Embed Size (px)
Citation preview

This article was downloaded by: [129.130.252.222]On: 19 July 2014, At: 06:52Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
FisheriesPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/ufsh20
Dispelling Some Myths about HatcheriesRobin S. Waples aa Conservation Biology Division, National Marine Fisheries Service, Northwest FisheriesScience Center , 2725 Montlake Boulevard East, Seattle, WA, 98112, USA Phone:206/860-3254Published online: 09 Jan 2011.
To cite this article: Robin S. Waples (1999) Dispelling Some Myths about Hatcheries, Fisheries, 24:2, 12-21, DOI:10.1577/1548-8446(1999)024<0012:DSMAH>2.0.CO;2
To link to this article: http://dx.doi.org/10.1577/1548-8446(1999)024<0012:DSMAH>2.0.CO;2
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

I1 FISH CULTURE-PERSPECTIVE
Dispelling Some Myths about HatcheriesBy Robin S. Waples
ABSTRACTContributing to the controversies that have surrounded fish hatcheries in recent years are a num-ber of misconceptions or myths about hatcheries and their effects on natural populations. Thesemyths impede productive dialogue among those with differing views about hatcheries. Most ofthe myths include a measure of truth, which makes it difficult to recognize the elements that arenot true. Consideration of these myths leads to the following conclusions: (1) Hatcheries are intrin-sically neither good nor bad-their value can be determined only in the context of clearly definedgoals; (2) genetic changes in cultured populations can be reduced but not eliminated entirely; (3)empirical evidence exists of many adverse effects of hatcheries, but some risks have been overstat-ed; and (4) monitoring and evaluation programs are important but should not be used as a substi-tute for developing risk-averse hatchery programs in the first place. A key step in resolving someof the controversies will be moving toward agreement on a common version (rather than two ormore separate versions) of the realities about hatcheries. More efforts are needed in four majorareas: identifying goals, conducting overall cost:benefit analyses to guide policy decisions,improving the information base, and dealing with uncertainty.
was interviewed recently by a college studentdoing a project for her journalism class aboutwhether hatcheries are a viable solution to thedecline of Pacific salmon (Oncorhynchus spp.)
populations. One question she asked was, "Why arehatcheries so controversial?" Because progress inresolving the controversy is unlikely until we under-stand its origins, I begin this article by summarizingmy response to her question. Next, I identify severalmyths or misconceptions about hatcheries and theireffects on natural populations that I believe are im-peding progress and contributing to the controversy.Finally, I suggest some actions that should help ad-vance the dialogue on hatchery issues and helpresolve some of the considerable uncertainty sur-rounding this topic.
Most of the examples I discuss involve salmon.However, many of the same principles should apply tohatchery programs for other fish species as well. Al-though controversies about hatcheries occur on manylevels, I focus on those that involve biological issues orthe intersection of biological and nonbiological issues.Further, I consider primarily genetic (as opposed toecological) effects of cultured fish on natural popula-tions. This does not mean that ecological considera-tions are unimportant; rather, the topic is too complexand my expertise in that area too limited to do it jus-tice here. Finally, because the key issues involve bothfish culture and fisheries management, I emphasizehatchery programs rather than hatcheries per se.
12 * Fisheries
Why Are Hatcheries so Controversial?Controversies regarding fish hatcheries arise for a
number of reasons:(1) Collectively, salmon hatchery programs in the
Pacific Northwest are very large: hundreds of millionsof juvenile fish are released each year from hundreds offacilities. Substantial programs for freshwater speciesalso occur in all regions of the country, and interest inmarine aquaculture is growing. Because of their largesize and strong support from user groups (who rely onhatcheries to provide fish for commercial, recreational,and tribal harvest as well as jobs, hatchery programstend to resist change. Furthermore, in the minds ofmany, the failure of society to address the basic causesfor the decline of fish populations often leaves fish cul-ture as the only realistic means of providing harvestopportunities. This fact (along with the economic,logistic, and societal impediments to meaningful habi-tat restoration) contributes to a reluctance to undertakemajor changes to production programs.
(2) For more than a century, fish hatcheries havebeen viewed as a substitute for addressing root causesof declines in abundance-primarily loss or degrada-tion of habitat, blockage of migratory routes, and over-harvest. As the concept of a mitigation hatchery evolved,it became acceptable to sacrifice habitat and/or naturalpopulations as long as hatchery production was in-creased to compensate for the loss. This was seen bymany as a sound tradeoff, with hatcheries allowing fulluse of the watersheds while still maintaining fish pro-duction (Lichatowich et al. 1996). (These tradeoffs werenot universally endorsed and in some cases wereimplemented over the strong objections of the resourceagencies.) More recently, there has been a growingappreciation that long-term sustainability of salmon
Vol. 24, No. 2
Robin S. Waples is director of the Conservation BiologyDivision, National Marine Fisheries Service, NorthwestFisheries Science Center, 2725 Montlake Boulevard East,Seattle, WA 98112; 206/860-3254; [email protected].
��-�� --- �-��- � -��-� -- ���-
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTUJRE-PERSPECTIVE _
requires conservation of natural populations and theirhabitats (e.g., National Research Council 1995). As aresult of this paradigm shift, many hatchery programsare now being asked to minimize impacts to naturalpopulations. Not surprisingly, such changes are oftenresisted, particularly if they threaten the ability toachieve the objectives for which the programs weredesigned. In other cases, reforms to hatchery programshave been delayed because they must await changes inrelated areas such as harvest management.
(3) It is easy to identify risks that hatcheries pose fornatural populations; it is not so easy to predict whetherdeleterious effects will occur in any given situation or,if they do, how serious the consequences will be. Muchof the controversy surrounding hatcheries can be attrib-uted to different views of where the burden of proofshould reside with respect to this uncertainty. Someargue that, in accordance with the scientific method,we should assume that hatcheries will not cause harmunless it can be demonstrated in an hypothesis-testingframework. Others argue that this approach is too riskyfor use with natural resource issues because if we waitfor conclusive proof of harm to natural populations, itmay be too late to reverse the effects.
(4) Critics of hatcheries often do not agree amongthemselves on the nature and severity of the riskshatcheries pose and ways to minimize them. This cre-ates confusion among hatchery supporters, many ofwhom are willing to "do the right thing" if only some-one could tell them with some certainty (and consisten-cy) exactly what that is.
