O3 is known to react with a diverse set ofmolecules that would be encountered within the cell walland on the plasma membrane surface Initial targets for O3
include plasma membrane lipids susceptible amino acids inplasma membrane proteins or apoplastic enzymes and avariety of organic metabolites localized in the cell wallPotentially ozonolysis products from the oxidation of lipidsand metabolites containing carbon-carbon double bonds(Heath 1987 Runeckles amp Chevone 1992) and alteredplasma membrane protein function (Dominy amp Heath1985 Castillo amp Heath 1990) could serve as the initial sig-nals leading to O3 responses However the primary O3
reaction products in plants have not been identified due todifficulties in following O3 reactions within the complexbiochemical network of the leaf apoplast Knowledge ofthese early events is critical to understanding the metabolicsignals that initiate O3 responses and may provide insightinto O3 tolerance mechanisms localized in the leaf apoplastOzone incorporation into the cells and extracellular lipid-protein complexes of rat lung tissue has been studied usinga combination of 18O3 and mass spectrometry (Gunnison ampHatch 1999) an approach that could be employed in con-junction with available methods for isolation of leaf apo-plast components and plasma membranes to addressquestions regarding the initial O3 reaction products inplants
Formation of reactive oxygen species (ROS) such assuperoxide hydrogen peroxide and hydroxyl radical arethought to be associated with the initial breakdown of O3
in the leaf apoplast ESR approaches based on spin trap-ping suggest that free radicals are involved in the earlystages of O3 response (Grimes et al 1983 Mehlhorn Tab-ner amp Wellburn 1990 Runeckles amp Vaartnou 1997) but itis not clear whether the observed signals originate fromdirect O3 breakdown or the initial reactions with cellularconstituents such as phenolic compounds Localizedregions of ROS formation either hydrogen peroxide orsuperoxide depending on the plant species are associatedwith O3 response in sensitive plants (Wohlgemuth et al2002 Pendall et al 2004) The kinetics of ROS formation inthe leaf apoplast following O3 exposure seem to involve twophases an initial phase that is associated with direct effectsof O3 and a second phase associated with a plant-derivedsecondary oxidative burst (Schraudner et al 1998) The sec-ondary oxidative burst is initially localized in the leaf apo-
Crop responses to ozone 1001
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
plast and cell wall but later expands into the cytoplasm andsubcellular compartments leading to the formation of visi-ble lesions (Pendall et al 2004)
The initial signals induced by O3 in the leaf apoplastmight then be translated into responses at the tissue levelResponses vary and include unregulated cell death hyper-sensitive response leading to programmed cell death andaccelerated senescence (Pell Schlagnhaufer amp Arteca 1997Sandermann et al 1998) Evidence is accumulating thatthese responses are modulated by ethylene jasmonic andsalicylic acid levels and the interactions among their signal-ling pathways (Kangasjaumlrvi et al 1994 Sharma et al 1996Pell et al 1997 Overmyer et al 2000 Rao et al 2000 Raoamp Davis 2001 Rao Lee amp Davis 2002 Overmyer Broscheamp Kangasjaumlrvi 2003 Tamaoki et al 2003 Vahala et al 2003see also Kangasjaumlrvi et al 2005 this issue) For exampleArabidopsis thaliana mutants that overproduce ethylene(eto1) or are insensitive to jasmonic acid (jar1) are moreextensively injured by O3 than wild-type plants (Overmyeret al 2000 Rao et al 2000 Tamaoki et al 2003) Ethyleneacts as a promoting factor for O3 injury whereas jasmonatesmight have a role in minimizing injury (Overmyer et al2000 2003)
Ozone effects at the tissue level are often characterizedas either acute or chronic responses subjective terms thatare related to the type of visible symptoms observed andwhether the O3 concentrations employed typify ambient O3
episodes For example unregulated cell death and pro-grammed cell death are usually considered acute responsesin which lesions occur within hours after exposure to rela-tively high O3 concentrations (typically gt150 nmol mol-1)In contrast chronic responses include lesions that developover days to weeks under lower O3 concentrations andaccelerated senescence where lesions might not formUnder field conditions accelerated senescence may be dif-ficult to identify in the absence of a clean air control orother reference point (eg O3-tolerant cultivar) because thecumulative effects on foliar senescence at the end of theseason are separated in time from the O3 events causingthe effects The timing of O3 episodes in relation to plantdevelopment can also have a major effect on the outcomeas in the case of soybean in which O3 exposure duringreproductive growth has a much greater impact on yieldcompared with exposure during vegetative growth (Heagleet al 1991 Miller et al 1991 Morgan et al 2003) The dis-tinction between acute and chronic responses is furthercomplicated by species and genotype differences that leadto a range of effects for a given combination of O3 concen-tration and environmental conditions
Genetic variation in O3 responses and potential O3 tolerance mechanisms
Studies of O3 effects on plants often identify genetic varia-tion as a contributing factor in the observed responseOzone-sensitive and tolerant cultivars or clones have beenreported for many plant species (Wellburn amp Wellburn1996) including crops such as soybean (Tingey Reinert amp
Carter 1972 Heagle amp Letchworth 1982 Robinson amp Britz2000) snap bean (Phaseolus vulgaris L) (Guzy amp Heath1993) tomato (Lycopersicon esculentum Miller) (Temple1990) clover (Trifolium repens L) (Heagle et al 1993)wheat (Heagle Miller amp Pursley 2000) and potato(Solanum tuberosum L) (Heagle Miller amp Pursley 2003)and natural vegetation such as black cherry (Prunus sero-tina Ehrh) (Lee et al 2002) and Plantago spp (Wolff Mor-gan-Richards amp Davison 2000) Studies have also shownthat O3 tolerance is a heritable trait (Damicone amp Manning1987 Reinert amp Eason 2000) Although much effort hasbeen made to identify the physiological and biochemicalelements of O3 tolerance no clear picture has emergedBased on the oxidative stress model formation of ROS andother oxidation products is the underlying process involvedin the generation and propagation of toxic compounds andabiotic elicitors in plants Therefore features of antioxidantmetabolism that scavenge or affect the perception of O3-derived ROS could contribute to O3 tolerance
Plants have evolved elaborate systems to combat generaloxidative stress based on enzymes that utilize ascorbic acidand glutathione (Noctor amp Foyer 1998 see also Foyer ampNoctor 2005 this issue) and these antioxidant systems havebeen linked to O3 stress tolerance (Runeckles amp Chevone1992 Smirnoff 1996 Chernikova et al 2000 Conklin ampBarth 2004) A critical role for ascorbic acid is demon-strated with the vtc1 mutant of Arabidopsis in which asignificant decrease in ascorbate content was associatedwith increased O3 sensitivity (Conklin Williams amp Last1996) A more detailed analysis of vtc1 suggested that inaddition to acting as an antioxidant ascorbic acid serves aregulatory function in the signalling networks that controlplant defence responses and leaf senescence (Conklin ampBarth 2004) Although it is clear that minimum levels ofascorbic acid and glutathione are required for normal leaffunction the concentrations of these antioxidant metabo-lites that naturally occur in leaf tissue are not always wellcorrelated with O3 tolerance (Guzy amp Heath 1993 Well-burn amp Wellburn 1996 Burkey et al 2000) suggesting amore complex mechanism that remains to be elucidated
Recent studies have found higher levels of ascorbic acidin the leaf apoplast of certain O3-tolerant snap bean geno-types compared with sensitive lines (Burkey Eason amp Fis-cus 2003) suggesting that localization of ascorbic acid inthe cell wall where initial O3 reactions occur might beimportant Leaf extracellular ascorbate content and redoxstatus are affected by O3 treatment (Castillo amp Greppin1988 Luwe amp Heber 1995 Burkey 1999) evidence thatextracellular ascorbic acid is involved in detoxification ofO3 and related ROS Ascorbic acid is initially synthesizedinside cells (Smirnoff Conklin amp Loewus 2001) followedby transport between the cytoplasm and extracellular spacevia specific carriers located in the plasma membrane (Hore-mans Foyer amp Asard 2000) A model developed by Cha-meides (1989) and expanded by others (Ploumlchl et al 2000Turcsanyi et al 2000) features ascorbic acid in the leaf apo-plast as a critical factor in the detoxification of extracellularO3 and ROS protecting plasma membranes from oxidative
1002 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
damage and preventing O3 injury Ascorbic acid potentiallycould serve as either a direct chemical scavenger of O3although this mechanism has been questioned (Jakob ampHeber 1998) or as a substrate for extracellular enzymes(eg ascorbate peroxidase) that attenuate ROS levels andthus affect the propagation of the initial O3 signal How-ever leaf apoplast ascorbic acid levels can be quite low incertain plant species [summarized in Burkey et al (2003)]suggesting that extracellular scavenging of O3 and ROS byascorbate may not be a significant factor in all plants Othercompounds with antioxidant properties also exist in the leafapoplast Identification of these compounds and theirimpact on ROS formation and propagation are subjects forfuture research on O3 tolerance mechanisms
Another approach to understanding O3 responses inplants involves investigations of the potential interactionbetween chemical signals and specific receptors in theplasma membrane Booker et al (2004) showed that theepinasty frequently observed for wild-type Arabidopsisleaves after O3 treatment did not occur in gpa1 mutantplants lacking the alpha subunit of the heterotrimeric Gprotein complex suggesting that G-proteins may beinvolved in the transduction of O3-derived signals Thebinding of O3 or more likely one of the postulated O3 reac-tion products to a G-protein receptor could activate theprotein and initiate signal cascade processes that activatetarget proteins in this signal transduction system Howeverfurther studies will be required to define the possible roleG-proteins have in O3-induced signal transduction andpropagation It is also possible that G-proteins modulate O3
responses via hormone signal transduction processes Ineither case elements of G-protein pathways involved in O3
response represent potential molecular targets that mightbe manipulated to enhance O3 tolerance although this isspeculative at present The challenge will be to identifytargets unique to O3 so that other important stressresponses remain intact (eg plant defence responsesagainst pathogen attack) Finally it is likely that any pro-tective mechanism will come with some metabolic cost thatwill require evaluation to determine its net benefits
IMPACTS ON PHOTOSYNTHESIS
Net assimilation
Other than crop yield and visible injury photosynthesis hasbeen the most broadly studied aspect of plant responses toO3 Generally O3 exposure results in decreased photosyn-thetic carbon assimilation (Miller 1987b Runeckles ampChevone 1992 Pell et al 1997 Long amp Naidu 2002 Morganet al 2003) In many cases loss of assimilation capacity wasshown to be due primarily to reduced carboxylation effi-ciency directly related to loss of Rubisco activity Theselosses of activity are thought to be due to decreases inRubisco concentration in the leaves rather than a decreasein activation state Messenger RNA transcripts for theRubisco small subunit (rbcS) have been noted to declinewith O3 treatment in potato leaves (Glick et al 1995) sug-
gesting that protein synthesis might be inhibited a notionconsistent with observations that Rubisco content in poplar(Populus spp) leaves treated with O3 does not rise as highas controls during leaf expansion (Pell et al 1997) How-ever Eckardt amp Pell (1994) under conditions where addi-tional synthesis should be minimal showed a substantialdecline in Rubisco content suggesting that the primarycause of the decline in Rubisco due to O3 exposure wasenhanced degradation rather than reduced production
Because photosynthetic measurements are typicallymade on the most recently fully expanded upper canopyleaves in determinate plants including many soybean linesit is often difficult to demonstrate differences in net photo-synthesis (An) due to O3 until near or after the time whenproduction of new upper canopy leaves has ceased Thenduring most of the reproductive stages An decreased earlierthan in control plants (accelerated senescence) This sug-gests that during vegetative growth either Rubisco synthe-sis in newly formed upper canopy leaves is not substantiallyaffected or that these leaves have not accumulated suffi-cient damage to elicit a response Vozzo et al (1995) dem-onstrated significant reductions in An on somewhat oldersoybean leaves that were relatively unaffected by shadingbut had received sufficient exposure to exhibit visible symp-toms thus lending support to this idea In contrast Morganet al (2004) using an indeterminate line were unable todemonstrate any differences due to dose accumulation in acohort of soybean leaves that developed during vegetativegrowth However they did observe decreased An in anothercohort that developed during reproductive growth andremained near the top of the canopy These results couldwell have been due to differences in exposure methodolo-gies particularly the greater atmospheric coupling to thelower canopy in OTCs as compared to FACE systems Infurther contrast to the above observations additionalreview of the data in two earlier soybean studies (Reid ampFiscus 1998 Reid et al 1998) illustrated in Fig 2 showsthat Rubisco content and activity chlorophyll content andAn were reduced by O3 early in the vegetative stage andthat all but An had reached or exceeded clean air levels byearly flowering Afterwards while chlorophyll content Anand Rubisco activity remained level or declined slightlyRubisco content continued to rise in the O3 treatmentFinally in early seed development all these parametersbegan to decline more precipitously Throughout this wholeexperiment chlorophyll content and Rubisco activitytracked each other very well This suggests that from earlyflowering through most of seed development O3 stimulatedRubisco content on a leaf area basis In addition the ques-tion of the role of accelerated senescence especially duringreproductive growth is one deserving more attention Forexample in Fig 2 the O3-induced decline in An duringreproduction may be due to the normal process of senes-cence that has been accelerated by O3 exposure or it maybe due simply to the loss of photosynthetically active leafarea due to accumulated O3 damage Thus especially inareas associated with visible lesions one would measure amixture of live and dead or dying cells and it seems rele-
Crop responses to ozone 1003
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
vant to know whether the remaining live cells are function-ing at or near normal levels for their age This possibility issuggested by the fact that from beginning of pod fill throughseed maturation when An per unit area is declining in thisstudy An per unit Rubisco remained high at 91 of the CFcontrol (difference not significant and data not shown)Further information is necessary to clarify this Anotherpoint deserving attention and bearing on the exposureaccumulation comments earlier in the paragraph is the sub-stantially reduced An early in the vegetative period Thismay have been caused by some inherent sensitivity of theseleaves or be the result of enhanced atmospheric couplingdue to the more open nature of the canopy at this growthstage
Photosynthetic electron transport
Detrimental effects of O3 on photosynthetic electron trans-port has received increased attention in recent years andwhile earlier it had been difficult to demonstrate significantdeclines in function evidence is accumulating that in someplants there are demonstrable effects both on PSII func-tion as well as the xanthophyll cycle components Forinstance significant and substantial declines in FvFm whichrepresents the efficiency of excitation energy capture byPSII in dark-adapted leaves have been reported for pump-kin (Cucurbita pepo L) (Ciompi et al 1997 Castagna et al2001) wheat (T durum and T aestivum) (Reichenauer et al1998) bean (P vulgaris L) (Guidi Di Cagno amp Soladatini2000a Guidi Tonini amp Soladatini 2000b Leipner Oxbor-ough amp Baker 2001 Guidi DeglrsquoInnocenti amp Soldatini2002) tomato (Calatayud amp Barreno 2001) and Festucapratensis and turnip (Brassica napus L) (Plazek Rapacz ampSkoczowski 2000) Of these particular studies three dem-
onstrated an increase in Fo which suggested damaged ordeactivated PSII centers (Guidi et al 2000a Leipner et al2001 Guidi et al 2002) all occurring in bean In additionlight saturated photosynthesis (ASAT) decreased in bean(Guidi et al 2000a Guidi et al 2002) pumpkin (Ciompiet al 1997 Castagna et al 2001) and wheat (Reichenaueret al 1998) although in the latter the effect was confined toolder leaves in two of the three lines and was partiallyreversible in some lines and fully reversible overnight inone It was unknown whether reversibility was due to anight-time epoxidation of the xanthophyll cycle decreasingsubsequent thermal quenching of PSII or to overnightrepair of damaged PSII centres This latter point becomesrelevant since a decrease in FvFm may be caused by both arelatively fast reversible down-regulation or a reversibledestruction (Osmond 1994) However the two processesmight be distinguished by monitoring effects on Fo in thatthermal dissipation by the xanthophyll cycle would lowerFo whereas damage to PSII would increase Fo as was foundin bean (Guidi et al 2000a 2002 Leipner et al 2001) EPRmeasurements also showed increases in free radicals whichReichenauer et al (1998) suggested might contribute todamage of chloroplast membranes in the sensitive wheatlines Runeckles amp Vaartnou (1997) also observed light-dependent EPR signals in bluegrass (Poa pratensis L)ryegrass (Lolium perenne L) and radish (Raphanus sativaL) consistent with superoxide anion characteristics onexposure to O3 The light dependence suggested that thesignal was localized in the chloroplast and significantly itsappearance was reduced in leaves with increased levels ofapoplastic ascorbic acid
