Embed Size (px)
Citation preview

JOURNAL OF SURGICAL RESEARCH 63, 220–224 (1996)ARTICLE NO. 0251
Dichloroacetate Enhanced Myocardial Functional RecoveryPost-ischemia: ATP and NADH Recovery
JOYCE A. WAHR, M.D., DOUGLAS OLSZANSKI, M.D., KEITH F. CHILDS, B.S., AND STEVEN F. BOLLING, M.D.
Departments of Anesthesiology and Surgery, Section of Thoracic Surgery, University of Michigan, Ann Arbor, Michigan 48104-0048
Presented at the Annual Meeting of the Association for Academic Surgery, Dearborn, Michigan, November 8–11, 1995
niques, myocardial metabolic requirements are notThis study was undertaken to determine the effect eliminated, and functional and metabolic recovery is
of dichloroacetate (DCA) on myocardial functional and often impaired at reperfusion. Depletion of ATP levelsmetabolic recovery following global ischemia. Isolated and/or slow restoration of ATP levels following isch-rabbit hearts were subjected to 120 min of mildly hypo- emia appears to be a critical factor in this poor func-thermic (347C), cardioplegic arrest with multidose, tional recovery, and agents that enhance repletion ofmodified St. Thomas’ cardioplegia. Hearts were reper- ATP following ischemia should improve recovery. Oxi-fused with either physiologic salt solution (PSS) as dative phosphorylation is a critical source of cellularcontrols, (CON, n Å 10) or PSS containing DCA (DCA, ATP; a key step in this metabolic pathway is the oxida-nÅ 6) at a concentration of 1 mM. Functional and meta-
tion of pyruvate to acetyl CoA by the multi-enzymebolic indices were determined at baseline and at 15,complex pyruvate dehydrogenase (PDH). Dichloroa-30, and 45 min of reperfusion. In four DCA and fourcetate (DCA) has been shown to enhance the activityCON hearts, myocardial biopsies were taken at base-of PDH in virtually all tissues [1], and has been pre-line, end-ischemia, 15 and 45 min for nucleotide levels.viously noted to limit the ST segment elevation associ-Functional recovery was significantly better in heartsated with abrupt coronary occlusion in dogs [2]. DCAreperfused with DCA as demonstrated by recovery ofhas also been shown to enhance glucose oxidation inbaseline developed pressure (DCA Å 69 { 5%, CON Åisolated, endotoxin-shocked rat hearts [3], and to im-45 { 9%) and dP/dt (DCA Å 64% { 10% versus CONprove cardiac index in patients with severe lactic aci-Å 48% { 10%). Coronary blood flow was not differentdosis [4, 5]. DCA has been used in humans with littlebetween groups either at baseline or during reperfu-
sion, but myocardial oxygen consumption (MVO2) was or no signs of toxicity [1, 4, 5] and thus has potentialincreased in the DCA versus CON hearts (79% { 20% for use in patients with myocardial ischemia, espe-of baseline vs 50% { 18%). Recovery of myocardial ade- cially the induced ischemia that occurs during cardiacnylate energy status was improved in the DCA versus surgery.CON hearts (ATP recovered to 45% { 20% versus 8% { We have previously demonstrated that DCA en-6% of baseline). Coronary sinus lactate concentration hances functional recovery following global ischemiawas decreased in DCA perfused hearts at 45 min of during mild hypothermic cardioplegic arrest in iso-reperfusion. Percent of baseline NADH values was sim- lated, perfused rabbit hearts [6]. We postulated thatilar at 15 min of reperfusion, but at 45 min, DCA hearts DCA, by increasing the activity of PDH, improves theshowed a decrease in NADH levels, while CON hearts recovery of myocellular metabolic energy status.showed an increase (DCA Å 48%; CON Å 121%). Theenhanced myocardial function and improved meta-
METHODSbolic status noted with DCA may result from increasedoxidative phosphorylation due to altered pyruvate de-hydrogenase (PDH) activity. q 1996 Academic Press, Inc. Preparation of isolated heart. Studies were performed in isolated,
perfused rabbit hearts. Male New Zealand rabbits weighing 3–4 kgwere anesthetized with intravenously administered sodium pento-barbital (10 mg/kg), after which the heart was removed through amedian sternotomy. The heart was rapidly excised and immersed inINTRODUCTIONice-cold physiologic salt solution (PSS, 118.0 mM NaCl, 4.0 mM KCl,22.3 mM NaHCO3, 11.1 mM glucose, 0.66 mM K2HPO4, 1.23 mM
