Embed Size (px)
Citation preview

Plant Ecology138: 137–147, 1998.© 1998Kluwer Academic Publishers. Printed in the Netherlands.
137
Development of black spruce growth forms at treeline
Daria Pereg & Serge PayetteCentre d’etudes nordiques, Universit´e Laval, Ste-Foy, QC, Canada, G1K 7P4
Received 4 November 1997; accepted in revised form 6 May 1998
Key words:Climate change, Dendrochronology, Forest tundra,Picea mariana, Stem analysis
Abstract
Most treeline populations in northeastern Canada are monospecific stands of black spruce (Picea mariana[Mill.]B.S.P.), a hardy, cold-tolerant species able to withstand harsh climatic conditions under different growth forms.In the forest tundra, black spruce thrives in protected areas and exhibits a normal arborescent growth form, butin exposed sites, upright stems are damaged above the snowpack by snow abrasion and wind. In this study, thedevelopment of damaged growth forms was examined in a moderately exposed habitat. Five developmental stageswere identified and described using detailed stem analysis of 13 spruce trees. Four different types of damagedgrowth forms were identified according to variations in supra-nival (above snow) stem height and number. Atthe site scale, the age structure of supra-nival shoots, based on a larger sample of 256 stems, was unimodal,suggesting a synchronous development of the spruce stand in which 46% of the shoots were initiated during the1960s and 1970s. Subfossil trunks on the ground were all depressed trees, indicating that the former vegetationwas a krummholz, not a forest. This indicates the recent development of the small-tree stand above the snowpack,probably triggered by recent milder conditions associated with snowier winters in the last decades.
Introduction
Across the circumpolar zone, treeline trees are ex-posed to harsh conditions and are often pushed to theirultimate tolerance limits (Savile 1972; Marchand &Chabot 1978; Marchand 1987). These extreme condi-tions are responsible for the discontinuous distributionof forest stands so characteristic of the forest-tundra.Most trees are confined to valley bottoms and otherprotected sites while exposed hilltops are covered bylichen-heath vegetation (Arseneault & Payette 1992).
In northern Québec (Canada), treeline sites aredominated by monospecific stands of black spruce(Picea mariana[Mill.] B.S.P.), a cold tolerant speciesable to survive as a shrub and to reproduce by lay-ering (clonal growth) when conditions are inimical tovertical growth. Black spruce also exhibits a large phe-notypic plasticity expressed by different growth forms(Figure 1) distributed along a wind exposure gradient(Payette et al. 1996) enabling the species to survivein a large array of habitats. Whereas in protectedsites black spruce develops a normal (symmetrical)
arborescent form, in exposed areas the trees are dam-aged by windblown snow and ice (Hadley & Smith1983; 1986, 1989; Marchand 1987; Payette et al.1996) and stems are defoliated above the snowpack.Also, vegetative buds are often killed by deep frostunder windchill conditions, hence triggering reitera-tive processes inducing the formation of new shoots(Bégin 1991). Reiteration consists in the transfer ofapical dominance from a dead or dying apical meris-tem to one or several latent lateral buds. Vegetativebuds will develop to form new shoots for the replace-ment of the original, moribund stem (Édelin 1981,1986; Barthélémy et al. 1989). Defoliation and re-iteration processes generate single-stemmed (skirted)or multi-stemmed (whorled) tree forms. The latterare characterized by heavily branched basal cushionscovered by healthy foliage below the snowpack, andstems that are progressively defoliated at their centralpart by wind erosion above the snowpack (Scott et al.1987a,b; Payette et al. 1989; Bégin 1991; Payette et al.1996). In the most exposed sites, black spruce thrivesas krummholz mats with no surviving stems above

138
Figure 1. Black spruce growth forms at treeline along a wind exposure gradient.
the snowpack because of severe snowdrift conditionspreventing vertical growth.
The wind exposure of a given site depends ontopography and vegetation, because the latter offer acertain degree of wind protection. Climatic conditionsstongly affect vegetation growth and thus, wind ex-posure and climatic conditions are related. Becausedamaged growth forms are directly related to site ex-posure and climatic conditions, they have been usedas bioindicators of past climatic variations. A mod-erate change in these conditions may trigger a shiftof dominant growth forms in a given stand from ar-borescent to depressed forms as evidenced by tree-ringanalysis of subfossil spruce stems (Payette et al. 1985,1989; Lavoie & Payette 1992, 1994). The most impor-tant shift in growth-form dominance occurred duringthe Little Ice Age (1580–1850 AD), a climatic periodcharacterized by drier winters and colder tempera-tures (Grove 1988). Since the end of the Little IceAge, warming was observed over the northern hemi-sphere with decadal variations of 0.1◦C, though withregional differences (Jones et al. 1986; Folland et al.1990; Jones & Briffa 1992; Briffa & Jones 1993).Increased stem growth above the snowpack was re-
ported in several spruce stands at tree-line in northernQuébec (Payette et al. 1985, 1989; Lavoie & Payette1992, 1994; Lescop-Sinclair & Payette 1995) and inthe Swedish Scandes (Kullman 1986), but in the lattercase, a recent cooling and recession of stem growthwas later observed (Kullman 1996).
