Embed Size (px)
Citation preview

Biologicals 33 (2005) 241e245www.elsevier.com/locate/biologicals
Determination of the effectiveness of Pseudorabies markervaccines in experiments and field trials
Annemarie Bouma*
Department of Farm Animal Health, Faculty of Veterinary Medicine, Utrecht University, Yalelaan 7, 3584 CL Utrecht, The Netherlands
Accepted 18 August 2005
Abstract
The aim of vaccination in an eradication campaign is not only to induce clinical protection, but primarily to stop transmission of infectionswithin and between herds by inducing herd immunity. Consequently, vaccines should be evaluated for their capacity to reduce virus transmissionin the population. Glycoprotein E (gE) negative marker vaccines against Pseudorabies virus (PRV) infections in pigs have been evaluated thisway in experiments and field studies. PRV infection in groups of (vaccinated) pigs was determined by measuring antibodies against gE of PRVfrom regularly taken serum samples. For the statistical analysis of the experiments a stochastic susceptibleeinfectiouseremoved (SIR) modelwas used. A measure for the transmission of virus is the reproduction ratio R, which is defined as the average number of secondary cases causedby one typical infectious individual. This implies that an infection will always fade out in a population when R! 1, but the infection can spreadmassively when RO 1. From several experiments it was shown that R! 1. Field studies showed that the R within herds was still O1, but byreducing further contacts the R could be reduced to a value below one. This would imply that PRV could be eradicated by means of vaccination.In The Netherlands, an eradication campaign was launched in 1993, and in 2002 the virus was eradicated, as shown by a negligible number ofgE-positive pigs. Farmers’ organizations have to decide whether or not to stop vaccination.� 2005 The International Association for Biologicals. Published by Elsevier Ltd. All rights reserved.
Keywords: Pseudorabies; Eradication; Marker vaccines; Vaccination
1. Introduction
Infectious diseases in animal husbandry are still common,and the interest for strategies to control them has increased.Control of infectious diseases can focus on the control of dis-ease or on prevention of spread of the pathogens between ani-mals. The strategy to be followed depends on the disease ofinterest, and may change over time.
Vaccines against viral diseases are often used in the controlof diseases, as they usually induce good clinical protectionagainst disease. The development of marker vaccines offeredthe opportunity to reconsider the control strategy. An exampleof such reconsideration is the change in the control strategy ofPseudorabies or Aujeszky’s disease in The Netherlands. This
* Tel.: C31 30 2531013; fax: C31 30 2521887.
E-mail address: [email protected]
1045-1056/05/$30.00 � 2005 The International Association for Biologicals. Pub
doi:10.1016/j.biologicals.2005.08.011
paper reviews the research carried out to evaluate the effec-tiveness of marker vaccines against Pseudorabies virus (PRV).
1.1. Historic background of the situation
Pseudorabies was described for the first time at the begin-ning of the 19th century in the United States, and is causedby a virus of the Herpes virus group. Pseudorabies is primarilyan infection of pigs, which represent its only known reservoirhost. Pseudorabies is characterized by a variety of clinicalsigns, involving the nervous and respiratory systems being par-ticularly prominent (e.g. [1]). The virus had not existed widelyoutside Eastern Europe before the 1960s but by 1989 it hada worldwide distribution affecting 43 countries, including sev-eral regions of Europe, South East Asia including China andCentral and South America.
Also in The Netherlands, most herds became infected. Thewithin and the between-herd prevalences were high, and the
lished by Elsevier Ltd. All rights reserved.

242 A. Bouma / Biologicals 33 (2005) 241e245
disease became an increasingly important disease, because ofthe economic losses for individual farmers. At first, the problemwas considered a farm problem, to be solved at the farm level.Vaccines were applied widely with the aim of inducing clinicalprotection of individual pigs. Some disease outbreaks occurred,despite vaccination, but in general, the vaccines could preventor reduce clinical signs due to PRV infections sufficiently.
