Embed Size (px)
Citation preview

Submitted M
anuscript
Diversity of Halimeda (Bryopsidales, Chlorophyta) in New
Caledonia: a combined morphological and molecular study.
Journal: Journal of Phycology
Manuscript ID: JPY-11-199-ART.R2
Manuscript Type: Regular Article
Date Submitted by the Author: 22-Mar-2012
Complete List of Authors: Dijoux, Laury; IRD, Coreus Verbruggen, Heroen; Ghent University, Phycology Research Group and Center for Molecular Phylogenetics and Evolution; Mattio, Lydiane; University of Cape Town, Botany department & Marine Research Institute Duong, Nathalie; IRD, Coreus
Payri, Claude; IRD, Coreus
Keywords: Biodiversity, Halimeda, New Caledonia, tufA, rbcL, coral reefs, taxonomic revision
Journal of Phycology

Submitted M
anuscript
1
Diversity of Halimeda (Bryopsidales, Chlorophyta) in New Caledonia: a combined 1
morphological and molecular study 1 2
Laury Dijoux ² 3
Institut de Recherche pour le Développement, U227, BPA5, 98848 Nouméa, New Caledonia 4
Tél. : (687) 26 07 32 6
Fax : (687) 26 43 26 7
Heroen Verbruggen 8
Phycology Research Group and Center for Molecular Phylogenetics and Evolution, Ghent 9
University, Krijgslaan 281 S8 (WE11), B-9000 Ghent, Belgium 10
Lydiane Mattio 11
Botany department & Marine Research Institute – University of Cape Town 7701 12
Rondebosch, South Africa. 13
Nathalie Duong 14
Institut de Recherche pour le Développement, U227, BPA5, 98848 Nouméa, New Caledonia 15
Claude Payri 16
Institut de Recherche pour le Développement, U227, BPA5, 98848 Nouméa, New Caledonia 17
18
Condensed running title: Halimeda diversity in New Caledonia19
Page 2 of 59 Journal of Phycology

Submitted M
anuscript
2
Abstract 20
21
Halimeda is a genus of calcified and segmented green macroalgae in the order Bryopsidales. 22
In New Caledonia, the genus is abundant and represents an important part of the reef flora. 23
Previous studies recorded 19 species identified using morphological criteria. The aim of the 24
present work was to reassess the genus' diversity in New Caledonia using morpho-anatomical 25
examinations and molecular analyses of the plastid tufA and rbcL genes. Our results suggest 26
the occurrence of 22 species. Three of these are reported for the first time from New 27
Caledonia: H. kanaloana, H. xishaensis, and an entity resembling H. stuposa. DNA analyses 28
revealed that the species H. fragilis exhibits cryptic or pseudocryptic diversity in New 29
Caledonia. We also show less conclusive evidence for cryptic species within H. taenicola. 30
31
Keywords: biodiversity, Halimeda, New Caledonia, tufA, rbcL, coral reefs, taxonomic 32
revision. 33
Abbreviation: tufA, elongation factor TU; rbcL, RUBISCO large subunit 34
35
Introduction 36
The genus Halimeda Lamouroux (Bryopsidales, Chlorophyta) is a well-studied group of 37
calcareous green macroalgae. It occurs worldwide in tropical and subtropical marine regions 38
but also in the Mediterranean Sea (Hillis-Colinvaux 1980, Kooistra et al. 2002). It is a 39
conspicuous component of coral reef ecosystems for its biomass and species richness, and 40
occurs from shallow habitats down to 150 m deep on the reef slope (Hillis-Colinvaux 1985, 41
Littler et al. 1986). Halimeda species are composed of calcareous segments connected to each 42
other by non-calcified medullar siphons at the nodal joint. The internal structure of segments 43
is composed of a network of medullar siphons ramifying into a cortex and terminating in 44
inflated peripheral utricles. The outermost utricles adhere to each other forming a continuous 45
Page 3 of 59Journal of Phycology

Submitted M
anuscript
3
membrane that envelops the internal space of the segment in which calcium carbonate 46
precipitates as aragonite (Hillis-Colinvaux 1980).They are important primary producers, 47
providing food and habitat to several coastal species of invertebrates and parrotfishes (Hillis-48
Colinvaux 1980, Hay et al. 1994). Halimeda is regarded as an important structural component 49
contributing to most of the biogenic carbonate of reefs from the Cenozoic to the present 50
(Payri, 1988, 1995; Taylor et al., 2009). The life history of Halimeda includes periodical 51
sexual reproduction during which the entire cytoplasm turns into reproductive cells and the 52
thallus dies, this is called the holocarpic reproduction (Meinesz, 1980). The calcified 53
segments then break down and crumble, contributing to sand formation. They also reproduce 54
clonally by elongating subterranean rhizoids and fragmentation (Walters et al. 2002). Species 55
display a diversity of forms and various habits colonizing sandy and rocky habitats. Some 56
species feature high intra-specific morphological variability, like H. heteromorpha N’Yeurt 57
which displays trilobed to reniform segment forms and has either a small bulbous attachment 58
structure anchoring it in sand or a small cushion of filaments fixing it to rock. Because of such 59
high levels polymorphism, identification of Halimeda species can be difficult. 60
After its description by Lamouroux (1812), the genus was subdivided in four species groups 61
based on external morphological criteria (Tunae, Pseudo-opuntia, Opuntia and Rhipsalis) by 62
J. G. Agardh (1887:80-89). The analyses of nodal structure and utricles initiated by Askenasy 63
(1888) and expanded by Barton (1901) and Hillis-Colinvaux (1959, 1980), led to the 64
recognition of five lineages: Rhipsalis (eight species), Opuntia (seven species), Halimeda (11 65
species), Micronesicae (three species) and Crypticae (one species). In the molecular 66
phylogenetic studies of Kooistra et al. (2002) and Verbruggen and Kooistra (2004), the 67
classification was further revised, including the reinstatement of section Pseudo-opuntia and 68
the merger of Crypticae with Micronesicae. Additionally, Halimeda melanesica Valet, which 69
was classified in Micronesicae by Hillis-Colinvaux (1980), was transferred to Rhipsalis. 70
Kooistra et al. (2002) further identified the presence of cryptic diversity in the genus. During 71
Page 4 of 59 Journal of Phycology

Submitted M
anuscript
4
the last decade, Verbruggen and co-workers carried out multi-gene phylogenetic studies to 72
further improve our understanding of species delimitation and evolutionary relationships in 73
Halimeda (Verbruggen et al., 2004, 2005a, b, 2006, 2007, 2009a, b). While the section-level 74
lineages have been well-characterized, many taxonomical ambiguities remain at the species 75
level. The number of formally recognized species is 44 (Guiry and Guiry, 2011), although 76
molecular tools indicate at least 52 evolutionarily significant units, some of which are 77
probably cryptic or pseudo-cryptic new species in need of description (Verbruggen et al. 78
2009b). 79
In New Caledonia, Halimeda is common to very abundant in many coralgal reef ecosystems. 80
Most of the sandy or muddy areas of lagoons and back reefs are colonized by species with 81
bulbous holdfasts that form extensive meadows that are sometimes mixed with seagrasses or 82
Caulerpa. These meadows have an important ecological function since they represent one of 83
the three major lagoon habitats structured by macrophytes together with seagrass and 84
Sargassum beds (Garrigue, 1995). Various species form thick sprawling mats or large 85
draperies on hard substrata of reef flats, coral pinnacles and the fore-reef slope. Halimeda 86
contributes significantly to the reef biogenic carbonate and represents one of the dominant 87
bioclastic constituents of lagoon sediments (Chevillon, 1996). The genus’ importance as a 88
structural component of Cenozoic reefs makes it a good indicator in sedimentary archives 89
and, along with coralline red algae, it is used to reconstruct paleoenvironment and climatic 90
histories (Massieux, 1976; Payri & Cabioch, 2004). When Valet (1966, 1968) published his 91
taxonomical work on Chlorophyta from New Caledonia he identified eight species including 92
the new species Halimeda melanesica Valet. Since then, several ecological and floristic 93
studies have been conducted (Garrigue and Tsuda, 1988; Garrigue, 1995) and during the past 94
decade, important new collections have been made in a wide variety of locations around New 95
Caledonia. The numerous specimens collected were identified to currently recognized taxa 96
based mostly on morphological and anatomical analyses. The most comprehensive inventory 97
Page 5 of 59Journal of Phycology

Submitted M
anuscript
5
to date lists 19 species (Payri 2007). However, the material available has been insufficiently 98
studied in many instances and the diversity may have been underestimated. Furthermore, very 99
little molecular systematic work has been done to verify species identification, and only 100
twelve sequences belonging to five species have been generated from New Caledonian 101
material (Kooistra et al., 2002, Verbruggen et al., 2005a, c , 2009a). 102
Considering the importance of Halimeda in the actual coralgal system and paleoecology of 103
New Caledonia it is clear that the available collections deserve to be completely re-evaluated. 104
Hence, in this paper we aim to revise the species diversity of Halimeda in New Caledonia by 105
characterizing and comparing the morphology and anatomy of New Caledonian morphotypes 106
and evaluating variation in the plastid DNA markers tufA and rbcL. Our strategy consists of 107
defining morphotypes in the collections based on morpho-anatomical analyses and 108
subsequently sequencing the tufA DNA barcodes of a majority of the specimens to evaluate 109
species boundaries, infer the validity of morphotypes and assess the diversity of the genus in 110
the study area. 111
112
Material and Methods 113
114
Site and specimens collections 115
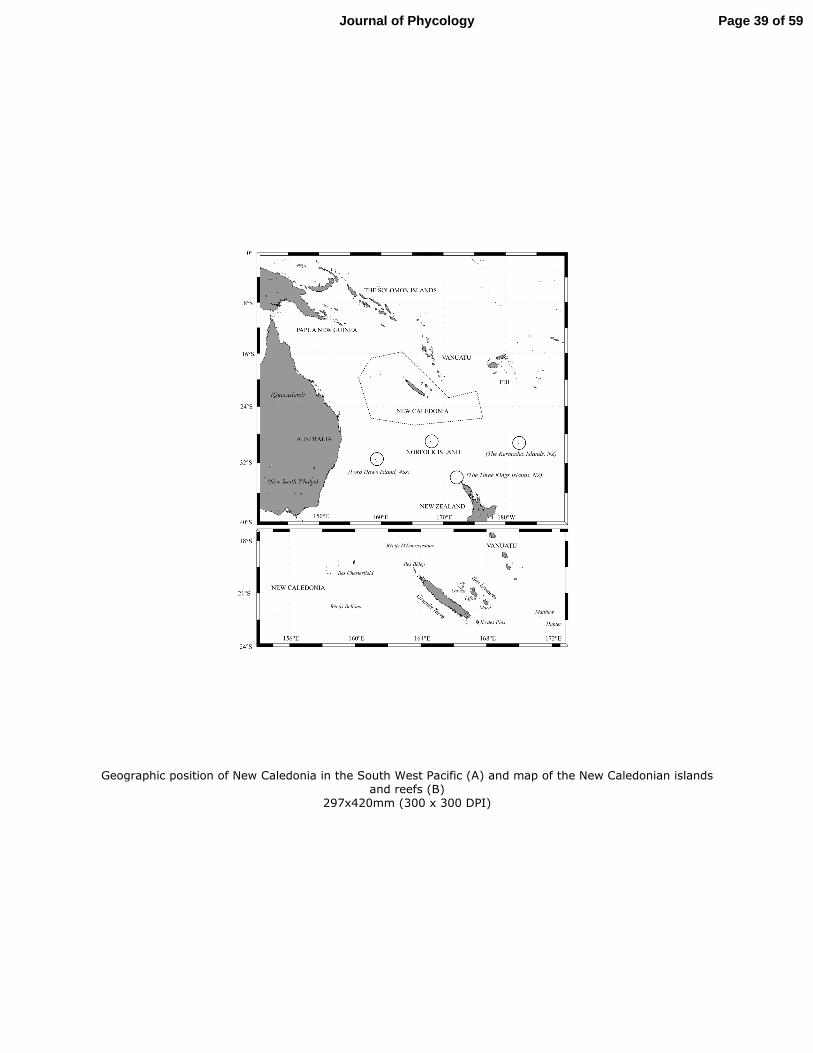
New Caledonia is located in the Southwest Pacific Ocean, between 18° and 23° S and 164° 116
and 167° E, about 1,500 km east of Australia (Fig. 1). It is composed of a main island, named 117
“Grande Terre” and several groups of smaller islands, coral reefs and lagoons. The New 118
Caledonian lagoon represents a total surface area of 24,000 km², which makes it one of the 119
biggest in the world (Andréfouët et al., 2004, 2009). It is surrounded by a continuous 1,600 120
km long barrier reef, located from 10 to 70 km away from the Grande Terre coast, and 121
expanding from the Entrecasteaux reefs to the northwest and the Isle of Pines to the south-122
Page 6 of 59 Journal of Phycology

Submitted M
anuscript
6
east. The isolated Chesterfield-Bellona reefs are situated towards the west, half way towards 123
the Australian coast, and the Loyalty Islands are located to the east of the main island. 124
A total of 155 New Caledonian samples pressed as herbarium vouchers and kept in the IRD-125
NOU phycological collection at IRD (Institut de Recherche pour le Développement) in 126
Noumea were considered for morphological analyses. They were collected mainly by SCUBA 127
from the intertidal to 60 m depth. All specimens were morphologically characterized and a 128
selection of them was used for DNA analyses from ethanol or silica gel preserved thallus 129
pieces. 130
The New Caledonian sampling sites included the southwest lagoon of Grande Terre (casual 131
collections between 2004 and 2010), Isle of Pines (BIODIP, November 2005), the Loyalty 132
Islands (BSM-Loyauté, March-April, 2005), La Côte Oubliée (CORALCAL1, March 2007); 133
the Chesterfield –Bellona-Bampton area (CORALCAL2, July 2008), and Le Grand Lagon 134
Nord (CORALCAL3, February 2009). 135
Additional collections from the Maldives (Baa Atoll, 2008), French Polynesia (Moorea, 2007, 136
Biocode Moorea Project –MBP) as well as samples lodged in the Ghent University algal 137
herbarium from Jamaica, Costa Rica, Philippines, Kenya, Indonesia, Maldives, Palau, 138
Australia, Tanzania, Nicaragua, Hawaii, French Polynesia, Thailand, Fiji, Micronesia, Wallis, 139
Chagos and Colombia were included in the analyses. 140
141
Morphology and anatomy 142
The morphological features of the samples were recorded following protocols described by 143
Hillis-Colinvaux (1980) and Verbruggen et al. (2005a, b). We registered both macroscopic 144
criteria such as holdfast type, shape of the thallus or segment form as well as anatomical 145
characters of the nodes and the utricles, which were observed with stereo and light 146
microscopy after dissection and decalcification using HCl 10%. Utricles from non-calcified 147
Page 7 of 59Journal of Phycology

Submitted M
anuscript
7
apical segments and basal segments were excluded from the analyses as recommended by 148
Verbruggen et al. (2005d). 149
Specimens were subsequently grouped into morphotypes (groups of similar morphology) 150
according to morphological analyses of the criteria listed above. All the morphological and 151
anatomical characters were evaluated simultaneously for all the specimens to place them into 152
their corresponding lineages, according to Verbruggen et al. (2004). Within each lineage, the 153
specimens were separated into different morphotypes looking the morphological and 154
anatomical discontinuities in the arrangement and size ranges of the segments, medullar 155
filaments, utricles and nodal fusions. Finally, the morphotype boundaries often, but not 156
always correspond to species delimitation. Subsequent DNA analyses were used to confirm or 157
reject species placement within these groups. 158
159
DNA extraction and PCR 160
A total of 141 samples preserved in ethanol, ten fresh samples and 60 dried samples from the 161
herbarium were processed. Samples preserved in ethanol were air dried for 12 hours prior to 162
DNA extraction. Few segments or part of one segment (approximately 0.5 cm long) were 163
crushed in liquid nitrogen and total DNA was extracted using the DNeasy Plant minikit 164
(Qiagen) according to the manufacturer’s instructions. PCR was carried out in a total volume 165
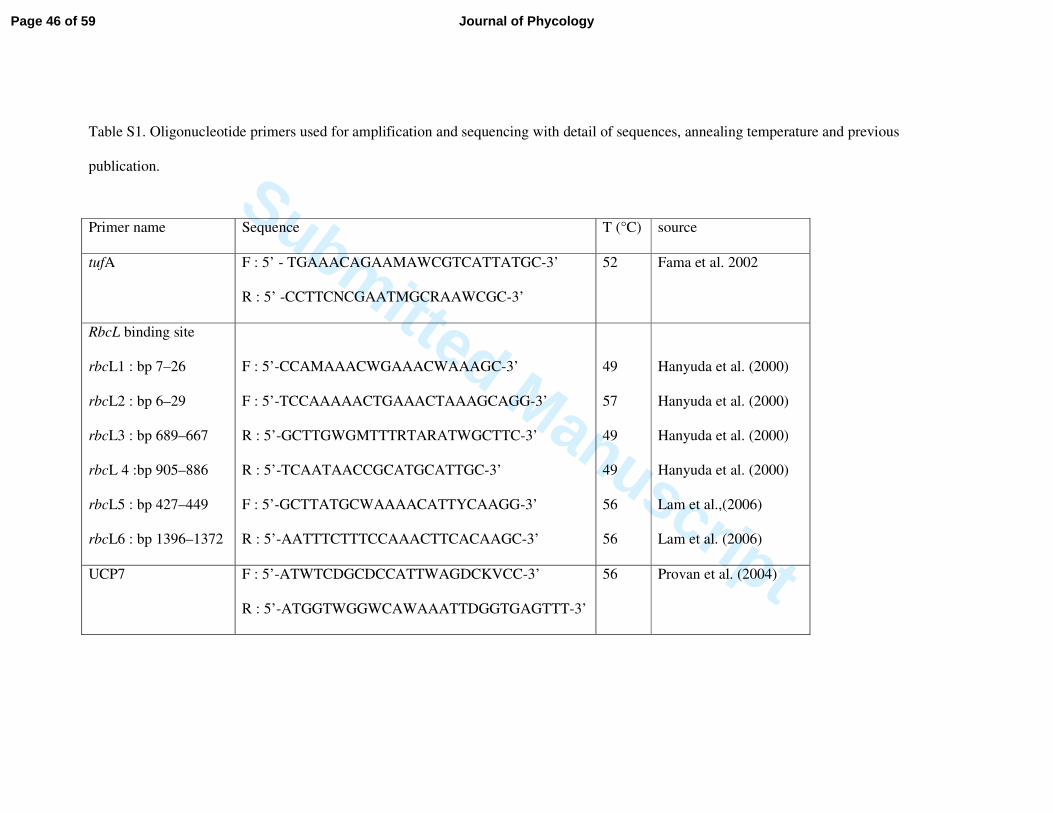
of 25 µL containing 2 µL of DNA extract, 0.2 µM of forward and reverse primers (Table S1), 166
0.2 mM of each dNTP, 2.5 µL of reaction buffer 10X (including 1.5 mM of MgCl2) and 1.25 167
U of Taq polymerase (Jump Start Red Taq, Sigma). PCR parameters included an initial 168
denaturation phase at 94°C for 2 min, 40 (tufA) or 35 (rbcL) cycles of 1 min at 94°C, 1 min at 169
annealing temperature (see Table S1), 1 min at 72°C and a final extension of 5 min at 72°C. 170
171
DNA sequencing 172
Page 8 of 59 Journal of Phycology

