Embed Size (px)
Citation preview

Development of Independence from the Motherin Gorilla gorilla gorilla
Angela A. Nowell & Alison W. Fletcher
Received: 8 April 2005 /Revised: 25 January 2006 /Accepted: 10 April 2006 /Published online: 2 May 2007# Springer Science + Business Media, LLC 2007
Abstract In gorillas, the mother takes full responsibility for carrying and feedingoffspring, and lactational amenorrhea prevents her from investing in anotheroffspring while suckling. Therefore, mothers should encourage immature gorillasto become gradually more independent until they are physically able to acquiresufficient resources, travel independently, and manage social relationships unaided.We investigated the development of independence in a population of wild westerngorillas (Gorilla gorilla gorilla) at Mbeli Bai, Nouabalé-Ndoki National Park,Republic of Congo. We observed the spatial proximity, the development of travel,patterns of suckling and weaning, and the nature of mother-offspring interactions for42 immature gorillas to investigate the nature of the relationship between mother andoffspring. Though mother-offspring distance and independent travel increased duringimmaturity, with few active interactions between the dyads, offspring still continuedto suckle until a median age of 4.6 yr, longer than in mountain gorillas. Thetolerance of the mother toward suckling by older offspring may reflect theimportance of continued nutritional investment: the temporal nature of the diet ofwestern gorillas means that succulent fruits—likely to be preferred weaning foods—are not always available. We propose that to buffer against potential fruit shortage,mothers continue to nurse their offspring to decrease the likelihood of death fromcare-dependent factors and to improve an offspring’s chances of survival.
Keywords gorilla . independence . infant development . maternal investment
Int J Primatol (2007) 28:441–455DOI 10.1007/s10764-007-9128-6
A. A. Nowell : A. W. Fletcher (*)Department of Biological Sciences, University of Chester, Parkgate Road, Chester, CH1 4BJ, UKe-mail: [email protected]
A. A. NowellMbeli Bai Study, Nouabalé-Ndoki National Park, B.P. 14537 Brazzaville, Republic of Congo

Introduction
In mammals, postpartum investment in offspring reduces chances of offspring deathfrom care–dependent factors (sensu Barrett et al. 2006) and consequently willindirectly increase the reproductive success of the parents (Trivers 1972). Primatesspend a longer period growing and maturing than other mammals with similar bodysizes (Harvey and Clutton-Brock 1985; Kappeler et al. 2003), and the period duringwhich offspring are dependent on the mother and rely on her care is comparablylonger in great apes than in other primates (Watts and Pusey 1993). The length of theperiod may reflect the effort required to learn the behavioral and social skillsnecessary to survive in complex and dynamic environments (Watts and Pusey 1993).Maternal investment in great apes is high, and the mother takes virtually fullresponsibility for feeding and carrying offspring (Delgado and van Schaik 2000;Watts 1996). Mothers also often act as mediators in their infants’ initial interactionswith other members of the species (Fletcher 2001; Horvat and Kraemer 1981;Nicolson 1987). Young primates do not achieve independence from the mother untilthey have sufficiently matured physically and behaviorally to enable them to meetthe demands of their environment.
Offspring sex is one of several factors that influence the age at which immatureindividuals achieve independence from the mother. Researchers expect mothers toinvest most heavily in the sex that experiences the greatest variability in reproductivesuccess (Trivers 1972), though tests of the hypothesis have produced conflictingresults (Berman 1988; Meikle et al. 1984; Nevison et al. 1996). Parity of the mothercan also influence the age at which immature individuals gain independence:primiparous mothers engage in more infant handling and are more restrictive of theirinfants’ movements (Brown and Dixson 2000; Manson 1999); other researchers havereported them to be more neglectful of their infants (Maestripieri and Carrol 1998;Nakamichi et al. 2004). Furthermore, the age and condition of the mother herself canalso influence the way she behaves toward her offspring (Nowell 2005; van Lawick-Goodall 1971).
Gorilla neonates are relatively helpless (Fossey 1979), and infants must develop theskills that will allow them to travel independently, find and process sufficient food,and integrate into or form a social group in which to reproduce. Thus, in addition tothe social environment in which a gorilla matures, the ecological environment mayaffect the speed with which immature gorillas achieve independence from the mother.Western gorillas (Gorilla gorilla gorilla) are seasonally frugivorous, incorporatingfruits into the diet when they are available (Remis 1997; Rogers et al. 2004), resultingin increased daily path lengths reflecting the greater costs of feeding on sparselydistributed and ephemerally available food items (Cipolletta 2004; Doran et al. 2004).During periods of fruit scarcity, individuals feed on greater quantities of more fibrousand less proteinaceous leaves and stems, and sweet, succulent seasonal fruits arereplaced by more fibrous fruits that are otherwise ignored (Doran et al. 2002; Remis1997; Rogers et al. 2004). Conversely, Visoke mountain gorillas (Gorilla beringeiberingei) are relatively specialized folivores that feed very selectively on a smallnumber of high-quality species with high protein and low tannin content (Watts 1984).Diet shows little seasonal variation, with the exception of bamboo, which groups withaccess to it use when it is available (Watts 1998). Mountain gorillas also show various
442 A.A. Nowell, A.W. Fletcher

craniodental adaptations that are associated with a folivorous diet (Taylor 1997, 2002;Uchida 1998). As a consequence of the adaptations and the ever-present nature ofhigh-quality food in mountain gorilla habitat, we hypothesize that the transition tonutritional dependence in mountain gorillas may be more predictable and occur at anearlier age than in western gorillas. Data from studies on other great apes also supportthe hypothesis. Chimpanzees and orangutans rely more heavily on fruits than westerngorillas do and exhibit a less flexible diet in response to fruit shortages (Stanford andNkurunungi 2003; White and Tutin 2001). In chimpanzees, interbirth intervals, whichare inextricably linked to age at weaning as a result of lactational amenorrhea—where asuckling stimulus prevents a mother from returning to estrus—vary between 5 and 7 yr(Boesch and Boesch-Ackerman 2000), later than in all gorillas. In orangutans,interbirth intervals are even longer than those of chimpanzees: 7.7–9.3 yr (Galdikasand Wood 1990; Wich et al. 2004; van Noordwijk and van Schaik 2005).Consequently, one can apply to other great apes our hypothesis that western gorillasshould achieve independence from the mother later than mountain gorillas as a resultof resource predictability.
