Embed Size (px)
Citation preview

Eur. J. Biochem. 29, 348-352 (1972)
Deoxyribonucleoside Triphosphate Pools of Normal and Transformed Baby -Hamster-Kidney Cells
K. Gunnar BJURSELL, Peter A. REICHARD, and K. Lambert S ~ o o a Medicinska Nobelinstitutet, Biokemiska Avdelningen, Karolinska Institutet, Stockholm
(Received March 30/June 16,1972)
Pools of the four deoxyribonucleoside triphosphates were determined during growth in tissue culture of baby hamster kidney normal cells and cells transformed either by wild-type polyoma virus or a temperature-sensitive mutant of polyoma (ts-3). The cells are transformed by ts-3 a t 31 "C but lose most of their transformed character at 39 "C. The three cell lines did not show any major differences with respect to pool sizes at 39 "C: during rapid growth, dCTP formed the largest and dGTP the smallest pool, while the dATP and dTTP pools were of intermediate size. On cessation of growth all pools decrease in size ; the decrease was very pronounced for dCTP but slight for dATP.
The results a t 31 "C were similar to those a t 39 "C with one exception: the two transformed lines contained about twice as much dTTP as the normal cell line under comparable growth conditions. Our results suggest that after transformation a change occurs in the control of one or several of the enzymes leading to the synthesis of dTTP. This change is manifested only a t the lower temperature.
Transformation of cells with oncogenic viruses results in alterations of many properties of the cell population [l, 21. In particular, the absence of contact inhibition observed in most transformed cell lines has attracted considerable attention [3,4]. The loss of growth inhibition has been related to a change in some control mechanism of DNA synthesis. Such speculations are supported by findings that the activity of several enzymes connected with DNA synthesis is increased after infection with oncogenic viruses [5,6].
Many of the enzymes involved in the synthesis of DNA appear to be controlled by deoxyribonucleoside triphosphates [7] and it is therefore of interest to determine cellular pools of these nucleotides in relation to DNA synthesis. I n previous experiments with secondary mouse embryo cells, we observed that pool levels showed considerable fluctuations which appeared to be correlated with the rate of DNA synthesis. This was found both in normal cells and in cells productively infected with poly- oma virus [S, 91.
Baby hamster kidney cells can be transformed with polyoma virus and such cells have altered growth properties[3]. It has been shown that the viral DNA is integrated into the genome of such transformed cells [lo, 111. I n the present study, we compared deoxyribonucleoside triphosphate pools
of normal and polyoma-transformed baby hamster kidney cells under different stages of growth. For this purpose, we also used a line of cells which had been transformed with a temperature-sensitive mutant of polyoma (ts-3). At the permissive temper- ature (31 "C), these cells exhibit the properties of tarnsformed cells while a t the nonpermissive temper- ature (39 "C) the cells have essentially a "normal" appearance [12,13]. This transformed cell line thus enabled us to study both states in the same cell line.
MATERIALS AND METHODS
Cell Cultures The following hamster cell lines were used:
baby hamster kidney C1 13, which is a clone of baby hamster kidney cells [3], C12A, a wild-type polyoma-transformed line of strain C1 13, and C1 7C, obtained from strain C1 13 by transformation with ts-3, a temperature-sensitive mutant of polyoma [12]. All cell lines were kindly supplied by Dr Walter Eckhart (The Salk Institute, La Jolla, U.S.A.). The cells were cultured in Dulbecco's modified Eagle medium (Grand Island Biological Company, Berkeley, California) supplemented with 5 fetal calf serum (Biocult, Glasgow, Scotland). Cultures were started by seeding 3x105cells per 100mm plastic petri
Vol.29, No.2,1972 K. G. BJURSELL, P. A. REICHARD, and K. L. SKOOO 349
dishes (Falcon), and incubated in an atmosphere of 5 O / , CO, in air a t the temperatures indicated.
Prewarmed medium was changed every second day when the cells were grown a t 37 "C and every third day when the cells were grown a t 31°C and 39 "C.
The cells were free from mycoplasma contamina- tion as monitored by cultivation in agar [14] and by absence of cytoplasmic incorporation of rH]thymi- dine as judged by autoradiography.
