Embed Size (px)
Citation preview

Eur. J. Biochem.251, 5162521 (1998) FEBS1998
Divalent cation dependence and inhibition ofSchistosoma mansoniATP diphosphohydrolase by fluorosulfonylbenzoyl adenosine
Christiane R. TORRES1, Eveline G. VASCONCELOS2, Sergio T. FERREIRA1 and Sergio VERJOVSKI-ALMEIDA3
1 Departamento de Bioquı´mica Medica, Instituto de Cieˆncias Biome´dicas, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil2 Departamento de Bioquı´mica, Instituto de Cieˆncias Biologicas, Universidade Federal de Juiz de Fora, Juiz de Fora, Brazil3 Departamento de Bioquı´mica, Instituto de Quı´mica, Universidade de Sa˜o Paulo, Sa˜o Paulo, Brazil
(Received 28 August/27 October1997) 2 EJB 971241/4
Hydrolysis of ATP or ADP catalyzed by the ATP diphosphohydrolase ofSchistosoma mansonitegu-ment was measured in the presence of different cations. ATP diphosphohydrolase was stimulated bymicromolar concentrations of either Ca21 or Mg21, Ca21 producing threefold higher maximal activitiesthan Mg21. Kinetic studies indicated that Ca21 and Mg21 compete for the same binding site on theenzyme. The effect of covalent labeling of ATP diphosphohydrolase with the ATP analog fluorosulfonyl-benzoyl adenosine (FSO2BzAdo) was studied. Schistosome tegument was passed through with SephadexG-50 filtration centrifugation columns to remove endogenous nucleotides, and this was followed by label-ing with FSO2BzAdo. Incubation of ATP diphosphohydrolase with1 mM FSO2BzAdo for 1 h inhibitedATPase or ADPase activities by 60% and 50%, respectively. Addition of ATP together with FSO2BzAdoprovided greater than 90% protection against FSO2BzAdo inactivation, indicating that FSO2BzAdo bindsto an ATP-binding site on the ATP diphosphohydrolase. Furthermore, addition of FSO2BzAdo to a me-dium containing intact worms caused 30% and 50% inhibition of ATPase and ADPase activities, respec-tively, indicating that the ATP-binding site of diphosphohydrolase is accessible to FSO2BzAdo from theexternal surface ofS. mansoniworms.
Keywords:ecto-ATPase; ATP diphosphohydrolase;Schistosoma mansoni ;fluorosulfonylbenzoyl adeno-sine; calcium and magnesium dependence.
Schistosoma mansoniis one of the major etiological agents responses of the host that involve platelet activation and cytotox-icity (Bout et al.,1986; Tran et al.,1992) and cytolytic T lym-of schistosomiasis (Capron et al.,1982). S. mansonitegument
has been implicated in protecting the parasite from immune and phocyte reactivity (Capron et al.,1982). These responses mightinvolve ADP and/or ATP, which are known to be released byhemostatic responses (Ruppel and McLaren,1986; Saito,1991),
and it is actively involved in uptake of solutes and nutrients activated platelets and cytolytic T lymphocytes (Vargaftig et al.,1981 ; Filippini et al., 1990). The ability to evade such defense(Coles, 1984). An ATP diphosphohydrolase was shown to be
present on the outer surface of the tegument ofS. mansoni mechanisms could thus depend on hydrolysis of nucleotides onthe surface of the parasites. Inhibitors have been successfullyworms (Vasconcelos et al.,1993, 1996) and in schistosomules,
the early stage of the worm which develops following penetra- used in general for the characterization of enzymatic functions.However, no specific or efficient ATP diphosphohydrolase in-tion of cercariae through the skin (Vasconcelos et al.,1997).
This enzyme belongs to a new family of ATP diphosphohydro- hibitor has been reported. Fluorosulfonylbenzoyl adenosine (5′-FSO2BzAdo), an ATP analog that specifically alkylates aminolases that was recently identified (Vasconcelos et al.,1996;
Handa and Guidotti,1996; Verjovski-Almeida et al.,1997). ATP acid residues present in ATP-binding sites of a wide range ofenzymes (Allison et al.,1986), has been used to covalently mod-diphosphohydrolases share several common features, namely a
broad nucleotide specificity and micromolar dependence on ify the active site and/or inhibit ATP diphosphohydrolases frommammalian and fish tissues (Cote´ et al., 1992; Sevigny et al.,Mg21 and Ca21 (Plesner,1995; Ferreira et al.,1995). One of the
goals of the present work was to carry out a detailed character-1995; Martı et al.,1996). FSO2BzAdo has a configuration andmolecular size similar to that of ADP and ATP (Bennett et al.,ization of Mg21 and Ca21 dependence ofS. mansonienzyme.
