Embed Size (px)
Citation preview

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 1/60
Department of Natural Sciences
University of St. La Salle
Bacolod City

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 2/60
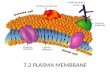
Plasma membrane is present in both eukaryoticand prokaryotic cells.
Despite their differing functions, all biological
membranes have a common general structure:
each is a very thin film of lipid and proteinmolecules, held together mainly by noncovalent
interactions.
Too thin to be seen with the light microscope, but
in EM, the membrane is 8.5-10 nm thick and shows
the trilaminar structure of the unit membrane. Specialized techniques, such as x-ray diffraction
and freeze-fracture electron microscopy, are
needed to reveal the details of its organization.
CELL MEMBRANE or PLASMALEMMA

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 3/60
Membrane
proteins are
solubilized with amild detergent.
The detergent
disrupts the lipid
bilayer and brings
the proteins into
solution as
protein-lipid-
detergent
complexes. Thephospholipids in
the membrane are
also solubilized by
the detergent.
STUDYING CELL
MEMBRANES

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 4/60
Freeze-fracture EM provides images of both the
hydrophobic interior of the cytosolic half of the bilayer (the
P face) and the hydrophobic interior of the external half of the bilayer (the E face). After the fracturing process, the
exposed fracture faces are shadowed with platinum and
carbon, the organic material is digested away, and the
resulting platinum replica is examined in EM.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 5/60
Models of Plasma Membrane Structure
Danielli and Davson model ² 2 layers of protein on
either side of a bimolecular leaflet of mixed lipid.
Fluid-mosaic model- phospholipid bilayer with
proteins floating on a ´sea of phospholipids.µ

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 6/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 7/60
There are 3 major classes of
membrane lipid molecules
phospholipids, cholesterol,
and glycolipids. The most abundant are the
phospholipids.
Lipid molecules in cell
membranes are amphipathic
or amphiphilic i.e., they havea hydrophilic or polar end
and a hydrophobic or
nonpolar end.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 8/60
Hydrophilic molecules dissolve readily in water because
they contain charged or uncharged polar groups that can
form either favorable electrostatic interactions or
hydrogen bonds with water molecules.
Hdrophobic molecules are insoluble in water because
almost all of their atoms are uncharged and nonpolar
and therefore cannot form energetically favorable
interactions with water molecules. If dispersed in water, they force the adjacent water
molecules to reorganize into icelike cages that surround
the hydrophobic molecule.
Because these cage structures are more ordered than
the surrounding water, their formation increases freeenergy.
This free energy cost is minimized if the hydrophobic
portions of amphipathic molecules cluster together so
that the smallest number of water molecules is affected.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 9/60
How hydrophilic and hydrophobic molecules interact differently
with water. (A) Because acetone is polar, it can form favorable
electrostatic interactions with water molecules, which are also
polar. Thus, acetone readily dissolves in water. (B) Insoluble
molecules such as 2-methyl propane is entirely hydrophobic &
cannot form favorable interactions with water, forcing adjacent water molecules to reorganize into icelike cage structures, which
increases the free energy. The symbol - indicates a partial
negative charge, and + indicates a partial positive charge. Polar
atoms are shown in color and nonpolar groups are shown in gray.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 10/60
Lipid composition can influence the activity of
particular membrane proteins. They can determine
the physical state of the membrane. Depending on their shape, lipid molecules can form
spherical micelles, with the tails inward; or they can
form bimolecular sheets, or bilayers, with the
hydrophobic tails sandwiched between the
hydrophilic head groups. Being cylindrical, phospholipid molecules
spontaneously form bilayers in aqueous
environments.
A lipid bilayer is responsible for the fluid
characteristic of the membrane. In this energetically most-favorable arrangement, the
hydrophilic heads face the water at each surface of
the bilayer, and the hydrophobic tails are shielded
from the water in the interior.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 11/60
(A) Wedge-shaped lipid molecules form micelles,
whereas cylinder-shaped phospholipid molecules
form bilayers. (B) A lipid micelle and a lipid bilayer
seen in cross section.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 12/60
PHOSPHOLIPIDS: have a polar head group and 2 hydrophobic
hydrocarbon tails. The tails are usually fatty acids, differing in length
from 14-24 C. Unsaturated FA have cis-double bands, producing a
kink in the tail. Differences in length and saturation influence thepacking of phospholipids , affecting the fluidity of the membrane.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 13/60
Types of phospholipids: phosphoglycerides have a glycerol
backbone; sphingolipids contain sphingosine instead of glycerol.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 14/60
The same forces that drive phospholipids to form
bilayers also provide a self-healing property.
A small tear in the bilayer creates a free edge withwater; because this is energetically unfavorable,
the lipids spontaneously rearrange to eliminate the
free edge. (In eukaryotic plasma membranes, larger
tears are repaired by the fusion of intracellularvesicles.)

