Embed Size (px)
Citation preview

Cumulus Oophorus-associated Glycodelin-C DisplacesSperm-bound Glycodelin-A and -F and StimulatesSpermatozoa-Zona Pellucida Binding*
Received for publication, August 7, 2006, and in revised form, November 1, 2006 Published, JBC Papers in Press, December 27, 2006, DOI 10.1074/jbc.M607482200
Philip C. N. Chiu‡, Man-Kin Chung‡, Riitta Koistinen§¶, Hannu Koistinen¶, Markku Seppala¶, Pak-Chung Ho‡,Ernest H. Y. Ng‡, Kai-Fai Lee‡, and William S. B. Yeung‡1
From the ‡Department of Obstetrics and Gynaecology, University of Hong Kong, Queen Mary Hospital, Pokfulam Road,Hong Kong SAR, China and the Departments of §Obstetrics and Gynaecology and ¶Clinical Chemistry,University of Helsinki and Helsinki University, Central Hospital, 00029 HUS Helsinki, Finland
Spermatozoa have to swim through the oviduct and thecumulus oophorus before fertilization in vivo. In the oviduct,spermatozoa are exposed to glycodelin-A and -F that inhibitspermatozoa-zona pellucida binding. In this study, we deter-mined whether these glycodelins would inhibit fertilization.The data showed that the spermatozoa without previousexposure to glycodelin-A and -F acquired glycodelin immu-noreactivity during their passage through the cumulusoophorus. On the other hand, when glycodelin-A or -F-pre-treated spermatozoa were exposed to the cumulus oophorus,the zona pellucida binding inhibitory activity of glycodelin-Aand -F was not only removed, but the spermatozoa acquiredenhanced zona pellucida binding ability. These actions of thecumulus oophorus were due to the presence of a cumulusisoform of glycodelin, designated as glycodelin-C. The cumu-lus cells could convert exogenous glycodelin-A and -F to gly-codelin-C, which was then released into the surroundingmedium. The protein core of glycodelin-C was identical tothat in other glycodelin isoforms, as demonstrated by massspectrum, peptide mapping, and affinity to anti-glycodelinantibody recognizing the protein core of glycodelin. In addi-tion to having a smaller size and a higher isoelectric point,glycodelin-C also had lectin binding properties differentfrom other isoforms. Glycodelin-C stimulated spermatozoa-zona pellucida binding in a dose-dependent manner, and iteffectively displaced sperm-bound glycodelin-A and -F. Inconclusion, the cumulus cells transform glycodelin-A and -Fto glycodelin-C, which in turn removes the spermatozoa-zona binding inhibitory glycodelin isoforms and enhancesthe zona binding capacity of spermatozoa passing throughthe cumulus oophorus.
After ovulation, the oocyte-cumulus complex and follicularfluid are transported to the oviduct. Therefore, the spermato-zoa have to traverse through the follicular fluid and the cumulusoophorus before they bind to the zona pellucida of the oocyte toinitiate the fertilization process. Failure of cumulus oophorusformation results in problems in fertilization (1–4). Both thefollicular fluid and the cumulus oophorus are known to modu-late sperm function (5, 6). The cumulus oophorus may selectspermatozoa with normal morphology (7, 8) and with intactacrosome (9) and create an optimal microenvironment to facil-itate fertilization (7, 10).Glycodelin is a lipocalin glycoprotein with three well known
isoforms, namely glycodelin-A (amniotic fluid isoform), gly-codelin-S (seminal plasma isoform), and glycodelin-F (follicu-lar fluid isoform) (11, 12). These isoforms have molecular sizesof 28–30 kDa. They have the same protein core and only differin their glycosylation (11, 13).Glycodelin isoforms are present in the reproductive tract
throughwhich the spermatozoamust pass before fertilizing theoocyte. Different isoforms have diverse actions on sperm func-tions (13). Glycodelin-Smaintains the spermatozoa in an unca-pacitated state before their entry into the uterine lumen (14).Glycodelin-A and -F inhibit spermatozoa-zona pellucida bind-ing (15–17). The latter, but not the former, inhibits progester-one-induced acrosome reaction, thus preventing prematureacrosome reaction of spermatozoa (12, 18).The fallopian tube produces glycodelin (19, 20) with increas-
ing expression during the luteal phase (21). Our unpublishedobservation2 shows that human oviductal cells secrete both gly-codelin-A and -F in vitro. Glycodelin-F is the main glycodelinisoform in the follicular fluid. Thus, spermatozoa are exposedto these glycodelins in the oviduct and follicular fluid beforethey fertilize an oocyte. It seems paradoxical that both glycode-lin-A and -F inhibit the binding of spermatozoa to the zonapellucida.Whether these glycodelin isoformswould inhibit fer-tilization in vivo remains unclear.Biological activities are often regulated by a fine balance
between agonist and antagonist actions. For instance, proges-terone induces acrosome reaction, but its activity is inhibited byhuman oviductal fluid (22) or glycodelin-F from human follic-
* This work was supported in part by grants from the Committee on Researchand Conference Grant; The University of Hong Kong; Hong Kong ResearchGrant Council Grants HKU7261/01M, HKU7408/03M, and HKU7647/06M,Helsinki University Central Hospital Research Funds; Federation of theFinnish Life and Pension Insurance Companies; the Cancer Society of Fin-land; the Academy of Finland; and the University of Helsinki. The costs ofpublication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked “advertisement” inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 To whom correspondence should be addressed: Dept. of Obstetrics andGynaecology, University of Hong Kong, Queen Mary Hospital, PokfulamRoad, Hong Kong SAR, China. Tel.: 852-28553405; Fax: 852-28175374;E-mail: [email protected].
2 P. C. N. Chiu, M.-K.Chung, R. Koistinen, H. Koistinen, M. Seppala, P.-C. Ho,E. H. Y. Ng, K.-F. Lee, and W. S. B. Yeung, unpublished observation.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 8, pp. 5378 –5388, February 23, 2007© 2007 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
5378 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 8 • FEBRUARY 23, 2007
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from

ular fluid (12, 18). Our recent data showed that the cumuluscells reduced the spermatozoazona binding inhibitory activityof human follicular fluid (23). Although the cumulus cells donot synthesize glycodelin, they take up glycodelin-A from thesurrounding medium (24). In the cell lysate of glycodelin-A-treated cumulus cells, a glycodelin-likemolecule of size smallerthan glycodelin-A was found. Whether this small glycodelin-like molecule represents product of taken up glycodelin-A orhas another function is unknown. Recently, we reported ourpreliminary observation on the presence of a similar-sized gly-codelin-like molecule in the cumulus matrix (13). Based onthese observations, we hypothesize that the cumulus cells pro-duce another glycodelin isoform to modulate the activity ofglycodelin-A and -F on spermatozoa-zona pellucida binding.The objectives of this report were 1) to determine the zonabinding capacity of spermatozoa that had penetrated throughthe cumulus oophorus, 2) to characterize the small glycodelin-like molecule in the extracellular matrix of cumulus oophorusaffecting spermatozoa-zona pellucida binding, and 3) to studythe action of the molecule on glycodelin-A and -F-treatedspermatozoa.
EXPERIMENTAL PROCEDURES
Semen Samples—The Ethics committee of the University ofHong Kong approved the research protocol. Samples with nor-mal semen parameters according to World Health Organiza-tion criteria (25) from men visiting the subfertility clinics atQueenMary Hospital, the University of Hong Kong were used.The semen samples were processed by Percoll (GE Healthcare)density gradient centrifugation (12). The processed spermato-zoa were capacitated in Earle’s balanced salt solution (EBSS3;Flow Laboratories, Irvine, UK) supplemented with sodiumpyruvate and 3% bovine serum albumin for 3 h before theywereresuspended in EBSS containing 0.3% BSA (EBSS/BSA).Purification of Glycodelin Isoforms—Amonoclonal anti-gly-
codelin antibody (clone F43-7F9)-Sepharose column was usedto purify glycodelin-A and -S from amniotic fluid and seminalplasma, respectively, as described (26). The bound glycodelinwas eluted with 0.1% trifluoroacetic acid and dialyzed against100 mM sodium phosphate buffer (pH 7.2). Glycodelin-S wasfurther purified by anion exchange chromatography (14). Gly-codelin-F was purified from human follicular fluid collectedfrom women undergoing assisted reproduction treatment inQueen Mary Hospital (Hong Kong). The follicular fluid waspassed successively through cibacron blue (HiTrap Blue), pro-tein-G, concanavalin A-Sepharose columns (GE Healthcare),Amicon-10 concentrator (Amicon Inc. Beverly, CA), Mono-Q,and Superose columns as described (17). The concentrations ofthe purified glycodelin isoforms were determined by a com-mercial kit (protein assay kit; Bio-Rad). The N-glycosidase Fdeglycosylation kit (Bio-Rad) was used to deglycosylate gly-codelins (18). The deglycosylated protein was further purifiedby gel filtration chromatography in an AKTA system (GEHealthcare).