Given the usefulness of treeline black spruces asbioindicators of climatic changes, it is important toinvestigate thoroughly the development of its differ-ent growth forms, in order to better understand themechanisms underlying the use of this bioindicator.Conversely, the first objective of this study was to ana-lyze the developmental patterns of damaged tree formsin moderately exposed sites, and the second objectivewas to examine the stand dynamics of a site apparentlyshowing a recent increase in stem growth.

139
Methods
Study site
A treeline black spruce stand dominated by damagedtree forms was selected about 35 km east of the Hud-son Bay coast (Figure 2). The spruce population waslocated on a hillside moderately exposed to westerlywinds (alt. ≈ 150 m) in the Rivière Boniface re-gion (57◦45’ N; 76◦20’ W). Climatic data from theInukjuak weather station (150 km north-west of thestudy site) indicate that the mean annual temperatureis −7◦C, ranging from−26◦C (in February) to 9◦C(in July) with a mean frost-free period of about 60days. Annual precipitation totals 400 mm, from which35% falls as snow (Environnement Canada 1989).The growing season extends from mid-June to mid-August. At the study site, the last fire occurred at1300 years B.P. as deduced from radiocarbon-datedcharcoal (Payette, unpubl. data). The vegetation coveris dominated by black spruce clones, lichens (Clad-ina and Cladonia), and shrubs (Betula glandulosa,Salix, LedumandVaccinium). The climatic data fromInukjuak weather station highly correlates with themore recent data obtained from two weather stationslocated in the Rivière Boniface region since 1988(R2 = 0.95, p < 0.0001) (Wang & Payette 1996),according to temperature and snowfall data. The useof Inukjuak’s data seems warranted and provides dataranging from 1944 till today.
A quadrat (15 m× 15 m) was positioned at randomwithin the krummholz stand, and microtopographicalfeatures of the ground surface were assessed with anelectronic level to ensure that no exposure gradientcould affect the distribution of growth forms in thequadrat. Each stem was mapped according to spatialcoordinates along with snowpack depth, stem heightand number of shoots above snowpack. The snow-pack level was estimated using the contrasting foliagecolor of black spruce (darker foliage below snow) andheight of dwarf birch (Betula glandulosa) which corre-sponds to the snowpack height in exposed sites. Thismethod of snowpack measurement is an estimate ofthe mean snow accumulation and is performed duringsummer using biological indicators. It involves a cer-tain level of imprecision due to the yearly variationsof the snowpack and it is admitted that its maximumprecision is approximately 10 cm. Unfortunately, theestimation of snow level height during summer canhardly be more precise on the field and this uncertaintyis accounted for in all further analyses.
Stem analysis
The developmental pattern of damaged growth formswas assessed using stem analysis of 13 trees sam-pled at random in the quadrat. The 13 trees weredrawn to scale and morphological features, such asfoliage quality and presence of scars or stem bend-ings indicating former reiterative events, were noted.Cross-sections were taken every 5 cm along the stemand all leaf biomass 2.5 cm above and below eachcross-section’s upper surface was collected. Stem sec-tions were sanded for dendrochronological analysis(Fritts & Swetnam 1989) and tree-ring counts andcrossdating were performed to ensure proper datingof the samples using light rings (Filion et al. 1986).Ring widths were measured under a binocular micro-scope at 40× with a Henson micrometer (±0.1 mm)interfaced with a computer to record the data. Rawdata was mathematically standardized to account forthe negative exponential growth trend in ring measure-ment caused by the geometrical constraint inherent totree growth (Fritts & Swetnam 1989). Stem analysisdiagrams were constructed following a recently de-scribed method (Payette & Delwaide 1994; Payetteet al. 1994, 1996). Ring width measurements of eachtree were grouped in five growth categories (with classintervals of 0.5σ ), with the two end categories gath-ered in one ‘strong growth’ category (>1.5σ ) and one‘weak growth’ category (<1.5σ ) and the three remain-ing categories centered around the mean. The growthdata were integrated in diagrams illustrating the spa-tial and temporal distribution of annual ring growth atevery 5 cm along the stem. Periods of increased anddecreased radial growth were identified for the 13 treesand a developmental model for damaged growth formswas constructed.
Stand dynamics
Stand development associated with recent climaticvariations was assessed based on growth form devel-opment. Four types of damaged growth forms wereidentified according to stem height above snowpack(arborescent:≥1 m and sub-arborescent:<1 m) andnumber of shoots above snowpack (single-stemmedor multi-stemmed). Stem defoliation was evaluated(percentage of whole stem without leaves) and yearof stem initiation above the snowpack (stem age) wasassessed by counting the age of cross-sections at thesnowpack line. A two-way MANOVA analysis wasused to verify the interaction between stem height andshoot number above snow level, and to determine the
140
Figure 2. Northern Quebec’s vegetation zones and location of study site.
contribution of each of these parameters in the defolia-tion and age differences observed between the groups(Sokal & Rohlf 1981). The distribution of damagedgrowth forms within the stand was then evaluated ac-cording to the four types previously described. Theage structure of supra-nival shoots was used to identifyperiods of shoot initiation and favorable growth abovethe snowpack. Stand dynamics was then comparedwith regional climatic data (Inukjuak weather station),but given the imprecision related to snow level estima-tion, and consequently to the date of stem initiation,the weather data were not statistically treated to relatethem to stand dynamics data.