Then, the economic importance of the disease changed.Since the intensification of the swine and pork production,the export of pigs and pig products became more importantfor Dutch farmers. In the mid-1980s of the 20th century theexport was endangered by the PRV-free status of other (partsof) countries in the EU, such as the United Kingdom and Den-mark. In these countries, the disease has been eradicated byusing large-scale serological surveys to identify pigs exposedto PRVand these pigs were subsequently culled. The main ad-vantage for those countries was that they could export to Japanand the United States. These countries did not allow import ofpork products from countries where the disease was endemicor where pigs were vaccinated. Because The Netherlandswas still vaccinating, and because it was impossible in vacci-nated herds to detect infected pigs, The Netherlands was afraidof losing their export market. Thus, Pseudorabies became anincreasingly important disease for farmers, not for direct los-ses, but for losses of export [2].
1.2. Reconsideration of the situation
The control of the disease, the clinical signs, was more orless effective by the application of vaccines. A major break-through in the control of Pseudorabies occurred when markervaccines were developed in the 1980s of the 20th century. Pigsvaccinated with a marker vaccine do not produce antibodiesagainst the marker protein, whereas pigs infected with wildtype PRV do develop antibodies directed against all viral gly-coproteins, including the marker. In combination with a sero-logical test to detect antibodies directed against the markerprotein, vaccinated pigs can be distinguished from infectedpigs [3e5]. Since then, discussions about a potential eradica-tion of PRV in The Netherlands began. Before starting anobligatory vaccination campaign, the efficacy of marker vac-cines had to be demonstrated.
2. Evaluation of the effectiveness of vaccines
Vaccines have usually been evaluated in vaccination-chal-lenge experiments, in which vaccinated pigs were challengedwith a virulent wild type (WT) strain to determine the clinicalprotection after infection and amount of excreted virus. Thesestudies often showed that vaccination could not preventchallenge-infection or virus excretion (e.g. [6e8]). The find-ings in these studies suggest that eradication of PRV was notpossible.
When considering an eradication campaign for PRV bymeansof vaccination, however, it is crucial that the vaccine can pre-vent the spread of PRV. The classical vaccination-challenge
experiments do not shed light on how transmission occurs,as they focus on characteristics of the infection that are impor-tant for the individual pig. Therefore, it should be studied howPRV may spread in the population and which factors may in-fluence the transmission of PRV [9].
Transmission of PRV has been studied using susceptiblesentinel pigs. In these studies, it was investigated not onlywhether PRV could spread [10e15], but also via which routestransmission occurred [16,17]. Although these studies demon-strated qualitatively that PRV could spread from infected tosentinel pigs, they did not address the crucial questions of:(1) to what extent does PRV spread, and (2) are the observeddifferences in transmission significant and relevant? Conse-quently, these studies were not suitable to evaluate the effectof control measures on the transmission of PRV.
To answer these crucial questions, transmission has to bequantified. Data that can be used for the quantification oftransmission can be obtained from laboratory experimentsand from field observations. The incidences of infection indifferent groups within the same population are dependent,generating non-linear infection dynamics [18]. To unravelthe complexity of the dynamic behaviour of infections inpopulations, non-linear mathematical models are useful.They are used to generate hypotheses about how transmissionoccurs, to understand epidemiological processes, and to eval-uate the possible effect of control measures [19]. Mathemat-ical models can also be used to estimate transmission ofinfections, and for a better understanding of the relation be-tween variables that determine the course of infection in indi-viduals and the variables that determine the pattern ofinfection in populations [20].
A measure for transmission of infections is the reproduc-tion ratio R. The R is defined as the average number of second-ary cases caused by one typical infectious individual, and isa measure for transmission from one generation to the nextgeneration [19]. R has a threshold value (RZ 1). When R isbelow one, a small part of the population may become in-fected, called a minor outbreak. When R is above one, a largepart of the population may become infected, called a majoroutbreak, although minor outbreaks can also occur due tochance processes [21].