Collection of Cumulus Oophorus and CumulusMatrix—TheEthicsCommittee of theUniversity ofHongKong approved theprotocol for collection anduse of cumulus oophorus and cumu-lus matrix in this study. Cumulus-oocyte complexes were col-lected after oocyte retrieval from women who came for treat-ment with intracytoplasmic sperm injection because of maleinfertility. Humanmenopausal gonadotropin (Serono, Geneva,Switzerland) was used for ovarian stimulation after down-reg-ulation with buserelin (27). For use in the capillary-cumulusmodel (6), intact cumulus oophorus was obtained by mechani-cal dissection from the cumulus-oocyte complex. The cumulusmatrix was collected after dispersion of the cumulus oophorusin 1 ml of 0.1% hyaluronidase at 37 °C followed by centrifuga-tion at 300 � g for 10 min to remove the cumulus cells.Zona Binding Capacity of Spermatozoa Penetrating through
the Cumulus Oophorus—A capillary-cumulus model was used(6). Briefly, EBSS/BSA and cumulus oophorus were succes-sively aspirated into a sterile glass capillary (Microcaps; Drum-mund Broomall, PA) to form columns of 3 and 2 cm in length,respectively. The end of the capillary with the cumulus columnwas dipped into a 100-�l droplet of sperm suspension contain-ing 10 � 106 motile spermatozoa/ml overlaid with mineral oil.Another capillary containing only EBSS/BSA serves as the con-trol. The setup was kept in a mobile in vitro fertilization cham-ber (Air-shields, Inc. Hatboro, PA) at 37 °C in an atmosphere of5% CO2 in air. After incubation for 1 h, the experimental cap-illary was cut at the interface between the cumulus column andthemedium column. Spermatozoa that had passed through thecumulus oophorus (penetrated spermatozoa) were collected inthe medium column. The control capillary was cut at a levelthat was similar to that of the experimental capillaries. Sperma-tozoa that had swum above the cutting level of the control cap-illary were collected as the control spermatozoa.The hemizona binding assay described (28) wasminiaturized
to determine the zona binding capacity of the above spermato-zoa. Unfertilized oocytes from assisted reproduction programwere microbisected into two identical hemizonae. In this mod-ified protocol, 6000 spermatozoa were incubated with eachhemizona in 30 �l of EBSS/BSA under mineral oil for 3 h at37 °C in an atmosphere of 5%CO2 in air. The numbers of tightlybound spermatozoa on the outer surface of the hemizonaewerecounted. The hemizona binding index (HZI) was defined as theratio of the number of bound spermatozoa in test droplet tothat in the control droplet times 100. The experiment wasrepeated 10 times using different sperm samples and cumulusmasses.Glycodelin immunoreactivity on penetrated spermatozoa
was determined by staining using monoclonal anti-glycodelinantibody (clone F43-7F9) recognizing the protein core of gly-codelins (12). Staining without the primary antibody was usedas control. The experiment was repeated five times with fivesemen samples. The presence of glycodelin immunoreactivityin these spermatozoa suggested the presence of glycodelin inthe cumulusmatrix (see “Results”).Western blot analysis of thecumulus matrix using the same antibody was used to confirmthe observation.IdentificationGlycodelin fromCumulusMatrix—Solubilized
cumulus matrix was diluted 1:5 (v/v) with Tris-buffered saline
3 The abbreviations used are: EBSS, Earle’s balanced salt solution; BSA, bovineserum albumin; HZI, hemizona binding index; TBS, Tris-buffered saline;FITC-PSA, fluorescein isothiocyanate-labeled peanut (P. sativum)agglutinin.
Glycodelin and Spermatozoa
FEBRUARY 23, 2007 • VOLUME 282 • NUMBER 8 JOURNAL OF BIOLOGICAL CHEMISTRY 5379
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from

(TBS). The glycodelin in the matrix were isolated by the mono-clonal anti-glycodelin antibody-Sepharose columnas describedabove. The bound glycodelin was eluted and further fraction-ated by aMono-Q column using a stepwise elution with 20 mMTris-HCl, 0.5 M NaCl (pH 7.0) at a flow rate of 100 �l/min. Theeluted fractions were concentrated by Microcon-10 (Milli-pore). Their purities were determined by SDS-PAGE.The bands in SDS-PAGE showing glycodelin immunoreac-
tivity were excised for in situ digestion with trypsin (0.1 mg/mlin 25 mM NH4HCO3, pH 8.0). The peptides were recoveredwith Millipore C18 ZipTips and 60% acetonitrile with 0.1% tri-fluoroacetic acid containing �-cyano-4-hydroxycinammic acidmatrix. The peptide-matrix samples were analyzed with thematrix-assisted laser desorption/ionization time-of-flight massspectrometry to obtain the peptide mass spectra, which werecompared with the protein spectra in the protein data bases atthe Swiss-Prot (www.ebi.ac.uk/swissprot/) as described (29).Biochemical Characterization of Cumulus Glycodelin—The
protein core of cumulus glycodelin was also studied by peptidemapping (12). In this experiment, 600 pmol of deglycosylatedglycodelin-A or -S or cumulus glycodelin was digested in 40 �lof 0.1 MNaHCO3with 2 units of agarose bound trypsin (Sigma)for 20 h at 37 °C. The resulting peptides in the supernatant wereanalyzed on a 15% SDS-PAGE.The isoelectric point of themolecule was determined by ana-
lytical isoelectric focusing on 5% polyacrylamide gel (30) in apH range of 3–10. The experiment was performed with 25 mMNaOH (cathode buffer) and 20mM acetic acid (anode buffer) at200 V for 1.5 h followed by 400 V for another 1.5 h. The proteinbands were stained by Coomassie Blue, and their pI valueswere estimated using the IEF calibration kits (AmershamBiosciences).Lectin binding assay (14) was used to study the glycosylation
of cumulus glycodelin. Briefly, the wells in 96-well plate werecoated with various lectins (10 �g/ml) overnight. Uncoatedsites in the well were blocked by incubation with 100 �l of 10mg/mlBSA in phosphate-buffered saline for 3 hwith slow shak-ing. Glycodelins (7.5 pmol in 25 �l) were then added and incu-bated overnight at 4 °C. After washing the wells twice with0.05% Tween 20-TBS, monoclonal anti-glycodelin antibody(clone F43-7F9, 2.5 �g in 200 �l) was added, and the mixturewas incubated for 2 h. The wells were washed four times and100�l of horseradish peroxidase-conjugated anti-mouse IgG ata dilution of 1:5000 (Sigma) was added. After 2 h of incubation,the wells were washed, and 100 �l/well of o-phenylenediamine(Sigma) was added. TheA490 was determinedwith amicroplatereader (MR5000; Dynatech, Embrach, Switzerland). The assaywas done at room temperature. The control wells containedBSA instead of glycodelin.The binding affinity of cumulus glycodelin to a monoclonal
anti-glycodelin antibody was compared with that of other gly-codelin isoforms in an enzyme-linked immunosorbent assay(12). Briefly, 0.03–6 pmol/ml of cumulus glycodelin and gly-codelin-A and -F were coated onto a 96-well plate for 3 h atroom temperature with slow shaking. Uncoated sites in eachwell were blocked by incubationwith 100�l of 3% BSA/TBS for2 h. The wells were then washed with Tween 20-TBS followedby the addition of a limiting concentration of anti-glycodelin
antibody (clone F43-7F9, 1000 ng/ml) in TBS (12). After over-night incubation at 4 °C, the wells were washed thrice withTween 20-TBS and fluorescein-conjugated goat anti-mouseIgG (Sigma) at a dilution of 1:300 was then added. The boundfluorescence was measured with a fluorometer (FL600;Bio-Tek).Biological Characterization of Cumulus Glycodelin—The
equilibrium binding of cumulus glycodelin to spermatozoa wasdetermined as described (14). Four semen samples were used(n � 4). Spermatozoa (2 � 106 motile spermatozoa/ml) in 100�l of EBSS/BSA were incubated with different concentrations(0.162343 pmol/ml) of iodinated cumulus glycodelin at 37 °Cfor 1 h. The binding was terminated by the addition of 1.5 ml ofice-cold phosphate-buffered saline followed by centrifugationat 300 g for 3 min. After washing the spermatozoa with freshEBSS/BSA, the sperm-bound radioactivity was counted with agamma counter (model 5500B; Beckman, Fullerton, CA). Thespecific binding of cumulus glycodelin was determined by sub-tracting the sperm-bound radioactivity in the presence of a50-fold higher concentration of unlabeled glycodelin from thatin the absence of unlabeled protein. The determinations of totalbinding and nonspecific binding were done in duplicate.Competition binding analysis was used to compare the affin-
