Embed Size (px)
Citation preview

Free Radical Biology & Medicine, Vol. 36, No. 4, pp. 507 –516, 2004Copyright D 2004 Elsevier Inc.
Printed in the USA. All rights reserved0891-5849/$-see front matter
doi:10.1016/j.freeradbiomed.2003.11.028
Original Contribution
DEPLETION OF DEOXYRIBONUCLEOSIDE TRIPHOSPHATE
POOLS IN TUMOR CELLS BY NITRIC OXIDE
BEATRICE ROY, OLIVIER GUITTET, CLAIRE BEUNEU, GENEVIEVE LEMAIRE, and MICHEL LEPOIVRE
UMR CNRS 8619, IBBMC, Universite de Paris XI, F-91405 Orsay Cedex, France
(Received 28 July 2003; Revised 10 November 2003; Accepted 26 November 2003)
Ad
Bat. 43
+(33-1
Abstract—Nitric oxide displays pro- and anti-tumor activities, prompting further studies to better understand its precise
role. Nitric oxide inhibits ribonucleotide reductase (RnR), the limiting enzyme for de novo dNTP synthesis. We report here
the first detailed analysis of dNTP variations induced in tumor cells by NO. NO prodrugs induced a depletion in dNTP
pools and an activation of the pyrimidine salvage pathway, as did hydroxyurea, the prototypic RnR inhibitor. In the
presence of dipyridamole, which blocked salvaged dNTP synthesis, depletion of dNTP pools was also observed in tumor
cells cocultured with macrophages expressing the high-output iNOS activity. This effect was rapid, reversible, blocked by
NO scavengers, and cGMP independent. It was quantitatively correlated to iNOS activity. In the absence of dipyridamole,
NO still induced a decrease in dATP concentration in tumor cells cocultured with macrophages, whereas surprisingly,
concentrations of dCTP and dTTP expanded considerably, resulting in a strong imbalance in dNTP pools. NO prodrugs
did not cause such an increase in pyrimidine dNTP, suggesting that pyrimidine nucleosides were released by NO-injured
macrophages. Altered dNTP levels have been reported to promote mutagenesis and apoptosis. It is suggested that
abnormal changes in dNTP pools in tumors might contribute to NO-dependent toxicity. D 2004 Elsevier Inc. All rights
reserved.
Keywords—Nitric oxide, Deoxyribonucleotide, Macrophage, Ribonucleotide reductase, Leukemia, Free radicals
INTRODUCTION
Ribonucleotide reductase (RnR) is crucial for DNA
replication and repair, because it is the only enzyme that
catalyzes the reduction of ribonucleotides into deoxyri-
bonucleotides in the de novo pathway for dNTP synthe-
sis [1]. A salvage pathway that phosphorylates deoxy-
nucleosides imported from the extracellular milieu
provides an alternative route for dNTP production, but
cannot compensate for RnR deficiency [2]. RnR inacti-
vation by a variety of pharmaceutical inhibitors such as
hydroxyurea and gemcitabine leads to dNTP depletion
and has been successfully applied to anticancer chemo-
therapy for many years [3,4].
dress correspondence to: Michel Lepoivre, UMR CNRS 8619,
0, University of Paris XI, F-91405 Orsay Cedex, France. Fax:
)-69-85-37-15; E-mail: [email protected].
507
Eukaryotic RnR is an a2h2 complex. The small h2
homodimer, called R2, is a storage form for a stable
free radical essential for activity and is supposed to be
transferred to the catalytic site on a2 (R1) to initiate a
radical-driven reduction of the substrate. The free
radical is located on a tyrosyl residue buried in the
core of protein R2 and close to a diferric iron center.
Previous studies have shown that the reactive radical
NO inactivates RnR [5,6]. Different inhibitory mecha-
nisms have been proposed, but the most probable is a
loss of the R2-centered free radical consecutive to a
radical–radical reaction between NO and the tyrosyl
free radical [7–9]. This event has been detected by
EPR spectroscopy in vitro and in cell culture. We
proposed that cessation of tumor cell proliferation
induced by the high-output iNOS activity arises in
part from RnR inhibition [5]. This would cause dNTP
depletion and hence, inhibition of DNA synthesis.
However, only one report from our laboratory docu-
mented the effects of NO on dNTP pools [10].

B. Roy et al.508
Moreover, several studies have demonstrated the exis-
tence of NO-dependent cytostatic mechanisms indepen-
dent of RnR inhibition [11–13]. Another aspect of the
question concerns the amounts of NO necessary to
exert antiproliferative effects. In fact, the role of NO in
tumor growth is a matter of debate (reviewed in
[14,15]). In several studies, NO has been shown to
promote carcinogenesis and tumor progression, in
particular via deleterious reactions on DNA [16] and
stimulation of angiogenesis [14]. Other reports have
established that sustained overproduction of NO, typ-
ically by cells expressing the iNOS isoform, can cause
cytostasis and apoptosis of tumor cells. Very different
concentrations of NO in the tumor microenvironment
might explain this discrepancy as well as differential
susceptibilities of tumor cells to NO-induced damage.
Clearly, more quantitative data are necessary to better
understand the paradoxical actions of NO on tumor
growth.
In the present report, we show that dNTP pools are
drastically modified by NO prodrugs or macrophage
iNOS activity. The extent of dNTP depletion is propor-
tional to NO output. In the presence of macrophages,
there is a profound alteration in the dNTP concentration
equilibrium. Imbalance in dNTP levels may help ex-
plain NO-dependent mutagenic effects.
EXPERIMENTAL PROCEDURES
Biochemicals and cytokines
LPS from Salmonella enteritidis, human oxyhemo-
globin, human recombinant IL-1h, hydroxyurea, and
dipyridamole were obtained from Sigma. Carboxy-
PTIO, S-ethylisothiourea (SEITU), and 1H-[1,2,4]oxa-
diazolo[4,3-a]quinoxalin-1-one (ODQ) were purchased
from Calbiochem. [5-3H]Deoxycytidine (sp act 18.4
Ci/mmol) was from Amersham Pharmacia Biotech.
(Z)-1-[N-(3-Aminopropyl)-N-(4-(3-aminopropylammo-
nio)butyl)amino]diazen-1-ium-1,2-diolate (SPER-NO)
and (Z)-1-[N-(2-aminoethyl)-N-(2-ammonioethyl)ami-
no]diazen-1-ium-1,2-diolate (DETA-NO) were obtained
from Cayman Chemicals. Human recombinant TNF-a
and IFN-g were generous gifts from Rhone-Poulenc
Rorer and Roussel-Uclaf (France), respectively. Mu-
rine recombinant IFN-g was provided by Dr. Adolf
(Ernst–Boehringer Institut fur Arzneimittel Forschung,
Austria).
Macrophages and cell lines
Murine peritoneal macrophages elicited by i.p.
injection of thioglycolate broth were obtained and
purified by adherence as previously described [17].