DNA Synthesis and Pool Measurements The methods for measuring the amounts of
DNA and deoxyribonucleotides present in the cul- tures and the percentage of cells synthesizing DNA, as judged by autoradiography after 60-min pulses with [SH]thymidine, were described previously [8,15,16].
RESULTS Pools in Normal Cells
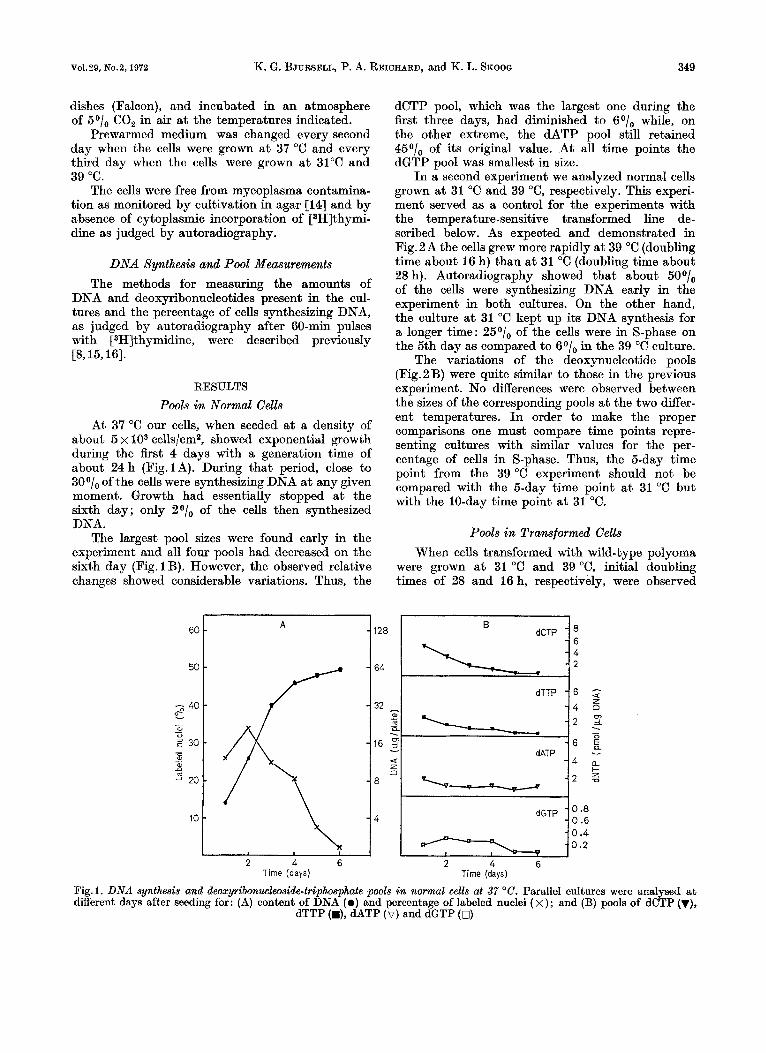
At 37 "C our cells, when seeded a t a density of about 5 x lo3 cells/cm2, showed exponential growth during the first 4 days with a generation time of about 24 h (Fig. 1A). During that period, close to 3001, of the cells were synthesizing DNA a t any given moment. Growth had essentially stopped a t the sixth day; only 2O/, of the cells then synthesized DNA.
The largest pool sizes were found early in the experiment and all four pools had decreased on the sixth day (Fig. 1 B). However, the observed relative changes showed considerable variations. Thus, the
dCTP pool, which was the largest one during the first three days, had diminished to 60/, while, on the other extreme, the dATP pool still retained 45O/, of its original value. At all time points the dGTP pool was smallest in size.