The majority of detailed data on gorilla behavior come from a small population ofmountain gorillas in the Virunga Volcanoes of Rwanda, Democratic Republic of Congoand Uganda, but at an altitudinal extreme of gorilla habitat, they cannot represent theentire genus (Robbins et al. 2004). It has been difficult to obtain detailed data onaspects of behavioral development in western gorillas, owing primarily to the densevegetation and lack of well habituated groups (Doran-Sheehy and Boesch 2004; Tutinand Fernandez 1991). Some habituation studies are now enjoying good levels ofcontact with their groups (Bermejo 2004; Cipolletta 2004; Doran et al. 2004), and it ishoped that more behavioral data on western gorillas in forest habitats will beforthcoming. In the meantime, the discovery of bais—open clearings used by manyspecies of forest mammal, primarily for feeding purposes—has provided new potentialfor answering questions regarding the demography and social behavior of westerngorillas, and one can collect data from an entire population of gorillas via the baiinstead of a small number of habituated groups. Important differences in thesocioecology of western and mountain gorillas are emerging as western gorilla studysites yield more detailed observations.
We conducted the first investigation of behavioral development in a large sample ofimmature wild western gorillas. We aim to provide insight into processes of behavioraldevelopment in the wild, focusing on proximity patterns between mother and offspring,the development of independent locomotion, suckling and weaning, and the nature ofmother-offspring interactions. In addition to exploring the extent to which femalewestern gorillas invest in their offspring, we consider the effects of resource availabilityon the development of independence from the mother.
Methods
Study Site
We collected data over 16 mo between February 2002 and August 2003 at MbeliBai, Nouabalé-Ndoki National Park, in the north of the Republic of Congo. Mbeli
Development of Independence in Gorillas 443

Bai is a 12.9-ha swampy clearing characterized by aquatic herbs, of which theCyperaceae together with Hydrocharis chevalieri are the preferred gorilla foods(Stokes et al. 2003).
Study Individuals
Thirteen social groups visited the bai, and we observed each group for 7–146 h(mean: 55±36.7 h). Observers individually recognized members of all groups. Werecorded data for all individuals <8 yr with the mother present in the social group—atotal of 42 immature individuals (Table I). We knew the approximate dates of birthfor the majority of subjects, which we classed as infants (0–3 yr), juveniles (3–6 yr),or subadults (6–8 yr). For others, we estimated dates of birth based on comparisonswith known-age individuals. Where possible, we categorized mothers as primiparous(n=4) or multiparous (n=14). All juveniles had multiparous mothers, so statisticalanalyses relating to parity of the mother are restricted to infants.
Data Collection
We observed gorillas from a platform on the bai edge, and subjects were present for atotal of ca. 550 h. Data collection (Altmann 1974) included scan sampling at 20-minintervals to record both the behavior of each subject and the distances between subjectsand other group members; focal sampling to record all behaviors continuouslyperformed by a randomly selected focal subject for 15-min periods; and sequencesampling to record all instances of less frequent behaviors, such as suckling and playinteractions. Immature subjects each contributed 1–200 scan samples (mean: 63.5) and0–8 h (mean: 2.7 h) of focal sampling (Table I). We used the following definitions ofbehavioral patterns that we recorded during focal and scan samples: feeding—ingestionof a recognized food item; processing—manipulation of a recognized food item;suckling—subject nursing from the mother; traveling—any form of locomotion
Table I Name, age (at start of the study), number of scans, and hours of focal observation for all subjects
Name(age in mo)
No. of scansamples
Focalhours
Name(age in months)
No. ofscans
Focalhours
Name(age in mo)
No. ofscans
Focalhours
DO (Niari) (–16) 8 0 Fang (1) 97 4.25 Mowgli (22) 57 1.75DO (Agatha)(–15) 8 .5 Hathi (2) 64 2.75 Ikki (28) 46 2.25Piment (–13) 69 2.75 Masai (4) 88 5 Peapod (30) 155 5.25Howsa (–12) 69 3 Sage (5) 228 8 Mysa (32) 56 2Lando (–12) 33 1.5 Nancy (6) 36 2.75 Brie (36) 147 4.5Hitchcock (–9) 12 1.25 Fig (8) 156 6 Kotick (38) 50 2Toomai (–9) 16 1.5 Likouala (8) 1 .25 Betsy (41) 26 1.75Jola (–8) 76 4.5 Luther (9) 26 1.5 Ndjeke (46) 18 1.25Kojak (–6) 31 2.75 Axel (10) 49 4.25 Sunbeam (48) 56 2Columbus (–4) 25 1.75 Kung (15) 95 3.75 Louis (50) 45 1.25Marple (–3) 9 .75 Whisky (15) 206 6.5 Darzee (53) 54 1.5Sherlock (–2) 12 .75 Bofi (18) 91 4.75 Pepper (63) 143 5.25Clyde (1) 29 2 Yoda (20) 38 .5 Ike (64) 55 3.5Custer (1) 25 1.5 Clover (21) 149 5 Ndoki (75) 16 .75
A minus age indicates an individual that was born after the start of the study.
444 A.A. Nowell, A.W. Fletcher

(position on mother if mother and offspring were in contact); play behavior—chasing,wrestling, or slapping with no apparent agonistic elements; other interactions—activebehavior directed from one individual to another, e.g., agonism, grooming (Table II);resting—no discernible activity performed by the study individual.
Data Analysis
We summarized data in 6-mo categories according to the age of the individual when wecollected the data. We calculated median values from each individual within a 6-mo agecategory, and then used the individual median values to give age-category medians(Fletcher 2001). We analyzed all data using nonparametric statistics. For all tests, thelevel of significance above which we rejected the null hypothesis was p=.05.
Results
Proximity to the Mother
As young gorillas increased in age, the percentage of time that they spent in contactwith the mother decreased significantly (Fig. 1: Spearman rank correlation: rs=−.966,n=20, p<.0001). We found no difference between males and females in their levelsof contact with the mother over the first 5 yr (Wilcoxon matched-pairs test: Z=−.296, n=11, p=.767), and observed no effect of the mother’s parity during infancy(Mann-Whitney U test: U=17.0, n=18, p=.238). We recorded no contact (exceptduring suckling bouts) during scan or focal samples between mother and offspring inany offspring >55 mo old.