High light is another factor that might interact with O3
in such a way as to increase or aggravate the effects of O3
alone through the process of photoinhibition with a result-
Figure 2 Rubisco quantity and activity An and chlorophyll content ratios for soybean (cv Essex) leaves treated with charcoal-filtered air (CF) 25 nmol O3 mol-1 and 15 yen ambient O3 (75 nmol mol-1 12 h average) in open-top field chambers in 1994 The ratios represent the response to O3 and are calculated as the quantity present in the O3 treatment compared with the control (O3CF) Developmental stages are given in the bar above the graph Data are from Reid et al (1998) and Reid amp Fiscus (1998)
1004 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
ant increase in free radicals in the chloroplast (Massman2004) Guidi et al (2002) demonstrated that both high lightand O3 decreased FvFm and increased Fo Furthermore thecombined effects were additive Farage et al (1991) uponexposing wheat leaves to high [O3] (200 and 400 nmolmol-1) could detect no difference in atrazine binding inisolated thylakoids nor any difference in D1 protein asindicated by western blots using D1-specific antibodiesHowever Godde amp Buchhold (1992) found that the turn-over rate for D1 was increased by exposure of spruce nee-dles to O3 suggesting both increased degradation andsynthesis In addition Ciompi et al (1997) found that inyoung leaves of pumpkin engagement of the xanthophyllcycle appeared to counteract the effects of O3 on the PSIIreaction center preventing a decline in D1 content How-ever in mature leaves even though there was activation ofthe xanthophyll cycle the effect appeared inadequate toprevent photoinhibition and loss of D1 protein The authorssuggested that this effect in mature leaves may have beendue to the normal decline in antioxidant activity in olderleaves Clearly high light has the potential to induce photo-inhibition which could be already sensitized by O3 exposurein field crops or inversely and is an area that requiresfurther investigation especially under more realistic fieldconditions
Direct effects on stomata
Another possible point of attack by O3 that cannot beignored is the guard cells Although much of the evidenceto date indicates that reduced gs is the result and not thecause of reduced assimilation (McKee et al 1995 Fiscuset al 1997 Long amp Naidu 2002) as suggested anddeduced on the basis of water use efficiency studies(Reich Schoettle amp Amundsen 1985) there is someintriguing work being done on possible direct effects ofO3 on stomatal response which links oxidative stress inthe form of H2O2 Ca2+ homeostasis of guard cells andABA response This work was recently reviewed byMcAinsh et al (2002) while the review by Schroeder et al(2001) provides a more detailed context for these studiesThe studies showed that H2O2 stimulated increased wholeplant cytosolic free Ca ions [Ca2+]cyt The stimulation washighly dose dependent with biphasic kinetics similar tothe Ca signature induced by exposure to O3 which wasalso highly dose dependent Using various techniquesincreased [Ca2+]cyt was also demonstrated in guard cells inresponse to both O3 and H2O2 In addition Pei et al(2000) demonstrated H2O2-induced stomatal closure andthat ABA induces H2O2 production in guard cells andactivates plasma membrane Ca-permeable channels Allof these responses were disrupted in the ABA-insensitiveArabidopsis mutant gca2 Taken together these and otherstudies reviewed (McAinsh et al 2002) raise the possibil-ity that O3 might directly affect stomata through theaction of H2O2 formed when O3 enters solution on theCa-permeable channels in the guard cell plasma mem-branes
O3ndashCO2 interactions on biomass and yield
Along with O3 atmospheric CO2 concentrations are alsorising and numerous studies indicate that elevated CO2
might enhance the productivity of current cropping sys-tems Long et al (2004) have shown that the magnitude ofthe response to elevated CO2 is highly variable not onlyacross but within species and cultivars Although thereappears to be a genetic component to this variability someof it may be due to environmental stresses that occurredduring plant growth Ozone is one such possible stress andit would not be surprising that there are significant physio-logical interactions between elevated CO2 and O3 The mostrelevant interaction for present purposes is the reductionin gs at elevated levels of CO2 that has the effect of prevent-ing reductions of An growth and yield in many crops grownin the presence of toxic levels of O3 (Fiscus et al 1997Olszyk et al 2000 Fiscus et al 2002 Morgan et al 2003Booker et al 2005) Figure 3 illustrates this interaction insoybean grown in both elevated CO2 and O3 (compare withAn ratio in Fig 2) and although it is expected that theassimilation ratios in elevated CO2 are substantially higherthan the control it is interesting to note that elevated CO2
did not seem to prevent the accelerated senescence charac-teristic of an O3 fumigation at current levels of CO2 Anal-ysis of compiled shoot biomass and yield data (Fiscus et al2002) show that increased An does get translated into anaverage 30 increase in biomass but often not into yieldin clean (charcoal-filtered) air (Fig 4) (Fiscus et al 2001)The variability in both biomass and yield is large with yieldsin doubled CO2 ranging from -37 to +41 in CF air witha mean of +6 across the crops studied However inincreasing O3 concentrations both the apparent biomassand yield due to elevated CO2 (CO2 fertilization effect)increased dramatically but the variability of the responsesremained large Of course in elevated CO2 gs will bereduced typically by 30 in these studies so O3 flux intothe leaves and the effective exposure is much reduced Thusthe apparent dramatic yield increases in some non-filteredair experiments might be caused in part by the fact thatyields are already suppressed by 5ndash15 by O3 in ambientair control plants and the best that elevated CO2 seemedto do in these experiments was to restore yields to the cleanair levels The wide range of variability in these experimentssuggests that in some CO2 was not the limiting resourceduring reproduction In fact it seems possible given thelarge increases in vegetative biomass that early resourcedepletion under elevated CO2 actually may have reducedyield One would expect that any environmental stress thataffects gs (eg drought) would also influence the directeffects of O3 and lead to large apparent CO2-stimulatedincreases in growth and yield The cause of the distinctvariability shown in these experiments is an area thatdeserves further research
Impacts on translocation and partitioning
Early in the season chronic exposure to O3 can inhibit An
(Fig 2) and plant growth (Fig 5) but often the decline in
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

1000 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
In the field canopy structure and foliage density affectthe concentration of O3 that leaves experience while envi-ronmental factors such as PPFD leaf temperature and VPDvary within the leaf canopy and can affect gs and thus O3
flux However after canopy closure for crops such as soy-bean cotton (Gossypium hirsutum L) and peanut (Ara-chis hypogaea L) leaves at the top of the canopy providethe majority of photosynthate for seed production Ozoneflux to the upper portion of the canopy is likely most criticalto impacts on yield For crops such as wheat (Triticum aes-tivum L) and rice (Oryza sativa L) flux to the entirecanopy might be important Efforts to parameterize canopyO3 flux have included leaf area index as a way to estimatecanopy gs (Massman 2004) which may be effective inaccounting for differences among crop canopy structuresHowever the ICP Vegetation protocol utilizes projectedleaf area namely leaf area of sunlit leaves at the top of thecanopy to calculate O3 flux (Pleijel et al 2004 UNECE2004b) Ozone flux to the most productive leaves is viewedas having the greatest impact on yield because even thoughthe soil and lower parts of the canopy consume largeamounts of O3 it is likely these fluxes have much less effecton yield (Pleijel et al 2004 Ashmore 2005)
Lastly the ICP Vegetation program (UNECE 2004a) iscurrently comparing economic crop loss assessments ofconcentration-based and flux-based methodologies Thisanalysis should suggest whether the flux approach providesa significantly better assessment of crop losses due to O3
pollution compared with concentration-based exposureindices
MODES OF ACTION
Apoplastic and symplastic reactions and responses
The internal air spaces within the leaf are a potential sitefor O3 reactions with volatile compounds produced by theplant Isoprene is one example of a compound emitted frommany angiosperms and conifers (Sharkey amp Yeh 2001) thatcould react with O3 However volatile compounds releasedby crop species are mainly monoterpenes which have beenlittle studied in this context although parallels betweenthese volatile organic compounds (VOCs) and O3 reactionswith isoprene are likely At present hypotheses are contra-dictory regarding the role of isoprene in plant responses toO3 Initially isoprene was thought to enhance O3 damagethrough the formation of hydroperoxides (Hewitt Kok ampRall 1990) More recently isoprene was found to exert aprotective effect either through action as a radical scaven-ger or membrane stabilizer (Loreto amp Velikova 2001) Theconcept of isoprene as an antioxidant is intriguing from theperspective of O3 scavenging reactions outside the leaf thatcould increase boundary layer resistance to O3 and effec-tively reduce the dose The potential benefits of such amechanism must be balanced against the metabolicdemand for carbon to support isoprene synthesis (Sharkeyamp Yeh 2001) and the consequences of increased plant-
derived VOCs on atmospheric chemistry leading to O3
formationOzone that has passed through leaf internal air spaces
will then dissolve in the aqueous layer surrounding leafcells The breakdown of O3 in pure water produces hydroxyland peroxyl radicals and superoxide although the reactionsproceed very slowly at neutral pH (Heath 1987) GrimesPerkins amp Boss (1983) demonstrated that the rate ofhydroxyl radical formation from O3 in aqueous solution issignificantly greater when phenolic compounds are presentsuggesting that biologically relevant compounds enhanceO3 reactions Based on in vitro and in vivo studies (Heath1987 Runeckles amp Chevone 1992 Pryor 1994 Kirichenkoet al 1996) O3 is known to react with a diverse set ofmolecules that would be encountered within the cell walland on the plasma membrane surface Initial targets for O3
include plasma membrane lipids susceptible amino acids inplasma membrane proteins or apoplastic enzymes and avariety of organic metabolites localized in the cell wallPotentially ozonolysis products from the oxidation of lipidsand metabolites containing carbon-carbon double bonds(Heath 1987 Runeckles amp Chevone 1992) and alteredplasma membrane protein function (Dominy amp Heath1985 Castillo amp Heath 1990) could serve as the initial sig-nals leading to O3 responses However the primary O3
reaction products in plants have not been identified due todifficulties in following O3 reactions within the complexbiochemical network of the leaf apoplast Knowledge ofthese early events is critical to understanding the metabolicsignals that initiate O3 responses and may provide insightinto O3 tolerance mechanisms localized in the leaf apoplastOzone incorporation into the cells and extracellular lipid-protein complexes of rat lung tissue has been studied usinga combination of 18O3 and mass spectrometry (Gunnison ampHatch 1999) an approach that could be employed in con-junction with available methods for isolation of leaf apo-plast components and plasma membranes to addressquestions regarding the initial O3 reaction products inplants
Formation of reactive oxygen species (ROS) such assuperoxide hydrogen peroxide and hydroxyl radical arethought to be associated with the initial breakdown of O3
in the leaf apoplast ESR approaches based on spin trap-ping suggest that free radicals are involved in the earlystages of O3 response (Grimes et al 1983 Mehlhorn Tab-ner amp Wellburn 1990 Runeckles amp Vaartnou 1997) but itis not clear whether the observed signals originate fromdirect O3 breakdown or the initial reactions with cellularconstituents such as phenolic compounds Localizedregions of ROS formation either hydrogen peroxide orsuperoxide depending on the plant species are associatedwith O3 response in sensitive plants (Wohlgemuth et al2002 Pendall et al 2004) The kinetics of ROS formation inthe leaf apoplast following O3 exposure seem to involve twophases an initial phase that is associated with direct effectsof O3 and a second phase associated with a plant-derivedsecondary oxidative burst (Schraudner et al 1998) The sec-ondary oxidative burst is initially localized in the leaf apo-
Crop responses to ozone 1001
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
plast and cell wall but later expands into the cytoplasm andsubcellular compartments leading to the formation of visi-ble lesions (Pendall et al 2004)
The initial signals induced by O3 in the leaf apoplastmight then be translated into responses at the tissue levelResponses vary and include unregulated cell death hyper-sensitive response leading to programmed cell death andaccelerated senescence (Pell Schlagnhaufer amp Arteca 1997Sandermann et al 1998) Evidence is accumulating thatthese responses are modulated by ethylene jasmonic andsalicylic acid levels and the interactions among their signal-ling pathways (Kangasjaumlrvi et al 1994 Sharma et al 1996Pell et al 1997 Overmyer et al 2000 Rao et al 2000 Raoamp Davis 2001 Rao Lee amp Davis 2002 Overmyer Broscheamp Kangasjaumlrvi 2003 Tamaoki et al 2003 Vahala et al 2003see also Kangasjaumlrvi et al 2005 this issue) For exampleArabidopsis thaliana mutants that overproduce ethylene(eto1) or are insensitive to jasmonic acid (jar1) are moreextensively injured by O3 than wild-type plants (Overmyeret al 2000 Rao et al 2000 Tamaoki et al 2003) Ethyleneacts as a promoting factor for O3 injury whereas jasmonatesmight have a role in minimizing injury (Overmyer et al2000 2003)
Ozone effects at the tissue level are often characterizedas either acute or chronic responses subjective terms thatare related to the type of visible symptoms observed andwhether the O3 concentrations employed typify ambient O3
episodes For example unregulated cell death and pro-grammed cell death are usually considered acute responsesin which lesions occur within hours after exposure to rela-tively high O3 concentrations (typically gt150 nmol mol-1)In contrast chronic responses include lesions that developover days to weeks under lower O3 concentrations andaccelerated senescence where lesions might not formUnder field conditions accelerated senescence may be dif-ficult to identify in the absence of a clean air control orother reference point (eg O3-tolerant cultivar) because thecumulative effects on foliar senescence at the end of theseason are separated in time from the O3 events causingthe effects The timing of O3 episodes in relation to plantdevelopment can also have a major effect on the outcomeas in the case of soybean in which O3 exposure duringreproductive growth has a much greater impact on yieldcompared with exposure during vegetative growth (Heagleet al 1991 Miller et al 1991 Morgan et al 2003) The dis-tinction between acute and chronic responses is furthercomplicated by species and genotype differences that leadto a range of effects for a given combination of O3 concen-tration and environmental conditions
Genetic variation in O3 responses and potential O3 tolerance mechanisms
Studies of O3 effects on plants often identify genetic varia-tion as a contributing factor in the observed responseOzone-sensitive and tolerant cultivars or clones have beenreported for many plant species (Wellburn amp Wellburn1996) including crops such as soybean (Tingey Reinert amp
Carter 1972 Heagle amp Letchworth 1982 Robinson amp Britz2000) snap bean (Phaseolus vulgaris L) (Guzy amp Heath1993) tomato (Lycopersicon esculentum Miller) (Temple1990) clover (Trifolium repens L) (Heagle et al 1993)wheat (Heagle Miller amp Pursley 2000) and potato(Solanum tuberosum L) (Heagle Miller amp Pursley 2003)and natural vegetation such as black cherry (Prunus sero-tina Ehrh) (Lee et al 2002) and Plantago spp (Wolff Mor-gan-Richards amp Davison 2000) Studies have also shownthat O3 tolerance is a heritable trait (Damicone amp Manning1987 Reinert amp Eason 2000) Although much effort hasbeen made to identify the physiological and biochemicalelements of O3 tolerance no clear picture has emergedBased on the oxidative stress model formation of ROS andother oxidation products is the underlying process involvedin the generation and propagation of toxic compounds andabiotic elicitors in plants Therefore features of antioxidantmetabolism that scavenge or affect the perception of O3-derived ROS could contribute to O3 tolerance
Plants have evolved elaborate systems to combat generaloxidative stress based on enzymes that utilize ascorbic acidand glutathione (Noctor amp Foyer 1998 see also Foyer ampNoctor 2005 this issue) and these antioxidant systems havebeen linked to O3 stress tolerance (Runeckles amp Chevone1992 Smirnoff 1996 Chernikova et al 2000 Conklin ampBarth 2004) A critical role for ascorbic acid is demon-strated with the vtc1 mutant of Arabidopsis in which asignificant decrease in ascorbate content was associatedwith increased O3 sensitivity (Conklin Williams amp Last1996) A more detailed analysis of vtc1 suggested that inaddition to acting as an antioxidant ascorbic acid serves aregulatory function in the signalling networks that controlplant defence responses and leaf senescence (Conklin ampBarth 2004) Although it is clear that minimum levels ofascorbic acid and glutathione are required for normal leaffunction the concentrations of these antioxidant metabo-lites that naturally occur in leaf tissue are not always wellcorrelated with O3 tolerance (Guzy amp Heath 1993 Well-burn amp Wellburn 1996 Burkey et al 2000) suggesting amore complex mechanism that remains to be elucidated