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 15/60
The only way for a
bilayer to avoid
having free edges is
by closing in on
itself and form a
sealed
compartment.The closed structure
is stable because it
avoids the exposure
of the hydrophobic
hydrocarbon tails towater, which would
be energetically
unfavorable.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 16/60
Cholesterol reduces membrane fluidity at moderate temperatures by
reducing phospholipid movement,
but at low temperatures it hinderssolidification by disrupting the
regular packing of phospholipids.
Phospholipids very rarely migrate from
the monolayer on one side to that onthe other ("flip-flop´). In contrast, lipid
molecules readily exchange placeswith their neighbors within a monolayer
giving rise to a rapid lateral diffusion.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 17/60
The structure of cholesterol. Cholesterol is represented (A) by a
formula, (B) by a schematic drawing, and (C) as a space-filling model.
Eukaryotic plasma membranes contain up to one
molecule of for every
phospholipid molecule which enhance thepermeability-barrier properties of the lipid bilayer.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 18/60
� They orient themselves in the
bilayer with their hydroxyl
groups close to the polar head
groups of the phospholipidmolecules.
� In this position, their rigid,
platelike steroid rings interact
with and partly immobilize
those regions of the hydro-
carbon chains closest to the polar head groups.� By decreasing the mobility of the first few CH2 groups of the
hydrocarbon chains of the phospholipid molecules,
cholesterol makes the lipid bilayer less deformable in this region
and thereby decreases the permeability of the bilayer to small
water-soluble molecules.� Although cholesterol tends to make lipid bilayers less fluid, at the
high concentrations found in most eukaryotic plasma
membranes, it also prevents the hydrocarbon chains from coming
together and crystallizing. In this way, it inhibits possible phase
transitions.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 19/60
Spur cell anemia is caused
by too much cholesterol in
RBC membrane. It is
common in alcoholics with
cirrhosis of liver. Serum
cholesterol gets very high
and goes into the outer
leaflet of RBC membrane.
This expands the outerleaflet and produce spurs.
The membrane loses
flexibility and cells
become round and spur
covered. The cells get
trapped in spleen
capillaries and are
destroyed. Total RBC
count decreases resulting
to anemia.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 20/60
Almost all glycolipids are derived from glycerol, as are most
phospholipids; in animal cells they are produced from serine, e.g.
phospholipid sphingomyelin. 3 uncharged sugars: Gal =
galactose; Glc = glucose, GalNAc = N- acetylgalactosamine
GLYCOLIPIDS: (A)
Galactocerebroside,
a neutral glycolipidbecause the sugar
that forms its head
group is uncharged.
(B) A ganglioside
always contains one
or more negativelycharged sialic acid
residues (also called
N-acetylneuraminic
acid, or NANA)
shown in (C).