Submitted M
anuscript
8
The majority of samples was sequenced in both directions with the PCR primers by Macrogen 173
(Macrogen Inc., Seoul, Korea) using the BigDye TM terminator method. The other samples 174
were purified on column MinElute (Qiagen) and sequenced with an ABI Prism BigDye 175
Terminator v.3.1 cycle sequencing kit (Applied Biosystems) at the Plateforme du Vivant 176
(IRD, Nouméa, New Caledonia). The 20 µL reaction volume contained 0.8 µL of ready 177
reaction premix, 4 µL of BigDye sequencing buffer, 4 µM of each primer and 10 ng of 178
purified DNA. The sequencing reaction included a denaturation phase at 96°C for 1 min, 60 179
cycles of 15 sec at 98°C, 10 sec at 50°C and 4 min at 60°C. The sequencing products were 180
purified on Cephadex G50. Automated sequencing was performed on an ABI Prism 310 181
Genetic Analyzer (Applied Biosystems). 182
183
Sequence alignment and phylogenetic analyses 184
Sequences were aligned manually using the BioEdit sequence alignment editor (Hall 1999). A 185
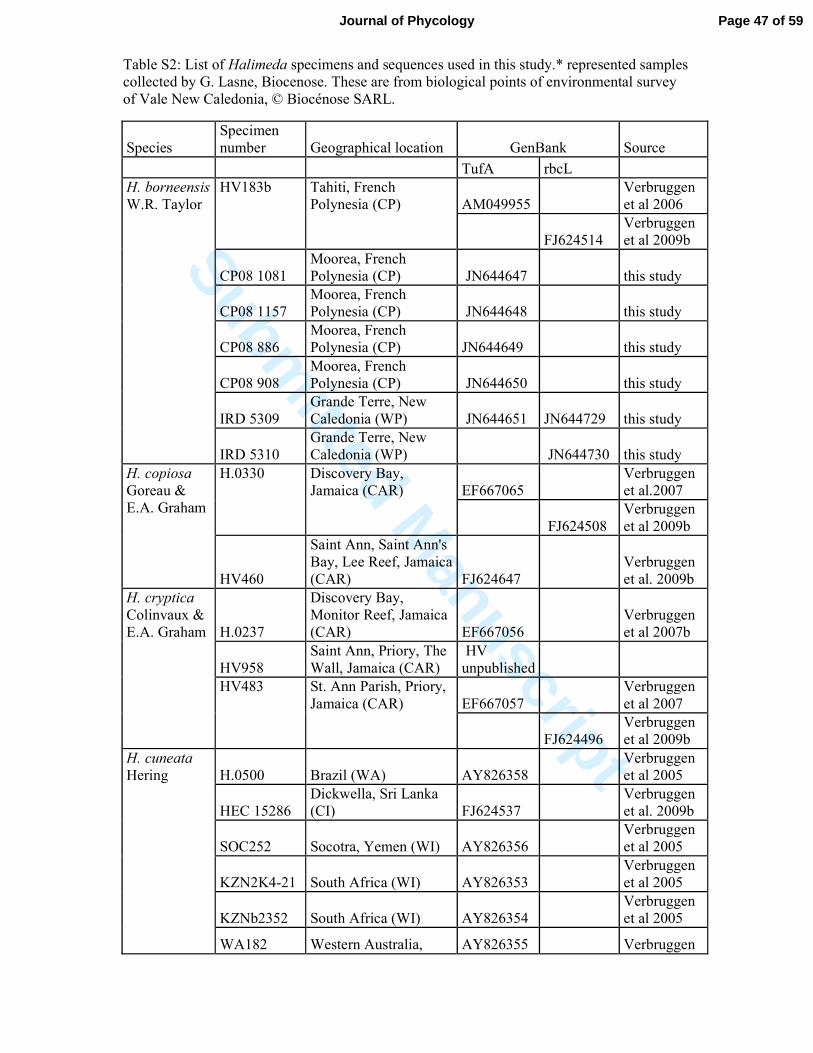
total of 124 tufA and rbcL sequences of Halimeda were downloaded from GenBank 186
(Hanyuda et al., 2000, Verbruggen et al., 2005 a, b, c, d, 2006, 2007 and 2009b, Lam et al., 187
2006, Curtis et al., 2008,Cocquyt et al., 2009) and added to our sequence alignments. 188
Our phylogenetic analyses consisted of two parts. First, a global phylogeny of the genus was 189
generated using a concatenated dataset of rbcL and tufA. The purpose of this analysis was to 190
show the major groupings in the genus and their relationships. Second, we performed analyses 191
of only tufA sequences for the five major groupings observed in the first step. These analyses 192
were performed to yield information about species delimitation and relationships within the 193
major groupings of the genus. 194
For the first analysis, tufA and rbcL were concatenated when both were available for a 195
sample. Phylogenies were inferred with neighbour joining (NJ), maximum parsimony (MP), 196
and maximum likelihood (ML) in MEGA4 (NJ, MP; Tamura et al. 2007) or PHYML (ML; 197
Guindon and Gascuel 2003). For ML analyses, the model of nucleotide substitution was 198
Page 9 of 59Journal of Phycology

Submitted M
anuscript
9
estimated using Findmodel (available at 199
http://www.hiv.lanl.gov/content/sequence/findmodel/findmodel.html). Support for nodes was 200
assessed by the bootstrap method (Felsenstein 1985) using 1,000 replicates for NJ and MP 201
analyses and 100 replicates for ML analyses. Nodes were considered as weak below 70%, 202
moderately supported between 71 and 84%, and strongly supported above 85%. A sequence 203
of Avrainvillea rawsonii was used as outgroup for the first analysis (Verbruggen et al. 2006). 204
For the lineage specific analyses of tufA, a sequence from a related lineage was used as 205
outgroup. The resulting lineage-specific haplotype trees were used to delimit species with a 206
method previously shown to yield accurate species boundaries in integrated morphological-207
molecular studies of Halimeda (e.g. Verbruggen et al 2005a,b). The method consists of 208
identifying well-supported clades that are preceded by a relatively long branch and have 209
comparably low sequence diversity within. Alignments and trees are available through 210
Treebase (http://www.treebase.org). 211
212
Results 213
214
Morphology 215
Morphological investigation of the 187 New Caledonian samples led to the recognition of 20 216
morphotypes (Table 1) into which each individual from New Caledonia could be 217
unambiguously assigned. These morphotypes, numbered A-T matched the descriptions of 218
respectively (A) H. cylindracea, (B) H. borneensis, (C) H. macroloba, (D) H. fragilis, (E) H. 219
micronesica, (F) H. lacunalis, (G) H. discoidea, (H) H. macrophysa, (I) H. magnidisca, (J) H. 220
taenicola, (K) H. gracilis, (L) H. minima, (M) H. opuntia, (N) H. distorta, (O) H. 221
heteromorpha, (P) H. kanaloana, (Q) H. xishaensis, (R) H. velasquezii, (S) H. gigas, and (T) 222
H. melanesica (Table S2). Morphological characters of the morphotypes are summarized in 223
Table 1. 224
Page 10 of 59 Journal of Phycology

Submitted M
anuscript
10
Morphotypes A, B, C, P could easily be distinguished from the others by the presence of 225
pores at the nodal fusion and a well-developed bulbous holdfast for anchorage in sandy 226
substrata.They could be distinguished from one another by their segment form. Morphotype A 227
showed cylindrical segments, simple or trilobed while morphotype B presented flat trilobed 228
segments and morphotype C had large reniform segments. Morphotype P presented basal 229
cylindrical segments similar to morphotype A but its segments were flat and trilobed. 230
Morphotype T also presented pores at the nodal fusion but lacked well-developed holdfasts. 231
Morphotypes D and E were distinguished by the absence of nodal fusion and by the 232
separation of the utricles after decalcification. Morphotype D had a basal segment with many 233
daughter segments in all directions (at 360° all around) whereas morphotype E showed a basal 234
“handlike” segment with numerous branching segments in its upper part (oriented about 235
180°). Both had a single holdfast. 236
Morphotypes F, G, H, I, J, Q and S featured erect thalli with a single holdfast and complete 237
nodal fusion of two or three siphons. Morphotype F could be distinguished by rounded, oval 238
or cuneate segments and utricles with a diameter less than 50 µm in surface view. 239
Morphotype G showed high polymorphism but constant and distinctive anatomical criteria 240
with inflated secondary utricles generally exceeding 100 µm in diameter. Morphotype H has a 241
thallus smaller than morphotype G and utricles visible to the bare eye and featuring a 242
characteristic honeycomb aspect before decalcification. Morphotype I had basal segments 243
forming a long and thin stipe with a well-developed bulbous holdfast. Morphotype J could be 244
distinguished from the other morphotypes by thick cuneate, cylindrical or oval segments and a 245
single rhizoidal holdfast; the anatomical studies pointed out an inflated tertiary layer of 246
utricles with a width over 110 µm. 247
Morphotype Q was very similar to morphotype G for the external morphology but 248
occasionally featured distinctive undulate segments. It had very large peripheral utricles and 249
Page 11 of 59Journal of Phycology

Submitted M
anuscript
11
relatively small secondary utricles compared to morphotype G (peripheral utricle length: 80 to 250
160 µm, width: 60 to 93 µm; secondary utricles length: 100 to 200 µm, width: 60 to 135 µm). 251
Morphotype S presented flat and glossy segments, most of which were reniform or oval, to 31 252
mm long and 42 mm broad. While the external morphology is reminiscent of morphotype G, 253
it could be easily distinguished by its internal anatomy because it possessed larger peripheral 254
utricles (peripheral utricle length: 92 to 187 µm, width: 130 to 240 µm). 255
Morphotype K presented a complete nodal fusion of pairs of siphons and could be 256
distinguished by a spreading, sprawling or bushy thallus, multiple attachment points, 257
numerous ramifications and long and thin branches with trilobed and cylindrical segments. 258
Morphotypes L, M, N and R shown identical nodal structures with partial fusion of two or 259
three siphons (rarely more) for a short distance. Morphotype L had an erect or pendent 260
curtain-like thallus with a single attachment to the substrata, branching in one plane except 261
sometimes at first segments. Morphotype R was similar to morphotype L but in surface view 262
its peripheral utricles were narrower than those of morphotype L (14 to 16 µm for R Vs 14 to 263
40 µm for L). Morphotype M was characterized by a bushy and compact thallus with multiple 264
attachment points to the substrata, small reniform segments heavily calcified sometimes 265
ribbed and brittle. Morphotype N possessed a sprawling thallus with prostrate and erect 266
portions, with multiple fixation points and reniform or trilobed segments of variable size and 267
organized in perpendicular planes. It can be distinguished from morphotype M by bigger and 268
twisted segments. 269
Morphotype O had a partial nodal fusion forming a single unit with very rare pores (small and 270
not clearly visible) and 3-4 utricle layers, a polymorphous thallus, a basal part sometimes 271
forming a pseudo-stalk, holdfast with matted rhizoids adapted to hard substrata. Segments 272
were small, mostly trilobed but occasionally reniform. 273
274
DNA analyses 275
Page 12 of 59 Journal of Phycology

Submitted M
anuscript
12
A total of 223 tufA and 52 partial rbcL sequences, including data downloaded from GenBank 276
(109 and 15 respectively), were aligned. New sequences were submitted to GenBank; 277
accessions are listed in Table S2. The tufA alignment was 857 bp, including gaps. The partial 278
rbcL sequences (first 689 bp) were obtained with the primer pair rbcL1-rbcL3 (Table S1). 279
Other primer pairs did not provide consistent results and were thus not analyzed further. NJ, 280
MP, and ML analyses produced similar tree topologies for both markers analyzed either 281
separately or in combination. A summary of the tree resulting from the ML analysis applied to 282
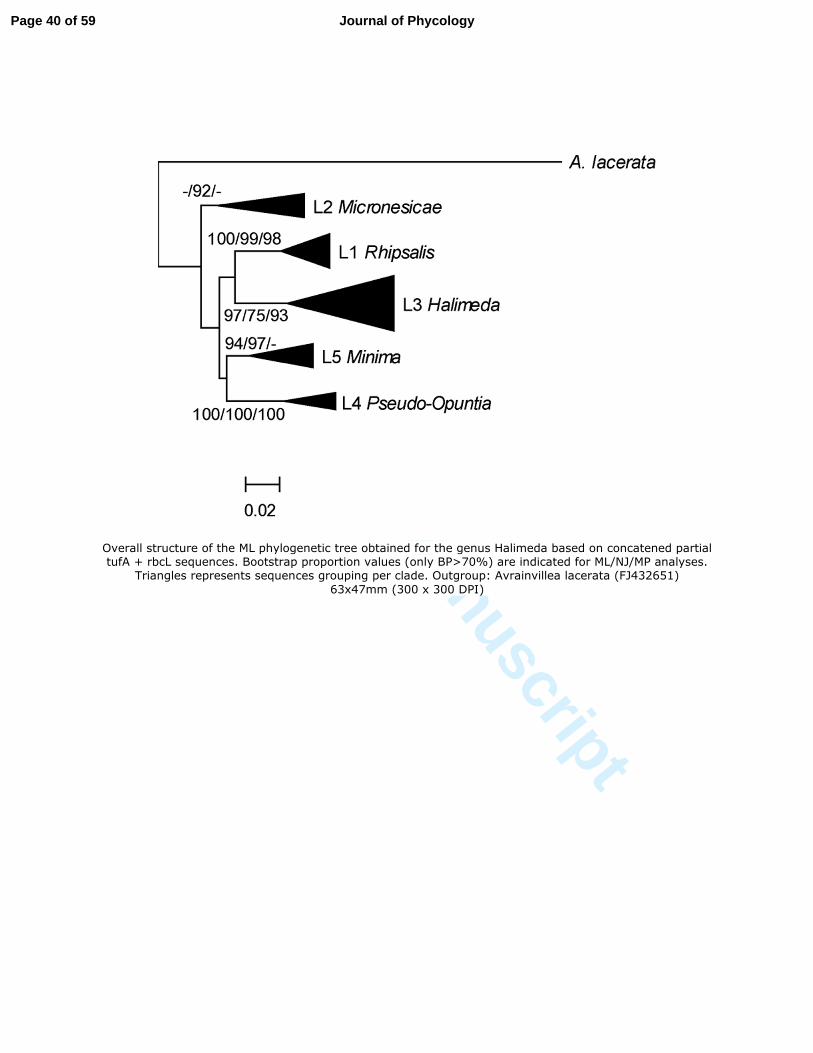
the concatenated (tufA + rbcL) sequence alignment is shown in Fig.2. While the backbone of 283
the tree did not receive acceptable support, five lineages representing the five sections of 284
Verbruggen and Kooistra (2004) were consistently recovered: Rhipsalis (lineage L1), 285
Micronesicae (L2), Halimeda (L3), Pseudo-opuntia (L4) and Opuntia (L5). The tufA gene, 286
for which the biggest dataset was available (see Table S2), was analyzed separately for each 287
section to get a more detailed picture of species boundaries within these lineages. 288
289
Lineage 1 (Rhipsalis) 290
Morphotypes A, B, C, O, P and T grouped within section Rhipsalis with GenBank sequences 291
available for H. heteromorpha, H. macroloba, H. kanaloana, H. incrassata, H. melanesica, H. 292
borneensis, H. cylindracea, H. monile and H. simulans (Fig. 3). The section was subdivided 293
into five moderately to strongly supported subclades (73% ≤ BP ≤100 %) denoted (i) to (v). 294
The sequences of H. borneensis grouped together (iv) but with two distinctive subgroups from 295
French Polynesia and New Caledonia. 296
Sequences of each morphotype grouped together and our identifications of these morphotypes 297
matched with the species names on the downloaded GenBank sequences. One exception is 298
specimen (IRD 5311, see arrow on Fig. 3), which we assigned to morphotype A (H. 299
cylindracea) but formed a lineage clearly separated from other species. While the sequence 300
grouped with GenBank sequences of H. monile, H. simulans and H. incrassata, the genetic 301
Page 13 of 59Journal of Phycology