A simple susceptibleeinfectiouseremoved (SIR) modelmakes it possible to determine the reproduction ratio fromthe incidence of new infections in a population. In the SIRmodel a population (N individuals) consists of susceptible(S ), infectious (I ) and recovered (R) individuals. Infection oc-curs with rate b and recovery with rate a [22]. The reproduc-tion ratio is b/a.
The main question in a vaccination campaign meant toeradicate PRV is whether the vaccine can reduce transmissionsufficiently, in other words whether the vaccine can reduce Rto a value below one.
This might be achieved since the effect of vaccination onthe individual is twofold. First, vaccination can reduce the sus-ceptibility of an individual against infection. This means thatan individual has a reduced chance of becoming infected,for example because the infectious dose needed to establish

243A. Bouma / Biologicals 33 (2005) 241e245
an infection is higher after vaccination [23,24]. Second, vacci-nation can reduce the infectivity after the occurrence of an in-fection [6,8]. Clinical protection and reduced susceptibility areof benefit to the individual. The reduced susceptibility reducesthe probability of an individual becoming infected after con-tact with an infectious individual; the clinical protection re-duces the severity of symptoms due to the disease when ananimal is infected. The reduced susceptibility and infectivity,called herd immunity, is of benefit to the population. Thisherd immunity is the reduced probability of an individual be-coming infected when it is part of a vaccinated population[25,26]. The chance of becoming infected in a population de-creases with an increasing density of individuals being vacci-nated. Note that herd immunity applies to vaccinatedindividuals as well as to unvaccinated individuals. Conse-quently, protection against disease can be induced at the indi-vidual level by vaccinating (a part of) any population with(possibly other) individuals that need protection. Even whenan individual is not vaccinated it has a reduced chance of be-coming infected, because of the herd immunity.
For PRV it is important that a vaccine can induce sufficientherd immunity. The herd immunity should be sufficient to re-duce the probability of not-yet infected animals becoming in-fected, i.e. prevent the transmission of pathogen within thepopulation to such an extent that the infection will eventuallygo extinct (R! 1). It will be illustrated that it is not only the-oretically but also practically possible to quantify herd immu-nity among farm animals with data from small-scaleexperiments as well as with data from field trials.
3. Transmission experiments
The first experiment in which the effectiveness of a gE-neg-ative vaccine against PRV transmission was determined wascarried out by De Jong and Kimman [27]. They performedan experiment with two groups of 10 pigs each, each grouphoused in an isolated unit. All pigs in one group were vacci-nated twice with a gE-deleted vaccine, whereas all pigs inthe other group remained unvaccinated. The infection chain,necessary to quantify transmission, was started by intranasalinoculation of half of each group with the mildly virulentWT PRV Sterksel strain. The other half of each group was con-tact-exposed to the inoculated pigs for 6 weeks. Transmissionof PRV was determined from the number of contact-infections,and the reproduction ratio was calculated in both groups andcompared. They estimated the reproduction ratio R in the vac-cinated group to be 0.5 whereas the R in the unvaccinatedgroup was estimated to be 10, which was a significantdifference.
The effectiveness of a vaccine to prevent transmission ofvirus should be determined in homogeneous groups. This im-plies that all animals in one group should be vaccinated,whereas all animals in the second group remain unvaccinated.The animals are assumed to be identical, i.e. ‘equally’ infec-tious or susceptible, and to mingle freely within the group.This experimental set-up differs from the approach to
determine virus transmission by adding susceptible sentinelanimals in a group of vaccinated animals [10e15]. Transmis-sion, however, cannot be estimated properly using such a het-erogeneous group, because of the following reason.Vaccination generally reduces both the infectivity and thesusceptibility of an animal [7,24]. Infecting unvaccinated an-imals in a group of vaccinated contact animals might result inextended virus spread (RO 1), because the unvaccinated in-oculated animals are more infectious than vaccinated inocu-lated animals would have been. Vice versa, unvaccinatedcontact animals in a group of vaccinated inoculated animalsmight result in extended virus spread (RO 1), because theunvaccinated contact animals are more susceptible than vac-cinated contact animals would have been. Moreover, it is pos-sible that the inoculated vaccinated animals infect forexample only one unvaccinated sentinel (Rv! 1), that, inturn, might infect the other unvaccinated contact animals(RnvO 1). Then, the effectiveness of vaccination is likelyto be underestimated.