Global myocardial ischemia may result in poor func- MgCl2, and 2.38 mM CaCl2) and was suspended from a perfusiontional recovery despite restoration of myocardial blood column by aortic cannulation within 30 sec of excision (Langendorff
preparation). Coronary perfusion was established at 80 mmHg withflow and repletion of substrate necessary for energyPSS equilibrated with 95% O2–5% CO2. The oxygen tension of theproduction. Hypothermic cardioplegic arrest is fre-perfusate was maintained between 450 and 550 mmHg, and thequently used to provide myocardial protection during perfusate temperature was kept at 377C by means of a heat ex-
the period of mandatory myocardial ischemia that oc- changer in the perfusion column.After coronary perfusion was initiated, the heart generally re-curs during cardiac surgery. Even with these tech-
2200022-4804/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
AID JSR 4819 / 6n0e$$$841 05-24-96 12:58:07 srga AP: Surg Res

221WAHR ET AL.: DCA AND MYOCARDIAL RECOVERY
gained sinus rhythm, but in the presence of persistent fibrillation, fused at 80 mmHg with oxygenated PSS at 377C and the water bathtemperature was increased to 377C. Control hearts (n Å 10) weredirect-current cardioversion was carried out. While the heart was
perfused on the column, a portion of the mitral valve with its chordae reperfused with the same PSS used during baseline, and DCA hearts(n Å 6) were reperfused with PSS containing 1 mM DCA. A freshwas excised, and a latex balloon, connected to tubing filled with saline
solution, was introduced into the left ventricle through the mitral solution of DCA (Sigma Chemicals, St. Louis, MO) was prepared foreach experiment, and the solution was adjusted to a pH of 7.35–orifice. The balloon was sewn in place in such a way as to allow
passive venting of the left ventricular (LV) cavity. The balloon was 7.45. Defibrillation was performed as needed during the initial 3 minof reperfusion. During the first 15 min of reperfusion the intraven-connected to a Statham P23Db pressure transducer (Gould, Akron,
OH) for continuous measurement of left ventricular pressure (LVP) tricular balloon was kept deflated to simulate the beating, nonwork-ing condition. After the initial 15 min of reperfusion, the LV balloonand the first derivative of LVP (dP/dt), which were recorded on an
IBM analog to digital system (Purchase, NY). The superior vena was refilled to the preischemic EDV and hemodynamic and metabolicmeasurements were made. The balloon remained inflated for thecava and inferior vena cava were ligated. The pulmonary artery was
cannulated to enable timed collection measurements of coronary flow remainder of the reperfusion period and measurements were ob-tained after 15, 30, and 45 min of reperfusion. Diastolic stiffnessand the cannula was connected to a Clark electrode (Chemical micro-
sensor, Diamond Electro-Tech, Inc., Ann Arbor, MI) for continuous (compliance) was determined as at baseline.After 45 min of reperfusion, all hearts were removed from themeasurement of the partial pressure of oxygen in the coronary efflu-
ent. Coronary flow was measured by performing timed collections of perfusion apparatus. Wet weight of the heart was determined at theconclusion of each experiment after trimming the great vessels andthe pulmonary effluent flow with a graduated cylinder.fat and blot drying. The myocardial water content was determinedHemodynamic and metabolic measurements. Developed pressureby weighing a fresh specimen of myocardium, allowing this sample(DP) was defined as peak systolic pressure (PSP) minus end-diastolicto desiccate for 48 hr at 807C, and then reweighing the sample. Per-pressure (EDP). The product of heart rate (HR) and DP (PRP, mmHg/cent water was calculated using the formula ([1 0 dry weight/wetmin) was calculated to provide an estimate of changes in myocardialweight] X 100 Å %H2O).work. Myocardial oxygen consumption (MVO2) was calculated as
This protocol was approved by the University of Michigan Institu-tional Animal Care and Use Committee and complied with the Prin-
MVO2 Å CF 1 [(PaO2 0 PvO2) 1 (c/760)] ciples of Laboratory Animal Care and the Guide for the Care and Useof Laboratory Animals (NIH Publication No. 80.23, revised 1978).