The function ofS. mansoniATP diphosphohydrolase is not 1978) and reacts covalently with several classes of amino acidsincluding serine, tyrosine, lysine, and histidine. Here we reportclear at present. Schistosoma adult worms may live for several
years in the mesenteric vasculature of infected human host in- the characterization of FSO2BzAdo as an inhibitor ofS. mansoniATP diphosphohydrolase, which might permit its further utiliza-dividuals (Vermund et al.,1983), effectively evading cytotoxiction in the characterization of the role(s) of this enzyme in the
Correspondence toS. Verjovski-Almeida, Departamento de Bio- parasite.quımica, Instituto de Quı´mica, Universidade de Sa˜o Paulo, Caixa Postal26077, 05599-970 Sa˜o Paulo, SP, Brazil
Fax: 155 11 818 7986.E-mail : [email protected] MATERIALS AND METHODSURL: http://www.iq.usp.br/dbq/verjo/index.htm
Materials. ATP, ADP, ouabain, dimethyl sulfoxide, proteaseAbbreviation.FSO2BzAdo, 5′-p-fluorosulfonylbenzoyl adenosine.Enzyme.Apyrase (ATP diphosphohydrolase) (EC 3.6.1.5). inhibitors, and FSO2BzAdo were from Sigma. Sephadex-G-50
517Torres et al. (Eur. J. Biochem. 251)
(medium) was from Pharmacia. All other reagents were of thehighest analytical grade available.
Parasites. Adult S. mansoniworms were perfused eitherfrom Syrian hamsters or from mice infected six weeks pre-viously with approximately 350 cercariae each (Simpson et al.,1981).
Isolation of tegument. Isolated tegument was obtained fromlive parasites by incubation for10 min at 37°C in medium con-taining 2.7 mM KCl, 0.9 mM CaCl2, 0.5 mM MgCl2, 0.5µg/mlleupeptin, 0.07µg/ml pepstatin, 50µg/l soybean trypsin inhibi-tor, and 2µg/ml phenylmethylsulfonyl fluoride, as previouslydescribed (Vasconcelos et al.,1993). Isolated tegument wasstored in liquid nitrogen in 5 mM Tris/HCl, pH 7.4, 8% sucroseplus the protease inhibitors described above. The protein con-centrations were determined according to Lowry et al. (1951).
Assay of ADPase or ATPase activities.Activity measure-ments were routinely performed as previously described (Vas-concelos et al.,1993) in standard reaction medium containing50 mM Mops, pH 7.4, 5 mM CaCl2, 2 mM MgCl2, 150 mMNaCl, 5 mM KCl, 1 mM ouabain,10 mM NaN3, 0.0520.1mgprotein/ml and 3 mM of either ADP or ATP. The amount ofPi liberated was determined spectrophotometrically according toTaussky and Shorr (1953).
When micromolar concentrations of free Mg21 were tested,1mM EDTA was added to the reaction medium and the amounts
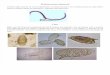
Fig.1. Ca2+ activation of S. mansoniATP diphosphohydrolase at dif-of MgCl2 and CaCl2 needed to give the desired free Mg21 and ferent Mg2+ concentrations.Hydrolytic activities were measured in theCa21 concentrations were calculated by an iterative computerpresence of 3 mM of either ATP (h) or ADP (s) with contaminantprogram that takes into account the binding constants of ATPMg21 (A), 5220 µM free Mg21 (B) or 1502200µM free Mg21 (C).(or ADP) and EDTA to Mg21 and Ca21 at pH 7.4 (Sorenson et Tegument (0.0320.05 mg protein/ml) was incubated in reaction medium
containing 50 mM Mops, pH 7.4,1mM ouabain,10 mM NaN3, 1mMal., 1986). EGTA was used instead of EDTA when micromolarEGTA, and the amount of CaCl2 needed to give the free Ca21 concentra-concentrations of free Ca21 were tested in the presence of milli-tions indicated in the figure. In B and C, the medium was supplementedmolar MgCl2, and the appropriate binding constants of EGTAwith MgCl2 to give the free Mg21 concentrations indicated above.were used in the program (Sorenson et al.,1986).
Labeling of tegument or intact adult worms with FSO2-BzAdo. To remove endogenous nucleotides, the isolated tegu-mental fraction was subjected to gel filtration through1-mlcentrifuge columns of Sephadex G-50 (medium) equilibratedwith 5 mM Mops, pH 7.4. The column effluents were collectedin microcentrifuge tubes (Kasahara and Penefsky,1978), andwere used for labeling with FSO2BzAdo and/or measurementsof enzyme activity.