Results
Developmental pattern of damaged tree forms
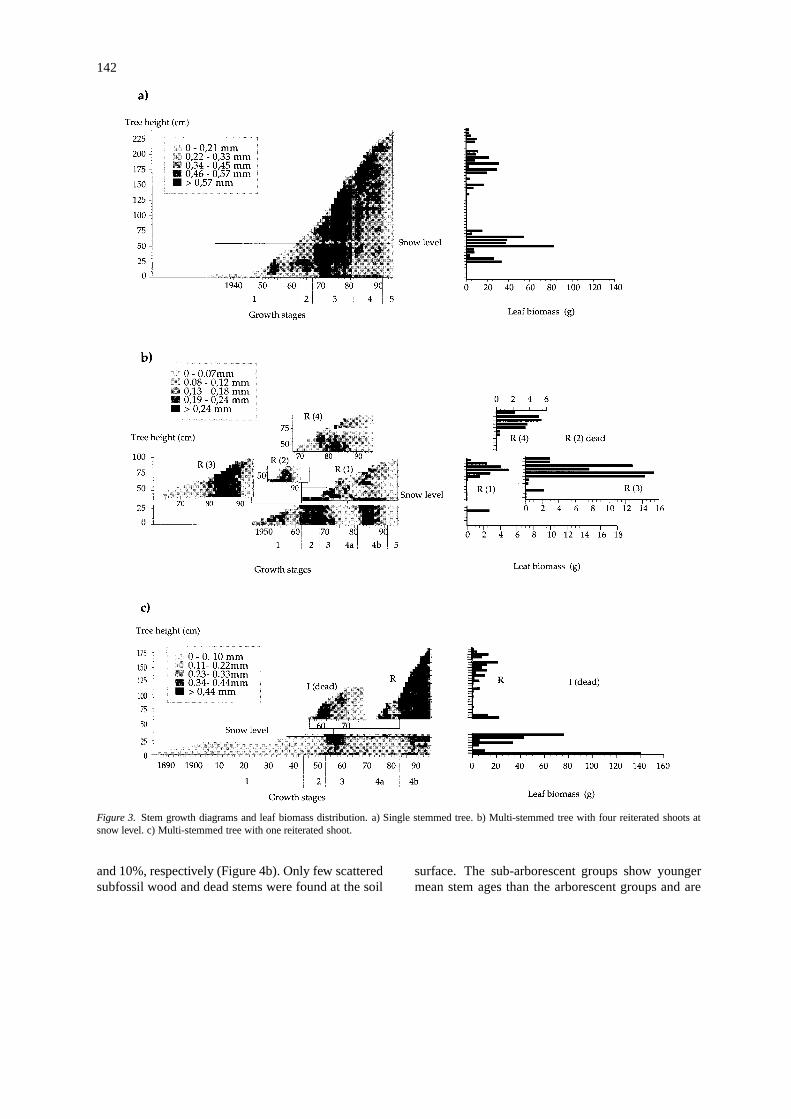
Thirteen trees were compared from diagrams of stemanalysis to decipher the pattern of growth form degra-dation under snowdrift conditions. Representativesingle-stemmed and multi-stemmed tree forms areshown by squares of different shades (Figure 3). Thehorizontal axis represents time in years (life span ofthe tree) and the vertical axis, tree height. Each squarecorresponds to a specific, annual ring growth (onetree ring measurement) at a given height along the

141
Table 1. Developmental pattern of damaged growth forms as identified with stem analysis
stem: the darker the square, the stronger the growth.The developmental patterns of stem growth character-ized by common growth periods in the diagrams aresummerized in Table 1.
The first stage (1) corresponds to variable ra-dial and vertical growth below the snowpack. Thisstage outlines the early development of small, snow-protected stems not yet affected by snowdrift condi-tions. The second stage (2) corresponds to positionof the stem’s apex at the snow-air interface showingreduced radial and vertical growth rates. The survivaland growth of the apical meristem above the snow-airinterface produces single-stemmed trees (Figure 3a).At this stage, multi-stemmed trees are also formedat the snow-air interface when reiterative processesare triggered following death of the apical meristem(Figure 3b). After their passage across the snow-airinterface, the developing stems (I : original orR: reit-erated) experience a period of rapid growth, i.e., theelongation stage (3), preceeding the degradation (4)and decline (5) stages. The elongation stage is char-acterized by strong growth throughout the stem, asshown by black squares in the diagrams (Figure 3).Stem degradation begins near the snowpack level,
where windblown snow abrasion is strongest, withdecreasing radial growth in stem parts most affectedby defoliation. This is well illustrated in the diagramsby the correspondance between foliage loss and de-creased radial growth along the stems. During thedegradation stage, multi-stemmed trees are created byreiterative processes (Figure 3c). The original stem (I )is being damaged during the first years of degradation(4a) and apical dominance is transferred to a branchbelow the snowpack. The radial and vertical growth ofthe reiterated shoot (R) is faster and replaces the dy-ing original stem. Then the reiterated shoot becomesdefoliated (4b) and may be replaced or not, follow-ing the same process. Decline (5) is characterized bythe pronounced, overall decrease in radial growth notcompensated by other reiterated shoots.