Recent studies have found higher levels of ascorbic acidin the leaf apoplast of certain O3-tolerant snap bean geno-types compared with sensitive lines (Burkey Eason amp Fis-cus 2003) suggesting that localization of ascorbic acid inthe cell wall where initial O3 reactions occur might beimportant Leaf extracellular ascorbate content and redoxstatus are affected by O3 treatment (Castillo amp Greppin1988 Luwe amp Heber 1995 Burkey 1999) evidence thatextracellular ascorbic acid is involved in detoxification ofO3 and related ROS Ascorbic acid is initially synthesizedinside cells (Smirnoff Conklin amp Loewus 2001) followedby transport between the cytoplasm and extracellular spacevia specific carriers located in the plasma membrane (Hore-mans Foyer amp Asard 2000) A model developed by Cha-meides (1989) and expanded by others (Ploumlchl et al 2000Turcsanyi et al 2000) features ascorbic acid in the leaf apo-plast as a critical factor in the detoxification of extracellularO3 and ROS protecting plasma membranes from oxidative
1002 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
damage and preventing O3 injury Ascorbic acid potentiallycould serve as either a direct chemical scavenger of O3although this mechanism has been questioned (Jakob ampHeber 1998) or as a substrate for extracellular enzymes(eg ascorbate peroxidase) that attenuate ROS levels andthus affect the propagation of the initial O3 signal How-ever leaf apoplast ascorbic acid levels can be quite low incertain plant species [summarized in Burkey et al (2003)]suggesting that extracellular scavenging of O3 and ROS byascorbate may not be a significant factor in all plants Othercompounds with antioxidant properties also exist in the leafapoplast Identification of these compounds and theirimpact on ROS formation and propagation are subjects forfuture research on O3 tolerance mechanisms
Another approach to understanding O3 responses inplants involves investigations of the potential interactionbetween chemical signals and specific receptors in theplasma membrane Booker et al (2004) showed that theepinasty frequently observed for wild-type Arabidopsisleaves after O3 treatment did not occur in gpa1 mutantplants lacking the alpha subunit of the heterotrimeric Gprotein complex suggesting that G-proteins may beinvolved in the transduction of O3-derived signals Thebinding of O3 or more likely one of the postulated O3 reac-tion products to a G-protein receptor could activate theprotein and initiate signal cascade processes that activatetarget proteins in this signal transduction system Howeverfurther studies will be required to define the possible roleG-proteins have in O3-induced signal transduction andpropagation It is also possible that G-proteins modulate O3
responses via hormone signal transduction processes Ineither case elements of G-protein pathways involved in O3
response represent potential molecular targets that mightbe manipulated to enhance O3 tolerance although this isspeculative at present The challenge will be to identifytargets unique to O3 so that other important stressresponses remain intact (eg plant defence responsesagainst pathogen attack) Finally it is likely that any pro-tective mechanism will come with some metabolic cost thatwill require evaluation to determine its net benefits
IMPACTS ON PHOTOSYNTHESIS
Net assimilation
Other than crop yield and visible injury photosynthesis hasbeen the most broadly studied aspect of plant responses toO3 Generally O3 exposure results in decreased photosyn-thetic carbon assimilation (Miller 1987b Runeckles ampChevone 1992 Pell et al 1997 Long amp Naidu 2002 Morganet al 2003) In many cases loss of assimilation capacity wasshown to be due primarily to reduced carboxylation effi-ciency directly related to loss of Rubisco activity Theselosses of activity are thought to be due to decreases inRubisco concentration in the leaves rather than a decreasein activation state Messenger RNA transcripts for theRubisco small subunit (rbcS) have been noted to declinewith O3 treatment in potato leaves (Glick et al 1995) sug-
gesting that protein synthesis might be inhibited a notionconsistent with observations that Rubisco content in poplar(Populus spp) leaves treated with O3 does not rise as highas controls during leaf expansion (Pell et al 1997) How-ever Eckardt amp Pell (1994) under conditions where addi-tional synthesis should be minimal showed a substantialdecline in Rubisco content suggesting that the primarycause of the decline in Rubisco due to O3 exposure wasenhanced degradation rather than reduced production
Because photosynthetic measurements are typicallymade on the most recently fully expanded upper canopyleaves in determinate plants including many soybean linesit is often difficult to demonstrate differences in net photo-synthesis (An) due to O3 until near or after the time whenproduction of new upper canopy leaves has ceased Thenduring most of the reproductive stages An decreased earlierthan in control plants (accelerated senescence) This sug-gests that during vegetative growth either Rubisco synthe-sis in newly formed upper canopy leaves is not substantiallyaffected or that these leaves have not accumulated suffi-cient damage to elicit a response Vozzo et al (1995) dem-onstrated significant reductions in An on somewhat oldersoybean leaves that were relatively unaffected by shadingbut had received sufficient exposure to exhibit visible symp-toms thus lending support to this idea In contrast Morganet al (2004) using an indeterminate line were unable todemonstrate any differences due to dose accumulation in acohort of soybean leaves that developed during vegetativegrowth However they did observe decreased An in anothercohort that developed during reproductive growth andremained near the top of the canopy These results couldwell have been due to differences in exposure methodolo-gies particularly the greater atmospheric coupling to thelower canopy in OTCs as compared to FACE systems Infurther contrast to the above observations additionalreview of the data in two earlier soybean studies (Reid ampFiscus 1998 Reid et al 1998) illustrated in Fig 2 showsthat Rubisco content and activity chlorophyll content andAn were reduced by O3 early in the vegetative stage andthat all but An had reached or exceeded clean air levels byearly flowering Afterwards while chlorophyll content Anand Rubisco activity remained level or declined slightlyRubisco content continued to rise in the O3 treatmentFinally in early seed development all these parametersbegan to decline more precipitously Throughout this wholeexperiment chlorophyll content and Rubisco activitytracked each other very well This suggests that from earlyflowering through most of seed development O3 stimulatedRubisco content on a leaf area basis In addition the ques-tion of the role of accelerated senescence especially duringreproductive growth is one deserving more attention Forexample in Fig 2 the O3-induced decline in An duringreproduction may be due to the normal process of senes-cence that has been accelerated by O3 exposure or it maybe due simply to the loss of photosynthetically active leafarea due to accumulated O3 damage Thus especially inareas associated with visible lesions one would measure amixture of live and dead or dying cells and it seems rele-
Crop responses to ozone 1003
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
vant to know whether the remaining live cells are function-ing at or near normal levels for their age This possibility issuggested by the fact that from beginning of pod fill throughseed maturation when An per unit area is declining in thisstudy An per unit Rubisco remained high at 91 of the CFcontrol (difference not significant and data not shown)Further information is necessary to clarify this Anotherpoint deserving attention and bearing on the exposureaccumulation comments earlier in the paragraph is the sub-stantially reduced An early in the vegetative period Thismay have been caused by some inherent sensitivity of theseleaves or be the result of enhanced atmospheric couplingdue to the more open nature of the canopy at this growthstage
Photosynthetic electron transport
Detrimental effects of O3 on photosynthetic electron trans-port has received increased attention in recent years andwhile earlier it had been difficult to demonstrate significantdeclines in function evidence is accumulating that in someplants there are demonstrable effects both on PSII func-tion as well as the xanthophyll cycle components Forinstance significant and substantial declines in FvFm whichrepresents the efficiency of excitation energy capture byPSII in dark-adapted leaves have been reported for pump-kin (Cucurbita pepo L) (Ciompi et al 1997 Castagna et al2001) wheat (T durum and T aestivum) (Reichenauer et al1998) bean (P vulgaris L) (Guidi Di Cagno amp Soladatini2000a Guidi Tonini amp Soladatini 2000b Leipner Oxbor-ough amp Baker 2001 Guidi DeglrsquoInnocenti amp Soldatini2002) tomato (Calatayud amp Barreno 2001) and Festucapratensis and turnip (Brassica napus L) (Plazek Rapacz ampSkoczowski 2000) Of these particular studies three dem-
onstrated an increase in Fo which suggested damaged ordeactivated PSII centers (Guidi et al 2000a Leipner et al2001 Guidi et al 2002) all occurring in bean In additionlight saturated photosynthesis (ASAT) decreased in bean(Guidi et al 2000a Guidi et al 2002) pumpkin (Ciompiet al 1997 Castagna et al 2001) and wheat (Reichenaueret al 1998) although in the latter the effect was confined toolder leaves in two of the three lines and was partiallyreversible in some lines and fully reversible overnight inone It was unknown whether reversibility was due to anight-time epoxidation of the xanthophyll cycle decreasingsubsequent thermal quenching of PSII or to overnightrepair of damaged PSII centres This latter point becomesrelevant since a decrease in FvFm may be caused by both arelatively fast reversible down-regulation or a reversibledestruction (Osmond 1994) However the two processesmight be distinguished by monitoring effects on Fo in thatthermal dissipation by the xanthophyll cycle would lowerFo whereas damage to PSII would increase Fo as was foundin bean (Guidi et al 2000a 2002 Leipner et al 2001) EPRmeasurements also showed increases in free radicals whichReichenauer et al (1998) suggested might contribute todamage of chloroplast membranes in the sensitive wheatlines Runeckles amp Vaartnou (1997) also observed light-dependent EPR signals in bluegrass (Poa pratensis L)ryegrass (Lolium perenne L) and radish (Raphanus sativaL) consistent with superoxide anion characteristics onexposure to O3 The light dependence suggested that thesignal was localized in the chloroplast and significantly itsappearance was reduced in leaves with increased levels ofapoplastic ascorbic acid
High light is another factor that might interact with O3
in such a way as to increase or aggravate the effects of O3
alone through the process of photoinhibition with a result-
Figure 2 Rubisco quantity and activity An and chlorophyll content ratios for soybean (cv Essex) leaves treated with charcoal-filtered air (CF) 25 nmol O3 mol-1 and 15 yen ambient O3 (75 nmol mol-1 12 h average) in open-top field chambers in 1994 The ratios represent the response to O3 and are calculated as the quantity present in the O3 treatment compared with the control (O3CF) Developmental stages are given in the bar above the graph Data are from Reid et al (1998) and Reid amp Fiscus (1998)
1004 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
ant increase in free radicals in the chloroplast (Massman2004) Guidi et al (2002) demonstrated that both high lightand O3 decreased FvFm and increased Fo Furthermore thecombined effects were additive Farage et al (1991) uponexposing wheat leaves to high [O3] (200 and 400 nmolmol-1) could detect no difference in atrazine binding inisolated thylakoids nor any difference in D1 protein asindicated by western blots using D1-specific antibodiesHowever Godde amp Buchhold (1992) found that the turn-over rate for D1 was increased by exposure of spruce nee-dles to O3 suggesting both increased degradation andsynthesis In addition Ciompi et al (1997) found that inyoung leaves of pumpkin engagement of the xanthophyllcycle appeared to counteract the effects of O3 on the PSIIreaction center preventing a decline in D1 content How-ever in mature leaves even though there was activation ofthe xanthophyll cycle the effect appeared inadequate toprevent photoinhibition and loss of D1 protein The authorssuggested that this effect in mature leaves may have beendue to the normal decline in antioxidant activity in olderleaves Clearly high light has the potential to induce photo-inhibition which could be already sensitized by O3 exposurein field crops or inversely and is an area that requiresfurther investigation especially under more realistic fieldconditions
Direct effects on stomata
Another possible point of attack by O3 that cannot beignored is the guard cells Although much of the evidenceto date indicates that reduced gs is the result and not thecause of reduced assimilation (McKee et al 1995 Fiscuset al 1997 Long amp Naidu 2002) as suggested anddeduced on the basis of water use efficiency studies(Reich Schoettle amp Amundsen 1985) there is someintriguing work being done on possible direct effects ofO3 on stomatal response which links oxidative stress inthe form of H2O2 Ca2+ homeostasis of guard cells andABA response This work was recently reviewed byMcAinsh et al (2002) while the review by Schroeder et al(2001) provides a more detailed context for these studiesThe studies showed that H2O2 stimulated increased wholeplant cytosolic free Ca ions [Ca2+]cyt The stimulation washighly dose dependent with biphasic kinetics similar tothe Ca signature induced by exposure to O3 which wasalso highly dose dependent Using various techniquesincreased [Ca2+]cyt was also demonstrated in guard cells inresponse to both O3 and H2O2 In addition Pei et al(2000) demonstrated H2O2-induced stomatal closure andthat ABA induces H2O2 production in guard cells andactivates plasma membrane Ca-permeable channels Allof these responses were disrupted in the ABA-insensitiveArabidopsis mutant gca2 Taken together these and otherstudies reviewed (McAinsh et al 2002) raise the possibil-ity that O3 might directly affect stomata through theaction of H2O2 formed when O3 enters solution on theCa-permeable channels in the guard cell plasma mem-branes
O3ndashCO2 interactions on biomass and yield
Along with O3 atmospheric CO2 concentrations are alsorising and numerous studies indicate that elevated CO2
might enhance the productivity of current cropping sys-tems Long et al (2004) have shown that the magnitude ofthe response to elevated CO2 is highly variable not onlyacross but within species and cultivars Although thereappears to be a genetic component to this variability someof it may be due to environmental stresses that occurredduring plant growth Ozone is one such possible stress andit would not be surprising that there are significant physio-logical interactions between elevated CO2 and O3 The mostrelevant interaction for present purposes is the reductionin gs at elevated levels of CO2 that has the effect of prevent-ing reductions of An growth and yield in many crops grownin the presence of toxic levels of O3 (Fiscus et al 1997Olszyk et al 2000 Fiscus et al 2002 Morgan et al 2003Booker et al 2005) Figure 3 illustrates this interaction insoybean grown in both elevated CO2 and O3 (compare withAn ratio in Fig 2) and although it is expected that theassimilation ratios in elevated CO2 are substantially higherthan the control it is interesting to note that elevated CO2
did not seem to prevent the accelerated senescence charac-teristic of an O3 fumigation at current levels of CO2 Anal-ysis of compiled shoot biomass and yield data (Fiscus et al2002) show that increased An does get translated into anaverage 30 increase in biomass but often not into yieldin clean (charcoal-filtered) air (Fig 4) (Fiscus et al 2001)The variability in both biomass and yield is large with yieldsin doubled CO2 ranging from -37 to +41 in CF air witha mean of +6 across the crops studied However inincreasing O3 concentrations both the apparent biomassand yield due to elevated CO2 (CO2 fertilization effect)increased dramatically but the variability of the responsesremained large Of course in elevated CO2 gs will bereduced typically by 30 in these studies so O3 flux intothe leaves and the effective exposure is much reduced Thusthe apparent dramatic yield increases in some non-filteredair experiments might be caused in part by the fact thatyields are already suppressed by 5ndash15 by O3 in ambientair control plants and the best that elevated CO2 seemedto do in these experiments was to restore yields to the cleanair levels The wide range of variability in these experimentssuggests that in some CO2 was not the limiting resourceduring reproduction In fact it seems possible given thelarge increases in vegetative biomass that early resourcedepletion under elevated CO2 actually may have reducedyield One would expect that any environmental stress thataffects gs (eg drought) would also influence the directeffects of O3 and lead to large apparent CO2-stimulatedincreases in growth and yield The cause of the distinctvariability shown in these experiments is an area thatdeserves further research
Impacts on translocation and partitioning
Early in the season chronic exposure to O3 can inhibit An
(Fig 2) and plant growth (Fig 5) but often the decline in
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

Crop responses to ozone 1001
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
plast and cell wall but later expands into the cytoplasm andsubcellular compartments leading to the formation of visi-ble lesions (Pendall et al 2004)