Labeling was performed for various times at 37°C withFSO2BzAdo concentrations ranging from1027 M to 1023 M instandard reaction medium containing filtered tegumental mem-branes (final protein concentration 0.03 mg/ml). Nucleotides(ATP or ADP) were then added to a final concentration of3 mM, and nucleotide hydrolysis was carried out for 45 min at37°C. Reaction was arrested by addition of 0.1 M HCl and theamount of Pi liberated was measured colorimetrically accordingto Taussky and Schorr (1953). Tegumental samples, incubatedin the presence of1% (by vol.) dimethyl sulfoxide (the solvent
Fig.2. Mg2+ dependence in the absence of Ca2+. ATP diphosphohydro-in which stock FSO2BzAdo solutions were prepared), were usedlase activity was measured in the presence of 3 mM of either ADP (s)as controls to determine enzyme activities.or ATP (h) in reaction medium containing 0.03 mg/ml tegument protein,The time course of labeling was followed from 0 min to50 mM Mops, pH 7.4,1mM ouabain,10 mM NaN3, 1 mM EDTA, and
120 min at a tegumental protein concentration of 0.3 mg/ml anddifferent amounts of MgCl2 to give the free Mg21 concentrations indi-100 µM FSO2BzAdo, in the absence or in the presence of 3 mMcated in the figure.ATP. After labeling for the desired times, aliquots were with-drawn, diluted to 0.03 mg/ml, and assayed for remaining ATPase
activities were compared with that of control intact wormsactivity. Alternatively, freshly obtainedS. mansoniadult wormswhich were not labeled with FSO2BzAdo.were immersed in standard reaction medium as described above
in the presence of 3 mM FSO2BzAdo for 2 h at 37°C. Afterlabeling, excess reagent was washed away and the intact worms
RESULTSwere assayed for hydrolytic activity by addition of exogenousATP or ADP. Inorganic phosphate liberated was measured di-Ca2+-activated hydrolysis of ATP or ADP. Activation of S.
mansoniATP diphosphohydrolase by Ca21 was found to be de-rectly in the suspension medium with the colorimetric assay. The
518 Torres et al. (Eur. J. Biochem. 251)
Table 1. Inactivation of ATP diphosphohydrolase by FSO2BzAdo atdifferent Mg 2+ concentrations.ATPase activities were measured after2 h incubation in the absence or presence of100 µM FSO2BzAdo. AT-Pase activity assay media contained100µM free Ca21 (unless otherwiseindicated) plus the indicated free Mg21 concentrations, which were ad-justed by addition of variable amounts of CaCl2 and MgCl2 in the pres-ence of1 mM EGTA.
Condition ATPase activity Inhibition
no 100 µMFSO2BzAdo FSO2BzAdo
nmol Pi mg21 min21 %
Contaminant Mg21 420a 48 8910 µM Mg21 230a 42 82200µM Mg21 80a 38 533.2 mM Ca21, 900µM Mg21 97 54 44
a Data from. Fig.1.
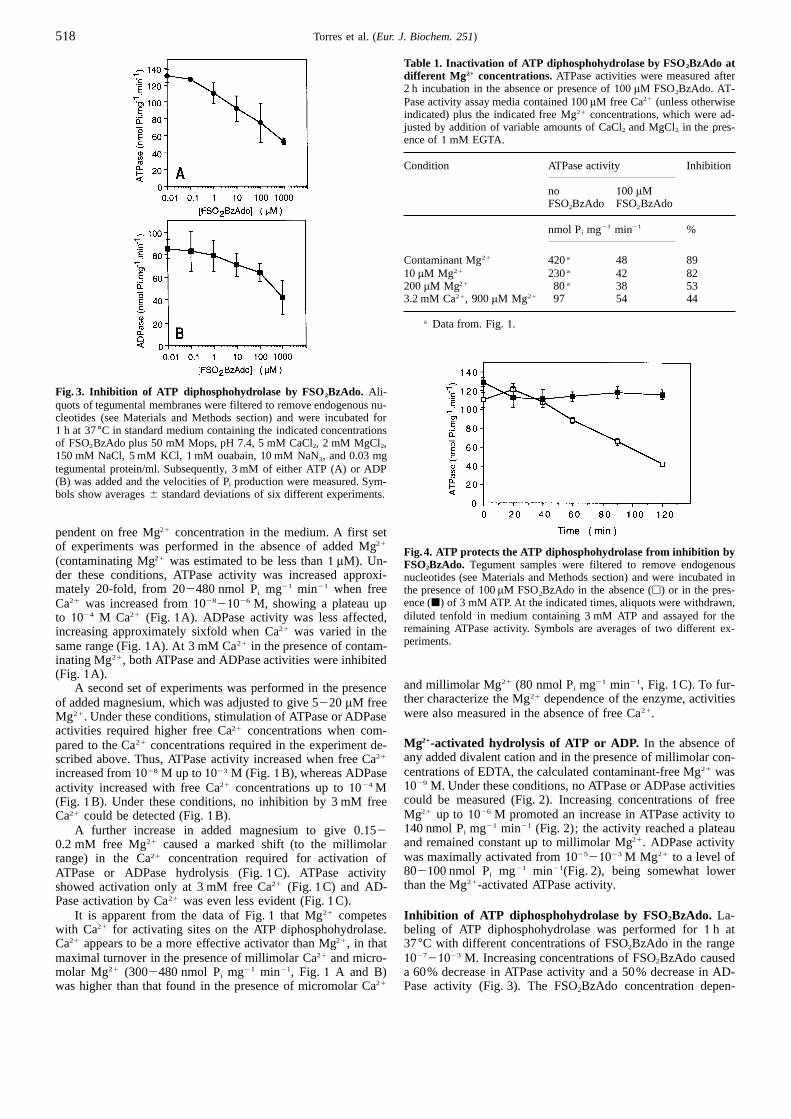
Fig. 3. Inhibition of ATP diphosphohydrolase by FSO2BzAdo. Ali-quots of tegumental membranes were filtered to remove endogenous nu-cleotides (see Materials and Methods section) and were incubated for1 h at 37°C in standard medium containing the indicated concentrationsof FSO2BzAdo plus 50 mM Mops, pH 7.4, 5 mM CaCl2, 2 mM MgCl2,150 mM NaCl, 5 mM KCl,1mM ouabain,10 mM NaN3, and 0.03 mgtegumental protein/ml. Subsequently, 3 mM of either ATP (A) or ADP(B) was added and the velocities of Pi production were measured. Sym-bols show averages6 standard deviations of six different experiments.