Submitted M
anuscript
13
distinctness of this sequence suggests that it represents a separate species. A closer 302
morphological analysis indicated similarities with H. stuposa. 303
304
Lineage 2 (Micronesicae) 305
Sequences available for morphotypes D and E grouped within section Micronesicae with 306
GenBank sequences for H. micronesica, H. fragilis, H. cryptica and H. pygmaea (Fig.4). 307
Morphotype E from the Maldives strongly grouped (BP>99%, Fig. 4) with H. micronesica. 308
Specimens initially ascribed to morphotype D (H. fragilis) appeared to be polyphyletic and 309
grouped in three well-supported subgroups (BP>99%, Fig. 4). One specimen from the 310
Chesterfield grouped with sequences for H. fragilis from Guam and the Northern Mariana 311
Islands (noted H. fragilis 1), whereas the three specimens from the Maldives formed a distinct 312
clade with other sequences for H. fragilis from Philippines, Tanzania and Maldives (noted H. 313
fragilis 2). Two other specimens formed a distinct lineage (H. fragilis 3) sister to H. pygmaea 314
(Fig. 4). 315
316
Lineage 3 (Halimeda) 317
Morphotypes G, I, Q, H, J, F and S grouped within section Halimeda, which included 318
GenBank sequences for H. discoidea, H. gigas, H. magnidisca, H. xishaensis, H. cuneata, H. 319
macrophysa, H. taenicola, H. lacunalis, H. tuna, H. hummii and H. scabra (Fig. 5). This 320
lineage was subdivided into ten moderately to strongly supported clades (78% < BP < 100 %, 321
Fig 45). Overall, the morphotype assignments of our samples were consistent with the 322
identified GenBank sequences with which the samples clustered. However, small deviations 323
did occur. Morphotype G (H. discoidea) is polyphyletic, with widely divergent clades 324
containing Indo-Pacific and Atlantic/Caribbean specimens (see also Kooistra et al 2002, 325
Verbruggen et al. 2005b, Verbruggen et al 2009b). Specimens in morphotype J (H. taenicola) 326
formed a moderately to well supported clade (82% ≤ BP ≤ 90%) but was subdivided into two 327
Page 14 of 59 Journal of Phycology

Submitted M
anuscript
14
divergent subgroups, one with individuals from French Polynesia and the Maldives, and the 328
second with specimens from Chesterfield. 329
330
Lineage 4 (Pseudo-opuntia) 331
Only sequences available for morphotype K (H. gracilis) grouped within the Pseudo-opuntia 332
lineage with GenBank sequences available for H. gracilis and H. lacrimosa (Fig. 6). 333
Morphotype K sequences formed a moderately supported clade (72 % ≤ BP ≤ 81%, Fig. 6) 334
with H. gracilis. As can be seen in Fig. 6, H. gracilis is a complex consisting of at least four 335
genetically distinct species (see also Verbruggen et al., 2009b). Our specimens from New 336
Caledonia and Maldives all fell in clade "H. gracilis 1", which is the clade also containing 337
material from Sri Lanka, the type locality. Interestingly, the H. gracilis 1 lineage exhibits 338
quite divergent subclades, hinting towards the possibility that additional cryptic diversity is 339
present. 340
341
Lineage 5 (Opuntia) 342
Sequences available for morphotypes L, M, N and R grouped within the Opuntia lineage with 343
GenBank sequences for H. renschii, H. minima, H. distorta , H. opuntia, H. velasquezii, H. 344
copiosa and H. goreauii (Fig. 7). As anticipated on morphological grounds, morphotype M 345
grouped with sequences of H. opuntia (75% ≤ BP ≤ 97%, Fig. 7). H. distorta appeared 346
polyphyletic with two clades noted H. distorta 1 and H. distorta 2 weakly to strongly 347
supported (BP = 94% and BP = 76% respectively). Sequences obtained for morphotype N 348
grouped within the subgroup H. distorta 2. Morphotype R grouped strongly with H. 349
velasquezii sequences (BP = 93%, Fig 6). Sequences for H. minima and H. renschii formed a 350
large clade (BP >89 %, Fig 6) including a subgroup for H. renschii (BP =78%). 351
Morphotype L was found in the H. minima / H. renschii group within two subclades: 352
sequences from the Maldives grouped moderately with H. minima 1 from Australia and 353
Page 15 of 59Journal of Phycology

Submitted M
anuscript
15
Tanzania (BP = 72%) and sequences from New Caledonia grouped strongly with H. minima 4 354
from the Philippines and Fiji. Three unidentified samples from Northern Mariana Islands and 355
Fiji (H. no ID) were strongly grouped together (BP = 100%) and distinctive from other 356
species. 357
358
359
Discussion 360
Our primary aim was to characterize the species diversity of the genus Halimeda in New 361
Caledonia using a combination of molecular tools and morphological investigations. Besides 362
addressing this goal, our results have implications for our knowledge of the taxonomy and 363
global diversity of the genus and warrant reflecting upon the suitability of conventional 364
morphological identification criteria and methods for delimiting species in the genus. 365
366
Approaches towards species delimitation 367
Species delimitation is one of the principal goals of systematic biology. Classically, species 368
were delimited based on morphological differences between perceived entities. Since the 369
early 1990's, DNA sequencing has made it increasingly clear that species boundaries based on 370
perceived morphological differences have not always reflected biological reality. They have 371
shown that morphologically defined entities can encompass various genetic species (cryptic 372
diversity) and that DNA clusters at the species level can be morphologically diverse (e.g., 373
Zuccarello and West 2003; Leliaert et al., 2009). As a consequence of the morphological 374
plasticity of algae and the fact that morphological species delimitation can be deceptive, DNA 375
sequences are very useful complements to morphological data in defining species boundaries. 376
In Halimeda, various approaches have been taken towards species delimitation. All work 377
predating the DNA sequencing era has used external morphological and anatomical features 378
to define species boundaries. More recently, Verbruggen et al. (2005a, 2006) started by 379
Page 16 of 59 Journal of Phycology

Submitted M
anuscript
16
defining species boundaries based on sets of DNA barcodes and subsequently looked for 380
morphological characters coinciding with the DNA-based species boundaries. In this paper, 381
we tried an intermediate approach. We defined morphotypes based on analyses of our external 382
morphological and anatomical datasets and subsequently sequenced the samples and verified 383
whether the morphotypes corresponded to DNA clusters. While this was true in most cases, 384
we were unable to capture the full species diversity in our morphotypes – some of them 385
showed cryptic diversity at the DNA level. 386
The recognition of species-level clades in this study followed previous work on Halimeda, in 387
which species limits are defined manually by inspecting haplotype trees to identify well-388
supported clades that are preceded by a relatively long branch and have comparably low 389
sequence diversity within. We acknowledge that this method is to some degree subjective, in 390
that different workers may disagree on what constitutes a "well-supported clade preceded by a 391
relatively long branch and comparably low sequence diversity within". This is where the 392
taxonomist's judgment comes in, which is based on the joint interpretation of the haplotype 393
tree and the morphological information. In recent years, more objective methods have been 394
developed to pinpoint species boundaries in DNA sequence datasets (Pons et al. 2006, 395
Monaghan et al. 2009, Leliaert et al. 2009). These methods, and the general mixed Yule-396
coalescence approach in particular, correspond well to the method we have used here. It relies 397
on differences between slower above-species tree branching (speciation model) and faster 398
within-species tree branching (haplotype coalescence), which is what is more intuitively 399
assessed with the method we use. 400
Over the years, several DNA markers have been used for questions near the species level in 401
Halimeda. The early work by Kooistra used 18S nrDNA sequences (Hillis et al., 1998, 402
Kooistra et al., 1999), which turned out to be too conservative for fine-grained work and were 403
replaced with the ITS region (Kooistra et al.,2002, Verbruggen et al., 2005a, b, 2006). 404
Slightly later, the plastid marker UCP7 (Provan et al., 2004), comprising partial rps19 and 405
Page 17 of 59Journal of Phycology

Submitted M
anuscript
17
rps3 sequences and the spacer between them, was introduced for species delimitation in 406
sections Rhipsalis and Halimeda (Verbruggen et al. 2005a, b, 2006, 2009b). Recently, the 407
plastid tufA gene was proposed as the DNA barcode of choice for the Ulvophyceae (Saunders 408
and Kucera 2010). This marker had already been used for phylogenetic purposes in the genus 409
Halimeda (Verbruggen et al., 2005) and for species delimitation in two of its sections 410
(Pseudo-opuntia and Opuntia: Verbruggen et al., 2009b). In the present study, the use of the 411
tufA barcode is extended to all sections of the genus. It clearly follows from our results that 412
the gene has potential as a barcoding marker as it shows enough resolution to clearly 413
distinguish between species-level clades, shows the presence of cryptic diversity within 414
morphotypes (e.g. H. fragilis), and indicates the presence of new species and/or species not 415
currently present in the DNA databases (e.g. H. cf. stuposa, H. no ID). 416
417
Morphological identification criteria 418
Among the nine morphological characters analyzed in this study, many appeared to show 419
overlap between species. For example, it can be difficult to discriminate between H. 420
discoidea, and H. xishaensis using only thallus and segment form. On the other 421
handHowever, the same two characters allow perfect discrimination between other species 422
(e.g. H. cylindracea vs. H. macroloba). Bulbous holdfasts were found to occur in sections 423
Rhipsalis (L1) (except for H. melanesica) as well as Halimeda (L3), which was interpreted by 424
Verbruggen and Kooistra (2004) to be a response to a specific adaptation to growth in sandy 425
or muddy substrata. Verbruggen et al. (2005d) noted that some deviant segments should not 426
be included in morphological analyses because they can mislead species assignment. 427
Following their conclusion that basal, apical and non-calcified segments may not be 428
representative of the average morphology of a species, these segment types were excluded 429
from the present study. It is clear that external morphology may be used for a primary 430
classification of individuals into morphological groups but this enables species identification 431
Page 18 of 59 Journal of Phycology

Submitted M
anuscript
18
only in rare cases. Anatomical characters appear more reliable to identify species. Features 432
like the nodal fusion type and the size and form of utricles show more variation among 433
species (Verbruggen et al., 2005a), and therefore represent the main characters for 434
morphological identification. However, these features show overlap between species, too. A 435
combination of external and anatomical features obviously extends the range of distinctions 436
that can be made and is the preferred strategy for morphological discrimination between 437
species. Even though a well-contemplated combination of external morphological and 438
anatomical characters can provide a powerful source of information for the identification of a 439
majority of Halimeda species, it is certainly possible that they will not permit distinction 440
between the most similar of cryptic species pairs. 441
In the present study, which defined morphotypes prior to DNA analyses, morphological and 442
anatomical identifications were consistent with molecular results in a majority of cases, 443
except for morphotypes A, D and L, which were present in multiple genetic clusters. A hybrid 444
identification approach based on morpho-anatomical features for more distinctive species and 445
DNA tools for the cryptic species complexes may thus be reliable, although an inherent risk 446
of such an approach is that it may leave cryptic diversity within morphologically distinctive 447
taxa undetected. 448
449
Taxonomical implications 450
As mentioned above, there was generally a clear distinction between sequence clusters at the 451
species level and 14 out of 20 morphotypes grouped unambiguously with sequences 452
previously identified to the same species: morphotype B = H. borneensis, C = H. macroloba, 453
E = H. micronesica, F = H. lacunalis, H = H. macrophysa, I = H. magnidisca, J = H. 454
taenicola, M = H. opuntia, O = H. heteromorpha, P = H. kanaloana, Q = H. xishaensis, R = 455
H. velasquezii, S = H. gigas and T= H. melanesica. Clearly, these species can be incorporated 456
unambiguously in the list of New Caledonian Halimeda species. 457
Page 19 of 59Journal of Phycology

Submitted M
anuscript
19
Conversely, for 6 out of the 20 morphotypes, the situation was more complex due to cryptic 458
or pseudocryptic diversity. We distinguish between two different scenarios. The first is that 459
within New Caledonia, specimens from one morphotype belong to different cryptic entities. 460
This is the case for morphotypes A = H. cylindracea (2 entities), and D = H. fragilis (2 461
entities). The second scenario is that the morphospecies in question is known to contain 462
cryptic diversity, but the New Caledonian samples belong to only one of the constituent 463
cryptic entities. This is the case for morphotypes G = H. discoidea (2 entities), K = H. gracilis 464
(4 entities), L = H. minima (4 entities), and N = H. distorta (2 entities). We will now discuss 465
each of these six problematic cases and their taxonomic consequences in more detail. 466
467
Morphotype A, which was thought to correspond to H. cylindracea, appeared polyphyletic in 468
our phylogenetic analysis. As could be expected the majority of the samples of this 469
morphotype matched Genbank sequences belonging to H. cylindracea. However, a single 470
sequence attributed to morphotype A (IRD 5311) formed a separate lineage related to H. 471
incrassata, H. simulans and H. monile in subgroup (iii) (Fig. 3). A closer morphological 472
analysis demonstrated similarities of this specimen to H. stuposa. First and considering the 473
identification key to Halimeda species of section Rhipsalis (Verbruggen et al., 2005a), some 474
details can help us to differentiate the specimen from H. cylindracea: it has a basal zone 475
(basal segments up to the holdfast and underneath the first ramifications) of 1.6 cm long, 476
segments thicker than H. cylindracea (in average, 1.5 to 2 mm vs less than 1.5 mm for H. 477
cylindracea) and its segments are not cylindrical throughout the thallus but most of them were 478
trilobed. Secondly, it possessed segments always less than 12.5 mm wide (most in the 2-6 mm 479
range), and its peripheral utricles averaged 35.8 µm in width and 58.8 µm in length, while the 480
width of its secondary utricles equaled 28 µm in width with supranodal siphons shorter than 481
300 µm (208 µm). In fact, following Verbruggen et al. (2005a), our specimen presented all 482
the criteria to be identified as H. stuposa, but since no DNA sequence data are available for 483
Page 20 of 59 Journal of Phycology

Submitted M
anuscript
20
confirmed samples of H. stuposa for verification, we converged on "H. cf. stuposa" as the 484
identification for this clade. 485
486
Morphotype D (Lineage 2), identified as H. fragilis, showed clear evidence for cryptic 487
diversity. The New Caledonian samples were only present in two of these (H. fragilis 1 and 488
H. fragilis 3) but our samples from the Maldives, Tanzania and the Philippines formed a third 489
cluster (H. fragilis 2). Closer examination did not reveal obvious morphological differences 490
between the clusters. 491
The morphospecies H. discoidea (lineage 3) appeared polyphyletic with two distinct and 492
unrelated clusters. The first (H. discoidea 1) contained all sequences from Indo-Pacific 493
material, including all our New Caledonian samples belonging to morphotype G. The second 494
cluster comprised all sequences from Atlantic specimens (H. discoidea 2). The genetic 495
distinctness of Atlantic and Indo-Pacific specimens belonging to the morphospecies has been 496
discussed in detail in the literature (Kooistra et al., 1999, Verbruggen et al. 2005b, 497
Verbruggen et al. 2009b). 498
The polyphyly of H. gracilis initially noted by Kooistra et al. (2002) has later been confirmed 499
with better taxon sampling (Verbruggen et al., 2009b, this study). It is now clear that this 500
morphospecies is a complex of four species (Fig. 6), two from the Indo-Pacific region (H. 501
gracilis ip 1 & ip 2) and two from the Atlantic Ocean (H. gracilis flo & car 2). Our specimens 502
from New Caledonia and the Maldives (morphotype K) all fell within H. gracilis ip 1, which 503
includes the specimen from the type locality and can safely be considered as the true H. 504
gracilis. The other groups, which are not the focus of this paper as they do not occur in New 505
Caledonia, should be subjected to more intensive morphological investigation in an attempt to 506
identify diagnostic characters. 507
Our molecular data resolved Halimeda minima into four clusters, one of which (H. minima 4) 508
is present in New Caledonia. The polyphyly of this species was initially noted by Kooistra et 509
Page 21 of 59Journal of Phycology

Submitted M
anuscript
21
al. (2002), who distinguished three distinct groups, while Verbruggen et al. (2009b) also 510
recognized the fourth cluster noted here. The new data presented here shows that while two of 511
the four clusters are clearly distinct (H. minima 2 and H. minima 4), sequences of H. minima 512
1, H. minima 3 and H. renschii are very closely related and do not always form clear-cut 513
clusters, indicating that the taxonomy of the species H. minima and H. renschii needs to be 514
reconsidered. The principal morphological criterion used to separate H. minima from H. 515
renschii is that the holdfast region of the latter consists of a region at least 1 cm high in which 516
the basal segments are covered in rhizoids and sand particles of several segments, whereas the 517
former has a simple holdfast at the basal segment (Hillis-Colinvaux, 1980). However, it is 518
known that holdfasts can show within-species variation depending on habitat (Verbruggen et 519
al. 2006). As the H. renschii – minima 1 – minima 3 complex does not appear to be present in 520
New Caledonia, we will not treat this any further, but this group needs a more critical 521
evaluation. 522
Our analyses resolved H. distorta in two groups (cf. Verbruggen et al., 2009b). While the 523
cluster noted H. distorta 1 appears to be restricted to the Pacific, H. distorta 2 grouped 524
specimens from the wider Indo-Pacific area, including our specimens from New Caledonia 525
(morphotype N). Kooistra and Verbruggen (2005) merged H. hederacea with H. distorta 526
based on the lack of separation of these taxa in ITS nrDNA sequences, and it is not clear how 527
the two clusters we find within H. distorta relate to these older names. Additional work is 528
clearly needed to sort out the taxonomy of this complex. 529
We wish to make a final taxonomic note about H. taenicola. Even though this species is 530
recovered as monophyletic in our study, it is composed of two distinct clusters one of which 531
has samples only from the Chesterfield reef area. We have not been able to find marked 532
anatomical or morphological differences between these two subgroups, although our 533
specimens from the Maldives did look bushier than specimens from Chesterfield. However 534
this character may not be valuable to discriminate the two subgroups because the analysis is 535
Page 22 of 59 Journal of Phycology