Values are reported as means { standard deviation, or as thewhere CF is coronary flow (ml/min/g), (PaO20 PvO2) is the differencemean percent of baseline value. Data were evaluated with repeatedin the partial pressure of oxygen (PO2, mmHg) between perfusatemeasures analysis of variance (ANOVA). When significant F valuesand coronary effluent flow, and c is the Bunsen solubility coefficientwere obtained, Scheffe’s test was used to distinguish which timeof O2 in perfusate at 377C (22.7 ml O2rATM01
rml01 perfusate).periods or groups differed from one another significantly. DifferencesIn four control (CON) and four DCA hearts, punch biopsies of thewere considered significant when P õ 0.05.free left ventricular wall were made for determination of nucleotide
levels. Biopsies were taken at baseline, at end-ischemia, and at 15min of reperfusion. Biopsied myocardium was frozen in liquid nitro- RESULTSgen and was assayed for the content of adenosine triphosphate (ATP),adenosine diphosphate (ADP), adenosine monophosphate (AMP),
Functional Recoveryinosine monophosphate (IMP), and NAD/NADH levels by high per-formance liquid chromatography. The total nondiffusible nucleotides
There were no significant differences at baseline for(TNN) were determined as the sum of ATP, ADP, AMP, and IMP.Adenylate energy charge (AEC) or energy status was determined by HR, DP, or dP/dt values between CON hearts and DCAthe formula (ATP / ADP (1
2))/(ATP / AMP / ADP). Results are hearts. Baseline DP averaged 110 mmHg for all hearts.expressed as percent of baseline concentration at end-ischemia and Left ventricular systolic and diastolic functional mea-15 min of reperfusion.
surements of DP and dP/dt made after 15 and 45 min ofProtocol. After completing instrumentation and performing cali-
reperfusion are shown as percent recovery of functionbrations, left ventricular balloon volumes were varied over a rangecompared to baseline values.of values to construct modified left ventricular function curves. In
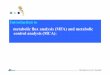
this manner, it was possible to define a specific balloon volume (end- After 120 min of hypothermic ischemia and 45 mindiastolic volume, EDV) that was associated with a developed pres- of reperfusion, CON hearts recovered 45 { 9% of base-sure between 100 and 140 mmHg. This volume was maintained the line DP, and 48 { 10% of dP/dt (Fig. 1). Hearts reper-same for each heart during baseline and reperfusion conditions. The