(5) There has been little systematic evaluation of bio-logical benefits of hatcheries and only sporadic evalua-tion of economic benefits. Evaluation also has beenhampered by a failure to clearly articulate programgoals. Basic questions-"Do hatcheries produce extrafish for harvest, or do they simply replace natural fishwith hatchery fish?" and "Are hatcheries cost-effectivein producing fish?"-have seldom been answered. Manynow express doubts about the value of hatcheriesbecause of uncertainties associated with these unan-swered questions. For mitigation hatcheries there is anadditional twist: Some view the issue of cost-effective-ness as irrelevant, arguing that the parties responsiblefor losses to natural populations should be obligated tocompensate for lost production regardless of the cost.
(6) A comprehensive assessment of the value ofhatcheries requires comparing fundamentally differentcurrencies: benefits to society (money, jobs, fishingopportunities) and to natural populations (conserva-tion) versus costs to society and risks to natural popu-lations (extinction, loss of fitness and diversity). Therehas been little effort to develop a framework underwhich these disparate ways of valuation can be jointlyconsidered in a comprehensive cost:benefit analysis.
(7) Hatchery supporters and detractors alike gener-ally agree there is considerable room for improvementin the operation of hatcheries to reduce effects on
February 1999
natural populations. However, no consensus exists onexactly what changes should be made and what theireffects would be. Some of the myths or misconceptionsdescribed in the next section contribute to the diver-gent views on this issue.
Common Myths and Misconceptionsabout HatcheriesAn error is more dangerous the more truth it contains.
-Henri-Fderdic Amiel, Journal Intime, 1883I won't attempt to address these complex issues com-
prehensively here; instead, I will focus on one aspect ofthe problem-misconceptions or myths about hatch-eries. These myths impede productive dialogue amongthose with differing views about hatcheries. As is gen-erally the case, most of these myths include a measureof truth-in some cases, quite a bit of truth. This makesit all the more difficult to recognize the elements thatare not true, and this factor has contributed to the cur-rent state of affairs, in which opposing sides are strong-ly entrenched in their own views of hatchery reality. Iwant to try to dispel some of these myths in the inter-est of advancing the dialogue about hatchery and nat-ural fish issues. The myths can be phrased in a varietyof ways, but each has been expressed more or less asstated here by some biologists, fish culturists, fisheriesmanagers, journalists, or other interested parties.
Myth 1: Hatcheries are inherently bad(or inherently good).
Neither of these positions leads to productive dia-logue, nor is either supported by a thoughtful consider-ation of the issue. Fish hatcheries are tools managerscan use to accomplish certain goals, and the value of ahatchery program can only be determined in the con-text of these objectives. A program that is well suited toachieve one type of goal (for example, harvest augmen-tation) might fail to achieve another (sustainability ofnatural populations). Unless goals are clearly articulat-ed and agreed on by the various parties involved, thereis little hope that arguments about the program's mer-its will be constructive. In my experience, both lack ofclarity about goals and lack of agreement regardinggoals are common to hatchery programs.
A related issue is the concept of identifying "appro-priate" uses for artificial propagation, with appropri-ateness evaluated with respect to the goals of the pro-gram. This is fine as far as it goes; the danger is that,once an appropriate goal is identified, scrutiny of theprogram often relaxes. This is an example of the mis-conception that "If a program has a worthy goal, itmust be beneficial." Clear goals are essential to adap-tive management, but they are not enough; hatcheryprograms must be evaluated rigorously as part of anongoing process to determine whether they are, infact, achieving their goals and are not compromisingother worthy goals.
Fisheries * 13
"-" --- '-"- l
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTURE-PERSPECTIVE
Myth 2: Risks posed by hatcheries can be avoided withbetter management.
This myth contains a good deal of truth. Considerableimprovements have been (and continue to be) made inboth fish culture and fisheries management, including bet-ter broodstock collection and mating protocols, more-nat-ural rearing conditions, wild-fish-friendly release strate-gies, and more focus on local broodstocks (Kapuscinskiand Miller 1993; Maynard et al. 1995). Doubtless, thesechanges can help reduce opportunities for direct and indi-rect effects on natural fish populations; the myth is thatthese changes will make the problems disappear altogeth-er. This is an illusion for two reasons: (1) Genetic changesin cultured populations cannot be avoided entirely and(2) many risks are negatively correlated, so efforts toreduce one risk simultaneously increase another. Becausethis myth is central to the controversy regarding hatch-eries, I will consider it in some detail.
Genetic changes in hatchery populations are associatedwith the terms domestication and domestication selection,which have been used in various (and sometimes confus-ing) ways in the literature. I'll offer definitions for theseterms here, not because these definitions are necessarilythe best, but because doing so should help clarify the
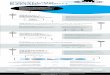
AHatchery v Wild Environments
Similarities* Water* Photoperiod
Differences* Food* Substrate* Density* Temperature* Flow regime* Competitors* Predators
B
Relativesurvival
0 Egg-smolt Smolt-adultLife stage
Figure 1 illustrates two reasons why some genetic change relative tothe wild population is inevitable as a result of fish culture. A. Theselective regime experienced by hatchery fish differs in many impor-tant respects from that experienced in the wild (after R. Reisenbichler,U.S. Geological Survey, Seattle, unpublished data). B. The mortalityprofile in cultured fish populations differs dramatically from that expe-rienced in the wild, both before and after release from the hatchery(schematic diagram after Waples 1991).
discussion that follows. Although some definitions ofdomestication focus on adaptation to a human-controlledenvironment (e.g., Kohane and Parsons 1988), it can beuseful to define the term more broadly to include anygenetic changes that result directly or indirectly fromhuman efforts to control the environment experienced bya population. Domestication thus describes a state or con-dition of a population and can be contrasted with theterm domestication selection, which refers to a process thatleads to domestication.
Campton (1995) identified three factors that lead togenetic change in cultured populations: (1) intentional orartificial selection for a desired trait (such as growth rate oradult body size), (2) selection resulting from nonrandomsampling of broodstock, and (3) unintentional or naturalselection that occurs in the hatchery environment. Camp-ton used the term artificial selection (selection due primarilyto humans) to distinguish factors 1 and 2 from factor 3,which is the only one he considered to represent domesti-cation selection. Busack and Currens (1995) also recog-nized the same three factors but considered all to be formsof domestication selection.