ity of cumulus glycodelin and other lipocalins to spermatozoa(14). The binding of 200 pmol/ml of 125I-cumulus glycodelin to2 � 105 motile spermatozoa (n � 3) in 100 �l of EBSS/BSAwasdetermined in the presence of increasing concentrations (2, 20,200, 1000, and 2000 pmol/ml) of unlabeled cumulus glycodelin,glycodelin-A, -F, and -S, deglycosylated glycodelin and bovine�-lactoglobulin A (Sigma), or buffer alone at 37 °C for 360 min.The cell-bound radioactivity was then determined. Each indi-vidual experiment was repeated twice.The binding of purified cumulus glycodelin to spermatozoa
was visualized by immunohistochemical staining after the incu-bation of capacitated spermatozoa (n � 5, 2 � 106 spermato-zoa/ml) with 200 pmol/ml cumulus glycodelin for 60 min at37°C under 5% CO2 in air. Binding of deglycosylated glycodelinwas used as a control.Hemizona binding assay was used to study the effect of
cumulus glycodelin on spermatozoa-zona pellucida binding.Briefly, capacitated spermatozoa (2� 106 spermatozoa/ml; n�5) were incubated with 0.3, 3, 30, 300, or 750 pmol/ml cumulusglycodelin or EBSS/BSA (control) at 37 °C in an atmosphere of5% CO2 in air. After 1 h of incubation, the spermatozoa werewashed with fresh EBSS/BSA. Standard protocol of hemizonabinding assay (28) was performed on the 20,000 treated sper-matozoa in 100 �l of EBSS/BSA. Glycodelin-A, -F, and -S anddeglycosylated glycodelin were used as controls.The effect of cumulus glycodelin on spermmotility, viability,
and acrosomal status were also determined. Fluorescein iso-thiocyanate-labeled peanut (Pisum sativum) agglutinin (FITC-PSA; Sigma) and Hoechst staining techniques were used toevaluate the acrosomal status of spermatozoa (16). The fluores-cence patterns of 300 spermatozoa in randomly selected fieldswere determined under a fluorescence microscope (Zeiss,Oberkochen, Germany) with 600� magnification. Spermato-zoawithoutHoechst and FITC-PSA staining orwith FITC-PSAstaining confined to the equatorial segment only were consid-
Glycodelin and Spermatozoa
5380 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 8 • FEBRUARY 23, 2007
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from

ered as acrosome reacted. Hobson Sperm Tracker System(Hobson Tracking Systems Ltd., Sheffield, UK) was used tostudy spermmotility. The set-up parameters of the system andthe procedures are described elsewhere (12).Effects of Cumulus Penetration on the Zona Binding Capacity
of Glycodelin-A- and -F-treated Spermatozoa—Capacitatedspermatozoa (2 � 106 spermatozoa/ml) were incubated with300 pmol/ml of glycodelin-F or -A, deglycosylated glycodelin orEBSS/BSA (control) at 37 °C in an atmosphere of 5% CO2 in airfor 2 h. They were then washed and allowed to penetrate thecumulus oophorus column in the capillary as described above.The penetrated spermatozoa were washed with fresh EBSS/BSA, and their zona binding capacities were determined byhemizona binding assay. The experimentwas repeated 10 timeswith different sperm samples and cumulus masses.The fate of sperm-bound glycodelin-F and -A after cumulus
oophorus penetration was investigated by repeating the aboveexperiment using iodinated or Alexa 488 (Alexa Fluor� 488protein labeling kit; Molecular Probes, Carlsbad, CA)-conju-gated glycodelin-F and -A. The penetrated spermatozoa werecollected and either smeared on slides to observe under a fluo-rescence microscope with an excitation filter BP 450–490,chromatic beam splitter FT510, and barrier filter LP520 orcounted for their associated radioactivity as above.Effects of Cumulus Glycodelin on Sperm-bound Glycodelin-A
and -F—Iodinated or Alexa 488-conjugated glycodelin-A or -F(300 pmol/ml) were incubated with 2 � 105 motile spermato-zoa (n � 3) in 100 �l of EBSS with 0.3% BSA at 37 °C for 120min. Unlabeled glycodelin-A or -F, cumulus glycodelin, or deg-lycosylated glycodelin (200 pmol/ml) was then added. After afurther incubation for 0, 2, 5, 10, 15, 20, 30, 60, or 90 min,ice-cold buffer was added to stop the dissociation. The sperma-tozoa were then washed, and their associated radioactivity orfluorescence was determined as described above.Processing of Glycodelin by Cumulus Cells—The dispersed
cumulus cells were washed twice in TC-199 medium (Sigma)and resuspended to a final concentration of 4� 106 viable cells/ml. Trypan blue exclusion test was employed to determine theviability of the cells. They were then cultured in 2ml of TC-199medium supplemented with 20% fetal bovine serum (Sigma) asdescribed (31). After 24 h, 300 pmol/ml glycodelin-A or -F wasadded to the culture medium. The spent media were collectedafter 10 h and diluted 1:5 (v/v) with TBS. 94% of the cellsremained viable at this time. The cell debris in the conditionedmedia was removed by centrifugation at 500 � g for 5 min.The glycodelin in the media were immunoaffinity-purifiedusing monoclonal anti-glycodelin antibody-Sepharose col-umn. The eluates from 10 runs were combined and analyzedby 15% SDS-PAGE.Data Analysis—All of the data were expressed as the
means � S.E. The data were analyzed by statistical softwares(SigmaPlot 8.02; Ligand Binding Analysis Module & SigmaStat2.03, Jandel Scientific, San Rafael, CA). For all experiments, thenonparametric analysis of variance on Rank test for multiplecomparisons followed by MannWhitneyU test were used. Aprobability value �0.05 was considered to be statisticallysignificant.
RESULTS
Cumulus Oophorus-penetrated Spermatozoa Had IncreasedZona Binding Capacity and Glycodelin Immunoreactivity—Eight semen samples were recruited. By comparing with sper-matozoa in the control capillary, the cumulus oophorus pene-trated spermatozoa had a higher zona binding capacity with thenumber of tightly bound spermatozoa on the hemizona (72.6�5.6), significantly higher than that of the control spermatozoa(58.3 � 6.2), and the HZI was increased from 112.4 � 3.8 to136.4 � 6.2. Indirect immunofluorescence staining revealedglycodelin immunoreactivity on the whole head of cumuluspenetrated spermatozoa, the signal being stronger on the equa-torial region (Fig. 1A). This staining pattern was different fromthat of the glycodelin-A or -F-treated spermatozoa (12). Thepercentages of spermatozoa with positive immunoreactivitywere�82–94%. In contrast, no immunoreactivitywas observedon spermatozoa in the control capillary or with the omission ofthe primary antibody (data not shown).Cumulus Matrix Contains a Glycodelin Isoform—The above
experiment suggested the presence of glycodelin in the cumu-lus matrix. This was confirmed by Western blot analysis of theextracellularmatrix of cumulus oophorus using anti-glycodelinantibody. The data revealed two major bands of sizes �30 and�25 kDa. The molecular size of the larger band was similar tothat of glycodelin-A (Fig. 1B) and -F (12).Liquid chromatographywas used to isolate the glycodelins in
the cumulus matrix. A total of 250 cumulus samples were usedfor the purification. A typical ion exchange chromatogram ofthe eluate of cumulus matrix components bound to anti-gly-codelin-Sepharose column is shown in Fig. 2A. There werethree major peaks, named fractions F1, F2, and F3 according totheir ionic strength. SDS-PAGE showed that all fractions con-tained a major protein band of sizes larger than that of thedeglycosylated glycodelin-A or -F (Fig. 2B, lanes 4 and 5). Themolecular masses of F1 (Fig. 2B, lane 1) and F2 (Fig. 2B, lane 2)were similar to that glycodelin-A and -F (�30 kDa). Based ontheir mass spectrometry pattern, ionic strength, molecularweights, lectin binding properties, and immunoreactivity (datanot shown), F1 and F2 were glycodelin-A and -F, respectively.F3 contained the cumulus glycodelin with size �25 kDa that
FIGURE 1. Glycodelin immunoreactivity in the extracellular matrix ofcumulus-oophorus. A, immunofluorescent staining of glycodelin in humanspermatozoa after cumulus penetration. B, Western blot analysis of cumulusmatrix using monoclonal anti-glycodelin antibody. First lane, cumulus matrix;second lane, 1 �g of glycodelin-A.