As immature study subjects increased in age, mother-offspring distance increased. Asoffspring began to move out of contact with the mother, the percentage of scan samplesspent <2 m of the mother increased until 2.5 yr, before decreasing significantly during
Table II Definitions of interaction types between mothers and offspring
Interaction type Definition
Positive interactionsProximity seeking One individual establishes contact with, or seeks the proximity of another,
moving to ≤2 m of them without feeding or initiating agonismGrooming Manipulation of the hair by another individual, using hands, teeth or lipsPlay Chasing, wrestling or slapping of an individual by another, without agonistic
elements, often accompanied by a play faceNegative interactionsAgonistic Instances of hitting, slapping, aggressive vocalizations, biting, pushing,
and displacement interactions (the approach of one individual followedby the departure of the approached individual)
Suckling termination Termination or refusal of a suckling boutOther interactions (neither wholly positive or negative)Food transfer The movement of solid food between individualsMaternal restraint The mother prevents her offspring from leaving her immediate proximity,
or brings her offspring back into physical contact with her by clutchingit to her, or grabbing an arm or leg to stop her offspring from moving away
Development of Independence in Gorillas 445
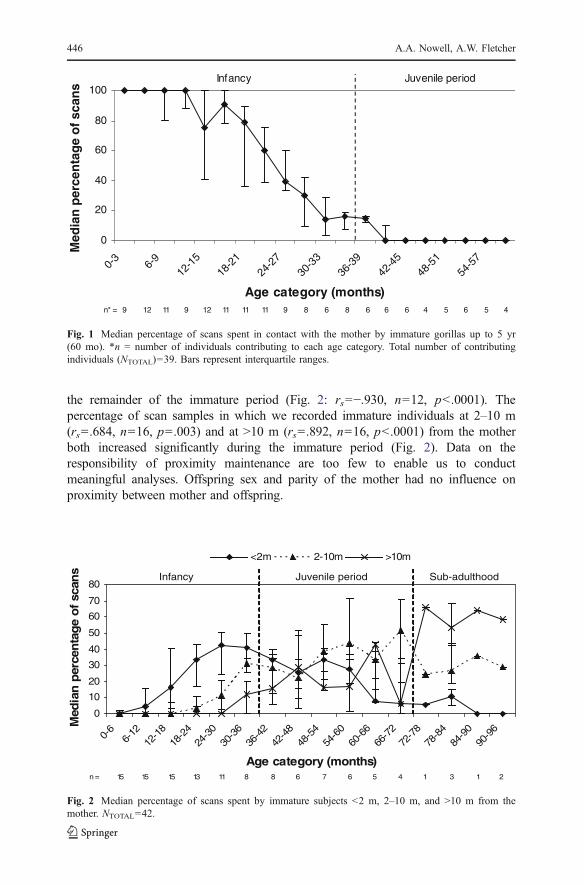
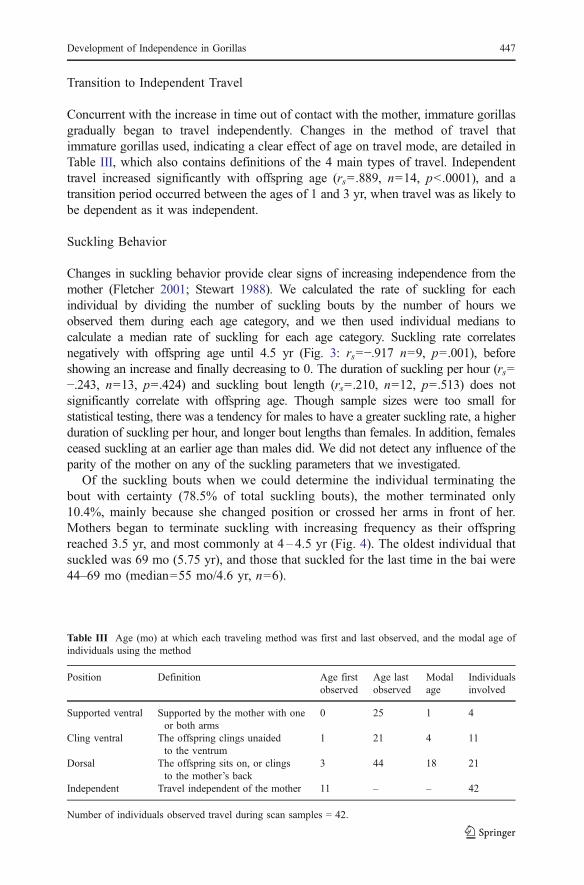
the remainder of the immature period (Fig. 2: rs=−.930, n=12, p<.0001). Thepercentage of scan samples in which we recorded immature individuals at 2–10 m(rs=.684, n=16, p=.003) and at >10 m (rs=.892, n=16, p<.0001) from the motherboth increased significantly during the immature period (Fig. 2). Data on theresponsibility of proximity maintenance are too few to enable us to conductmeaningful analyses. Offspring sex and parity of the mother had no influence onproximity between mother and offspring.
0
20
40
60
80
100
0-3
6-9
12-1
518
-21
24-2
730
-33
36-3
942
-45
48-5
154
-57
Age category (months)
Med
ian
per
cen
tag
e o
f sc
ans
n* = 9 12 11 9 12 11 11 11 9 8 6 8 6 6 6 4 5 6 5 4
Infancy Juvenile period
Fig. 1 Median percentage of scans spent in contact with the mother by immature gorillas up to 5 yr(60 mo). *n = number of individuals contributing to each age category. Total number of contributingindividuals (NTOTAL)=39. Bars represent interquartile ranges.
0
10
20
30
40
50
60
70
80
0-6
6-12
12-1
818
-24
24-3
030
-36
36-4
242
-48
48-5
454
-60
60-6
666
-72
72-7
878
-84
84-9
090
-96
Age category (months)
Med
ian
per
cen
tag
e o
f sc
ans
<2m 2-10m >10m
n = 15 15 15 13 11 8 8 6 7 6 5 4 1 3 1 2
Infancy Juvenile period Sub-adulthood
Fig. 2 Median percentage of scans spent by immature subjects <2 m, 2–10 m, and >10 m from themother. NTOTAL=42.