The distinctions made by Campton can be useful toseparate human and environmental causes of geneticchange; however, it also is important to remember that theoverall effects on cultured and natural populations are thesame regardless of how the components are categorized.For the latter reason, I favor the approach by Busack andCurrens because it includes a broader range of factorsunder a single term. In fact, I would go further and identi-fy a fourth component of domestication selection: (4) tem-porary relaxation during the culture phase of selection thatotherwise would occur in the wild. Busack and Currensconsidered this to be a component of their factor 3, but itcan be useful to identify this factor explicitly. Temporaryrelaxation of selection may not lead to genetic changewithin the hatchery population, but it does lead to geneticchange compared with the high mortality (some randomdeaths, but others selectively mediated) in the early lifehistory phases in natural populations. Taking all of thesefactors into consideration, I define domestication selection asany change in the selection regime of a cultured popula-tion relative to that experienced by the natural population.
Although it may be possible to eliminate intentionalselection from hatchery programs, it generally will not bepossible to eliminate factors 2-4 entirely because of twoinescapable facts: (1) The hatchery environment differs inmany ways from the natural environment, and (2) a suc-cessful hatchery program profoundly changes the mortali-ty profile of the population and results in more fish sur-viving than would have survived in the wild (Figure 1).Because of these factors, Busack and Currens (1995) con-cluded that some level of domestication selection isinevitable in a captive population. Although many factorscan help reduce the nature and extent of the resultinggenetic changes, they cannot be avoided entirely.
There are several corollaries to the myth regardingdomestication.
14 * Fisheries Vol. 24, No. 2
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTURE-PERSPECTIVE
Corollary 1: Domestication selection can be avoided ifthere is no mortality in culture.
This is a misconception because it fails to recognize thatthe genetic effects of fish culture can transcend the cultureperiod. Even if all progeny survive in a hatchery until timeof release, they will exhibit a range of values for traits suchas size, morphology, aggressiveness, swimming speed,metabolic rate, etc., and these characteristics can have aprofound effect on post-release survival and reproductivesuccess. For example, some fish will be larger than othersat the time of release because they hatch earlier, growfaster, or are better able to compete for food in the hatch-ery. In turn, fish that are larger at release may survive toadulthood at a higher rate than smaller fish (for example,because they are better at avoiding predation). Thisprocess will select for genotypes that produce large juve-nile fish under hatchery conditions, even if all fish surviveuntil time of release. Therefore, domestication selection forhatchery-adapted traits can occur even in the absence ofmortality during the culture phase. Furthermore, in addi-tion to any contribution to adaptation for hatchery condi-tions, high survival in culture represents a substantialrelaxation of selection that would occur in the wild.
Corollary 2: Domestication selection can be avoided iffamily size is equalized.
Equalizing family size in cultured populations can helpreduce domestication selection (Allendorf 1993), but limitsexist to the effectiveness of this strategy. Although in somecases it may be possible to equalize family size during thecaptive phase, the key to reducing domestication selectionis equalizing reproductive output into the next generation,and that is much more difficult. In a typical salmon hatch-ery, for example, 90% or more of the mortality occurs afterrelease of the juveniles, so efforts to equalize family size incaptivity can easily be nullified by events that occur laterin the life cycle (e.g., Geiger et al. 1997). Even in caseswhere it is possible to equalize reproductive success acrossfamilies, the result still will be genetic change relative tothe natural population, which typically will experiencestrong sexual selection for reproductive success. Finally,equalizing family size does not address the within-familycomponent of domestication selection.
Corollary 3: Any effects of domestication will be reversedby natural selection that occurs after thefish are released.
The process of domestication is easy to understand inspecies such as cattle and sheep, and few also doubt thatmost hatchery trout populations are domesticated. How-ever, many question whether domestication really occursin Pacific salmon hatcheries, which typically culture fishfor only 2-18 months of a life cycle that lasts severalyears. When comparing salmon to domesticated popula-tions using broodstock that spends its entire life in cap-tivity, it seems reasonable to ask, "How can hatcherysalmon be domesticated when, after release into the wild,they migrate thousands of miles to the sea and back everygeneration?"
No doubt natural selection will operate on the post-release population to help eliminate individuals that arenot well suited to survive in the wild, and this can helpoffset the effects of domestication selection. However, thetraits exposed to selection in the post-release juvenile-to-adult phase will generally not be the same as the early-life-history traits for which selection was relaxed duringthe period in culture. It is mathematically possible for themortality experienced after release to exactly compensatefor genetic change that occurs in the captive phase, but thechances that this will happen are extremely small. Further-more, even if this did occur, it would cancel out any bene-fit of the hatchery program. Therefore, we are led to theconclusion that a successful hatchery program-one thatproduces more fish than would have been produced in thewild-will always result in some genetic change to thehatchery population. As noted below, whether thesechanges will affect natural populations depends on severaladditional factors.
Corollary 4: Domestication selection can be avoided iffishare propagated for only a single generation.
This belief appears to be surprisingly widespreadamong hatchery managers and fisheries managers, but Ican find no theoretical or empirical support for it. Geneticchange can occur at many points within a single genera-tion as well as between generations. There is no mecha-nism that automatically erases genetic changes that occurwithin a single generation. Theory and empirical studiesagree that, in general, cumulative genetic changes willincrease with the length of domestication, but the changeswill not be zero for a single generation of culture.
A variation of this theme is the belief that geneticchanges can be avoided if only natural fish are taken intothe hatchery each generation for broodstock, thus allowingall returning hatchery fish to spawn naturally. The ideahere is that any effects of domestication from a single gen-eration in the hatchery will be erased in the ensuing gen-eration in the wild. This concept is similar to Corollary 3,except that the purifying selection does not occur until thesubsequent generation. This would be convenient if itwere true, but I am not aware of any empirical data tosupport it, and its theoretical basis appears to be weak.Genetic theory for populations that experience differentselective regimes but are connected by gene flow (e.g.,Karlin and McGregor 1972; Lythgoe 1997) indicates thatthe result of alternating generations between captive andnatural environments could lead to adaptation of theoverall population to the hatchery environment (andreduced fitness in the wild) as easily as it could preserveadaptations to the natural environment. In practice, thenet result would almost certainly be some combination ofthe two effects.
It may be that alternating hatchery and wild genera-tions is the best way to minimize effects of domesticationselection-this topic deserves additional study. However,there does not appear to be any scientific reason to believethat this strategy will eliminate domestication entirely.
February 1999 Fisheries * 15
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTURE-PERSPECTIVE
Corollary 5: Divergence of the hatchery population can beavoided if random sampling of broodstock is practiced.
Regarding this myth, it is important to recognize thateven a random sample is just that-a sample. Taking arandom sample will, on average, avoid directional bias,but in any particular case there is no guarantee that thesampled individuals will be representative of the pop-ulation as a whole (Hard et al. 1992). By chance, sometypes will generally be overrepresented while others areunderrepresented. The effect can be pronounced insmall samples, and diminishing as the sample sizeincreases. Of course, effects of broodstock sampling canbe avoided entirely by taking the whole population intocaptivity, but this extreme strategy substantially increasesother risks.