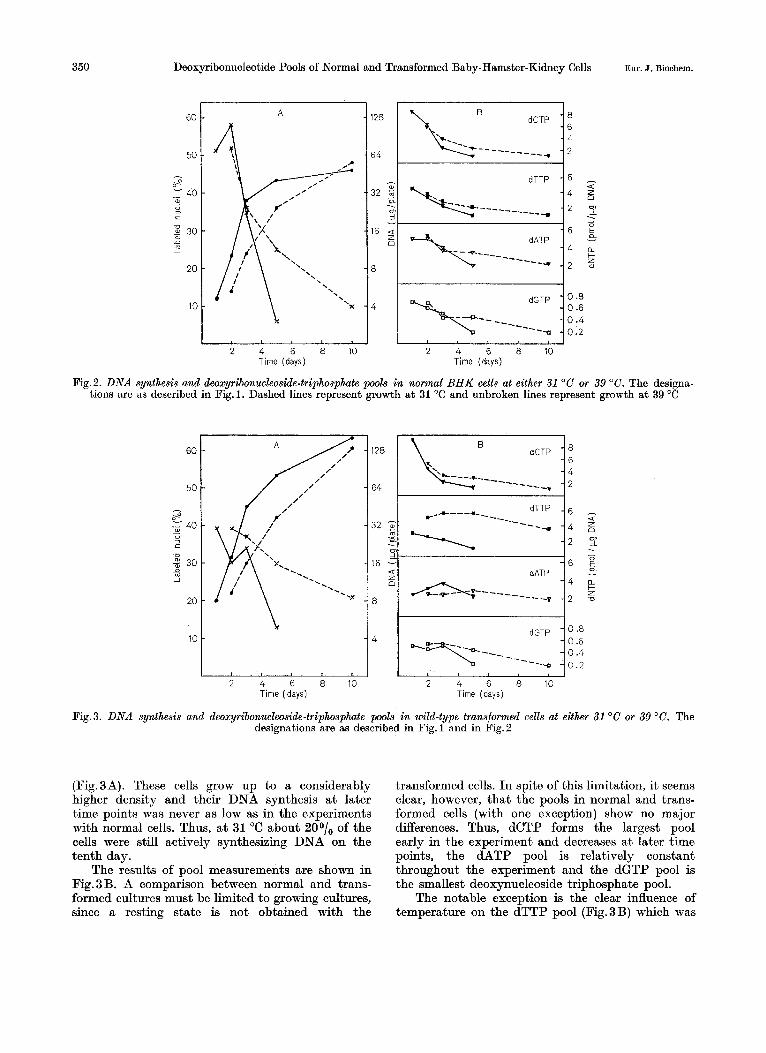
In a second experiment we analyzed normal cells grown a t 31 "C and 39 "C, respectively. This experi- ment served as a control for the experiments with the temperature-sensitive transformed line de- scribed below. As expected and demonstrated in Fig. 2A the cells grew more rapidly at 39 "C (doubling time about 16 h) than a t 31 "C (doubling time about 28 h). Autoradiography showed that about 50°/, of the cells were synthesizing DNA early in the experiment in both cultures. On the other hand, the culture a t 31 "C kept up its DNA synthesis for a longer time: 25O/, of the cells were in S-phase on the 5th day as compared to 6O/, in the 39 "C culture.
The variations of the deoxynucleotide pools (Fig. 2 B) were quite similar to those in the previous experiment. No differences were observed between the sizes of the corresponding pools at the two differ- ent temperatures. I n order to make the proper comparisons one must compare time points repre- senting cultures with similar values for the per- centage of cells in S-phase. Thus, the 5-day time point from the 39 "C experiment should not be compared with the 5-day time point a t 31 "C but with the 10-day time point a t 31 "C.
Pools in Transformed Cells When cells transformed with wild-type polyoma
were grown a t 31 "C and 39 "C, initial doubling times of 28 and 16 h, respectively, were observed
Fig. 1. DNA sunthesis and deoxvribonucleoside-trcphosphate pools in normal cells at 37 "C. ParalleI cultures were analvsed at different daysafter seeding for: (A) content of DNA ( 0 ) ind percentage of labeled nuclei ( x); and (B) pools of dC-?P (v),
dTTP (a), dATP (v) and dGTP (0)