The experiments by De Jong and Kimman [27] were car-ried out with a mildly virulent strain using SPF pigs. Basedon knowledge of the interaction between PRV and individualpigs, one can imagine that other factors might affect the trans-mission of PRV, e.g. the virulence of the WT strain, thepresence of maternally derived antibodies (MDA), or the vac-cine used. More researche was done to evaluate these riskfactors.
Vaccination of pigs with maternal immunity to PRV wasnot less effective than vaccination of SPF pigs, and in bothgroups R! 1 [28]. Two vaccines were compared, that differedin the capacity, to reduce virus excretion after challenge. Al-though it seemed that one vaccine was more able to reduce in-fectiousness than the other vaccine, both vaccines reduced theR sufficiently [29]. Virulent strains are often assumed tospread more or more quickly than less virulent strains. A com-parison of transmission between NIA3, a very virulent strain,and Sterksel, however, did not show a significant difference ingroups of vaccinated pigs [30], although NIA3 was excreted ina higher amount than Sterksel. Probably, the virulence ofNIA3 is mainly determined by the capacity of the virus to in-duce meningitis, rather than in a high level of excretion. Thisfinding indicates that virulent strains do not necessarily spreadmore than less virulent strains.
In all these transmission experiments, the R value wasbelow one, although not always significant. These findingssuggest that eradication of PRV by vaccination should bepossible.
4. Field studies
Experiments are hampered by problems associated with ex-trapolating results to the field. Consequently, a field study inthe target population is the ultimate test for a vaccine, as vac-cines have to exert their effects in the field. Again, the devel-opment of the marker vaccines offered the opportunity toevaluate the effectiveness of vaccines under field conditions.
244 A. Bouma / Biologicals 33 (2005) 241e245
Van Nes et al. [31] studied infection chains in sow herds. Inthese herds, it was expected that the virus might persist easily,because of the constant influx of new, susceptible piglets. Thesows were vaccinated every 4 months with 783 in OW. Trans-mission was determined by measuring seroconversion for gE 2weeks after each transport of a sow to another unit. They ob-served only minor outbreaks of PRV, and estimated the R insow herds to be 0.7, probably because of the limited contactsbetween sows. From these results one could conclude thateradication of PRV from sow herds was possible.
Stegeman et al. [32] compared the effectiveness of vacci-nating pigs once, 4 weeks after arrival at the farm (approxi-mately 14 weeks of age) with vaccinating pigs twice, 1 and5 weeks after arrival (approximately 11 and 15 weeks ofage). They showed that in groups of finishing pigs that werevaccinated once, the R was 3.5, and in groups of pigs thatwere vaccinated twice R was 1.5. A possible explanation isthe difference between SPF and conventional pigs. Van Neset al. [33] showed that vaccination of conventional pigs wasless effective than vaccination of SPF pigs. This suggeststhat eradication by the application of vaccines would not bepossible. Moreover, the findings seemed conflicting with theexperimental work mentioned before. Explanations are thepresence of other risk factors such as a sub-optimal vaccina-tion, climate, other diseases, etc.
Although the transmission within finishing herd was not re-duced sufficiently to eradicate the virus, eradication of PRVona region level could still be possible. Van Nes et al. [34]showed in a mathematical model the relation between thewithin-herd transmission and between-herd transmission.The application of marker vaccines, in combination with addi-tional management measures, such as reduced contacts be-tween herds, could result in a reproduction ratio below one.These findings suggest that it might be possible to eradicatePRV by means of vaccination [2].
5. Current situation in The Netherlands
The Dutch farmers’ organization adopted control policies.A three-stage program was announced in 1993. The first stagestarted in 1995, comprised compulsory vaccination with gE-deleted vaccines. This should lead to a disease-free certifica-tion program for individual herd, accompanied by regularmonitoring. In subsequent stages, remaining sources of infec-tion (gE-positive animals) were traced and eliminated, al-though it can be questioned whether these gE-positive pigsare a threat with respect to the occurrence of new outbreaksas reactivation does not seem to play an important role inthe spread of PRV [34].