pendent on free Mg21 concentration in the medium. A first setof experiments was performed in the absence of added Mg21
Fig.4. ATP protects the ATP diphosphohydrolase from inhibition by(contaminating Mg21 was estimated to be less than1 µM). Un- FSO2BzAdo. Tegument samples were filtered to remove endogenousder these conditions, ATPase activity was increased approxi-nucleotides (see Materials and Methods section) and were incubated inmately 20-fold, from 202480 nmol Pi mg21 min21 when free the presence of100µM FSO2BzAdo in the absence (h) or in the pres-
ence (j) of 3 mM ATP. At the indicated times, aliquots were withdrawn,Ca21 was increased from102821026 M, showing a plateau updiluted tenfold in medium containing 3 mM ATP and assayed for theto 1024 M Ca21 (Fig. 1A). ADPase activity was less affected,remaining ATPase activity. Symbols are averages of two different ex-increasing approximately sixfold when Ca21 was varied in theperiments.same range (Fig.1A). At 3 mM Ca21 in the presence of contam-
inating Mg21, both ATPase and ADPase activities were inhibited(Fig. 1A).
and millimolar Mg21 (80 nmol Pi mg21 min21, Fig.1C). To fur-A second set of experiments was performed in the presencether characterize the Mg21 dependence of the enzyme, activitiesof added magnesium, which was adjusted to give 5220 µM freewere also measured in the absence of free Ca21.Mg21. Under these conditions, stimulation of ATPase or ADPase
activities required higher free Ca21 concentrations when com-Mg2+-activated hydrolysis of ATP or ADP. In the absence ofpared to the Ca21 concentrations required in the experiment de-any added divalent cation and in the presence of millimolar con-scribed above. Thus, ATPase activity increased when free Ca21
centrations of EDTA, the calculated contaminant-free Mg21 wasincreased from1028 M up to1023 M (Fig. 1B), whereas ADPase1029 M. Under these conditions, no ATPase or ADPase activitiesactivity increased with free Ca21 concentrations up to1024 Mcould be measured (Fig. 2). Increasing concentrations of free(Fig. 1B). Under these conditions, no inhibition by 3 mM freeMg21 up to 1026 M promoted an increase in ATPase activity toCa21 could be detected (Fig.1B).140 nmol Pi mg21 min21 (Fig. 2); the activity reached a plateauA further increase in added magnesium to give 0.152and remained constant up to millimolar Mg21. ADPase activity0.2 mM free Mg21 caused a marked shift (to the millimolarwas maximally activated from102521023 M Mg21 to a level ofrange) in the Ca21 concentration required for activation of802100 nmol Pi mg21 min21(Fig. 2), being somewhat lowerATPase or ADPase hydrolysis (Fig.1C). ATPase activitythan the Mg21-activated ATPase activity.showed activation only at 3 mM free Ca21 (Fig.1C) and AD-
Pase activation by Ca21 was even less evident (Fig.1C).It is apparent from the data of Fig.1 that Mg21 competes Inhibition of ATP diphosphohydrolase by FSO2BzAdo. La-
beling of ATP diphosphohydrolase was performed for1 h atwith Ca21 for activating sites on the ATP diphosphohydrolase.Ca21 appears to be a more effective activator than Mg21, in that 37°C with different concentrations of FSO2BzAdo in the range
102721023 M. Increasing concentrations of FSO2BzAdo causedmaximal turnover in the presence of millimolar Ca21 and micro-molar Mg21 (3002480 nmol Pi mg21 min21, Fig. 1 A and B) a 60% decrease in ATPase activity and a 50% decrease in AD-
Pase activity (Fig. 3). The FSO2BzAdo concentration depen-was higher than that found in the presence of micromolar Ca21

519Torres et al. (Eur. J. Biochem. 251)