350 Deoxyribonucleot,ide Pools of Normal and Transformed Baby-Hamster-Kidney Cells Eur. J. Biochem.
60
50
- ,--" - 40 .- b,
" 3 c
D
-
t 30 1) m 1
20
10
A
2 4 6 8 1 0 Time (days)
dGTP
0.4 -- 4
I I
Time (days) 2 4 6 8 1 0
Fig.2. DNA synthesis and deoxyribonucleoside-triphosphate pools in normal BHK cells at either 31 "C or 39 "C. The designa- tions are as described in Fig.l. Dashed lines represent growth a t 31 "C and unbroken lines represent growth a t 39 "C
8 1
4
---------p i 2 &--
Time (days)
Fig.3. DNA synthesis and deoxyribonucleoside-triphosphate pools in wild-type transformed cells at either 31 "C or 39 "C. The designations are as described in Fig. 1 and in Fig. 2
(Fig.3A). These cells grow up to a considerably higher density and their DNA synthesis a t later time points was never as low as in the experiments with normal cells. Thus, a t 31 "C about 20°/, of the cells were still actively synthesizing DNA on the tenth day.
The results of pool measurements are shown in Fig.3B. A comparison between normal and trans- formed cultures must be limited to growing cultures, since a resting state is not obtained with the
transformed cells. In spite of this limitation, it seems clear, however, that the pools in normal and trans- formed cells (with one exception) show no major differences. Thus, dCTP forms the largest pool early in the experiment and decreases a t later time points, the dATP pool is relatively constant throughout the experiment and the dGTP pool is the smallest deoxynucleoside triphosphate pool.
The notable exception is the clear influence of temperature on the dTTP pool (Fig.3B) which was
Vo1.29, No.2,1972 K. G. BJURSELL, P. A. REIOHARD, and I<. L. SKOOG 351
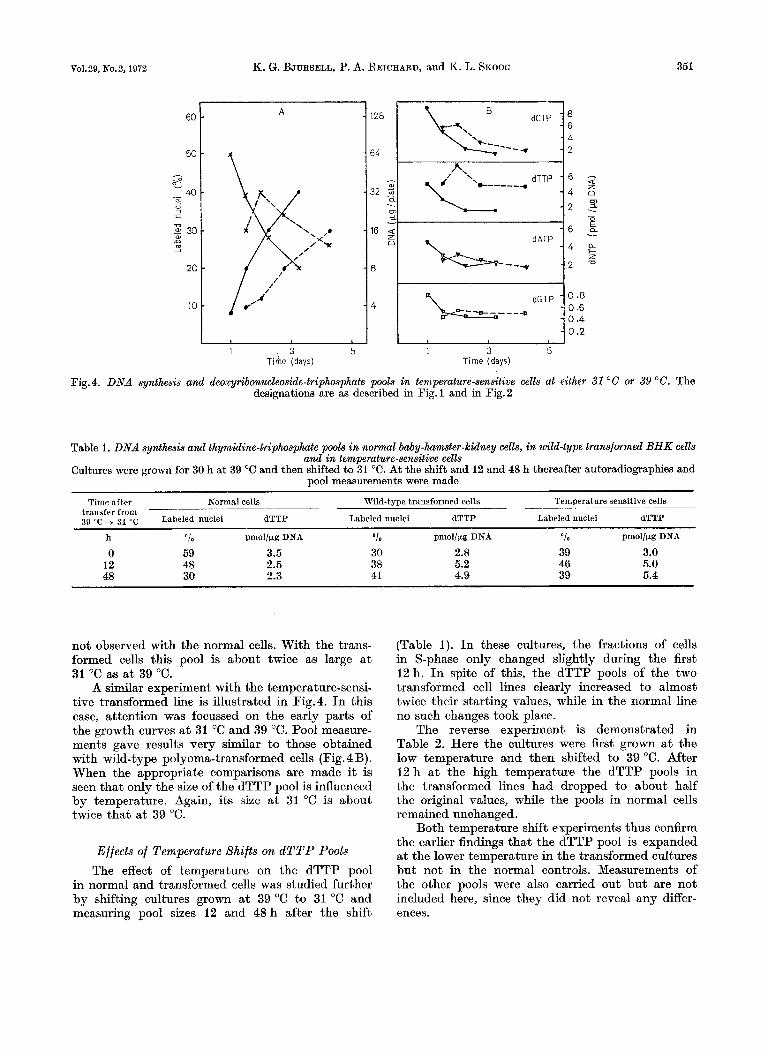
Fig. 4. DNA synthesis and deoxyribonucleoside-triphosphate pools in temperature-sensitive cells at either 31 "C or 39 "G. The designations are as described in Fig. 1 and in Fig. 2
Table 1. DNA synthesis and thymidine-triphsphate pools in normal baby-hamster-kidney cells, in wild-type transformed BHK cells and in temperature-sensitive cells
Cultures were grown for 30 h at 39 O C and then shifted to 31 "C. At the shift and 12 and 48 h thereafter autoradiographies and pool measurements were made
Time after Normal cells Wild-type transformed cells Temperature sensitive cells
~ J ~ O C ' . I, O P Labeled nuclei dTTP Labeled nuclei dTTP Labeled nuclei dTTP transfer from -
h "0 pmol/Wg DNA 'la pmol/wg DNA "0 pmol/Wg DNA
0 59 3.5 30 2.8 39 3.0 12 48 2.5 38 5.2 46 5.0 48 30 2.3 41 4.9 39 5.4
not observed with the normal cells. With the trans- formed cells this pool is about twice as large a t 31 "C as a t 39 "C.