Glycodelin and Spermatozoa
FEBRUARY 23, 2007 • VOLUME 282 • NUMBER 8 JOURNAL OF BIOLOGICAL CHEMISTRY 5381
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from
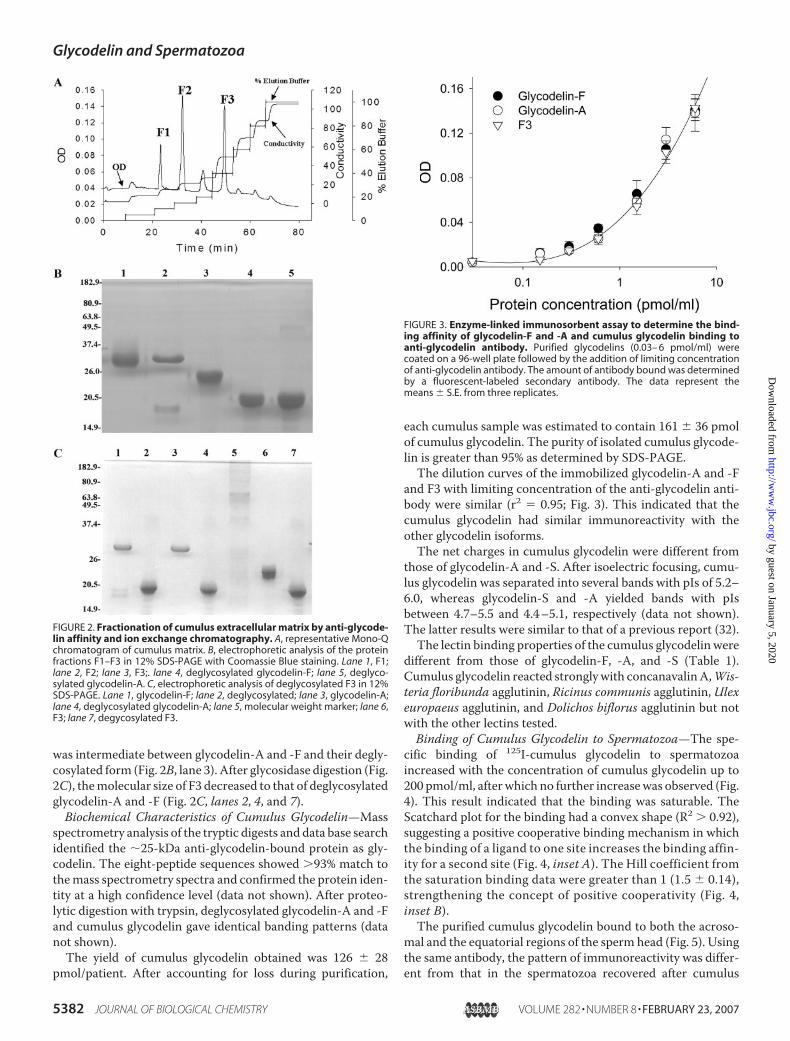
was intermediate between glycodelin-A and -F and their degly-cosylated form (Fig. 2B, lane 3). After glycosidase digestion (Fig.2C), themolecular size of F3 decreased to that of deglycosylatedglycodelin-A and -F (Fig. 2C, lanes 2, 4, and 7).Biochemical Characteristics of Cumulus Glycodelin—Mass
spectrometry analysis of the tryptic digests and data base searchidentified the �25-kDa anti-glycodelin-bound protein as gly-codelin. The eight-peptide sequences showed �93% match tothemass spectrometry spectra and confirmed the protein iden-tity at a high confidence level (data not shown). After proteo-lytic digestion with trypsin, deglycosylated glycodelin-A and -Fand cumulus glycodelin gave identical banding patterns (datanot shown).The yield of cumulus glycodelin obtained was 126 � 28
pmol/patient. After accounting for loss during purification,
each cumulus sample was estimated to contain 161 � 36 pmolof cumulus glycodelin. The purity of isolated cumulus glycode-lin is greater than 95% as determined by SDS-PAGE.The dilution curves of the immobilized glycodelin-A and -F
and F3 with limiting concentration of the anti-glycodelin anti-body were similar (r2 � 0.95; Fig. 3). This indicated that thecumulus glycodelin had similar immunoreactivity with theother glycodelin isoforms.The net charges in cumulus glycodelin were different from
those of glycodelin-A and -S. After isoelectric focusing, cumu-lus glycodelin was separated into several bands with pIs of 5.2–6.0, whereas glycodelin-S and -A yielded bands with pIsbetween 4.7–5.5 and 4.4–5.1, respectively (data not shown).The latter results were similar to that of a previous report (32).The lectin binding properties of the cumulus glycodelin were
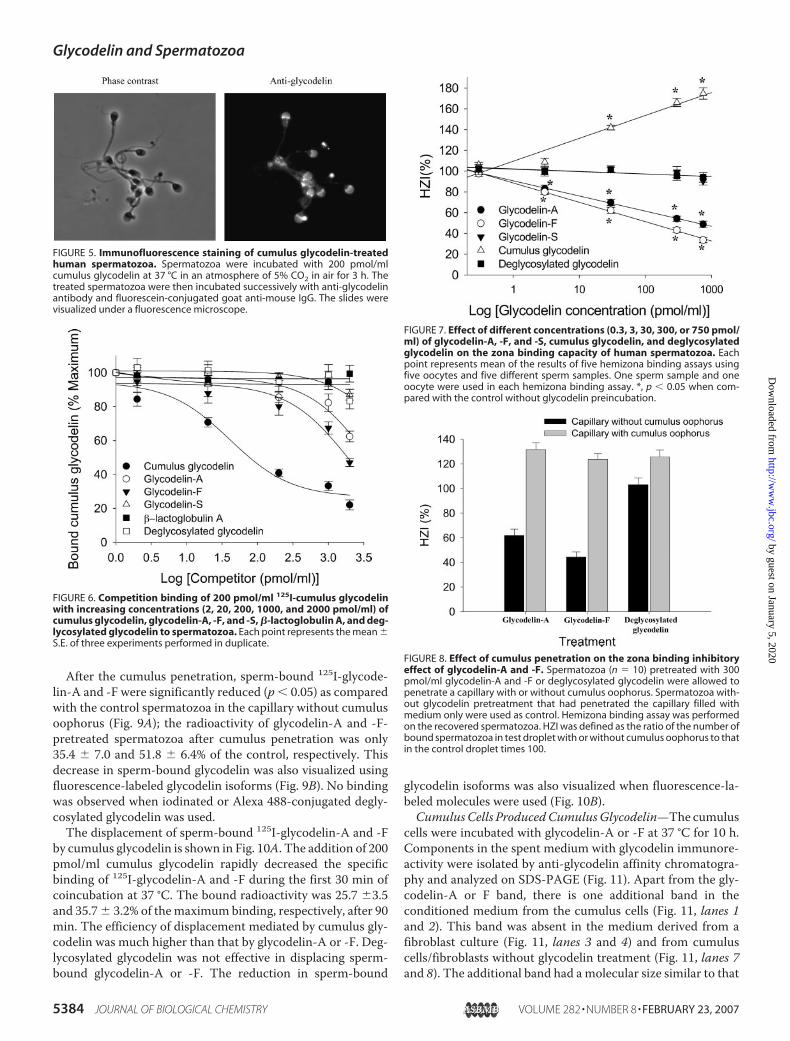
different from those of glycodelin-F, -A, and -S (Table 1).Cumulus glycodelin reacted stronglywith concanavalinA,Wis-teria floribunda agglutinin, Ricinus communis agglutinin, Ulexeuropaeus agglutinin, and Dolichos biflorus agglutinin but notwith the other lectins tested.Binding of Cumulus Glycodelin to Spermatozoa—The spe-
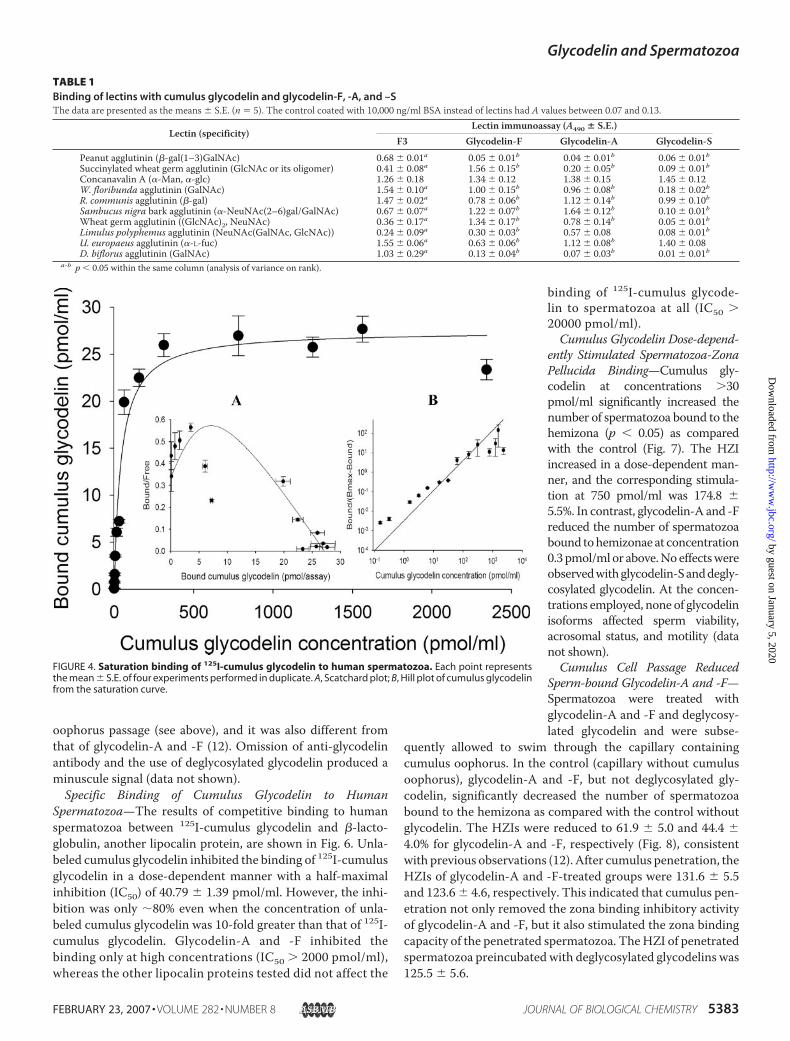
cific binding of 125I-cumulus glycodelin to spermatozoaincreased with the concentration of cumulus glycodelin up to200 pmol/ml, afterwhich no further increasewas observed (Fig.4). This result indicated that the binding was saturable. TheScatchard plot for the binding had a convex shape (R2 � 0.92),suggesting a positive cooperative binding mechanism in whichthe binding of a ligand to one site increases the binding affin-ity for a second site (Fig. 4, inset A). The Hill coefficient fromthe saturation binding data were greater than 1 (1.5 � 0.14),strengthening the concept of positive cooperativity (Fig. 4,inset B).The purified cumulus glycodelin bound to both the acroso-
mal and the equatorial regions of the sperm head (Fig. 5). Usingthe same antibody, the pattern of immunoreactivity was differ-ent from that in the spermatozoa recovered after cumulus
FIGURE 2. Fractionation of cumulus extracellular matrix by anti-glycode-lin affinity and ion exchange chromatography. A, representative Mono-Qchromatogram of cumulus matrix. B, electrophoretic analysis of the proteinfractions F1–F3 in 12% SDS-PAGE with Coomassie Blue staining. Lane 1, F1;lane 2, F2; lane 3, F3;. lane 4, deglycosylated glycodelin-F; lane 5, deglyco-sylated glycodelin-A. C, electrophoretic analysis of deglycosylated F3 in 12%SDS-PAGE. Lane 1, glycodelin-F; lane 2, deglycosylated; lane 3, glycodelin-A;lane 4, deglycosylated glycodelin-A; lane 5, molecular weight marker; lane 6,F3; lane 7, degycosylated F3.