The L-1210 murine lymphoblastic leukemia and the
human T4-lymphoblastoid CEM-SS cell lines were
maintained in RPMI 1640 medium (Invitrogen) sup-
plemented with 25 mM Hepes, pH 7.4, L-glutamine,
antibiotics, and 5% heat-inactivated fetal calf serum
(FCS). The human colon carcinoma cell line DLD-1
was maintained in Dulbecco’s modified Eagle’s medi-
um with added antibiotics and 10% FCS [18].
Coculture of macrophages with tumor cells for dNTP
measurements
In experiments designed to measure dNTP concen-
tration in tumor cells, macrophages were seeded into
six-well culture plates (Nunc) at a nonsaturating density
(typically 2 � 106 cells/well) in culture medium. After
adherence and washings, they were activated overnight
with 20 U/ml murine IFN-g and 100 ng/ml LPS, in the
presence of 50 AM SEITU, to reduce NO production
and NO-dependent toxicity. Then, nonadherent cells
were eliminated by two vigorous washings and 2 �106 tumor cells were introduced into 5 ml of fresh
culture medium. When required, carboxy-PTIO (150
AM), HbO2 (175 AM), aminoguanidine (2 mM), or
dipyridamole (1.2 AM) was also added. At the end of
the coculture period, nonadherent cells were gently
removed and collected. An aliquot was counted after
trypan blue staining to evaluate cytotoxicity and the
remaining cell pellet was used for nucleotide analysis.
These cells were washed twice in ice-cold phosphate-
buffered saline. The cell pellet was rapidly resuspended
in cold 60% methanol (250 Al) and the resulting
suspension was allowed to stand for 2 min in boiling
water. Then, the cell extract was frozen at �80jC for at
least 1 day before analysis.
Coculture of macrophages with tumor cells for
ribonucleotide measurements
Experiments performed to measure ribonucleotide
pools were done under the conditions used for dNTP as-
say, but modified to handle a higher number of tumor cells.
Macrophages were initially seeded in larger culture dishes
(80 mm; Nunc) at a similar density. After macrophage
activation and elimination of nonadherent cells, 13 � 106
CEM-SS cells were added in 40 ml of culture medium.
Coculture of DLD-1 and CEM-SS cells
Experiments were carried out as already reported for
dNTP assay, except that 2.4 � 106 DLD-1 cells were
initially seeded into six-well culture plates. After 1 day,
cells were activated overnight with IL-1h (2 ng/ml),
human TNF-a (102 U/ml), and IFN-g (103 U/ml). Then,
2 � 106 CEM-SS cells were added and processed as
described above for dNTP extraction.

Depletion of dNTP pools by NO 509
Analysis of ribonucleotides
A Waters 600 multisolvent delivery system, a Waters
486 absorbance detector set at 260 nm, and Winflow
radiochromatography software (Berthold) were used for
the separation and quantification of ribonucleotides. Cell
extract (100–150 Al) was injected onto a Whatman
Partisil SAX 5 AM (250 � 4.6 mm) equilibrated in
buffer A (10 mM ammonium phosphate, pH 5.4), at a
flow rate of 2 ml/min. Elution started in 100% buffer A
for 7 min, then proceeded into 98% buffer A, 2% buffer
B (700 mM ammonium phosphate, pH 3.5) over 3 min,
and then switched to 100% buffer B using a linear
gradient over 40 min. The column was then regenerated
in buffer C (350 mM ammonium phosphate, pH 5.4) for
15 min before the next analysis was begun. Nucleotides
were identified by their retention time, compared to those
of authentic standards.
Analysis of dNTP
Quantification of dNTP in cell extracts was carried
out according to an enzymatic assay already described
[19]. This assay is based on the elongation of 32P-5V-end-labeled oligonucleotide primers annealed to com-
plementary oligonucleotide templates. Incorporation
within the primer/template was catalyzed by the Klenow
fragment of Escherichia coli, under conditions under
which the concentration of the dNTP to be analyzed is
limiting. The elongated products were separated after
electrophoresis on a denaturing 10% polyacrylamide
gel.
Determination of the pyrimidine salvage pathway activity
CEM-SS cells (3 � 106) were incubated for up to 6
h in culture medium supplemented with 0.1 AM 4 ACi[3H]deoxycytidine (dC*) and 100 AM SPER-NO, alone
or in combination. At different time intervals, cells were
collected and processed as described above for dNTP
analysis. About one-fifth of the material was used for
dCTP determination. The rest was used for the quantifi-
cation of radioactive dC metabolites (dC* and the
corresponding mono-, di-, and triphosphate derivatives).
HPLC separations were performed on an ion-exchange
Partisil 10-SAX column. The eluting buffers were 0.01
M potassium phosphate, pH 3.5 (buffer A), and 0.5 M
potassium phosphate, pH 3.5, containing 0.8 M KCl
(buffer B). The elution program of dC* and metabolites
was 8 min of buffer A followed by a 20 min linear
gradient to 100% buffer B and a 10 min isocratic elution
with 100% buffer B. The flow rate of the elution buffer
was 1 ml/min. Radioactivity was monitored by running
Quickszint Flow 306 scintillator (Zinsser Analytic)
through a radiomatic detector at a ratio of 2:1.
Measurement of nitrite production
Nitrite concentration was determined in cell culture
supernatants with the Griess reagent [7].
RESULTS
Depletion of dNTP pools induced by NO prodrugs
In a first set of experiments, structurally different
NO prodrugs were compared to HU in their ability to
deplete dNTP pools in human leukemia CEM-SS cells
(Fig. 1). Because RnR activity controls de novo
deoxynucleotide synthesis, dNTP concentrations are
indicative of intracellular RnR activity. HU (100 AM)
induced a pronounced and persistent decrease in dCTP
and dATP levels. The dTTP pool increased during the
first 4 h and then returned progressively to control
level. The dGTP concentration was not modified
during the first 6 h. All these variations were consis-
tent with previous observations [2,20]. The diazenium-
diolate DETA-NO is a long-lived NO donor (t1/2 = 20
h). When used at 1 mM, it induced dNTP variations
that closely resembled the HU profile. A small differ-
ence was a delay in maximal dTTP increase or in
dCTP decrease, probably reflecting the time before
sufficient NO amounts were released by DETA-NO
to inhibit RnR. The S-nitrosothiol SPER-NO at 100
AM had a transient effect on dNTP pools, which was
in accordance with its short half-life (t1/2 = 115 min in
RPMI medium at 37jC). This was particularly evident
on dCTP and dATP concentrations, which contracted
during the first 4 to 6 h and then returned to nearly
normal levels at 9 h. From these experiments, we
concluded that the NO prodrugs DETA-NO and
SPER-NO inhibited RnR activity, with a time course
related to their respective half-lives.
Activation of the pyrimidine salvage pathway by
SPER-NO
Inhibition of RnR activity by HU induces an
activation of the pyrimidine salvage pathway [2]. We
investigated whether such a regulatory mechanism was
induced by NO, by measuring incorporation of radio-
labeled dC into the dCTP pool. Addition of 0.1 AMradiolabeled dC to feed the salvage pathway attenuated
notably the decrease in dCTP concentration induced
by SPER-NO (Fig. 2A). An enhancement of the
amount of radiolabeled metabolites of dC was visible
in cells cultivated with tritiated dC and SPER-NO for
1 to 3 h, compared to controls with dC alone (Fig.