446 A.A. Nowell, A.W. Fletcher
Transition to Independent Travel
Concurrent with the increase in time out of contact with the mother, immature gorillasgradually began to travel independently. Changes in the method of travel thatimmature gorillas used, indicating a clear effect of age on travel mode, are detailed inTable III, which also contains definitions of the 4 main types of travel. Independenttravel increased significantly with offspring age (rs=.889, n=14, p<.0001), and atransition period occurred between the ages of 1 and 3 yr, when travel was as likely tobe dependent as it was independent.
Suckling Behavior
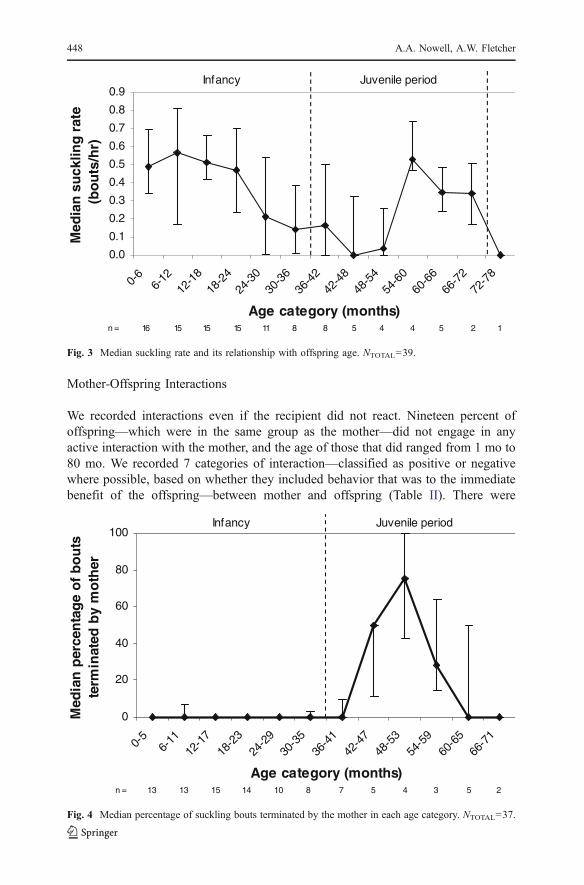
Changes in suckling behavior provide clear signs of increasing independence from themother (Fletcher 2001; Stewart 1988). We calculated the rate of suckling for eachindividual by dividing the number of suckling bouts by the number of hours weobserved them during each age category, and we then used individual medians tocalculate a median rate of suckling for each age category. Suckling rate correlatesnegatively with offspring age until 4.5 yr (Fig. 3: rs=−.917 n=9, p=.001), beforeshowing an increase and finally decreasing to 0. The duration of suckling per hour (rs=−.243, n=13, p=.424) and suckling bout length (rs=.210, n=12, p=.513) does notsignificantly correlate with offspring age. Though sample sizes were too small forstatistical testing, there was a tendency for males to have a greater suckling rate, a higherduration of suckling per hour, and longer bout lengths than females. In addition, femalesceased suckling at an earlier age than males did. We did not detect any influence of theparity of the mother on any of the suckling parameters that we investigated.
Of the suckling bouts when we could determine the individual terminating thebout with certainty (78.5% of total suckling bouts), the mother terminated only10.4%, mainly because she changed position or crossed her arms in front of her.Mothers began to terminate suckling with increasing frequency as their offspringreached 3.5 yr, and most commonly at 4 – 4.5 yr (Fig. 4). The oldest individual thatsuckled was 69 mo (5.75 yr), and those that suckled for the last time in the bai were44–69 mo (median=55 mo/4.6 yr, n=6).
Table III Age (mo) at which each traveling method was first and last observed, and the modal age ofindividuals using the method
Position Definition Age firstobserved
Age lastobserved
Modalage
Individualsinvolved
Supported ventral Supported by the mother with oneor both arms
0 25 1 4
Cling ventral The offspring clings unaidedto the ventrum
1 21 4 11
Dorsal The offspring sits on, or clingsto the mother’s back
3 44 18 21
Independent Travel independent of the mother 11 – – 42
Number of individuals observed travel during scan samples = 42.
Development of Independence in Gorillas 447
Mother-Offspring Interactions
We recorded interactions even if the recipient did not react. Nineteen percent ofoffspring—which were in the same group as the mother—did not engage in anyactive interaction with the mother, and the age of those that did ranged from 1 mo to80 mo. We recorded 7 categories of interaction—classified as positive or negativewhere possible, based on whether they included behavior that was to the immediatebenefit of the offspring—between mother and offspring (Table II). There were
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
0-6
6-12
12-1
818
-24
24-3
030
-36
36-4
242
-48
48-5
454
-60
60-6
666
-72
72-7
8
Age category (months)
Med
ian
su
cklin
g r
ate
(bo
uts
/hr)
n = 16 15 15 15 11 8 8 5 4 4 5 2 1
Infancy Juvenile period
Fig. 3 Median suckling rate and its relationship with offspring age. NTOTAL=39.
0
20
40
60
80
100
0-5
6-11
12-1
718
-23
24-2
930
-35
36-4
142
-47
48-5
354
-59
60-6
566
-71
Age category (months)
Med
ian
per
cen
tag
e o
f b
ou
tste
rmin
ated
by
mo
ther
n = 13 13 15 14 10 8 7 5 4 3 5 2
Infancy Juvenile period
Fig. 4 Median percentage of suckling bouts terminated by the mother in each age category. NTOTAL=37.