FIGURE 3. Enzyme-linked immunosorbent assay to determine the bind-ing affinity of glycodelin-F and -A and cumulus glycodelin binding toanti-glycodelin antibody. Purified glycodelins (0.03– 6 pmol/ml) werecoated on a 96-well plate followed by the addition of limiting concentrationof anti-glycodelin antibody. The amount of antibody bound was determinedby a fluorescent-labeled secondary antibody. The data represent themeans � S.E. from three replicates.
Glycodelin and Spermatozoa
5382 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 8 • FEBRUARY 23, 2007
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from
oophorus passage (see above), and it was also different fromthat of glycodelin-A and -F (12). Omission of anti-glycodelinantibody and the use of deglycosylated glycodelin produced aminuscule signal (data not shown).Specific Binding of Cumulus Glycodelin to Human
Spermatozoa—The results of competitive binding to humanspermatozoa between 125I-cumulus glycodelin and �-lacto-globulin, another lipocalin protein, are shown in Fig. 6. Unla-beled cumulus glycodelin inhibited the binding of 125I-cumulusglycodelin in a dose-dependent manner with a half-maximalinhibition (IC50) of 40.79 � 1.39 pmol/ml. However, the inhi-bition was only �80% even when the concentration of unla-beled cumulus glycodelin was 10-fold greater than that of 125I-cumulus glycodelin. Glycodelin-A and -F inhibited thebinding only at high concentrations (IC50 � 2000 pmol/ml),whereas the other lipocalin proteins tested did not affect the
binding of 125I-cumulus glycode-lin to spermatozoa at all (IC50 �20000 pmol/ml).Cumulus Glycodelin Dose-depend-
ently Stimulated Spermatozoa-ZonaPellucida Binding—Cumulus gly-codelin at concentrations �30pmol/ml significantly increased thenumber of spermatozoa bound to thehemizona (p � 0.05) as comparedwith the control (Fig. 7). The HZIincreased in a dose-dependent man-ner, and the corresponding stimula-tion at 750 pmol/ml was 174.8 �5.5%. In contrast, glycodelin-A and -Freduced the number of spermatozoabound tohemizonaeat concentration0.3pmol/mlor above.Noeffectswereobservedwithglycodelin-Sanddegly-cosylated glycodelin. At the concen-trations employed, none of glycodelinisoforms affected sperm viability,acrosomal status, and motility (datanot shown).Cumulus Cell Passage Reduced
Sperm-bound Glycodelin-A and -F—Spermatozoa were treated withglycodelin-A and -F and deglycosy-lated glycodelin and were subse-
quently allowed to swim through the capillary containingcumulus oophorus. In the control (capillary without cumulusoophorus), glycodelin-A and -F, but not deglycosylated gly-codelin, significantly decreased the number of spermatozoabound to the hemizona as compared with the control withoutglycodelin. The HZIs were reduced to 61.9 � 5.0 and 44.4 �4.0% for glycodelin-A and -F, respectively (Fig. 8), consistentwith previous observations (12). After cumulus penetration, theHZIs of glycodelin-A and -F-treated groups were 131.6 � 5.5and 123.6� 4.6, respectively. This indicated that cumulus pen-etration not only removed the zona binding inhibitory activityof glycodelin-A and -F, but it also stimulated the zona bindingcapacity of the penetrated spermatozoa. TheHZI of penetratedspermatozoa preincubated with deglycosylated glycodelins was125.5 � 5.6.
FIGURE 4. Saturation binding of 125I-cumulus glycodelin to human spermatozoa. Each point representsthe mean � S.E. of four experiments performed in duplicate. A, Scatchard plot; B, Hill plot of cumulus glycodelinfrom the saturation curve.
TABLE 1Binding of lectins with cumulus glycodelin and glycodelin-F, -A, and –SThe data are presented as the means � S.E. (n � 5). The control coated with 10,000 ng/ml BSA instead of lectins had A values between 0.07 and 0.13.
Lectin (specificity)Lectin immunoassay (A490 � S.E.)
F3 Glycodelin-F Glycodelin-A Glycodelin-SPeanut agglutinin (�-gal(1–3)GalNAc) 0.68 � 0.01a 0.05 � 0.01b 0.04 � 0.01b 0.06 � 0.01bSuccinylated wheat germ agglutinin (GlcNAc or its oligomer) 0.41 � 0.08a 1.56 � 0.15b 0.20 � 0.05b 0.09 � 0.01bConcanavalin A (�-Man, �-glc) 1.26 � 0.18 1.34 � 0.12 1.38 � 0.15 1.45 � 0.12W. floribunda agglutinin (GalNAc) 1.54 � 0.10a 1.00 � 0.15b 0.96 � 0.08b 0.18 � 0.02bR. communis agglutinin (�-gal) 1.47 � 0.02a 0.78 � 0.06b 1.12 � 0.14b 0.99 � 0.10bSambucus nigra bark agglutinin (�-NeuNAc(2–6)gal/GalNAc) 0.67 � 0.07a 1.22 � 0.07b 1.64 � 0.12b 0.10 � 0.01bWheat germ agglutinin ((GlcNAc)2, NeuNAc) 0.36 � 0.17a 1.34 � 0.17b 0.78 � 0.14b 0.05 � 0.01bLimulus polyphemus agglutinin (NeuNAc(GalNAc, GlcNAc)) 0.24 � 0.09a 0.30 � 0.03b 0.57 � 0.08 0.08 � 0.01bU. europaeus agglutinin (�-L-fuc) 1.55 � 0.06a 0.63 � 0.06b 1.12 � 0.08b 1.40 � 0.08D. biflorus agglutinin (GalNAc) 1.03 � 0.29a 0.13 � 0.04b 0.07 � 0.03b 0.01 � 0.01b
a-b p � 0.05 within the same column (analysis of variance on rank).
Glycodelin and Spermatozoa
FEBRUARY 23, 2007 • VOLUME 282 • NUMBER 8 JOURNAL OF BIOLOGICAL CHEMISTRY 5383
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from
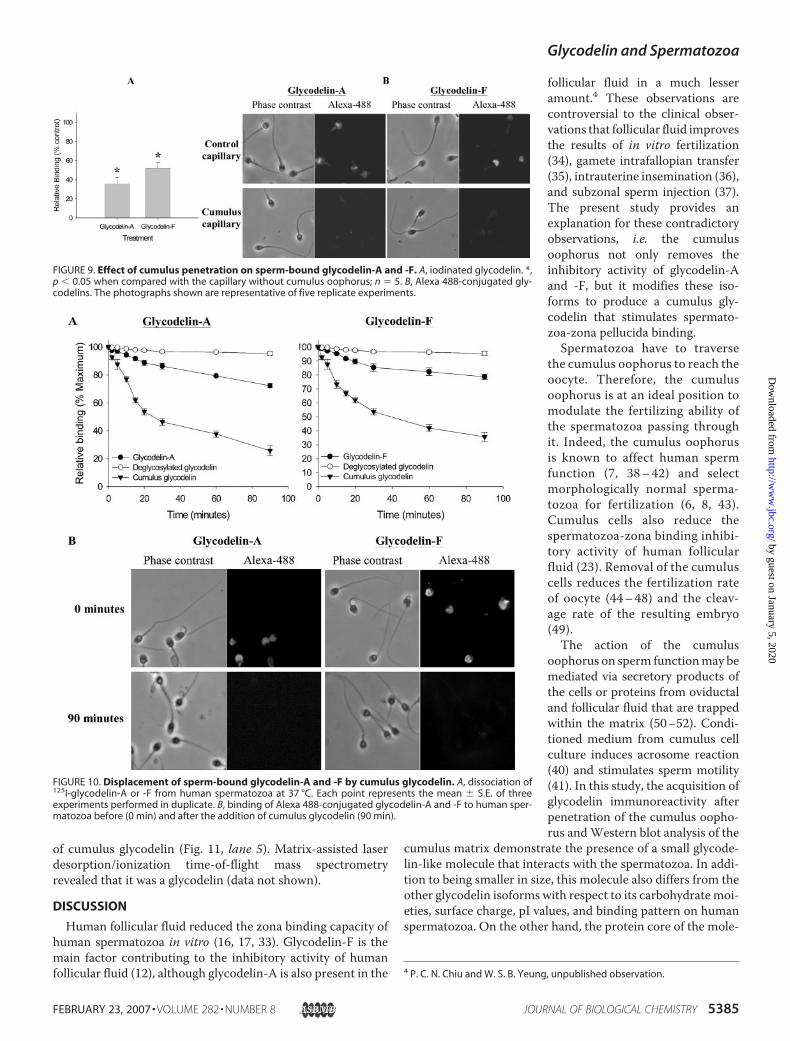
After the cumulus penetration, sperm-bound 125I-glycode-lin-A and -F were significantly reduced (p � 0.05) as comparedwith the control spermatozoa in the capillary without cumulusoophorus (Fig. 9A); the radioactivity of glycodelin-A and -F-pretreated spermatozoa after cumulus penetration was only35.4 � 7.0 and 51.8 � 6.4% of the control, respectively. Thisdecrease in sperm-bound glycodelin was also visualized usingfluorescence-labeled glycodelin isoforms (Fig. 9B). No bindingwas observed when iodinated or Alexa 488-conjugated degly-cosylated glycodelin was used.The displacement of sperm-bound 125I-glycodelin-A and -F
by cumulus glycodelin is shown in Fig. 10A. The addition of 200pmol/ml cumulus glycodelin rapidly decreased the specificbinding of 125I-glycodelin-A and -F during the first 30 min ofcoincubation at 37 °C. The bound radioactivity was 25.7 �3.5and 35.7� 3.2% of themaximum binding, respectively, after 90min. The efficiency of displacement mediated by cumulus gly-codelin was much higher than that by glycodelin-A or -F. Deg-lycosylated glycodelin was not effective in displacing sperm-bound glycodelin-A or -F. The reduction in sperm-bound
glycodelin isoforms was also visualized when fluorescence-la-beled molecules were used (Fig. 10B).CumulusCells ProducedCumulusGlycodelin—The cumulus
cells were incubated with glycodelin-A or -F at 37 °C for 10 h.Components in the spent medium with glycodelin immunore-activity were isolated by anti-glycodelin affinity chromatogra-phy and analyzed on SDS-PAGE (Fig. 11). Apart from the gly-codelin-A or F band, there is one additional band in theconditioned medium from the cumulus cells (Fig. 11, lanes 1and 2). This band was absent in the medium derived from afibroblast culture (Fig. 11, lanes 3 and 4) and from cumuluscells/fibroblasts without glycodelin treatment (Fig. 11, lanes 7and 8). The additional band had amolecular size similar to that
FIGURE 5. Immunofluorescence staining of cumulus glycodelin-treatedhuman spermatozoa. Spermatozoa were incubated with 200 pmol/mlcumulus glycodelin at 37 °C in an atmosphere of 5% CO2 in air for 3 h. Thetreated spermatozoa were then incubated successively with anti-glycodelinantibody and fluorescein-conjugated goat anti-mouse IgG. The slides werevisualized under a fluorescence microscope.