A related approach uses a form of stratified randomsampling to collect adults from throughout the duration ofthe spawning season in proportion to their occurrence inthe natural population. This strategy has some attractivefeatures. The difficulty is that although it may be possibleto collect a sample of broodstock that is more or less repre-sentative of the population as a whole for one trait (e.g.,run timing), it generally will not be possible to collect asample that is simultaneously representative of the popu-lation for all traits of interest (sex ratio, age, size, etc.).Therefore, it should be recognized that broodstock sam-pling will result in a captive population whose geneticcomposition differs from that of the natural population forone or more traits of interest. At best, one can hope to min-imize the extent of these genetic differences.
Risk TradeoffsInescapable risk tradeoffs associated with alternative
management strategies also make it impossible to avoid allundesirable effects of fish hatcheries. For example, al-though opportunities for genetic change in the hatcheryare reduced if fish are released early in their life cycle (e.g.,as fry or parr rather than as smolts), doing so increasesopportunities for competition with natural fish and alsoreduces the survival benefit provided by the hatchery.Similarly, taking a large fraction of the population forbroodstock minimizes founder effect in the hatchery popu-lation but exposes a larger proportion of the population tothe risks of fish culture and could affect the remaining nat-ural population demographically or genetically.
These inherent tradeoffs in risk are a major reason whyit is difficult to develop comprehensive guidelines forbroodstock collection, fish culture, and release strategies.There are no simple, universal answers to questions suchas, "Is it better to mark all hatchery fish in a supplementa-tion program to facilitate monitoring and enhance man-agers' ability to meet program guidelines (such as control-ling hatchery-wild spawning ratios), or is it better tominimize marking to reduce harm to the fish?" In thisand many other cases, no strategies exist that will simul-taneously minimize every type of risk. Guidelines can,and should, outline the various risks and tradeoffsinvolved, but which strategy should be preferred in any
16 + Fisheries
given situation must be evaluated on a case-by-case basiswithin the context of the program goals.
SummarySome level of genetic change relative to the natural
population cannot be avoided in a cultured population.The difference in selective environments experienced byhatchery and natural fish also means that changes that dooccur as a result of fish culture are unlikely to aid survivaland reproduction in the wild. It is important that fish cul-turists, fisheries managers, and fishery biologists considerthese realities in evaluating the appropriate nature andscale for hatchery programs.
What the above analysis does not tell us is how exten-sive the genetic changes in cultured populations will beand how strongly they will affect natural populations.This is a complex topic that needs further research, anddeveloping the information that will help us address thiskey information gap should be a major focus of effort inthe near future.
Myth 3: Hatcheries will always have unintended anddeleterious effects on natural populations.
Whether genetic change in cultured populations willaffect natural populations will depend on the nature of theprogram. In supplementation programs, which involve theintentional integration of natural and hatchery production,some genetic change to the natural population also isinevitable. Therefore, the net value of a supplementationprogram must be determined by weighing these geneticchanges (and potential ecological effects such as competi-tion, predation, etc.) against benefits (such as alleviatingsevere demographic and genetic risks of small popula-tions) that a successful program may be able to provide tonatural populations.
In contrast, harvest augmentation hatchery programscan (at least in theory) avoid deleterious effects on naturalpopulations if strong enough isolation of hatchery andnatural fish can be maintained. Whether such isolation isfeasible will vary widely from program to program andspecies to species. Here, I identify two incidental risks tonatural fish populations posed by hatcheries that canoccur but sometimes have been overstated in the popularpress. These are examples of a blurring of fact and specu-lation about the effects of hatcheries mentioned by Camp-ton (1995).
Corollary 1: Hatcheryfish stray more than wild fish.This assertion is not necessarily true. What is known
about straying in salmon can be summarized as follows(see Quinn 1993, 1997 for reviews and discussion):
* The proportion of cultured fish that return to sitesother than the hatchery or release site varies greatlyamong programs; some hatcheries consistently produce arelatively high proportion of strays, while others producefew. Limited data for natural fish suggest that they alsoexhibit considerable variation among populations in ratesof straying.
Vol. 24, No. 2
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTURE-PERSPECTIVE
* Whether, on average, hatchery fish stray more fre-quently than natural fish is an open question. Studies ofstraying in natural populations have been too limited toresolve this issue.
* Homing and straying are complex phenomena thatare imperfectly understood. Fish culture can be a factor inthe level of straying, but stock transfers and choice ofrelease site also can have a strong influence. Programs thatinvolve translocated stocks can lead to straying on a geo-graphic scale much greater than would occur naturally.
* Effects on natural populations are a function of theproportion of natural spawners that are hatchery fish, notthe fraction of the hatchery population that strays (Grant1997). Therefore, a hatchery program with a relatively lowstray rate can still substantially affect natural populations,particularly if the hatchery is large and/or the naturalpopulations are depressed. Conversely, a relatively highstray rate from a small hatchery might not substantiallyaffect a large natural population. Furthermore, the geneticimpact of stray hatchery fish can be less than their num-bers would indicate if they have reduced reproductivesuccess in the wild.
Corollary 2: Hatcheryfish will transfer disease andparasites to natural populations.
It is by no means inevitable that such transmission willtake place. For example, although several pathogens areabundant and cause chronic problems in Pacific salmonhatcheries, little or no direct evidence exists of transferfrom cultured to natural populations in spite of apparent-ly widespread opportunities for this to occur. That thesediseases also are endemic to natural populations, whichappear to have evolved strategies for dealing with them, isprobably a factor in this result. A cautionary note is that wehave relatively little information about the incidence of dis-ease in natural salmon populations and almost no historicalinformation about pristine populations. Therefore, it is diffi-cult to be sure that salmon hatcheries have not contributedto the current pathogen levels in natural populations.
In contrast, clear evidence exists that stock transfers offish can lead to the spread of exotic pathogens and para-sites into natural fish populations well out of the historicrange of the disease. In two recent examples [spread ofGyrodactylus salaris into populations of Atlantic salmon (Sal-mo salar) in Norway and spread of whirling disease (Myxo-bolus cerebralis) into populations of rainbow trout (Oncor-hynchus mykiss) in several areas of the western UnitedStates], pathogens were brought into contact with naturalpopulations that had no previous exposure to them, in eachcase with devastating results. Stock transfers of Atlanticsalmon and rainbow trout, respectively, appear to havebeen involved in the spread of these two diseases. Even inthese cases, however, the exact nature of the link withhatchery propagation may not be clear. For example, therecent appearance of whirling disease in Montana, whichdoes not currently have a trout stocking program, suggeststhat factors other than artificial propagation (e.g., anglers,their boats or equipment, or fish-eating birds) may have
February 1999
contributed to the spread of this pathogen, at least in thatstate (Bergersen and Anderson 1997; Potera 1997).