Five marker vaccines were accepted in the market. Finish-ing pigs and gilts were vaccinated twice before the age of 16weeks; sows were vaccinated four times per year. Serum sam-ples were taken regularly to monitor the prevalence on farmand regional levels.
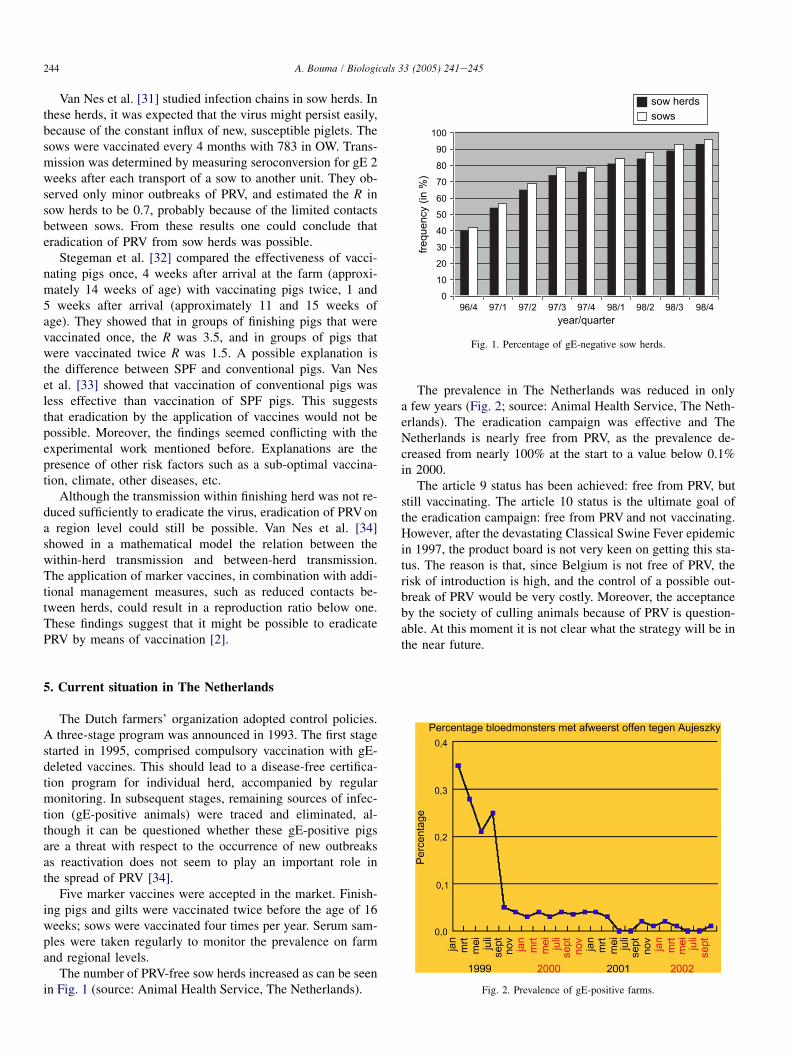
The number of PRV-free sow herds increased as can be seenin Fig. 1 (source: Animal Health Service, The Netherlands).
The prevalence in The Netherlands was reduced in onlya few years (Fig. 2; source: Animal Health Service, The Neth-erlands). The eradication campaign was effective and TheNetherlands is nearly free from PRV, as the prevalence de-creased from nearly 100% at the start to a value below 0.1%in 2000.
The article 9 status has been achieved: free from PRV, butstill vaccinating. The article 10 status is the ultimate goal ofthe eradication campaign: free from PRV and not vaccinating.However, after the devastating Classical Swine Fever epidemicin 1997, the product board is not very keen on getting this sta-tus. The reason is that, since Belgium is not free of PRV, therisk of introduction is high, and the control of a possible out-break of PRV would be very costly. Moreover, the acceptanceby the society of culling animals because of PRV is question-able. At this moment it is not clear what the strategy will be inthe near future.