448 A.A. Nowell, A.W. Fletcher
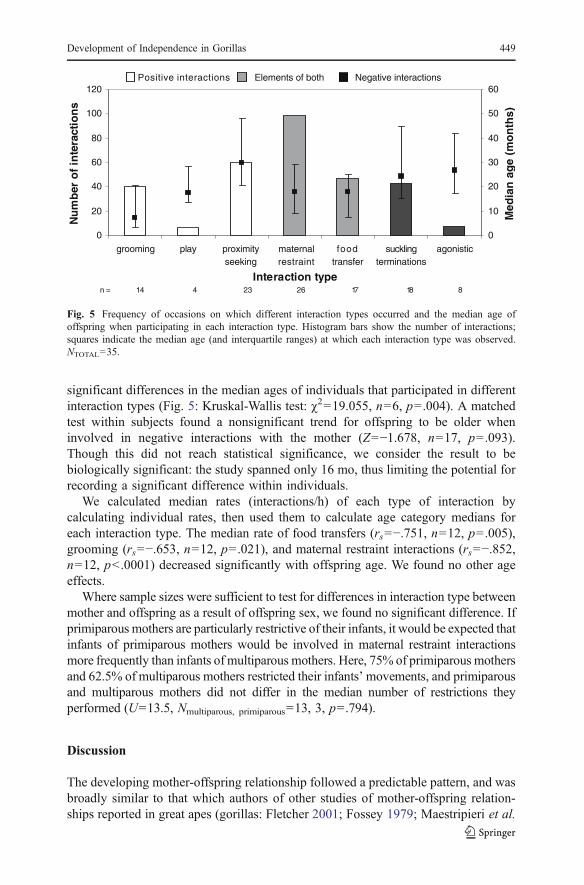
significant differences in the median ages of individuals that participated in differentinteraction types (Fig. 5: Kruskal-Wallis test: #2=19.055, n=6, p=.004). A matchedtest within subjects found a nonsignificant trend for offspring to be older wheninvolved in negative interactions with the mother (Z=−1.678, n=17, p=.093).Though this did not reach statistical significance, we consider the result to bebiologically significant: the study spanned only 16 mo, thus limiting the potential forrecording a significant difference within individuals.
We calculated median rates (interactions/h) of each type of interaction bycalculating individual rates, then used them to calculate age category medians foreach interaction type. The median rate of food transfers (rs=−.751, n=12, p=.005),grooming (rs=−.653, n=12, p=.021), and maternal restraint interactions (rs=−.852,n=12, p<.0001) decreased significantly with offspring age. We found no other ageeffects.
Where sample sizes were sufficient to test for differences in interaction type betweenmother and offspring as a result of offspring sex, we found no significant difference. Ifprimiparous mothers are particularly restrictive of their infants, it would be expected thatinfants of primiparous mothers would be involved in maternal restraint interactionsmore frequently than infants of multiparous mothers. Here, 75% of primiparous mothersand 62.5% of multiparous mothers restricted their infants’movements, and primiparousand multiparous mothers did not differ in the median number of restrictions theyperformed (U=13.5, Nmultiparous, primiparous=13, 3, p=.794).
Discussion
The developing mother-offspring relationship followed a predictable pattern, and wasbroadly similar to that which authors of other studies of mother-offspring relation-ships reported in great apes (gorillas: Fletcher 2001; Fossey 1979; Maestripieri et al.
0
20
40
60
80
100
120
grooming play proximityseeking
maternalrestraint
foodtransfer
sucklingterminations
agonistic
Interaction type
Nu
mb
er o
f in
tera
ctio
ns
0
10
20
30
40
50
60
Med
ian
ag
e (m
on
ths)
n = 14 4 23 26 17 18 8
Positive interactions Elements of both Negative interactions
Fig. 5 Frequency of occasions on which different interaction types occurred and the median age ofoffspring when participating in each interaction type. Histogram bars show the number of interactions;squares indicate the median age (and interquartile ranges) at which each interaction type was observed.NTOTAL=35.
Development of Independence in Gorillas 449

2002; chimpanzees: van Lawick-Goodall 1971; Watts and Pusey 1993; orangutans:van Noordwijk and van Schaik 2005) and other primates (baboons: Nash 1978;vervets: Fairbanks 1993). With increasing offspring age, a decline in contact andproximity between mothers and offspring, development of independent travel,cessation of suckling, and a subtle change in the nature of interactions betweenmother and offspring occurred, all pointing toward decreasing investment in thecurrent offspring as the mother prepares to invest in the next. Though offspring areselected to demand more than mothers are selected to provide (Trivers 1974), wefound little evidence of open conflict between mother and offspring.
More data are required to determine whether western gorillas have a prolongedinvestment in sons instead of daughters, but the tendency for males to have a highersuckling rate, a higher duration of suckling per hour, longer bout lengths, and tocontinue suckling to a later age than females suggests that mothers may be moretolerant of demands to suckle by sons than by daughters. There is a high degree ofsexual dimorphism between adult male and female gorillas (Taylor 1997), and a lowratio of males to females within reproductive groups, pointing to a greater degree ofvariability in the reproductive success of males (Watts 1991). If male reproductivesuccess in gorillas is influenced by body size (Clutton-Brock et al. 1984), then amother’s increased investment in her sons, e.g., by tolerating suckling for longer, ispotentially a way in which she can increase her indirect reproductive success(Boesch 1997; Trivers and Willard 1973).
We predicted that the parity of the mother would influence the nature of therelationship between mother and offspring, and to influence the age at whichoffspring reached independence (Hooley and Simpson 1981; Manson 1999), but wefound no evidence of primiparous mothers behaving differently toward theiroffspring. It is likely that the bai environment may have influenced the behaviorof mothers, by increasing their tendency to restrict offspring movement. Mbeli Bai isswampy and difficult to move about in, and the high percentage of multiparousmothers that restricted their offspring’s movements may suggest that all motherswere more protective in the environment, thereby masking any effect of parity of themother.
The age at which western gorillas displayed behavior characteristic of anindependent immature appeared later than in wild mountain gorillas (Fletcher 2001;Stewart 1981). Increases in independence also seemed to develop differently tocaptive western gorillas (Maestripieri et al. 2002) wherein, e.g., the most significantchange in the time offspring spent in contact with the mother occurred during the firstyear of life, with relatively little change between 2 and 4 yr. In our study, contact withthe mother was almost constant until the age of 1 yr, with changes occurring beyondthen. Again, one must take the effect of the bai as an environment into considerationwhen interpreting the observed effects: gorillas spend as little as 1% of their total timein bais (Magliocca and Gautier-Hion 2002), and the behavior during this period maynot entirely represent their behavior in the more familiar forest environment. Younggorillas may remain closer to their mothers in unfamiliar situations (sensu Fossey1979), thus prolonging the apparent period of dependence on the mother in westerngorillas when measured in a bai. With many different gorilla groups and solitarysilverbacks using bais, the possibility of encountering other groups here may beincreased (Parnell 2002), prompting more control of offspring by nervous mothers,
450 A.A. Nowell, A.W. Fletcher

and a consequent apparent increase in the age at which offspring achieveindependence from the mother. However, the bai is an open habitat, and offersgreater visibility than the forest, potentially limiting the dangers of predation andinfanticide. Locomotion is terrestrial, and though the substrate is swampy, lower bodyweight may enable smaller individuals to negotiate travel with less difficulty thanadults, and therefore have less need of transport by the mother. Similarly, after adultssettled to feed, little movement occurred until the group left the bai (Nowell, pers.obs.), again resulting in little necessity for transportation by the mother. In addition,group spread tends to be less than the spreads of ≤500 m that occur in the forest(Bermejo 2004; Tutin 1996). The bai-specific observations may suggest reduceddependence on the mother recorded earlier than one would otherwise expect in foresthabitats.