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 21/60
� Glycolipids are confined to the exposed apical surface,
where they help protect the membrane against the
harsh conditions such as low pH and degradativeenzymes.
� Charged glycolipids, such as gangliosides, alter the
electrical field across the membrane and the
concentrations of ions especially Ca+2 at the
membrane surface.� They function in cell-recognition processes, in which
membrane-bound carbohydrate-binding proteins
(lectins) bind to the sugar groups on both glycolipids
and glycoproteins in the process of cell-cell adhesion
� Mutant mice that are deficient in all of their complex
gangliosides show no obvious abnormalities, although
the males cannot transport testosterone normally in
the testes and are consequently sterile.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 22/60
Tay-Sach·s disease is a fatal
inherited condition that occurs
when harmful quantitiesof gangliosides accumulate in
the nerve cells of the brain
because of deficiency in -N-
acetylhexosaminidase A, an
enzyme which catalyzes thebiodegradation of
gangliosides, eventually
leading to the premature death
of those cells.
One of the symptoms of Tay-
Sach·s disease is the reddish
dot in the retina of the eye
which was first noticed byWarren Tay in the year 1881.
The disease is caused by the genetic mutation of HEXA
gene on chromosome 15 which provides instructions for
making the GM2 activator protein, a cofactor required for normal function of the -N-acetylhexosaminidase A.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 23/60
Different mixtures of lipids are found in the membranes of
cells of different types, as well as in the various
membranes of a single eucaryotic cell. Functions include:
1. Certain membrane proteins accumulate in lipid rafts
which are specialized areas in membranes where some
lipids (primarily sphingolipids and cholesterol) and
proteins (green) are concentrated. Rafts concentrate
them for transport in small vesicles or to enable the
proteins to function together, such as when they convert
extracellular signals into intracellular ones.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 24/60
Caveolae- special type of lipid rafts specialized for
endocytosis and compartmentalization of signal
transduction at the cell surface. One pathway is
through formation of vesicles that bud off from the PM
and travel to endosomes via microtubules. Another
pathway is through cyclic budding at the cell surface
to deliver small molecules and ions. The 3rd type form
tubules from the cell surface to the center.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 25/60
2. Some membrane-bound enzymes require specific
lipid head groups in order to function.
3. The head groups
of some lipids
form docking sites for specific
cytosolic proteins.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 26/60
4.Some extracellular signals that act through membrane
receptor proteins activate phospholipases that cleave
selected phospholipid molecules in the plasmamembrane, thereby generating fragments that act as
intracellular signaling molecules.
� For example, when animal cells undergo programmed
cell death, or apoptosis, phosphorylated inositol
phospholipids act as binding sites that recruit specificproteins from the cytosol to the membrane.
� Phosphatidylserine, which is normally confined to the
cytosolic monolayer of the plasma membrane lipid
bilayer, rapidly translocates to the extracellular
monolayer� The phosphatidylserine exposed on the cell surface
serves as a signal to induce neighboring cells, such as
macrophages, to phagocytose and digest the dead cell.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 27/60
Some functions of membrane phospholipids in cell signaling. (A)
Extracellular signals can activate PI 3-kinase, which phosphorylatesinositol phospholipids in the plasma membrane. Signaling molecules
then bind to these phosphorylated lipids & are thus recruited to the
membrane, where they can interact and help relay the signal into the
cell. (B)Other extracellular signals activate phospholipases that cleave
phospholipids. The lipid fragments then act as signaling molecules to
relay the signal into the cell.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 28/60
Freeze-fracturing splits the bilayer, showing proteins to be entirely within
the lipid bilayer (integral/ transmembrane/ intrinsic), or projecting from
the outer surface to varying degrees (peripheral/ extrinsic).
PROTEINS

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 29/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 30/60
1.Transmembrane, integral or intrinsic proteins extend
through the lipid bilayer, with part of their mass on either
side. are amphiphatic molecules possessing one or more
hydrophobic regions that exhibit an affinity for the
hydrophobic interior of the lipid bilayer; hence these
molecules are difficult to remove from membranes
They are notoriously difficult to crystallize; few have beenstudied in their entirety by x-ray crystallography.
DNA cloning and sequencing techniques, have revealed
their amino acid sequences, and it is often possible to
predict from an analysis of the protein's sequence which
parts of the polypeptide chain extend across the lipidbilayer.
Proteins are capable of lateral movement within the
bilayer; apical to basolateral movement is hindered by cell
junctions, or their attachment to cytoskeletal elements.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 31/60
2.Peripheral or extrinsic proteins function on
only one side of the lipid bilayer; are often
associated exclusively with either the lipidmonolayer or a protein domain on that side.
Some of these are anchored to the cytosolic
surface by an amphipathic helix that
partitions into the cytosolic monolayer of the
lipid bilayer through the hydrophobic face of
the helix.
� For example, some of the proteins involved in
intracellular signaling are bound to the
cytosolic half of the plasma membrane by
one or more covalently attached lipid groups.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 32/60
Peripheral proteins of RBC plasma membrane are
spectrin and ankyrin, and a protein called band.
These proteins are bound to the inner surface of PMand form as a meshwork that support the PM and
help maintain the shape of RBC. Gene mutations for
these proteins result to RBC shape anomalies.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 33/60
� How a membrane protein associates with the lipid
bilayer reflects the function of the protein.
� Only transmembrane proteins (e.g. cell-surfacereceptors can function on both sides of the bilayer
or transport molecules across it.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 34/60
Various ways in which membrane proteins associate with the lipid bilayer.
Some of these "single-pass" and "multipass" transmembrane proteins havea covalently attached fatty acid chain inserted in the cytosolic lipid
monolayer (1). Most are thought to extend across the bilayer as (1) a single
a helix, (2) as multiple a helices, (3) as a rolled-up sheet or a barrel. Others(5) are attached to the bilayer by a covalently attached lipid chain either a
fatty acid chain or a prenyl group in the cytosolic monolayer, (6) via an
oligosaccharide linker, or to phosphatidylinositol in the noncytosolicmonolayer. (7, 8) Many proteins are attached to the membrane only by
noncovalent interactions with other membrane proteins.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 35/60
The cell coat is made up of the oligosaccharide side chains
of glycolipids and integral membrane glycoproteins and the
polysaccharide chains on integral membrane
proteoglycans. In addition, adsorbed glycoproteins and
adsorbed proteoglycans contribute to the glycocalyx in
many cells. Note that all of the carbohydrate is on the
noncytosolic surface of the membrane.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 36/60
It plays a role in the selective uptake of
substances, recognition of self, a cellular ID
system.
It confers an electrostatic charge on the cell
surface, important in cell-to-cell adhesion.
� Disturbances in the cell coat and surface charge
maybe responsible for the failure of cancer cellsto adhere to one another.
� Histogenesis is achieved by modulating cell-
adhesion molecules (CAMs) which are integral
protein-carbohydrate (e.g., sialic acid)complexes.
� Antigens are located within the cell coat.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 37/60
Human ABO blood-
group antigens.
These antigens are
oligosaccharidechains covalently
attached to
glycolipids or glyco-
proteins in the
plasma membrane.
The terminal oligo-saccharide sugars
distinguish the 3
antigens. The
presence or absence
of the glycosyl-
transferases that addgalactose (Gal) or N-
acetylgalactosamine
(GalNAc) toO antigen
determine a person·s
blood type.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 38/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 39/60
TRANSPORT MECHANISMS