FIGURE 6. Competition binding of 200 pmol/ml 125I-cumulus glycodelinwith increasing concentrations (2, 20, 200, 1000, and 2000 pmol/ml) ofcumulus glycodelin, glycodelin-A, -F, and -S, �-lactoglobulin A, and deg-lycosylated glycodelin to spermatozoa. Each point represents the mean �S.E. of three experiments performed in duplicate.
FIGURE 7. Effect of different concentrations (0.3, 3, 30, 300, or 750 pmol/ml) of glycodelin-A, -F, and -S, cumulus glycodelin, and deglycosylatedglycodelin on the zona binding capacity of human spermatozoa. Eachpoint represents mean of the results of five hemizona binding assays usingfive oocytes and five different sperm samples. One sperm sample and oneoocyte were used in each hemizona binding assay. *, p � 0.05 when com-pared with the control without glycodelin preincubation.
FIGURE 8. Effect of cumulus penetration on the zona binding inhibitoryeffect of glycodelin-A and -F. Spermatozoa (n � 10) pretreated with 300pmol/ml glycodelin-A and -F or deglycosylated glycodelin were allowed topenetrate a capillary with or without cumulus oophorus. Spermatozoa with-out glycodelin pretreatment that had penetrated the capillary filled withmedium only were used as control. Hemizona binding assay was performedon the recovered spermatozoa. HZI was defined as the ratio of the number ofbound spermatozoa in test droplet with or without cumulus oophorus to thatin the control droplet times 100.
Glycodelin and Spermatozoa
5384 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 8 • FEBRUARY 23, 2007
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from
of cumulus glycodelin (Fig. 11, lane 5). Matrix-assisted laserdesorption/ionization time-of-flight mass spectrometryrevealed that it was a glycodelin (data not shown).
DISCUSSION
Human follicular fluid reduced the zona binding capacity ofhuman spermatozoa in vitro (16, 17, 33). Glycodelin-F is themain factor contributing to the inhibitory activity of humanfollicular fluid (12), although glycodelin-A is also present in the
follicular fluid in a much lesseramount.4 These observations arecontroversial to the clinical obser-vations that follicular fluid improvesthe results of in vitro fertilization(34), gamete intrafallopian transfer(35), intrauterine insemination (36),and subzonal sperm injection (37).The present study provides anexplanation for these contradictoryobservations, i.e. the cumulusoophorus not only removes theinhibitory activity of glycodelin-Aand -F, but it modifies these iso-forms to produce a cumulus gly-codelin that stimulates spermato-zoa-zona pellucida binding.Spermatozoa have to traverse
the cumulus oophorus to reach theoocyte. Therefore, the cumulusoophorus is at an ideal position tomodulate the fertilizing ability ofthe spermatozoa passing throughit. Indeed, the cumulus oophorusis known to affect human spermfunction (7, 38–42) and selectmorphologically normal sperma-tozoa for fertilization (6, 8, 43).Cumulus cells also reduce thespermatozoa-zona binding inhibi-tory activity of human follicularfluid (23). Removal of the cumuluscells reduces the fertilization rateof oocyte (44–48) and the cleav-age rate of the resulting embryo(49).The action of the cumulus
oophorus on sperm functionmay bemediated via secretory products ofthe cells or proteins from oviductaland follicular fluid that are trappedwithin the matrix (50–52). Condi-tioned medium from cumulus cellculture induces acrosome reaction(40) and stimulates sperm motility(41). In this study, the acquisition ofglycodelin immunoreactivity afterpenetration of the cumulus oopho-rus andWestern blot analysis of the
cumulus matrix demonstrate the presence of a small glycode-lin-like molecule that interacts with the spermatozoa. In addi-tion to being smaller in size, this molecule also differs from theother glycodelin isoforms with respect to its carbohydratemoi-eties, surface charge, pI values, and binding pattern on humanspermatozoa. On the other hand, the protein core of the mole-
4 P. C. N. Chiu and W. S. B. Yeung, unpublished observation.
FIGURE 9. Effect of cumulus penetration on sperm-bound glycodelin-A and -F. A, iodinated glycodelin. *,p � 0.05 when compared with the capillary without cumulus oophorus; n � 5. B, Alexa 488-conjugated gly-codelins. The photographs shown are representative of five replicate experiments.
FIGURE 10. Displacement of sperm-bound glycodelin-A and -F by cumulus glycodelin. A, dissociation of125I-glycodelin-A or -F from human spermatozoa at 37 °C. Each point represents the mean � S.E. of threeexperiments performed in duplicate. B, binding of Alexa 488-conjugated glycodelin-A and -F to human sper-matozoa before (0 min) and after the addition of cumulus glycodelin (90 min).
Glycodelin and Spermatozoa
FEBRUARY 23, 2007 • VOLUME 282 • NUMBER 8 JOURNAL OF BIOLOGICAL CHEMISTRY 5385
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from

cule is identical to the other glycodelin isoforms as evident bytheir similarity in molecular size after deglycosylation, massspectrometry pattern, affinity to anti-glycodelin antibody, andprofile of the peptide fragments after trypsin digestion. Basedon these observations, it is reasonable to name this molecule asglycodelin-C (cumulus matrix isoform of glycodelin) to be inagreement with the existing nomenclature of glycodelin.The luteinized granulosa cells but not the cumulus cells pos-
sess glycodelin mRNA, suggesting that the former is the site ofbiosynthesis of glycodelin (24). However, the cumulus cells cantake up specifically exogenous glycodelin but not the otherlipocalin family members, thereby modifying the uptaken gly-codelin to glycodelin-C (24). The present study confirms theseprevious observations and further demonstrates that the gly-codelin-C herein produced is released back into the culturemedium. Cumulus cells contain cytoplasmic glycodelin immu-noreactivity but not glycodelin mRNA (24). Therefore, it islikely that exogenous glycodelin is internalized in cumulus cellsprobably by cell surface receptor-mediated endocytosis similarto that found in other lipocalin familymembers (53, 54). Cumu-lus cells have well developed machinery for glycan manipula-tion and produce plenty of carbohydrate-containing extracel-lular matrix molecules, such as proteoglycan (55), at theperiovulatory period. Thus, the conversion of exogenous gly-codelin to glycodelin-C is likely to occur intracellularly by amechanism yet to be defined. However, these observations donot exclude the possibility that part of the conversion could bedone on the plasma membrane of the cumulus cells. Glycohy-drolase activities have been demonstrated on the plasmamem-brane of many cell types (56–59), although their presence onthe surface of cumulus has not been explored. These possibili-ties are under investigation in our laboratory.