Fig. 5. Labeling of intact S. mansoniadult worms by FSO2BzAdo.Freshly perfused adult parasites were incubated for 2 h at 37°C in stan-dard medium in the absence (hatched bars) or in the presence (solidFig.6. Three-dimensional structures of FSO2BzAdo and ATP. Thebars) of 3 mM FSO2BzAdo. After incubation, the reaction medium wasstructures of FSO2BzAdo and ATP were obtained by energy minimiza-removed and worms were placed in fresh assay medium without FSO2- tion of the potential energy of their configurations using BIOSYM/Mo-BzAdo for measuring ATP diphosphohydrolase activity in the presencelecular Simulations (San Diego, CA) software run on a Silicon Graphicsof either ATP (left bars) or ADP (right bars). The control activities inworkstation. A dielectric constant of 80 was used. The two moleculesthe absence of FSO2BzAdo were measured in a standard medium eitherwere placed in the figure with their adenosine moieties (leftmost part)without (light hatched bars) or with 3% (by vol.) dimethyl sulfoxidein the same orientation. It can be seen that the triphosphate moiety of(heavy hatched bars) which is the solvent used for preparing FSO2- ATP lies in an almost perpendicular orientation with respect to the fluo-BzAdo. rosulfonylbenzoyl moiety of FSO2BzAdo.
dence was similar for inactivation of either ATPase or ADPaseof S. mansoniATP diphosphohydrolase. Activity measurementsactivities.in the presence of contaminant Mg21 and high Ca21 (5 mM)Enzyme activities reported in Fig. 3 were determined in theresulted in a low ATPase/ADPase ratio (<1), whereas in thepresence of millimolar concentrations of Mg21 and Ca21. Addi-presence of contaminant Mg21 and low Ca21 (1 µM) the ratiotional experiments were performed to determine the degree ofwas<5 (Fig.1A). We have previously shown that two isoformsinactivation of ATP diphosphohydrolase by FSO2BzAdo whenof ATP diphosphohydrolase are present in the tegument, whichthe enzyme was maximally activated by micromolar Ca21 in thediffer in their sensitivities to non-ionic detergents (Vasconcelospresence of low Mg21. Table1 shows that labeling the enzymeet al., 1996). It is possible that the isoforms also differ in theirwith 100 µM FSO2BzAdo caused marked (53289%) inhibitiondivalent metal ion requirements, which would explain the com-of the activity when the assay was performed with contaminantplex pattern of activation and inhibition by metal ions exhibitedMg21, 10 µM and 200µM Mg21.by the enzyme (Fig.1).
ATP protects against inactivation by FSO2BzAdo. Inactiva- Physiological total ion concentrations of magnesium andtion of S. mansoniATP diphosphohydrolase by FSO2BzAdo was calcium in human serum are 0.521 mM and 2.322.7 mM,measured as a function of time in the presence of100 µM respectively. The assay medium in the experiments withFSO2BzAdo in the absence or presence of 3 mM ATP (Fig. 4).FSO2BzAdo-labeled ATP diphosphohydrolase (Figs 325) wasAddition of 3 mM ATP together with FSO2BzAdo during incu- used to represent ionic conditions similar to those encounteredbation provided greater than 90% protection against inactivationby the parasites in the host circulation. It is not known whether(Fig. 4), suggesting that the binding site for FSO2BzAdo was an the concentrations of ATP and/or ADP at the surface of parasitesATP-binding site on the ATP diphosphohydrolase. reach a high value, but it is conceivable that ADP released upon
activation of platelets (Day and Holmsen,1971) may reach localFSO2BzAdo inactivates ATP diphosphohydrolase on the concentrations as high as millimolar. Thus, changes in concen-outer surface of intact worms. Intact worms that were labeledtrations of free Ca21 and/or nucleotide might play a role in regu-for 2 h with 3 mM FSO2BzAdo were compared to control wormslating the ATPase and ADPase activities of the ATP diphospho-
that were kept in the absence of FSO2BzAdo, either with stan- hydrolase on the surface of parasites. The family of ATP diphos-dard medium or with medium supplemented with 3% dimethyl
phohydrolases (Vasconcelos et al.,1996; Handa and Guidotti,sulfoxide, the solvent used for preparing FSO2BzAdo solutions.1996; Verjovski-Almeida et al.,1997) includes enzymes with
After 2 h of reaction, the worms were resuspended in fresh me-different subcellular localizations and nucleotide specificities.dium containing ATP or ADP (without FSO2BzAdo) and the
Further studies involving conserved sequence analysis and deter-remaining ATP diphosphohydrolase activities were determinedmination of individual kinetic characteristics of distinct enzymes
by measuring the amount of inorganic phosphate liberated uponin the family will probably help in understanding the functionaladdition of ATP or ADP to the medium. Comparison of the