Stand dynamics
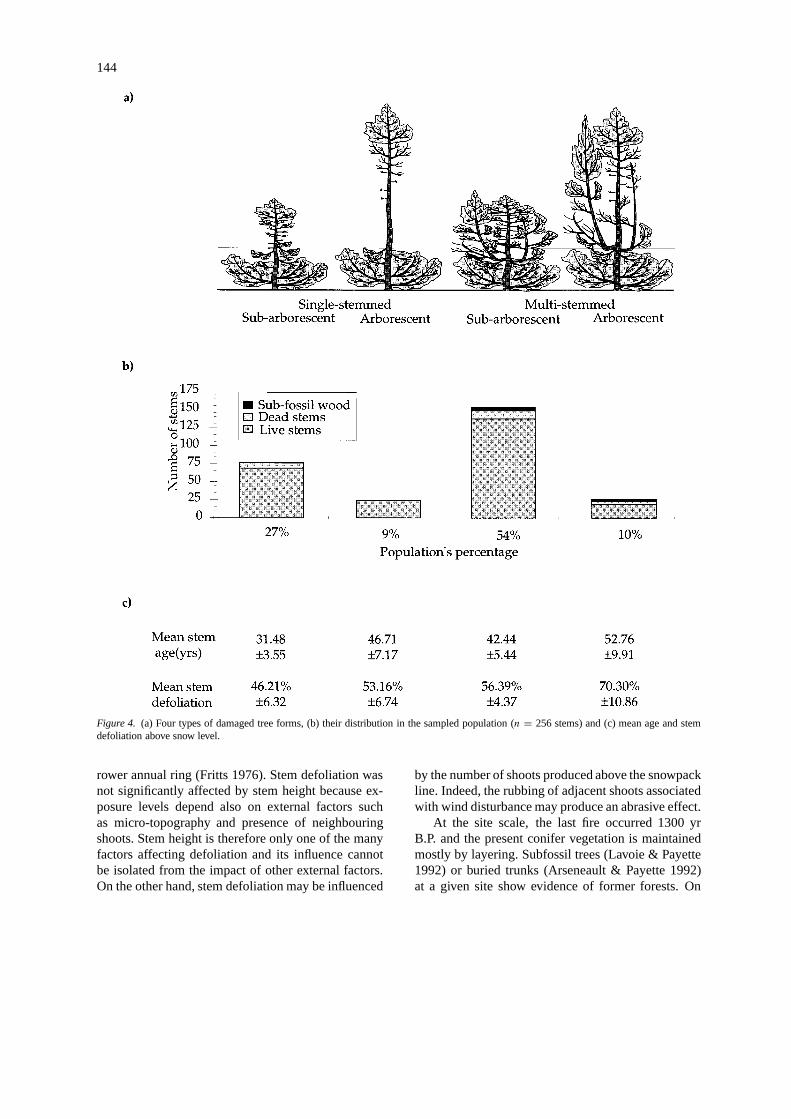
The distribution (%) of the stems in the four groupsof damaged growth forms previously described (Fig-ure 4a) indicates the dominance of sub-arborescent,multi-stemmed (54%) and single-stemmed (27%)trees in the stand, while arborescent multi-stemmedand single-stemmed trees are less frequent, i.e., 9%
142
Figure 3. Stem growth diagrams and leaf biomass distribution. a) Single stemmed tree. b) Multi-stemmed tree with four reiterated shoots atsnow level. c) Multi-stemmed tree with one reiterated shoot.
and 10%, respectively (Figure 4b). Only few scatteredsubfossil wood and dead stems were found at the soil
surface. The sub-arborescent groups show youngermean stem ages than the arborescent groups and are

143
generally less defoliated (Figure 4c). There is a signif-icant influence of stem height (p = 0.0001) and stemnumber (p = 0.0016) on stem defoliation and age,but no significant interaction was found between thetwo factors (p = 0.2340). Stem defoliation was notsignificantly affected by stem height (p = 0.0605) butit was influenced by the number of shoots producedabove snow level (p = 0.0008). Stem age was mostrelated to stem height (p = 0.0001) and not to stemnumber above snow level (p = 0.0740) (Two-wayMANOVA, α = 0.05).