Patterns of suckling, especially the age at which individuals last suckle, are morelikely to be independent of habitat type and thus should provide a more reliableindication of the reliance of offspring on their mothers than proximity patterns. Themedian age of weaning in our study was 4.6 yr, compared with 3.6 yr in mountaingorillas (Fletcher 2001; Stewart 1981), showing that western gorilla motherscontinue to invest in their offspring for longer than their mountain gorillacounterparts do (Nowell 2005). Weaning seemed to be a process determined bymothers, as shown by the increase with which they terminated suckling bouts astheir offspring approached 4 yr. It also seemed a gentle process: increasedterminations appeared to lead to increased attempts to suckle which were permittedwhen offspring persisted, hence the temporary increase in suckling rate around4.5 yr. The dynamic assessment theory proposed by Bateson (1994) predicts thatmother and offspring monitor each other closely, and the mother responds to genuineneed in her offspring (Bateson 1994; Devinney et al. 2001), so we consider thatpersistent attempts to suckle by offspring represented an honest need. Rather thanopen conflict between mother and offspring over the extent to which maternalinvestment continued, the results suggest that dynamic assessment theory may moreaccurately describe weaning behavior between mothers and offspring in gorillas(sensu Bateson 1994; Devinney et al. 2001).
The large size of western gorillas allows them to digest the increased amounts ofterrestrial herbaceous vegetation that is incorporated into their diet during periods offruit scarcity (Remis 2000). Fruits are likely to be suitable weaning foods, but the gutof a small, young gorilla may not be of a sufficient size or maturity (Lee 1996) toprocess enough herbaceous vegetation to sustain it through periods of fruit scarcity.We propose that the ephemerally available and widely dispersed nature of thewestern gorilla diet (Remis 1997; Rogers et al. 2004) drives the need for continuedinvestment by the mother. Researchers have found similar results in studies ofbaboons (Lycett et al. 1998; Barrett et al. 2006), seals (Trillmich 1990), andelephants (Lee and Moss 1986), in which food limitations and drought years haveextended the period of lactation, indicating the dangers of the period during whichthe transition from suckling to self-sufficiency in food acquisition occurs (Fragaszyand Bard 1997; Janson and van Schaik 1993). If availability of fruit is linked toweaning, one might predict that there should be differences in seasonal sucklingrates. A month-by-month analysis of suckling rate (Nowell, unpub. data) showed nosignificant correlation with season, but ideally suckling data should correlate with
Development of Independence in Gorillas 451

phenological data to investigate more fully the influence of fruit availability onsuckling, and ultimately its effect on an immature individual’s attainment ofindependence from the mother.
A heavy reliance on fruit and an increasingly inflexible diet in response to fruitshortage may also be linked to variation in the period of dependence on the motherin other great apes, not only accounting for differences between frugivorous westerngorillas and the more folivorous mountain gorillas, but also explaining the extremelylong interbirth intervals of orangutans (van Noordwick and van Schaik 2005).
Despite immature gorillas being capable of traveling independently, showing lowlevels of contact and engaging in few affiliative interactions with the mother,suckling is still tolerated, suggesting that a late age at weaning in western gorillas isnecessary to allow them to proceed to the next stage of immaturity. Though certainindependent feeding skills are in place by the end of infancy and undergo littlesignificant further change (Nowell 2005; Nowell and Fletcher, in press) similarly tomountain gorillas (Byrne and Byrne 1993; Watts 1985), we propose that theavailability of food items within the environment can ultimately determine whenan individual can achieve independence from the mother. By responding togenuine needs of her current offspring and permitting the continuation of sucklingat the expense of investment in subsequent offspring, a mother may be able toreduce the probability of offspring death from care-dependent factors, thusimproving the chances of offspring survival and increased reproductive success.
Acknowledgments We thank the Ministère de l’Economie Forestière of Congo-Brazzaville and theWildlife Conservation Society for permission to work in the Nouabalé-Ndoki National Park. We thank allstaff of the Nouabalé-Ndoki Project, particularly Thomas Breuer, Emma Stokes, Mark Gately, and BryanCurran. A University of Chester Gladstone Fellowship provided funding for A. Nowell.
References
Altmann, J. (1974). Observational study of behaviour: Sampling methods. Behaviour, 49, 227–267.Barrett, L., Henzi, S. P., Lycett, J. E. (2006). Whose life is it anyway? Maternal investment, developmental
trajectories and life history strategies in baboons. In L. Swedell & S. R. Leigh (Eds.), BaboonReproductive Strategies and Life History (pp. 199–224). London: Kluwer Academic Publishers.
Bateson, P. (1994). The dynamics of parent-offspring relationships in mammals. Trends in Ecology &Evolution, 9, 399–403.
Berman, C. M. (1988). Maternal condition and offspring sex ratio in a group of free ranging rhesusmonkeys: An eleven year study. American Naturalist, 131, 307–328.
Bermejo, M. (2004). Home-range use and intergroup encounters in western gorillas (Gorilla g. gorilla) atLossi Forest, North Congo. American Journal of Primatology, 64, 223–232.
Boesch, C. (1997). Evidence for dominant wild female chimpanzees investing more in sons. AnimalBehaviour, 54, 811–815.
Boesch, C., & Boesch-Achermann, H. (2000). The Chimpanzees of the Taï Forest: Behavioural Ecologyand Evolution. Oxford University Press, Oxford.
Brown, G. R., & Dixson, A. F. (2000). The development of behavioural sex differences in infant rhesusmacaques (Macaca mulatta). Primates, 41, 63–77.