The initial signals induced by O3 in the leaf apoplastmight then be translated into responses at the tissue levelResponses vary and include unregulated cell death hyper-sensitive response leading to programmed cell death andaccelerated senescence (Pell Schlagnhaufer amp Arteca 1997Sandermann et al 1998) Evidence is accumulating thatthese responses are modulated by ethylene jasmonic andsalicylic acid levels and the interactions among their signal-ling pathways (Kangasjaumlrvi et al 1994 Sharma et al 1996Pell et al 1997 Overmyer et al 2000 Rao et al 2000 Raoamp Davis 2001 Rao Lee amp Davis 2002 Overmyer Broscheamp Kangasjaumlrvi 2003 Tamaoki et al 2003 Vahala et al 2003see also Kangasjaumlrvi et al 2005 this issue) For exampleArabidopsis thaliana mutants that overproduce ethylene(eto1) or are insensitive to jasmonic acid (jar1) are moreextensively injured by O3 than wild-type plants (Overmyeret al 2000 Rao et al 2000 Tamaoki et al 2003) Ethyleneacts as a promoting factor for O3 injury whereas jasmonatesmight have a role in minimizing injury (Overmyer et al2000 2003)
Ozone effects at the tissue level are often characterizedas either acute or chronic responses subjective terms thatare related to the type of visible symptoms observed andwhether the O3 concentrations employed typify ambient O3
episodes For example unregulated cell death and pro-grammed cell death are usually considered acute responsesin which lesions occur within hours after exposure to rela-tively high O3 concentrations (typically gt150 nmol mol-1)In contrast chronic responses include lesions that developover days to weeks under lower O3 concentrations andaccelerated senescence where lesions might not formUnder field conditions accelerated senescence may be dif-ficult to identify in the absence of a clean air control orother reference point (eg O3-tolerant cultivar) because thecumulative effects on foliar senescence at the end of theseason are separated in time from the O3 events causingthe effects The timing of O3 episodes in relation to plantdevelopment can also have a major effect on the outcomeas in the case of soybean in which O3 exposure duringreproductive growth has a much greater impact on yieldcompared with exposure during vegetative growth (Heagleet al 1991 Miller et al 1991 Morgan et al 2003) The dis-tinction between acute and chronic responses is furthercomplicated by species and genotype differences that leadto a range of effects for a given combination of O3 concen-tration and environmental conditions
Genetic variation in O3 responses and potential O3 tolerance mechanisms
Studies of O3 effects on plants often identify genetic varia-tion as a contributing factor in the observed responseOzone-sensitive and tolerant cultivars or clones have beenreported for many plant species (Wellburn amp Wellburn1996) including crops such as soybean (Tingey Reinert amp
Carter 1972 Heagle amp Letchworth 1982 Robinson amp Britz2000) snap bean (Phaseolus vulgaris L) (Guzy amp Heath1993) tomato (Lycopersicon esculentum Miller) (Temple1990) clover (Trifolium repens L) (Heagle et al 1993)wheat (Heagle Miller amp Pursley 2000) and potato(Solanum tuberosum L) (Heagle Miller amp Pursley 2003)and natural vegetation such as black cherry (Prunus sero-tina Ehrh) (Lee et al 2002) and Plantago spp (Wolff Mor-gan-Richards amp Davison 2000) Studies have also shownthat O3 tolerance is a heritable trait (Damicone amp Manning1987 Reinert amp Eason 2000) Although much effort hasbeen made to identify the physiological and biochemicalelements of O3 tolerance no clear picture has emergedBased on the oxidative stress model formation of ROS andother oxidation products is the underlying process involvedin the generation and propagation of toxic compounds andabiotic elicitors in plants Therefore features of antioxidantmetabolism that scavenge or affect the perception of O3-derived ROS could contribute to O3 tolerance
Plants have evolved elaborate systems to combat generaloxidative stress based on enzymes that utilize ascorbic acidand glutathione (Noctor amp Foyer 1998 see also Foyer ampNoctor 2005 this issue) and these antioxidant systems havebeen linked to O3 stress tolerance (Runeckles amp Chevone1992 Smirnoff 1996 Chernikova et al 2000 Conklin ampBarth 2004) A critical role for ascorbic acid is demon-strated with the vtc1 mutant of Arabidopsis in which asignificant decrease in ascorbate content was associatedwith increased O3 sensitivity (Conklin Williams amp Last1996) A more detailed analysis of vtc1 suggested that inaddition to acting as an antioxidant ascorbic acid serves aregulatory function in the signalling networks that controlplant defence responses and leaf senescence (Conklin ampBarth 2004) Although it is clear that minimum levels ofascorbic acid and glutathione are required for normal leaffunction the concentrations of these antioxidant metabo-lites that naturally occur in leaf tissue are not always wellcorrelated with O3 tolerance (Guzy amp Heath 1993 Well-burn amp Wellburn 1996 Burkey et al 2000) suggesting amore complex mechanism that remains to be elucidated
Recent studies have found higher levels of ascorbic acidin the leaf apoplast of certain O3-tolerant snap bean geno-types compared with sensitive lines (Burkey Eason amp Fis-cus 2003) suggesting that localization of ascorbic acid inthe cell wall where initial O3 reactions occur might beimportant Leaf extracellular ascorbate content and redoxstatus are affected by O3 treatment (Castillo amp Greppin1988 Luwe amp Heber 1995 Burkey 1999) evidence thatextracellular ascorbic acid is involved in detoxification ofO3 and related ROS Ascorbic acid is initially synthesizedinside cells (Smirnoff Conklin amp Loewus 2001) followedby transport between the cytoplasm and extracellular spacevia specific carriers located in the plasma membrane (Hore-mans Foyer amp Asard 2000) A model developed by Cha-meides (1989) and expanded by others (Ploumlchl et al 2000Turcsanyi et al 2000) features ascorbic acid in the leaf apo-plast as a critical factor in the detoxification of extracellularO3 and ROS protecting plasma membranes from oxidative
1002 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
damage and preventing O3 injury Ascorbic acid potentiallycould serve as either a direct chemical scavenger of O3although this mechanism has been questioned (Jakob ampHeber 1998) or as a substrate for extracellular enzymes(eg ascorbate peroxidase) that attenuate ROS levels andthus affect the propagation of the initial O3 signal How-ever leaf apoplast ascorbic acid levels can be quite low incertain plant species [summarized in Burkey et al (2003)]suggesting that extracellular scavenging of O3 and ROS byascorbate may not be a significant factor in all plants Othercompounds with antioxidant properties also exist in the leafapoplast Identification of these compounds and theirimpact on ROS formation and propagation are subjects forfuture research on O3 tolerance mechanisms
Another approach to understanding O3 responses inplants involves investigations of the potential interactionbetween chemical signals and specific receptors in theplasma membrane Booker et al (2004) showed that theepinasty frequently observed for wild-type Arabidopsisleaves after O3 treatment did not occur in gpa1 mutantplants lacking the alpha subunit of the heterotrimeric Gprotein complex suggesting that G-proteins may beinvolved in the transduction of O3-derived signals Thebinding of O3 or more likely one of the postulated O3 reac-tion products to a G-protein receptor could activate theprotein and initiate signal cascade processes that activatetarget proteins in this signal transduction system Howeverfurther studies will be required to define the possible roleG-proteins have in O3-induced signal transduction andpropagation It is also possible that G-proteins modulate O3
responses via hormone signal transduction processes Ineither case elements of G-protein pathways involved in O3
response represent potential molecular targets that mightbe manipulated to enhance O3 tolerance although this isspeculative at present The challenge will be to identifytargets unique to O3 so that other important stressresponses remain intact (eg plant defence responsesagainst pathogen attack) Finally it is likely that any pro-tective mechanism will come with some metabolic cost thatwill require evaluation to determine its net benefits
IMPACTS ON PHOTOSYNTHESIS
Net assimilation
Other than crop yield and visible injury photosynthesis hasbeen the most broadly studied aspect of plant responses toO3 Generally O3 exposure results in decreased photosyn-thetic carbon assimilation (Miller 1987b Runeckles ampChevone 1992 Pell et al 1997 Long amp Naidu 2002 Morganet al 2003) In many cases loss of assimilation capacity wasshown to be due primarily to reduced carboxylation effi-ciency directly related to loss of Rubisco activity Theselosses of activity are thought to be due to decreases inRubisco concentration in the leaves rather than a decreasein activation state Messenger RNA transcripts for theRubisco small subunit (rbcS) have been noted to declinewith O3 treatment in potato leaves (Glick et al 1995) sug-
gesting that protein synthesis might be inhibited a notionconsistent with observations that Rubisco content in poplar(Populus spp) leaves treated with O3 does not rise as highas controls during leaf expansion (Pell et al 1997) How-ever Eckardt amp Pell (1994) under conditions where addi-tional synthesis should be minimal showed a substantialdecline in Rubisco content suggesting that the primarycause of the decline in Rubisco due to O3 exposure wasenhanced degradation rather than reduced production
Because photosynthetic measurements are typicallymade on the most recently fully expanded upper canopyleaves in determinate plants including many soybean linesit is often difficult to demonstrate differences in net photo-synthesis (An) due to O3 until near or after the time whenproduction of new upper canopy leaves has ceased Thenduring most of the reproductive stages An decreased earlierthan in control plants (accelerated senescence) This sug-gests that during vegetative growth either Rubisco synthe-sis in newly formed upper canopy leaves is not substantiallyaffected or that these leaves have not accumulated suffi-cient damage to elicit a response Vozzo et al (1995) dem-onstrated significant reductions in An on somewhat oldersoybean leaves that were relatively unaffected by shadingbut had received sufficient exposure to exhibit visible symp-toms thus lending support to this idea In contrast Morganet al (2004) using an indeterminate line were unable todemonstrate any differences due to dose accumulation in acohort of soybean leaves that developed during vegetativegrowth However they did observe decreased An in anothercohort that developed during reproductive growth andremained near the top of the canopy These results couldwell have been due to differences in exposure methodolo-gies particularly the greater atmospheric coupling to thelower canopy in OTCs as compared to FACE systems Infurther contrast to the above observations additionalreview of the data in two earlier soybean studies (Reid ampFiscus 1998 Reid et al 1998) illustrated in Fig 2 showsthat Rubisco content and activity chlorophyll content andAn were reduced by O3 early in the vegetative stage andthat all but An had reached or exceeded clean air levels byearly flowering Afterwards while chlorophyll content Anand Rubisco activity remained level or declined slightlyRubisco content continued to rise in the O3 treatmentFinally in early seed development all these parametersbegan to decline more precipitously Throughout this wholeexperiment chlorophyll content and Rubisco activitytracked each other very well This suggests that from earlyflowering through most of seed development O3 stimulatedRubisco content on a leaf area basis In addition the ques-tion of the role of accelerated senescence especially duringreproductive growth is one deserving more attention Forexample in Fig 2 the O3-induced decline in An duringreproduction may be due to the normal process of senes-cence that has been accelerated by O3 exposure or it maybe due simply to the loss of photosynthetically active leafarea due to accumulated O3 damage Thus especially inareas associated with visible lesions one would measure amixture of live and dead or dying cells and it seems rele-
Crop responses to ozone 1003
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
vant to know whether the remaining live cells are function-ing at or near normal levels for their age This possibility issuggested by the fact that from beginning of pod fill throughseed maturation when An per unit area is declining in thisstudy An per unit Rubisco remained high at 91 of the CFcontrol (difference not significant and data not shown)Further information is necessary to clarify this Anotherpoint deserving attention and bearing on the exposureaccumulation comments earlier in the paragraph is the sub-stantially reduced An early in the vegetative period Thismay have been caused by some inherent sensitivity of theseleaves or be the result of enhanced atmospheric couplingdue to the more open nature of the canopy at this growthstage
Photosynthetic electron transport
Detrimental effects of O3 on photosynthetic electron trans-port has received increased attention in recent years andwhile earlier it had been difficult to demonstrate significantdeclines in function evidence is accumulating that in someplants there are demonstrable effects both on PSII func-tion as well as the xanthophyll cycle components Forinstance significant and substantial declines in FvFm whichrepresents the efficiency of excitation energy capture byPSII in dark-adapted leaves have been reported for pump-kin (Cucurbita pepo L) (Ciompi et al 1997 Castagna et al2001) wheat (T durum and T aestivum) (Reichenauer et al1998) bean (P vulgaris L) (Guidi Di Cagno amp Soladatini2000a Guidi Tonini amp Soladatini 2000b Leipner Oxbor-ough amp Baker 2001 Guidi DeglrsquoInnocenti amp Soldatini2002) tomato (Calatayud amp Barreno 2001) and Festucapratensis and turnip (Brassica napus L) (Plazek Rapacz ampSkoczowski 2000) Of these particular studies three dem-
onstrated an increase in Fo which suggested damaged ordeactivated PSII centers (Guidi et al 2000a Leipner et al2001 Guidi et al 2002) all occurring in bean In additionlight saturated photosynthesis (ASAT) decreased in bean(Guidi et al 2000a Guidi et al 2002) pumpkin (Ciompiet al 1997 Castagna et al 2001) and wheat (Reichenaueret al 1998) although in the latter the effect was confined toolder leaves in two of the three lines and was partiallyreversible in some lines and fully reversible overnight inone It was unknown whether reversibility was due to anight-time epoxidation of the xanthophyll cycle decreasingsubsequent thermal quenching of PSII or to overnightrepair of damaged PSII centres This latter point becomesrelevant since a decrease in FvFm may be caused by both arelatively fast reversible down-regulation or a reversibledestruction (Osmond 1994) However the two processesmight be distinguished by monitoring effects on Fo in thatthermal dissipation by the xanthophyll cycle would lowerFo whereas damage to PSII would increase Fo as was foundin bean (Guidi et al 2000a 2002 Leipner et al 2001) EPRmeasurements also showed increases in free radicals whichReichenauer et al (1998) suggested might contribute todamage of chloroplast membranes in the sensitive wheatlines Runeckles amp Vaartnou (1997) also observed light-dependent EPR signals in bluegrass (Poa pratensis L)ryegrass (Lolium perenne L) and radish (Raphanus sativaL) consistent with superoxide anion characteristics onexposure to O3 The light dependence suggested that thesignal was localized in the chloroplast and significantly itsappearance was reduced in leaves with increased levels ofapoplastic ascorbic acid
High light is another factor that might interact with O3
in such a way as to increase or aggravate the effects of O3
alone through the process of photoinhibition with a result-
Figure 2 Rubisco quantity and activity An and chlorophyll content ratios for soybean (cv Essex) leaves treated with charcoal-filtered air (CF) 25 nmol O3 mol-1 and 15 yen ambient O3 (75 nmol mol-1 12 h average) in open-top field chambers in 1994 The ratios represent the response to O3 and are calculated as the quantity present in the O3 treatment compared with the control (O3CF) Developmental stages are given in the bar above the graph Data are from Reid et al (1998) and Reid amp Fiscus (1998)
1004 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
ant increase in free radicals in the chloroplast (Massman2004) Guidi et al (2002) demonstrated that both high lightand O3 decreased FvFm and increased Fo Furthermore thecombined effects were additive Farage et al (1991) uponexposing wheat leaves to high [O3] (200 and 400 nmolmol-1) could detect no difference in atrazine binding inisolated thylakoids nor any difference in D1 protein asindicated by western blots using D1-specific antibodiesHowever Godde amp Buchhold (1992) found that the turn-over rate for D1 was increased by exposure of spruce nee-dles to O3 suggesting both increased degradation andsynthesis In addition Ciompi et al (1997) found that inyoung leaves of pumpkin engagement of the xanthophyllcycle appeared to counteract the effects of O3 on the PSIIreaction center preventing a decline in D1 content How-ever in mature leaves even though there was activation ofthe xanthophyll cycle the effect appeared inadequate toprevent photoinhibition and loss of D1 protein The authorssuggested that this effect in mature leaves may have beendue to the normal decline in antioxidant activity in olderleaves Clearly high light has the potential to induce photo-inhibition which could be already sensitized by O3 exposurein field crops or inversely and is an area that requiresfurther investigation especially under more realistic fieldconditions
Direct effects on stomata
Another possible point of attack by O3 that cannot beignored is the guard cells Although much of the evidenceto date indicates that reduced gs is the result and not thecause of reduced assimilation (McKee et al 1995 Fiscuset al 1997 Long amp Naidu 2002) as suggested anddeduced on the basis of water use efficiency studies(Reich Schoettle amp Amundsen 1985) there is someintriguing work being done on possible direct effects ofO3 on stomatal response which links oxidative stress inthe form of H2O2 Ca2+ homeostasis of guard cells andABA response This work was recently reviewed byMcAinsh et al (2002) while the review by Schroeder et al(2001) provides a more detailed context for these studiesThe studies showed that H2O2 stimulated increased wholeplant cytosolic free Ca ions [Ca2+]cyt The stimulation washighly dose dependent with biphasic kinetics similar tothe Ca signature induced by exposure to O3 which wasalso highly dose dependent Using various techniquesincreased [Ca2+]cyt was also demonstrated in guard cells inresponse to both O3 and H2O2 In addition Pei et al(2000) demonstrated H2O2-induced stomatal closure andthat ABA induces H2O2 production in guard cells andactivates plasma membrane Ca-permeable channels Allof these responses were disrupted in the ABA-insensitiveArabidopsis mutant gca2 Taken together these and otherstudies reviewed (McAinsh et al 2002) raise the possibil-ity that O3 might directly affect stomata through theaction of H2O2 formed when O3 enters solution on theCa-permeable channels in the guard cell plasma mem-branes