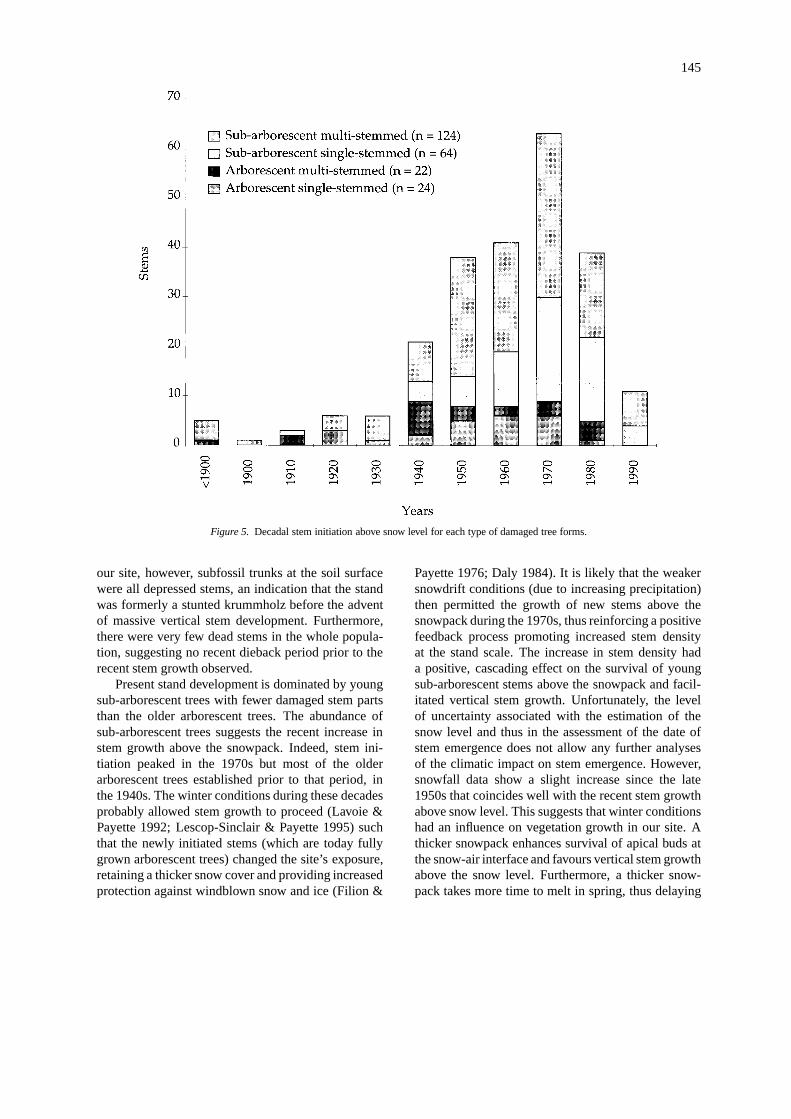
Stem emergence above snow was initiated dur-ing the late 1930s and 1940s for arborescent trees(Figure 5). Stem initiation mainly formed by youngsub-arborescent trees peaked in the 1970s. The liveperiod of the few dead stems in the stand was similarto that of the living stems, suggesting no particulardieback period. Furthermore, subfossil trunks on theground were all prostrate growth forms, indicating thatthe former vegetation was a krummholz stand, not aforest.
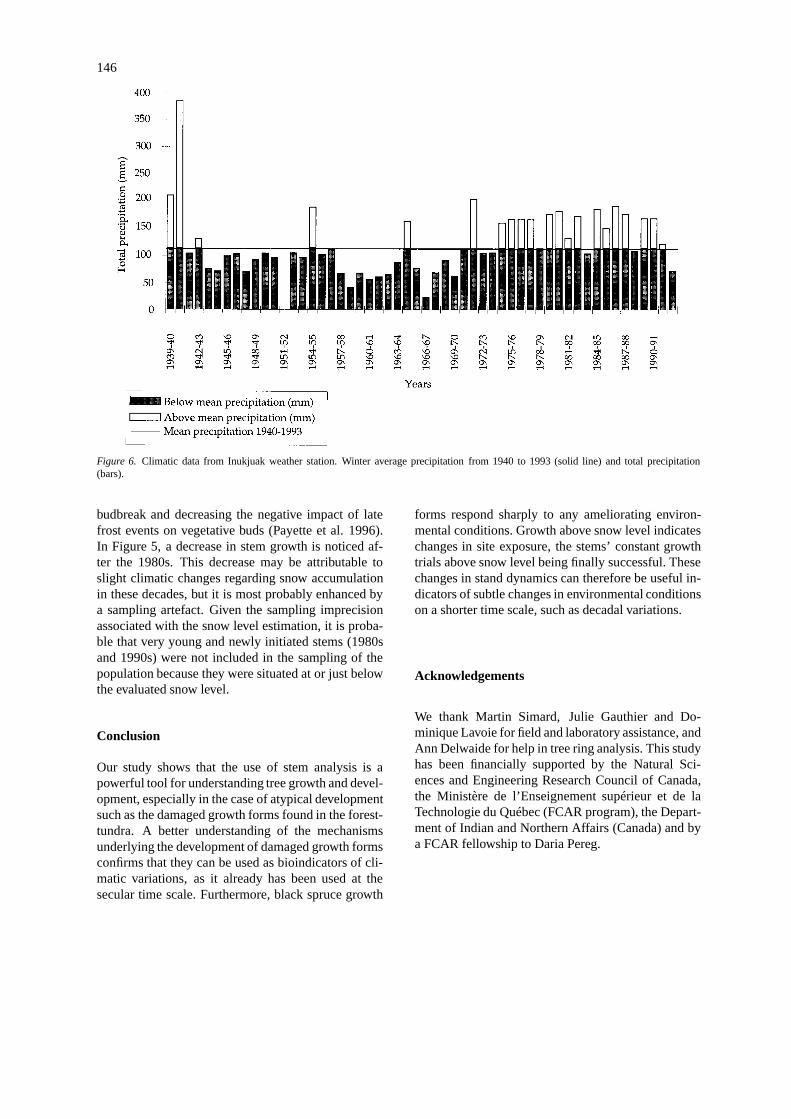
This suggests the recent development of the stempopulation above the snowpack. The stand develop-ment through the mediation of stem growth abovethe snowpack may be related to climatic conditionsprevailing in the nearby weather station at Inukjuak(Figure 6). Winter data reveal slightly milder andsnowier winters since the 1970s as shown by a totalsnow accumulation constantly above the mean (1944–1994). These slight changes in winter conditions wereapparently favourable to stem growth above the snow-pack and coincided well with the increase in stemgrowth observed in the 1970s. However, no pro-nounced trends were observed in the recent decadesconcerning weather conditions, and it appears diffi-cult to obviously relate the recent change in standdynamics shown by our data to the decadal climaticvariations.