Recent efforts in many areas to limit the number andgeographic scale of stock transfers of fish should helpreduce the risk of disease transmission to natural popula-tions, but (as is apparent from the above example) vigi-lance on other fronts is necessary as well.
Myth 4: Objections to hatcheries are purely theoreticaland have no empirical basis.
This has been a theme of some recent pieces complain-ing about critics of hatcheries (e.g., Incerpi 1996; Rensel1997). I don't believe this view is supported by a review ofthe evidence. It is true that our understanding of thegenetic and ecological effects of hatcheries on natural pop-ulations is far from perfect. Substantial uncertaintiesremain about virtually every major issue, and periodicallyit is important, as Campton (1995) has done, to take stockof the situation and summarize the empirical data to helprefocus the debate. Nevertheless, I believe a review of thisbody of information shows that, in spite of the manyuncertainties, every major concern raised about hatcherieshas some empirical basis. Extensive literature exists on thistopic (e.g., see Waples 1991; Hindar et al. 1991; Campton1995 and other papers in the same volume), but I will givea few more recent examples.
* For several decades, Washington State has used amanagement strategy for winter steelhead (0. mykiss) thatinvolves advancing the run timing of hatchery popula-tions by several months (Crawford 1979). Although initial-ly designed to allow production of yearling smolts, the ad-vancement in run timing is now viewed as a fisheriesmanagement tool that minimizes opportunities for inter-breeding with natural fish and allows selective harvest ofhatchery fish. The program is based on a few domesticat-ed stocks that are widely distributed throughout the state.In a comprehensive allozyme study of steelhead popula-tions in Washington State, Phelps et al. (1994) found evi-dence for introgression of the nonnative hatchery stock intoa number of natural populations but found no evidence ofintrogression in other populations. The fitness consequencesof the interbreeding have not been critically evaluated.
* In an analysis of spawner-recruit relationships for 26steelhead populations in Oregon, Chilcote (1997) found astrong negative correlation between the proportion of nat-urally spawning hatchery fish and stock productivity. Al-though this correlation is striking and suggests that hatch-ery fish may be depressing fitness of natural populations,it does not necessarily represent a causal relationship sinceother factors (e.g., larger hatchery programs in areas wherefreshwater habitat cannot support healthy natural popula-tions) may have influenced the results. Chilcote did notfind a relationship between stock origin and productivity.
* In the Tucannon River in southeastern Washington, asupplementation program for the depressed run of springchinook salmon (0. tshawytscha) was initiated in the mid-1980s. Founded with local broodstock, this program aimsto maintain genetic integrity of the natural population and
Fisheries * 17
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTURE-PERSPECTIVE
has a strong research and evaluation component. In spiteof these efforts, data for the early 1990s showed that, com-pared to natural adults, returning hatchery fish wereyounger, were smaller for the same age, and had lowerfecundity for the same size (Bugert et al. 1992). The under-lying causes of these somewhat surprising phenotypicchanges are not known; however, even if the changes wereentirely an environmental response to hatchery conditions,they still would represent a significant single-generationreduction in productivity of the population.
* Reisenbichler (1997) has compiled available informa-tion regarding the effects of hatchery culture on fitness ofanadromous Pacific salmonids in the wild. These data-allfrom controlled experimental studies-are imperfect: Theyapply to a single species (steelhead) and in some casesconfound the effects of fish culture and stock transfers.Nevertheless, they are the best available data on this issue,and they demonstrate that fitness of hatchery-reared fishin the wild can be substantially reduced compared withnatural fish. Furthermore, the reductions in fitnessoccurred across all life stages and increased with the num-ber of generations of fish culture.
These examples are important but not entirely satisfy-ing; they raise as many questions as they answer. Never-theless, they emphasize a point made by Busack and Cur-rens (1995:77) regarding effects of hatchery fish on naturalpopulations: "We are unaware of rigorous researchdesigned to detect genetic impacts that has failed to findthem." What is lacking is a consensus on what constitutesa reasonable approach to this issue given the substantialuncertainty involved and the potentially major conse-quences of whatever actions are (or are not) taken.
Myth 5: It is not a hatchery problem; it is a fisheriesmanagement problem.
Campton (1995) and others (e.g., Rensel 1997) haveargued that critics of hatcheries often confound biologicalfactors intrinsic to hatcheries with effects of fisheries man-agement (stock transfers, selective breeding, extensive har-vest rates in mixed-stock fisheries). As a result, manyhatchery managers believe they have "become scapegoatsfor virtually every perceived negative biological effect associ-ated with the artificial propagation, release, and managementof anadromous salmonid fish" (Campton 1995:338). Thisargument has some merit. Adaptive management is mosteffective when the consequences of individual actions canbe evaluated and modified as appropriate. To the extent thatdistinguishing between biological and management fac-tors facilitates this process, it can be useful and productive.
However, we should be careful not to exaggerate thedichotomy between biology and management. No fishhatchery exists in a vacuum isolated from fisheries man-agement concerns; rather, every hatchery program isdesigned to meet one or more management objectives(e.g., harvest enhancement, mitigation, conservation).Some factors identified by Campton such as stock trans-fers and mixed-stock fisheries are primarily a function offisheries management rather than fish culture, but many
18 * Fisheries
others involve both. For example, selective breeding, whenit occurs, is carried out by fish culturists to achieve a fish-eries management objective. The effects on natural popula-tions from an action such as selective breeding are thesame whether one chooses to allocate this action to fishculture or fisheries management. In this case, the onlymeaningful unit to consider is the overall hatchery pro-gram, which encompasses both fish culture and fisheriesmanagement. Put another way, "Fish culture is as much apart of management as management is of fish culture"(Incerpi 1996).
It is unfortunate if hatchery managers and fish cultur-ists believe they are scapegoats in the controversies abouthatcheries. The concerns about hatcheries I have raisedhere and elsewhere are not directed at those involved inthe culture of fish-professionals conscientiously trying todo their job the best way they know how-but rathertoward the effects of hatchery programs on natural fishpopulations. To the extent that we can depersonalize thedebate about hatcheries and redirect energy to solving theproblems rather than trying to assess blame, the resourceas a whole will benefit.