O3ndashCO2 interactions on biomass and yield
Along with O3 atmospheric CO2 concentrations are alsorising and numerous studies indicate that elevated CO2
might enhance the productivity of current cropping sys-tems Long et al (2004) have shown that the magnitude ofthe response to elevated CO2 is highly variable not onlyacross but within species and cultivars Although thereappears to be a genetic component to this variability someof it may be due to environmental stresses that occurredduring plant growth Ozone is one such possible stress andit would not be surprising that there are significant physio-logical interactions between elevated CO2 and O3 The mostrelevant interaction for present purposes is the reductionin gs at elevated levels of CO2 that has the effect of prevent-ing reductions of An growth and yield in many crops grownin the presence of toxic levels of O3 (Fiscus et al 1997Olszyk et al 2000 Fiscus et al 2002 Morgan et al 2003Booker et al 2005) Figure 3 illustrates this interaction insoybean grown in both elevated CO2 and O3 (compare withAn ratio in Fig 2) and although it is expected that theassimilation ratios in elevated CO2 are substantially higherthan the control it is interesting to note that elevated CO2
did not seem to prevent the accelerated senescence charac-teristic of an O3 fumigation at current levels of CO2 Anal-ysis of compiled shoot biomass and yield data (Fiscus et al2002) show that increased An does get translated into anaverage 30 increase in biomass but often not into yieldin clean (charcoal-filtered) air (Fig 4) (Fiscus et al 2001)The variability in both biomass and yield is large with yieldsin doubled CO2 ranging from -37 to +41 in CF air witha mean of +6 across the crops studied However inincreasing O3 concentrations both the apparent biomassand yield due to elevated CO2 (CO2 fertilization effect)increased dramatically but the variability of the responsesremained large Of course in elevated CO2 gs will bereduced typically by 30 in these studies so O3 flux intothe leaves and the effective exposure is much reduced Thusthe apparent dramatic yield increases in some non-filteredair experiments might be caused in part by the fact thatyields are already suppressed by 5ndash15 by O3 in ambientair control plants and the best that elevated CO2 seemedto do in these experiments was to restore yields to the cleanair levels The wide range of variability in these experimentssuggests that in some CO2 was not the limiting resourceduring reproduction In fact it seems possible given thelarge increases in vegetative biomass that early resourcedepletion under elevated CO2 actually may have reducedyield One would expect that any environmental stress thataffects gs (eg drought) would also influence the directeffects of O3 and lead to large apparent CO2-stimulatedincreases in growth and yield The cause of the distinctvariability shown in these experiments is an area thatdeserves further research
Impacts on translocation and partitioning
Early in the season chronic exposure to O3 can inhibit An
(Fig 2) and plant growth (Fig 5) but often the decline in
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

1002 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
damage and preventing O3 injury Ascorbic acid potentiallycould serve as either a direct chemical scavenger of O3although this mechanism has been questioned (Jakob ampHeber 1998) or as a substrate for extracellular enzymes(eg ascorbate peroxidase) that attenuate ROS levels andthus affect the propagation of the initial O3 signal How-ever leaf apoplast ascorbic acid levels can be quite low incertain plant species [summarized in Burkey et al (2003)]suggesting that extracellular scavenging of O3 and ROS byascorbate may not be a significant factor in all plants Othercompounds with antioxidant properties also exist in the leafapoplast Identification of these compounds and theirimpact on ROS formation and propagation are subjects forfuture research on O3 tolerance mechanisms
Another approach to understanding O3 responses inplants involves investigations of the potential interactionbetween chemical signals and specific receptors in theplasma membrane Booker et al (2004) showed that theepinasty frequently observed for wild-type Arabidopsisleaves after O3 treatment did not occur in gpa1 mutantplants lacking the alpha subunit of the heterotrimeric Gprotein complex suggesting that G-proteins may beinvolved in the transduction of O3-derived signals Thebinding of O3 or more likely one of the postulated O3 reac-tion products to a G-protein receptor could activate theprotein and initiate signal cascade processes that activatetarget proteins in this signal transduction system Howeverfurther studies will be required to define the possible roleG-proteins have in O3-induced signal transduction andpropagation It is also possible that G-proteins modulate O3
responses via hormone signal transduction processes Ineither case elements of G-protein pathways involved in O3
response represent potential molecular targets that mightbe manipulated to enhance O3 tolerance although this isspeculative at present The challenge will be to identifytargets unique to O3 so that other important stressresponses remain intact (eg plant defence responsesagainst pathogen attack) Finally it is likely that any pro-tective mechanism will come with some metabolic cost thatwill require evaluation to determine its net benefits
IMPACTS ON PHOTOSYNTHESIS
Net assimilation
Other than crop yield and visible injury photosynthesis hasbeen the most broadly studied aspect of plant responses toO3 Generally O3 exposure results in decreased photosyn-thetic carbon assimilation (Miller 1987b Runeckles ampChevone 1992 Pell et al 1997 Long amp Naidu 2002 Morganet al 2003) In many cases loss of assimilation capacity wasshown to be due primarily to reduced carboxylation effi-ciency directly related to loss of Rubisco activity Theselosses of activity are thought to be due to decreases inRubisco concentration in the leaves rather than a decreasein activation state Messenger RNA transcripts for theRubisco small subunit (rbcS) have been noted to declinewith O3 treatment in potato leaves (Glick et al 1995) sug-
gesting that protein synthesis might be inhibited a notionconsistent with observations that Rubisco content in poplar(Populus spp) leaves treated with O3 does not rise as highas controls during leaf expansion (Pell et al 1997) How-ever Eckardt amp Pell (1994) under conditions where addi-tional synthesis should be minimal showed a substantialdecline in Rubisco content suggesting that the primarycause of the decline in Rubisco due to O3 exposure wasenhanced degradation rather than reduced production
Because photosynthetic measurements are typicallymade on the most recently fully expanded upper canopyleaves in determinate plants including many soybean linesit is often difficult to demonstrate differences in net photo-synthesis (An) due to O3 until near or after the time whenproduction of new upper canopy leaves has ceased Thenduring most of the reproductive stages An decreased earlierthan in control plants (accelerated senescence) This sug-gests that during vegetative growth either Rubisco synthe-sis in newly formed upper canopy leaves is not substantiallyaffected or that these leaves have not accumulated suffi-cient damage to elicit a response Vozzo et al (1995) dem-onstrated significant reductions in An on somewhat oldersoybean leaves that were relatively unaffected by shadingbut had received sufficient exposure to exhibit visible symp-toms thus lending support to this idea In contrast Morganet al (2004) using an indeterminate line were unable todemonstrate any differences due to dose accumulation in acohort of soybean leaves that developed during vegetativegrowth However they did observe decreased An in anothercohort that developed during reproductive growth andremained near the top of the canopy These results couldwell have been due to differences in exposure methodolo-gies particularly the greater atmospheric coupling to thelower canopy in OTCs as compared to FACE systems Infurther contrast to the above observations additionalreview of the data in two earlier soybean studies (Reid ampFiscus 1998 Reid et al 1998) illustrated in Fig 2 showsthat Rubisco content and activity chlorophyll content andAn were reduced by O3 early in the vegetative stage andthat all but An had reached or exceeded clean air levels byearly flowering Afterwards while chlorophyll content Anand Rubisco activity remained level or declined slightlyRubisco content continued to rise in the O3 treatmentFinally in early seed development all these parametersbegan to decline more precipitously Throughout this wholeexperiment chlorophyll content and Rubisco activitytracked each other very well This suggests that from earlyflowering through most of seed development O3 stimulatedRubisco content on a leaf area basis In addition the ques-tion of the role of accelerated senescence especially duringreproductive growth is one deserving more attention Forexample in Fig 2 the O3-induced decline in An duringreproduction may be due to the normal process of senes-cence that has been accelerated by O3 exposure or it maybe due simply to the loss of photosynthetically active leafarea due to accumulated O3 damage Thus especially inareas associated with visible lesions one would measure amixture of live and dead or dying cells and it seems rele-
Crop responses to ozone 1003
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
vant to know whether the remaining live cells are function-ing at or near normal levels for their age This possibility issuggested by the fact that from beginning of pod fill throughseed maturation when An per unit area is declining in thisstudy An per unit Rubisco remained high at 91 of the CFcontrol (difference not significant and data not shown)Further information is necessary to clarify this Anotherpoint deserving attention and bearing on the exposureaccumulation comments earlier in the paragraph is the sub-stantially reduced An early in the vegetative period Thismay have been caused by some inherent sensitivity of theseleaves or be the result of enhanced atmospheric couplingdue to the more open nature of the canopy at this growthstage
Photosynthetic electron transport
Detrimental effects of O3 on photosynthetic electron trans-port has received increased attention in recent years andwhile earlier it had been difficult to demonstrate significantdeclines in function evidence is accumulating that in someplants there are demonstrable effects both on PSII func-tion as well as the xanthophyll cycle components Forinstance significant and substantial declines in FvFm whichrepresents the efficiency of excitation energy capture byPSII in dark-adapted leaves have been reported for pump-kin (Cucurbita pepo L) (Ciompi et al 1997 Castagna et al2001) wheat (T durum and T aestivum) (Reichenauer et al1998) bean (P vulgaris L) (Guidi Di Cagno amp Soladatini2000a Guidi Tonini amp Soladatini 2000b Leipner Oxbor-ough amp Baker 2001 Guidi DeglrsquoInnocenti amp Soldatini2002) tomato (Calatayud amp Barreno 2001) and Festucapratensis and turnip (Brassica napus L) (Plazek Rapacz ampSkoczowski 2000) Of these particular studies three dem-
onstrated an increase in Fo which suggested damaged ordeactivated PSII centers (Guidi et al 2000a Leipner et al2001 Guidi et al 2002) all occurring in bean In additionlight saturated photosynthesis (ASAT) decreased in bean(Guidi et al 2000a Guidi et al 2002) pumpkin (Ciompiet al 1997 Castagna et al 2001) and wheat (Reichenaueret al 1998) although in the latter the effect was confined toolder leaves in two of the three lines and was partiallyreversible in some lines and fully reversible overnight inone It was unknown whether reversibility was due to anight-time epoxidation of the xanthophyll cycle decreasingsubsequent thermal quenching of PSII or to overnightrepair of damaged PSII centres This latter point becomesrelevant since a decrease in FvFm may be caused by both arelatively fast reversible down-regulation or a reversibledestruction (Osmond 1994) However the two processesmight be distinguished by monitoring effects on Fo in thatthermal dissipation by the xanthophyll cycle would lowerFo whereas damage to PSII would increase Fo as was foundin bean (Guidi et al 2000a 2002 Leipner et al 2001) EPRmeasurements also showed increases in free radicals whichReichenauer et al (1998) suggested might contribute todamage of chloroplast membranes in the sensitive wheatlines Runeckles amp Vaartnou (1997) also observed light-dependent EPR signals in bluegrass (Poa pratensis L)ryegrass (Lolium perenne L) and radish (Raphanus sativaL) consistent with superoxide anion characteristics onexposure to O3 The light dependence suggested that thesignal was localized in the chloroplast and significantly itsappearance was reduced in leaves with increased levels ofapoplastic ascorbic acid
High light is another factor that might interact with O3
in such a way as to increase or aggravate the effects of O3
alone through the process of photoinhibition with a result-
Figure 2 Rubisco quantity and activity An and chlorophyll content ratios for soybean (cv Essex) leaves treated with charcoal-filtered air (CF) 25 nmol O3 mol-1 and 15 yen ambient O3 (75 nmol mol-1 12 h average) in open-top field chambers in 1994 The ratios represent the response to O3 and are calculated as the quantity present in the O3 treatment compared with the control (O3CF) Developmental stages are given in the bar above the graph Data are from Reid et al (1998) and Reid amp Fiscus (1998)
1004 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
ant increase in free radicals in the chloroplast (Massman2004) Guidi et al (2002) demonstrated that both high lightand O3 decreased FvFm and increased Fo Furthermore thecombined effects were additive Farage et al (1991) uponexposing wheat leaves to high [O3] (200 and 400 nmolmol-1) could detect no difference in atrazine binding inisolated thylakoids nor any difference in D1 protein asindicated by western blots using D1-specific antibodiesHowever Godde amp Buchhold (1992) found that the turn-over rate for D1 was increased by exposure of spruce nee-dles to O3 suggesting both increased degradation andsynthesis In addition Ciompi et al (1997) found that inyoung leaves of pumpkin engagement of the xanthophyllcycle appeared to counteract the effects of O3 on the PSIIreaction center preventing a decline in D1 content How-ever in mature leaves even though there was activation ofthe xanthophyll cycle the effect appeared inadequate toprevent photoinhibition and loss of D1 protein The authorssuggested that this effect in mature leaves may have beendue to the normal decline in antioxidant activity in olderleaves Clearly high light has the potential to induce photo-inhibition which could be already sensitized by O3 exposurein field crops or inversely and is an area that requiresfurther investigation especially under more realistic fieldconditions
Direct effects on stomata
Another possible point of attack by O3 that cannot beignored is the guard cells Although much of the evidenceto date indicates that reduced gs is the result and not thecause of reduced assimilation (McKee et al 1995 Fiscuset al 1997 Long amp Naidu 2002) as suggested anddeduced on the basis of water use efficiency studies(Reich Schoettle amp Amundsen 1985) there is someintriguing work being done on possible direct effects ofO3 on stomatal response which links oxidative stress inthe form of H2O2 Ca2+ homeostasis of guard cells andABA response This work was recently reviewed byMcAinsh et al (2002) while the review by Schroeder et al(2001) provides a more detailed context for these studiesThe studies showed that H2O2 stimulated increased wholeplant cytosolic free Ca ions [Ca2+]cyt The stimulation washighly dose dependent with biphasic kinetics similar tothe Ca signature induced by exposure to O3 which wasalso highly dose dependent Using various techniquesincreased [Ca2+]cyt was also demonstrated in guard cells inresponse to both O3 and H2O2 In addition Pei et al(2000) demonstrated H2O2-induced stomatal closure andthat ABA induces H2O2 production in guard cells andactivates plasma membrane Ca-permeable channels Allof these responses were disrupted in the ABA-insensitiveArabidopsis mutant gca2 Taken together these and otherstudies reviewed (McAinsh et al 2002) raise the possibil-ity that O3 might directly affect stomata through theaction of H2O2 formed when O3 enters solution on theCa-permeable channels in the guard cell plasma mem-branes
O3ndashCO2 interactions on biomass and yield
Along with O3 atmospheric CO2 concentrations are alsorising and numerous studies indicate that elevated CO2
might enhance the productivity of current cropping sys-tems Long et al (2004) have shown that the magnitude ofthe response to elevated CO2 is highly variable not onlyacross but within species and cultivars Although thereappears to be a genetic component to this variability someof it may be due to environmental stresses that occurredduring plant growth Ozone is one such possible stress andit would not be surprising that there are significant physio-logical interactions between elevated CO2 and O3 The mostrelevant interaction for present purposes is the reductionin gs at elevated levels of CO2 that has the effect of prevent-ing reductions of An growth and yield in many crops grownin the presence of toxic levels of O3 (Fiscus et al 1997Olszyk et al 2000 Fiscus et al 2002 Morgan et al 2003Booker et al 2005) Figure 3 illustrates this interaction insoybean grown in both elevated CO2 and O3 (compare withAn ratio in Fig 2) and although it is expected that theassimilation ratios in elevated CO2 are substantially higherthan the control it is interesting to note that elevated CO2