fused with DCA showed significantly better recovery ofintraventricular balloon volumes were not adjusted to produce spe-function, to 69 { 5% of DP and 64 { 10% of baselinecific EDPs (rather, a defined level of systolic pressure development),
but EDPs at baseline greater than 10 mmHg were not considered dP/dt (P õ 0.05 compared to CON hearts).acceptable. Hearts characterized by developed pressures less than100 mmHg or greater than 140 mmHg were not used. After determi-
Metabolic Recoverynation of the best EDV for each heart, the balloon volume was in-creased in increments, permitting estimation of diastolic stiffness at
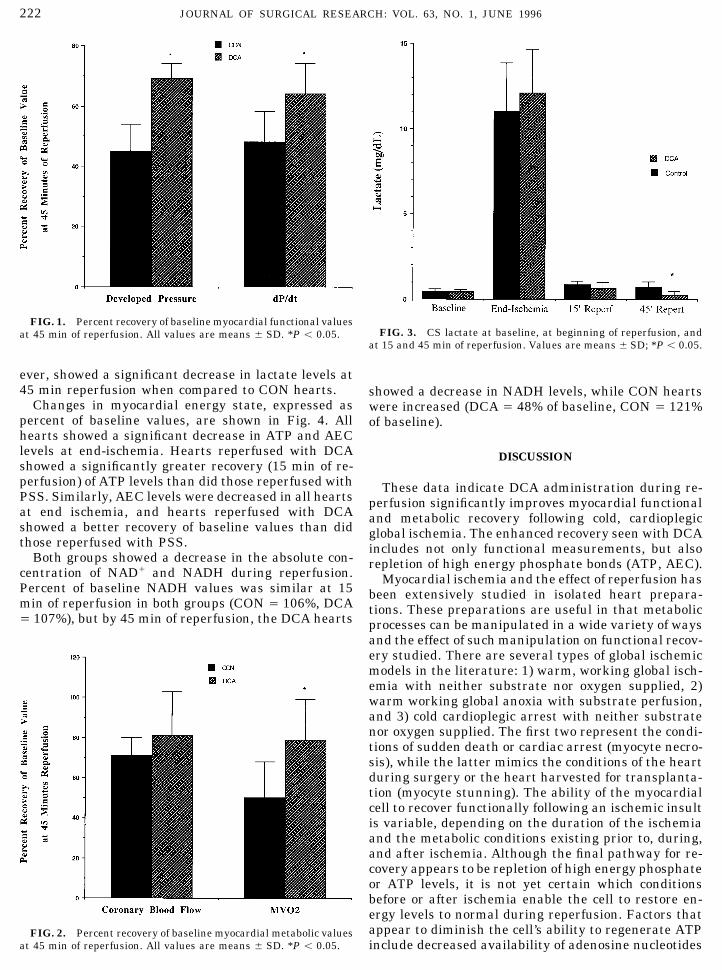
Metabolic recovery is shown in Fig. 2 (percent recov-baseline and reperfusion by measuring the slope of linearized EDPery of baseline values). Recovery of coronary blood flowversus EDV curves for each heart.
After a 30-min stabilization period, baseline measurements were was not different between CON hearts and those reper-made in each heart. Hearts were then rendered globally ischemic by fused with DCA. Myocardial oxygen consumption, how-interruption of the perfusion column immediately above the aortic ever, recovered to 79 { 20% of baseline value in DCAcannulation site. The intraventricular balloon was deflated, and 15
perfused hearts, while CON hearts recovered to onlyml of cardioplegia (modified St. Thomas’s solution, 47C) was adminis-50 { 18%. There were no differences between DCA andtered. All hearts were maintained in mild hypothermic arrest (347C)
by means of a circulating water jacket during 120 min of total isch- CON hearts in myocardial water content.emia. All hearts received 10 ml of cardioplegic solution every 30 min Coronary sinus (CS) lactate levels are shown in Fig.during ischemia. Multidose administration of cardioplegic solution 3. There were no differences between DCA and CONis used commonly in the clinical arena and is consistent with the
hearts in CS lactate levels at baseline, at end-ischemia,experimental studies we and others have reported previously.When the 2-hr ischemic period was ended, the hearts were reper- or at 15 min of reperfusion. DCA-perfused hearts, how-
AID JSR 4819 / 6n0e$$$842 05-24-96 12:58:07 srga AP: Surg Res
222 JOURNAL OF SURGICAL RESEARCH: VOL. 63, NO. 1, JUNE 1996
FIG. 1. Percent recovery of baseline myocardial functional valuesFIG. 3. CS lactate at baseline, at beginning of reperfusion, andat 45 min of reperfusion. All values are means { SD. *P õ 0.05.