This is the first report on characterization of the binding ofglycodelin-C to human spermatozoa. The binding is specific,and it is time- and concentration-dependent. Three observa-tions indicate that the binding sites for glycodelin-C are differ-ent from those of the other glycodelin isoforms (14, 18, 60).First, the equilibrium binding of glycodelin-C to spermatozoasuggests a positive cooperative binding mechanism that doesnot occur in the binding glycodelin-A, -F, and -S to spermato-zoa. Second, the binding sites of glycodelin-A and -F are foundevenly on the acrosomal region (12), whereas those of glycode-lin-C have differential distribution with the highest density andaffinity at the equatorial region. The binding sites of glycode-lin-S spread evenly over the whole head and had a low affinityfor glycodelin-S (14). Third, the other glycodelin isoforms can-not significantly compete with glycodelin-C for its binding sitesin the competition binding assay. Likewise, another lipocalinfamily member, �-lactoglobulin, is ineffective in competingwith glycodelin-C.In this study, the binding pattern of purified glycodelin-C
on spermatozoa is different from that of glycodelin immu-noreactivity on spermatozoa that had penetrated the cumu-lus oophorus; the former is confined to the acrosome andequatorial region, whereas the latter also includes the post-acrosomal region. The question of how the cumulus oopho-rus modifies the binding of glycodelin to spermatozoa is sofar unknown. The hyaluronan, proteoglycans, and proteinsin the cumulus extracellular matrix (55) modulate protein-receptor binding (61, 62). Therefore, it is possible that thecumulus oophorus induces the exposure and/or functioningof glycodelin receptors on the post-acrosomal region ofspermatozoa. This hypothesis is under investigation in ourlaboratory.Interestingly, treatment of spermatozoa with either glycode-
lin-A or -F prior to cumulus penetration did not affect the zonapellucida binding capacity of the penetrated spermatozoa. Infact, cumulus penetration has two effects on the penetratedspermatozoa, namely removal of the inhibitory effect of gly-codelin-A and -F and stimulation of the zona binding capacityof the penetrated spermatozoa. The former effect is consistentwith significant reduction of sperm-bound glycodelin-A and -F.Glycodelin-C is likely to be at least partly responsible for reduc-ing the bound glycodelin isoforms because it could effectivelydisplace sperm-bound glycodelin-A and -F to �30 and 40% ofthe maximum binding, respectively, after 60 min. Indeed, itsdisplacement activity is more effective than that of glycode-lin-A or -F. An efficient mechanism for removal of bound gly-codelin-A and -F is likely to be important for fertilization,because human spermatozoa can penetrate the cumulusoophorus within 1 h in vitro (6, 63, 64).Glycodelin-C is also likely to be responsible for the second
effect of cumulus penetration on the penetrated spermatozoabecause it stimulates spermatozoa zona-pellucida binding in adose-dependent manner. No other glycodelin isoform has asimilar stimulatory activity. The stimulatory activity of glycode-lin-C is not due to the effect on acrosome reaction and spermmotility, and it is no exception to the general observation thatthe activity of glycodelin isoforms on spermatozoa-zona pellu-cida interaction is glycosylation-dependent (12, 32, 65). Gly-
FIGURE 11. Detection of cumulus glycodelin in condition medium ofcumulus cells by anti-glycodelin affinity chromatography and 15% SDS-PAGE. Lane 1, conditioned medium of human cumulus cells after glycode-lin-A treatment; lane 2, conditioned medium of human cumulus cells afterglycodelin-F treatment; lane 3, conditioned medium of human fibroblastafter glycodelin-A treatment. lane 4, conditioned medium of human fibro-blast after glycodelin-F treatment; lane 5, purified cumulus glycodelin; lane 6,purified glycodelin-A; lane 7, conditioned medium of human cumulus cellswithout glycodelin treatment; lane 8, conditioned medium of human fibro-blast without glycodelin treatment.
Glycodelin and Spermatozoa
5386 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 8 • FEBRUARY 23, 2007
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from

codelin-Swith glycosylation different from that of glycodelin-Aand -F lacks the spermatozoa-zona binding inhibitory activity(12, 32, 65).Carbohydrate recognition is important in spermatozoa-zona
pellucida binding, although the details of the molecular inter-actions between spermmembrane and zona pellucida proteinsremain elusive (66–68). The zona binding stimulatory activityof glycodelin-C suggests that changes in glycosylation of gly-codelin can have significant effects on spermatozoa-zona pel-lucida binding. Comparison of the glycosylation of differentglycodelin isoforms is ongoing to identify the role of differentglycans in the regulation of spermatozoa-zona pellucida bind-ing. Modification of biological activity resulting from thechanges in glycosylation has been demonstrated in differentglycohormones and glycoproteins (69).How glycodelin-C stimulates spermatozoa-zona binding is
still largely unknown. It may act as a “bridge” between sperma-tozoa and zona pellucida, using its unique carbohydrate moi-eties to promote binding between the spermmembrane recep-tor(s) and the zona pellucida proteins. A recent reportdemonstrated that when glycodelin binds to its receptor on Tcells, CD45, it can act as a calcium-dependent lectin to bindother T cell surface glycoproteins to mediate its immunoregu-latory activities (70). Gonadotropin-releasing hormoneincreases spermatozoa-zona binding via an elevation of intra-cellular calcium through T-type voltage-operated calciumchannels (71). It is also possible that glycodelin-C induces signaltransduction leading to the formation of more functional zonapellucida receptors. In mouse, tyrosine phosphorylation-asso-ciated change of endoplasmin and heat shock protein 60 inspermatozoa facilitates the formation of functional zona pellu-cida receptors, resulting in increased zona pellucida bindingability (72). These possibilities are being investigated in ourlaboratory.Based on the results of this report and previous results, we
propose that the following sequence of events take place at thefertilization site: Glycodelin is synthesized in the granulosa cellsof the late secondary follicle. During the periovulatory period,the cumulus cells use glycodelin-A and -F in the follicular fluidas substrate for the production of the glycodelin-C, which thenis released into the extracellular matrix during cumulus expan-sion or shortly after ovulation. Glycodelin-F and probably ovi-ductal-derived glycodelin (19) binds to spermatozoa and pro-tects them from premature acrosome reaction. Duringcumulus penetration, glycodelin-C in the matrix displacessperm-bound glycodelin isoforms and promotes the zona bind-ing capacity of the penetrated spermatozoa. Besides, theremoval of glycodelin-F may allow a more efficient zona-in-duced acrosome reaction to occur on the surface of the zonapellucida.
Acknowledgment—We thank Dr. S. J. Hong for contribution of theminiaturized hemizona binding assay.
REFERENCES1. Hizaki, H., Segi, E., Sugimoto, Y., Hirose, M., Saji, T., Ushikubi, F., Mat-
suoka, T., Noda, Y., Tanaka, T., Yoshida, N., Narumiya, S., and Ichikawa,A. (1999) Proc. Natl. Acad. Sci. U. S. A. 96, 10501–10506
2. Zhuo, L., Yoneda, M., Zhao, M., Yingsung, W., Yoshida, N., Kitagawa, Y.,Kawamura, K., Suzuki, T., and Kimata, K. (2001) J. Biol. Chem. 276,7693–7696
3. Fulop, C., Szanto, S., Mukhopadhyay, D., Bardos, T., Kamath, R. V., Rugg,M. S., Day, A. J., Salustri, A., Hascall, V. C., Glant, T. T., and Mikecz, K.(2003) Development 130, 2253–2261
4. Salustri, A., Garlanda, C., Hirsch, E., De, A.M.,Maccagno, A., Bottazzi, B.,Doni, A., Bastone, A., Mantovani, G., Beck, P. P., Salvatori, G., Mahoney,D. J., Day, A. J., Siracusa, G., Romani, L., and Mantovani, A. (2004) Devel-opment 131, 1577–1586
5. Yao, Y., Ho, P., and Yeung, W. S. (1999) Fertil. Steril. 72, 1079–10846. Hong, S. J., Chiu, P. C., Lee, K. F., Tse, J. M., Ho, P. C., and Yeung, W. S.
(2004) Hum. Reprod. 19, 1562–15697. Tanghe, S., Van, S. A., Nauwynck, H., Coryn,M., and de, K. A. (2002)Mol.
Reprod. Dev. 61, 414–4248. Carrell, D. T.,Middleton, R.G., Peterson, C.M., Jones, K. P., andUrry, R. L.
(1993) Arch. Androl. 31, 133–1379. Yanagimachi, R. (1994) in The Physiology of Mamalian Reproduction
(Knobil, E., and Neill, J. D.) 2nd Ed., pp. 189–318, Raven Press Ltd.,New York
10. Tesarik, J. (1989) J. Reprod. Fertil. 87, 193–19811. Seppala, M., Taylor, R. N., Koistinen, H., Koistinen, R., and Milgrom, E.