Byrne, R. W., & Byrne, J. M. E. (1993). The complex leaf gathering skills of mountain gorillas (Gorilla g.beringei)—variability and standardization. American Journal of Primatology, 31, 241–261.
Cipolletta, C. (2004). Effects of group dynamics and diet on the ranging patterns of a western gorillagroup (Gorilla gorilla gorilla) at Bai Hokou, Central African Republic. American Journal ofPrimatology, 64, 193–205.
452 A.A. Nowell, A.W. Fletcher

Clutton-Brock, T., Albon, S. D., & Guinness, F. E. (1984). Maternal dominance, breeding success, andbirth sex ratios in red deer. Nature, 308, 358–360.
Delgado, R. A., & van Schaik, C. P. (2000). The behavioral ecology and conservation of the orangutan(Pongo pygmaeus): A tale of two islands. Evolutionary Anthropology, 9, 201–218.
Devinney, B. J., Berman, C. M., & Rasmussen, K. L. R. (2001). Changes in yearling rhesus monkeys’relationships with their mothers after sibling birth. American Journal of Primatology, 54, 193–210.
Doran, D., Greer, D., Mongo, P., & Schwindt, D. (2004). Impact of ecological and social factors onranging in western gorillas. American Journal of Primatology, 64, 207–222.
Doran, D., McNeilage, A., Greer, D., Mocian, C., Mehlman, P., & Shah, N. (2002). Western lowlandgorilla diet and resource availability: New evidence, cross-site comparisons, and reflections onindirect sampling methods. American Journal of Primatology, 58, 91–116.
Doran-Sheehy, D. M., & Boesch, C. (2004). Behavioral ecology of western gorillas: New insights fromthe field. American Journal of Primatology, 64, 139–143.
Fairbanks, L. A. (1993). Juvenile vervet monkeys: Establishing relationships and practising skills for thefuture. In M. E. Pereira & L. A. Fairbanks (Eds.), Juvenile Primates: Life History, Development, andBehavior (pp. 211–227). New York: Oxford University Press.
Fletcher, A. W. (2001). Development of infant independence from the mother in wild mountain gorillas. InM. M. Robbins, P. Sicotte & K. J. Stewart (Eds.), Mountain Gorillas: Three Decades of Research atKarisoke (pp. 153–182). Cambridge: Cambridge University Press.
Fossey, D. (1979). Development of the mountain gorilla (Gorilla gorilla beringei): The first 36 months. InD. A. Hamburg & E. R. McCown (Eds.), The Great Apes (pp. 139–186). Menlo Park, CA: Benjamin/Cummings.
Fragaszy, D., & Bard, K. A. (1997). Comparison of development and life history in Pan and Cebus.International Journal of Primatology, 18, 683–701.
Galdikas, B. F. M., & Wood, J. W. (1990). Birth spacing patterns in humans and apes. American Journalof Physical Anthropology, 83, 185–191.
Harvey, P. H., & Clutton-Brock, T. H. (1985). Life history variation in primates. Evolution, 39, 559–581.Hooley, J. M., & Simpson, M. J. A. (1981). A comparison of primiparous and multiparous mother-infant
dyads in Macaca mulatta. Primates, 22, 379–392.Horvat, J. R., & Kraemer, H. C. (1981). Infant socialization and maternal influence in chimpanzees. Folia
Primatologica, 36, 99–110.Janson, C. H., & van Schaik, C. P. (1993). Ecological risk aversion in juvenile primates: Slow and steady
wins the race. In M. E. Pereira & L. A. Fairbanks, (Eds.), Juvenile Primates; Life History,Development and Behavior (pp. 62–73.). New York: Oxford University Press.
Kappeler, P. M., Pereira, M. E., & van Schaik, C. P. (2003). Primate life histories and socioecology. InP. M. Kappeler & M. E. Pereira (Eds.), Primate Life Histories and Socioecology (pp. 1–20). Chicago:University of Chicago Press.
Lee, P. C. (1996). The meanings of weaning: growth, lactation and life history. EvolutionaryAnthropology, 5, 87–96.
Lee, P. C., & Moss, C. J. (1986). Early maternal investment in male and female African elephant calves.Behavioural Ecology and Sociobiology, 18, 353–361.
Lycett, J. E., Henzi, S. P., & Barrett, L. (1998). Maternal investment in mountain baboons and thehypothesis of reduced care. Behavioural Ecology and Sociobiology, 42, 49–56.
Maestripieri, D., & Carrol, K. A. (1998). Risk factors for infant abuse and neglect in group-living rhesusmonkeys. Psychological Science, 9, 143–145.
Maestripieri, D., Ross, S. K., & Megna, N. L. (2002). Mother-infant interactions in western lowlandgorillas (Gorilla gorilla gorilla): Spatial relationships, communication, and opportunities for sociallearning. Journal of Comparative Psychology, 116, 219–227.
Magliocca, F., & Gautier-Hion, A. (2002). Mineral content as a basis for food selection by westernlowland gorillas in a forest clearing. American Journal of Primatology, 57, 67–77.
Manson, J. H. (1999). Infant handling in wild Cebus capuchinus: Testing bonds between females? AnimalBehaviour, 57, 911–921.
Meikle, D. B., Tilford, B. L., & Vessey, S. H. (1984). Dominance rank, secondary sex ratio andreproduction in polygynous primates. American Naturalist, 124, 173–188.
Nakamichi, M., Silldor, A., Bringham, C., & Sexton, P. (2004). Baby-transfer and other interactionsbetween its mother and grandmother in a captive social group of lowland gorillas. Primates, 45,73–77.
Nash, L. T. (1978). The development of the mother-infant relationship in wild baboons (Papio anubis).Animal Behaviour, 26, 746–759.
Development of Independence in Gorillas 453

Nevison, C. M., Rayment, F. D. G., & Simpson, M. J. A. (1996). Birth sex ratios and maternal socialrank in a captive colony of rhesus monkeys (Macaca mulatta). American Journal of Primatology,39, 123–138.
Nicolson, N. A. (1987). Infants, mother and other females. In B. B. Smuts, D. L. Cheney, R. M. Seyfarth,R. W. Wrangham & T. T. Struhsaker (Eds.), Primate Societies (pp. 330–342). Chicago: University ofChicago Press.