0
10
20
30
40
50
60
70
80
90
100
96/4 97/1 97/2 97/3 97/4 98/1 98/2 98/3 98/4year/quarter
frequ
ency
(in
%)
sow herds sows
Fig. 1. Percentage of gE-negative sow herds.
Fig. 2. Prevalence of gE-positive farms.

245A. Bouma / Biologicals 33 (2005) 241e245
References
[1] Pensaert MB, Kluge JP. Pseudorabies (Aujeszky’s disease). In:
Pensaert MB, editor. Virus infections of porcines. Amsterdam, The Neth-
erlands: Elsevier Science Publishers; 1989. p. 39e64.[2] Stegeman A, De Jong MCM, Van Nes A, Bouma A. Dynamics of pseu-
dorabies virus infections in vaccinated pig populations: a review. Vet Q
1997;19:117e22.
[3] Van Oirschot JT. Present and future of veterinary viral vaccinology: a re-
view. Vet Q 2001;23:100e8.
[4] Van Oirschot JT. Bovine viral vaccines, diagnostics, and eradication:
past, present, and future. Adv Vet Med 1999;41:197e216.[5] Van Oirschot JT, Houwers DJ, Rziha HJ, Moonen PJLM. Development of
an ELISA for the detection of antibodies to glycoprotein I of Aujeszky’s
disease virus: a method for the serologic differentiation between infected
and vaccinated pigs. J Virol Methods 1988;22:191e206.[6] De Leeuw PW, Van Oirschot JT. Vaccines against Aujeszky’s disease:
evaluation of their efficacy under standardized laboratory conditions.
Vet Q 1985;7:191e7.
[7] Pensaert MB, De Smet K, De Waele K. Extent and duration of virulent
virus excretion upon challenge of pigs vaccinated with different glyco-
protein-deleted Aujeszky’s disease vaccines. Vet Microbiol 1990;22:
107e17.[8] Vannier P, Hutet E, Cariolet R. Levels of virulent virus excreted by
infected pigs previously vaccinated with different glycoprotein deleted
Aujeszky’s disease vaccines. Vet Microbiol 1991;29:213e23.
[9] De Jong MCM, Bouma A. Herd immunity after vaccination: how
to quantify it and how to use it to halt disease. Vaccine 2001;19:
2722e8.
[10] Zuffa A, Salaj J, Cernik K. Immunity of pigs vaccinated by live or inac-
tivated Aujeszky vaccines against experimental infection. Zentralbl
Veterinarmed B 1982;29:663e75.
[11] Maes L, Pensaert M. Examination for virus persistence on swine fatten-
ing and breeding farms after an outbreak of Aujeszky’s disease. Tijdschr
Diergeneesk 1984;109:439e45.
[12] Thawley DG, Solorzano RF, Johnson ME. Confirmation of pseudorabies
virus infection, using virus recrudescence by dexamethasone treatment
and in vitro lymphocyte stimulation. Am J Vet Res 1984;47:2099e103.[13] Williams PP, Sacks JM, Yang PC, Pirtle EC, Erickson GA, Beran GW.
Immunological surveillance in a pseudorabies quarantine herd using gilts
and their progeny as sentinels. Viral Immunol 1989;2:185e93.
[14] Van Oirschot JT, Daus F, Kimman TG, Van Zaane D. Antibody response
to glycoprotein I in maternally immune pigs exposed to a mildly virulent
strain of pseudorabies virus. Am J Vet Res 1991;52:1788e93.
[15] Motha MJX, Horner GW, Tham KM, Ralston JC. A comparison of the
efficacy of two commercial Aujeszky’s disease vaccines with glycopro-
tein-I deletion in pigs. N Z Vet J 1993;41:1e6.
[16] Donaldson AI, Wardley RC, Martin S, Ferris NP. Experimental Aujesz-
ky’s disease in pigs: excretion, survival and transmission of the virus.
Vet Rec 1983;113:490e4.