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 40/60
1. Passive transport down an electrochemical gradient
occurs spontaneously, either by simple diffusion
through the lipid bilayer or by facilitated diffusion
through channels and passive carriers.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 41/60
A model of how a conformational change in a carrier protein
could mediate the PASSIVE TRANSPORT of a solute. The carrier
protein can exist in 2 conformational states: in state A, the bindingsites for solute are exposed on the outside of the lipid bilayer; in
state B, the same sites are exposed on the other side of the
bilayer. The transition between the two states can occur randomly,
completely reversible and does not depend on whether the solute
binding site is occupied.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 42/60
2. Active transport requires an input of metabolic energy and
is always mediated by carriers that harvest metabolic
energy to pump the solute against its electrochemical
gradient.
� An electrochemical gradient combines the membrane
potential and the concentration gradient, which can work
additively to increase the driving force on an ion across
the membrane or can work against each other.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 43/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 44/60
The sodium-potassium pump: a specific case of active transport

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 45/60
http://highered.mcgraw-
hill.com/olc/dl/120068/bio03.swf

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 46/60
transports the solute in (or out) and the co-
transported solute the opposite direction.
http://highered.mcgraw-
hill.com/olc/
dl/120068/bi
o04.swf

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 47/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 48/60
http://highered.mcgraw-
hill.com/olc/dl/120068/bio05.swf

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 49/60
Co-transport
� Glucose is
pumped into the
cell through the
apical domain of
the membrane bya Na+ -poweredglucose symport.
� Glucose passes
out of the cell by
passive transport
mediated by adifferent glucosecarrier protein in
the basal and
lateral membrane
domains.
Transcellular transport of glucose across an intestinal epithelial cell

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 50/60
4.Signal-Reception- messenger molecules are
membrane-bound or may pass through
membrane channels to mediatecommunication among cells.
� G-proteins- activated by the binding of a 1st
messenger molecule to a membrane receptor.
� The GTP-G protein exchanges act onmembrane-bound enzyme effectors, which in
turn activate a 2nd messenger.
� This triggers a cascade of molecular
reactions that lead to changes in cell behavior(e.g., LH activates G5 adenylyl cyclase
increased synthesis of estrogen and
progesterone).

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 51/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 52/60
The gating of ion channels. Different kinds of stimuli open ion
channels. Mechanically gated channels often have cytoplasmic
extensions that link the channel to the cytoskeleton. Ion Channel Diseases:
Myasthenia Gravis ² Muscle weakness due to autoantibodies
against the acetylcholine receptor
Cystic Fibrosis ² Defect in the Cl- channel CFTR ² leads to
excessive phlegm and static infections

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 53/60
when transport is out of the cell.

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 54/60
http://highered
.mcgraw-
hill.com/olc/dl/120068/bio02.swf

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 55/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 56/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 57/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 58/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 59/60

8/6/2019 Cytology Plasma Membrane
http://slidepdf.com/reader/full/cytology-plasma-membrane 60/60




![Plasma Membrane [7.2] Goals: Understand the concept of homeostasis in relation to the plasma membrane Demonstrate and understand how the plasma membrane](https://img.pdfslide.net/doc/110x75/5697c01d1a28abf838cd0a9a/plasma-membrane-72-goals-understand-the-concept-of-homeostasis-in-relation.jpg)