did not seem to prevent the accelerated senescence charac-teristic of an O3 fumigation at current levels of CO2 Anal-ysis of compiled shoot biomass and yield data (Fiscus et al2002) show that increased An does get translated into anaverage 30 increase in biomass but often not into yieldin clean (charcoal-filtered) air (Fig 4) (Fiscus et al 2001)The variability in both biomass and yield is large with yieldsin doubled CO2 ranging from -37 to +41 in CF air witha mean of +6 across the crops studied However inincreasing O3 concentrations both the apparent biomassand yield due to elevated CO2 (CO2 fertilization effect)increased dramatically but the variability of the responsesremained large Of course in elevated CO2 gs will bereduced typically by 30 in these studies so O3 flux intothe leaves and the effective exposure is much reduced Thusthe apparent dramatic yield increases in some non-filteredair experiments might be caused in part by the fact thatyields are already suppressed by 5ndash15 by O3 in ambientair control plants and the best that elevated CO2 seemedto do in these experiments was to restore yields to the cleanair levels The wide range of variability in these experimentssuggests that in some CO2 was not the limiting resourceduring reproduction In fact it seems possible given thelarge increases in vegetative biomass that early resourcedepletion under elevated CO2 actually may have reducedyield One would expect that any environmental stress thataffects gs (eg drought) would also influence the directeffects of O3 and lead to large apparent CO2-stimulatedincreases in growth and yield The cause of the distinctvariability shown in these experiments is an area thatdeserves further research
Impacts on translocation and partitioning
Early in the season chronic exposure to O3 can inhibit An
(Fig 2) and plant growth (Fig 5) but often the decline in
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005
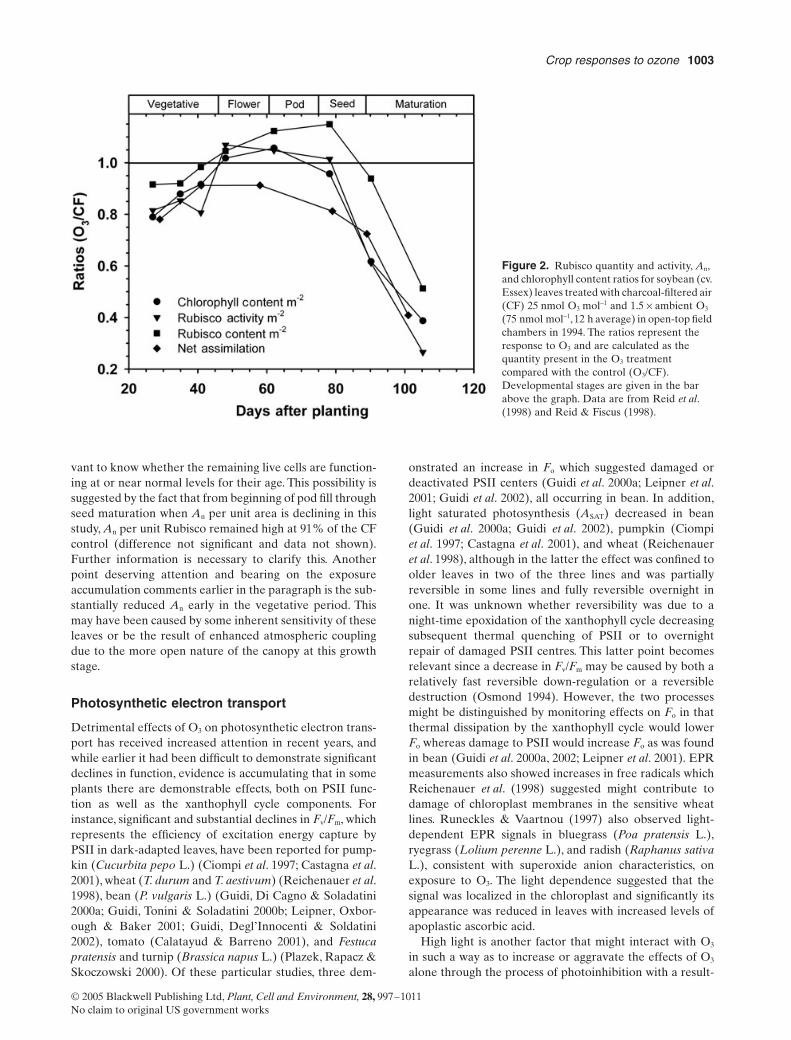
Crop responses to ozone 1003
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
vant to know whether the remaining live cells are function-ing at or near normal levels for their age This possibility issuggested by the fact that from beginning of pod fill throughseed maturation when An per unit area is declining in thisstudy An per unit Rubisco remained high at 91 of the CFcontrol (difference not significant and data not shown)Further information is necessary to clarify this Anotherpoint deserving attention and bearing on the exposureaccumulation comments earlier in the paragraph is the sub-stantially reduced An early in the vegetative period Thismay have been caused by some inherent sensitivity of theseleaves or be the result of enhanced atmospheric couplingdue to the more open nature of the canopy at this growthstage
Photosynthetic electron transport
Detrimental effects of O3 on photosynthetic electron trans-port has received increased attention in recent years andwhile earlier it had been difficult to demonstrate significantdeclines in function evidence is accumulating that in someplants there are demonstrable effects both on PSII func-tion as well as the xanthophyll cycle components Forinstance significant and substantial declines in FvFm whichrepresents the efficiency of excitation energy capture byPSII in dark-adapted leaves have been reported for pump-kin (Cucurbita pepo L) (Ciompi et al 1997 Castagna et al2001) wheat (T durum and T aestivum) (Reichenauer et al1998) bean (P vulgaris L) (Guidi Di Cagno amp Soladatini2000a Guidi Tonini amp Soladatini 2000b Leipner Oxbor-ough amp Baker 2001 Guidi DeglrsquoInnocenti amp Soldatini2002) tomato (Calatayud amp Barreno 2001) and Festucapratensis and turnip (Brassica napus L) (Plazek Rapacz ampSkoczowski 2000) Of these particular studies three dem-
onstrated an increase in Fo which suggested damaged ordeactivated PSII centers (Guidi et al 2000a Leipner et al2001 Guidi et al 2002) all occurring in bean In additionlight saturated photosynthesis (ASAT) decreased in bean(Guidi et al 2000a Guidi et al 2002) pumpkin (Ciompiet al 1997 Castagna et al 2001) and wheat (Reichenaueret al 1998) although in the latter the effect was confined toolder leaves in two of the three lines and was partiallyreversible in some lines and fully reversible overnight inone It was unknown whether reversibility was due to anight-time epoxidation of the xanthophyll cycle decreasingsubsequent thermal quenching of PSII or to overnightrepair of damaged PSII centres This latter point becomesrelevant since a decrease in FvFm may be caused by both arelatively fast reversible down-regulation or a reversibledestruction (Osmond 1994) However the two processesmight be distinguished by monitoring effects on Fo in thatthermal dissipation by the xanthophyll cycle would lowerFo whereas damage to PSII would increase Fo as was foundin bean (Guidi et al 2000a 2002 Leipner et al 2001) EPRmeasurements also showed increases in free radicals whichReichenauer et al (1998) suggested might contribute todamage of chloroplast membranes in the sensitive wheatlines Runeckles amp Vaartnou (1997) also observed light-dependent EPR signals in bluegrass (Poa pratensis L)ryegrass (Lolium perenne L) and radish (Raphanus sativaL) consistent with superoxide anion characteristics onexposure to O3 The light dependence suggested that thesignal was localized in the chloroplast and significantly itsappearance was reduced in leaves with increased levels ofapoplastic ascorbic acid
High light is another factor that might interact with O3
in such a way as to increase or aggravate the effects of O3
alone through the process of photoinhibition with a result-
Figure 2 Rubisco quantity and activity An and chlorophyll content ratios for soybean (cv Essex) leaves treated with charcoal-filtered air (CF) 25 nmol O3 mol-1 and 15 yen ambient O3 (75 nmol mol-1 12 h average) in open-top field chambers in 1994 The ratios represent the response to O3 and are calculated as the quantity present in the O3 treatment compared with the control (O3CF) Developmental stages are given in the bar above the graph Data are from Reid et al (1998) and Reid amp Fiscus (1998)
1004 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
ant increase in free radicals in the chloroplast (Massman2004) Guidi et al (2002) demonstrated that both high lightand O3 decreased FvFm and increased Fo Furthermore thecombined effects were additive Farage et al (1991) uponexposing wheat leaves to high [O3] (200 and 400 nmolmol-1) could detect no difference in atrazine binding inisolated thylakoids nor any difference in D1 protein asindicated by western blots using D1-specific antibodiesHowever Godde amp Buchhold (1992) found that the turn-over rate for D1 was increased by exposure of spruce nee-dles to O3 suggesting both increased degradation andsynthesis In addition Ciompi et al (1997) found that inyoung leaves of pumpkin engagement of the xanthophyllcycle appeared to counteract the effects of O3 on the PSIIreaction center preventing a decline in D1 content How-ever in mature leaves even though there was activation ofthe xanthophyll cycle the effect appeared inadequate toprevent photoinhibition and loss of D1 protein The authorssuggested that this effect in mature leaves may have beendue to the normal decline in antioxidant activity in olderleaves Clearly high light has the potential to induce photo-inhibition which could be already sensitized by O3 exposurein field crops or inversely and is an area that requiresfurther investigation especially under more realistic fieldconditions
Direct effects on stomata
Another possible point of attack by O3 that cannot beignored is the guard cells Although much of the evidenceto date indicates that reduced gs is the result and not thecause of reduced assimilation (McKee et al 1995 Fiscuset al 1997 Long amp Naidu 2002) as suggested anddeduced on the basis of water use efficiency studies(Reich Schoettle amp Amundsen 1985) there is someintriguing work being done on possible direct effects ofO3 on stomatal response which links oxidative stress inthe form of H2O2 Ca2+ homeostasis of guard cells andABA response This work was recently reviewed byMcAinsh et al (2002) while the review by Schroeder et al(2001) provides a more detailed context for these studiesThe studies showed that H2O2 stimulated increased wholeplant cytosolic free Ca ions [Ca2+]cyt The stimulation washighly dose dependent with biphasic kinetics similar tothe Ca signature induced by exposure to O3 which wasalso highly dose dependent Using various techniquesincreased [Ca2+]cyt was also demonstrated in guard cells inresponse to both O3 and H2O2 In addition Pei et al(2000) demonstrated H2O2-induced stomatal closure andthat ABA induces H2O2 production in guard cells andactivates plasma membrane Ca-permeable channels Allof these responses were disrupted in the ABA-insensitiveArabidopsis mutant gca2 Taken together these and otherstudies reviewed (McAinsh et al 2002) raise the possibil-ity that O3 might directly affect stomata through theaction of H2O2 formed when O3 enters solution on theCa-permeable channels in the guard cell plasma mem-branes
O3ndashCO2 interactions on biomass and yield
Along with O3 atmospheric CO2 concentrations are alsorising and numerous studies indicate that elevated CO2
might enhance the productivity of current cropping sys-tems Long et al (2004) have shown that the magnitude ofthe response to elevated CO2 is highly variable not onlyacross but within species and cultivars Although thereappears to be a genetic component to this variability someof it may be due to environmental stresses that occurredduring plant growth Ozone is one such possible stress andit would not be surprising that there are significant physio-logical interactions between elevated CO2 and O3 The mostrelevant interaction for present purposes is the reductionin gs at elevated levels of CO2 that has the effect of prevent-ing reductions of An growth and yield in many crops grownin the presence of toxic levels of O3 (Fiscus et al 1997Olszyk et al 2000 Fiscus et al 2002 Morgan et al 2003Booker et al 2005) Figure 3 illustrates this interaction insoybean grown in both elevated CO2 and O3 (compare withAn ratio in Fig 2) and although it is expected that theassimilation ratios in elevated CO2 are substantially higherthan the control it is interesting to note that elevated CO2
did not seem to prevent the accelerated senescence charac-teristic of an O3 fumigation at current levels of CO2 Anal-ysis of compiled shoot biomass and yield data (Fiscus et al2002) show that increased An does get translated into anaverage 30 increase in biomass but often not into yieldin clean (charcoal-filtered) air (Fig 4) (Fiscus et al 2001)The variability in both biomass and yield is large with yieldsin doubled CO2 ranging from -37 to +41 in CF air witha mean of +6 across the crops studied However inincreasing O3 concentrations both the apparent biomassand yield due to elevated CO2 (CO2 fertilization effect)increased dramatically but the variability of the responsesremained large Of course in elevated CO2 gs will bereduced typically by 30 in these studies so O3 flux intothe leaves and the effective exposure is much reduced Thusthe apparent dramatic yield increases in some non-filteredair experiments might be caused in part by the fact thatyields are already suppressed by 5ndash15 by O3 in ambientair control plants and the best that elevated CO2 seemedto do in these experiments was to restore yields to the cleanair levels The wide range of variability in these experimentssuggests that in some CO2 was not the limiting resourceduring reproduction In fact it seems possible given thelarge increases in vegetative biomass that early resourcedepletion under elevated CO2 actually may have reducedyield One would expect that any environmental stress thataffects gs (eg drought) would also influence the directeffects of O3 and lead to large apparent CO2-stimulatedincreases in growth and yield The cause of the distinctvariability shown in these experiments is an area thatdeserves further research
Impacts on translocation and partitioning
Early in the season chronic exposure to O3 can inhibit An
(Fig 2) and plant growth (Fig 5) but often the decline in
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

1004 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
ant increase in free radicals in the chloroplast (Massman2004) Guidi et al (2002) demonstrated that both high lightand O3 decreased FvFm and increased Fo Furthermore thecombined effects were additive Farage et al (1991) uponexposing wheat leaves to high [O3] (200 and 400 nmolmol-1) could detect no difference in atrazine binding inisolated thylakoids nor any difference in D1 protein asindicated by western blots using D1-specific antibodiesHowever Godde amp Buchhold (1992) found that the turn-over rate for D1 was increased by exposure of spruce nee-dles to O3 suggesting both increased degradation andsynthesis In addition Ciompi et al (1997) found that inyoung leaves of pumpkin engagement of the xanthophyllcycle appeared to counteract the effects of O3 on the PSIIreaction center preventing a decline in D1 content How-ever in mature leaves even though there was activation ofthe xanthophyll cycle the effect appeared inadequate toprevent photoinhibition and loss of D1 protein The authorssuggested that this effect in mature leaves may have beendue to the normal decline in antioxidant activity in olderleaves Clearly high light has the potential to induce photo-inhibition which could be already sensitized by O3 exposurein field crops or inversely and is an area that requiresfurther investigation especially under more realistic fieldconditions
Direct effects on stomata
Another possible point of attack by O3 that cannot beignored is the guard cells Although much of the evidenceto date indicates that reduced gs is the result and not thecause of reduced assimilation (McKee et al 1995 Fiscuset al 1997 Long amp Naidu 2002) as suggested anddeduced on the basis of water use efficiency studies(Reich Schoettle amp Amundsen 1985) there is someintriguing work being done on possible direct effects ofO3 on stomatal response which links oxidative stress inthe form of H2O2 Ca2+ homeostasis of guard cells andABA response This work was recently reviewed byMcAinsh et al (2002) while the review by Schroeder et al(2001) provides a more detailed context for these studiesThe studies showed that H2O2 stimulated increased wholeplant cytosolic free Ca ions [Ca2+]cyt The stimulation washighly dose dependent with biphasic kinetics similar tothe Ca signature induced by exposure to O3 which wasalso highly dose dependent Using various techniquesincreased [Ca2+]cyt was also demonstrated in guard cells inresponse to both O3 and H2O2 In addition Pei et al(2000) demonstrated H2O2-induced stomatal closure andthat ABA induces H2O2 production in guard cells andactivates plasma membrane Ca-permeable channels Allof these responses were disrupted in the ABA-insensitiveArabidopsis mutant gca2 Taken together these and otherstudies reviewed (McAinsh et al 2002) raise the possibil-ity that O3 might directly affect stomata through theaction of H2O2 formed when O3 enters solution on theCa-permeable channels in the guard cell plasma mem-branes
O3ndashCO2 interactions on biomass and yield
Along with O3 atmospheric CO2 concentrations are alsorising and numerous studies indicate that elevated CO2
might enhance the productivity of current cropping sys-tems Long et al (2004) have shown that the magnitude ofthe response to elevated CO2 is highly variable not onlyacross but within species and cultivars Although thereappears to be a genetic component to this variability someof it may be due to environmental stresses that occurredduring plant growth Ozone is one such possible stress andit would not be surprising that there are significant physio-logical interactions between elevated CO2 and O3 The mostrelevant interaction for present purposes is the reductionin gs at elevated levels of CO2 that has the effect of prevent-ing reductions of An growth and yield in many crops grownin the presence of toxic levels of O3 (Fiscus et al 1997Olszyk et al 2000 Fiscus et al 2002 Morgan et al 2003Booker et al 2005) Figure 3 illustrates this interaction insoybean grown in both elevated CO2 and O3 (compare withAn ratio in Fig 2) and although it is expected that theassimilation ratios in elevated CO2 are substantially higherthan the control it is interesting to note that elevated CO2
did not seem to prevent the accelerated senescence charac-teristic of an O3 fumigation at current levels of CO2 Anal-ysis of compiled shoot biomass and yield data (Fiscus et al2002) show that increased An does get translated into anaverage 30 increase in biomass but often not into yieldin clean (charcoal-filtered) air (Fig 4) (Fiscus et al 2001)The variability in both biomass and yield is large with yieldsin doubled CO2 ranging from -37 to +41 in CF air witha mean of +6 across the crops studied However inincreasing O3 concentrations both the apparent biomassand yield due to elevated CO2 (CO2 fertilization effect)increased dramatically but the variability of the responsesremained large Of course in elevated CO2 gs will bereduced typically by 30 in these studies so O3 flux intothe leaves and the effective exposure is much reduced Thusthe apparent dramatic yield increases in some non-filteredair experiments might be caused in part by the fact thatyields are already suppressed by 5ndash15 by O3 in ambientair control plants and the best that elevated CO2 seemedto do in these experiments was to restore yields to the cleanair levels The wide range of variability in these experimentssuggests that in some CO2 was not the limiting resourceduring reproduction In fact it seems possible given thelarge increases in vegetative biomass that early resourcedepletion under elevated CO2 actually may have reducedyield One would expect that any environmental stress thataffects gs (eg drought) would also influence the directeffects of O3 and lead to large apparent CO2-stimulatedincreases in growth and yield The cause of the distinctvariability shown in these experiments is an area thatdeserves further research