2B). The specific activity of dCTP (Fig. 2C, calculated
from the data in Figs. 2A and 2B) exhibited a
maximum between 1 and 3 h, coincident with the
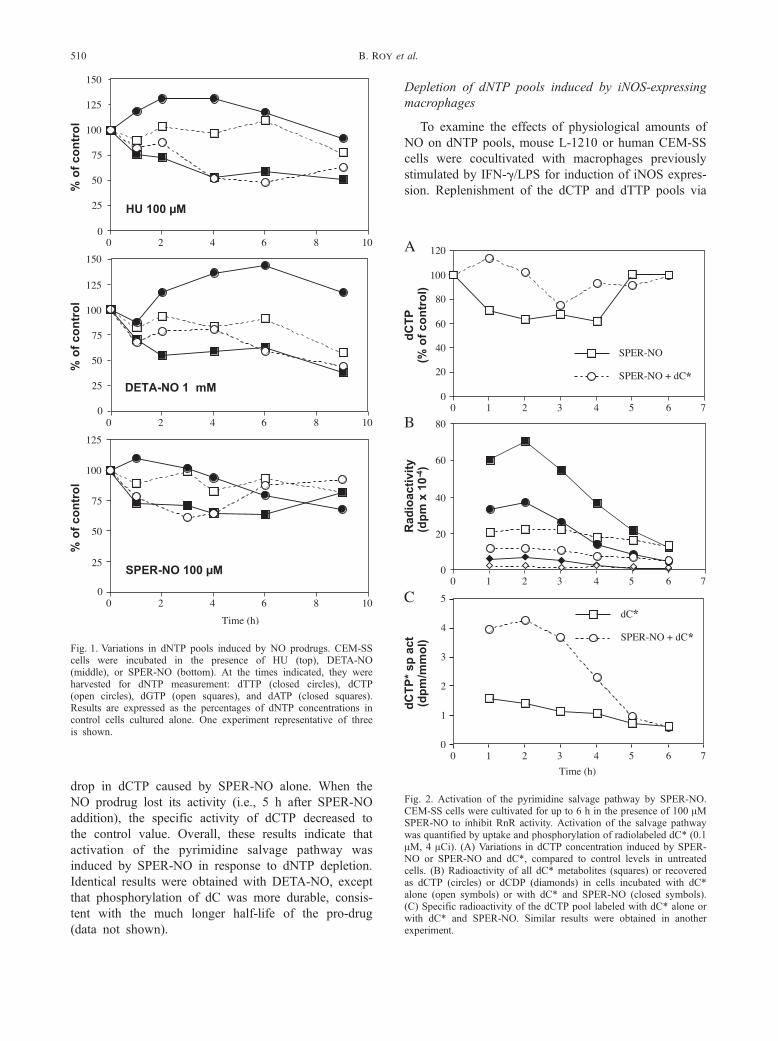
Fig. 2. Activation of the pyrimidine salvage pathway by SPER-NO.CEM-SS cells were cultivated for up to 6 h in the presence of 100 AMSPER-NO to inhibit RnR activity. Activation of the salvage pathwaywas quantified by uptake and phosphorylation of radiolabeled dC* (0.1AM, 4 ACi). (A) Variations in dCTP concentration induced by SPER-NO or SPER-NO and dC*, compared to control levels in untreatedcells. (B) Radioactivity of all dC* metabolites (squares) or recoveredas dCTP (circles) or dCDP (diamonds) in cells incubated with dC*alone (open symbols) or with dC* and SPER-NO (closed symbols).(C) Specific radioactivity of the dCTP pool labeled with dC* alone orwith dC* and SPER-NO. Similar results were obtained in anotherexperiment.
Fig. 1. Variations in dNTP pools induced by NO prodrugs. CEM-SScells were incubated in the presence of HU (top), DETA-NO(middle), or SPER-NO (bottom). At the times indicated, they wereharvested for dNTP measurement: dTTP (closed circles), dCTP(open circles), dGTP (open squares), and dATP (closed squares).Results are expressed as the percentages of dNTP concentrations incontrol cells cultured alone. One experiment representative of threeis shown.
B. Roy et al.510
drop in dCTP caused by SPER-NO alone. When the
NO prodrug lost its activity (i.e., 5 h after SPER-NO
addition), the specific activity of dCTP decreased to
the control value. Overall, these results indicate that
activation of the pyrimidine salvage pathway was
induced by SPER-NO in response to dNTP depletion.
Identical results were obtained with DETA-NO, except
that phosphorylation of dC was more durable, consis-
tent with the much longer half-life of the pro-drug
(data not shown).
Depletion of dNTP pools induced by iNOS-expressing
macrophages
To examine the effects of physiological amounts of
NO on dNTP pools, mouse L-1210 or human CEM-SS
cells were cocultivated with macrophages previously
stimulated by IFN-g/LPS for induction of iNOS expres-
sion. Replenishment of the dCTP and dTTP pools via
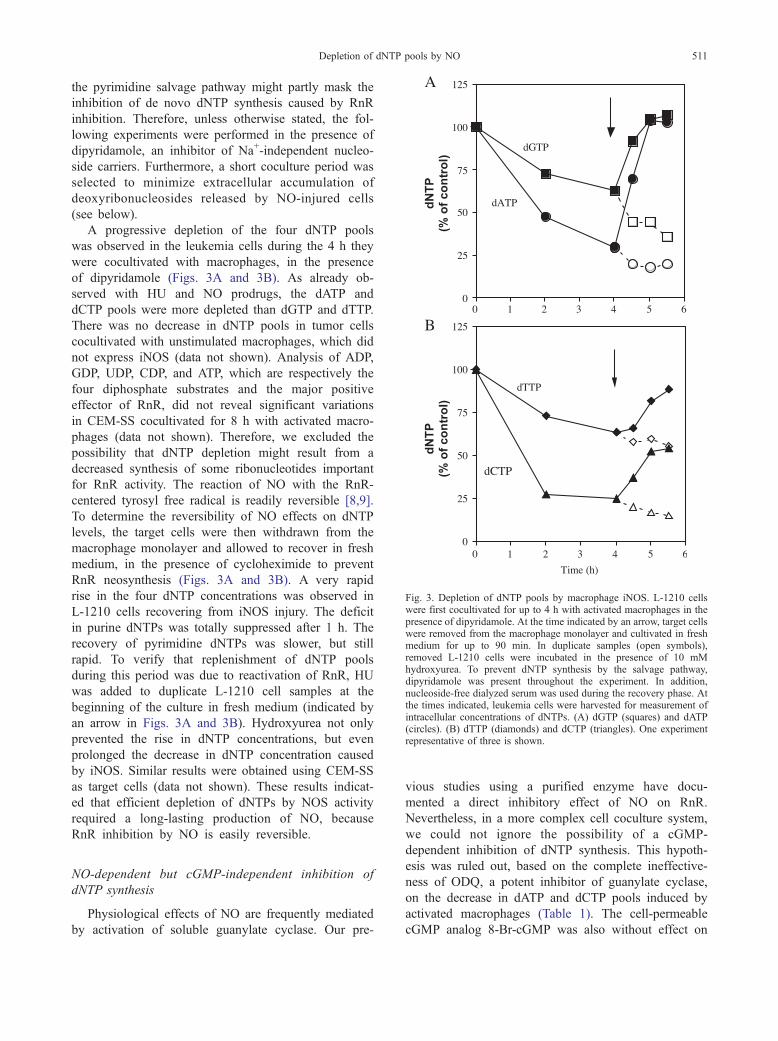
Fig. 3. Depletion of dNTP pools by macrophage iNOS. L-1210 cellswere first cocultivated for up to 4 h with activated macrophages in thepresence of dipyridamole. At the time indicated by an arrow, target cellswere removed from the macrophage monolayer and cultivated in freshmedium for up to 90 min. In duplicate samples (open symbols),removed L-1210 cells were incubated in the presence of 10 mMhydroxyurea. To prevent dNTP synthesis by the salvage pathway,dipyridamole was present throughout the experiment. In addition,nucleoside-free dialyzed serum was used during the recovery phase. Atthe times indicated, leukemia cells were harvested for measurement ofintracellular concentrations of dNTPs. (A) dGTP (squares) and dATP(circles). (B) dTTP (diamonds) and dCTP (triangles). One experimentrepresentative of three is shown.