at 15 and 45 min of reperfusion. Values are means { SD; *P õ 0.05.
ever, showed a significant decrease in lactate levels at45 min reperfusion when compared to CON hearts. showed a decrease in NADH levels, while CON hearts
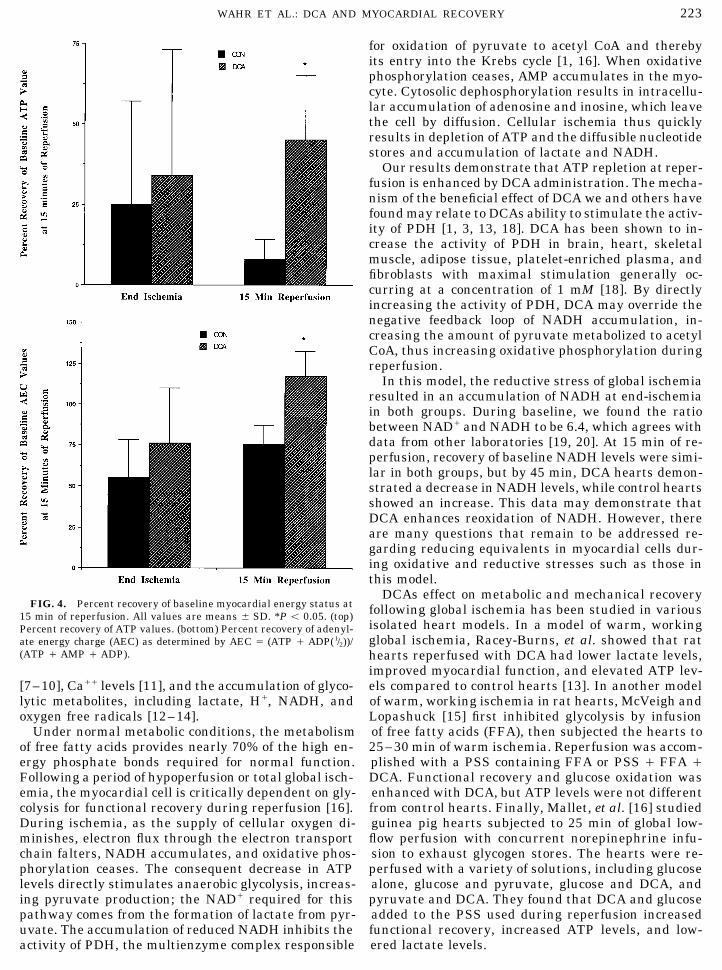
Changes in myocardial energy state, expressed as were increased (DCA Å 48% of baseline, CON Å 121%percent of baseline values, are shown in Fig. 4. All of baseline).hearts showed a significant decrease in ATP and AEClevels at end-ischemia. Hearts reperfused with DCA DISCUSSIONshowed a significantly greater recovery (15 min of re-perfusion) of ATP levels than did those reperfused with These data indicate DCA administration during re-PSS. Similarly, AEC levels were decreased in all hearts perfusion significantly improves myocardial functionalat end ischemia, and hearts reperfused with DCA and metabolic recovery following cold, cardioplegicshowed a better recovery of baseline values than did global ischemia. The enhanced recovery seen with DCAthose reperfused with PSS. includes not only functional measurements, but also
Both groups showed a decrease in the absolute con- repletion of high energy phosphate bonds (ATP, AEC).centration of NAD/ and NADH during reperfusion. Myocardial ischemia and the effect of reperfusion hasPercent of baseline NADH values was similar at 15 been extensively studied in isolated heart prepara-min of reperfusion in both groups (CON Å 106%, DCA tions. These preparations are useful in that metabolicÅ 107%), but by 45 min of reperfusion, the DCA hearts processes can be manipulated in a wide variety of ways
and the effect of such manipulation on functional recov-ery studied. There are several types of global ischemicmodels in the literature: 1) warm, working global isch-emia with neither substrate nor oxygen supplied, 2)warm working global anoxia with substrate perfusion,and 3) cold cardioplegic arrest with neither substratenor oxygen supplied. The first two represent the condi-tions of sudden death or cardiac arrest (myocyte necro-sis), while the latter mimics the conditions of the heartduring surgery or the heart harvested for transplanta-tion (myocyte stunning). The ability of the myocardialcell to recover functionally following an ischemic insultis variable, depending on the duration of the ischemiaand the metabolic conditions existing prior to, during,and after ischemia. Although the final pathway for re-covery appears to be repletion of high energy phosphateor ATP levels, it is not yet certain which conditionsbefore or after ischemia enable the cell to restore en-ergy levels to normal during reperfusion. Factors thatappear to diminish the cell’s ability to regenerate ATPFIG. 2. Percent recovery of baseline myocardial metabolic values
at 45 min of reperfusion. All values are means { SD. *P õ 0.05. include decreased availability of adenosine nucleotides