(2002) Endocr. Rev. 23, 401–43012. Chiu, P. C., Koistinen, R., Koistinen, H., Seppala,M., Lee, K. F., and Yeung,
W. S. (2003) Biol. Reprod. 69, 365–37213. Yeung,W. S., Lee, K. F., Koistinen, R., Koistinen, H., Seppala,M., Ho, P. C.,
and Chiu, P. C. (2006)Mol. Cell. Endocrinol. 250, 149–15614. Chiu, P. C., Chung, M. K., Tsang, H. Y., Koistinen, R., Koistinen, H., Sep-
pala, M., Lee, K. F., and Yeung, W. S. (2005) J. Biol. Chem. 280,25580–25589
15. Oehninger, S., Coddington, C. C., Hodgen, G. D., and Seppala, M. (1995)Fertil. Steril. 63, 377–383
16. Chiu, P. C., Ho, P. C., Ng, E. H., and Yeung,W. S. (2002)Mol. Reprod. Dev.61, 205–212
17. Yao, Y. Q., Chiu, C. N., Ip, S. M., Ho, P. C., and Yeung, W. S. (1998)Hum.Reprod. 13, 2541–2547
18. Chiu, P. C., Koistinen, R., Koistinen, H., Seppala,M., Lee, K. F., and Yeung,W. S. (2003) J. Biol. Chem. 278, 13570–13577
19. Julkunen, M., Wahlstrom, T., and Seppala, M. (1986) Am. J. Obstet.Gynecol. 154, 1076–1079
20. Laird, S. M., Hill, C. J., Warren, M. A., Tuckerman, E. M., and Li, T. C.(1995) Hum. Reprod. 10, 1346–1351
21. Brown, S. E., Mandelin, E., Oehninger, S., Toner, J. P., Seppala, M., andJones, H. W., Jr. (2000) Fertil. Steril. 74, 130–133
22. Zhu, J., Barratt, C. L., Lippes, J., Pacey, A. A., and Cooke, I. D. (1994) Fertil.Steril. 61, 1129–1135
23. Hong, S. J., Tse, J. Y., Ho, P. C., and Yeung, W. S. (2003) Fertil. Steril. 79,(Suppl. 1) 802–807
24. Tse, J. Y., Chiu, P. C., Lee, K. F., Seppala, M., Koistinen, H., Koistinen, R.,Yao, Y. Q., and Yeung, W. S. (2002)Mol. Hum. Reprod. 8, 142–148
25. World Health Organization (1999) Laboratory Manual for the Examina-tion of Human Semen and Semen-CervicalMucus Interaction, CambridgeUniversity Press, Cambridge, UK
26. Riittinen, L., Narvanen, O., Virtanen, I., and Seppala, M. (1991) J. Immu-nol. Methods 136, 85–90
27. Ng, E. H., Ajonuma, L. C., Lau, E. Y., Yeung, W. S., and Ho, P. C. (2000)Hum. Reprod. 15, 772–777
28. Yao, Y. Q., Yeung, W. S., and Ho, P. C. (1996) Hum. Reprod. 11,1516–1519
29. Lee, Y. L., Lee, K. F., Xu, J. S., He, Q. Y., Chiu, J. F., Lee, W. M., Luk, J. M.,and Yeung, W. S. (2004) J. Biol. Chem. 279, 12763–12768
30. Robertson, E. F., Dannelly, H. K., Malloy, P. J., and Reeves, H. C. (1987)Anal. Biochem. 167, 290–294
31. Chian, R. C., Ao, A., Clarke, H. J., Tulandi, T., and Tan, S. L. (1999) Fertil.Steril. 71, 61–66
32. Koistinen, H., Koistinen, R., Dell, A., Morris, H. R., Easton, R. L., Patankar,M. S., Oehninger, S., Clark, G. F., and Seppala, M. (1996) Mol. Hum.Reprod. 2, 759–765
Glycodelin and Spermatozoa
FEBRUARY 23, 2007 • VOLUME 282 • NUMBER 8 JOURNAL OF BIOLOGICAL CHEMISTRY 5387
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from

33. Munuce, M. J., Caille, A. M., Botti, G., and Berta, C. L. (2004) Andrologia36, 395–401
34. Giorgetti, C., Hans, E., Spach, J. L., Auquier, P., and Roulier, R. (1992)Hum. Reprod. 7, 1121–1125
35. Fakih, H., and Vijayakumar, R. (1990) Fertil. Steril. 53, 515–52036. Blumenfeld, Z., and Nahhas, F. (1989) Fertil. Steril. 51, 863–86837. Obasaju, M. F., Venier, W. C., Carson, R. S., and Ying, Y. (1993) Hum.
Reprod. 8, 1886–189138. Bastias, M. C., Kamijo, H., and Osteen, K. G. (1993) Hum. Reprod. 8,
1670–167739. Hossain, A. M., Rizk, B., Huff, C., Helvacioglu, A., and Thorneycroft, I. H.
(1996) Arch. Androl. 37, 7–1040. Sullivan, R., Duchesne, C., Fahmy, N., Morin, N., and Dionne, P. (1990)
Hum. Reprod. 5, 830–83441. Fetterolf, P. M., Jurisicova, A., Tyson, J. E., and Casper, R. F. (1994) Biol.
Reprod. 51, 184–19242. Westphal, L. M., el, D. I., Shimizu, S., Tadir, Y., and Berns, M. W. (1993)
Hum. Reprod. 8, 1083–108643. Pereda, J., and Coppo, M. (1985) Anat. Embryol. (Berl.) 173, 247–25244. Magier, S., van, d. V., Diedrich, K., and Krebs, D. (1990) Hum. Reprod. 5,
847–85245. Tajik, P., Niwa, K., and Murase, T. (1993) Theriogenology 40, 949–95846. Dubey, A. K.,Wang, H. A., Duffy, P., and Penzias, A. S. (1995) Fertil. Steril.
64, 787–79047. Zhang, L., Jiang, S., Wozniak, P. J., Yang, X., and Godke, R. A. (1995)Mol.
Reprod. Dev. 40, 338–34448. Suzuki, H., Jeong, B. S., and Yang, X. (2000) Biol. Reprod. 63, 723–72949. Fatehi, A. N., Zeinstra, E. C., Kooij, R. V., Colenbrander, B., and Bevers,
M. M. (2002) Theriogenology 57, 1347–135550. Fraser, L. R. (1984) J. Reprod. Fertil. 72, 373–38451. Virji, N., Phillips, D. M., and Dunbar, B. S. (1990) Mol. Reprod. Dev. 25,
339–34452. Rodgers, R. J., Irving-Rodgers, H. F., and Russell, D. L. (2003)Reproduction
126, 415–42453. Flower, D. L. (2000) Biochim. Biophys. Acta 1482, 327–33654. Wojnar, P., Lechner, M., and Redl, B. (2003) J. Biol. Chem. 278,
16209–16215
55. Zhuo, L., and Kimata, K. (2001) Cell Struct. Funct. 26, 189–19656. Miyagi, T.,Wada, T., Iwamatsu, A., Hata, K., Yoshikawa, Y., Tokuyama, S.,
and Sawada, M. (1999) J. Biol. Chem. 274, 5004–501157. Goi, G., Bairati, C., Massaccesi, L., Lovagnini, A., Lombardo, A., and Tet-
tamanti, G. (2000) FEBS Lett. 473, 89–9458. Cordero, O. J., Merino, A., Paez de la Cadena, M., Bugia, B., Nogueira, M.,
Vinuela, J. E., Martinez-Zorzano, V. S., de Carlos, A., and Rodriguez-Ber-rocal, F. J. (2001) Eur. J. Biochem. 268, 3321–3331
59. Mencarelli, S., Cavalieri, C., Magini, A., Tancini, B., Basso, L., Lemansky,P., Hasilik, A., Li, Y.T., Chigorno, V., Orlacchio, A., Emiliani, C., and Son-nino, S. (2005) FEBS Lett. 579, 5501–5506
60. Chiu, P. C., Tsang, H. Y., Koistinen, R., Koistinen, H., Seppala, M., Lee,K. F., and Yeung, W. S. (2004) Biol. Reprod. 70, 1710–1719
61. Shishido, Y., Sharma, K. D., Higashiyama, S., Klagsbrun, M., and Mekada,E. (1995) J. Biol. Chem. 270, 29578–29585
62. Chang, Z., Meyer, K., Rapraeger, A. C., and Friedl, A. (2000) FASEB J. 14,137–144
63. Chen, C., and Sathananthan, A. H. (1986) Arch. Androl. 16, 183–19764. Dirnfeld, M., Bider, D., Koifman, M., Calderon, I., and Abramovici, H.
(1999) Hum. Reprod. 14, 2562–256465. Morris, H. R., Dell, A., Easton, R. L., Panico, M., Koistinen, H., Koistinen,
R., Oehninger, S., Patankar, M. S., Seppala, M., and Clark, G. F. (1996)J. Biol. Chem. 271, 32159–32167
66. Chapman, N. R., and Barratt, C. L. (1996)Mol. Hum. Reprod. 2, 767–77467. Wassarman, P. M. (1999) Cell 96, 175–18368. Ozgur, K., Patankar, M. S., Oehninger, S., and Clark, G. F. (1998) Mol.
Hum. Reprod. 4, 318–32469. Stanton, P. G., Burgon, P. G., Hearn, M. T., and Robertson, D. M. (1996)
Mol. Cell. Endocrinol. 125, 133–14170. Ish-Shalom, E., Gargir, A., Andre, S., Borovsky, Z., Ochanuna, Z., Gabius,
H. J., Tykocinski, M. L., and Rachmilewitz, J. (2006) Glycobiology 16,173–183
71. Morales, P., Pizarro, E., Kong, M., Kerr, B., Ceric, F., and Vigil, P. (2000)Biol. Reprod. 63, 635–642
72. Asquith, K. L., Baleato, R. M., McLaughlin, E. A., Nixon, B., and Aitken,R. J. (2004) J. Cell Sci. 117, 3645–3657
Glycodelin and Spermatozoa
5388 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 8 • FEBRUARY 23, 2007
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from

Seppala, Pak-Chung Ho, Ernest H. Y. Ng, Kai-Fai Lee and William S. B. YeungPhilip C. N. Chiu, Man-Kin Chung, Riitta Koistinen, Hannu Koistinen, Markku
and -F and Stimulates Spermatozoa-Zona Pellucida BindingCumulus Oophorus-associated Glycodelin-C Displaces Sperm-bound Glycodelin-A
doi: 10.1074/jbc.M607482200 originally published online December 27, 20062007, 282:5378-5388.J. Biol. Chem.
10.1074/jbc.M607482200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/282/8/5378.full.html#ref-list-1
This article cites 70 references, 15 of which can be accessed free at
by guest on January 5, 2020http://w
ww
.jbc.org/D
ownloaded from





