Discussion
During the first stage of development, layeringbranches adopt an upright position and gradually be-come independant stems (Légère & Payette 1981).This process slows down stem growth below the snow-pack, even though the stem is not yet influenced bysnowdrift conditions. This stage lasts until the stemreaches the snow-air interface and enters into stage 2of the developmental pattern. If snowdrift conditionsare severe enough to prevent stem growth above the
snowpack line, the tree develops as a depressed shrub(Hadley & Smith 1987; Payette et al. 1996) and, there-fore, will never reach stage 3 of the developmentalpattern. For single-stemmed individuals, the period ofgrowth decrease at the snowpack line is shorter andnot very pronounced (Figure 3a). For multi-stemmedtrees experiencing reiterative activities at the snow-air interface, the first developmental stage is longer(Figure 3c), and some trees may be affected by sev-eral unsuccessful reiterative trials before initiating newshoots above the snowpack (Figure 3b). When a stemis initiated above the snowpack line, it undergoes aperiod of strong growth until progressive defoliationtriggers the degradation stage (Payette et al. 1996).For single-stemmed trees, stem degradation above thesnowpack may cause stem decline (Figure 3a) or stemreplacement (Bégin 1991) leading to the formation ofmulti-stemmed trees (Figure 3c). The latter showed se-vere degradation of the initial stem (I ) until 1973, i.e.,the year during which a reiterated shoot was initiatedand exhibited fast and strong growth concomitantlywith the decline of the initial stem. For multi-stemmedtrees formed by reiterative processes at the snowpackline, degradation was synchronous for all stems orfavoured the growth of one stem to the detriment ofothers (Figure 3b). Stem age was most related to stemheight and not to the number of stems produced by oneindividual above snow level, which supports the factthat multi-stemmed trees may originate from reiteratedshoots formed either by young stems at snowpack lineor by old declining stems through the stem replace-ment process. Decline occurred when stem growthdecrease was not followed by the rise of a new shoot.
Stem analysis revealed that stages of strong growthwere associated with stem elongation before the oc-currence of foliage loss, either on initial or reiter-ated shoots. On the other hand, growth decrease isrelated to traumatic events such as bud death andreiterative activities (Bégin 1991), stem breakage ordefoliation (Payette et al. 1994, 1996). The relationbetween leaf loss and radial growth (Swetnam et al.1988; Jardon et al. 1994; Dvorak et al. 1996) wasoutlined by sustained growth decrease in most de-foliated stem parts (Payette et al. 1996). Our dataclearly shows this concordance when stem growthdiagrams are compared with leaf biomass. Neverthe-less, nutrient and water transport allow minimal radialgrowth in defoliated stem parts according to the pipemodel theory (Robichaud & Methven 1992), but thelesser need for water and nutrients may also reducexylem production, resulting in the formation of a nar-
144
Figure 4. (a) Four types of damaged tree forms, (b) their distribution in the sampled population (n = 256 stems) and (c) mean age and stemdefoliation above snow level.
rower annual ring (Fritts 1976). Stem defoliation wasnot significantly affected by stem height because ex-posure levels depend also on external factors suchas micro-topography and presence of neighbouringshoots. Stem height is therefore only one of the manyfactors affecting defoliation and its influence cannotbe isolated from the impact of other external factors.On the other hand, stem defoliation may be influenced
by the number of shoots produced above the snowpackline. Indeed, the rubbing of adjacent shoots associatedwith wind disturbance may produce an abrasive effect.
At the site scale, the last fire occurred 1300 yrB.P. and the present conifer vegetation is maintainedmostly by layering. Subfossil trees (Lavoie & Payette1992) or buried trunks (Arseneault & Payette 1992)at a given site show evidence of former forests. On
145
Figure 5. Decadal stem initiation above snow level for each type of damaged tree forms.
our site, however, subfossil trunks at the soil surfacewere all depressed stems, an indication that the standwas formerly a stunted krummholz before the adventof massive vertical stem development. Furthermore,there were very few dead stems in the whole popula-tion, suggesting no recent dieback period prior to therecent stem growth observed.