Depletion of dNTP pools by NO 511
the pyrimidine salvage pathway might partly mask the
inhibition of de novo dNTP synthesis caused by RnR
inhibition. Therefore, unless otherwise stated, the fol-
lowing experiments were performed in the presence of
dipyridamole, an inhibitor of Na+-independent nucleo-
side carriers. Furthermore, a short coculture period was
selected to minimize extracellular accumulation of
deoxyribonucleosides released by NO-injured cells
(see below).
A progressive depletion of the four dNTP pools
was observed in the leukemia cells during the 4 h they
were cocultivated with macrophages, in the presence
of dipyridamole (Figs. 3A and 3B). As already ob-
served with HU and NO prodrugs, the dATP and
dCTP pools were more depleted than dGTP and dTTP.
There was no decrease in dNTP pools in tumor cells
cocultivated with unstimulated macrophages, which did
not express iNOS (data not shown). Analysis of ADP,
GDP, UDP, CDP, and ATP, which are respectively the
four diphosphate substrates and the major positive
effector of RnR, did not reveal significant variations
in CEM-SS cocultivated for 8 h with activated macro-
phages (data not shown). Therefore, we excluded the
possibility that dNTP depletion might result from a
decreased synthesis of some ribonucleotides important
for RnR activity. The reaction of NO with the RnR-
centered tyrosyl free radical is readily reversible [8,9].
To determine the reversibility of NO effects on dNTP
levels, the target cells were then withdrawn from the
macrophage monolayer and allowed to recover in fresh
medium, in the presence of cycloheximide to prevent
RnR neosynthesis (Figs. 3A and 3B). A very rapid
rise in the four dNTP concentrations was observed in
L-1210 cells recovering from iNOS injury. The deficit
in purine dNTPs was totally suppressed after 1 h. The
recovery of pyrimidine dNTPs was slower, but still
rapid. To verify that replenishment of dNTP pools
during this period was due to reactivation of RnR, HU
was added to duplicate L-1210 cell samples at the
beginning of the culture in fresh medium (indicated by
an arrow in Figs. 3A and 3B). Hydroxyurea not only
prevented the rise in dNTP concentrations, but even
prolonged the decrease in dNTP concentration caused
by iNOS. Similar results were obtained using CEM-SS
as target cells (data not shown). These results indicat-
ed that efficient depletion of dNTPs by NOS activity
required a long-lasting production of NO, because
RnR inhibition by NO is easily reversible.
NO-dependent but cGMP-independent inhibition of
dNTP synthesis
Physiological effects of NO are frequently mediated
by activation of soluble guanylate cyclase. Our pre-
vious studies using a purified enzyme have docu-
mented a direct inhibitory effect of NO on RnR.
Nevertheless, in a more complex cell coculture system,
we could not ignore the possibility of a cGMP-
dependent inhibition of dNTP synthesis. This hypoth-
esis was ruled out, based on the complete ineffective-
ness of ODQ, a potent inhibitor of guanylate cyclase,
on the decrease in dATP and dCTP pools induced by
activated macrophages (Table 1). The cell-permeable
cGMP analog 8-Br-cGMP was also without effect on
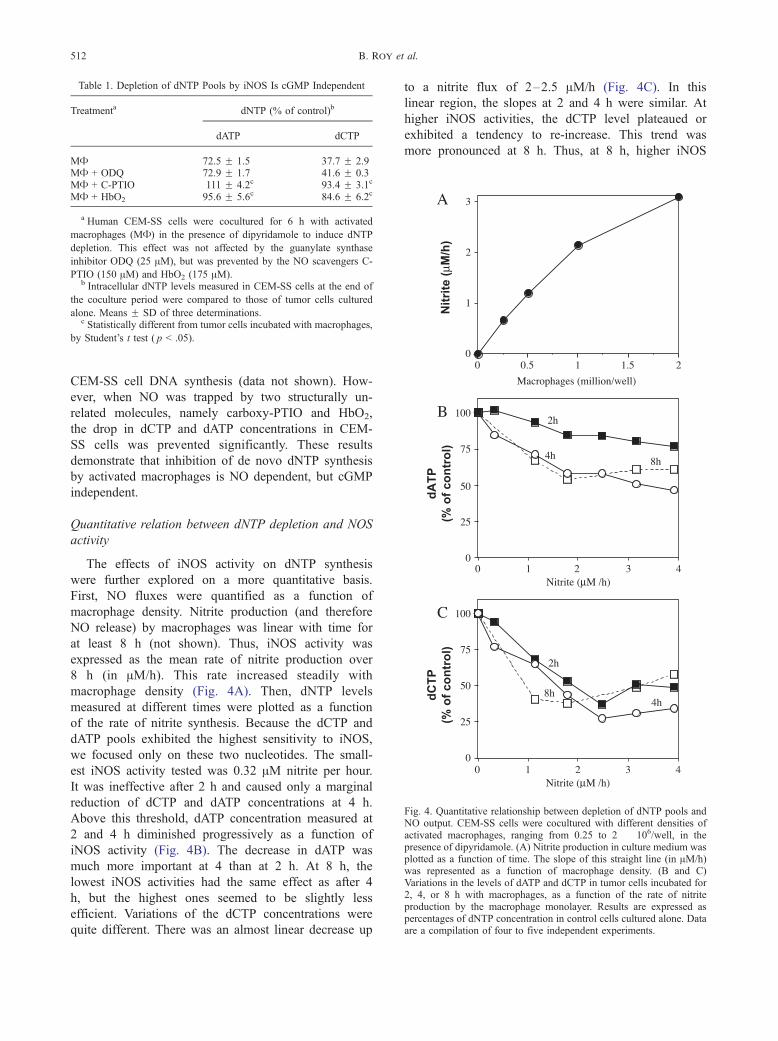
Table 1. Depletion of dNTP Pools by iNOS Is cGMP Independent
Treatmenta dNTP (% of control)b
dATP dCTP
MA 72.5 F 1.5 37.7 F 2.9MA + ODQ 72.9 F 1.7 41.6 F 0.3MA + C-PTIO 111 F 4.2c 93.4 F 3.1c
MA + HbO2 95.6 F 5.6c 84.6 F 6.2c
a Human CEM-SS cells were cocultured for 6 h with activated
macrophages (MA) in the presence of dipyridamole to induce dNTP
depletion. This effect was not affected by the guanylate synthase
inhibitor ODQ (25 AM), but was prevented by the NO scavengers C-
PTIO (150 AM) and HbO2 (175 AM).b Intracellular dNTP levels measured in CEM-SS cells at the end of
the coculture period were compared to those of tumor cells cultured
alone. Means F SD of three determinations.c Statistically different from tumor cells incubated with macrophages,
by Student’s t test ( p < .05).