Impacts on translocation and partitioning
Early in the season chronic exposure to O3 can inhibit An
(Fig 2) and plant growth (Fig 5) but often the decline in
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005
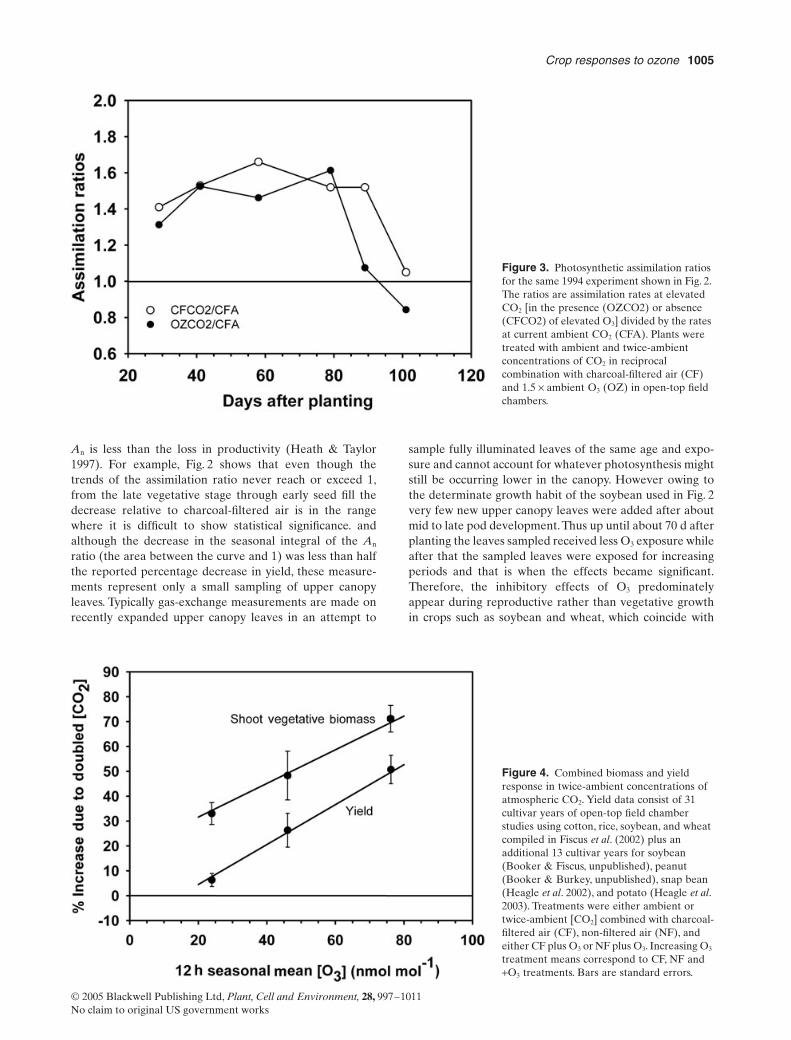
Crop responses to ozone 1005
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
An is less than the loss in productivity (Heath amp Taylor1997) For example Fig 2 shows that even though thetrends of the assimilation ratio never reach or exceed 1from the late vegetative stage through early seed fill thedecrease relative to charcoal-filtered air is in the rangewhere it is difficult to show statistical significance andalthough the decrease in the seasonal integral of the An
ratio (the area between the curve and 1) was less than halfthe reported percentage decrease in yield these measure-ments represent only a small sampling of upper canopyleaves Typically gas-exchange measurements are made onrecently expanded upper canopy leaves in an attempt to
sample fully illuminated leaves of the same age and expo-sure and cannot account for whatever photosynthesis mightstill be occurring lower in the canopy However owing tothe determinate growth habit of the soybean used in Fig 2very few new upper canopy leaves were added after aboutmid to late pod development Thus up until about 70 d afterplanting the leaves sampled received less O3 exposure whileafter that the sampled leaves were exposed for increasingperiods and that is when the effects became significantTherefore the inhibitory effects of O3 predominatelyappear during reproductive rather than vegetative growthin crops such as soybean and wheat which coincide with
Figure 3 Photosynthetic assimilation ratios for the same 1994 experiment shown in Fig 2 The ratios are assimilation rates at elevated CO2 [in the presence (OZCO2) or absence (CFCO2) of elevated O3] divided by the rates at current ambient CO2 (CFA) Plants were treated with ambient and twice-ambient concentrations of CO2 in reciprocal combination with charcoal-filtered air (CF) and 15 yen ambient O3 (OZ) in open-top field chambers
Figure 4 Combined biomass and yield response in twice-ambient concentrations of atmospheric CO2 Yield data consist of 31 cultivar years of open-top field chamber studies using cotton rice soybean and wheat compiled in Fiscus et al (2002) plus an additional 13 cultivar years for soybean (Booker amp Fiscus unpublished) peanut (Booker amp Burkey unpublished) snap bean (Heagle et al 2002) and potato (Heagle et al 2003) Treatments were either ambient or twice-ambient [CO2] combined with charcoal-filtered air (CF) non-filtered air (NF) and either CF plus O3 or NF plus O3 Increasing O3 treatment means correspond to CF NF and +O3 treatments Bars are standard errors
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005
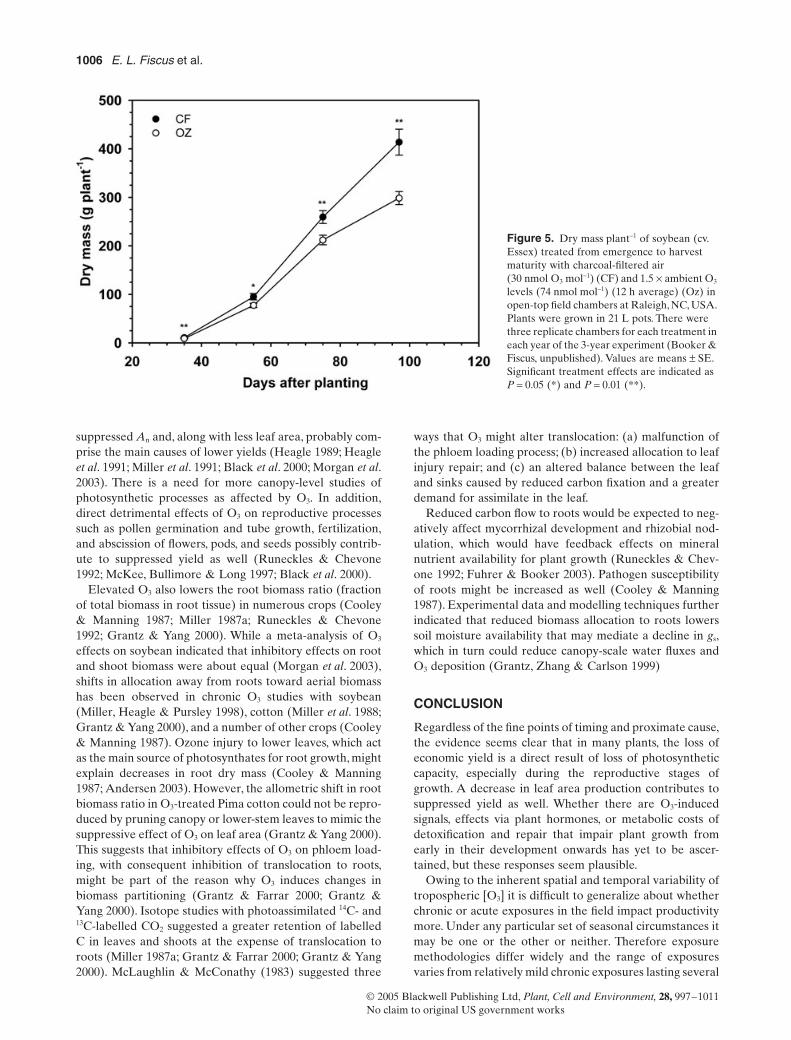
1006 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
suppressed An and along with less leaf area probably com-prise the main causes of lower yields (Heagle 1989 Heagleet al 1991 Miller et al 1991 Black et al 2000 Morgan et al2003) There is a need for more canopy-level studies ofphotosynthetic processes as affected by O3 In additiondirect detrimental effects of O3 on reproductive processessuch as pollen germination and tube growth fertilizationand abscission of flowers pods and seeds possibly contrib-ute to suppressed yield as well (Runeckles amp Chevone1992 McKee Bullimore amp Long 1997 Black et al 2000)
Elevated O3 also lowers the root biomass ratio (fractionof total biomass in root tissue) in numerous crops (Cooleyamp Manning 1987 Miller 1987a Runeckles amp Chevone1992 Grantz amp Yang 2000) While a meta-analysis of O3
effects on soybean indicated that inhibitory effects on rootand shoot biomass were about equal (Morgan et al 2003)shifts in allocation away from roots toward aerial biomasshas been observed in chronic O3 studies with soybean(Miller Heagle amp Pursley 1998) cotton (Miller et al 1988Grantz amp Yang 2000) and a number of other crops (Cooleyamp Manning 1987) Ozone injury to lower leaves which actas the main source of photosynthates for root growth mightexplain decreases in root dry mass (Cooley amp Manning1987 Andersen 2003) However the allometric shift in rootbiomass ratio in O3-treated Pima cotton could not be repro-duced by pruning canopy or lower-stem leaves to mimic thesuppressive effect of O3 on leaf area (Grantz amp Yang 2000)This suggests that inhibitory effects of O3 on phloem load-ing with consequent inhibition of translocation to rootsmight be part of the reason why O3 induces changes inbiomass partitioning (Grantz amp Farrar 2000 Grantz ampYang 2000) Isotope studies with photoassimilated 14C- and13C-labelled CO2 suggested a greater retention of labelledC in leaves and shoots at the expense of translocation toroots (Miller 1987a Grantz amp Farrar 2000 Grantz amp Yang2000) McLaughlin amp McConathy (1983) suggested three
ways that O3 might alter translocation (a) malfunction ofthe phloem loading process (b) increased allocation to leafinjury repair and (c) an altered balance between the leafand sinks caused by reduced carbon fixation and a greaterdemand for assimilate in the leaf
Reduced carbon flow to roots would be expected to neg-atively affect mycorrhizal development and rhizobial nod-ulation which would have feedback effects on mineralnutrient availability for plant growth (Runeckles amp Chev-one 1992 Fuhrer amp Booker 2003) Pathogen susceptibilityof roots might be increased as well (Cooley amp Manning1987) Experimental data and modelling techniques furtherindicated that reduced biomass allocation to roots lowerssoil moisture availability that may mediate a decline in gswhich in turn could reduce canopy-scale water fluxes andO3 deposition (Grantz Zhang amp Carlson 1999)
CONCLUSION
Regardless of the fine points of timing and proximate causethe evidence seems clear that in many plants the loss ofeconomic yield is a direct result of loss of photosyntheticcapacity especially during the reproductive stages ofgrowth A decrease in leaf area production contributes tosuppressed yield as well Whether there are O3-inducedsignals effects via plant hormones or metabolic costs ofdetoxification and repair that impair plant growth fromearly in their development onwards has yet to be ascer-tained but these responses seem plausible
Owing to the inherent spatial and temporal variability oftropospheric [O3] it is difficult to generalize about whetherchronic or acute exposures in the field impact productivitymore Under any particular set of seasonal circumstances itmay be one or the other or neither Therefore exposuremethodologies differ widely and the range of exposuresvaries from relatively mild chronic exposures lasting several
Figure 5 Dry mass plant-1 of soybean (cv Essex) treated from emergence to harvest maturity with charcoal-filtered air (30 nmol O3 mol-1) (CF) and 15 yen ambient O3 levels (74 nmol mol-1) (12 h average) (Oz) in open-top field chambers at Raleigh NC USA Plants were grown in 21 L pots There were three replicate chambers for each treatment in each year of the 3-year experiment (Booker amp Fiscus unpublished) Values are means plusmn SE Significant treatment effects are indicated as P = 005 () and P = 001 ()
Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

Crop responses to ozone 1007
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
days weeks or the entire growing season to acute exposureslasting from a few hours to a few days Experimental objec-tives will determine the preferred approach and whenassessing productivity losses in crops or natural systems itmay even be desirable to combine the two overlaying acuteexposures at developmental periods suspected of beingparticularly sensitive But acute exposures alone may bemore revealing for uncovering damage mechanisms espe-cially those suspected of being on the front line of defenceHowever acute exposures especially to plants that havegrown in relatively O3-free air may be difficult to interpretFor example a short high O3 exposure may overwhelmplant defences and disrupt membrane lipids and proteinscausing extensive leakage of cellular components anduncontrolled cell or tissue death which rarely occurs in thecurrent environment although similar responses mightoccur on a micro-scale in substomatal cells and appear as ahypersensitive response to pathogens The choice of O3
treatment methodology depends on the goal of theresearch whether that is to determine the effects of cur-rent ambient O3 concentrations on yield [eg NCLANin which open top chambers were used to treat plantswith a range of O3 concentrations that encompassed amb-ient air concentrations (Heagle 1989)] or to predict ef-fects of future levels of ambient O3 compared with currentlevels [eg SoyFACE in which plants are treated with am-bient air and ambient air plus O3 in a free air system(httpwwwsoyfaceuiuceduindexhtm)] or to investigatemechanisms of O3 damage with the aim of improving croptolerance to O3 stress (eg identification of genetic traitscontrolling sensitivity to O3) Each approach has its advan-tages and drawbacks and choices depend on objectives andeconomic feasibility
However there is unmistakable evidence that increasingconcentrations of atmospheric CO2 will ameliorate O3 dam-age to a number of crops and woody plants This interactionwill likely continue in the foreseeable future as atmosphericCO2-enrichment and emission of tropospheric O3 precur-sors continue to increase While this might be seen as afortunate coincidence at present it is difficult to predicthow interactions between these factors and other possiblechanges in global climate will play out At present it isreasonable to conclude that ambient O3 detracts from thetotal crop productivity possible in clean air in many regionsof the world and thus may diminish the quantity of carboneventually sequestered in soils Ozone flux models willlikely require modification to account for the influence ofelevated CO2 on gs and possibly metabolic changes thataffect injury threshold levels Further O3 flux models havethe potential to help identify key physiological and bio-chemical processes in the network of plant responses to O3Advances in our ability to genetically dissect mechanismsinvolved in O3 toxicity should lead to better understandingof differential sensitivity to O3 and oxidative stress in gen-eral among crop species and cultivars with the goal ofdeveloping crops with improved performance in a changingglobal climate This research also will aid in better assess-ments of the costs in terms of potential world-wide food
production associated with current and future levels of tro-pospheric O3 pollution that underpin governmental airquality regulations
REFERENCES
Adams RM amp Horst RL Jr (2003) Future directions in air qual-ity research economic issues Environment International 29289ndash302
Andersen C (2003) Source-sink balance and carbon allocationbelow ground in plants exposed to ozone New Phytologist 157213ndash228
Ashmore M (2005) Assessing the future global impacts of ozoneon vegetation Plant Cell and Environment 28 doi101111j1365-3040200501341x
Black VJ Black CR Roberts JA amp Stewart CA (2000)Impact of ozone on the reproductive development of plantsNew Phytologist 147 421ndash447
Booker FL Burkey KO Overmyer K amp Jones AM (2004)Differential responses of G-protein Arabidopsis thalianamutants to ozone New Phytologist 162 633ndash641
Booker FL Fiscus EL amp Miller JE (2004) Combined effects ofelevated atmospheric carbon dioxide and ozone on soybeanwhole-plant water use Environmental Management 33 S355ndashS362
Booker FL Miller JE Fiscus EL Pursley WA amp StefanskiLA (2005) Comparative responses of container- versus ground-grown soybean to elevated CO2 and O3 Crop Science 45 883ndash895
Burkey KO (1999) Effects of ozone on apoplastcytoplasm parti-tioning of ascorbic acid in snap bean Physiologia Plantarum107 188ndash193
Burkey KO Eason G amp Fiscus EL (2003) Factors that affectleaf extracellular ascorbic acid content and redox status Physi-ologia Plantarum 117 51ndash57
Burkey KO Wei C Palmer G Ghosh P amp Fenner GP (2000)Antioxidant metabolite levels in ozone-sensitive and tolerantgenotypes of snap bean Physiologia Plantarum 110 195ndash200
Calatayud A amp Barreno E (2001) Chlorophyll a fluorescenceantioxidant enzymes and lipid peroxidation in tomato inresponse to ozone and benomyl Environmental Pollution 115283ndash289
Castagna A Nali C Ciompi S Lorenzini G Soldatini GF ampRanieri A (2001) Ozone exposure affects photosynthesis ofpumpkin (Cucurbita pepo) plants New Phytologist 152 223ndash229
Castillo FJ amp Greppin H (1988) Extracellular ascorbic acid andenzyme activities related to ascorbic acid metabolism in Sedumalbum L leaves after ozone exposure Environmental andExperimental Botany 28 231ndash2387
Castillo FJ amp Heath RL (1990) Ca2+ transport in membranevesicles from pinto bean leaves and its alteration after ozoneexposure Plant Physiology 94 788ndash795
Chameides WL (1989) The chemistry of ozone deposition toplant leaves role of ascorbic acid Environmental Science andTechnology 23 595ndash600
Chernikova T Robinson JM Lee EH amp Mulchi CL (2000)Ozone tolerance and antioxidant enzyme activity in soybeancultivars Photosynthesis Research 64 15ndash26
Ciompi S Castagna A Ranieri A Nali C Lorenzini G amp Sol-datini FG (1997) CO2 assimilation xanthophyll cycle pigmentsand PSII efficiency in pumpkin plants as affected by ozone fumi-gation Physiologia Plantarum 101 881ndash889
Conklin PL amp Barth C (2004) Ascorbic acid a familiar smallmolecule intertwined in the response of plants to ozone patho-gens and the onset of senescence Plant Cell and Environment27 959ndash970
1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

1008 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Conklin PL Williams EH amp Last RL (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant Proceedings of the National Academy of Sciences of theUSA 93 9970ndash9974
Cooley DR amp Manning WJ (1987) The impact of ozone onassimilate partitioning in plants a review Environmental Pollu-tion 47 95ndash113
Damicone JP amp Manning WJ (1987) Foliar sensitivity of soy-beans from early maturity groups to ozone and inheritance ofinjury response Plant Disease 71 332ndash336
Dominy PJ amp Heath RL (1985) Inhibition of the K+-stimulatedATPase of the plasmalemma of pinto bean leaves by ozonePlant Physiology 77 43ndash45
Eckardt NA amp Pell E (1994) O3-induced degradation of Rubiscoprotein and loss of Rubisco mRNA in relation to leaf age inSolanum tuberosum L New Phytologist 127 741ndash748
Emberson LD Ashmore MR Cambridge HM Simpson D ampTuovinen JP (2000) Modelling stomatal ozone flux acrossEurope Environmental Pollution 109 403ndash413
Farage PK Long SP Lechner EG amp Baker NR (1991) Thesequence of change within the photosynthetic apparatus ofwheat following short-term exposure to ozone Plant Physiology95 529ndash535
Fiscus EL Booker FL Reid CD amp Miller JE (2001) Uncon-sidered environmental stresses may cause overestimates of theCO2-fertilization effect Paper presented at the 12th InternationalCongress on Photosynthesis Brisbane Australia httpwwwpublishcsiroaupaperSA0403642htm
Fiscus EL Miller JE Booker FL Heagle AS amp Reid CD(2002) The impact of ozone and other limitations on the cropproductivity response to CO2 Technology 8 181ndash192
Fiscus EL Reid CD Miller JE amp Heagle AS (1997) ElevatedCO2 reduces O3 flux and O3-induced yield losses in soybeanspossible implications for elevated CO2 studies Journal of Exper-imental Botany 48 307ndash313
Foyer C H amp Noctor G (2005) Oxidant and antioxidant signallingin plants a re-evaluation of the concept of oxidative stress in aphysiological context Plant Cell and Environment 28doi101111j1365-3040200501327x
Fuhrer J amp Booker FL (2003) Ecological issues related toozone agricultural issues Environment International 29 141ndash154
Fuhrer J Skarby L amp Ashmore M (1997) Critical levels for ozoneeffects on vegetation in Europe Environmental Pollution 97 91ndash106
Glick RE Schlagnhaufer CD Arteca RN amp Pell EJ (1995)Ozone-induced ethylene emission accelerates the loss of ribu-lose-15-bisphosphate carboxylaseoxygenase and nuclear-encoded mRNAs in senescing potato leaves Plant Physiology109 891ndash898
Godde D amp Buchhold J (1992) Effect of long term fumigationwith ozone on the turnover of the D-1 reaction center polypep-tide of photosystem II in spruce (Picea abies) Physiologia Plan-tarum 86 568ndash574
Grantz DA amp Farrar JF (2000) Ozone inhibits phloem load-ing from a transport pool Compartmental efflux analysis inPima cotton Australian Journal of Plant Physiology 27 859ndash868
Grantz DA amp Yang S (2000) Ozone impacts on allometry androot hydraulic conductance are not mediated by source limita-tion nor developmental age Journal of Experimental Botany 51919ndash927
Grantz DA Zhang X amp Carlson T (1999) Observations andmodel simulations link stomatal inhibition to impaired hydraulicconductance following ozone exposure in cotton Plant Cell andEnvironment 22 1201ndash1210
Grimes HD Perkins KK amp Boss WF (1983) Ozone degradesinto hydroxyl radical under physiological conditions Plant Phys-iology 72 1016ndash1020
Gruumlnhage L Haenel HD amp Jaumlger HJ (2000) The exchange ofozone between vegetation and atmosphere micrometeorologi-cal measurement techniques and models Environmental Pollu-tion 109 373ndash392
Guidi L DeglrsquoInnocenti E amp Soldatini GF (2002) Assimilationof CO2 enzyme activation and photosynthetic electron trans-port in bean leaves as affected by high light and ozone NewPhytologist 156 377ndash388
Guidi L Di Cagno R amp Soladatini GF (2000a) Screening of beancultivars for their response to ozone as evaluated by visiblesymptoms and leaf chlorophyll fluorescence EnvironmentalPollution 107 349ndash355
Guidi L Tonini M amp Soladatini GF (2000b) Effects of high lightand ozone fumigation on photosynthesis in Phaseolus vulgarisPlant Physiology and Biochemistry 38 717ndash725
Gunnison AF amp Hatch GE (1999) O3-induced inflammation inprepregnant pregnant and lactating rats correlates with O3 doseestimated by 18O American Journal of Physiology 276 L332ndashL340
Guzy MR amp Heath RL (1993) Responses to ozone of varietiesof common bean (Phaseolus vulgaris L) New Phytologist 124617ndash625