Submitted M
anuscript
22
based on few specimens and the general shape of the thallus may be strongly influenced by 536
the environment. 537
538
Diversity of Halimeda in New Caledonia 539
This study identified 22 species of Halimeda in New Caledonia, representing all five sections 540
of the genus. 541
Eighteen species were confirmed using a combination of DNA analyses and morphological 542
investigation, i.e. H. borneensis, H. cylindracea, H. discoidea, H. distorta, H. fragilis 1, H. 543
fragilis 3, H. gracilis, H. heteromorpha, H. kanaloana, H. lacunalis, H. macroloba, H. 544
macrophysa, H. magnidisca, H. minima, H.melanesica, H. opuntia, H. taenicola and H. 545
velasquezii. Some morpho-species were shown to harbor cryptic diversity. In one case, two 546
cryptic species were present in New Caledonia (H. fragilis 1 and H. fragilis 3). In four other 547
cases, the species in question was shown to contain several cryptic species but only one of 548
these cryptic species was present in New Caledonia. This was the case for H. discoidea (only 549
the Indo-Pacific entity in New Caledonia), H. gracilis (only entity #1 in New Caledonia), H. 550
minima (only entity #4 in New Caledonia) and H. distorta (only entity #2 in New Caledonia). 551
Four Three other species were identified based on morphological criteria only: H. 552
micronesica, H. xishaensis and H. gigas. Regrettably, we were unable to generate DNA data 553
for these species. However, because these species represent clear-cut morphotypes that have 554
been shown to form distinct DNA clusters (Verbruggen et al., 2005b, 2005c; this study) with 555
no reports of cryptic diversity, a high degree of confidence can be attached to these records. 556
We also identified a DNA cluster (H. cf stuposa) that forms a distinct lineage in molecular 557
analyses and thus represents a proper species. While the specimen in question bears 558
resemblance to H. stuposa, this identification is not certain as the morphology does not match 559
exactly and no sequences of typical H. stuposa samples are available. 560
Page 23 of 59Journal of Phycology

Submitted M
anuscript
23
The report of the species H. kanaloana for New Caledonia represents a major extension of the 561
known distribution range of this species, as this species was previously known only from 562
Hawaii and possibly Japan (Ryu Kyu Islands). The New Caledonian specimens had 563
previously been identified as H. incrassata (Garrigue 1985), which is morphologically close 564
to H. kanaloana in various aspects, but can be distinguished on the basis of six features 565
including the dimensions of subperipheral and peripheral utricles, and the distance between 566
the base of the nodal fusions and the first siphon ramification above the node (Verbruggen et 567
al., 2006). They also differ in their distribution: H. incrassata is restricted to the Atlantic 568
Ocean while H. kanaloana is a Pacific Ocean species (Verbruggen et al., 2006, this study). 569
Similarly, the recognition of H. xishaensis in New Caledonia marks a major extension of the 570
known range of this species, because, to our knowledge, it was previously only recorded from 571
the Xi Sha or Paracel Islands in the South China Sea. Finally, we obtained here the first 572
sequence of H. melanesica from its type locality (Lifou, New Caledonia), which will facilitate 573
accurate molecular identification of specimens of this species from other parts of the world. 574
575
Acknowledgments 576
We acknowledge the IRD divers team and boat pilots for their contribution to this work 577
through kind and careful assistance. Thanks also to all collectors who have contributed to the 578
study: G. Lasne (BIOCENOSE), E. Fontan, O. Cornubert and D. Ponton. Thanks to C. 579
Fauvelot for providing samples from French Polynesia and Antoine de N’Yeurt for help with 580
identification of specimens collected in Moorea. Thanks to Coppejans E., Dargent O., 581
Tyberghein L., Leliaert F., Diaz R., Galanza C., Huisman J., De Clerck O., Schils T., Payo 582
D.A. and Pauly K. for collecting Ghent samples. We sincerely thank C. Majorel (Plateforme 583
du vivant) and S. D'hondt (Ghent University) for assisting with the molecular work. HV is a 584
postdoctoral fellow of the Research Foundation – Flanders. LD is a PhD fellow of the 585
Province Sud- New Caledonia. 586
Page 24 of 59 Journal of Phycology

Submitted M
anuscript
24
587
References 588
Agardh, J. G. 1887. Till algernes systematik. Nya bidrag. (Femte afdelningen.). Acta 589
Universitatis Lundensis. 23(2): 1-174, 5 plates. 590
591
Andréfouët, S., Torres-Pulliza, D., Dosdane, M. (collab.), Kranenburg, C. (collab.), Murch, B. 592
(collab.), Muller-Karger, F. E. (collab.), Robinson, J. A. (collab.) 2004. Atlas des récifs 593
coralliens de Nouvelle Calédonie. Nouméa (NCL) ; Nouméa : IRD ; IFRECOR, 26 p. + 22 p. 594
595
Andréfouët, S., Cabioch, G., Flamand, B., Pelletier, B. 2009. A reappraisal of the diversity of 596
geomorphological and genetic processes of New Caledonian coral reefs: a synthesis from 597
optical remote sensing, coring and acoustic multibeam observations. Coral Reefs. 28:691-707. 598
599
Askenasy, E. 1888. Algen mit Unterstützung der Herren E. Bornet, E. Grunow, P. Hariot, M. 600
Moebius, O. Nordstedt bearbeitet. In: Forschungsreise S.M.S. "Gazelle". Theil 4: Bot. 601
(Engler, A. Eds), Berlin, pp. 1-58. 602
603
Barton, E. S. 1901. The genus Halimeda. Siboga-Expeditie Monographe LX of: Uitkomsten 604
op Zoologisch, Botanisch, Oceanographisch en Geologisch Gebied verzameld in 605
Nederlandsch Oost-Indie 1899-1900 60: 1-32, 2 figs, 4 pls. 606
Chevillon, C. 1996. Skeletal composition of modern lagoon sediments in New Caledonia: 607
coral, a minor constituent. Coral Reefs. 15:199-207. 608
Cocquyt, E., Verbruggen, H., Leliaert, F., Zechman, F. W., Sabbe, K. & De Clerck, O. 2009. 609
Gain and loss of elongation factor genes in green algae. BMC Evol. Biol. 9:39. 610
Page 25 of 59Journal of Phycology

Submitted M
anuscript
25
Curtis, N., Dawes, C. & Pierce, S. 2008. Phylogenetic analysis of the large subunit Rubisco 611
gene supports the exclusion of Avrainvillea and Cladocephalus from the Udoteaceae 612
(Bryopsidales, Chlorophyta). J. Phycol. 44:761-67. 613
Dong, M. L., Tseng, C. K. 1980. Studies on the Chlorophyceae from Xi Sha Islands, China. 614
Studia Marina Sinica 17:1-10. 615
Famà, P., Wysor, B., Kooistra, W. H. C. F. & Zuccarello, G. C. 2002. Molecular phylogeny of 616
the genus Caulerpa (Caulerpales, Chlorophyta) inferred from chloroplast tufA gene. J. 617
Phycol. 38:1040-50. 618
Felsenstein, J. 1985. Confidence limits on phylogenies: An approach using the Bootstrap. 619
Evolution 39:783-91. 620
Garrigue, C., & Tsuda, R. O. Y. T. 1988. Catalog of Marine Benthic Algae from New 621
Caledonia. Micronesica 21:53-70. 622
Garrigue, C. 1995. Macrophyte associations on the soft bottoms of the south-west lagoon of 623
New Caledonia: Description, structure and biomass. Bot. Mar. 38:481–92. 624
Guindon, S. & Gascuel, O. 2003. A simple, fast, and accurate algorithm to estimate large 625
phylogenies by maximum likelihood. Syst. Biol. 52:696-704. 626
Guiry, M. D. & Guiry, G. M. 2011. AlgaeBase. World-wide electronic publication, National 627
University of Ireland, Galway. http://www.algaebase.org; searched on 23 August 2011. 628
Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis 629
program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 41:95-98. 630
Hanyuda, T., Arai, S. & Ueda, K. 2000. Variability in the rbcL introns of Caulerpalean algae 631
(Chlorophyta, Ulvophyceae). J. Plant Res. 113:403-413. 632
Page 26 of 59 Journal of Phycology

Submitted M
anuscript
26
Use the "Insert Citation" button to add citations to this document. 633
Zuccarello, G.C. & West, J.A., 2003. Multiple cryptic species: molecular diversity and 634
reproductive isolation in the Bostrychia radicans/ B. moritziana complex (Rhodomelaceae, 635
Rhodophyta) with focus on North American isolates. J. Phycol., 39(5):948-959. 636
637
638
639
Figure legends: 640
641
642
Figure 1. Geographic position of New Caledonia in the South West Pacific (A) and map of the 643
New Caledonian islands and reefs (B) 644
645
Figure 2. Overall structure of the ML phylogenetic tree obtained for the genus Halimeda 646
based on concatened partial tufA + rbcL sequences. Bootstrap proportion values (only 647
BP>70%) are indicated for ML/NJ/MP analyses. Triangles represents sequences grouping per 648
clade. Outgroup: Avrainvillea lacerata (FJ432651) 649
650
Figure 3. NJ analysis for section Rhipsalis (L1) based on tufA sequence alignment. Bootstrap 651
values (BP>70%) are indicated for NJ/MP/ML analyses. Outgroup: Halimeda micronesica. 652
Localities are noted as follows, FP: French Polynesia, Phi: Philippines, NC: New Caledonia, 653
Tanz: Tanzania, H: Hawaii, Ye: Yemen, Pan: Panama, Jam: Jamaica, Bah: Bahamas and Car: 654
Caribbean. Asterisks indicate type localities and the arrow points to an unexpected grouping 655
and boxes highlight New Caledonian samples 656
657
Page 27 of 59Journal of Phycology

Submitted M
anuscript
27
Figure 4. NJ analysis of section Micronesicae (L2) based on tufA sequence alignment. 658
Bootstrap values (BP>70%) are indicated for NJ/MP/ML analyses. Outgroup: Halimeda 659
distorta. Localities are noted as follows, Mal: Maldives; Phi: Philippines; Tanz: Tanzania; 660
NC: Grande terre, New Caledonia; Chest-NC: Chesterfield, New Caledonia; Jam: Jamaica; 661
NM: North Marianna; Aust: Australia; Wa: Wallis and Cha: Chagos. Asterisks indicate type 662
localities, arrows point at unexpected groupings and boxes highlight New Caledonian 663
samples. 664
665
Figure 5. MP analysis of section Halimeda (L3) based on tufA sequence alignment. Bootstrap 666
values (BP>70%) are indicated for MP/ NJ/ML analyses. Outgroup: Halimeda incrassata. 667
Localities are noted as followed, Nic : Nicaragua; FP: French Polynesia; Mal: Maldives; 668
Chest-NC: Chesterfield; New Caledonia; NC: New Caledonia; Ye: Yemen; SrL: Sri-Lanka; 669
Aust W: Western Australia; Sth Af: South Africa; Tanz: Tanzania; Bre: Brasil; It: Italia; Can: 670
Canaries; Jam: Jamaica; Pan: Panama; Mex: Mexico; Bah: Bahamas; Thai: Thailand; Phi : 671
Philippines; CR: Costa Rica. Asterisks indicate type localities and boxes highlight New 672
Caledonian samples. 673
674
Figure 6. NJ analysis of section Pseudo-opuntia (L4) based on tufA sequence alignments. 675
Bootstrap values (BP>70%) are indicated for NJ/MP/ML analyses. Outgroup: Halimeda 676
macrophysa. Localities are noted as follows, Mal: Maldives; SrL: Sri Lanka; Ha: Hawaii; 677
Chest-NC: Chesterfield, New Caledonia; PF: French Polynesia; Aust W: West Australia; Phi: 678
Philippines; Sth Af: South Africa;Tanz: Tanzania; Flo: Florida; Jam: Jamaica; Bah: Bahamas. 679
Asterisks indicate type localities and the box highlight New Caledonian sample. 680
681
Figure 7. ML analysis of section Opuntia (L5) based on tufA sequence alignments. Bootstrap 682
values (BP>70%) are indicated for ML/ NJ/MP analyses. Outgroup: Halimeda micronesica. 683
Page 28 of 59 Journal of Phycology

Submitted M
anuscript
28
Localities are noted as followed, FP: French Polynesia, Aust: Australia, Phi: Philippines, 684
Tanz: Tanzania, C. Isl: Cook Islands, Mal: Maldives, Soc: Socotra, NC: New Caledonia, Wa: 685
Wallis, Micro: Micronesia, Pal: Palau, NMI: North Marianna Island, Jam: Jamaica, Pan: 686
Panama. Asterisks indicate type localities and boxes highlight New Caledonian samples. 687
. 688
689
Page 29 of 59Journal of Phycology