Fig. 4. Quantitative relationship between depletion of dNTP pools andNO output. CEM-SS cells were cocultured with different densities ofactivated macrophages, ranging from 0.25 to 2 � 106/well, in thepresence of dipyridamole. (A) Nitrite production in culture medium wasplotted as a function of time. The slope of this straight line (in AM/h)was represented as a function of macrophage density. (B and C)Variations in the levels of dATP and dCTP in tumor cells incubated for2, 4, or 8 h with macrophages, as a function of the rate of nitriteproduction by the macrophage monolayer. Results are expressed aspercentages of dNTP concentration in control cells cultured alone. Dataare a compilation of four to five independent experiments.
B. Roy et al.512
CEM-SS cell DNA synthesis (data not shown). How-
ever, when NO was trapped by two structurally un-
related molecules, namely carboxy-PTIO and HbO2,
the drop in dCTP and dATP concentrations in CEM-
SS cells was prevented significantly. These results
demonstrate that inhibition of de novo dNTP synthesis
by activated macrophages is NO dependent, but cGMP
independent.
Quantitative relation between dNTP depletion and NOS
activity
The effects of iNOS activity on dNTP synthesis
were further explored on a more quantitative basis.
First, NO fluxes were quantified as a function of
macrophage density. Nitrite production (and therefore
NO release) by macrophages was linear with time for
at least 8 h (not shown). Thus, iNOS activity was
expressed as the mean rate of nitrite production over
8 h (in AM/h). This rate increased steadily with
macrophage density (Fig. 4A). Then, dNTP levels
measured at different times were plotted as a function
of the rate of nitrite synthesis. Because the dCTP and
dATP pools exhibited the highest sensitivity to iNOS,
we focused only on these two nucleotides. The small-
est iNOS activity tested was 0.32 AM nitrite per hour.
It was ineffective after 2 h and caused only a marginal
reduction of dCTP and dATP concentrations at 4 h.
Above this threshold, dATP concentration measured at
2 and 4 h diminished progressively as a function of
iNOS activity (Fig. 4B). The decrease in dATP was
much more important at 4 than at 2 h. At 8 h, the
lowest iNOS activities had the same effect as after 4
h, but the highest ones seemed to be slightly less
efficient. Variations of the dCTP concentrations were
quite different. There was an almost linear decrease up
to a nitrite flux of 2–2.5 AM/h (Fig. 4C). In this
linear region, the slopes at 2 and 4 h were similar. At
higher iNOS activities, the dCTP level plateaued or
exhibited a tendency to re-increase. This trend was
more pronounced at 8 h. Thus, at 8 h, higher iNOS

Depletion of dNTP pools by NO 513
activities were less effective than lower ones on dCTP
and dATP pools. This could be due to a residual
activity of the pyrimidine salvage pathway despite the
presence of dipyridamole, in conjunction with an
increased release of nucleosides by macrophages at
the most toxic iNOS activities (see below).
In one series of experiments, murine macrophages
were replaced by human DLD-1 cells, which, after
activation by cytokines, produced nitrite at a 10-fold
lower rate that murine macrophages on a per-cell basis.
There was no effect of DLD-1 cells on dNTP levels in
CEM-SS cells, for up to 8 h (data not shown).
Contribution of the salvage pathway to dNTP pool
imbalance
Pyrimidine deoxynucleosides are recycled into
dNTPs by the salvage pathway much more efficiently
than purine nucleosides. Therefore, inhibition of de novo
dNTP synthesis on one hand, and activation of the
salvage pathway on the other hand, may induce an
imbalance in intracellular dNTP pools if large amounts
of pyrimidine nucleosides are available in the medium.
Such an imbalance has been linked to apoptosis [21]. To
investigate whether RnR inhibition by NO caused a
dNTP imbalance, we analyzed dNTP variations in tumor
cells with an active salvage pathway (that is, in the
absence of dipyridamole). Results are presented in Table
2. As expected, we observed a decrease in dATP
concentration (by 60%) during the 16 h coculture period
with macrophages, which was consistent with our pre-
vious results. However, there was a huge increase in
pyrimidine nucleotide pools in the tumor cells and no
variation of dGTP level. That dGTP concentration was
Table 2. NO-Dependent RnR Inhibition and Activation of the PyrimidineSalvage Pathway
Additivea dNTP (% of control)b Nitrite
dCTP dTTP dATP dGTP
(AM)c
None 638 F 37d 681 F 53d 34 F 4d 103 F 15 31 F 0.3AG 148 F 10d 143 F 11d 104 F 6 136 F 5d NDe
DP 71 F 3d 122 F 11 24 F 1d 38 F 7d 31 F 0.9AG + DP 109 F 8 88 F 5 88 F 9 86 F 5 ND
a Human CEM-SS cells were cocultured for 16 h with activated
macrophages at a 1:1 ratio, without (None) and with the NOS inhibitor
aminoguanidine (AG), the nucleoside transporter inhibitor dipyridamole
(DP), or both (AG + DP).b Intracellular dNTP levels measured in CEM-SS cells at the end of
the coculture period were compared to those of tumor cells cultured
alone. Means F SD of triplicates.c Nitrite concentration in the culture medium.d Statistically different from tumor cells cultured alone, by Student’s
t test ( p < .05).e ND = not detectable.
not diminished might be explained by the 6-fold in-
crease in dTTP, because dTTP is a positive effector of
RnR for dGTP synthesis. All dNTP concentrations were
normalized by aminoguanidine, which completely
inhibited macrophage iNOS activity, as judged by un-
detectable nitrite production (Table 2). In particular, the
considerable expansion of dCTP and dTTP pools was
almost suppressed, suggesting that NO-injured cells
could be an important source of salvaged nucleosides.