Myth 6: Everything will be okay if we have a goodmonitoring program.
Although no one doubts the importance of monitoringand evaluation (M&E) for adaptive management, there is avery real danger in relying too heavily on M&E as a sub-stitute for meaningful and comprehensive risk manage-ment. The danger arises from three factors.
First, M&E for hatchery programs has limitations withrespect to both reliability and timeliness. Statistical powerto detect genetic effects of hatcheries can be relatively highfor selectively neutral molecular markers, which can pro-vide key information on gene flow between hatchery andnatural populations, individual reproductive success, andeffective population size (e.g., Waples et al. 1993). Howev-er, the most serious concerns regarding fish hatcheries in-volve fitness effects on phenotypic, behavioral, and lifehistory traits in natural populations. As discussed by Hard(1995; see also Peterman 1990), the power of even the mostambitious M&E program to statistically detect a fish cul-ture effect on traits such as these is likely to be very lowbecause the background noise level (natural variability inthe same traits) is typically very high. Furthermore, even ifsuch an effect is detected, it will generally occur only afterseveral (fish) generations of monitoring. This means thatartificial propagation could substantially harm naturalpopulations long before there is any reasonable expectationof being able to detect it. Of course, this does not mean thatdeleterious effects will always occur; however, it shouldgive pause to those willing to embark on a high-risk pro-gram in the expectation that they will be able to quicklyand surgically intervene at the first sign of an undesirableoutcome. Unfortunately, in most cases that is a myth.
Second, even if harm is found, there is no guaranteethat effective remedial action will be taken. To be effective,an adaptive management program should include a
Vol. 24, No. 2
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTURE-PERSPECTIVE
framework describing the range of possible outcomesalong with management actions that would be triggeredby each outcome. Too frequently, this has proven difficultto accomplish for hatchery programs. Once begun, hatch-ery programs have considerable momentum and may bedifficult to terminate or substantially change, even in theface of clear evidence that they are not accomplishing theirgoals. For example, Hilborn (1992) described a salmonhatchery in Oregon that, in spite of all attempts at remedi-al action, had been shown to be ineffective in producingadult fish. Nevertheless, it proved to be impossible toclose the hatchery because of a fear that doing so wouldindicate a lack of commitment to the resource. As dis-cussed by Hilborn, this is an example of displacement ofthe original goal of producing fish to the goal of demon-strating an effort to produce fish.
Third, fishery biologists often find themselves in a M&Ecatch-22 with respect to hatcheries: If they find no evidenceof deleterious effects, it is difficult to argue for restrictivemeasures; if they do find harm, some will argue that restric-tive measures are unnecessary because there is no pure nat-ural population to worry about any more. A variation of thisscenario is that by the time adverse effects become apparent,they (in combination with other factors) may have depressedthe natural population to such an extent that aggressivesupplementation is considered essential anyway.
These factors by no means diminish the importance ofM&E for hatchery programs. However, they demonstratethe folly of using M&E as an excuse for not rigorouslyaddressing all risks up front and designing a program thatis as risk-averse as possible from the outset.
Where Do We Go from Here?A key step in resolving some of the controversies regard-
ing fish hatcheries will be moving toward agreement on acommon version (rather than two or more separate versions)of the realities about hatcheries. In my view, actions are need-ed in four major areas: identifying goals, conducting over-all cost:benefit analyses to guide policy decisions, improv-ing the information base, and dealing with uncertainty.
GoalsTo begin with, we must not only achieve greater clarity
in articulating the goals of hatchery programs, but also en-sure the programs are developed to address fisheries man-agement needs. A given program may have a single majorgoal (e.g., to replace production lost through habitat block-age, to supplement an at-risk natural population), or it mayhave multiple goals (increase the abundance of naturallyspawning fish while augmenting harvest opportunities).
Second, it is important to determine whether the explic-it goals are inclusive enough. The following question shouldbe asked: "Is pursuit of the major goal of the program like-ly to compromise the ability to achieve other fisheries man-agement or societal goals?" If so, then the goals of the pro-gram should be modified or expanded to reflect these otherconsiderations. For example, the 1982 Fish and WildlifeProgram developed by the Northwest Power Planning
February 1999
Council (NWPPC 1982) identified as its interim goaldoubling the run size of salmon and steelhead in the Colum-bia River basin. As revised in 1994 (NWPPC 1994), the cur-rent goal is to double run size without loss of biologicaldiversity. This revision of the goal statement reflects the beliefthat long-term sustainability of fisheries resources withinthe basin depends on conservation of natural populations.
Third, fisheries managers, fish culturists, fishery biolo-gists, and conservation biologists should review the goalsand, if possible, agree on general principles that will guidefuture management decisions. This will establish a solidbasis for conducting the program under an adaptive manage-ment framework-if possible, using performance indicatorsto measure whether the program is adhering to the princi-ples and meeting the goals. Such a framework also shouldacknowledge that goals may change through time and pro-vide a mechanism for reevaluating the program if that occurs.
Comprehensive Cost:benefit AnalysisOnce the fundamental goals of a hatchery program are
agreed on, the next step is to identify its nature and scope.A wide variety of approaches is available, each of whichmay be appropriate under certain conditions. To deter-mine the most suitable type of program for a particular sit-uation, it is necessary to do a comprehensive cost:benefitanalysis. This analysis takes the following basic form:
Net benefit = [benefits to society + benefits to naturalpopulations + benefits to ecosystems] - [costs to society+ costs to natural populations + costs to ecosystems]
Conducting such an analysis is exceedingly challeng-ing. In the simplest scenario (use of artificial propagationto alleviate short-term extinction risk to natural popula-tions), the primary costs and benefits are in a common cur-rency, both being evaluated with respect to the natural pop-ulation. Even so, determining whether a particular programwill have a net benefit to the natural population is far fromeasy. Furthermore, a comprehensive analysis of supplemen-tation also should include additional costs such as effectson other species or the environment, or fiscal expendituresthat might otherwise be used for another purpose.
The analysis becomes much more complex if the goalsof the program include benefits to society as a wholerather than (or in addition to) benefits to natural popula-tions. Societal benefits might include those derived direct-ly from the hatchery program (more fish to harvest; morejobs for fishers, fish culturists, and fishery biologists; andmeeting legal or tribal treaty obligations) as well as bene-fits that accrue indirectly from activities (hydropower, log-ging, agriculture) that are permitted because they arecompensated for by the hatchery program. An analysis ofthis type requires comparing at least three types of curren-cies: fiscal expenditures to operate the program (measuredin dollars), benefits to society (only some of which can eas-ily be measured in dollars), and costs and benefits to nat-ural populations and the environment. Placing an econom-ic value on living natural resources is a complex process,and literature on this topic is limited.