role and mechanism of regulation of the enzyme.amount of Pi liberated by intact control worms (Fig. 5) with thatThe requirement for low or contaminant Mg21 has been de-
liberated by intact worms labeled with FSO2BzAdo showed that scribed as a common characteristic of all ATP diphosphohydro-the ATPase activity was reduced to 70% of its original value
lases (Plesner,1995; Ferreira et al.,1995), which is quite distinctand that the ADPase activity was reduced to 50% of its originalfrom the requirement of millimolar Mg21 for activation of
value. These results suggest that the ATP-binding site of theplasma membrane Ca21-ATPase (Carafoli,1991). In the assaydiphosphohydrolase is accessible to FSO2BzAdo from the outer
of ATP diphosphohydrolase activities from different tissues andsurface ofS. mansoniparasites.species, it has been found that millimolar Mg21 may activatecontaminant Na1-K1 ATPase and adenylate kinase (Sarkis et al.,
DISCUSSION 1995). InS. mansonitegumental ATP diphosphohydrolase (Vas-concelos et al.,1993), no inhibition was observed with1mMThis study shows that the ionic composition of the medium
causes a marked change in the ratio of ATPase/ADPase activities ouabain, an inhibitor of Na1/K1-ATPase (Periyasamy et al.,

520 Torres et al. (Eur. J. Biochem. 251)
1983) and 3 mM vanadate, an inhibitor of ion-transport ATPase sent in human placenta, lung, skeletal muscle, kidney, and heart(Kaczmarek et al.,1996) it is possible that the physiological role(Carafoli,1991) or with 100µM Ap5A, an inhibitor of adenylate
kinase (Melnick et al.,1979). of the enzyme may differ among different tissues.Although the use of FSO2BzAdo is limited by the distinctSeveral possible reversible inhibitors of ATP diphosphohy-
drolase have been tested with limited success (Plesner,1995). possibility of its interaction with ATP-binding sites of other en-zymes on the surface of the parasite, the availability of such anOn the other hand, covalent irreversible modification of the en-
zyme with the ATP analog FSO2BzAdo gave a significant inhibi- irreversible inhibitor of the schistosomal ATP diphosphohydro-lase will likely turn this analog into an important tool for furthertion of S. mansoniATP diphosphohydrolase (Fig. 3, Table1), in
analogy to what has been reported for the ecto-ATPase of cyto- functional assays that may elucidate the role(s) of this enzymeon the surface ofS. mansoni.lytic T lymphocytes (Filippini et al.,1990), bovine aorta ATP
diphosphohydrolase (Coˆte et al.,1992), andTorpedoelectric or-This work was supported by grants fromFundacao de Amparo agan ATP diphosphohydrolase (Martı´ et al.,1996).
Pesquisa do Estado de Sa˜o Paulo (FAPESP),Programa de Apoio aoOur studies with intact worms labeled with FSO2BzAdoDesenvolvimento Cientı´fico e Tecnolo´gico (PADCT),Conselho Nacional(Fig. 5) showed that ATPase and ADPase activities were 30%de Desenvolvimento Cientıfico e Tecnolo´gico (CNPq) andFinanciadora
and 50% inhibited, respectively, confirming an external ATP-de Estudos e Projetos(FINEP). STF is a Howard Hughes Medical Insti-binding site onS. mansoni. Ecto-localization ofS. mansoniATP tute International Research Scholar.diphosphohydrolase has been shown by electron microscopy,which identified electron-dense lead phosphate deposits on the
REFERENCESouter surface ofS. mansonitegument upon hydrolysis of ATP orADP (Vasconcelos et al.,1993), and was confirmed by confocalAllison, W. S., Bullough, D. A. & Andrews, W. W. (1986) Identification
of essential residues in the F1-ATPases by chemical modification,immunofluorescence microscopy (Vasconcelos et al.,1996). AMethods Enzymol. 126, 7412761.possible explanation for the partial inhibition of ATPase and
Bennett, J. S., Colman, R. F. & Colman, R. W. (1978) Identification ofADPase activities on intact worms by FSO2BzAdo (Fig. 5) couldadenine nucleotide binding proteins in human platelet membranes bybe the turnover of tegumental proteins which would reduce theaffinity labeling with 5′-p-fluorosulfonylbenzoyl adenosine,J. Biol.amount of FSO2BzAdo-labeled ATP diphosphohydrolase. Alter-Chem. 253, 734627354.natively, an heterogeneity of ATP diphosphohydrolase isoformsBout, D., Joseph, M., Pontet, M., Vorng, H., Desle´e, D. & Capron, A.