AID JSR 4819 / 6n0e$$$842 05-24-96 12:58:07 srga AP: Surg Res
223WAHR ET AL.: DCA AND MYOCARDIAL RECOVERY
for oxidation of pyruvate to acetyl CoA and therebyits entry into the Krebs cycle [1, 16]. When oxidativephosphorylation ceases, AMP accumulates in the myo-cyte. Cytosolic dephosphorylation results in intracellu-lar accumulation of adenosine and inosine, which leavethe cell by diffusion. Cellular ischemia thus quicklyresults in depletion of ATP and the diffusible nucleotidestores and accumulation of lactate and NADH.
Our results demonstrate that ATP repletion at reper-fusion is enhanced by DCA administration. The mecha-nism of the beneficial effect of DCA we and others havefound may relate to DCAs ability to stimulate the activ-ity of PDH [1, 3, 13, 18]. DCA has been shown to in-crease the activity of PDH in brain, heart, skeletalmuscle, adipose tissue, platelet-enriched plasma, andfibroblasts with maximal stimulation generally oc-curring at a concentration of 1 mM [18]. By directlyincreasing the activity of PDH, DCA may override thenegative feedback loop of NADH accumulation, in-creasing the amount of pyruvate metabolized to acetylCoA, thus increasing oxidative phosphorylation duringreperfusion.
In this model, the reductive stress of global ischemiaresulted in an accumulation of NADH at end-ischemiain both groups. During baseline, we found the ratiobetween NAD/ and NADH to be 6.4, which agrees withdata from other laboratories [19, 20]. At 15 min of re-perfusion, recovery of baseline NADH levels were simi-lar in both groups, but by 45 min, DCA hearts demon-strated a decrease in NADH levels, while control heartsshowed an increase. This data may demonstrate thatDCA enhances reoxidation of NADH. However, thereare many questions that remain to be addressed re-garding reducing equivalents in myocardial cells dur-ing oxidative and reductive stresses such as those inthis model.
DCAs effect on metabolic and mechanical recoveryFIG. 4. Percent recovery of baseline myocardial energy status at following global ischemia has been studied in various15 min of reperfusion. All values are means { SD. *P õ 0.05. (top)
isolated heart models. In a model of warm, workingPercent recovery of ATP values. (bottom) Percent recovery of adenyl-global ischemia, Racey-Burns, et al. showed that ratate energy charge (AEC) as determined by AEC Å (ATP / ADP(1/2))/
(ATP / AMP / ADP). hearts reperfused with DCA had lower lactate levels,improved myocardial function, and elevated ATP lev-els compared to control hearts [13]. In another model[7–10], Ca// levels [11], and the accumulation of glyco-
lytic metabolites, including lactate, H/, NADH, and of warm, working ischemia in rat hearts, McVeigh andLopashuck [15] first inhibited glycolysis by infusionoxygen free radicals [12–14].