Fisheries + 19
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTURE-PERSPECTIVE
To make significant progress on this issue will requirean effective, two-pronged attack. First, scientists must do abetter job quantifying the risks and potential benefits ofartificial propagation. For example, a typical question totackle would be, "If hatchery program A were implement-ed, what would be the likely effects (changes in fitness,changes in within- and among-population diversity) onpopulations B, C, and D, and what would be the short-and long-term consequences of these changes for sustain-ability of the resource?" These questions are difficult toanswer, but until scientists progress in this area, we cannotprovide managers with the information they need to prop-erly evaluate the biological consequences of alternativemanagement actions (Table 1).
Second, a framework needs to be developed to helpfisheries managers, policy makers, and social scientistsevaluate the disparate types of costs and benefits associat-ed with artificial propagation. Again, this will not be sim-ple, but some of the approaches from the field of decisiontheory could be applied usefully to this issue.
Research
Major gaps in our understanding occur for most of thekey processes associated with fish culture. Some criticalissues (e.g., domestication selection, the relative impor-tance of inbreeding depression and outbreeding depres-sion) are regularly identified by scientific workshops andpanels as high-priority research topics; however, such re-search is seldom funded because the necessary experimentsare expensive, time-consuming (often requiring severalfish generations), and logistically difficult.
Major new funding initiatives are unlikely in the cur-rent political climate. However, one approach is readilyavailable and could make a significant impact: Devote morehatchery facilities to research. This could involve either
20 Fisheries
setting aside portions of a number of hatcheries or devot-ing one or more facilities entirely to research. Doing sowould not only alleviate many logistical problems associ-ated with this type of research, it also would reduce the ex-pense because in most cases retrofitting existing hatcheriesto conduct research should be simpler and cheaper thanconstructing new facilities. On a smaller scale, productionprograms could incorporate more cooperative researchprojects at relatively modest cost. Many hatchery man-agers and fish culturists are willing, even eager, to partici-pate in research projects provided they understand theimportance of the project and have a stake in its success.
UncertaintyMore and better research is necessary but is not by itself
sufficient. Because new research will not resolve all uncer-tainties, and because in any event some critical informationis not likely to be available for many years, it also is essen-tial that we develop workable methods for dealing withuncertainty. For production hatcheries, the most criticalquestions are, "Given the inevitable uncertainty, whereshould the burden of proof reside? Should hatcheries bepresumed harmless unless proven otherwise (thus riskingirreversible losses to biotic integrity if deleterious effectsdo occur), or should hatcheries be used only very cautiously(thus risking major sacrifices of societal benefits that mayturn out to be unnecessary)?" Underscoring the impor-tance of this issue is the fact that, in many cases, it will beimpossible to accommodate all concerns regarding hatch-eries without major changes to existing programs and, insome cases, sacrificing legal mandates to produce fish. Forsupplementation programs this question has an additionaltwist: "Should hatcheries be used aggressively in supple-mentation because of their demonstrated ability to pro-duce more fish (thus risking deleterious effects to natural
populations that may outweigh thebenefits), or should they be usedsparingly because of a lack of empiri-cal evidence for long-term benefitsto natural populations (thus riskingloss of an opportunity to alleviatehigh short-term risk of extinction orloss of diversity)?" Judicious appli-cation of the precautionary prin-ciple in these situations requires thewisdom of King Solomon.
A considerable amount hasbeen written on how to deal withuncertainty in the fields of decisionanalysis and ecological risk assess-ment (e.g., Raiffa 1968; Lackey 1997),but it has not been used effectivelywith hatchery issues (see discussionof this issue in Currens and Busack1995). Two key points are worth men-tioning here. First, both risks andpotential benefits should be calcu-lated as a distribution of possible
Vol. 24, No. 2
Table 1 shows the type of quantitative analysis necessary to make a direct link between aparticular hatchery-wild interaction and the primary and secondary consequences for naturalpopulations. For simplicity, only the modal values (in bold) for stray rate and change in fit-ness are used to evaluate subsequent consequences. For example, this hypothetical hatcheryprogram is most likely to lead to a stray rate between 11% and 20%; if straying at that leveloccurs, the most likely consequence is a 6%-10% reduction in fitness of the natural popula-tion, and the most likely consequence of a fitness reduction of this magnitude is a510%-100% increase in chance of extinction in 100 years. All values shown are hypothetical.
ConsequencesEvent Primary SecondaryHatchery fish stray into Change in fitness Change in extinction riskbnatural populationsMagnitudea Probability Magnitude Probability Magnitude Probability
0 .05 > +10% .03 > 1,000% .051-5% .15 +6-+10% .07 501-1,000% .10
6-10% .25 -5 - + 5% .20 101- 500% .2511-20% .30 -6--10% .30 51- 100% .3521-40% .20 -11 - -20% .25 21- 50% .20
> 40% .05 < -20% .15 < 20% .05
aPercentage of naturally spawning fish that were reared in a hatcherybChance of extinction in 100 years
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014

FISH CULTURE-PERSPECTIVE
outcomes, not (as is more typically the case) as a single pointestimate (Table 1). Only by considering the full range of pos-sible outcomes and their associated probabilities can in-formed management and policy decisions be made. Second,when there is substantial uncertainty, the chances of making awrong decision increase, making it more important to careful-ly evaluate the consequences of being wrong. A key questionto ask is, "If we find that a mistake was made, can we reversethe consequences of the error and, if so, at what cost?").
AcknowledgmentsJeff Hard, Mike Ford, Craig Busack, and Don Campton
contributed significantly to many of the ideas expressed here.This document also benefited from discussions with, and com-ments by, Susan Bishop, John Colt, Ken Currens, Walt Dick-hoff, Ross Fuller, Howard Fuss, Linda Jones, Loren Miller,Mike Schiewe, Steve Smith, and two anonymous reviewers.
ReferencesAllendorf, E 1993. Delay of adaptation to captive breeding by
equalizing family size. Conserv. Biol. 7:416-419.Bergersen, E. P, and D. E. Anderson. 1997. The distribution and
spread of Myxobolus cerebralis in the United States. Fisheries22(7):6-7.