Heagle AS (1989) Ozone and crop yield Annual Review of Phy-topathology 27 397ndash423
Heagle AS amp Letchworth MB (1982) Relationships amonginjury growth and yield responses of soybean cultivars exposedto ozone at different light intensities Journal of EnvironmentalQuality 11 690ndash694
Heagle AS Miller JE Booker FL amp Pursley WA (1999)Ozone stress carbon dioxide enrichment and nitrogen fertilityinteractions in cotton Crop Science 39 731ndash741
Heagle AS Miller JE Burkey KO Eason G amp Pursley WA(2002) Growth and yield responses of snap bean to mixtures ofcarbon dioxide and ozone Journal of Environmental Quality 312008ndash2014
Heagle AS Miller JE amp Pursley WA (2000) Growth and yieldresponses of winter wheat to mixtures of ozone and carbondioxide Crop Science 40 1656ndash1664
Heagle AS Miller JE amp Pursley WA (2003) Growth andyield responses of potato to mixtures of carbon dioxideand ozone Journal of Environmental Quality 32 1603ndash1610
Heagle AS Miller JE Rawlings JO amp Vozzo SF (1991) Effectof growth stage on soybean response to chronic ozone exposureJournal of Environmental Quality 20 562ndash570
Heagle AS Miller JE Sherrill DE amp Rawlings JO (1993)Effects of ozone and carbon dioxide mixtures on two clones ofwhite clover New Phytologist 123 751ndash762
Heath RL (1987) The biochemistry of ozone attack on the plasmamembrane of plant cells Recent Advances in Phytochemistry 2129ndash54
Heath RL amp Taylor GE Jr (1997) Physiological processes andplant responses to ozone exposure In Forest Decline and Ozonea Comparison of Controlled Chamber and Field Experiments(eds H Sandermann AR Wellburn amp RL Heath) pp 317ndash368 Springer-Verlag Berlin Germany
Heck WW Adams RM Cure WW Heagle AS HeggestadHE Kohut RJ Kress LW Rawlings JO amp Taylor OC(1983) A reassessment of crop loss from ozone EnvironmentalScience and Technology 17 572Andash581A
Hewitt CN Kok GL amp Rall R (1990) Hydroperoxides in plantsexposed to ozone mediate air pollution damage to alkene emit-ters Nature 344 56ndash58
Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

Crop responses to ozone 1009
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Horemans N Foyer CH amp Asard H (2000) Transport and actionof ascorbate at the plasma membrane Trends in Plant Science5 263ndash267
Humphries SW amp Long SP (1995) WIMOVAC a software pack-age for modelling the dynamics of plant leaf and canopy photo-synthesis CABIOS (Computer Applications in BioSciences) 11361ndash371
Jakob B amp Heber U (1998) Apoplastic ascorbate does not pre-vent the oxidation of fluorescent amphiphilic dyes by ambientand elevated concentrations of ozone in leaves Plant and CellPhysiology 39 313ndash322
Jarvis PG (1976) The interpretation of the variations in leaf waterpotential and stomatal conductance found in canopies in thefield Philosophical Transactions of the Royal Society of LondonSeries B Biology Sciences 273 593ndash610
Kangasjaumlrvi J Talvinen J Utriainen M amp Karjalainen R (1994)Plant defence systems induced by ozone Plant Cell and Envi-ronment 17 783ndash794
Kangasjaumlrvi J Jaspers P amp Kollist H (2005) Signalling and celldeath in ozone-exposed plants Plant Cell and Environment 28doi101111j1365-3040200501325x
Kirichenko A Li L Morandi MT amp Holian A (1996) 4-Hydroxy-2-nonenal-protein adducts and apoptosis in murinelung cells after acute ozone exposure Toxicology and AppliedPharmacology 141 416ndash424
Kuik OJ Helming JFM Dorland C amp Spaninks FA (2000)The economic benefits to agriculture of a reduction of low levelozone pollution in the Netherlands European Review of Agri-cultural Economics 27 75ndash90
Lee JC Steiner KC Zhang JW amp Skelly JM (2002) Herita-bility of ozone sensitivity in open-pollinated families of blackcherry (Prunus serotina Ehrh) Forest Science 48 111ndash117
Lefohn AS (1992) The characterization of ambient ozone expo-sures In Surface Level Ozone Exposures and Their Effects onVegetation (ed AS Lefohn) pp 31ndash92 Lewis Publishers IncChelsea MI USA
Leipner J Oxborough K amp Baker NR (2001) Primary sites ofozone-induced perturbations of photosynthesis in leaves iden-tification and characterization in Phaseolus vulgaris using highresolution chlorophyll fluorescence imaging Journal of Experi-mental Botany 52 1689ndash1696
Long SP Ainsworth EA Rogers A amp Ort DR (2004) Risingatmospheric carbon dioxide plants FACE the future AnnualReview of Plant Biology 55 591ndash628
Long SP amp Naidu SL (2002) Effects of oxidants at the biochem-ical cell and physiological levels with particular reference toozone In Air Pollution and Plant Life (eds JN B Bell amp MTreshow) pp 69ndash88 John Wiley amp Sons Ltd West Sussex
Loreto F amp Velikova V (2001) Isoprene produced by leaves pro-tects the photosynthetic apparatus against ozone damagequenches ozone products and reduces lipid peroxidation of cel-lular membranes Plant Physiology 127 1781ndash1787
Luwe M amp Heber U (1995) Ozone detoxification in the apoplasmand symplasm of spinach broad bean and beech leaves at ambi-ent and elevated concentrations of ozone in air Planta 197 448ndash455
Martin MJ Farage PK Humphries SW amp Long SP (2000) Canthe stomatal changes caused by acute ozone exposure be pre-dicted by changes occurring in the mesophyll A simplificationfor models of vegetation response to the global increase in tro-pospheric elevated ozone episodes Australian Journal of PlantPhysiology 27 211ndash219
Massman WJ (2004) Toward an ozone standard to protectvegetation based on effective dose a review of deposition resis-tances and a possible metric Atmospheric Environment 382323ndash2337
Mauzerall DL amp Wang XP (2001) Protecting agricultural cropsfrom the effects of tropospheric ozone exposure Reconcilingscience and standard setting in the United States Europe andAsia Annual Review of Energy and the Environment 26 237ndash268
McAinsh MR Evans NH Montgomery LT amp North KA(2002) Calcium signalling in stomatal responses to pollutantsNew Phytologist 153 441ndash447
McKee IF Bullimore JF amp Long SP (1997) Will elevated CO2
concentrations protect the yield of wheat from O3 damagePlant Cell and Environment 20 77ndash84
McKee IF Farage PK amp Long SP (1995) The interactive effectsof elevated CO2 and O3 concentration on photosynthesis inspring wheat Photosynthesis Research 45 111ndash119
McLaughlin SB amp McConathy RK (1983) Effects of SO2 and O3
on allocation of 14C-labeled photosynthate in Phaseolus vulgarisPlant Physiology 73 630ndash635
Mehlhorn H Tabner BJ amp Wellburn AR (1990) Electron spinresonance evidence for the formation of free radicals in plantsexposed to ozone Physiologia Plantarum 79 377ndash383
Miller JE (1987a) Effects of ozone and sulfur dioxide stress ongrowth and carbon allocation in plants Recent Advances in Phy-tochemistry 21 55ndash100
Miller JE (1987b) Effects on photosynthesis carbon allocationand plant growth associated with air pollutant stress In Assess-ment of Crop Loss from Air Pollutants (eds WW Heck OCTaylor amp DT Tingey) pp 287ndash314 Elsevier Applied ScienceLondon UK
Miller JE Heagle AS amp Pursley WA (1998) Influence of ozonestress on soybean response to carbon dioxide enrichment IIBiomass and development Crop Science 38 122ndash128
Miller JE Patterson RP Heagle AS Pursley WA amp HeckWW (1988) Growth of cotton under chronic ozone stress attwo levels of soil moisture Journal of Environmental Quality 17635ndash643
Miller JE Pursley WA Vozzo SF amp Heagle AS (1991)Response of net carbon exchange rate of soybean to ozone atdifferent stages of growth and its relation to yield Journal ofEnvironmental Quality 20 571ndash575
Morgan PB Ainsworth EA amp Long SP (2003) How does ele-vated ozone impact soybean A meta-analysis of photosynthesisgrowth and yield Plant Cell and Environment 26 1317ndash1328
Morgan PB Bernacchi CJ Ort DR amp Long SP (2004) An invivo analysis of the effect of season-long open-air elevation ofozone to anticipated 2050 levels on photosynthesis in soybeanPlant Physiology 135 2348ndash2357
Murphy JJ Delucchi MA McCubbin DR amp Kim HJ (1999)The cost of crop damage caused by ozone air pollution frommotor vehicles Journal of Environmental Management 55 365ndash376
Musselman RC amp Massman WJ (1999) Ozone flux to vegetationand its relationship to plant response and ambient air qualitystandards Atmospheric Environment 33 65ndash73
Noctor G amp Foyer CH (1998) Ascorbate and glutathione keep-ing active oxygen under control Annual Review of Plant Phys-iology and Plant Molecular Biology 49 249ndash279
Olszyk DM Tingey DT Watrud L Seidler R amp Andersen C(2000) Interactive effects of O3 and CO2 implications for terres-trial ecosystems In Trace Gas Emissions and Plants (ed SNSingh) pp 97ndash136 Kluwer Academic Publishers DordrechtThe Netherlands
Osmond CB (1994) What is photoinhibition Some insights fromcomparison of shade and sun plants In Photoinhibition of Pho-tosynthesis ndash from Molecular Mechanisms to the Field (eds NRBaker amp JR Bowyer) pp 1ndash24 Bios Scientific PublishersOxford UK
1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

1010 E L Fiscus et al
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Overmyer K Brosche M amp Kangasjaumlrvi J (2003) Reactive oxy-gen species and hormonal control of cell death Trends in PlantScience 8 335ndash342
Overmyer K Tuominen H Kettunen R Betz C LangebartelsC Sandermann HJ amp Kangasjaumlrvi J (2000) Ozone-sensitiveArabidopsis rcd1 mutant reveals opposite roles for ethyleneand jasmonate signaling pathways in regulating superoxide-dependent cell death Plant Cell 12 1849ndash1862
Pell E Schlagnhaufer CD amp Arteca RN (1997) Ozone-inducedoxidative stress mechanisms of action and reaction PhysiologiaPlantarum 100 264ndash273
Pendall E Bridgham S Hanson PJ et al (2004) Below-groundprocess responses to elevated CO2 and temperature a discussionof observations measurement methods and models New Phy-tologist 162 311ndash322
Pei Z-M Murata Y Benning G Thomine S Kluumlsener B AllenGJ Grill E amp Schroeder JI (2000) Calcium channels acti-vated by hydrogen peroxide mediate abscisic acid signalling inguard cells Nature 406 731ndash734
Plazek A Rapacz M amp Skoczowski A (2000) Effects of ozonefumigation on photosynthesis and membrane permeability inleaves of spring barley meadow fescue and winter rape Photo-synthetica 38 409ndash413
Pleijel H Danielsson H Ojanpera K Temmerman LD HogyP Badiani M amp Karlsson PE (2004) Relationships betweenozone exposure and yield loss in European wheat and potato ndasha comparison of concentration- and flux-based exposure indicesAtmospheric Environment 38 2259ndash2269
Ploumlchl M Lyons T Ollerenshaw J amp Barnes J (2000) Simulatingozone detoxification in the leaf apoplast through the direct reac-tion with ascorbate Planta 210 454ndash467
Prather M Ehhalt D Dentener F et al (2001) Atmosphericchemistry and greenhouse gases In Climate Change 2001 theScientific Basis Contribution of Working Group I to the ThirdAssessment Report of the Intergovernmental Panel on ClimateChange (eds JT Houghton Y Ding DJ Griggs MNoguer PJ van der Linden X Dai K Maskell amp CAJohnson) pp 239ndash287 Cambridge University Press Cam-bridge UK
Pryor WA (1994) Mechanisms of radical formation from reac-tions of ozone with target molecules in the lung Free RadicalBiology and Medicine 17 451ndash465
Rao MV amp Davis KR (2001) The physiology of ozone inducedcell death Planta 213 682ndash690
Rao MV Lee H-I Creelman RA Mullet JE amp Davis KR(2000) Jasmonic acid signaling modulates ozone-induced hyper-sensitive cell death Plant Cell 12 1633ndash1646
Rao MV Lee HI amp Davis KR (2002) Ozone-induced ethyleneproduction is dependent on salicylic acid and both salicylic acidand ethylene act in concert to regulate ozone-induced cell deathPlant Journal 32 447ndash456
Reich PB Schoettle AW amp Amundsen RG (1985) Effects oflow concentrations of O3 Leaf age and water stress on leafdiffusive conductance and water use efficiency in soybean Phys-iologia Plantarum 63 58ndash64
Reichenauer TG Goodman BA Kostecki P amp Soja G (1998)Ozone sensitivity in Triticum durum and T aestivum with respectto leaf injury photosynthetic activity and free radical contentPhysiologia Plantarum 104 681ndash686
Reid CD amp Fiscus EL (1998) Effects of elevated [CO2] andorozone on limitations to CO2 assimilation in soybean (Glycinemax) Journal of Experimental Botany 49 885ndash895
Reid CD Fiscus EL amp Burkey KO (1998) Combined effects ofchronic ozone and elevated CO2 on Rubisco activity and leafcomponents in soybean (Glycine max) Journal of ExperimentalBotany 49 1999ndash2011
Reid CD Fiscus EL amp Burkey KO (1999) Effects of chronicozone and elevated atmospheric CO2 concentrations on ribu-lose-15-bisphosphate in soybean (Glycine max) PhysiologiaPlantarum 106 378ndash385
Reinert RA amp Eason G (2000) Genetic control of O3 sensitivityin a cross between two cultivars of snap bean Journal of theAmerican Society of Horticultural Science 125 222ndash227
Robinson JM amp Britz SJ (2000) Tolerance of a field grownsoybean cultivar to elevated ozone level is concurrent withhigher leaflet ascorbic acid level higher ascorbate-dehy-droascorbate redox status and long term photosynthetic pro-ductivity Photosynthesis Research 64 77ndash87
Runeckles VC (1992) Uptake of ozone by vegetation In SurfaceLevel Ozone Exposures and Their Effects on Vegetation (ed ASLefohn) pp 157ndash188 Lewis Publishers Inc Chelsea MI USA
Runeckles VC amp Chevone BI (1992) Crop responses to ozoneIn Surface Level Ozone Exposures and Their Effects on Vegeta-tion (ed AS Lefohn) pp 189ndash270 Lewis Publishers IncChelsea MI USA
Runeckles VC amp Vaartnou M (1997) EPR evidence for superox-ide anion formation in leaves during exposure to low levels ofozone Plant Cell and Environment 20 306ndash314
Sandermann H Jr Ernst D Heller W amp Langebartels C (1998)Ozone an abiotic elicitor of plant defence reactions Trends inPlant Science 3 47ndash50
Schraudner M Moeder W Wiese C Van Camp W Inzeacute DLangebartels C amp Sandermann H Jr (1998) Ozone-inducedoxidative burst in the ozone biomonitor plant tobacco Bel W3Plant Journal 16 235ndash245
Schroeder JI Allen GJ Hugouvieux V Kwak JM amp WanerD (2001) Guard cell signal transduction Annual Review ofPlant Physiology and Molecular Biology 52 627ndash658
Sharkey TD amp Yeh S (2001) Isoprene emission from plantsAnnual Review of Plant Physiology and Molecular Biology 52407ndash436
Sharma YK Leon J Raskin I amp Davis KR (1996) Ozone-induced responses in Arabidopsis thaliana The role of salicylicacid in the accumulation of defense-related transcripts andinduced resistance Proceedings of the National Academy of Sci-ences of the USA 93 5099ndash5104
Smirnoff N (1996) The function and metabolism of ascorbic acidin plants Annals of Botany 78 661ndash669
Smirnoff N Conklin PL amp Loewus FA (2001) Biosynthesis ofascorbic acid in plants a renaissance Annual Review of PlantPhysiology and Molecular Biology 52 437ndash467
Tamaoki M Matsuyama T Kanna M Nakajima N Kubo AAono M amp Saji J (2003) Differential ozone sensitivity amongArabidopsis accessions and its relevance to ethylene synthesisPlanta 216 552ndash560
Temple PJ (1990) Growth and yield of processing tomato (Lyco-persicon esculentum Mill) cultivars to ozone Environmentaland Experimental Botany 30 283ndash291
Tingey DT amp Hogsett WE (1985) Water stress reduces ozoneinjury via a stomatal mechanism Plant Physiology 77 944ndash947
Tingey DT Reinert R A amp Carter HB (1972) Soybean culti-vars acute foliar response to ozone Crop Science 12 268ndash270
Tuovinen JP (2000) Assessing vegetation exposure to ozoneproperties of the AOT40 index and modifications by depositionmodelling Environmental Pollution 109 361ndash372
Turcsanyi E Lyons T Ploumlchl M amp Barnes J (2000) Does ascor-bate in the mesophyll cell walls form the first line of defenceagainst ozone Testing the concept using broad bean (Vicia fabaL) Journal of Experimental Botany 51 901ndash910
US Environmental Protection Agency (1996) Air Quality Criteriafor Ozone and Other Photochemical Oxidants Vol III Reportno EPA600P-93004bF US Environmental Protection Agency
Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005

Crop responses to ozone 1011
copy 2005 Blackwell Publishing Ltd Plant Cell and Environment 28 997ndash1011No claim to original US government works
Office of Air Quality Planning and Standards Research TrianglePark NC USA
UNECE (2000) Review of Strategies and Policies for Air PollutionAbatement UNECE Convention on LongndashRange Trans Bound-ary Air Pollution httpwwwuneceorgenvlrtapconvcon-clusihtm
UNECE (2004a) Air Pollution and Vegetation ICP VegetationAnnual Report 20032004 UNECE ICP Vegetation Coordina-tion Centre CEH Bangor UK
UNECE (2004b) Manual on Methodologies and Criteria for Mod-elling and Mapping Critical Loads amp Levels and Air PollutionEffects Risks and Trends Umweltbundesamt Berlin Germany
Vahala J Ruonala R Keinanen M Tuominen H amp KangasjaumlrviJ (2003) Ethylene insensitivity modulates ozone-induced celldeath in birch Plant Physiology 132 185ndash195
Vozzo SF Miller JE Pursley WA amp Heagle AS (1995) Effectsof ozone and water deficit on field-grown soybean I Leaf gasexchange Journal of Environmental Quality 24 663ndash670
Wellburn FAM amp Wellburn AR (1996) Variable patterns ofantioxidant protection but similar ethene emission differencesin several ozone-sensitive and ozone-tolerant plant selectionsPlant Cell and Environment 19 754ndash760
Wohlgemuth H Mittelstrass K Kschieschan S Bender J WeigelH-J Overmyer K Kangasjaumlrvi J Sandermann HJ amp Lange-bartels C (2002) Activation of an oxidative burst is a generalfeature of sensitive plants exposed to the air pollutant ozonePlant Cell and Environment 25 717ndash726
Wolff K Morgan-Richards M amp Davison AW (2000) Patternsof molecular genetic variation in Plantago major and P interme-dia in relation to ozone resistance New Phytologist 145 501ndash509
Received 19 November 2004 received in revised form 24 February2005 accepted for publication 24 February 2005














![Physiological and Transcriptomic Aspects of Urea … · Physiological and Transcriptomic Aspects of Urea Uptake and Assimilation in Arabidopsis Plants1[W] Patricia Me ´rigout,MaudLelandais,Frede´rique](https://img.pdfslide.net/doc/110x75/5bc37b1909d3f299608cbb80/physiological-and-transcriptomic-aspects-of-urea-physiological-and-transcriptomic.jpg)