could give rise to distinct conformations having ATP-binding (1986) Rat resistance to schistosomiasis: platelet-mediated cytotox-sites protected from FSO2BzAdo inactivation. In this respect, it icity induced by C-reactive protein,Science 231, 1532156.is relevant to analyze the three-dimensional models for FSO2- Candinas, D., Koyamada, N., Miyatake, T., Siegel, J., Hancock, W. W.,
Bach, F. H. & Robson, S. C. (1996) Loss of rat glomerular ATPBzAdo and ATP obtained by energy minimization (Fig. 6).diphosphohydrolase activity during reperfusion injury is associatedWhen the adenosine moieties of both analogs are placed in thewith oxidative stress reactions,Thromb Haemostasis 76, 8072812.same orientation (Fig. 6) it can be seen that the orientation of
Capron, A., Dessaint, J. P., Capron, M., Joseph, M. & Torpier, G. (1982)the fluorosulfonyl-benzoyl moiety of FSO2BzAdo is differentEffector mechanisms of immunity to schistosomes and their regula-from the orientation of the triphosphate moiety of ATP (Fig. 6).tion, Immunol. Rev. 61, 41266.Although the three-dimensional structure of FSO2BzAdo ob- Carafoli, E. (1991) Calcium pump of the plasma membrane,Physiol.
tained by energy minimization might be different from the struc- Rev. 71, 1292153.ture of the analog bound to the enzyme, the marked bend causedColes, G. C. (1984) Recent advances in schistosome biochemistry,Para-by the carbonyl moiety at the bond between the ribose and the sitology 89, 6032637.
Cote, Y. P., Ouellet, S. & Beaudoin, A. R. (1992) Kinetic properties ofbenzoyl moieties of FSO2BzAdo cannot be extended; thus it istype-II ATP diphosphohydrolase from the tunica media of the bovinenot expected that the fluorosulfonylbenzoyl moiety can ever lieaorta,Biochim. Biophys. Acta 1160, 2462250.in the same direction as the triphosphate moiety of ATP when
Day, H. J. & Holmsen, H. (1971) Concepts of the blood platelet releasethe two adenosines are superimposed. It is apparent that FSO2B-reaction,Ser. Haematol. 4, 3227.zAdo reactivity is directed towards the ATP-binding site of ATP
Ferreira, S. T., Vasconcelos, E. G. & Verjovski-Almeida, S. (1995) Thediphosphohydrolase (Figs 3 and 4) and the analog is probably ecto-ATPase of endo-parasites and of blood cells and vessels,Cien-orientated in the site by coordination through the adenosine moi- cia Cultura 47, 1512166.ety. The distinct orientation of the reactive fluorosulfonyl groupFilippini, A., Taffs, R. E., Agui, T. & Sitkovsky, M. V. (1990) Ecto-(Fig. 6) and eventually a lack of appropriate functional side ATPase activity in cytolytic T-lymphocytes,J. Biol. Chem. 265,
3342340.chain(s) at the ATP-binding site will probably cause the analogHanda, M. & Guidotti, G. (1996) Purification and cloning of solubleto react with the side chain of amino acids in the vicinity of the
ATP diphosphohydrolase (apyrase) from potato tuber (Solanum tu-ATP-binding site but outside the protein domain that normallyberosum), Biochem. Biophys. Res. Commun. 218, 9162923.coordinates the triphosphate moiety of ATP. It is conceivable
Kaczmarek, E., Koziak, K., Se´vigny, J., Siegel, J. B., Anrather, J., Beau-that partial inhibition by FSO2BzAdo results from the combineddoin, A. R., Bach, F. H. & Robson, S. C. (1996) Identification and
effects described above permitting that an ATP or ADP occupy characterization of CD39/vascular ATP diphosphohydrolase,J. Biol.the triphosphate-binding domain of an FSO2BzAdo-labeled di- Chem. 271, 33116233122.phosphohydrolase. Kasahara, M. & Penefsky, H. S. (1978) High affinity binding of monova-
S. mansoniis an endo-parasite that lives in the human vascu- lent Pi by beef heart mitochondrial adenosine triphosphatase,J. Biol.Chem. 253, 418024187.lature. Understanding the role of ATP diphosphohydrolase in
Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. (1951)vascular tissues may contribute to understanding the mechanismProtein measurement with the Folin phenol reagent,J. Biol. Chem.and functional significance of the enzyme on the surface of the193, 2652275.parasite. Recent work has provided evidence for the importance
Martı, E., Aranda, I. G. & Solsona, C. (1996) Inhibition of ATP-diphos-of ATP diphosphohydrolase in modulating vascular thrombosisphohydrolase (apyrase) of Torpedo electric organ by 5′-p-fluorosul-