Bugert, R., K. Petersen, G. Mendel, L. Ross, D. Milks, J. Ded-loff, and M. Alexandersdottir. 1992. Lower Snake River com-pensation plan, Tucannon River spring chinook salmon hatch-ery evaluation plan. U.S. Fish and Wildlife Service, Boise, ID.
Busack, C. A., and K. P. Currens. 1995. Genetic risks and hazardsin hatchery operations: fundamental concepts and issues. Am.Fish. Soc. Symp. 15:71-80.
Campton, D. E. 1995. Genetic effects of hatchery fish on wildpopulations of Pacific salmon and steelhead: What do we real-ly know? Am. Fish. Soc. Symp. 15:337-353.
Chilcote, M. 1997. Conservation status of steelhead in Oregon.Oregon Department of Fish and Wildlife, Portland.
Crawford, B. A. 1979. The origin and history of the trout broodstocks of the Washington Department of Game. WashingtonState Game Department Fishery Research Report, Olympia.
Currens, K. P., and C. A. Busack. 1995. A framework for assess-ing genetic vulnerability. Fisheries 20(12):24-31.
Geiger, H. J., W. W. Smoker, L. A. Zhivotovsky, and A. J. Ghar-rett. 1997. Variability of family size and marine survival in pinksalmon (Oncorhynchus gorbuscha) have implications for con-servation biology and human use. Can. J. Fish. Aquatic Sci.54:2,684-2,690.
Grant, S., ed. 1997; Genetic effects of straying of non-nativehatchery fish into natural populations. U.S. Department ofCommerce, NOAA Tech. Mem. NMFS-NWFSC-30, Seattle, WA.
Hard, J. J. 1995. Genetic monitoring of life-history characters insalmon supplementation: problems and opportunities. Am.Fish. Soc. Symp. 15:212-225.
Hard, J. J., R. P. Jones, Jr., M. R. Delarm, and R. S. Waples. 1992.Pacific salmon and artificial propagation under the EndangeredSpecies Act. U.S. Department of Commerce, NOAA Tech.Mem. NMFS-NWFSC-2, Seattle, WA.
Hilborn, R. 1992. Can fisheries agencies learn from experience?Fisheries 17(4):6-14.
Hindar, K., N. Ryman, and E Utter. 1991. Genetic effects of cul-tured fish on natural fish populations. Can. J. Fish. Aquat. Sci.48:945-957.
Incerpi, A. 1996. Hatchery-bashing: a useless pastime. Fisheries21(5):28.
Kapuscinski, A. R., and L. M. Miller. 1993. Genetic hatcheryguidelines for the Yakima/Klickitat Fisheries Project. Co-Aqua, 2369 Bourne Avenue, St. Paul, MN.
Karlin, S., and J. McGregor. 1972. Application of method of smallparameters to multi-niche population genetics models. Theor.Pop. Biol. 3:186-209.
Kohane, M. J., and P. A. Parsons. 1988. Domestication: evolution-ary change under stress. Evol. Biol. 23:31-48.
Lackey, R. T. 1997. Is ecological risk assessment useful for resolv-ing complex ecological problems? Pages 525-540 in Stouder,D. J., P. A. Bisson, and R. J. Naiman, eds. Pacific salmon andtheir ecosystems: status and future opportunities. Chapmanand Hall, New York.
Lichatowich, J. A., L. E. Mobrand, and R. J. Costello, and T. S.Vogel. 1996. A history of frameworks used in the managementof Columbia River chinook salmon. In L. Mobrand, ed.Applied ecosystem analysis. Bonneville Power AdministrationReport DOE/BP 33243-1, Portland, OR.
Lythgoe, K. A. 1997. Consequences of gene flow in spatiallystructured populations. Genet. Res. 69:49-60.
Maynard, D. J., T. A. Flagg, and C. V. W. Mahnken. 1995. Areview of semi-natural culture strategies for enhancing thepostrelease survival of anadromous salmonids. Am. Fish. Soc.Symp. 15:307-314.
NRC (National Research Council). 1996. Upstream: salmon andsociety in the Pacific Northwest. National Academy Press,Washington, DC.
NWPPC (Northwest Power Planning Council). 1982, 1994.Columbia River Basin Fish and Wildlife Program, Portland, OR.
Peterman, R. M. 1990. Statistical power analysis can improvefisheries research and management. Can. J. Fish. Aquat. Sci.47:2-15.
Phelps, S. R., B. M. Baker, P. L. Hulett, and S. A. Leider. 1994.Genetic analysis of Washington steelhead: initial electrophore-tic analysis of wild and hatchery steelhead and rainbow trout.Fisheries Management Program Report 94-9, WashingtonDepartment of Fish and Wildlife, Olympia.
Potera, C. 1997. Parasitology: fishing for answers to whirling dis-ease. Science 278:225-226.
Quinn, T. P. 1993. A review of homing and straying of wild andhatchery-produced salmon. Fish. Res. 18:29-44.
1997. Homing, straying, and colonization. Pages 73-85 inGrant, S., ed. Genetic effects of straying of non-native hatcheryfish into natural populations. U.S. Department of Commerce,NOAA Tech. Memo. NMFS-NWFSC-30, Seattle, WA.
Raiffa, H. 1968. Decision analysis: introductory lectures on choiceunder uncertainty. Addison-Wesley Publishing Co., Reading, MA.
Reisenbichler, R. R. 1997. Genetic factors contributing to declinesof anadromous salmonids in the Pacific Northwest. Pages223-244 in Stouder, D. J., P. A. Bisson, and R. J. Naiman, eds.Pacific salmon and their ecosystems: status and future oppor-tunities. Chapman and Hall, New York.
Rensel, J. 1997. Comment on March 1997 North Pacific Interna-tional Chapter of the American Fisheries Society plenary ses-sion: incorporating hatcheries in ecosystem management. NorthPacific International Chapter AFS Newsletter 19(1):14-15.
Waples, R. S. 1991. Genetic interactions between hatchery andwild salmonids: lessons from the Pacific Northwest. Can. J.Fish. Aquat. Sci. 48(Suppl. 1):124-133.
Waples, R. S., O. W. Johnson, P. B. Aebersold, C. K. Shiflett,D. M. VanDoornik, D. J. Teel, and A. E. Cook. 1993. A geneticmonitoring and evaluation program for supplemented popu-lations of salmon and steelhead in the Snake River basin.Annual report of research. Bonneville Power Administration,Portland, OR.
February 1999 Fisheries * 21
Dow
nloa
ded
by [
] at
06:
52 1
9 Ju
ly 2
014