Submitted M
anuscript
29
690
Page 30 of 59 Journal of Phycology
Submitted Manuscript
30
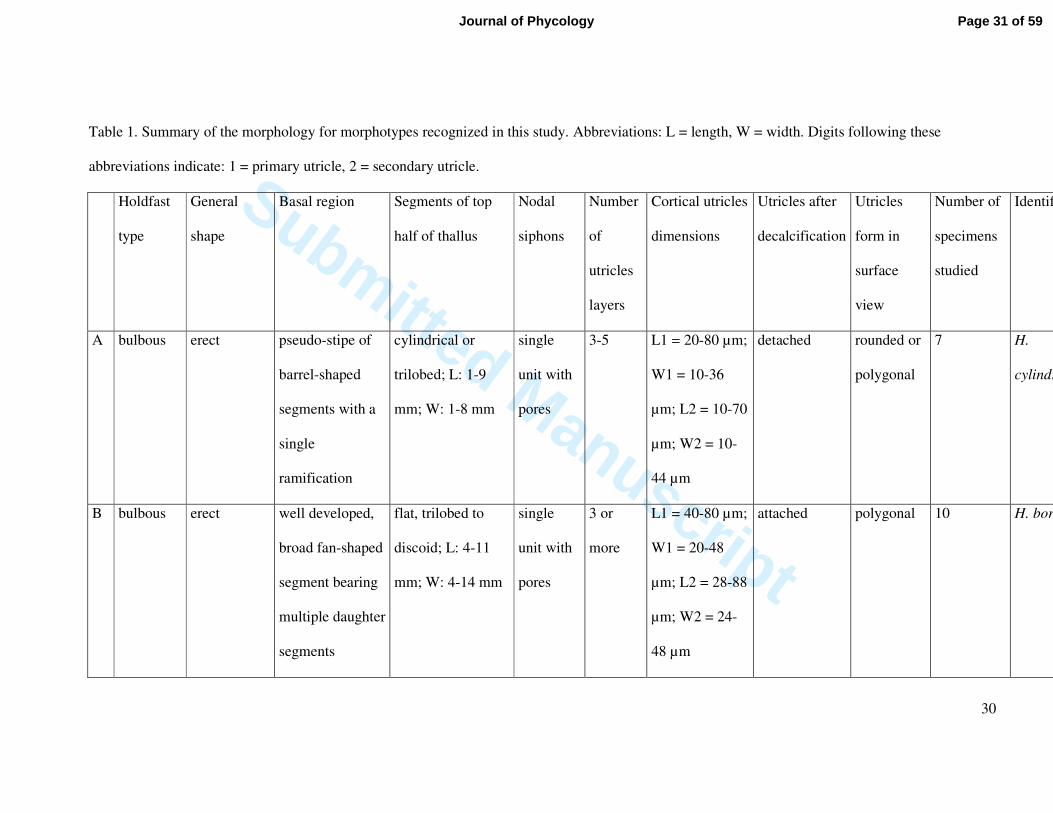
Table 1. Summary of the morphology for morphotypes recognized in this study. Abbreviations: L = length, W = width. Digits following these
abbreviations indicate: 1 = primary utricle, 2 = secondary utricle.
Holdfast
type
General
shape
Basal region Segments of top
half of thallus
Nodal
siphons
Number
of
utricles
layers
Cortical utricles
dimensions
Utricles after
decalcification
Utricles
form in
surface
view
Number of
specimens
studied
Identification
A bulbous erect pseudo-stipe of
barrel-shaped
segments with a
single
ramification
cylindrical or
trilobed; L: 1-9
mm; W: 1-8 mm
single
unit with
pores
3-5 L1 = 20-80 µm;
W1 = 10-36
µm; L2 = 10-70
µm; W2 = 10-
44 µm
detached rounded or
polygonal
7 H.
cylindracea
B bulbous erect well developed,
broad fan-shaped
segment bearing
multiple daughter
segments
flat, trilobed to
discoid; L: 4-11
mm; W: 4-14 mm
single
unit with
pores
3 or
more
L1 = 40-80 µm;
W1 = 20-48
µm; L2 = 28-88
µm; W2 = 24-
48 µm
attached polygonal 10 H. borneensis
Page 31 of 59Journal of Phycology
Submitted Manuscript
31
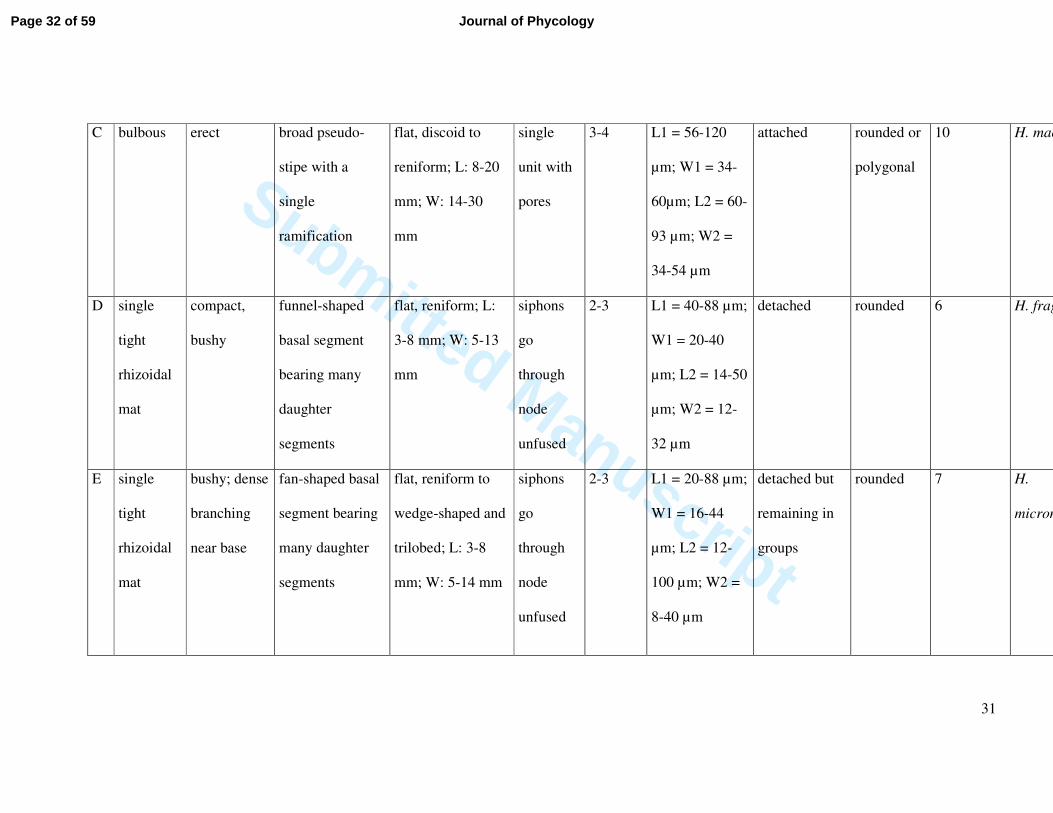
C bulbous erect broad pseudo-
stipe with a
single
ramification
flat, discoid to
reniform; L: 8-20
mm; W: 14-30
mm
single
unit with
pores
3-4 L1 = 56-120
µm; W1 = 34-
60µm; L2 = 60-
93 µm; W2 =
34-54 µm
attached rounded or
polygonal
10 H. macroloba
D single
tight
rhizoidal
mat
compact,
bushy
funnel-shaped
basal segment
bearing many
daughter
segments
flat, reniform; L:
3-8 mm; W: 5-13
mm
siphons
go
through
node
unfused
2-3 L1 = 40-88 µm;
W1 = 20-40
µm; L2 = 14-50
µm; W2 = 12-
32 µm
detached rounded 6 H. fragilis
E single
tight
rhizoidal
mat
bushy; dense
branching
near base
fan-shaped basal
segment bearing
many daughter
segments
flat, reniform to
wedge-shaped and
trilobed; L: 3-8
mm; W: 5-14 mm
siphons
go
through
node
unfused
2-3 L1 = 20-88 µm;
W1 = 16-44
µm; L2 = 12-
100 µm; W2 =
8-40 µm
detached but
remaining in
groups
rounded 7 H.
micronesica
Page 32 of 59 Journal of Phycology
Submitted Manuscript
32
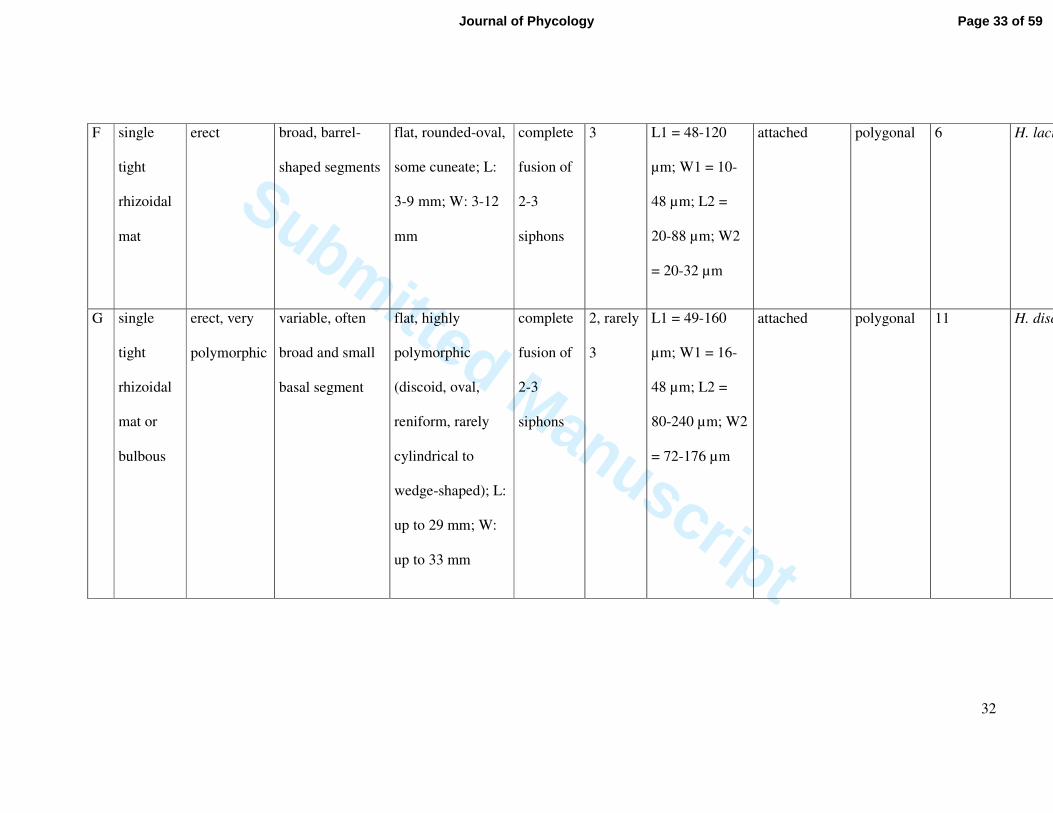
F single
tight
rhizoidal
mat
erect broad, barrel-
shaped segments
flat, rounded-oval,
some cuneate; L:
3-9 mm; W: 3-12
mm
complete
fusion of
2-3
siphons
3 L1 = 48-120
µm; W1 = 10-
48 µm; L2 =
20-88 µm; W2
= 20-32 µm
attached polygonal 6 H. lacunalis
G single
tight
rhizoidal
mat or
bulbous
erect, very
polymorphic
variable, often
broad and small
basal segment
flat, highly
polymorphic
(discoid, oval,
reniform, rarely
cylindrical to
wedge-shaped); L:
up to 29 mm; W:
up to 33 mm
complete
fusion of
2-3
siphons
2, rarely
3
L1 = 49-160
µm; W1 = 16-
48 µm; L2 =
80-240 µm; W2
= 72-176 µm
attached polygonal 11 H. discoidea
Page 33 of 59Journal of Phycology
Submitted Manuscript
33
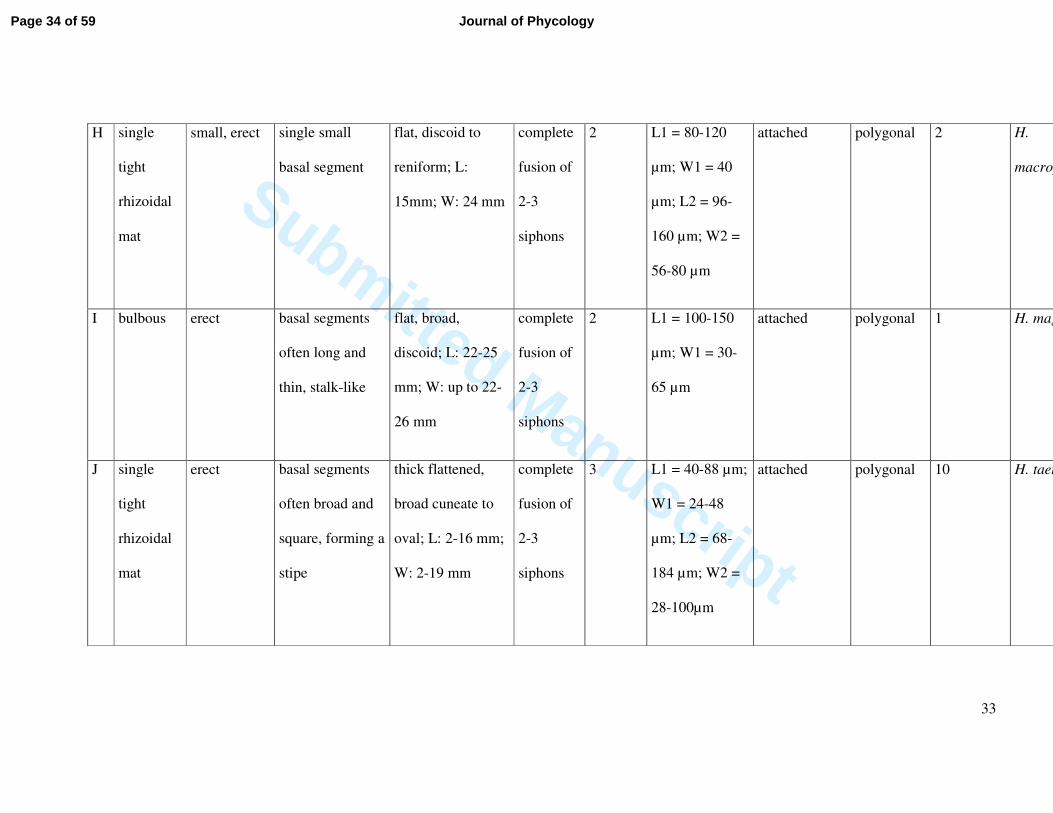
H single
tight
rhizoidal
mat
small, erect single small
basal segment
flat, discoid to
reniform; L:
15mm; W: 24 mm
complete
fusion of
2-3
siphons
2 L1 = 80-120
µm; W1 = 40
µm; L2 = 96-
160 µm; W2 =
56-80 µm
attached polygonal 2 H.
macrophysa
I bulbous erect basal segments
often long and
thin, stalk-like
flat, broad,
discoid; L: 22-25
mm; W: up to 22-
26 mm
complete
fusion of
2-3
siphons
2 L1 = 100-150
µm; W1 = 30-
65 µm
attached polygonal 1 H. magnidisca
J single
tight
rhizoidal
mat
erect basal segments
often broad and
square, forming a
stipe
thick flattened,
broad cuneate to
oval; L: 2-16 mm;
W: 2-19 mm
complete
fusion of
2-3
siphons
3 L1 = 40-88 µm;
W1 = 24-48
µm; L2 = 68-
184 µm; W2 =
28-100µm
attached polygonal 10 H. taenicola
Page 34 of 59 Journal of Phycology
Submitted Manuscript
34
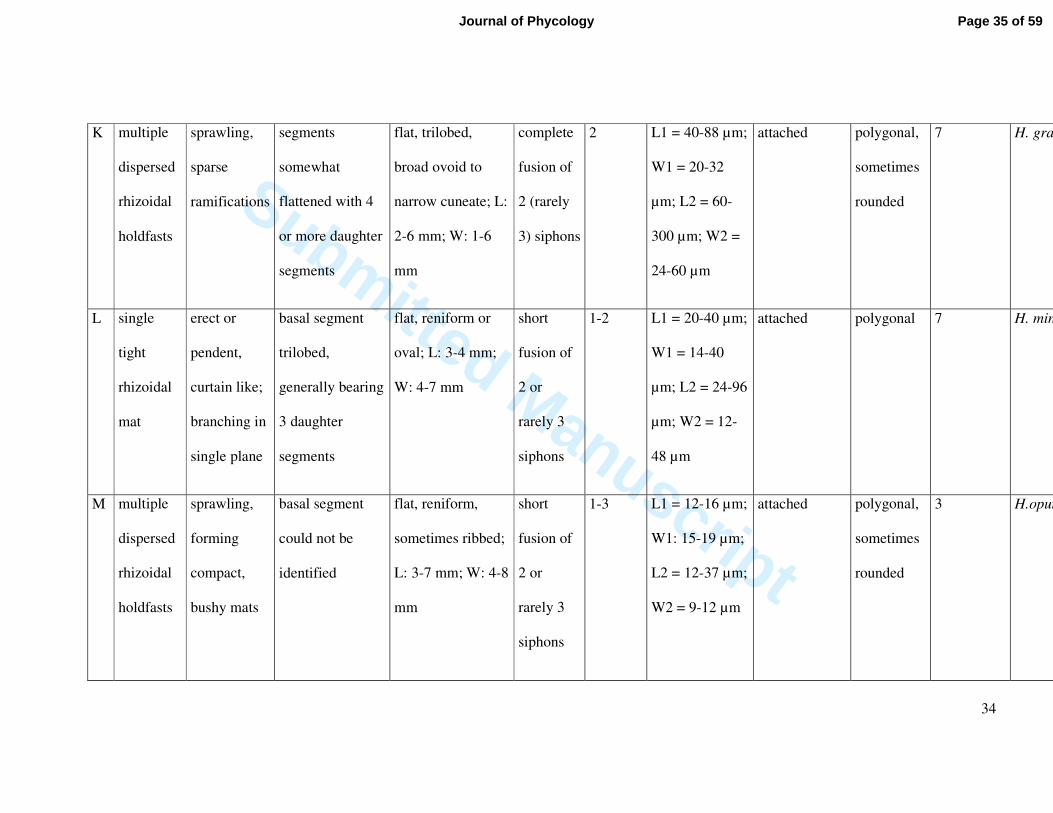
K multiple
dispersed
rhizoidal
holdfasts
sprawling,
sparse
ramifications
segments
somewhat
flattened with 4
or more daughter
segments
flat, trilobed,
broad ovoid to
narrow cuneate; L:
2-6 mm; W: 1-6
mm
complete
fusion of
2 (rarely
3) siphons
2 L1 = 40-88 µm;
W1 = 20-32
µm; L2 = 60-
300 µm; W2 =
24-60 µm
attached polygonal,
sometimes
rounded
7 H. gracilis
L single
tight
rhizoidal
mat
erect or
pendent,
curtain like;
branching in
single plane
basal segment
trilobed,
generally bearing
3 daughter
segments
flat, reniform or
oval; L: 3-4 mm;
W: 4-7 mm
short
fusion of
2 or
rarely 3
siphons
1-2 L1 = 20-40 µm;
W1 = 14-40
µm; L2 = 24-96
µm; W2 = 12-
48 µm
attached polygonal 7 H. minima
M multiple
dispersed
rhizoidal
holdfasts
sprawling,
forming
compact,
bushy mats
basal segment
could not be
identified
flat, reniform,
sometimes ribbed;
L: 3-7 mm; W: 4-8
mm
short
fusion of
2 or
rarely 3
siphons
1-3 L1 = 12-16 µm;
W1: 15-19 µm;
L2 = 12-37 µm;
W2 = 9-12 µm
attached polygonal,
sometimes
rounded
3 H.opuntia
Page 35 of 59Journal of Phycology
Submitted Manuscript
35
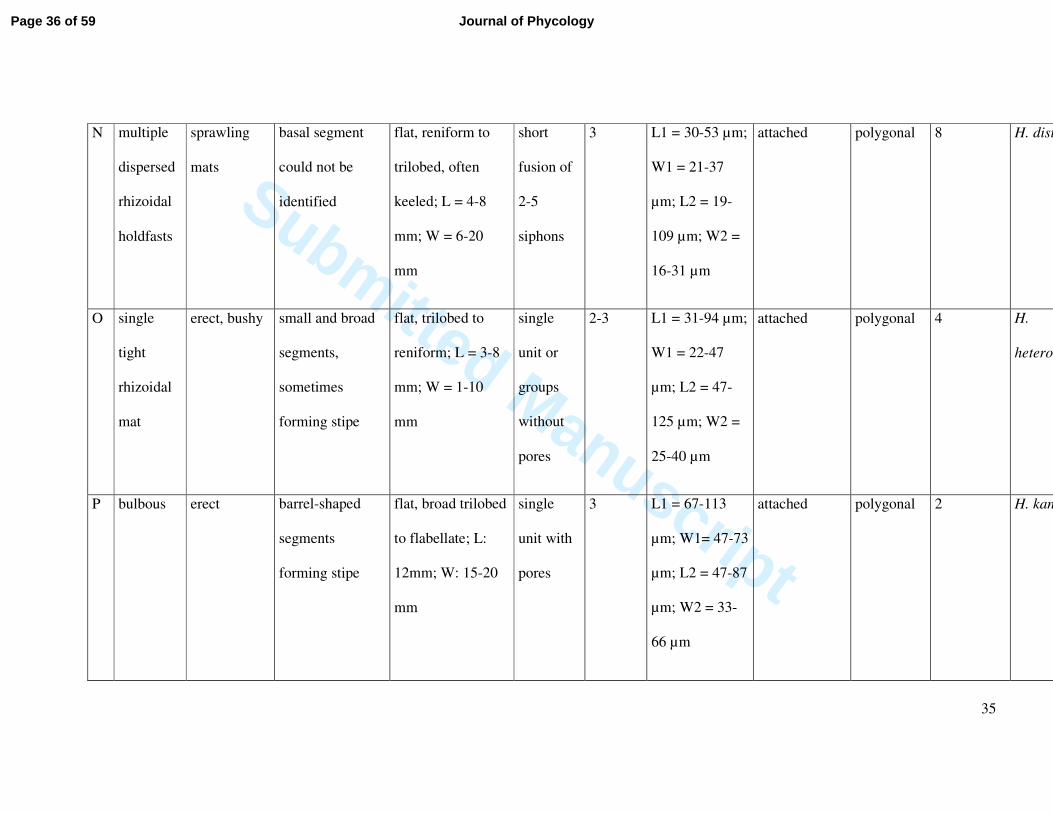
N multiple
dispersed
rhizoidal
holdfasts
sprawling
mats
basal segment
could not be
identified
flat, reniform to
trilobed, often
keeled; L = 4-8
mm; W = 6-20
mm
short
fusion of
2-5
siphons
3 L1 = 30-53 µm;
W1 = 21-37
µm; L2 = 19-
109 µm; W2 =
16-31 µm
attached polygonal 8 H. dist
O single
tight
rhizoidal
mat
erect, bushy small and broad
segments,
sometimes
forming stipe
flat, trilobed to
reniform; L = 3-8
mm; W = 1-10
mm
single
unit or
groups
without
pores
2-3 L1 = 31-94 µm;
W1 = 22-47
µm; L2 = 47-
125 µm; W2 =
25-40 µm
attached polygonal 4 H.
heteromorpha
P bulbous erect barrel-shaped
segments
forming stipe
flat, broad trilobed
to flabellate; L:
12mm; W: 15-20
mm
single
unit with
pores
3 L1 = 67-113
µm; W1= 47-73
µm; L2 = 47-87
µm; W2 = 33-
66 µm
attached polygonal 2 H. kanaloana
Page 36 of 59 Journal of Phycology
Submitted Manuscript
36
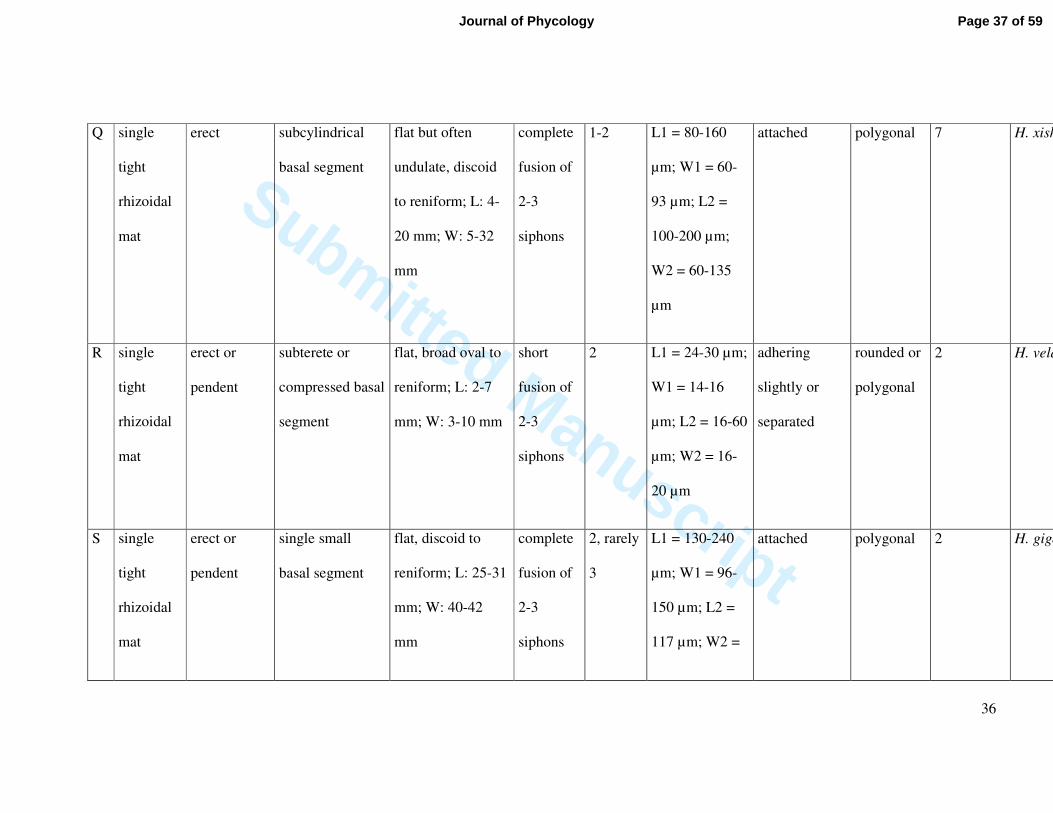
Q single
tight
rhizoidal
mat
erect subcylindrical
basal segment
flat but often
undulate, discoid
to reniform; L: 4-
20 mm; W: 5-32
mm
complete
fusion of
2-3
siphons
1-2 L1 = 80-160
µm; W1 = 60-
93 µm; L2 =
100-200 µm;
W2 = 60-135
µm
attached polygonal 7 H. xishaensis
R single
tight
rhizoidal
mat
erect or
pendent
subterete or
compressed basal
segment
flat, broad oval to
reniform; L: 2-7
mm; W: 3-10 mm
short
fusion of
2-3
siphons
2 L1 = 24-30 µm;
W1 = 14-16
µm; L2 = 16-60
µm; W2 = 16-
20 µm
adhering
slightly or
separated
rounded or
polygonal
2 H. velasquezii
S single
tight
rhizoidal
mat
erect or
pendent
single small
basal segment
flat, discoid to
reniform; L: 25-31
mm; W: 40-42
mm
complete
fusion of
2-3
siphons
2, rarely
3
L1 = 130-240
µm; W1 = 96-
150 µm; L2 =
117 µm; W2 =
attached polygonal 2 H. gigas
Page 37 of 59Journal of Phycology
Submitted Manuscript
37
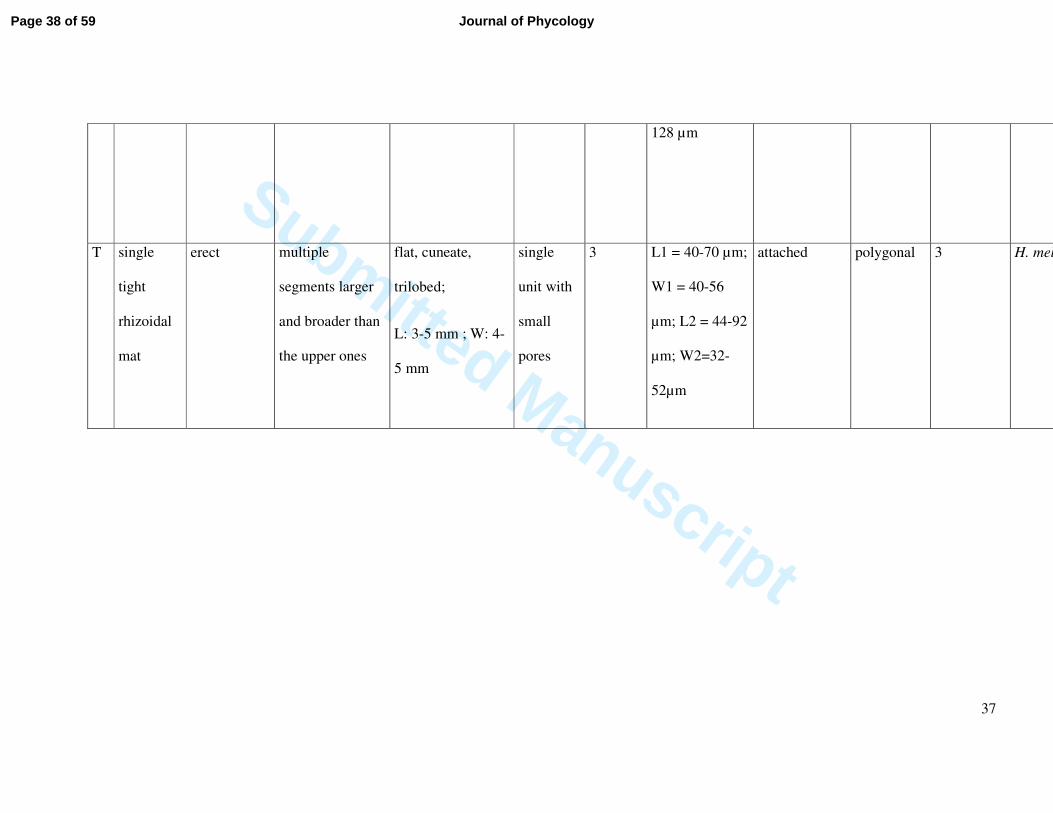
128 µm
T single
tight
rhizoidal
mat
erect multiple
segments larger
and broader than
the upper ones
flat, cuneate,
trilobed;
L: 3-5 mm ; W: 4-
5 mm
single
unit with
small
pores
3 L1 = 40-70 µm;
W1 = 40-56
µm; L2 = 44-92
µm; W2=32-
52µm
attached polygonal 3 H. melanesica
Page 38 of 59 Journal of Phycology
Submitted M
anuscript
Geographic position of New Caledonia in the South West Pacific (A) and map of the New Caledonian islands and reefs (B)
297x420mm (300 x 300 DPI)
Page 39 of 59Journal of Phycology
Submitted M
anuscript
Overall structure of the ML phylogenetic tree obtained for the genus Halimeda based on concatened partial tufA + rbcL sequences. Bootstrap proportion values (only BP>70%) are indicated for ML/NJ/MP analyses.
Triangles represents sequences grouping per clade. Outgroup: Avrainvillea lacerata (FJ432651)
63x47mm (300 x 300 DPI)
Page 40 of 59 Journal of Phycology