Present stand development is dominated by youngsub-arborescent trees with fewer damaged stem partsthan the older arborescent trees. The abundance ofsub-arborescent trees suggests the recent increase instem growth above the snowpack. Indeed, stem ini-tiation peaked in the 1970s but most of the olderarborescent trees established prior to that period, inthe 1940s. The winter conditions during these decadesprobably allowed stem growth to proceed (Lavoie &Payette 1992; Lescop-Sinclair & Payette 1995) suchthat the newly initiated stems (which are today fullygrown arborescent trees) changed the site’s exposure,retaining a thicker snow cover and providing increasedprotection against windblown snow and ice (Filion &
Payette 1976; Daly 1984). It is likely that the weakersnowdrift conditions (due to increasing precipitation)then permitted the growth of new stems above thesnowpack during the 1970s, thus reinforcing a positivefeedback process promoting increased stem densityat the stand scale. The increase in stem density hada positive, cascading effect on the survival of youngsub-arborescent stems above the snowpack and facil-itated vertical stem growth. Unfortunately, the levelof uncertainty associated with the estimation of thesnow level and thus in the assessment of the date ofstem emergence does not allow any further analysesof the climatic impact on stem emergence. However,snowfall data show a slight increase since the late1950s that coincides well with the recent stem growthabove snow level. This suggests that winter conditionshad an influence on vegetation growth in our site. Athicker snowpack enhances survival of apical buds atthe snow-air interface and favours vertical stem growthabove the snow level. Furthermore, a thicker snow-pack takes more time to melt in spring, thus delaying
146
Figure 6. Climatic data from Inukjuak weather station. Winter average precipitation from 1940 to 1993 (solid line) and total precipitation(bars).
budbreak and decreasing the negative impact of latefrost events on vegetative buds (Payette et al. 1996).In Figure 5, a decrease in stem growth is noticed af-ter the 1980s. This decrease may be attributable toslight climatic changes regarding snow accumulationin these decades, but it is most probably enhanced bya sampling artefact. Given the sampling imprecisionassociated with the snow level estimation, it is proba-ble that very young and newly initiated stems (1980sand 1990s) were not included in the sampling of thepopulation because they were situated at or just belowthe evaluated snow level.
Conclusion
Our study shows that the use of stem analysis is apowerful tool for understanding tree growth and devel-opment, especially in the case of atypical developmentsuch as the damaged growth forms found in the forest-tundra. A better understanding of the mechanismsunderlying the development of damaged growth formsconfirms that they can be used as bioindicators of cli-matic variations, as it already has been used at thesecular time scale. Furthermore, black spruce growth
forms respond sharply to any ameliorating environ-mental conditions. Growth above snow level indicateschanges in site exposure, the stems’ constant growthtrials above snow level being finally successful. Thesechanges in stand dynamics can therefore be useful in-dicators of subtle changes in environmental conditionson a shorter time scale, such as decadal variations.
Acknowledgements
We thank Martin Simard, Julie Gauthier and Do-minique Lavoie for field and laboratory assistance, andAnn Delwaide for help in tree ring analysis. This studyhas been financially supported by the Natural Sci-ences and Engineering Research Council of Canada,the Ministère de l’Enseignement supérieur et de laTechnologie du Québec (FCAR program), the Depart-ment of Indian and Northern Affairs (Canada) and bya FCAR fellowship to Daria Pereg.

147
References
Arseneault, D. & Payette S. 1992. A postfire shift from lichen-spruce to lichen-tundra vegetation at tree line. Ecology 73:1067–1081.
Barthélémy, D., Édelin C. & Hallé F. 1989. Architectural conceptsfor tropical trees. Pp. 89–100. In: Holm-Nielsen, L.B., NielsenI. C. & H. Baslev (eds), Tropical forest. Botanical dynamics,speciation and diversity. Academic Press Ltd.
Bégin, C. 1991. Analyse architecturale et dendroécologique d’unepessière à lichen à la limite des forêts. PhD Thesis, UniversitéLaval, Ste-Foy.
Briffa, K. R. & Jones P. D. 1993. Global surface air temperaturevariation during the twentieth century: Part 2, implications forlarge scale high frequency palaeoclimatic studies. The Holocene3: 77–88.
Daly, C. 1984. Snow distribution patterns in the alpine krummholzzone. Prog. Phys. Geogr. 8: 157–175.
Dvorak, V., Oplustilova M. & Janous D. 1996. Relation between leafbiomass and annual ring sapwood of Norway spruce according toneedle age-class. Can. J. For. Res. 26: 1822–1827.
Édelin, C. 1981. Quelques aspects de l’architecture végétative desconifères. Bull. Soc. Bot. Française - Lettres Bot. 128: 177–188.
Édelin, C. 1986. Stratégie de réitération et édification de la cimechez les conifères. Pp. 139–158. Colloque international surl’arbre. Montpellier.
Environnement Canada. 1989. Sommaire climatique et donnéesmétéorologiques de la station d’Inukjuak, Québec, Canada.
Filion, L. & Payette S. 1976. La dynamique de l’enneigementen région hémi-arctique, Poste-de-la-Baleine, Nouveau-Québec.Cahiers Géogr. Québec 20: 275–302.
Filion, L., Payette S., Gauthier L. & Boutin Y. 1986. Light rings insubarctic conifers as a dendrochronological tool. Quat. Res. 26:272–279.
Folland, C. K., Karl T. R. & Vinnikov K. Y. A. 1990. Observedclimate variations and changes. Pp. 195–239. In Houghton, J. T.,G. J. Jenkins & J. J. Ephraums (eds), Climate change: The IPCCscientific assesment. Cambridge University Press, New York.