Nowell, A. A. (2005). Behavioural Development in Wild Western Lowland Gorillas (Gorilla gorillagorilla). Ph.D. thesis, University of Liverpool.
Nowel, A. A., & Fletcher, A. W. (in press) The development of feeding behaviour in wild western lowlandgorillas (Gorilla gorrilla gorilla).
Parnell, R. J. (2002). The Social Structure and Behaviour of Western Lowland Gorillas (Gorilla gorillagorilla). at Mbeli Bai, Republic of Congo. Ph.D. thesis, University of Stirling.
Remis, M. J. (1997). Western lowland gorillas (Gorilla gorilla gorilla) as seasonal frugivores: Use ofvariable resources. American Journal of Primatology, 43, 87–109.
Remis, M. J. (2000). Initial studies on the contributions of body size and gastrointestinal passage rates todietary flexibility among gorillas. American Journal of Physical Anthropology, 112, 171–180.
Robbins, M. M., Bermejo, M., Cipolletta, C., Magliocca, F., Parnell, R. J., & Stokes, E. J. (2004). Socialstructure and life-history patterns in western gorillas (Gorilla gorilla gorilla). American Journal ofPrimatology, 64, 145–159.
Rogers, M. E., Abernethy, K., Bermejo, M., Cipolletta, C., Doran, D., McFarland, K., Nishihara, T.,Remis, M. J., & Tutin, C. E. G. (2004). Western gorilla diet: A synthesis from six sites. AmericanJournal of Primatology, 64, 173–192.
Stanford, C. B., & Nkurunungi, J. B. (2003). Behavioral ecology of sympatric chimpanzees and gorillas inBwindi Impenetrable National Park, Uganda: diet. International Journal of Primatology, 24, 901–918.
Stewart, K. J. (1981). Social Development of Wild Mountain Gorillas. Ph.D. thesis, University ofCambridge.
Stewart, K. J. (1988). Suckling and lactational anestrus in wild gorillas (Gorilla Gorilla). Journal ofReproduction and Fertility, 83, 627–634.
Stokes, E. J., Parnell, R. J., & Olejniczak, C. (2003). Female dispersal and reproductive success in westernlowland gorillas (Gorilla gorilla gorilla). Behavioural Ecology and Sociobiology, 54, 329–339.
Taylor, A. B. (1997). Relative growth, ontogeny and sexual dimorphism in Gorilla (Gorilla gorilla gorillaand G. g. beringei): Evolutionary and ecological considerations. American Journal of Primatology,43, 1–31.
Taylor, A. B. (2002). Masticatory form and function in the African apes. American Journal of PhysicalAnthropology, 117, 133–156.
Trillmich, F. (1990). The behavioural ecology of maternal effort in fur seals and sea lions. Behaviour, 114,3–20.
Trivers, R. (1972). Parental investment and sexual selection. In B. Campbell (Ed.), Sexual Selection andthe Descent of Man 1871–1971 (pp. 136–179). Chicago: Heinemann Educational Books.
Trivers, R. (1974). Parent-offspring conflict. American Zoologist, 14, 249–264.Trivers. R., & Willard, D. (1973). Natural selection of parent ability to vary the sex ratio of offspring.
Science, 179, 90–92.Tutin, C. E. G. (1996). Ranging and social structure of lowland gorillas in the Lope Reserve, Gabon. In W.
C. McGrew, L. C. Marchant & T. Nishida (Eds.), Great Ape Societies (pp. 58–70). Cambridge:Cambridge University Press.
Tutin, C. E. G., & Fernandez, M. (1991). Responses of wild chimpanzees and gorillas to the arrival ofprimatologists: Behaviour observed during habituation. In H. O. Box (Ed.), Primate Responses toEnvironmental Change (pp. 187–197). Cambridge: Chapman and Hall.
Uchida, A. (1998). Variation in tooth morphology of Gorilla gorilla. Journal of Human Evolution, 34,55–70.
van Lawick-Goodall, J. (1971). Some aspects of mother-infant relationships in a group of wildchimpanzees. In H. R. Schaffer (Ed.), The Origins of Human Social Relations (pp. 115–128).London: Academic Press.
van Noordwijk, M. A., & van Schaik, C. P. (2005). Development of ecological competence in Sumatranorangutans. American Journal of Physical Anthropology, 127, 79–94.
Watts, D. P. (1984). Composition and variability of mountain gorilla diets in the Central Virungas.American Journal of Primatology, 7, 323–356.
Watts, D. P. (1985). Observations on the ontogeny of feeding behavior in mountain gorillas (Gorillagorilla beringei). American Journal of Primatology, 8, 1–10.
454 A.A. Nowell, A.W. Fletcher

Watts, D. P. (1991). Mountain gorilla reproduction and sexual behavior. American Journal of Primatology,24, 211–225.
Watts, D. P. (1996). Comparative socio-ecology of gorillas. In: W. C. McGrew, L. C. Marchant, & T.Nishida (Eds.), Great Ape Societies (pp. 16–29). Cambridge: Cambridge University Press.
Watts, D. P. (1998). Seasonality in the ecology and life histories of mountain gorillas (Gorilla gorillaberingei). International Journal of Primatology, 19, 929–948.
Watts, D. P., & Pusey, A. E. (1993). Behavior of juvenile and adolescent great apes. In M. E. Pereira & L.A. Fairbanks (Eds.) Juvenile Primates; Life History, Development and Behavior (pp. 148–167). NewYork: Oxford University Press.
White, L. J. T., & Tutin, C. E. G. (2001). Why gorillas and chimpanzees react diffirently to logging: acautionary tale from Gabon. In: W. Weber, L. J. T. White, A. Vedder, & L. Naugthon–Treves (Eds.),African Rainforest Ecology and Conservation: An Interdisciplinary Perspective (pp. 449–462). NewHaven, CT: Yale University Press.
Wich, S. A., Utami-Atmoko, S. S., Setia, T. M., Rijksen, H. D., Schurmann, C., & van Schaik, C. (2004).Life history of wild Sumatran orangutans (Pongo abelii). Journal of Human Evolution, 47, 385–398.
Development of Independence in Gorillas 455





