Under normal metabolic conditions, the metabolism of free fatty acids (FFA), then subjected the hearts to25–30 min of warm ischemia. Reperfusion was accom-of free fatty acids provides nearly 70% of the high en-
ergy phosphate bonds required for normal function. plished with a PSS containing FFA or PSS / FFA /DCA. Functional recovery and glucose oxidation wasFollowing a period of hypoperfusion or total global isch-
emia, the myocardial cell is critically dependent on gly- enhanced with DCA, but ATP levels were not differentfrom control hearts. Finally, Mallet, et al. [16] studiedcolysis for functional recovery during reperfusion [16].
During ischemia, as the supply of cellular oxygen di- guinea pig hearts subjected to 25 min of global low-flow perfusion with concurrent norepinephrine infu-minishes, electron flux through the electron transport
chain falters, NADH accumulates, and oxidative phos- sion to exhaust glycogen stores. The hearts were re-perfused with a variety of solutions, including glucosephorylation ceases. The consequent decrease in ATP
levels directly stimulates anaerobic glycolysis, increas- alone, glucose and pyruvate, glucose and DCA, andpyruvate and DCA. They found that DCA and glucoseing pyruvate production; the NAD/ required for this
pathway comes from the formation of lactate from pyr- added to the PSS used during reperfusion increasedfunctional recovery, increased ATP levels, and low-uvate. The accumulation of reduced NADH inhibits the
activity of PDH, the multienzyme complex responsible ered lactate levels.
AID JSR 4819 / 6n0e$$$843 05-24-96 12:58:07 srga AP: Surg Res

224 JOURNAL OF SURGICAL RESEARCH: VOL. 63, NO. 1, JUNE 1996
and ouabain in isolated hearts from endotoxin-shocked rats. J.These results expand on our earlier findings, whichCardiovasc. Pharmacol.2(4): 379–386, 1988.demonstrated that functional recovery is enhanced
4. Stacpoole, P. W., Harman, E. M., Curry, S. H., Baumgartner,with DCA in a model of cold, cardioplegic arrest [6].T. G., Misbin, R. I. Treatment of lactic acidosis with dichloroa-
This earlier study also demonstrated that this benefi- cetate. New Engl. J. Med. 309: 390–396, 1983.cial effect is not due to changes in myocardial pH, and 5. Stacpoole, P. W., Lorenz, A. C., Thomas, R. G., Harman, E. M.indicated that the beneficial effect of DCA is not tem- Dichloroacetate in the treatment of lactic acidosis. Ann. Intern.
Med. 108: 58–631, 1988.perature dependent. This current study offers evidence6. Wahr, J. A., Childs, K. F., Bolling, S. F. Dichloroacetate en-that DCA enhances oxidative phosphorylation during
hances myocardial functional and metabolic recovery followingreperfusion.global ischemia. J. Cardiothorac. Vasc. Anes. 8: 192–197, 1994.
DCA has previously been shown to enhance lactate 7. Foker, J. E., Einzig, S., Wang, T. Adenosine metabolism andmetabolism in a wide variety of lactate acidosis models myocardial preservation. J. Thorac. Cardiovasc. Surg. 80: 506–[4, 5, 15, 17]. While we found that reperfusion with 516, 1980.