Because such a dramatic increase in dTTP and dCTP
pools was not induced by NO prodrugs (Fig. 1), macro-
phages were probably the major source of nucleosides.
Indeed, significant amounts of dC (0.04–0.05 AM) and
dT (0.08–0.11 AM) were detected after 8 h in culture
supernatants from activated macrophages. Dipyridamole
efficiently reduced the rise in dTTP concentration and
allowed a marked decrease in the dCTP pool (Table 2),
confirming the important activity of the salvage pathway
in cells exposed to NO. Dipyridamole also induced a
decrease in the dGTP level and further reduced dATP
concentration.
We can now summarize the series of events affecting
deoxynucleotide metabolism in NO-injured cells. First,
sustained NO production inhibited de novo dNTP syn-
thesis at the level of ribonucleotide reduction. This in
turn activated the pyrimidine salvage pathway, to com-
pensate for RnR inhibition [2]. Activation of pyrimidine
recycling caused an increase in dCTP and dTTP levels.
The rise in pyrimidine dNTPs caused by macrophage
iNOS (Table 2) is much higher than the small increase
induced by NO prodrugs (Fig. 1), probably because
activated macrophages release significant amounts of
pyrimidine dNs. Because purine pools are not efficiently
replenished by the salvage pathway, the ratios
dCTP:dGTP and dTTP:dATP, close to 1 in CEM-SS
cells, were augmented respectively by a 5.6- and a 18-
fold factor in cells cocultured with macrophages. It
would be interesting to know whether this marked
asymmetry in dNTP concentration could contribute to
NO-dependent genotoxicity.
DISCUSSION
This study is the first detailed analysis of dNTP
variations in tumor cells exposed to NO generated either
from chemical NO prodrugs or from macrophage iNOS
activity. In both cases, there was a rapid depletion of
dNTP pools, especially dATP, that in turn activated the
pyrimidine salvage pathway. Because we excluded the
possibility that dNTP depletion might result from alter-
ations of ribonucleotide metabolism by NO, this work
provides direct evidence for NO-mediated RnR inactiva-
tion in intact cells. Similar variations in dNTP levels are
caused by pharmacological inhibitors of RnR [2,4,22].

B. Roy et al.514
However, the considerable expansion of dCTP and dTTP
concentrations induced by iNOS (but not by NO pro-
drugs) is unusual. Dipyridamole and NOS inhibitors
prevented this augmentation, suggesting that NO-injured
macrophages could release into the culture medium large
amounts of nucleosides that are salvaged by tumor cells.
Nitric oxide is known to induce apoptosis in macro-
phages [23,24]. Hence, hydrolysis of apoptotic DNA
fragments might produce pyrimidine deoxynucleosides.
Also, macrophages have been already reported to syn-
thesize important amounts of thymidine [25]. Thus,
macrophages could significantly contribute to pyrimidine
release in the medium and, therefore, to dNTP imbalance
in the target cells. Considering the promutagenic effects
of an abnormal expansion of dNTP pools [26] and the
presence of infiltrating macrophages in many tumors, it
would be valuable to know whether enhanced pyrimidine
availability and marked dNTP asymmetry could also
occur in a tumor tissue.
Several reports have established the importance of
RnR in tumor proliferation, invasiveness, and aggressiv-
ity. For instance, elevation of RnR activity has been
observed in malignant cells and is positively correlated
with tumor growth rate in rodents [27,28]. Increased
levels of R2 protein and message have been found in
malignant tumor tissues [29]. Deregulated expression of
protein R2 in cooperation with oncogenes has been
shown to enhance tumor transformation and malignant
potential [30]. In this context, it could be anticipated that
even partial inhibition of RnR could be detrimental to
tumors. It was therefore important to define the condi-
tions leading to RnR inhibition in intact cells. The
reversibility of RnR inactivation by NO suggested that
only sustained iNOS activities would be inhibitory, as
opposed to constitutive NOSs activated transiently. Our
results indicated that the low iNOS activity of human
DLD-1 cells is ineffective against RnR. Therefore, RnR
inhibition by NO could be induced by those cells able to
produce a high flux of NO. Many mouse and rat cells
have this capacity, upon stimulation with cytokines or
bacterial components. Several, but fewer, human cell
types have been also found to produce NO at a compa-
rable rate, including hepatocytes [31], airway epithelial
cells [32], and alveolar macrophages [33]. Endogenous
iNOS activities found in human tumors are much weaker
[34–37]. They are probably too low to influence RnR
activity and, hence, to affect tumor proliferation in an
RnR-dependent manner. Nevertheless, iNOS overexpres-
sion induced by immunomodulation or gene transfection
in some experimental cancers is accompanied frequently
by NO-mediated anti-tumor cytostatic effects [15,38,39].
The strong iNOS activities measured in these models are
in the range of those that would impair RnR activity.
Considering our results, we envisage that a reduction in
RnR activity might partly support the antiproliferative
effects observed in these models.
NO-mediated depletion of dNTP pools could lead to
cellular damage beyond cessation of cell growth. Several
studies have suggested a link between RnR inhibition and
apoptosis. Structurally unrelated RnR inhibitors were
shown to induce or enhance apoptosis in different exper-
imental models [22,40,41], strongly suggesting that RnR
inhibition could be a proapoptotic event. But the most
convincing data linking dNTP deficiency to apoptotic cell
death came from studies on the recently discovered
homolog of R2, named p53R2 because its expression is
p53 dependent [42–44]. This protein is implicated spe-
cifically in DNA repair. Inhibition of p53R2 in vitro or
genetic inactivation of p53R2 in mice caused enhanced
apoptosis [42,43,45]. There is a remarkable sequence
homology between R2 and p53R2. In particular the
tyrosyl residue carrying the free radical and the amino
acid ligands for the diiron center are conserved [44].
Because the radical is the primary target of NO in R2, it
is tempting to imagine a similar inactivation of p53R2 by
NO at the level of its free radical. This hypothesis is
currently under investigation in our laboratory. The pres-
ent work demonstrates that NO is a potent inhibitor of
RnR in tumor cells. Its effects on dNTP pools are nearly
identical to those of chemotherapeutic RnR inhibitors.
Notwithstanding its pleiotropic roles in the immune
system, NO might be thus considered as a natural equiv-
alent of, for instance, hydroxyurea. Other endogenous
inhibitors of RnR have already been identified. In yeast,
the protein Sml1 binds to the R1 subunit, thereby inacti-
vating the enzyme when dNTP synthesis is not required
[46]. In many other organisms, protein R1 binds dATP, a
potent feedback inhibitor of all RnR activities [1]. The
inhibitors Sml1 and dATP exert a regulatory role. They
are intrinsically different from iNOS activity, which
perturbs RnR function. In this regard, NO belongs to a
new category of natural RnR inhibitors, whose mode of
action resembles those of pharmacological inhibitors.