[17] Gillespie RR, Hill MA, Kanitz CL. Infection of pigs by aerosols of Aujesz-
ky’s diseasevirus and their sheddingof virus.ResVet Sci 1996;60:228e33.
[18] Koopman JS, Longini IM. The ecological effects of individual exposure
and nonlinear disease dynamics in populations. Am J Public Health
1994;84:836e41.[19] Diekmann O, Heesterbeek JAP, Metz JAJ. On the definition and compu-
tation of the basic reproduction ratio R0 in models for infectious diseases
in heterogeneous populations. J Math Biol 1990;28:365e82.[20] Anderson RM, May RM. Infectious diseases of humans: dynamics and
control. Oxford, UK: Oxford University Press; 1991.
[21] De Jong MCM. Mathematical modelling in veterinary epidemiology:
why model building is important. Prev Vet Med 1995;25:183e93.[22] Becker NG. Analysis of infectious data. London, UK: Chapman & Hall;
1989. p. 139e74.
[23] Van Oirschot JT, Gielkens ALJ, Moormann RJM, Berns AJM. Marker
vaccines, virus protein-specific antibody assays and the control of Au-
jeszky’s disease. Vet Microbiol 1990;23:85e101.
[24] De Smet K, De Waele K, Pensaert M. Virulent Aujeszky’s disease virus
infectious dose in vaccinated and non-vaccinated pigs. In: Proceedings of
the 12th international pig veterinary society congress. The Hague, The
Netherlands; 1992. p. 65.
[25] Halloran ME, Haber ML, Longini IM, Struchiner CJ. Direct and indirect
effects in vaccine efficacy and effectiveness. Am J Epidemiol 1991;133:
323e31.
[26] Anderson RM. The concept of herd immunity and the design of commu-
nity-based immunization programmes. Vaccine 1992;10:928e35.
[27] De Jong MCM, Kimman TG. Experimental quantification of vaccine-
induced reduction in virus transmission. Vaccine 1994;12:761e6.
[28] Bouma A, De Jong MCM, Kimman TG. The influence of maternal im-
munity on the transmission of pseudorabies virus and on the effectiveness
of vaccination. Vaccine 1997;15:287e94.
[29] Bouma A, De Jong MCM, Kimman TG. Comparison of two pseudora-
bies virus vaccines, that differ in capacity to reduce virus excretion after
a challenge infection, in their capacity of reducing transmission of pseu-
dorabies virus. Vet Microbiol 1997;54:113e22.
[30] Bouma A, De Jong MCM, Kimman TG. Transmission of two pseudora-
bies virus strains, that differ in virulence and virus excretion, in groups of
vaccinated pigs. Am J Vet Res 1996;57:43e7.[31] Van Nes A, Stegeman JA, De Jong MCM, Loeffen WLA, Kimman TG,
Verheijden JHM. No major outbreaks of pseudorabies virus in well-im-
munised sow herds. Vaccine 1996;14:1042e4.
[32] Stegeman JA, Van Nes A, De Jong MCM, Bolder WMM. Assessment of
the effectiveness of vaccination against pseudorabies in finishing pigs.
Am J Vet Res 1995;56:573e8.
[33] Van Nes A, De JongMCM, Schoevers EJ, Van Oirschot JT, Verheijden JH.
Pseudorabies virus is transmitted among vaccinated conventional pigs, but
not among vaccinated SPF pigs. Vet Microbiol 2001;80:303e12.
[34] Van Nes A. Mathematical modelling of pseudorabies virus (syn. Aujesz-
ky’s disease virus) outbreaks aids eradication programmes: a review. Vet
Q 2001;23:21e6.



![Types of vaccines 1 - First generation vaccines are whole-organism vaccines - either live and weakened, or killed forms. [1] Live, attenuated vaccines,](https://img.pdfslide.net/doc/110x75/56649e155503460f94afff0b/types-of-vaccines-1-first-generation-vaccines-are-whole-organism-vaccines.jpg)

