A similar experiment with the temperature-sensi- tive transformed line is illustrated in Fig.4. In this case, attention was focussed on the early parts of the growth curves a t 31 "C and 39 "C. Pool measure- ments gave results very similar to those obtained with wild-type polyoma-transformed cells (Fig. 4B). When the appropriate comparisons are made it is seen that only the size of the dTTP pool is influenced by temperature. Again, its size a t 31 "C is about twice that a t 39 "C.
Effects of Temperature Shifts on dTTP Pools The effect of temperature on the dTTP pool
in normal and transformed cells was studied further by shifting cultures grown a t 39 "C to 31 "C and measuring pool sizes 12 and 48 h after the shift
(Table 1). I n these cultures, the fractions of cells in S-phase only changed slightly during the first 12 h. I n spite of this, the dTTP pools of the two transformed cell lines clearly increased to almost twice their starting values, while in the normal line no such changes took place.
The reverse experiment is demonstrated in Table 2. Here the cultures were first grown a t the low temperature and then shifted to 39 "C. After 12 h a t the high temperature the dTTP pools in the transformed lines had dropped to about half the original values, while the pools in normal cells remained unchanged.
Both temperature shift experiments thus confirm the earlier findings that the dTTP pool is expanded a t the lower temperature in the transformed cultures but not in the normal controls. Measurements of the other pools were also carried out but are not included here, since they did not reveal any differ- ences.

352 BJURSELL et al. : Deoxyribonucleotide Pools of Normal and Transformed Baby-Hamster-Kidney Cells Eur. J. Biochem.
Table 2. DNA synthesis and thymidine-triphsphate pools in normal cells, in wild-type transformed cells and in temperature- sensitive cells
Cultures were grown for 30 h at 31 "C and then shifted to 39 "C. At the shift and 12 and 48 h thereafter autoradiographies and pool measurements were made
Time after Normal cells Wild-type transformed cells Temperature-sensitive cells transfer from RI o r ! tn RO O P Labeled nuclei dTTP Labeled nuclei dTTP Labeled nuclei dTTP
h O/O pmol/wg DNA "la pmol/pg DNA "k pmol/pg DNA
0 52 3.8 39 5.0 32 5.5 12 59 3.5 36 3.0 29 2.0 48 25 2.7 30 2.0 31 1.5
DISCUSSION The principal question asked in our experiments
was whether the pattern of deoxyribonucleoside triphosphate pools differed between normal and transformed cell cultures. The availability of a cell line which had been transformed by a temperature- sensitive polyoma mutant and which was reported to retain most of its transformed characters only a t the permissive temperature appeared to be of particular use.