Submitted M
anuscript
NJ analysis for section Rhipsalis (L1) based on tufA sequence alignment. Bootstrap values (BP>70%) are indicated for NJ/MP/ML analyses. Outgroup: Halimeda micronesica. Localities are noted as follows, FP:
French Polynesia, Phi: Philippines, NC: New Caledonia, Tanz: Tanzania, H: Hawaii, Ye: Yemen, Pan: Panama, Jam: Jamaica, Bah: Bahamas and Car: Caribbean. Asterisks indicate type localities and the arrow points to
an unexpected grouping and boxes highlight New Caledonian samples 215x176mm (300 x 300 DPI)
Page 41 of 59Journal of Phycology

Submitted M
anuscript
NJ analysis of section Micronesicae (L2) based on tufA sequence alignment. Bootstrap values (BP>70%) are indicated for NJ/MP/ML analyses. Outgroup: Halimeda distorta. Localities are noted as follows, Mal:
Maldives; Phi: Philippines; Tanz: Tanzania; NC: Grande terre, New Caledonia; Chest-NC: Chesterfield, New Caledonia; Jam: Jamaica; NM: North Marianna; Aust: Australia; Wa: Wallis and Cha: Chagos. Asterisks
indicate type localities, arrows point at unexpected groupings and boxes highlight New Caledonian samples. 204x151mm (300 x 300 DPI)
Page 42 of 59 Journal of Phycology

Submitted M
anuscript
MP analysis of section Halimeda (L3) based on tufA sequence alignment. Bootstrap values (BP>70%) are indicated for MP/ NJ/ML analyses. Outgroup: Halimeda incrassata. Localities are noted as followed, Nic : Nicaragua; FP: French Polynesia; Mal: Maldives; Chest-NC: Chesterfield; New Caledonia; NC: New
Caledonia; Ye: Yemen; SrL: Sri-Lanka; Aust W: Western Australia; Sth Af: South Africa; Tanz: Tanzania; Bre: Brasil; It: Italia; Can: Canaries; Jam: Jamaica; Pan: Panama; Mex: Mexico; Bah: Bahamas; Thai: Thailand; Phi : Philippines; CR: Costa Rica. Asterisks indicate type localities and boxes highlight New
Caledonian samples. 125x289mm (300 x 300 DPI)
Page 43 of 59Journal of Phycology

Submitted M
anuscript
NJ analysis of section Pseudo-opuntia (L4) based on tufA sequence alignments. Bootstrap values (BP>70%) are indicated for NJ/MP/ML analyses. Outgroup: Halimeda macrophysa. Localities are noted as follows, Mal: Maldives; SrL: Sri Lanka; Ha: Hawaii; Chest-NC: Chesterfield, New Caledonia; PF: French Polynesia; Aust W: West Australia; Phi: Philippines; Sth Af: South Africa;Tanz: Tanzania; Flo: Florida; Jam: Jamaica; Bah:
Bahamas. Asterisks indicate type localities and the box highlight New Caledonian sample.
189x130mm (300 x 300 DPI)
Page 44 of 59 Journal of Phycology