Fritts, H. C. 1976. Tree rings and climate. Academic Press, London.Fritts, H. C. & Swetnam T. W. 1989. Dendroecology: A tool for
evaluating variations in past and present forest environments.Adv. Ecol. Res. 19: 111–188.
Grove, J. M. 1988. The Little Ice Age. Methuen & Co. Ltd., London.Hadley, J. L. & Smith W. K. 1983. Influence of wind exposure
on needle dessication and mortality for timberline conifers inWyoming, USA. Arctic Alpine Res. 15: 127–135.
Hadley, J. L. & Smith W. K. 1986. Wind effects on needles oftimberline conifers: seasonal influence on mortality. Ecology 67:12–19.
Hadley, J. L. & Smith W. K. 1987. Influence of krummholz matmicroclimate on needle physiology and survival. Oecologia 73:82–90.
Hadley, J. L. & Smith W. K. 1989. Wind erosion of leaf surface waxin alpine timberline conifers. Arctic Alpine Res. 21: 392–398.
Jardon, Y., Filion L. & Cloutier C. 1994. Long-term impact of insectdefoliation on growth and mortality of eastern larch in borealQuébec. Écoscience 1: 231–238.
Jones, P. D. & Briffa K. R. 1992. Global surface air temperaturevariations during the twentieth century: Part 1, spatial, temporaland seasonal details. The Holocene 2: 165–179.
Jones, P. D., Wigley T. M. L. & Wright P. B. 1986. Globaltemperature variations between 1861 and 1984. Nature 322:430–434.
Kullman, L. 1986. Recent tree-limit history ofPicea abiesin thesouthern Swedish Scandes. Can. J. For. Res. 16: 761–771.
Kullman, L. 1996. Recent cooling and recession of Norway spruce(Picea abies(L.) Karst.) in the forest-alpine tundra ecotone ofthe Swedish Scandes. J. Biog. 23: 843–854.
Lavoie, C. & Payette S. 1992. Black Spruce growth forms as arecord of a changing winter environment at treeline, Québec,Canada. Arctic Alpine Res. 24: 40–49.
Lavoie, C. & Payette S. 1994. Recent fluctuations of the lichen-spruce forest limit in subarctic Québec. J. Ecol. 82: 725–734.
Légère, A. & Payette S. 1981. Ecology of a black spruce (Piceamariana) clonal population in the hemiarctic zone, northernQuebec: population dynamics and spatial development. ArcticAlpine Res. 13: 261–276.
Lescop-Sinclair, K. & Payette S. 1995. Recent advance of the arctictreeline along the eastern coast of Hudson Bay. J. Ecol. 83: 929–936.
Marchand, P. J. 1987. Life in the cold. An introduction to winterecology. University Press of New England, Hanover.
Marchand, P. J. & Chabot P. F. 1978. Winter water relations af tree-line plant species of Mt. Washington, New Hampshire. ArcticAlpine Res. 10: 105–116.
Payette, S. & Delwaide A. 1994. Growth of black spruce at its north-ern range limit in arctic Quebec, Canada. Arctic Alpine Res. 26:174–179.
Payette, S., Delwaide A., Morneau C. & Lavoie C. 1994. Stemanalysis of a long lived black spruce clone at treeline. ArcticAlpine Res. 26: 56–59.
Payette, S., Delwaide A., Morneau C. & Lavoie C. 1996. Patterns oftree stem decline along a snow-drift gradient at tree line: a casestudy using stem analysis. Can. J. Bot. 74: 1671–1683.
Payette, S., Filion L., Delwaide A. & Bégin C. 1989. Reconstruc-tion of tree-line vegetation response to long-term climate change.Nature 341: 429–432.
Payette, S., Filion L., Gauthier L. & Boutin Y. 1985. Secular climatechange in old-growth tree-line vegetation of northern Quebec.Nature 315: 135–138.
Robichaud, E. & Methven I. R. 1992. The applicability of the pipemodel theory for the prediction of foliage biomass in trees fromnatural, untreated black spruce stands. Can. J. For. Res. 22:1118–1123.
Savile, D. B. O. 1972. Arctic adaptations in plants. AgricultureCanada Monograph #6.
Scott, P. A., Bentley C. V., Fayle D. C. F. & Hansell R. I. C. 1987a.Crown forms and shoot elongation of white spruce at the treeline,Churchill, Manitoba, Canada. Arctic Alpine Res. 19: 175–186.
Scott, P. A., Hansell R. I. C. & Fayle D. C. F 1987b. Establishmentof white spruce populations and responses to climatic change atthe treeline, Churchill, Manitoba, Canada. Arctic Alpine Res. 19:45–51.
Sokal, R. R. & Rohlf F. J. 1981. Biometry. The principles andpractice of statistics in biological research. W.H. Freeman andCompany, New York.
Swetnam, T. W., Thompson M. A. & Kennedy Sutherland E. 1988.Using dendrochronology to measure radial growth of defoliatedtrees. United States Dept. of Agriculture, Forest Service Co-operative State Research Service, Agriculture Handbook #639,Washington D.C.
Wang, L. & Payette, S. 1996. Tree ring density of black spruceat treeline (subarctic Quebec). Ecological Society of Americaannual meeting. Providence, Rhode Island.






