We did not find clear differences in the behaviour of any of the four pools when normal or trans- formed cultures were grown a t 39 "C. On the other hand, a t 31 "C we consistently observed a difference for one of the deoxynucleotides. At this temper- ature, the dTTP pool of the transformed cells was almost twice the size of that in normal cells. The results were quite clear-cut in spite of the difficulties in interpretation due to changes in pool sizes during the growth of the different cultures. I n particular, the temperature shift experiments reported in Tables 1 and 2 appear convincing. They also show that the changes in size of the dTTP pools in the transformed cells occurred within less than one generation time.
The underlying mechanism for the observed phenomenon is by no means clear. The increase in pool size at 31 "C may reflect a change in the relative activities of the enzymes which contribute to the synthesis of dTTP. In this connection an increased activity of the thymidine kinase comes to mind. Alternatively it may be caused by a change in the control mechanism of one or several of the enzymes, a change which is manifested only a t the lower temperature.
It should be stressed that a comparison of pool sizes between different cell cultures must consider the very large variations of the pools during the growth of the culture. Earlier experiments with secondary mouse embryo cells had already clearly demonstrated the flexibility of all four pools and this
is again indicated in our present experiments with baby hamster kidney cells. These studies, further- more, stress the point that the size of the dCTP pool is most directly related to the rate of DNA synthesis in the culture, while the size of the dATP pool is much more rigidly maintained a t the same level during all stages of growth. Thus, the relative size of the dATP pool, in relation to the other three pools, is largest in resting cultures. Similar results have been observed with other cell lines and will be reported elsewhere.
This work was supported by grants from the Swedish Cancer Society.
1. 2. 3. 4. 5.
6.
I . 8.
9.
10. 11.
12.
13.
14. 15. 16.
REFERENCES MacPherson, I. (1970) Advan. Cancer Res. 13, 169. Eckhart, W. (1968) Physiol. Rev. 48, 513. MacPherson, I. & Stoker, M. (1962) Virology, 16, 147. Dulbecco, R. (1969) Science (Washington) 166, 962. Dulbecco, R., Hartwell, L. H. & Vogt, M. (1965) Proc.
Nut. Acad. Sci. U . 8. A . 53, 403. Weil, R., Petursson, G., Kara, J. & Diggelman, H. (1967).
The Molecular Biology of Vireses, p. 517, Academic Press, New York.
Koerner, J. F. (1970) Ann. Rev. Biochem. 39, 723. Nordenskjold, B. A., Skoog, L., Brown, N. & Reichard,
Skoog, L. & Nordenskjold, B. (1970) Eur. J. Biochem.
Benjamin, T. L. (1966) J . MoZ. Biol. 16, 359. WestDhal. H. & Dulbecco. R. 119681 Proc. Nut. A d .
P. (1970) J . Biol. Chem. 245, 5360.
19, 81.
Sc;f U:S. A . 59, 1158. Dulbecco, R. & Eckhart, W. (1970) Proc. Nut. Acad.
. I
Sci. U.'S. A . 67, 1715. .
Nut. A d . Sci. U. S. A . 68, 283. Eckhart, W., Dulbecco, R. & Burger, M. (1971) Proc.
Haytlic, L. (1965) Texas Rep. Biol. Med. 23, 285. Lindberg, U. & Skoog, L. (1970) Anal. Biochem. 34, 152. Skoog, L. (1970) Eur. J . Biochem 17, 202.
K. G. Bjursell, P. A. Reichard, and K. L. Skoog Medicinska Nobelinstitutet, Biokemiska Avdelningen Karolinska Institutet Solnavagen 1, S-10401 Stockholm 60, Sweden



![Gemcitabine and Doxorubicin Combination Enhance the Cytotoxic … · 2017-10-26 · ther phosphorylate dFdCDP to its another active triphosphate (dFdCTP) forms [6]. Deoxyribonucleoside](https://img.pdfslide.net/doc/110x75/5e3ea43e2e3439223d37c393/gemcitabine-and-doxorubicin-combination-enhance-the-cytotoxic-2017-10-26-ther.jpg)

