8. Reibel, D. K., Rovetto, M. J. Myocardial adenosine salvage ratesPSS containing DCA does decrease CS lactate levels,and restoration of ATP content following ischemia. Am. J. Phys-this effect was not seen until 45 min of reperfusion,iol. 237(2): H247–H252, 1979.while the functional and metabolic (ATP repletion) ef-
9. Bolling, S. F., Bies, L. E., Gallagher, K. P., Bove, E. L. Enhancedfects were already evident at 15 min of reperfusion.myocardial protection with adenosine. Ann. Thorac. Surg. 47:
DCA may also have additional inotropic properties. 809–815, 1989.When studies in a model of endotoxin-shocked rat 10. Bolling, S. F., Bove, E. L., Gallagher, K. P. ATP precursor deple-hearts without ischemia, DCA enhanced the inotropic tion and postischemic myocardial recovery. J. Surg. Res. 50:
629–633, 1991.effect of both amrinone and ouabain and independently11. Naylor, W. G. The role of calcium in the ischemic myocardium.increased glucose oxidation in these hearts [3]. The
Am. J. Pathol. 102: 262–270, 1981.enhanced oxygen consumption we noted with DCA ad-12. Neely, J. R., Grotyohann, L. W. Role of glycolytic products inministration could be either the cause or the effect of damage to ischemic myocardium: Dissociation of adenosine tri-
improved function. phosphate levels and recovery of function of reperfused ischemicIn summary, DCA is a promising new agent capable hearts. Circ. Res. 55: 816–824, 1984.
13. Racey-Burns, L. A., Burns, A. H., Summer, W. R., Shepherd,of enhancing myocardial functional recovery followingR. E. The effect of dichloroacetate on the isolated no flow ar-induced global ischemia. The improvement in func-rested rat heart. Life Sci. 44: 2015–2023, 1989.tional recovery appears to be due to enhanced restora-
14. Myers, M. L., Bolli, R., Lekich, R. F., Hartley, C. J., Roberts,tion of myocardial ATP levels, presumably through in-R. Enhancement of recovery of myocardial function by oxygen
creased oxidative phosphorylation. While many agents free-radical scavengers after reversible regional ischemia. Cir-have been found to be protective when given prior to culation 72(4): 915–921, 1985.the ischemia, DCA appears to be unique in that it is 15. McVeigh, J. J., Lopaschuk, G. D. Dichloroacetate stimulation
of glucose oxidation improves recovery of ischemic rat hearts.able to exert its beneficial effect when given at reperfu-Am. J. Physiol. 28: H1079–1085, 1990.sion. Further study will be required to delineate the
16. Mallet, R. T., Hartman, D. A., Bunger, R. Glucose requirementexact nature of its beneficial effect.for postischemic recovery of perfused working heart. Eur. J.Biochem. 188: 481–493, 1990.
17. Wahr, J. A., Ullrich, K., Bolling, S. F. Metabolic and hemody-REFERENCESnamic effects of dichloroacetate in the treatment of hypoxic lac-tic acidosis. J. Cardiothorac. Vasc. Anes. 8: 64–69, 1993.
1. Stacpoole, P. W. The pharmacology of dichloroacetate. Metabo- 18. Whitehouse, S., Cooper, R. H., Randle, P. J. Mechanism of acti-lism 38(11): 1124–1144, 1989. vation of pyruvate dehydrogenase by dichloroacetate and other
2. Mjos, O. D., Miller, N. E., Riemersma, R. A., and Oliver, M. F. halogenated carboxylic acids. Biochem. J. 142: 761, 1974.Effects of dichloroacetate on myocardial substrate extraction, 19. Kehrer, J. P., Lund, L. G. Cellular reducing equivalents andepicardial ST-segment elevation, and ventricular blood flow fol- oxidative stress. Free Radical Biol. Med. 17: 65–75, 1994.lowing coronary occlusion in dogs. Cardiovasc. Res. 10: 427– 20. Janero, D. R., Hreniuk, D., Sharif, H. M., Prout, K. C. Hydroper-436, 1976. oxide-induced oxidative stress alters pyridine nucleotide metab-
olism in neonatal heart muscle cells. Am. J. Physiol. 264:3. Burns, A. H., Summer, W. R., Burns, L. A. R., Shepherd,R. E. Inotropic interactions of dichloroacetate with amrinone C1401–1410, 1993.
AID JSR 4819 / 6n0e$$$843 05-24-96 12:58:07 srga AP: Surg Res