Acknowledgments—We thank B. Wolfersberger for excellent technicalassistance and J.-C. Drapier (ICSN, CNRS, Gif-sur-Yvette, France) forgiving us the DLD-1 cells. We also gratefully acknowledge M. Hours(IFR 46 CNRS) and M.-P. Courageot (UMR 8619 CNRS) for assistancewith cytometric analysis. This work was supported by Grant 5329 froml’Association pour la Recherche contre le Cancer (to G.L.) and twogrants from La Ligue Nationale contre le Cancer (to M.L. and O.G.).
REFERENCES
[1] Reichard, P. Ribonucleotide reductases: the evolution of allostericregulation. Arch. Biochem. Biophys. 397:149–155; 2002.
[2] Bianchi, V.; Pontis, E.; Reichard, P. Changes of deoxyribonucleo-side triphosphate pools induced by hydroxyurea and their relationto DNA synthesis. J. Biol. Chem. 261:16037–16042; 1986.

Depletion of dNTP pools by NO 515
[3] Yarbro, J. W. Mechanism of action of hydroxyurea. Semin. Oncol.19:1–10; 1992.
[4] Plunkett, W.; Huang, P.; Xu, Y. Z.; Heinemann, V.; Grunewald,R.; Gandhi, V. Gemcitabine: metabolism, mechanisms of action,and self-potentiation. Semin. Oncol. 22:3–10; 1995.
[5] Lepoivre, M.; Fieschi, F.; Coves, J.; Thelander, L.; Fontecave, M.Inactivation of ribonucleotide reductase by nitric oxide. Biochem.Biophys. Res. Commun. 179:442–448; 1991.
[6] Kwon, N. S.; Stuehr, D. J.; Nathan, C. F. Inhibition of tumor cellribonucleotide reductase by macrophage-derived nitric oxide.J. Exp. Med. 174:761–767; 1991.
[7] Lepoivre, M.; Flaman, J.-M.; Henry, Y. Early loss of the tyrosylradical in ribonucleotide reductase of adenocarcinoma cells pro-ducing nitric oxide. J. Biol. Chem. 267:22994–23000; 1992.
[8] Roy, B.; Lepoivre, M.; Henry, Y.; Fontecave, M. Inhibition of ri-bonucleotide reductase by nitric oxide derived from thionitrites:reversible modifications of both subunits. Biochemistry 34:5411–5418; 1995.
[9] Guittet, O.; Ducastel, B.; Salem, J. S.; Henry, Y.; Rubin, H.;Lemaire, G.; Lepoivre, M. Differential sensitivity of the tyrosylradical of mouse ribonucleotide reductase to nitric oxide and per-oxynitrite. J. Biol. Chem. 273:22136–22144; 1998.
[10] Lemaire, G.; Alvarez-Pachon, F. J.; Beuneu, C.; Lepoivre, M.;Petit, J. F. Differential cytostatic effects of NO donors andNO producing cells. Free Radic. Biol. Med. 26:1274–1283;1999.
[11] Burney, S.; Tamir, S.; Gal, A.; Tannenbaum, S. R. A mechanisticanalysis of nitric oxide-induced cellular toxicity. Nitric Oxide1:130–144; 1997.
[12] Poluha, W.; Schonhoff, C. M.; Harrington, K. S.; Lachyankar,M. B.; Crosbie, N. E.; Bulseco, D. A.; Ross, A. H. A novel,nerve growth factor-activated pathway involving nitric oxide,p53, and p21(WAF1) regulates neuronal differentiation ofPC12 cells. J. Biol. Chem. 272:24002–24007; 1997.
[13] Pervin, S.; Singh, R.; Chaudhuri, G. Nitric oxide-induced cyto-stasis and cell cycle arrest of a human breast cancer cell line(MDA-MB-231): potential role of cyclin D1. Proc. Natl. Acad.Sci. USA 98:3583–3588; 2001.
[14] Moochhala, S.; Rajnakova, A. Role of nitric oxide in cancer biol-ogy. Free Radical Res. 31:671–679; 1999.
[15] Xie, K. P.; Fidler, I. J. Therapy of cancer metastasis by activationof the inducible nitric oxide synthase. Cancer Metastasis Rev.17:55–75; 1998.
[16] Burney, S.; Caulfield, J. L.; Niles, J. C.; Wishnok, J. S.; Tannen-baum, S. R. The chemistry of DNA damage from nitric oxide andperoxynitrite. Mutat. Res. 424:37–49; 1999.
[17] Lepoivre, M.; Flaman, J. M.; Bobe, P.; Lemaire, G.; Henry, Y.Quenching of the tyrosyl free radical of ribonucleotide reductaseby nitric oxide—relationship to cytostasis induced in tumor cellsby cytotoxic macrophages. J. Biol. Chem. 269:21891–21897;1994.
[18] Schmidt, K.; WernerFelmayer, G.; Mayer, B.; Werner, E. R. Pref-erential inhibition of inducible nitric oxide synthase in intact cellsby the 4-amino analogue of tetrahydrobiopterin. Eur. J. Biochem.259:25–31; 1999.
[19] Roy, B.; Beuneu, C.; Roux, P.; Buc, H.; Lemaire, G.; Lepoivre, M.Simultaneous determination of pyrimidine or purine deoxyribo-nucleoside triphosphates using a polymerase assay. Anal. Bio-chem. 269:403–409; 1999.
[20] Gao, W. Y.; Zhou, B. S.; Johns, D. G.; Mitsuya, H.; Yen, Y. Roleof the M2 subunit of ribonucleotide reductase in regulation byhydroxyurea of the activity of the anti-HIV-1 agent 2V,3V-di-deoxyinosine. Biochem. Pharmacol. 56:105–112; 1998.
[21] Oliver, F. J.; Collins, M. K. L.; Lopez-Rivas, A. dNTP pools im-balance as a signal to initiate apoptosis. Experientia 52:995–1000;1996.
[22] Grusch, M.; Fritzer-Szekeres, M.; Fuhrmann, G.; Rosenberger, G.;Luxbacher, C.; Elford, H. L.; Smid, K.; Peters, G. J.; Szekeres, T.;Krupitza, G. Activation of caspases and induction of apoptosis by
novel ribonucleotide reductase inhibitors amidox and didox. Exp.Hematol. 29:623–632; 2001.
[23] Brune, B.; vonKnethen, A.; Sandau, K. B. Nitric oxide and its rolein apoptosis. Eur. J. Pharmacol. 351:261–272; 1998.
[24] Eu, J. P.; Liu, L. M.; Zeng, M.; Stamler, J. S. An apoptotic modelfor nitrosative stress. Biochemistry 39:1040–1047; 2000.
[25] Stadecker, M. J.; Calderon, J.; Karnovsky, M. L.; Unanue, E. R.Synthesis and release of thymidine by macrophages. J. Immunol.119:1738–1743; 1977.
[26] Martomo, S. A.; Mathews, C. K. Effects of biological DNA pre-cursor pool asymmetry upon accuracy of DNA replication invitro. Mutat. Res. 499:197–211; 2002.