Submitted M
anuscript
ML analysis of section Opuntia (L5) based on tufA sequence alignments. Bootstrap values (BP>70%) are indicated for ML/ NJ/MP analyses. Outgroup: Halimeda micronesica. Localities are noted as followed, FP:
French Polynesia, Aust: Australia, Phi: Philippines, Tanz: Tanzania, C. Isl: Cook Islands, Mal: Maldives, Soc:
Socotra, NC: New Caledonia, Wa: Wallis, Micro: Micronesia, Pal: Palau, NMI: North Marianna Island, Jam: Jamaica, Pan: Panama. Asterisks indicate type localities and boxes highlight New Caledonian samples.
.
194x289mm (300 x 300 DPI)
Page 45 of 59Journal of Phycology
Submitted Manuscript
Table S1. Oligonucleotide primers used for amplification and sequencing with detail of sequences, annealing temperature and previous
publication.
Primer name Sequence T (°C) source
tufA F : 5’ - TGAAACAGAAMAWCGTCATTATGC-3’
R : 5’ -CCTTCNCGAATMGCRAAWCGC-3’
52 Fama et al. 2002
RbcL binding site
rbcL1 : bp 7–26
rbcL2 : bp 6–29
rbcL3 : bp 689–667
rbcL 4 :bp 905–886
rbcL5 : bp 427–449
rbcL6 : bp 1396–1372
F : 5’-CCAMAAACWGAAACWAAAGC-3’
F : 5’-TCCAAAAACTGAAACTAAAGCAGG-3’
R : 5’-GCTTGWGMTTTRTARATWGCTTC-3’
R : 5’-TCAATAACCGCATGCATTGC-3’
F : 5’-GCTTATGCWAAAACATTYCAAGG-3’
R : 5’-AATTTCTTTCCAAACTTCACAAGC-3’
49
57
49
49
56
56
Hanyuda et al. (2000)
Hanyuda et al. (2000)
Hanyuda et al. (2000)
Hanyuda et al. (2000)
Lam et al.,(2006)
Lam et al. (2006)
UCP7 F : 5’-ATWTCDGCDCCATTWAGDCKVCC-3’
R : 5’-ATGGTWGGWCAWAAATTDGGTGAGTTT-3’
56 Provan et al. (2004)
Page 46 of 59 Journal of Phycology
Submitted M
anuscript
Table S2: List of Halimeda specimens and sequences used in this study.* represented samples
collected by G. Lasne, Biocenose. These are from biological points of environmental survey
of Vale New Caledonia, © Biocénose SARL.
Species
Specimen
number Geographical location GenBank Source
TufA rbcL
H. borneensis
W.R. Taylor
HV183b Tahiti, French
Polynesia (CP) AM049955
Verbruggen
et al 2006
FJ624514
Verbruggen
et al 2009b
CP08 1081
Moorea, French
Polynesia (CP) JN644647 this study
CP08 1157
Moorea, French
Polynesia (CP) JN644648 this study
CP08 886
Moorea, French
Polynesia (CP) JN644649 this study
CP08 908
Moorea, French
Polynesia (CP) JN644650 this study
IRD 5309
Grande Terre, New
Caledonia (WP) JN644651 JN644729 this study
IRD 5310
Grande Terre, New
Caledonia (WP) JN644730 this study
H. copiosa
Goreau &
E.A. Graham
H.0330 Discovery Bay,
Jamaica (CAR) EF667065
Verbruggen
et al.2007
FJ624508
Verbruggen
et al 2009b
HV460
Saint Ann, Saint Ann's
Bay, Lee Reef, Jamaica
(CAR) FJ624647
Verbruggen
et al. 2009b
H. cryptica
Colinvaux &
E.A. Graham H.0237
Discovery Bay,
Monitor Reef, Jamaica
(CAR) EF667056
Verbruggen
et al 2007b
HV958
Saint Ann, Priory, The
Wall, Jamaica (CAR)
HV
unpublished
HV483 St. Ann Parish, Priory,
Jamaica (CAR) EF667057
Verbruggen
et al 2007
FJ624496
Verbruggen
et al 2009b
H. cuneata
Hering H.0500 Brazil (WA) AY826358
Verbruggen
et al 2005
HEC 15286
Dickwella, Sri Lanka
(CI) FJ624537
Verbruggen
et al. 2009b
SOC252 Socotra, Yemen (WI) AY826356
Verbruggen
et al 2005
KZN2K4-21 South Africa (WI) AY826353
Verbruggen
et al 2005
KZNb2352 South Africa (WI) AY826354
Verbruggen
et al 2005
WA182 Western Australia, AY826355 Verbruggen
Page 47 of 59Journal of Phycology
Submitted M
anuscript
Australia (EI) et al 2005
JH12
Western Australia,
Australia (EI) FJ624538
Verbruggen
et al. 2009b
H.0550
Western Australia,
Rottnest Island,
Australia (EI) FJ624525
Verbruggen
et al 2009b
H.0502
Espirito Santo State,
Manguinhos, Praia,
Brazil (WA) FJ624518
Verbruggen
et al 2009b
KZN-b 2263
KwaZulu-Natal,
Sodwana, Jesser Point,
South Africa (WI) FJ624533
Verbruggen
et al 2009b
SOC 348 Socotra, Yemen (WI) FJ624532
Verbruggen
et al 2009b
HV560
Cebu, Daanbantayan,
Malapascua,
Philippines (WP) FJ624531
Verbruggen
et al 2009b
HV823
Luzon, Sorsogon,
Bulusan, Dancalan,
Philippines (WP) FJ624528
Verbruggen
et al 2009b
H.0195-2
KwaZulu-Natal,
Scottsburgh, South
Africa (WI) FJ624527
Verbruggen
et al 2009b
H.
cylindracea
Decaisne
SOC364 Socotra, Yemen (WI)
AM049956
Verbruggen
et al 2006
FJ624510
Verbruggen
et al 2009b
IRD 5325
Grande Terre, New
Caledonia (WP) JN644652 this study
IRD 5326
Grande Terre, New
Caledonia (WP) JN644653 JN644731 this study
H. discoidea
1 Decaisne IRD 5349*
Grande Terre, New
Caledonia (WP) JN644654 JN644732 this study
IRD 5320
Chesterfield, New
Caledonia (WP) JN644655 JN644733 this study
CP08 413
Moorea, French
Polynesia (CP) JN644656 this study
CP08 454
Moorea, French
Polynesia (CP) JN644657 this study
IRD 5303
Grande Terre, New
Caledonia (WP) JN644658 JN644734 this study
IRD 5305
Grande Terre, New
Caledonia (WP) JN644659 JN644735 this study
IRD 5306
Grande Terre, New
Caledonia (WP) JN644660 JN644736 this study
IRD 5307
Grande Terre, New
Caledonia (WP) JN644661 JN644737 this study
G.678
Bahía Culebra: Bahía
Virador, Costa Rica
(EP)
HV
unpublished
Page 48 of 59 Journal of Phycology
Submitted M
anuscript
G.677
Bahía Culebra: Punta
Arenita, Costa Rica
(EP)
HV
unpublished
G.672
Bajo Tres Hermanas,
Costa Rica (EP)
HV
unpublished
ODC1745
Basco, Philippines
(WP)
HV
unpublished
ODC1525
Diani Beach, Kenya
(WI)
HV
unpublished
G.673
Isla Ballena, Costa
Rica (EP)
HV
unpublished
G.195
Kepulauan Seribu, Air,
Indonesia (WP)
HV
unpublished
G.679
León: Playa El
Tránsito, Nicaragua
(EP)
HV
unpublished
G.687
León: Playa Hermosa,
Nicaragua (EP)
HV
unpublished
HV585
Logon Bay,
Malapascua Island,
Philippines (WP)
HV
unpublished
G.686
Managua: Masachapa,
Nicaragua (EP)
HV
unpublished
G.510
Molokai, Penguin
Bank, Hawaii, USA
(CP)
HV
unpublished
G.683
Nicaragua: El Hueco,
Nicaragua (EP)
HV
unpublished
OMII118 Oman (WI) AY826359
Verbruggen
et al 2005
HV58
Paea,Tahiti, French
Polynesia (CP)
HV
unpublished
G.684
Rivas: Guacalito,
Nicaragua (EP)
HV
unpublished
G.681
Rivas: La Peña Rota,
Nicaragua (EP)
HV
unpublished
G.676
San Josecito, Costa
Rica (EP)
HV
unpublished
SOC299 Socotra Yemen (WI) AY826360
Verbruggen
et al 2006
G.644 Thailand (WP)
HV
unpublished
H. discoidea
2 Decaisne H.0207
Gran Canaria, Spain
(EA) AY826361
Verbruggen
et al 2005
HV 495 Jamaica (CAR)
HV
unpublished
HV396 Jamaica (CAR) AY826362
Verbruggen
et al 2005
H.discoidea
Decaisne DML61724
Barrier Reef, Belize
(CAR) FJ624519
Verbruggen
et al 2009b
Page 49 of 59Journal of Phycology
Submitted M
anuscript
HV431
Saint Ann, Priory Bay,
Jamaica (CAR) FJ624520
Verbruggen
et al 2009b
AB038488
Hanyuda et
al 2000
H. distorta 1
(Yamada)
Hillis-
Colinvaux
WLS422.02
Wallis Island, Wallis
and Futuna (WP) FJ624644
Verbruggen
et al. 2009b
LL0378 Fiji (WP)
HV
unpublished
HV767
Luzon: Sorsogon,
Bulusan, Dapdap,
Philippines (WP) FJ624645
Verbruggen
et al. 2009b
HV275
Rangiroa Atoll, Tiputa,
French Polynesia (CP) FJ624709 FJ624505
Verbruggen
et al. 2009b
HV263
Rangiroa, Avatoru,
French Polynesia (CP) FJ624643
Verbruggen
et al. 2009b
H. distorta 2
(Yamada)
Hillis-
Colinvaux
H.0507
Cebu, Mactan Island,
Lapu Lapu, Philippines
(WP) FJ624653
Verbruggen
et al. 2009b
TZ0118
Kunduchi, Tanzania
(WI) FJ624658
Verbruggen
et al. 2009b
HV600
Malapascua,
Philippines (WP) FJ624654
Verbruggen
et al. 2009b
HV572
Malapascua, Shark
Point, Philippines
(WP) FJ624649
Verbruggen
et al. 2009b
HV1817 Maldives (CI)
HV
unpublished
W0135 Micronesia (WP)
HV
unpublished
W0165 Micronesia (WP)
HV
unpublished
HV127
Moorea, Entre deux
baies, French Polynesia
(CP) FJ624655
Verbruggen
et al. 2009b
W0028 Palau (WP)
HV
unpublished
W0069 Palau (WP)
HV
unpublished
HV199
Tahiti, Faaa, French
Polynesia (CP) FJ624656
Verbruggen
et al. 2009b
H.0287
Tahiti, French
Polynesia (CP) FJ624651
Verbruggen
et al. 2009b
HV188
Tahiti, Punaauia,
French Polynesia (CP) FJ624657
Verbruggen
et al. 2009b
WLS391.02
Wallis Island, Wallis
and Futuna (WP)
HV
unpublished
CP08 450
Moorea, French
Polynesia (CP) JN644662 this study
CP08 910
Moorea, French
Polynesia (CP) JN644663 this study
Page 50 of 59 Journal of Phycology
Submitted M
anuscript
IRD 4926 Maldives (CI) JN644664 this study
IRD 4915 Maldives (CI) JN644665 this study
IRD 5350*
Grande Terre, New
Caledonia (WP) JN644666 this study
IRD 4875 Maldives (CI) JN644667 this study
H.0280 New Caledonia (WP) FJ624652
Verbruggen
et al. 2009b
H.0475
Queensland, Australie
(WP) FJ624648
H. fragilis 1
W.R. Taylor
H.0125 Bile Bay, Guam (WP)
EF667058
Verbruggen
et al 2007
FJ624498
Verbruggen
et al 2009b
G.014
Tinian, Northern
Mariana Islands (WP) FJ624640
Verbruggen
et al. 2009b
IRD 5318
Chesterfield, New
Caledonia (WP) JN644668 JN644738 this study
H. fragilis 2
W.R. Taylor HV 1816 Maldives (CI)
HV
unpublished
HV1828 Maldives (CI)
HV
unpublished
TZ0233
Mtwara, Mnazi Bay,
Ruvula, Tanzania (WI) FJ624642
Verbruggen
et al. 2009b
HV766
Palawan, Busuanga,
Tangat, Philippines
(WP) FJ624641
Verbruggen
et al. 2009b
ODC1772 Philippines (WP)
HV
unpublished
IRD 4917 Maldives (CI) JN644669 this study
IRD 4916 Maldives (CI) JN644670 this study
IRD 4874 Maldives (CI) JN644671 this study
H. fragilis 3
W.R. Taylor IRD 5319
Chesterfield, New
Caledonia JN644672 JN644739 this study
IRD 5300
Grande Terre, New
Caledonia (WP) JN644673 JN644740 this study
H. gigas
W.R. Taylor HV1820
Dhaaparuh,Thiladhun
mathi, Maldives (CI)
HV
unpublished
HEC12910 Guam (WP) AY826364
Verbruggen
et al 2005
G.752
James Price
Point,Western
Australia, Australia
(EI)
HV
unpublished
HV769
Luzon, Sorsogon,
Bulusan, Dapdap,
Philippines (WP)
HV
unpublished
TZ0323
Mikindani, Tanzania
(WI) FJ624539
Verbruggen
et al. 2009b
IRD 4927 Maldives (CI) JN644674 this study
Page 51 of 59Journal of Phycology
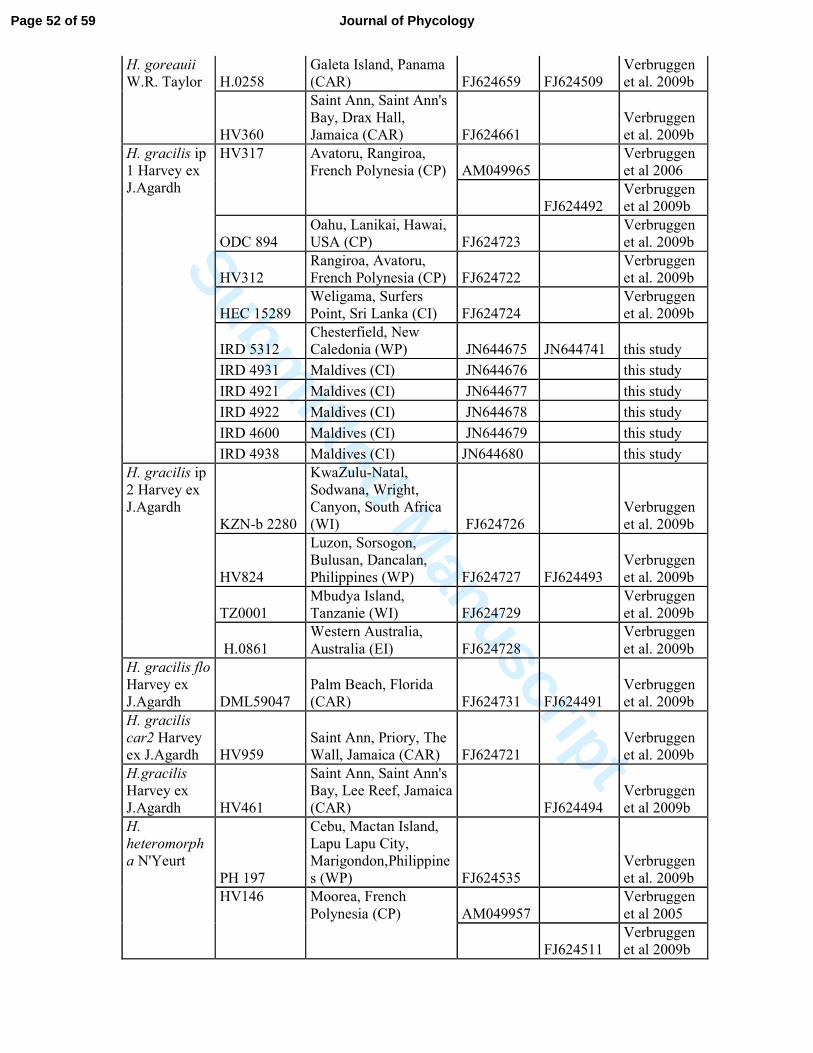
Submitted M
anuscript
H. goreauii
W.R. Taylor H.0258
Galeta Island, Panama
(CAR) FJ624659 FJ624509
Verbruggen
et al. 2009b
HV360
Saint Ann, Saint Ann's
Bay, Drax Hall,
Jamaica (CAR) FJ624661
Verbruggen
et al. 2009b
H. gracilis ip
1 Harvey ex
J.Agardh
HV317 Avatoru, Rangiroa,
French Polynesia (CP) AM049965
Verbruggen
et al 2006
FJ624492
Verbruggen
et al 2009b
ODC 894
Oahu, Lanikai, Hawai,
USA (CP) FJ624723
Verbruggen
et al. 2009b
HV312
Rangiroa, Avatoru,
French Polynesia (CP) FJ624722
Verbruggen
et al. 2009b
HEC 15289
Weligama, Surfers
Point, Sri Lanka (CI) FJ624724
Verbruggen
et al. 2009b
IRD 5312
Chesterfield, New
Caledonia (WP) JN644675 JN644741 this study
IRD 4931 Maldives (CI) JN644676 this study
IRD 4921 Maldives (CI) JN644677 this study
IRD 4922 Maldives (CI) JN644678 this study
IRD 4600 Maldives (CI) JN644679 this study
IRD 4938 Maldives (CI) JN644680 this study
H. gracilis ip
2 Harvey ex
J.Agardh
KZN-b 2280
KwaZulu-Natal,
Sodwana, Wright,
Canyon, South Africa
(WI) FJ624726
Verbruggen
et al. 2009b
HV824
Luzon, Sorsogon,
Bulusan, Dancalan,
Philippines (WP) FJ624727 FJ624493
Verbruggen
et al. 2009b
TZ0001
Mbudya Island,
Tanzanie (WI) FJ624729
Verbruggen
et al. 2009b
H.0861
Western Australia,
Australia (EI) FJ624728
Verbruggen
et al. 2009b
H. gracilis flo
Harvey ex
J.Agardh DML59047
Palm Beach, Florida
(CAR) FJ624731 FJ624491
Verbruggen
et al. 2009b
H. gracilis
car2 Harvey
ex J.Agardh HV959
Saint Ann, Priory, The
Wall, Jamaica (CAR) FJ624721
Verbruggen
et al. 2009b
H.gracilis
Harvey ex
J.Agardh HV461
Saint Ann, Saint Ann's
Bay, Lee Reef, Jamaica
(CAR) FJ624494
Verbruggen
et al 2009b
H.
heteromorph
a N'Yeurt
PH 197
Cebu, Mactan Island,
Lapu Lapu City,
Marigondon,Philippine
s (WP) FJ624535
Verbruggen
et al. 2009b
HV146 Moorea, French
Polynesia (CP) AM049957
Verbruggen
et al 2005
FJ624511
Verbruggen
et al 2009b
Page 52 of 59 Journal of Phycology
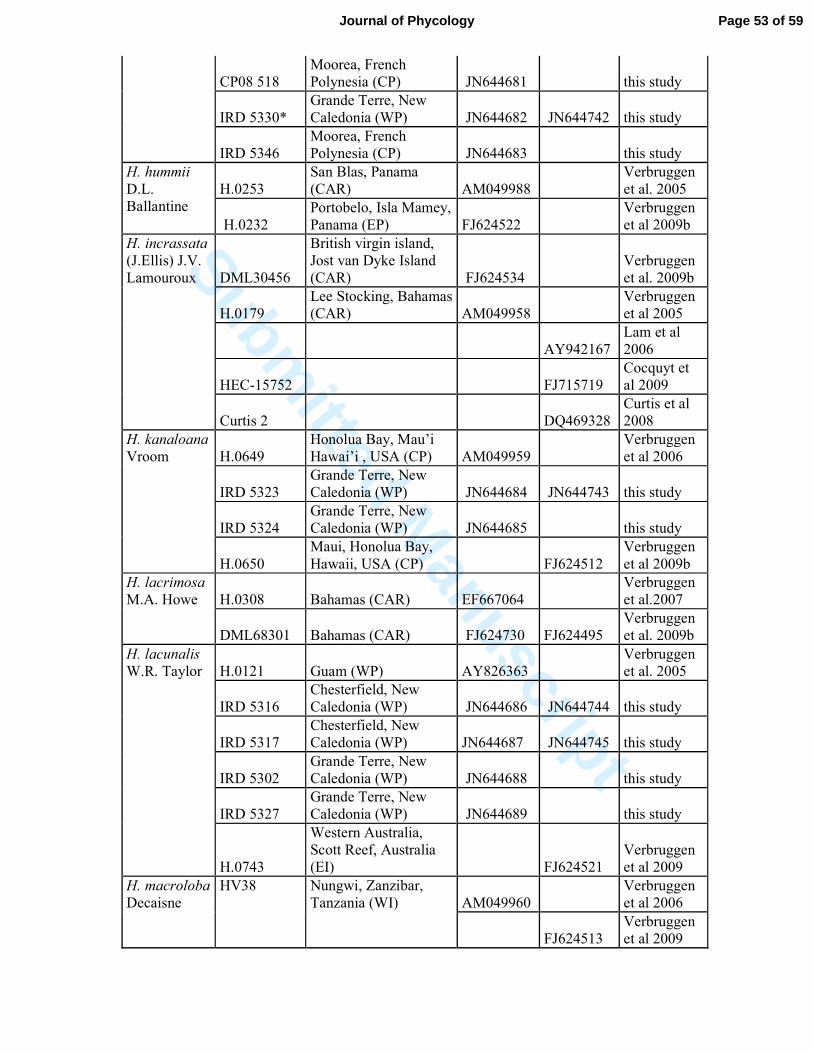
Submitted M
anuscript
CP08 518