in mammalians. Endothelial cell ATP diphosphohydrolase has fonylbenzoyladenosine,Biochim. Biophys. Acta 1282, 17224.the capacity to degrade the extracellular inflammatory mediatorsMelnick, R. L., Rubenstein, C. P. & Motzkin, S. M. (1979) MeasurementATP and ADP to AMP, thereby inhibiting platelet activation and of mitochondrial oxidative phosphorylation: selective inhibition ofmodulating vascular thrombosis (Candinas et al.,1996; Robson adenylate kinase activity by P1, P5-di(adenosine-5′)-pentaphosphate,
Anal. Biochem. 96, 7211.et al., 1997). Because ATP diphosphohydrolase mRNA is pre-

521Torres et al. (Eur. J. Biochem. 251)
Periyasamy, S. M., Huang, W. & Askari, A. (1983) Origins of the dif- Tran, A., Vanhe´e, D., Capron, A., Vorng, H., Braquet, P. & Joseph, M.(1992) Separate induction of human blood platelet aggregation orferent sensitivities of (Na11K1)-dependent adenosine triphosphatase
preparations to ouabain,Comp. Biochem. Physiol. 76B, 4492454. cytotoxicity by different concentrations of PAF-acether and throm-bin, Agents Actions 36, 39243.Plesner, L. (1995) Ecto-ATPases: identities and functions,Int. Rev. Cy-
tol. 158, 1412214. Vasconcelos, E. G., Nascimento, P. S., Meirelles, M. N., Verjovski-Al-meida, S. & Ferreira, S. T. (1993) Characterization and localizationRobson, S. C., Kaczmarek, E., Siegel, J. B., Candinas, D., Koziak, K.,
Millan, M., Hancock, W. W. & Bach, F. H. (1997) Loss of ATP of an ATP diphosphohydrolase on the external surface of the tegu-ment ofS. mansoni, Mol. Biochem. Parasitol. 58, 2052214.diphosphohydrolase activity with endothelial cell activation,J. Exp.
Med. 185, 1532163. Vasconcelos, E. G., Ferreira, S. T., de Carvalho, T. M. U., De Souza,W., Kettlun, A. M., Mancilla, M., Valenzuela, M. A. & Verjovski-Ruppel, A. & McLaren, D. J. (1986) Schistosoma mansoni: surface
membrane stability in vitro andin vivo. Exp. Parasitol. 62, 2232 Almeida, S. (1996) Partial purification and immunohistochemical lo-calization of ATP-diphosphohydrolase fromS. mansoni. Immunolog-236.
Saito, H. (1991) Evasion of host hemostasis by parasites: modulation of ical cross reactivities with potato apyrase andToxoplasma gondiinucleoside triphosphate hydrolase,J. Biol. Chem. 271, 221392blood coagulation and platelet function,J. Lab. Clin. Med. 117,
3412342. 22145.Vasconcelos, E. G., Torres, C. R., Martins, S. M., Verjovski-Almeida,Sarkis, J. J. F., Batastini, A. M. O., Oliveira, E. M., Frasseto, S. S. &
Dias, R. D. (1995) ATP diphosphohydrolase: an overview,Ciencia S. & Ferreira, S. T. (1997) The ATP diphosphohydrolase ofS. man-soni: ecto-localization and possible roles in host-parasite interac-Cultura 47, 1312136.
Sevigny, J., Cote, Y. P. & Beaudoin, A. R. (1995) Purification of pan- tions, inEcto-ATPases(Plesner, L., Kirley, T. L. & Knowles, A. F.,eds) pp. 2212226, Plenum Press, New York.creas type-I ATP diphosphohydrolase and identification by affinity
labeling with the 5′-p-fluorosulphonylbenzoyladenosine ATP ana- Vargaftig, B. B., Chignard, M. & Beneviste, J. (1981) Present conceptson the mechanism of platelet aggregation,Biochem. Pharmacol. 30,logue,Biochem. J. 312, 3512356.
Simpson, A. G. J., Schryer, M. P., Cesari, I. M., Evans, H. & Smithers, 2632271.Verjovski-Almeida, S., Vasconcelos, E. G., Ferreira, S. T., Kettlun, A.S. R. (1981) Isolation and partial characterization of the tegumental
outer membrane of adultSchistosoma mansoni, Parasitology 83, M., Mancilla, M. & Valenzuela, M. A. (1997) ATP diphosphohydro-lase fromSchistosoma mansonibelongs to a new family of apyrases,1632177.
Sorenson, M. M., Coelho, H. S. L. & Reuben, J. P. (1986) Caffeine inEcto-ATPases(Plesner, L., Kirley, T. L. & Knowles, A. F., eds)pp.1532159, Plenum Press, New York.inhibition of Ca accumulation by the sarcoplasmic reticulum in mam-
malian skinned fibers,J. Membr. Biol. 90, 2192230. Vermund, S. H., Bradley, D. J. & Ruiz-Tiben, E. (1983) Survival ofS. mansoniin the human host: estimates from a community-basedTaussky, H. M. & Shorr, E. (1953) A microcolorimetric method for the
determination of inorganic phosphorus,J. Biol. Chem. 202, 6752 prospective study in Puerto Rico,Am. J. Trop. Med. Hyg. 32, 104021048.685.