[27] Elford, H. L.; Freese, M.; Passamani, E.; Morris, H. P. Ribonu-cleotide reductase and cell proliferation. I. Variations of ribonu-cleotide reductase activity with tumor growth rate in a series of rathepatomas. J. Biol. Chem. 245:5228–5233; 1970.
[28] Weber, G. Biochemical strategy of cancer cells and the design ofchemotherapy. Cancer Res. 43:3466–3492; 1983.
[29] Jensen, R. A.; Page, D. L.; Holt, J. T. Identification of genesexpressed in premalignant breast disease by microscopy-direc-ted cloning. Proc. Natl. Acad. Sci. USA 91:9257 –9261;1994.
[30] Fan, H. Z.; Villegas, C.; Huang, A.; Wright, J. A. The mammalianribonucleotide reductase R2 component cooperates with a varietyof oncogenes in mechanisms of cellular transformation. CancerRes. 58:1650–1653; 1998.
[31] Nussler, A. K.; Di Silvio, M.; Billiar, T. R.; Hoffman, R. A.;Geller, D. A.; Selby, R.; Madariaga, J.; Simmons, R. L. Stimula-tion of the nitric oxide synthase pathway in human hepatocytes bycytokines and endotoxin. J. Exp. Med. 176:261–264; 1992.
[32] Guo, F. H.; Deraeve, H. R.; Rice, T. W.; Stuehr, D. J.; Thun-nissen, F. B. J. M.; Erzurum, S. C. Continuous nitric oxidesynthesis by inducible nitric oxide synthase in normal humanairway epithelium in vivo. Proc. Natl. Acad. Sci. USA 92:7809–7813; 1995.
[33] Hickman-Davis, J. M.; O’Reilly, P.; Davis, I. C.; Peti-Peterdi, J.;Davis, G.; Young, K. R.; Devlin, R. B.; Matalon, S. Killing ofKlebsiella pneumoniae by human alveolar macrophages. Am. J.Physiol. 282:L944–L956; 2002.
[34] Reveneau, S.; Arnould, L.; Jolimoy, G.; Hilpert, S.; Lejeune, P.;SaintGiorgio, V.; Belichard, C.; Jeannin, J. F. Nitric oxide syn-thase in human breast cancer is associated with tumor grade,proliferation rate, and expression of progesterone receptors. Lab.Invest. 79:1215–1225; 1999.
[35] Thomsen, L. L.; Miles, D. W.; Happerfield, L.; Bobrow, L. G.;Knowles, R. G.; Moncada, S. Nitric oxide synthase activity inhuman breast cancer. Br. J. Cancer 72:41–44; 1995.
[36] Jansson, O. T.; Morcos, E.; Brundin, L.; Bergerheim, U. S. R.;Adolfsson, J.; Wiklund, N. P. Nitric oxide synthase activity inhuman renal cell carcinoma. J. Urol. 160:556–560; 1998.
[37] Koh, E.; Noh, S. H.; Lee, Y. D.; Lee, H. Y.; Han, J. W.; Lee, H. W.;Hong, S. Differential expression of nitric oxide synthase in humanstomach cancer. Cancer Lett. 146:173–180; 1999.
[38] Onier, N.; Hilpert, S.; Reveneau, S.; Arnould, L.; SaintGior-gio, V.; Exbrayat, J. M.; Jeannin, J. E. Expression of induciblenitric oxide synthase in tumors in relation with their regres-sion induced by lipid A in rats. Int. J. Cancer 81:755–760;1999.
[39] Hajri, A.; Metzger, E.; Vallat, F.; Coffy, S.; Flatter, E.; Evrard, S.;Marescaux, J.; Aprahamian, M. Role of nitric oxide in pancreatictumour growth: in vivo and in vitro studies. Br. J. Cancer 78:841–849; 1998.
[40] Nandy, P.; Lien, E. J.; Avramis, V. I. Inhibition of ribonucleotidereductase by a new class of isoindole derivatives: drug synergismwith cytarabine (Ara-C) and induction of cellular apoptosis. Anti-cancer Res. 19:1625–1633; 1999.
[41] Anand, S.; Verma, H.; Kumar, L.; Singh, N. Induction of apop-tosis in chronic myelogenous leukemia lymphocytes by hydro-xyurea and adriamycin. Cancer Lett. 88:101–105; 1995.

B. Roy et al.516
[42] Tanaka, H.; Arakawa, H.; Yamaguchi, T.; Shiraishi, K.; Fukuda,S.; Matsui, K.; Takei, Y.; Nakamura, Y. A ribonucleotide reductasegene involved in a p53-dependent cell-cycle checkpoint for DNAdamage. Nature 404:42–49; 2000.
[43] Yamaguchi, T.; Matsuda, K.; Sagiya, Y.; Iwadate, M.; Fujino,M. A.; Nakamura, Y.; Arakawa, H. p53R2-dependent pathway forDNA synthesis in a p53-regulated cell cycle checkpoint. CancerRes 61:8256–8262; 2001.
[44] Guittet, O.; Hakansson, P.; Voevodskaya, N.; Fridd, S.; Graslund,A.; Arakawa, H.; Nakamura, Y.; Thelander, L. Mammalian p53R2protein forms an active ribonucleotide reductase in vitro with theR1 protein, which is expressed both in resting cells in response toDNA damage and in proliferating cells. J. Biol. Chem. 276:40647–40651; 2001.
[45] Kimura, T.; Takeda, S.; Sagiya, Y.; Gotoh, M.; Nakamura, Y.;Arakawa, H. Impaired function of p53R2 in Rrm2b-null micecauses severe renal failure through attenuation of dNTP pools.Nat. Genet. 34:440–445; 2003.
[46] Zhao, X.; Muller, E. G.; Rothstein, R. A suppressor of two essen-tial checkpoint genes identifies a novel protein that negativelyaffects dNTP pools. Mol. Cell 2:329–340; 1998.
ABBREVIATIONS
dNTP—deoxyribonucleoside triphosphate
NOS—NO synthase
iNOS—inducible NOS
RnR—ribonucleotide reductase
SEITU—S-ethylisothiourea
HU—hydroxyurea
![Triphosphate Tunnel Metalloenzyme Function in Senescence ... · Triphosphate Tunnel Metalloenzyme Function in Senescence Highlights a Biological Diversification of This Protein Superfamily1[OPEN]](https://img.pdfslide.net/doc/110x75/5e1eadbfbc21573d060be539/triphosphate-tunnel-metalloenzyme-function-in-senescence-triphosphate-tunnel.jpg)




![Gemcitabine and Doxorubicin Combination Enhance the Cytotoxic … · 2017-10-26 · ther phosphorylate dFdCDP to its another active triphosphate (dFdCTP) forms [6]. Deoxyribonucleoside](https://img.pdfslide.net/doc/110x75/5e3ea43e2e3439223d37c393/gemcitabine-and-doxorubicin-combination-enhance-the-cytotoxic-2017-10-26-ther.jpg)