Moorea, French
Polynesia (CP) JN644681 this study
IRD 5330*
Grande Terre, New
Caledonia (WP) JN644682 JN644742 this study
IRD 5346
Moorea, French
Polynesia (CP) JN644683 this study
H. hummii
D.L.
Ballantine
H.0253
San Blas, Panama
(CAR) AM049988
Verbruggen
et al. 2005
H.0232
Portobelo, Isla Mamey,
Panama (EP) FJ624522
Verbruggen
et al 2009b
H. incrassata
(J.Ellis) J.V.
Lamouroux DML30456
British virgin island,
Jost van Dyke Island
(CAR) FJ624534
Verbruggen
et al. 2009b
H.0179
Lee Stocking, Bahamas
(CAR) AM049958
Verbruggen
et al 2005
AY942167
Lam et al
2006
HEC-15752 FJ715719
Cocquyt et
al 2009
Curtis 2 DQ469328
Curtis et al
2008
H. kanaloana
Vroom H.0649
Honolua Bay, Mau’i
Hawai’i , USA (CP) AM049959
Verbruggen
et al 2006
IRD 5323
Grande Terre, New
Caledonia (WP) JN644684 JN644743 this study
IRD 5324
Grande Terre, New
Caledonia (WP) JN644685 this study
H.0650
Maui, Honolua Bay,
Hawaii, USA (CP) FJ624512
Verbruggen
et al 2009b
H. lacrimosa
M.A. Howe H.0308 Bahamas (CAR) EF667064
Verbruggen
et al.2007
DML68301 Bahamas (CAR) FJ624730 FJ624495
Verbruggen
et al. 2009b
H. lacunalis
W.R. Taylor H.0121 Guam (WP) AY826363
Verbruggen
et al. 2005
IRD 5316
Chesterfield, New
Caledonia (WP) JN644686 JN644744 this study
IRD 5317
Chesterfield, New
Caledonia (WP) JN644687 JN644745 this study
IRD 5302
Grande Terre, New
Caledonia (WP) JN644688 this study
IRD 5327
Grande Terre, New
Caledonia (WP) JN644689 this study
H.0743
Western Australia,
Scott Reef, Australia
(EI) FJ624521
Verbruggen
et al 2009
H. macroloba
Decaisne
HV38 Nungwi, Zanzibar,
Tanzania (WI) AM049960
Verbruggen
et al 2006
FJ624513
Verbruggen
et al 2009
Page 53 of 59Journal of Phycology
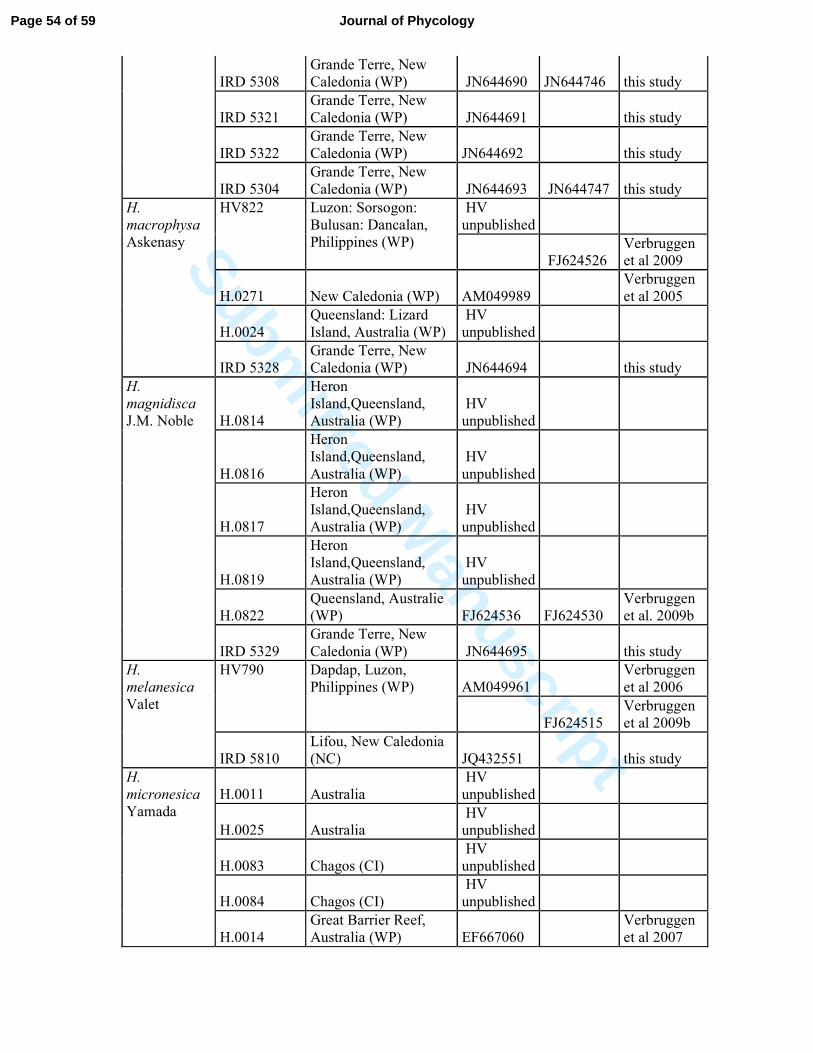
Submitted M
anuscript
IRD 5308
Grande Terre, New
Caledonia (WP) JN644690 JN644746 this study
IRD 5321
Grande Terre, New
Caledonia (WP) JN644691 this study
IRD 5322
Grande Terre, New
Caledonia (WP) JN644692 this study
IRD 5304
Grande Terre, New
Caledonia (WP) JN644693 JN644747 this study
H.
macrophysa
Askenasy
HV822 Luzon: Sorsogon:
Bulusan: Dancalan,
Philippines (WP)
HV
unpublished
FJ624526
Verbruggen
et al 2009
H.0271 New Caledonia (WP) AM049989
Verbruggen
et al 2005
H.0024
Queensland: Lizard
Island, Australia (WP)
HV
unpublished
IRD 5328
Grande Terre, New
Caledonia (WP) JN644694 this study
H.
magnidisca
J.M. Noble H.0814
Heron
Island,Queensland,
Australia (WP)
HV
unpublished
H.0816
Heron
Island,Queensland,
Australia (WP)
HV
unpublished
H.0817
Heron
Island,Queensland,
Australia (WP)
HV
unpublished
H.0819
Heron
Island,Queensland,
Australia (WP)
HV
unpublished
H.0822
Queensland, Australie
(WP) FJ624536 FJ624530
Verbruggen
et al. 2009b
IRD 5329
Grande Terre, New
Caledonia (WP) JN644695 this study
H.
melanesica
Valet
HV790 Dapdap, Luzon,
Philippines (WP) AM049961
Verbruggen
et al 2006
FJ624515
Verbruggen
et al 2009b
IRD 5810
Lifou, New Caledonia
(NC) JQ432551 this study
H.
micronesica
Yamada
H.0011 Australia
HV
unpublished
H.0025 Australia
HV
unpublished
H.0083 Chagos (CI)
HV
unpublished
H.0084 Chagos (CI)
HV
unpublished
H.0014
Great Barrier Reef,
Australia (WP) EF667060
Verbruggen
et al 2007
Page 54 of 59 Journal of Phycology
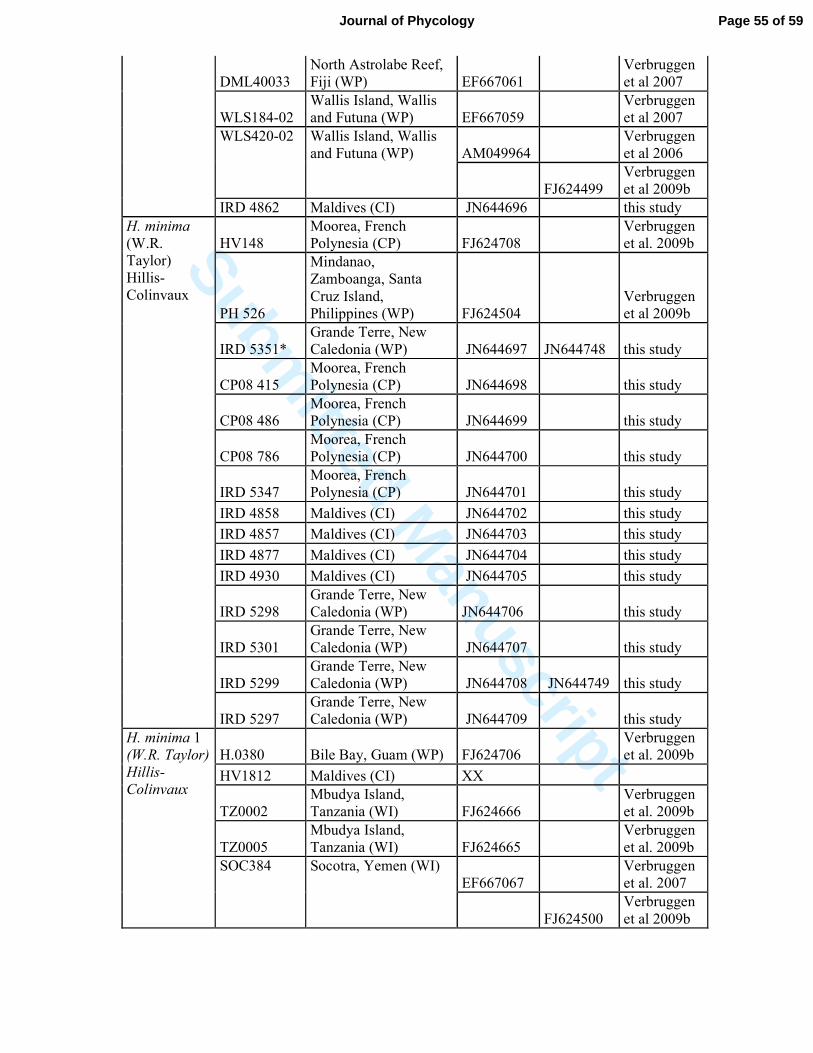
Submitted M
anuscript
DML40033
North Astrolabe Reef,
Fiji (WP) EF667061
Verbruggen
et al 2007
WLS184-02
Wallis Island, Wallis
and Futuna (WP) EF667059
Verbruggen
et al 2007
WLS420-02 Wallis Island, Wallis
and Futuna (WP) AM049964
Verbruggen
et al 2006
FJ624499
Verbruggen
et al 2009b
IRD 4862 Maldives (CI) JN644696 this study
H. minima
(W.R.
Taylor)
Hillis-
Colinvaux
HV148
Moorea, French
Polynesia (CP) FJ624708
Verbruggen
et al. 2009b
PH 526
Mindanao,
Zamboanga, Santa
Cruz Island,
Philippines (WP) FJ624504
Verbruggen
et al 2009b
IRD 5351*
Grande Terre, New
Caledonia (WP) JN644697 JN644748 this study
CP08 415
Moorea, French
Polynesia (CP) JN644698 this study
CP08 486
Moorea, French
Polynesia (CP) JN644699 this study
CP08 786
Moorea, French
Polynesia (CP) JN644700 this study
IRD 5347
Moorea, French
Polynesia (CP) JN644701 this study
IRD 4858 Maldives (CI) JN644702 this study
IRD 4857 Maldives (CI) JN644703 this study
IRD 4877 Maldives (CI) JN644704 this study
IRD 4930 Maldives (CI) JN644705 this study
IRD 5298
Grande Terre, New
Caledonia (WP) JN644706 this study
IRD 5301
Grande Terre, New
Caledonia (WP) JN644707 this study
IRD 5299
Grande Terre, New
Caledonia (WP) JN644708 JN644749 this study
IRD 5297
Grande Terre, New
Caledonia (WP) JN644709 this study
H. minima 1
(W.R. Taylor)
Hillis-
Colinvaux
H.0380 Bile Bay, Guam (WP) FJ624706
Verbruggen
et al. 2009b
HV1812 Maldives (CI) XX
TZ0002
Mbudya Island,
Tanzania (WI) FJ624666
Verbruggen
et al. 2009b
TZ0005
Mbudya Island,
Tanzania (WI) FJ624665
Verbruggen
et al. 2009b
SOC384 Socotra, Yemen (WI)
EF667067
Verbruggen
et al. 2007
FJ624500
Verbruggen
et al 2009b
Page 55 of 59Journal of Phycology
Submitted M
anuscript
H.0848
Western Australia,
Rowley Shoals,
Mermaid Reef,
Australia (EI) FJ624664
Verbruggen
et al. 2009b
H.0746
Western Australia,
Scott Reef,
Australia(EI) FJ624663
Verbruggen
et al. 2009b
H. minima 2
(W.R.
Taylor)
Hillis-
Colinvaux
H.0382
Apra Harbour, Guam
(WP) FJ624669 FJ624503
Verbruggen
et al. 2009b
H.0381 Guam (WP) FJ624707
Verbruggen
et al. 2009b
G.167 Indonesia (WP)
HV
unpublished
HV765
Palawan, Busuanga,
Tangat, Philippines
(WP) FJ624668
Verbruggen
et al. 2009b
H. minima 3
(W.R.
Taylor)
Hillis-
Colinvaux
HV656
Bohol, Cabilao,
Philippines (WP) FJ624710
Verbruggen
et al. 2009b
HV693
Bohol, Panglao, Alona
Beach, Philippines
(WP) FJ624682
Verbruggen
et al. 2009b
HV695
Bohol, Panglao, Alona
Beach, Philippines
(WP) FJ624676
Verbruggen
et al. 2009b
H.0395 Cook Islands (CP) FJ624683
Verbruggen
et al. 2009b
HV63
Moorea, Cook Bay,
French Polynesia (CP) FJ624671
Verbruggen
et al. 2009b
HV720
Panglao, Philippines
(WP) FJ624675
Verbruggen
et al. 2009b
H.0336
Queensland, Lizard
Island, Australia (WP) FJ624684
Verbruggen
et al. 2009b
H.0332
Queensland, Lizard
Island, Australia (WP) FJ624674
Verbruggen
et al. 2009b
HV163
Tahiti, Arue, French
Polynesia (CP) FJ624677
Verbruggen
et al. 2009b
H.0286
Tahiti, French
Polynesia (CP) FJ624685
Verbruggen
et al. 2009b
HV189
Tahiti, Punaauia,
French Polynesia (CP) FJ624681
Verbruggen
et al. 2009b
H. minima.4
(W.R.
Taylor)
Hillis-
Colinvaux
HV791
Luzon: Sorsogon,
Bulusan, Dapdap,
Philippines (WP) FJ624687 FJ624502
Verbruggen
et al. 2009b
H.0990
Vanua Levu,
Cakaulevu reef, Fiji
(WP) FJ624686
Verbruggen
et al. 2009b
H.0991
Vanua Levu,
Cakaulevu reef, Fiji
(WP) FJ624688
Verbruggen
et al. 2009b
H. monile
(J.Ellis &
H.0034 Galeta, Panama (CAR)
AM049962
Verbruggen
et al 2005
Page 56 of 59 Journal of Phycology
Submitted M
anuscript
Solander)
J.V.
Lamouroux
FJ624516
Verbruggen
et al 2009b
Curtis 3
Florida, Tarpon
Springs, USA (CAR) DQ469329
Verbruggen
et al 2009b
H. opuntia
(Linnaeus)
Lamouroux HV591
Cebu, Daanbantayan,
Malapascua, Logon
Bay, Philippines (WP) FJ624690
Verbruggen
et al. 2009b
HV61
Cook Bay, Moorea
French, Polynesia (CP) AM049967
Verbruggen
et al 2005
CP08 414
Moorea, French
Polynesia (CP) JN644710 this study
CP08 484
Moorea, French
Polynesia (CP) JN644711 this study
CP08 909
Moorea, French
Polynesia (CP) JN644712 this study
IRD 5352*
Grande Terre, New
Caledonia (WP) JN644713 this study
AY942174
Lam et al
2006
AB038489
Hanyuda et
al 2000
H. pygmaea
Verbruggen,
D.S.Littler &
M.M.Littler
DML40143 North Astrolabe Reef,
Fiji (WP) EF667062
Verbruggen
et al 2007
FJ624497
Verbruggen
et al 2009b
H. renschii
Hauck ODC1671 Philippines (WP)
HV
unpublished
ODC1743 Philippines (WP)
HV
unpublished
ODC1744 Philippines (WP)
HV
unpublished
HV7
Zanzibar, Mnemba
Atoll, Tanzania (CP) FJ624691 FJ624501
Verbruggen
et al. 2009b
TZ0468
Zanzibar, Mnemba,
Tanzania (WI) FJ624693
Verbruggen
et al. 2009b
TZ0480
Zanzibar, Mnemba,
Tanzania (WI) FJ624717
Verbruggen
et al. 2009b
H. scabra
M.A.Howe DML68229
Green Turtle Cay,
Bahamas (CAR) FJ624540
Verbruggen
et al. 2009b
DML68311
Bahamas: Green Turtle
Cay (CAR) FJ624523
Verbruggen
et al 2009b
H. simulans
M.A.Howe
HV449 Discovery Bay,
Jamaica (CAR) AM049963
Verbruggen
et al 2005
FJ624517
Verbruggben
et al 2009
H. cf stuposa IRD 5311
Grande Terre, New
Caledonia (WP) JN644714 this study
H. taenicola
W.R. Taylor HV285-1
Rangiroa, French
Polynesia (CP) AY826365
Verbruggen
et al 2005
Page 57 of 59Journal of Phycology
Submitted M
anuscript
HV1813
Thiladhunmathi:
Hanimaadhoo,
Maldives (CI)
HV
unpublished
HV285-1 Tiputa, Rangiroa
French Polynesia (CP) AM049966
Verbruggen
et al 2005
FJ624529
Verbruggen
et al 2009b
IRD 5314
Chesterfield, New
Caledonia (WP) JN644715 JN644750 this study
IRD 5315
Chesterfield, New
Caledonia JN644716 JN644751 this study
IRD 5313
Chesterfield, New
Caledonia JN644717 JN644752 this study
CP08 411
Moorea, French
Polynesia (CP) JN644718 this study
CP08 457
Moorea, French
Polynesia (CP) JN644719 this study
CP08 458
Moorea, French
Polynesia (CP) JN644720 this study
CP08 709
Moorea, French
Polynesia (CP) JN644721 this study
IRD 4860 Maldives (CI) JN644722 this study
IRD 4861 Maldives (CI) JN644723 JN644753 this study
H. tuna
(J.Ellis &
Solander)
J.V.Lamouro
ux
H.0113 Italy (MED) AY826366
Verbruggen
et al 2005
H.0231
Puerto Morelos,
Mexico (CAR) AY826367
Verbruggen
et al 2005
H.0231
Zanzibar: Pongwe,
Tanzania (WI) AY826367
Verbruggen
et al 2005
DML51688
Jamaica: Morant Cays
(CAR) FJ624524
Verbruggen
et al 2009b
AY942177
Verbruggen
et al 2009b
H.
velasquezii
W.R. Taylor HV768
Luzon: Sorsogon,
Bulusan, Dapdap,
Philippines (WP) FJ624699
Verbruggen
et al. 2009b
HV779
Luzon: Sorsogon,
Bulusan, Dapdap,
Philippines (WP) FJ624698
Verbruggen
et al. 2009
TZ0066
Mbudya Island,
Tanzania (WI) FJ624700
Verbruggen
et al. 2009b
HEC14003
Mtwara, Mnazi Bay,
Mana Hawanja Island,
Tanzania (WI) FJ624696
Verbruggen
et al. 2009b
TZ0163
Mtwara, Mnazi Bay,
Ruvula, Tanzania (WI) FJ624701
Verbruggen
et al. 2009b
HV746
Palawan, Busuanga,
Cagbatan, Philippines
(WP) FJ624697
Verbruggen
et al. 2009b
G.015 Talofofo Bay, Guam FJ624695 Verbruggen
Page 58 of 59 Journal of Phycology
Submitted M
anuscript
(WP) et al. 2009b
IPAN010
Talofofo, Mana Bay,
Ipan, Guam (WP) FJ624711
Verbruggen
et al. 2009b
TZ0464
Zanzibar, Mnemba,
Tanzania (WI) FJ624702
Verbruggen
et al. 2009b
TZ0547
Zanzibar, Nungwi,
Tanzania (WI) FJ624703
Verbruggen
et al. 2009b
TZ0839
Zanzibar, Paje,
Tanzania (WI) FJ624715
Verbruggen
et al. 2009b
TZ0745
Zanzibar, Pongwe,
Tanzania (WI) FJ624718
Verbruggen
et al. 2009b
IRD 5296
Grande Terre, New
Caledonia (WP) JN644724 JN644754 this study
IRD 4935 Maldives (CI) JN644725
HV779
Luzon, Sorsogon,
Bulusan, Dapdap,
Philippines (WP) FJ624507
Verbruggen
et al 2009b
H. xishaensis
Tseng and
M.L. Dong
H.0122 Guam (WP) AY826357
Verbruggen
et al 2005
ODC1564
Kinondo Reef, Kenya
(WI)
HV
unpublished
TZ0188
Mtwara: Ruvula,
Tanzania (WI)
HV
unpublished
HV731
Palawan: Busuanga:
Uson, Philippines
(WP)
HV
unpublished
H.0389
Providencia, Colombia
(CAR)
HV
unpublished
HV1814
Thiladhunmathi:
Hanimaadhoo,
Maldives (CI)
HV
unpublished
HV33
Zanzibar: Pongwe,
Tanzania (WI)
HV
unpublished
IRD 4932 Maldives (CI) JN644726 this study
IRD 4867 Maldives (CI) JN644727
this study
IRD 4933 Maldives (CI) JN644728 this study
H. no.ID.1 G.007
Maug, Northern
Mariana Islands (WP) FJ624704
Verbruggen
et al. 2009b
H. no.ID.1 G.011
Asuncion, Northern
Mariana Islands (WP) FJ624705
Verbruggen
et al. 2009b
H. no.ID.1 HVLL0367 Fiji (WP)
HV
unpublished
Page 59 of 59Journal of Phycology





























